 Belén Ponce
Belén Ponce Agustín Zamora-Quiroz
Agustín Zamora-Quiroz Ernesto González
Ernesto González Rodrigo Andler
Rodrigo Andler Alvaro Díaz-Barrera
Alvaro Díaz-Barrera- 1Escuela de Ingeniería Bioquímica, Pontificia Universidad Católica de Valparaíso, Valparaíso, Chile
- 2Universidad Técnica Federico Santa María, Valparaíso, Chile
- 3Departamento de Ingeniería Química y de Materiales, Facultad de Ciencias Químicas, Universidad Complutense de Madrid, Madrid, Spain
- 4Escuela de Ingeniería en Biotecnología, Universidad Católica del Maule, Talca, Chile
Alginates are polysaccharides composed of (1–4)-β-D-mannuronic acid (M) and α-L-guluronic acid (G), whose proportions influence their rheological properties and a wide range of applications in the food, pharmaceutical, and biomedical industries. Azotobacter vinelandii, a Gram-negative bacterium, has been studied for its ability to produce alginate due to its capacity to fix atmospheric nitrogen and its high respiratory activity. The biosynthesis of alginate in Azotobacter vinelandii involves precursor synthesis, polymerization, modification, and secretion, which are regulated by complex mechanisms, including the secondary messenger c-di-GMP. This regulatory network links cellular respiration with alginate yield and molecular characteristics. Fermentation strategies show that high oxygen transfer rates (OTRs) enhance alginate production, whereas low OTRs favor the synthesis of alginate with higher molecular weights and higher G/M ratios, which are crucial for advanced applications such as hydrogels and drug delivery systems. Insights into these biosynthetic and regulatory processes enable scalable production of high-quality alginate, bridging laboratory research with industrial applications and expanding its potential in the biotechnological and medical fields.
Introduction
Alginates are linear biopolymers present in the cell walls of brown algae and are produced by different bacteria, such as Pseudomonas spp. and Azotobacter sp. (Zhang et al., 2022; Hendawy et al., 2019; Bonartseva et al., 2017). These bacteria produce alginate as an exopolysaccharide in response to certain physiological processes, such as surface/cell adhesion, resistance to cytotoxic compounds, and dormant cell differentiation. Azotobacter vinelandii can also produce this biopolymer under vegetative growth conditions (Núñez et al., 2022; Ma et al., 2022; Castillo et al., 2013). The U.S. Food and Drug Administration (FDA) has cataloged the safe use of sodium alginate obtained from brown algae for diverse applications. Alginate can be used in the food, pharmaceutical, and cosmetic industries because of its high biocompatibility and advantageous physical–chemical properties, which encourage its use as a stabilizer and viscosifier agent, in addition to its ability to form a strong and stable hydrogel when interacting with divalent ions such as calcium (Hoefer et al., 2015).
Alginate is chemically composed of two monomers, β-D mannuronic acid (M) and its C-5 epimer α-L guluronic acid (G) (Figure 1). Its structure and conformation are influenced by the linking of these monomers in different sequences, forming patterns called blocks (Bojorges et al., 2023; Sanchez-Ballester et al., 2021; Hecht and Srebnik, 2016). M-residues can be acetylated on C-2 and/or C-3 during alginate synthesis in bacteria, which prevents the action of alginate-modifying enzymes (Cheng et al., 2020; Hecht and Srebnik, 2016). The chemical composition of alginate determines its properties and, therefore, its possible applications; for example, alginate with a relatively high molecular weight (between 400 and 3,420 kDa) has a higher viscosifying capacity, which is favorable for its use as a food additive (Hoefer et al., 2015; Schmid and Picker-Freyer, 2009). In the case of biomedical applications, a higher G/M ratio (with a ratio from 2.3 to 9) is associated with the presence of longer GG-blocks which interacts with divalent cations such as Ca2+ allowing crosslinking in between the polymeric chains favoring more physical stable and flexible hydrogels, that could be used to formulate wound dressings or other biomedical devices that demand higher physical resistance in contrast alginates with higher M/G ratio could be associated with lower GG-block presences thus preventing crosslinking resulting in more brittle hydrogel which disfavors its application in biomedical products (Sanchez-Ballester et al., 2021; Ramos et al., 2018; Pritchard et al., 2017; Hoefer et al., 2015). Monomeric characterization of alginates begins with controlled depolymerization by alkaline hydrolysis to obtain short oligomers. These short oligomers can then be analyzed by HPLC, often coupled with refractive index detection, or by colorimetric assays such as the carbazole reaction. Furthermore, to assess polymer sequential analysis, spectroscopic methods such as High Resolution Nuclear Magnetic Resonance Spectroscopy (HNMR), which distinguishes the sequence and block distribution of M and G residues, and Fourier Transform Infrared Spectroscopy (FTIR-ATR) spectroscopy, which confirms the characteristic vibrational bands of carboxylate and C–O–C, allow the differential detection of each monomer by peak intensity. Nevertheless, NMR is considered the preferred method for alginate characterization due to its ability to assess monomer frequencies and the presence of dyads (MM, MG/GM, or GG) and even their possible triads (Díaz-Barrera et al., 2021; Ertesvåg, 2015; Gaytán et al., 2012; Peña et al., 2006).
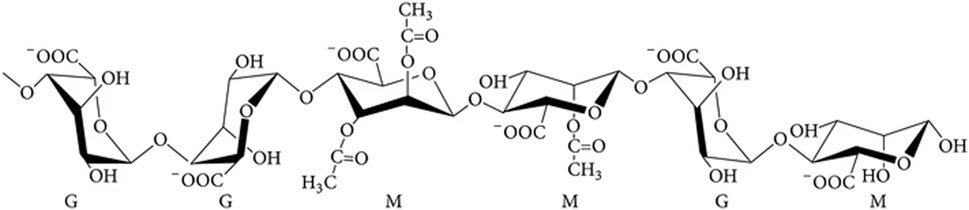
Figure 1. Chemical composition of bacterial alginate. Mannuronic acid (M) and guluronic acid (G) residues in the polymer.
Alginate has high biocompatibility and in vivo biodegradability (Martău et al., 2019), and it and its derivatives has been studied in diverse biomedical and biotechnological applications (Baldassarre et al., 2020; Hecht and Srebnik, 2016; Topuz et al., 2012; Yang et al., 2011), such as 3D cellular scaffolds in tissue engineering, wound dressings with better physical performance and delivery systems of bioactive compounds or as carriers for small therapeutic proteins (Qamar et al., 2024; Tan et al., 2023; Mallakpour and Mohammadi, 2022; Aguero et al., 2021; Mallakpour et al., 2021; Szekalska et al., 2016). The biomedical industry requires the use of materials with defined chemical compositions and of high purity. In the case of alginate, chemical characteristics such as molecular weight, the G/M ratio, and the presence of O-acetyl groups are highly relevant for its application in biomedical devices. In this context, different processes for obtaining and purifying alginate cannot ensure constant alginate quality in the previously presented terms (Sondermeijer et al., 2016; Ertesvåg, 2015; Bales et al., 2013; Langlois et al., 2009). Although most commercial alginates are of algal origin, various drawbacks are associated with their source, extraction, and refinement (Torres et al., 2019; Paredes-Juarez et al., 2014; Qi et al., 2009; Dusseault et al., 2006). In fact, Dusseault et al. (2006) demonstrated the presence of contaminants in various samples of algal and commercial alginates treated with different purification processes, obtaining products free of endotoxins, polyphenols, and proteins. Nevertheless, evidence from treatments with alginate beads has still revealed the presence of immunogenic proteins that compromise their therapeutic use. For this reason, it has been proposed that producing bacterial alginate in bioreactors using defined media would prevent the occurrence of these immunogenic proteins, while also ensuring the absence of endotoxins, polyphenols, and other contaminants. Futhermore, the chemical structure of algal alginate can vary due to climatic conditions, making it difficult to ensure a consistently well-characterized product. Additionally, the presence of phenolic compounds, proteins, lipids, and even mannitol bound to alginate requires additional purification steps such as precipitation which, although technically simple, can increase production costs and reduce overall process efficiency for many applications (food, pharma, and biomedical) (Bojorges et al., 2023; Saji et al., 2022). Finally, there is great concern about the ecological impact on marine fauna due to the harvesting of algal species (Bularz et al., 2022). These problems have been addressed by the study and elaboration of bioprocesses that involve the use of different biocatalysts, such as alginate-modifying enzymes, and fermentation with alginate-producing bacteria to ensure the production of high-quality alginates (Chuacharoen et al., 2022; Díaz-Barrera et al., 2021; Ertesvåg et al., 2017).
Although the properties of alginates have some limitations in terms of their G/M composition, it is interesting the current possibility of making in vitro modifications of alginate. Thus, in vitro modification of alginate can be carried out using alginate-lyase and C-5 mannuronan epimerases. Alginate-lyases are enzymes that catalyze the β-elimination of the 4-O-glycosidic bond between the inner residues of alginate, and the activity of this enzyme varies according to the neighboring residues of the bond. These enzymes can be used during structural sequencing of the polymer constituents; additionally, they are useful for obtaining bioactive molecules such as phenolic compounds from algae and help reduce alginate viscosity for dental applications. In addition, alginate epimerases catalyze the epimerization of M-residues to G-residues, which increases the numbers of G- and GM-blocks inside the alginate chain (Mollah et al., 2021). Alginate epimerases have been shown to produce alginates with high G contents and to modify the monomeric composition of alginates extracted from different algae to maintain a constant chemical composition (Tøndervik et al., 2020). Despite this background, even these processes are in the development stage and face problems with scaling up to enzymatic reactor quantities.
Azotobacter vinelandii is a bacterium capable of fixing atmospheric nitrogen and produces alginate during vegetative growth; during its differentiation process to a dormant state, it forms cysts (Gimmestad et al., 2009). Azotobacter vinelandii secretes alginate under vegetative growth conditions as part of the protection mechanism against oxygen inhibition of the nitrogenase complex (Inomura et al., 2017). During encystment, A. vinelandii produces an alginate layer that surrounds the dormant cell, called the central body. The two layers surrounding this cell are composed of alginate, alkylresorcinol and proteins (Rosales-Cruz et al., 2024). The outer layer, named the exine layer, is thin, rigid, and structured due to the inclusion of G-blocks by the action of C-5 mannuronan epimerases. In contrast, the inner layer (intine) has more MG- and MM-blocks, favoring an amorphous structure (Gawin et al., 2020).
The alginate biosynthetic pathways are similar in A. vinelandii and Pseudomonas aeruginosa and can be divided into four stages: precursor synthesis, polymerization, modification, and secretion (Dudun et al., 2022; Urtuvia et al., 2017; Maleki et al., 2017; Rehman et al., 2013; Franklin et al., 2011). Even though this polymer plays different roles in both microorganisms, the cluster of genes and regulatory events involved in alginate biosynthesis differ in each microorganism, affecting its chemical structure and thus its quality for biotechnological applications. These regulatory mechanisms are coordinated concerning the physiological state of the cell, which in turn depends on the environmental stimuli in which the cell develops (Núñez et al., 2022). Consequently, the complete description of the alginate biosynthetic pathway is complex, and the pathways must be separated into stages to better understand their mechanisms.
Furthermore, this review will analyze some strategies to optimize and improve alginate production. This review aims to present and discuss recent advances in biosynthesis, regulatory mechanisms, and the impact of oxygen transfer on alginate production.
Alginate applications
Alginate is a well-known biopolymer with a variety of applications in different industries (Table 1). Traditionally, alginate has been applied mainly as a viscosifier in the food industry or because of its gelifying capacity. During the last few years, novel developments in the biomedical, pharmacological, and cosmetic industries have drawn attention to the vast importance of biopolymers. In this context, alginate has emerged as a useful resource in the development of new technologies, being most relevant as part of novel drug delivery systems and in the development of biomedical devices (Tomić et al., 2023).
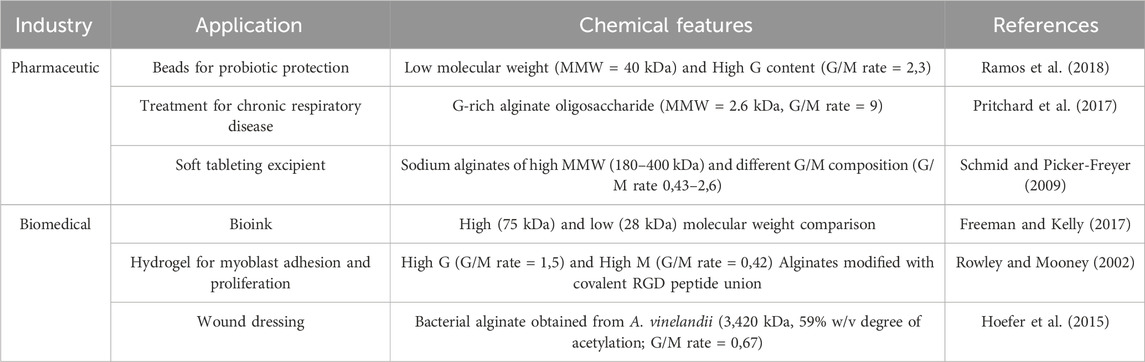
Table 1. Applications of alginate.
The use of alginate as a biomaterial for developing biomedical devices is closely linked to its gelling properties. Although the simplest way to generate alginate hydrogels is to use divalent cations for cross-linking, several studies have proposed chemical modification techniques or the combination with other polymers to increase the degree of cross-linking between alginate chains. These strategies aim to enhance the structural stability and mechanical strength of the hydrogels while maintaining sufficient flexibility and conformability, which are essential for applications such as wound dressings (Ramos et al., 2018; Freeman and Kelly, 2017; Pritchard et al., 2017; Hoefer et al., 2015; Schmid and Picker-Freyer, 2009; Rowley and Mooney, 2002).
Compared with algal alginate, the resulting bacterial alginate had a greater molecular weight, ranging from 400 to 3,420 kDa, a higher G/M ratio (with a ratio from 2.3 to 9), and a high degree of acetylation (>50% w/w), explaining the greater water retention capacity and physical stability of the hydrogels made with bacterial alginate (Rashidzadeh et al., 2020; Pritchard et al., 2017; Hoefer et al., 2015; Schmid and Picker-Freyer, 2009). The authors developed alginate-based antibacterial hydrogel beads with a magnetic drug delivery response due to the inclusion of ferroferric oxide (Fe3O4) and silver (Ag) nanoparticles, which presented high antimicrobial activity against Escherichia coli and Staphylococcus aureus (Rashidzadeh et al., 2020).
In a pharmacological context, alginates have attracted attention as raw materials for many applications (Sanchez-Ballester et al., 2021). Alginate oligosaccharides are considered bioactive compounds with wide-ranging biological activities, such as antihypertensive, hypolipidemic, immunoregulatory, and hypoglycemic properties, among other possible applications (Xing et al., 2020). A clinical study revealed that daily and oral administration of 10 mg of 1 kDa alginate oligosaccharide reduced the progression of osteosarcoma by improving patients’ anti-inflammatory and antioxidant capacities and relieving cancer-related inflammation (Chen et al., 2017). Additionally, alginate is more extensively used in the pharmaceutical industry as a disintegrant in compressed tablets, tablet binders, tastemakers, and controlled-release matrices in soft tableting (Sanchez-Ballester et al., 2021). In this case, the primary structure of alginate becomes a determining factor for its drug release capability. In phosphate buffer (pH 7.2), the release of chlorpheniramine maleate and metronidazole is greater in M-rich alginate tablets. In contrast, G-rich alginates present a higher drug release rate in acidic media (Sriamornsak et al., 2007).
These studies reinforce the need to obtain high-quality alginates (with a G/M ratio greater than 2.3), with a well-defined chemical composition and a low presence of contaminants, for their application in pharmaceutical formulations or the manufacture of biomedical devices. The requirements of these industries cannot be met using algal alginate as a raw material, as its chemical features are commonly negatively affected during the separation and purification processes (Bojorges et al., 2023). Hence, other alternatives, such as bacterial production and enzyme-driven structural modification, have emerged as opportunities to obtain tailor-made alginates (Moradali et al., 2018). As both alternatives present advantages in obtaining high-end alginates, these processes participate in different stages during alginate production and should be assessed separately.
Biosynthesis of alginate
The alginate biosynthesis process has been studied in Azotobacter sp. and Pseudomonas sp. (Núñez et al., 2022; Urtuvia et al., 2017; Hay et al., 2013; Franklin et al., 2011). The genes involved in biosynthesis are very similar, but some differ in their regulatory mechanisms. These genes are located in an operon that has been previously described: algD, alg8, alg44, algK, algJ, algG, algX, algL, algI, algV, algF and algA (Pacheco-Leyva et al., 2016; Hay et al., 2013; Chitnis and Ohman, 1993) (Table 2). A schematic representation of the biosynthesis of alginate synthesized by A. vinelandii is shown in Figure 2, integrating recent findings on this biosynthetic pathway.
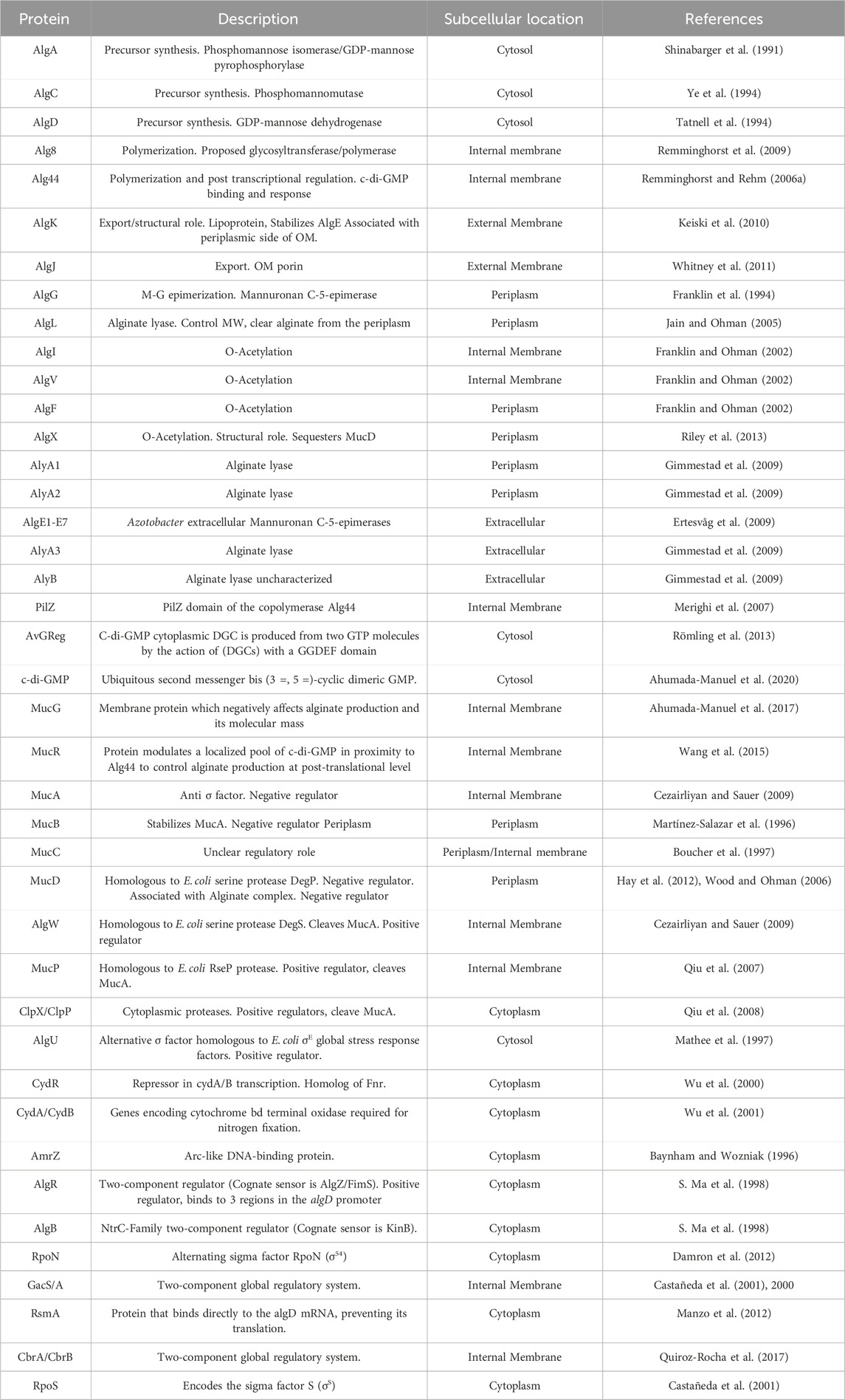
Table 2. Proteins involved in alginate biosynthesis and regulation.
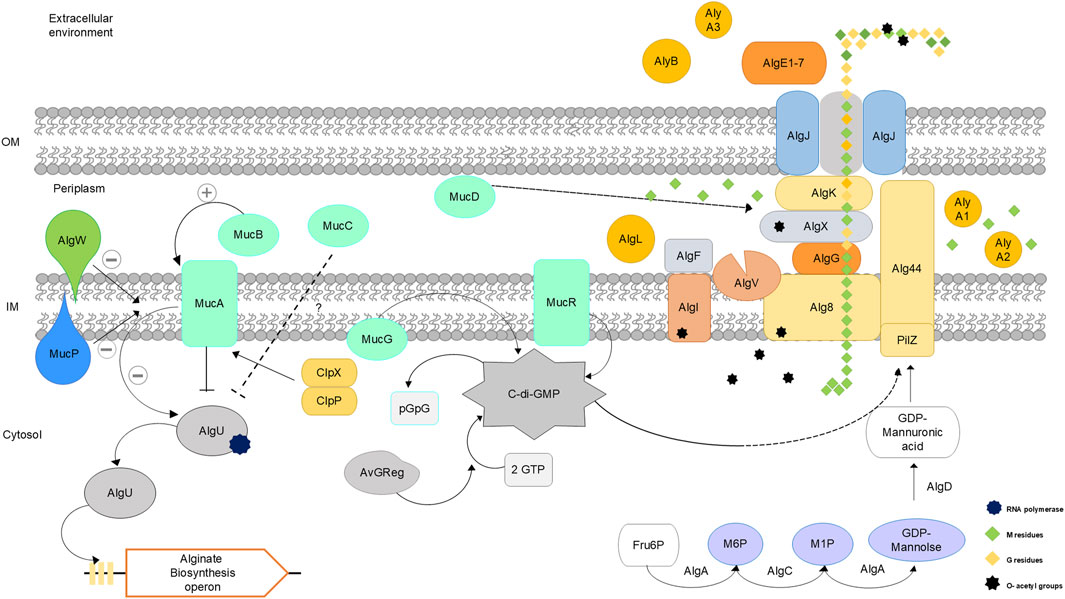
Figure 2. Schematic representation of alginate biosynthesis in Azotobacter vinelandii. Adapted from (Ahumada-Manuel et al., 2020; Urtuvia et al., 2017; Pacheco-Leyva et al., 2016; Ertesvåg, 2015; Flores et al., 2015; Hay et al., 2013).
Table 2 shows the proteins involved in alginate biosynthesis. Alginate biosynthesis can be divided into four steps: precursor synthesis, polymerization, modification, and secretion of alginate.
Alginate precursor synthesis
The first step in the synthesis of alginate is the generation of the monomer mannose-6-phosphate (M6P). This step begins with the import of hexoses from the medium to the cytosol. Glucose undergoes enzymatic reactions via the Entner–Doudoroff pathway until the formation of glyceraldehyde-3-phosphate/di-hidroxyacetone 3-phosphate by the enzyme glyceraldehyde-3-phosphate dehydrogenase; these molecules are phosphorylated and then reconverted into fructose 6-phosphate (F6P). Through the coordinated action of three enzymes located in the cytosol, AlgA, AlgC and AlgD, four reactions are catalyzed, allowing the synthesis of the precursor GDP-mannuronic acid (Shinabarger et al., 1991; Ye et al., 1994; Tatnell et al., 1994). In this way, F6P is converted to M6P by the action of phosphomannose isomerase (AlgA, a bifunctional enzyme) and then converted to mannose-1-phosphate (M1P), which is catalyzed by phosphomannomutase (AlgC), followed by conversion to GDP-mannose, which is again catalyzed by AlgA (Zielinski et al., 1992). The last step is the conversion to GDP-mannuronic acid catalyzed by GDP-mannose dehydrogenase (AlgD), which is the irreversible step in the formation of the alginate precursor and involves the use of carbon atoms toward the formation of the polymeric chain (Hay et al., 2013; Gaona et al., 2004; Tavares et al., 1999).
Polymerization
GDP-mannuronic acid is polymerized through two enzymes that are located inside the internal membrane (IM), glycosyltransferase/polymerase (Alg8) and copolymerase (Alg44), both of which are indispensable for the production of alginate (Urtuvia et al., 2017; Moradali et al., 2015; Hay et al., 2013; Oglesby et al., 2008; Galindo et al., 2007; Mejı́a-Ruı́z et al., 1997). The process of polymerization and transit of the newborn alginate chain through the membrane has been partially described through studies in both Pseudomonas aeruginosa and A. vinelandii (Remminghorst et al., 2009; Oglesby et al., 2008; Remminghorst and Rehm, 2006b). The polymerization of alginate is regulated at the posttranslational level through the interaction of the PilZ domain of Alg44, which binds the secondary messenger bis-(3′-5′)-cyclic dimeric guanosine monophosphate (c-di-GMP), and allows the interaction with other proteins that are involved in the next steps of alginate polymerization and transport through the membrane (Ahumada-Manuel et al., 2020).
C-di-GMP is a global regulatory molecule involved in several cellular processes, such as motility, exopolysaccharide production and biofilm formation (Ahumada-Manuel et al., 2020; 2017; Römling et al., 2013). Intracellular levels of c-di-GMP are controlled by the activities of diguanylate cyclase with a conserved domain containing the GGDEF motif and by phosphodiesterases (PDEs) bearing the EAL or HD-GYP motif. Proteins involved in signal transduction cascades have c-di-GMP-binding domains, which include PilZ (Römling et al., 2013; Merighi et al., 2007).
Modification, transport and secretion
During polymerization, the newborn chain of alginate is synthesized as a polymannuronic acid polysaccharide, which is translocated through the periplasm by the activity of some periplasmic proteins, such as AlgX, AlgG and AlgK (Riley et al., 2013; Keiski et al., 2010; Robles-Price et al., 2004; Jain et al., 2003; Franklin et al., 1994). During translocation, the poly-M alginate chain interacts with the protein complex composed of AlgI, AlgF, AlgV and AlgX, which presents acetyltransferase activity (Franklin and Ohman, 2002). This complex is capable of transferring an acetyl group from acetyl-CoA, which acts as a donor group, to the poly-M chain, which is the first event of chemical modification of the alginate chain, which is made on the C-2 and/or C-3 of the M-residues. AlgG has been identified as a protein with mannuronnan C-5 epimerase activity. Nevertheless, its activity is deficient in A. vinelandii, and its presence has a structural role in the alginate biosynthesis complex (Núñez et al., 2022; Ertesvåg, 2015). Therefore, the main event for the addition of G-residues into the alginate chain occurs after the secretion of the polymer into the extracellular medium, where the activity of seven different epimerases (AlgE1–7) in A. vinelandii cells can catalyze the epimerization of nonacetylated M-residues to G-residues (Stanisci et al., 2020).
AlgL is an alginate degradation protein with lyase activity that is encoded within the biosynthesis operon located in the periplasm. The role of AlgL is relevant for the viability of mucoid strains, exercising a maintenance function by degrading the alginate trapped in the periplasm (Jain and Ohman, 2005). AlgL can also actively control the polymer length and contribute to periplasmic translocation (Albrecht and Schiller, 2005; Bakkevig et al., 2005).
AlgK, together with AlgX, are structural proteins relevant for the process of alginate translocation through the periplasm; these proteins act as scaffolds for the assembly of functional alginate biosynthesis machinery (Keiski et al., 2010). In this context, AlgK (a lipoprotein) acts as an anchor for the protein complex in both the internal and external membranes.
The alginate chain, while transported through the periplasmic space, may undergo O-acetylation by the addition of hydroxyl O-2 and O-3 acetyl groups by four essential enzymes, AlgI, AlgV, AlgF, and AlgX, which can significantly alter the material properties of the resulting alginate (Fabich et al., 2012; Donati et al., 2009; Nivens et al., 2001). In addition, the O-acetylation of alginate is exclusive to bacterial alginates, which significantly increases the water retention capacity of alginate and the rigidity and structural homogeneity of alginate hydrogels, thus promoting the use of bacterial alginate as a biomaterial (Fabich et al., 2012; Nivens et al., 2001).
AlgI and AlgV are found in the IM of A. vinelandii cells, whereas AlgF and AlgX are found in the periplasm (Ertesvåg et al., 2017; Franklin and Ohman, 1996). AlgI transports acetyl-CoA groups from the cytoplasm to the periplasmic space where acetylation takes place. Acetyl groups are transferred by the actions of AlgF, AlgX, and AlgV to acetylate the M subunits in the alginate chain.
AlgG is responsible for the epimerization of nonacetylated M-residues in P. aeruginosa to convert them into G-residues, leading to the formation of alginate poly-MG (Maleki et al., 2017; Wolfram et al., 2014). In A. vinelandii, AlgG shares 60% structural homology with AlgG from P. aeruginosa and is essential in the alginate biosynthetic complex, although the main incorporation of G-residues in the polymer is made by soluble epimerases at the extracellular level (Nuñez et al., 2022).
Azotobacter vinelandii encodes six alginate lyases: alginate lyase (AlgL); mannuronan C-5 extracellular bifunctional epimerase and alginate lyase AlgE7 (which degrades the G-MM and G-GM bonds); three AlyA lyases, AlyA1, AlyA2 and AlyA3 (GM- and MG-cleavage); and an exolyase, AlyB (not characterized) (Ertesvåg, 2015; Setubal et al., 2009; Gimmestad et al., 2009; Ertesvåg et al., 2009). AlyA1 and AlyA3 are known to belong to the PL7 family of alginate lyases. AlyA1 and AlyA2 are related to growth.
AlgE1–7 are Ca2+-dependent enzymes that possess a regulatory domain (R-domain) sensitive to Ca2+, which regulates the activity of these enzymes through substrate affinity and catalytic modulation. These enzymes play different roles in A. vinelandii, as they are expressed under different conditions and have different catalytic activities in terms of the incorporation of diverse monomeric patterns through the alginate chain. For example, AlgE3 and AlgE1 can generate long MG patterns in the alginate chain, and AlgE6 can introduce long G-blocks; moreover, AlgE4 includes only individual G-residues, which promotes the generation of more flexible hydrogels because of the presence of short MG-blocks and GG-blocks. In terms of its physiological relevance, AlgE7 functions during cell germination from a cyst-dormant state because of its bifunctional activity (epimerase and lyase); AlgE3 and AlgE1 are related to vegetative growth, and AlgE2 is expressed during cyst differentiation (Moreno et al., 2018; Ertesvåg, 2015; Høidal et al., 2000).
In the last stage, after the polymeric chain is translocated through the periplasmic space, the porin protein AlgJ acts as a channel that allows the secretion of the alginate chain to the extracellular environment (Whitney et al., 2015; Baker et al., 2014; Hay et al., 2010).
Regulation related to alginate biosynthesis and modification
The regulation of alginate biosynthesis is a well-conserved process in both A. vinelandii and P. aeruginosa and is also highly regulated because of the energy input and carbon flux required for its production, secretion and modification (Inomura et al., 2017; Pacheco-Leyva et al., 2016; Castillo et al., 2013). In A. vinelandii, the genes associated with alginate biosynthesis are clustered together in an operon away from the cluster of genes related to epimerization. Other biosynthetic genes, such as algC, or depolymerases, such as AlyA1, AlyA2, and AlyA3, can be found in different locations. The overall regulatory events related to alginate biosynthesis can be separated into three levels: transcriptional, posttranscriptional, and posttranslational (Núñez et al., 2022). In the case of alginate chemical modification, most of the regulation occurs at the transcriptional and posttranslational levels through events controlled by the ubiquitous secondary messenger c-di-GMP and by the action of global genetic regulators such as AlgR and RpoS (Ma et al., 1998; Mathee et al., 1997).
A schematic representation of the regulation of alginate biosynthesis is shown in Figure 3.
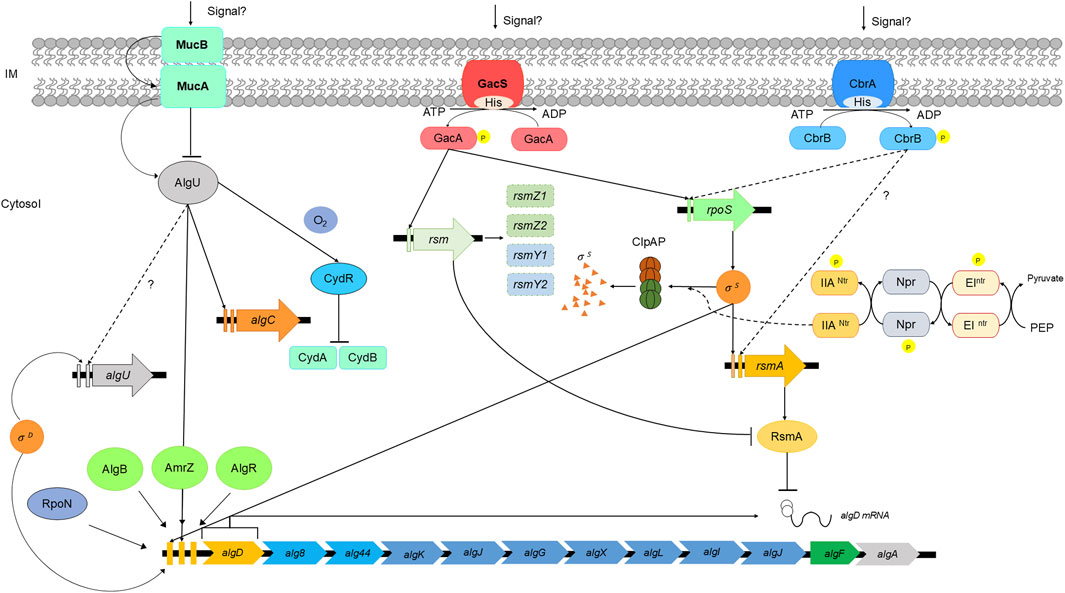
Figure 3. Schematic representation of alginate regulation in Azotobacter vinelandii. Adapted from (López-Pliego et al., 2020; 2018; Quiroz-Rocha et al., 2017; Romero et al., 2016; Pacheco-Leyva et al., 2016; Damron and Goldberg, 2012; Galindo et al., 2007; Castañeda et al., 2001).
The transcriptional regulation of alginate biosynthesis begins with the gene algD, which encodes the enzyme that mediates the carbon flux from sugars into the alginate precursor GDP-mannuronic acid. Three promoters have been found to exert positive effects on algD expression; two of these promoters are the sigma factors RpoS and AlgU, which also activate algC expression, and the third promoter activation factor (P3algD) remains unknown (Nuñez et al., 2022). In A. vinelandii, the sigma factor RpoS leads to a general response against oxidative stress and nutrient starvation and is essential during cell differentiation to the cyst dormant state. Moreover, the RpoE-like sigma factor AlgU also participates in the response against oxidative stress and is critical for alginate production during vegetative growth and encystment (Chowdhury-Paul et al., 2023; Martínez-Salazar et al., 1996). MucA is an antisigma factor that sequesters AlgU, preventing its action as a promoter of algD expression. Interestingly, AlgU and MucA, along with other antisigma factors, such as MucB, MucC, and MucD, are encoded within the same operon, suggesting a possible role in alginate biosynthesis regulation. Nevertheless, the regulatory role of MucC and MucD has not been elucidated to date (Núñez et al., 2022; Hay et al., 2012; Cezairliyan and Sauer, 2009; Wood and Ohman, 2006; Boucher et al., 1997; Martínez-Salazar et al., 1996). RpoS action is controlled by the response regulators RssB/RssC, which promote RpoS degradation through ClpXP during vegetative growth, acting as a repressor of the RpoS-related phenotype until the stationary phase (Rodríguez-Martínez et al., 2023).
After transcription, algD mRNA is regulated at the posttranscriptional level by the protein RsmA (CsrA). Rsm systems are RNA-binding proteins that act as posttranscriptional regulators belonging to the RsmA/CsrA family. In A. vinelandii, RsmA binds to the ribosome-binding site of algD mRNA, preventing its translation and thus negatively affecting alginate production. The expression of algD can be restored by the expression of small regulatory RNAs (rsmZ1–8 and rsmY), and these sRNAs saturate the RsmA-binding site, allowing algD mRNA translation. The two-component system (TCS) GacA/GacS oversees the expression of rsmZ1–7 and rsmY, and knocking out gacA or gacS prevents algD expression through rsmA action, annulling alginate biosynthesis (Rosales-Cruz et al., 2024; López-Pliego et al., 2020). Another TCS involved in alginate biosynthesis is the histidine kinase CbrA and its response regulator CbrB; these TCSs, together with the carbon catabolite repression system (Crc), control carbon flux in A. vinelandii and Pseudomonas aeruginosa. CbrA/CbrB mutants in the A. vinelandii wild-type strain present an alginate-overproducing phenotype. Other studies have shown that this TCS is required for high expression of rsmA and rpoS in A. vinelandii (Quiroz-Rocha et al., 2017; Manzo et al., 2012; Castañeda et al., 2000; 2001).
C-di-GMP is a universal secondary messenger in bacteria that controls many physiological processes, such as motility, pathogenesis, cellular differentiation, biofilm formation, and exopolysaccharide production (Valentini and Filloux, 2016). In A. vinelandii (as in P. aeruginosa), alginate polymerization through the polymerase complex Alg8/Alg44 is dependent on the presence of c-di-GMP through binding to the PilZ domain of Alg44. In the absence of this secondary messenger, alginate production is abolished; otherwise, its accumulation overstimulates alginate production (Merighi et al., 2007). The c-di-GMP pool is regulated through synthesis/degradation by specific enzymes. Diguanylate cyclases (DGCs) catalyze c-di-GMP production by the union of two GTP molecules at its active site, which is characterized by its GGDEF domain; commonly, DGCs contain an allosteric inhibition site that recognizes c-di-GMP; thus, these enzymes are inhibited by its product. Degradation of c-di-GMP occurs by the action of phosphodiesterases (PDEs) at specific EAL or HD-GYP domains, which catalyze the hydrolysis of this secondary messenger (Liang et al., 2022; Moradali and Rehm, 2020; Whitney et al., 2015). In the context of posttranslational regulation of alginate biosynthesis in A. vinelandii, AvGReg (DGC), MucR (DGC), and MucG (DGC-PDE) are the principal enzymes that control the c-di-GMP pool and thus promote or curb alginate production.
AvGReg is a globin-coupled sensor with DGC activity that directly binds oxygen and is considered the main contributor to the synthesis of c-di-GMP for alginate production. MucG can synthesize or degrade c-di-GMP due to the presence of both GGDEF and EAL domains (Ahumada-Manuel et al., 2017; Römling et al., 2013). Nevertheless, its PDE activity is the only one related to the inhibition of alginate production, as ΔMucG mutants can overproduce this polymer. Notably, MucG has a PAS domain that binds flavin adenine dinucleotide (FAD) cofactor, acting as a redox status sensor. This evidence suggests a relationship between respiratory metabolism, the redox status of the cell, and alginate production in A. vinelandii, as it has been shown that mutant strains affected by components of the respiratory chain show increased alginate production (Núñez et al., 2022; Ahumada-Manuel et al., 2020; Germani et al., 2020; Ahumada-Manuel et al., 2017). Interestingly, the PDE MucR is not essential for alginate polymerization, but it is essential during the encystment process, as ΔMucR mutants are unable to resist desiccation due to the lack of a well-assembled alginate capsule (Martínez-Ortiz et al., 2020).
Azotobacter vinelandii cysts are composed of two alginate layers named the intine and exine, and the guluronic acid contents of these two layers differ. The exine presents a higher G/M ratio than the intine does; consequently, the exine presents a rigid and structured shape in contrast to the intine, which presents a more amorphous structural characteristic (Martínez-Ortiz et al., 2020). In addition, Martínez-Ortiz et al. (2020) highlighted that MucR positively regulates the transcription of algE1–6 genes through c-di-GMP signaling, probably through the transcriptional regulator FleQ, which presents binding sites in the regulatory region of the algE1–7 genes. Additionally, MucR expression is regulated through AlgR, thus also being relevant in regulating alginate chemical modification, as ΔAlgR strains impair proper cyst coat organization. Moreover, c-di-GMP signaling also contributes to determining the molecular weight of the alginate chain, as an increase in the intracellular c-di-GMP pool promotes the production of alginates with higher mean molecular weights (MMWs), and AvGReg and MucG regulate this phenotype.
Epimerization of the alginate chain occurs at the extracellular level due to the action of the alginate epimerases AlgE1–7, which incorporate different patterns of MG- or GG-blocks of varying lengths depending on the individual activity of each epimerase. For example, AlgE4 modification incorporates long MG blocks through the alginate chain, whereas AlgE7 promotes long GG-blocks in the alginate chain (Petersen et al., 2023). The secretion of these epimerases requires the expression of the type 1 transport system eexDEF, which facilitates the transport of these enzymes through the membrane (Gimmestad et al., 2006).
The EexDEF transport system is encoded by an operon, and two promoters control its expression. P2eexD is RpoS-dependent and is related to cyst differentiation, as RpoS also positively controls the expression of AlgE1–7 genes, as AeRpoS (ΔRpoS-modified strain) can produce an alginate coat that is unable to resist desiccation, probably because of the lack of an organized structure capable of protecting dormant cells. P1eexD is another promoter region that is an unknown activator. The guluronic content in alginate can be altered through changes in the oxygen concentration in the culture (Sabra et al., 2000), and P1eexD is likely involved as a key control point, as it is necessary for epimerase excretion under other conditions not related to RpoS control (Moreno et al., 2018; Cocotl-Yañez et al., 2014).
Oxygen transfer in alginate production
Oxygen plays a key role in the growth, maintenance and production of metabolites, such as during the synthesis of alginate (Flores et al., 2015). Studies have shown that alginate production by A. vinelandii is influenced by factors such as dissolved oxygen tension (DOT), agitation rate, aeration, and OTR (Ponce et al., 2021; Díaz-Barrera et al., 2021; Castillo et al., 2020; Díaz-Barrera et al., 2017; Flores et al., 2015; Díaz-Barrera et al., 2014; 2007).
Bacterial alginate production is a highly energy-demanding process that requires a high oxygen uptake rate (OUR) to synthesize this polymer (Castillo et al., 2020; Lozano et al., 2011). Azotobacter vinelandii has a high respiration rate (Oelze, 2000), which generates oxygen-limiting conditions during its growth (Ponce et al., 2021; Díaz-Barrera et al., 2021; 2014). Under these conditions, the dissolved oxygen content is almost zero (dO2/dt = 0), and according to the oxygen balance, the OTR is equal to the OUR, which is also known as microaerophilic conditions. Respiration can be assessed by measuring OTR, which remains constant during growth (Ponce et al., 2021; Díaz-Barrera et al., 2021; 2014; Garcia-Ochoa et al., 2010). These parameters are related to alginate production, alginate molecular weight, and the expression of biosynthetic genes (Díaz-Barrera et al., 2012).
In A. vinelandii cultures, a relatively high oxygen transfer rate (OTR) prevents the generation of high MMW alginates, as a decrease in the OTR promotes alginate polymerization through increased c-di-GMP accumulation (Ahumada-Manuel et al., 2020). Oxygen-sensitive enzymes can regulate c-di-GMP: AvGReg promotes c-di-GMP accumulation through DGC activity in the presence of the intracellular binding of O2, and MucG reduces the c-di-GMP pool through PDE activity, which is regulated through the binding of FAD as a redox state sensor (Noar and Bruno-Bárcena, 2018; Ahumada-Manuel et al., 2017; Peña C et al., 2011).
One way to evaluate the role of oxygen in alginate production is to assess the effect of OTR on the maximum alginate concentration. Figure 4 shows a collection of scientific papers (n = 16) reporting data on OTR and alginate production under different culture conditions: batch cultures in flasks and bioreactors and continuous cultures.
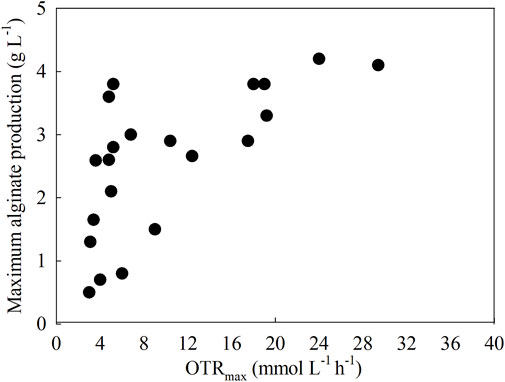
Figure 4. Relationship between the OTR and the alginate production in A. vinelandii under different culture conditions: batch cultures in flasks and bioreactors and continuous cultures.
The OTRmax is the highest rate at which oxygen is transferred from the gaseous phase to the liquid phase in a bioreactor and is limited by the aeration capacity and solubility of oxygen in the medium. It is a key parameter in aerobic culture systems to ensure an adequate oxygen supply (Maier and Büchs, 2001). The increase in OTRmax may be related to the increase in alginate concentration. At OTRmax values between 3 and 16 (mmol L−1 h−1), the value of the maximum alginate concentration increases from 0.8 to 3 (g L−1). Conversely, an increase in OTRmax from 16 to 30 (mmol L−1 h−1) does not significantly increase maximum alginate production. Two conclusions emerge from this relationship: an increase in OTRmax can modulate the maximum alginate production, which would help to generate defined culture strategies to produce and increase alginate production using a given OTRmax. However, it could be argued that there is a limit at which OTRmax increases alginate production, so increasing OTR beyond the limit proposed in this review to improve alginate production is not an appropriate strategy.
Effect of the oxygen transfer rate on the alginate molecular weight, degree of acetylation and G/M ratio
Several studies have measured OTR during A. vinelandii fermentation in flasks and bioreactors. These studies suggest that the OTR is related to the molecular weight of alginate. Some reports indicate that the highest molecular weight of alginate is obtained in batch cultures performed at a low OTR. However, as the OTR increases in cultures, the exact effect of OTR on the molecular weight of the alginate chain is still unknown.
Figure 5 shows the relationship between OTRmax and the maximum alginate MMW. This figure shows a compilation of different studies (n = 16), establishing a relationship between OTR and different culture strategies and how this respiration parameter is related to alginate MMW in different A. vinelandii strains. All molecular weight data compiled in this analysis were determined using gel permeation chromatography (GPC) coupled to HPLC, ensuring consistency and comparability across studies (Díaz-Barrera et al., 2021; García et al., 2020; Díaz-Barrera et al., 2007).
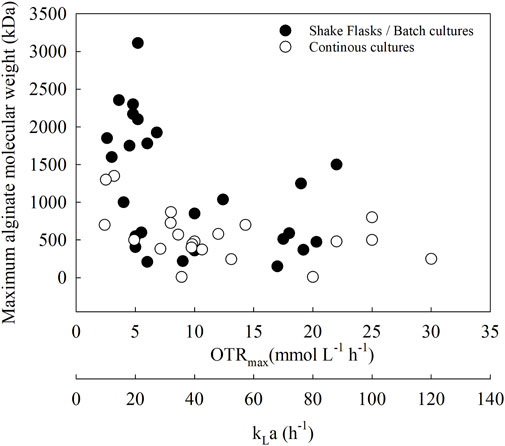
Figure 5. Relationship between the OTR and the alginate molecular weight in A. Vinelandii under different culture conditions: batch cultures in flasks and bioreactors and continuous cultures.
At a low OTRmax (less than 10 mmol L−1 h−1), alginate greater than 1,000 kDa can be obtained in flasks, batch cultures (black circles), and continuous cultures (white circles). Conversely, in cultures with OTRmax values greater than 10 mmol L−1 h−1, alginate MMW does not change markedly and appears to be independent of OTR in all culture modalities.
Another relationship that has been established is the volumetric mass transfer coefficient (kLa) versus the maximum alginate MMW. Typically, kLa is a key parameter used for the optimization of oxygen availability in aerobic cultures and is also a parameter for scaling up to bioreactors (Garcia-Ochoa and Gomez, 2009).
In A. vinelandii, the high respiratory rate limits intracellular oxygen availability, thereby altering the OTRmax value. This, in turn, can modify the kLa of the culture, leading to changes in both alginate concentration and its mean molecular weight (MMW). Therefore, by adjusting kLa, it may be possible to control alginate size across different culture scales. Although no studies have yet used kLa as a scaling parameter to reproduce alginate MMW, some previous reports have managed to obtain similar alginate production at different scales using the OTR parameter in the culture, which is determined by kLa (Ponce et al., 2021; Díaz-Barrera et al., 2021; 2014). All this would help to produce a customized polymer according to the required application.
To understand the relationship between OTR and alginate production at the metabolic level, modified strains of A. vinelandii have been developed. A study carried out by García et al. (2020) revealed that the mutant strain AT9, with a truncated copy of the mucG gene (a negative regulator of alginate biosynthesis), produced alginates with greater MMWs (3,312 ± 150 kDa) than its parental strain ATCC9046 (2,170 ± 260 kDa) under microaerophilic conditions. Although both strains presented similar OTRs, the absence of MucG favored the formation of longer alginate chains, suggesting that PDE activity could regulate alginate polymerization in response to oxygen availability or the cellular respiratory status (García et al., 2020).
Other authors have highlighted the role of the secondary messenger c-di-GMP in regulating the production of exopolysaccharides in many bacteria. Recent studies have shown that the enzymes MucG, MucR and AvGReg control the biosynthesis and modification of alginate by affecting the levels of c-di-GMP, which is essential for activating the polymerase through the PilZ domain of Alg44. Furthermore, the strain A. vinelandii ATCC9046 presented greater accumulation of c-di-GMP, which is associated with the overproduction of alginate of higher molecular weight, regardless of changes in the sequence or expression of MucG (Moradali and Rehm, 2020; Wang et al., 2015).
As previously mentioned, c-di-GMP signaling is part of a control modulus that affects alginate production and its chemical characteristics in A. vinelandii cultures, and this signaling modulus can respond to changes in OTR. Figure 4 shows a plateau type of behavior that reaches its highest point at an OTR of approximately 16 (mmol L−1 h−1). These results could be attributed to the fact that c-di-GMP signaling is involved in the production of alginate, as an increase in OTR reduces the levels of c-di-GMP and, therefore, decreases the concentration of alginate; however, there is still a need to elucidate c-di-GMP.
As the OTR increases from approximately 4 to 16 (mmol L−1 h−1), c-di-GMP levels could decrease to a concentration sufficient for the binding and activation of Alg44, and an increase in alginate production can be reached through a more efficient distribution of carbon for alginate biosynthesis.
As the OTR increases until it reaches 16 (mmol L−1 h−1) or higher, the respiratory protection complex starts to increase the consumption of reducing power, which increases carbon flux through the TCA cycle to maintain NADH consumption by uncoupled electron transport terminals; consequently, more carbon is distributed into CO2 production over alginate. Finally, the alginate biosynthetic pathway could maintain this constant level of production, as it allows the production of two NADH molecules for every mannuronic acid molecule synthesized, which could help to maintain the redox balance of A. vinelandii under high-aeration conditions (García et al., 2018; Castillo et al., 2013).
In the opposite direction, as OTRmax is reduced, c-di-GMP levels increase, leading to the generation of alginates with higher MMW (Ahumada-Manuel et al., 2020). In agreement with these results, Figure 5 shows that longer alginate chains can be obtained in OTRmax ranges between 2 and 8 (mmol L−1 h−1), which could be related to control by this secondary messenger. Although a low OTRmax allows the generation of higher MMW alginates due to an increase in c-di-GMP levels, these conditions do not allow greater polymer production. An OTRmax of 5 (mmol L−1 h−1) is associated with an OTR of 1.5 (g L−1) of alginate, in contrast to conditions with higher oxygen availability, where the alginate concentration could increase to 4 (g L−1), with an OTRmax = 18 (mmol L−1 h−1).
As elevated c-di-GMP levels can alter many cellular processes, such as motility, differentiation, and polymer secretion, in this case, the reduced alginate production at low OTRmax can be explained through metabolic behavior. At low OTRmax, oxygen limitation in A. vinelandii cultures negatively impacts carbon source intake due to the absence of oxygen as a terminal electron acceptor. This ultimately reduces the overall capacity of various biosynthetic pathways, including alginate production (Castillo et al., 2020). Notably, the viscosity of the broth increases due to the increased growth and secretion of alginates with high MMW values (>1,000 kDa), which could also exacerbate diffusional limitations from the medium to the intracellular level, affecting cellular growth and thus productivity.
In this scenario, modified strains such as A. vinelandii AT9 (mucG::mTn5; SpI) are useful tools for obtaining long alginate chains under high-OTR conditions (>16 mmol L−1 h−1) because of the ability of this strain to maintain elevated c-di-GMP levels without being significantly affected by OTRmax. Finally, it becomes necessary to further investigate the relationship between the control modules of c-di-GMP at OTRmax values greater than 5 (mmol L−1 h−1), as previously reported by Ahumada-Manuel et al. (2020). To elucidate and validate these phenomena, alginate production and its modification are analyzed under conditions that favor these processes, such as non-diazotrophic cultures, alginate epimerization during vegetative growth, and the effects of global regulators on c-di-GMP levels, among others (Ahumada-Manuel et al., 2020).
In terms of the effect of oxygen on alginate production, oxygen influences the degree of alginate acetylation. The incorporation of acetyl groups confers amphiphilicity to alginate and improves the physical‒chemical properties of the formulated products (Tao et al., 2024; Wang et al., 2023).
Table 3 shows the acetylation degree under different conditions of cultivation (Medina et al., 2023; Jiménez et al., 2016; Castillo et al., 2013; Gaytán et al., 2012; Peña et al., 2006; Clementi et al., 1999; Peña et al., 1997). García et al. (2020) show that by using a mutant strain of A. vinelandii AT9 (mucG::miniTn5), in a bioreactor with sucrose as a carbon source and yeast extract as a nitrogen source, an increase in the OTRmax in the range of 3.6 to – 12.4 (mmol L−1 h−1) promoted cell growth and alginate production, generating an increase in the degree of acetylation (0.56%–2.13%) and, therefore, an increase in the viscosifying power. Castillo et al. (2013) evaluated the effects of DOT and the specific growth rate (μ) on the degree of acetylation in continuous cultures of A. vinelandii ATCC 9046, which were limited by glucose. They reported that high DOT (9%) resulted in a high degree of alginate acetylation (6.88%). In contrast, increasing μ had negative effects on polymer production and acetylation. At a high DOT content (9%) and low μ, there was a decrease in the respiration rate, suggesting that the flow of acetyl-CoA (the acetyl donor) was diverted toward alginate acetylation.
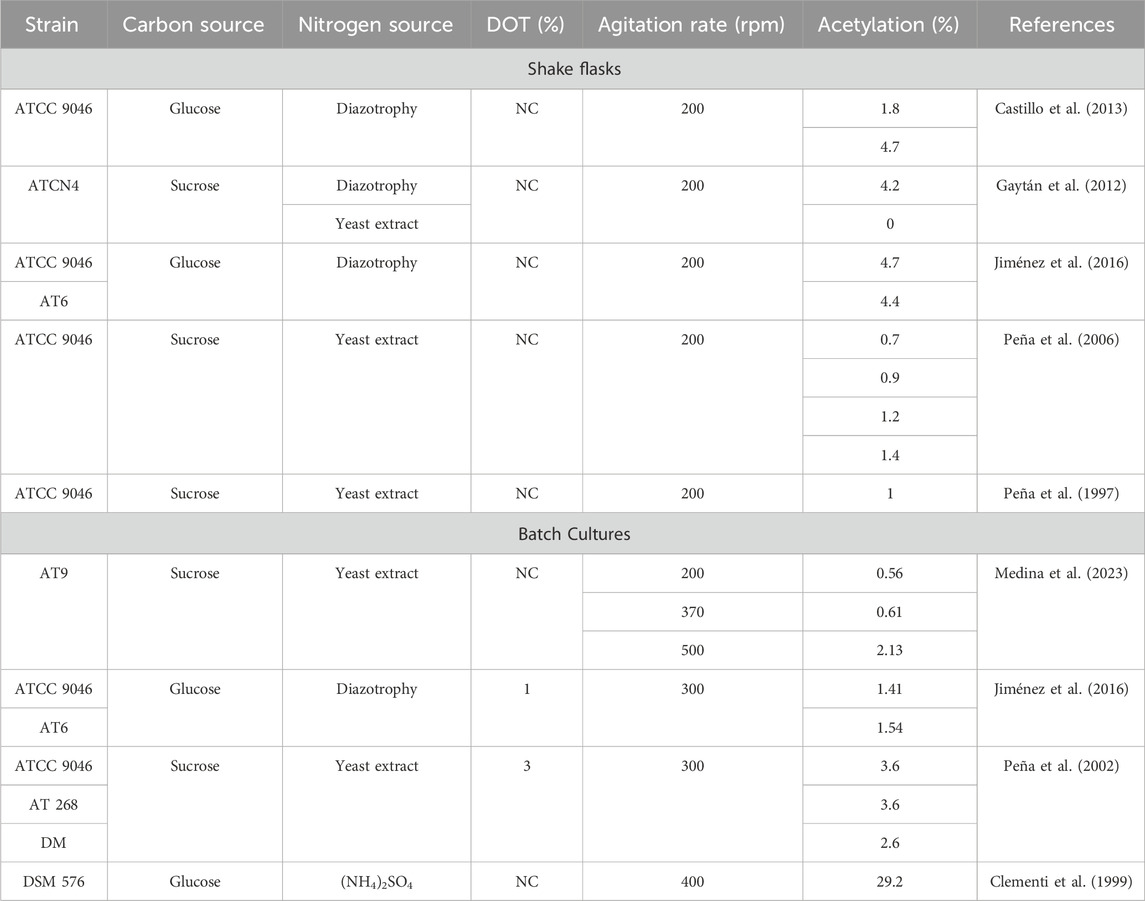
Table 3. Acetylation degree by Azotobacter vinelandii under different conditions of cultivation.
Another key parameter is the G/M ratio, which is crucial for defining the physical properties of these hydrogels (Díaz-Barrera et al., 2021). Table 4 shows various reports on the G/M ratio performed in flasks as well as in bioreactors using different types of A. vinelandii with different types of nitrogen sources (Díaz-Barrera et al., 2021; Gaytán et al., 2012; Kıvılcımdan Moral and Sanin, 2012; Peña et al., 2006; Clementi et al., 1999). Díaz-Barrera et al. (2021) evaluated the molecular weight and G/M ratio in batch cultures subjected to diazotrophic conditions at different OTRs. In this study, it was reported that the OTR affects the G/M ratio, as this ratio can be increased from 0.7 to 0.86 by increasing the OTRmax from 5 to 10.4 mmol L−1 h−1 through changes in the agitation rate. Gaytán et al. (2012) studied an alginate-overproducing strain called A. vinelandii ATCN4, which contains an insertion of the nqrE gene that encodes a subunit of the NADH-dependent sodium translocator complex: ubiquinone oxidoreductase (Na+-NQR). In this study, under nitrogen fixation conditions, this strain is capable of producing alginates with a high G/M ratio (close to 2); interestingly, when yeast extract was supplemented in the culture medium, this ratio increased to the highest G/M ratio (4.5) currently reported for alginates produced in A. vinelandii cultures.
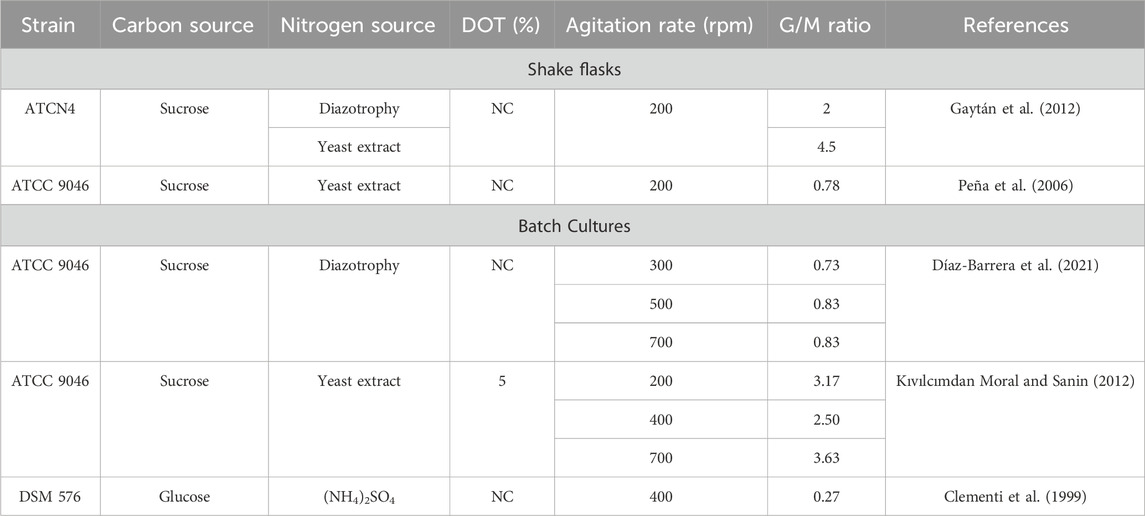
Table 4. G/M ratio by Azotobacter vinelandii under different conditions of cultivation.
Recently, interest in the use of alginate, which has a relatively high gelling capacity and rigidity, for applications in the biomedical and pharmaceutical industries has increased. Therefore, it is necessary to develop strategies to ensure the production of alginates with a high G/M ratio through the production of alginate-overproducing mutant strains and different culture methods.
Concluding remarks
This review highlights the importance of oxygen availability and its influence on intracellular c-di-GMP levels, regulating key enzymes that are involved in alginate polymerization and its modification. A high oxygen transfer rate (OTR) enhances the alginate concentration, whereas a lower OTR promotes the synthesis of alginate with a higher molecular weight and improved guluronic/mannuronic (G/M) ratios, which are critical for applications such as hydrogels and drug delivery systems. Future advancements should focus on engineering fermentation processes to fine-tune oxygen levels and use c-di-GMP signaling as a medication target because of its effect on the global regulation of alginate production and its chemical structure, which could favor the generation of alginates with better structural features. Genetic modifications targeting c-di-GMP metabolism could further enable precise control over alginate characteristics. By leveraging this regulatory pathway, it is possible to develop scalable and reproducible production systems that ensure consistency in biopolymer quality while maintaining economic and environmental sustainability.
Author contributions
BP: Formal Analysis, Writing – original draft, Investigation, Conceptualization, Validation, Visualization, Writing – review and editing. AZ-Q: Formal Analysis, Writing – review and editing, Writing – original draft, Investigation. EG: Writing – review and editing, Supervision. RA: Writing – review and editing, Supervision. ÁD-B: Writing – review and editing, Funding acquisition, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported and funded by the Agencia Nacional de Investigación y Desarrollo, Chile (ANID) through FONDECYT Regular project 1231075 (AD-B) and National Doctoral Scholarship N°21201148 (BP).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aguero, L., Alpdagtas, S., Ilhan, E., Zaldivar-Silva, D., and Gunduz, O. (2021). Functional role of crosslinking in alginate scaffold for drug delivery and tissue engineering: a review. Eur. Polym. J. 160, 110807. doi:10.1016/j.eurpolymj.2021.110807
Ahumada-Manuel, C. L., Guzmán, J., Peña, C., Quiroz-Rocha, E., Espín, G., and Núñez, C. (2017). The signaling protein MucG negatively affects the production and the molecular mass of alginate in Azotobacter vinelandii. Appl. Microbiol. Biotechnol. 101 (4), 1521–1534. doi:10.1007/s00253-016-7931-8
Ahumada-Manuel, C. L., Martínez-Ortiz, I. C., Hsueh, B. Y., Guzmán, J., Waters, C. M., Zamorano-Sánchez, D., et al. (2020). Increased c-di-GMP levels lead to the production of alginates of high molecular mass in Azotobacter vinelandii. J. Bacteriol. 202 (24), e00134-20. doi:10.1128/jb.00134-20
Albrecht, M. T., and Schiller, N. L. (2005). Alginate lyase (AlgL) activity is required for alginate biosynthesis in Pseudomonas aeruginosa. J. Bacteriol. 187 (11), 3869–3872. doi:10.1128/jb.187.11.3869-3872.2005
Baker, P., Ricer, T., Moynihan, P. J., Kitova, E. N., Walvoort, M. T. C., Little, D. J., et al. (2014). P. aeruginosa SGNH hydrolase-like proteins AlgJ and AlgX have similar topology but separate and distinct roles in alginate acetylation. PLOS Pathog. 10 (8), e1004334. doi:10.1371/journal.ppat.1004334
Bakkevig, K., Sletta, H., Gimmestad, M., Aune, R., Ertesvåg, H., Degnes, K., et al. (2005). Role of the Pseudomonas fluorescens alginate lyase (AlgL) in clearing the periplasm of alginates not exported to the extracellular environment. J. Bacteriol. 187 (24), 8375–8384. doi:10.1128/jb.187.24.8375-8384.2005
Baldassarre, M. E., Di Mauro, A., Pignatelli, M. C., Fanelli, M., Salvatore, S., Di Nardo, G., et al. (2020). Magnesium alginate in gastro-esophageal reflux: a randomized multicenter cross-over study in infants. Int. J. Environ. Res. Public Health 17 (1), 83. doi:10.3390/ijerph17010083
Bales, P. M., Renke, E. M., May, S. L., Shen, Y., and Nelson, D. C. (2013). Purification and characterization of biofilm-associated EPS exopolysaccharides from ESKAPE organisms and other pathogens. PLoS One 8 (6), e67950. doi:10.1371/journal.pone.0067950
Baynham, P. J., and Wozniak, D. J. (1996). Identification and characterization of AlgZ, an AlgT-dependent DNA-binding protein required for Pseudomonas aeruginosa algD transcription. Mol. Microbiol. 22 (1), 97–108. doi:10.1111/j.1365-2958.1996.tb02659.x
Bojorges, H., López-Rubio, A., Martínez-Abad, A., and Fabra, M. J. (2023). Overview of alginate extraction processes: impact on alginate molecular structure and techno-functional properties. Trends Food Sci. Technol. 140, 104142. doi:10.1016/j.tifs.2023.104142
Bonartseva, G. A., Akulina, E. A., Myshkina, V. L., Voinova, V. V., Makhina, T. K., and Bonartsev, A. P. (2017). Alginate biosynthesis by Azotobacter bacteria. Appl. Biochem. Microbiol. 53 (1), 52–59. doi:10.1134/S0003683817010070
Boucher, J. C., Schurr, M. J., Yu, H., Rowen, D. W., and Deretic, V. (1997). Pseudomonas aeruginosa in cystic fibrosis: role of mucC in the regulation of alginate production and stress sensitivity. Microbiology 143 (11), 3473–3480. doi:10.1099/00221287-143-11-3473
Bularz, B., Fernández, M., Subida, M. D., Wieters, E. A., and Pérez-Matus, A. (2022). Effects of harvesting on subtidal kelp forests (Lessonia trabeculata) in central Chile. Ecosphere 13 (3), e3958. doi:10.1002/ecs2.3958
Campos, M., Martínez-Salazar, J. M., Lloret, L., Moreno, S., Núñez, C., Espín, G., et al. (1996). Characterization of the gene coding for GDP-mannose dehydrogenase (algD) from Azotobacter vinelandii. J. Bacteriol. 178 (7), 1793–1799. doi:10.1128/jb.178.7.1793-1799.1996
Castañeda, M., Guzmán, J., Moreno, S., and Espín, G. (2000). The GacS sensor kinase regulates alginate and poly-β-hydroxybutyrate production in Azotobacter vinelandii. J. Bacteriol. 182 (9), 2624–2628. doi:10.1128/jb.182.9.2624-2628.2000
Castañeda, M., Sánchez, J., Moreno, S., Núñez, C., and Espı́n, G. (2001). The global regulators GacA and ςSForm part of a Cascade that controls alginate production in Azotobacter vinelandii. J. Bacteriol. 183 (23), 6787–6793. doi:10.1128/jb.183.23.6787-6793.2001
Castillo, T., García, A., Padilla-Córdova, C., Díaz-Barrera, A., and Peña, C. (2020). Respiration in Azotobacter vinelandii and its relationship with the synthesis of biopolymers. Electron. J. Biotechnol. 48, 36–45. doi:10.1016/j.ejbt.2020.08.001
Castillo, T., Heinzle, E., Peifer, S., Schneider, K., and Peña M, C. F. (2013). Oxygen supply strongly influences metabolic fluxes, the production of poly(3-hydroxybutyrate) and alginate, and the degree of acetylation of alginate in Azotobacter vinelandii. Process Biochem. 48 (7), 995–1003. doi:10.1016/j.procbio.2013.04.014
Cezairliyan, B. O., and Sauer, R. T. (2009). Control of Pseudomonas aeruginosa AlgW protease cleavage of MucA by peptide signals and MucB. Mol. Microbiol. 72 (2), 368–379. doi:10.1111/j.1365-2958.2009.06654.x
Chen, J., Hu, Y., Zhang, L., Wang, Y., Wang, S., Zhang, Y., et al. (2017). Alginate oligosaccharide DP5 exhibits antitumor effects in osteosarcoma patients following surgery. Front. Pharmacol. 8, 623. doi:10.3389/fphar.2017.00623
Cheng, D., Jiang, C., Xu, J., Liu, Z., and Mao, X. (2020). Characteristics and applications of alginate lyases: a review. Int. J. Biol. Macromol. 164, 1304–1320. doi:10.1016/j.ijbiomac.2020.07.199
Chitnis, C. E., and Ohman, D. E. (1993). Genetic analysis of the alginate biosynthetic gene cluster of Pseudomonas aeruginosa shows evidence of an operonic structure. Mol. Microbiol. 8 (3), 583–590. doi:10.1111/j.1365-2958.1993.tb01602.x
Chowdhury-Paul, S., Martínez-Ortíz, I. C., Pando-Robles, V., Moreno, S., Espín, G., Merino, E., et al. (2023). The Azotobacter vinelandii AlgU regulon during vegetative growth and encysting conditions: a proteomic approach. PLoS One 18 (11), e0286440. doi:10.1371/journal.pone.0286440
Chuacharoen, T., Aroonsong, S., and Chysirichote, T. (2022). Alginate production of Azotobacter vinelandii using sugar cane juice as the main carbon source in an airlift bioreactor. Industrial Eng. Chem. Res. 61 (34), 12329–12337. doi:10.1021/acs.iecr.2c02693
Clementi, F., Crudele, M. A., Parente, E., Mancini, M., and Moresi, M. (1999). Production and characterisation of alginate from Azotobacter vinelandii. J. Sci. Food Agric. 79 (4), 602–610. doi:10.1002/(SICI)1097-0010(19990315)79:4<602::AID-JSFA224>3.0.CO;2-N
Cocotl-Yañez, M., Moreno, S., Encarnación, S., López-Pliego, L., Castañeda, M., and Espín, G. (2014). A small heat-shock protein (Hsp20) regulated by RpoS is essential for cyst desiccation resistance in Azotobacter vinelandii. Microbiology 160 (3), 479–487. doi:10.1099/mic.0.073353-0
Damron, F. H., and Goldberg, J. B. (2012). Proteolytic regulation of alginate overproduction in Pseudomonas aeruginosa. Mol. Microbiol. 84 (4), 595–607. doi:10.1111/j.1365-2958.2012.08049.x
Damron, F. H., Owings, J. P., Okkotsu, Y., Varga, J. J., Schurr, J. R., Goldberg, J. B., et al. (2012). Analysis of the Pseudomonas aeruginosa regulon controlled by the sensor kinase KinB and sigma factor RpoN. J. Bacteriol. 194 (6), 1317–1330. doi:10.1128/jb.06105-11
Díaz-Barrera, A., Gutierrez, J., Martínez, F., and Altamirano, C. (2014). Production of alginate by Azotobacter vinelandii grown at two bioreactor scales under oxygen-limited conditions. Bioprocess Biosyst. Eng. 37 (6), 1133–1140. doi:10.1007/s00449-013-1084-2
Díaz-Barrera, A., Maturana, N., Pacheco-Leyva, I., Martínez, I., and Altamirano, C. (2017). Different responses in the expression of alginases, alginate polymerase and acetylation genes during alginate production by Azotobacter vinelandii under oxygen-controlled conditions. J. Industrial Microbiol. Biotechnol. 44 (7), 1041–1051. doi:10.1007/s10295-017-1929-9
Díaz-Barrera, A., Peña, C., and Galindo, E. (2007). The oxygen transfer rate influences the molecular mass of the alginate produced by Azotobacter vinelandii. Appl. Microbiol. Biotechnol. 76 (4), 903–910. doi:10.1007/s00253-007-1060-3
Díaz-Barrera, A., Sanchez-Rosales, F., Padilla-Córdova, C., Andler, R., and Peña, C. (2021). Molecular weight and guluronic/mannuronic ratio of alginate produced by Azotobacter vinelandii at two bioreactor scales under diazotrophic conditions. Bioprocess Biosyst. Eng. 44 (6), 1275–1287. doi:10.1007/s00449-021-02532-8
Díaz-Barrera, A., Soto, E., and Altamirano, C. (2012). Alginate production and alg8 gene expression by Azotobacter vinelandii in continuous cultures. J. Industrial Microbiol. Biotechnol. 39 (4), 613–621. doi:10.1007/s10295-011-1055-z
Donati, I., Mørch, Y. A., Strand, B. L., Skjåk-Bræk, G., and Paoletti, S. (2009). Effect of elongation of alternating sequences on swelling behavior and large deformation properties of natural alginate gels. J. Phys. Chem. B 113 (39), 12916–12922. doi:10.1021/jp905488u
Dudun, A. A., Akoulina, E. A., Zhuikov, V. A., Makhina, T. K., Voinova, V. V., Belishev, N. V., et al. (2022). Competitive biosynthesis of bacterial alginate using Azotobacter vinelandii 12 for tissue engineering applications. Polymers 14 (1), 131. doi:10.3390/polym14010131
Dusseault, J., Tam, S. K., Ménard, M., Polizu, S., Jourdan, G., Yahia, L., et al. (2006). Evaluation of alginate purification methods: effect on polyphenol, endotoxin, and protein contamination. J. Biomed. Mater. Res. Part A 76A (2), 243–251. doi:10.1002/jbm.a.30541
Ertesvåg, H. (2015). Alginate-modifying enzymes: biological roles and biotechnological uses. Front. Microbiol. 6, 523. doi:10.3389/fmicb.2015.00523
Ertesvåg, H., Sletta, H., Senneset, M., Sun, Y.-Q., Klinkenberg, G., Konradsen, T. A., et al. (2017). Identification of genes affecting alginate biosynthesis in Pseudomonas fluorescens by screening a transposon insertion library. BMC Genomics 18 (1), 11. doi:10.1186/s12864-016-3467-7
Ertesvåg, H., Valla, S., and Skjåk-Bræk, G. (2009). “Enzymatic alginate modification,” in Alginates: Biology and Applications. Microbiology Monographs. Editor B. Rehm (Berlin, Heidelberg: Springer), 13, 95–115. doi:10.1007/978-3-540-92679-5_4
Fabich, H. T., Vogt, S. J., Sherick, M. L., Seymour, J. D., Brown, J. R., Franklin, M. J., et al. (2012). Microbial and algal alginate gelation characterized by magnetic resonance. J. Biotechnol. 161 (3), 320–327. doi:10.1016/j.jbiotec.2012.04.016
Flores, C., Díaz-Barrera, A., Martínez, F., Galindo, E., and Peña, C. (2015). Role of oxygen in the polymerization and de-polymerization of alginate produced by Azotobacter vinelandii. J. Chem. Technol. Biotechnol. 90 (3), 356–365. doi:10.1002/jctb.4548
Franklin, M. J., Chitnis, C. E., Gacesa, P., Sonesson, A., White, D. C., and Ohman, D. E. (1994). Pseudomonas aeruginosa AlgG is a polymer level alginate C5-mannuronan epimerase. J. Bacteriol. 176 (7), 1821–1830. doi:10.1128/jb.176.7.1821-1830.1994
Franklin, M. J., Nivens, D. E., Weadge, J. T., and Howell, P. L. (2011). Biosynthesis of the Pseudomonas aeruginosa extracellular polysaccharides, alginate, Pel, and psl. Front. Microbiol. 2, 167. doi:10.3389/fmicb.2011.00167
Franklin, M. J., and Ohman, D. E. (1996). Identification of algI and algJ in the Pseudomonas aeruginosa alginate biosynthetic gene cluster which are required for alginate O acetylation. J. Bacteriol. 178 (8), 2186–2195. doi:10.1128/jb.178.8.2186-2195.1996
Franklin, M. J., and Ohman, D. E. (2002). Mutant analysis and cellular localization of the AlgI, AlgJ, and AlgF proteins required for O acetylation of alginate in Pseudomonas aeruginosa. Journal of bacteriology. J. Bacteriol. 184 (11), 3000–3007. doi:10.1128/jb.184.11.3000-3007.2002
Freeman, F. E., and Kelly, D. J. (2017). Tuning alginate bioink stiffness and composition for controlled growth factor delivery and to spatially direct MSC fate within bioprinted tissues. Sci. Rep. 7 (1), 17042. doi:10.1038/s41598-017-17286-1
Galindo, E., Peña, C., Núñez, C., Segura, D., and Espín, G. (2007). Molecular and bioengineering strategies to improve alginate and polydydroxyalkanoate production by Azotobacter vinelandii. Microb. Cell Factories 6 (1), 7. doi:10.1186/1475-2859-6-7
Gaona, G., Núñez, C., Goldberg, J. B., Linford, A. S., Nájera, R., Castañeda, M., et al. (2004). Characterization of the Azotobacter vinelandii algC gene involved in alginate and lipopolysaccharide production. FEMS Microbiol. Lett. 238 (1), 199–206. doi:10.1111/j.1574-6968.2004.tb09756.x
García, A., Castillo, T., Ramos, D., Ahumada-Manuel, C. L., Núñez, C., Galindo, E., et al. (2020). Molecular weight and viscosifying power of alginates produced by mutant strains of Azotobacter vinelandii under microaerophilic conditions. Biotechnol. Rep. 26, e00436. doi:10.1016/j.btre.2020.e00436
García, A., Ferrer, P., Albiol, J., Castillo, T., Segura, D., and Peña, C. (2018). Metabolic flux analysis and the NAD(P)H/NAD(P)+ ratios in chemostat cultures of Azotobacter vinelandii. Microb. Cell Factories 17 (1), 10. doi:10.1186/s12934-018-0860-8
Garcia-Ochoa, F., and Gomez, E. (2009). Bioreactor scale-up and oxygen transfer rate in microbial processes: an overview. Biotechnol. Adv. 27 (2), 153–176. doi:10.1016/j.biotechadv.2008.10.006
Garcia-Ochoa, F., Gomez, E., Santos, V. E., and Merchuk, J. C. (2010). Oxygen uptake rate in microbial processes: an overview. Biochem. Eng. J. 49 (3), 289–307. doi:10.1016/j.bej.2010.01.011
Gawin, A., Tietze, L., Aarstad, O. A., Aachmann, F. L., Brautaset, T., and Ertesvåg, H. (2020). Functional characterization of three Azotobacter chroococcum alginate-modifying enzymes related to the Azotobacter vinelandii AlgE mannuronan C-5-epimerase family. Sci. Rep. 10 (1), 12470. doi:10.1038/s41598-020-68789-3
Gaytán, I., Peña, C., Núñez, C., Córdova, M. S., Espín, G., and Galindo, E. (2012). Azotobacter vinelandii lacking the Na+-NQR activity: a potential source for producing alginates with improved properties and at high yield. World J. Microbiol. Biotechnol. 28 (8), 2731–2740. doi:10.1007/s11274-012-1084-4
Germani, F., Nardini, M., De Schutter, A., Cuypers, B., Berghmans, H., Van Hauwaert, M.-L., et al. (2020). Structural and functional characterization of the globin-coupled sensors of Azotobacter vinelandii and Bordetella pertussis. Antioxidants Redox Signal. 32 (6), 378–395. doi:10.1089/ars.2018.7690
Gimmestad, M., Ertesvåg, H., Heggeset, T. M. B., Aarstad, O., Svanem, B. I. G., and Valla, S. (2009). Characterization of three new Azotobacter vinelandii alginate lyases, one of which is involved in cyst germination. J. Bacteriol. 191 (15), 4845–4853. doi:10.1128/jb.00455-09
Gimmestad, M., Steigedal, M., Ertesvåg, H., Moreno, S., Christensen, B. E., Espín, G., et al. (2006). Identification and characterization of an Azotobacter vinelandii type I secretion system responsible for export of the AlgE-Type mannuronan C-5-Epimerases. J. Bacteriol. 188 (15), 5551–5560. doi:10.1128/jb.00236-06
Hay, I. D., Rehman, Z. U., Moradali, M. F., Wang, Y., and Rhem, B. H. A. (2013). Microbial alginate production, modification and its applications. Microbial Biotechnology, 6 (6), 637–650. doi:10.1111/1751-7915.12076
Hay, I. D., Schmidt, O., Filitcheva, J., and Rehm, B. H. A. (2012). Identification of a periplasmic algk–algx–mucd multiprotein complex in Pseudomonas aeruginosa involved in biosynthesis and regulation of alginate. Appl. Microbiol. Biotechnol. 93 (1), 215–227. doi:10.1007/s00253-011-3430-0
Hay, I. D., Ur Rehman, Z., Ghafoor, A., and Rehm, B. H. A. (2010). Bacterial biosynthesis of alginates. J. Chem. Technol. Biotechnol. 85 (6), 752–759. doi:10.1002/jctb.2372
Hecht, H., and Srebnik, S. (2016). Structural characterization of sodium alginate and calcium alginate. Biomacromolecules 17 (6), 2160–2167. doi:10.1021/acs.biomac.6b00378
Hendawy, W. S., El Kanani, Y. M., El Batawy, O. I., and El Mongy, T. M. (2019). Optimization the alginate production conditions in sweet whey medium by Azotobacter chrococcum. Arab Univ. J. Agric. Sci. 27 (1), 479–489. doi:10.21608/ajs.2019.43586
Hoefer, D., Schnepf, J. K., Hammer, T. R., Fischer, M., and Marquardt, C. (2015). Biotechnologically produced microbial alginate dressings show enhanced gel forming capacity compared to commercial alginate dressings of marine origin. J. Mater. Sci. Mater. Med. 26 (4), 162. doi:10.1007/s10856-015-5492-5
Høidal, H. K., Svanem, B. I. G., Gimmestad, M., and Valla, S. (2000). Mannuronan C-5 epimerases and cellular differentiation of Azotobacter vinelandii. Environ. Microbiol. 2 (1), 27–38. doi:10.1046/j.1462-2920.2000.00074.x
Inomura, K., Bragg, J., and Follows, M. J. (2017). A quantitative analysis of the direct and indirect costs of nitrogen fixation: a model based on Azotobacter vinelandii. ISME J. 11 (1), 166–175. doi:10.1038/ismej.2016.97
Jain, S., Franklin, M. J., Ertesvåg, H., Valla, S., and Ohman, D. E. (2003). The dual roles of AlgG in C-5-epimerization and secretion of alginate polymers in Pseudomonas aeruginosa. Mol. Microbiol. 47 (4), 1123–1133. doi:10.1046/j.1365-2958.2003.03361.x
Jain, S., and Ohman, D. E. (2005). Role of an alginate lyase for alginate transport in mucoid Pseudomonas aeruginosa. Infect. Immun. 73 (10), 6429–6436. doi:10.1128/iai.73.10.6429-6436.2005
Jiménez, L., Castillo, T., Flores, C., Segura, D., Galindo, E., and Peña, C. (2016). Analysis of respiratory activity and carbon usage of a mutant of Azotobacter vinelandii impaired in poly-β-hydroxybutyrate synthesis. J. Industrial Microbiol. Biotechnol. 43 (8), 1167–1174. doi:10.1007/s10295-016-1774-2
Keiski, C.-L., Harwich, M., Jain, S., Neculai, A. M., Yip, P., Robinson, H., et al. (2010). AlgK is a TPR-containing protein and the periplasmic component of a novel exopolysaccharide secretin. Structure 18 (2), 265–273. doi:10.1016/j.str.2009.11.015
Kıvılcımdan Moral, Ç., and Sanin, F. D. (2012). An investigation of agitation speed as a factor affecting the quantity and monomer distribution of alginate from Azotobacter vinelandii ATCC® 9046. J. Industrial Microbiol. Biotechnol. 39 (3), 513–519. doi:10.1007/s10295-011-1043-3
Langlois, G., Dusseault, J., Bilodeau, S., Tam, S. K., Magassouba, D., and Hallé, J.-P. (2009). Direct effect of alginate purification on the survival of islets immobilized in alginate-based microcapsules. Acta Biomater. 5 (9), 3433–3440. doi:10.1016/j.actbio.2009.05.029
Liang, Z., Rybtke, M., Kragh, K. N., Johnson, O., Schicketanz, M., Zhang, Y. E., et al. (2022). Transcription of the alginate operon in Pseudomonas aeruginosa is regulated by c-di-GMP. Microbiol. Spectr. 10 (4), e0067522–e0067522. doi:10.1128/spectrum.00675-22
López-Pliego, L., García-Ramírez, L., Cruz-Gómez, E. A., Domínguez-Ojeda, P., López-Pastrana, A., Fuentes-Ramírez, L. E., et al. (2018). Transcriptional study of the RsmZ-sRNAs and their relationship to the biosynthesis of alginate and alkylresorcinols in Azotobacter vinelandii. Mol. Biotechnol. 60 (9), 670–680. doi:10.1007/s12033-018-0102-7
López-Pliego, L., Mena-Muñoz, G., Terán-Melo, J. L., Fuentes, L. E., Núñez, C. E., and Castañeda, M. (2020). Study of the sRNA RsmY involved in the genetic regulation of the synthesis of alginate and alkyl resorcinols in Azotobacter vinelandii. Archives Microbiol. 202 (3), 579–589. doi:10.1007/s00203-019-01769-y
Lozano, E., Galindo, E., and Peña, C. F. (2011). Oxygen transfer rate during the production of alginate by Azotobacter vinelandii under oxygen-limited and non oxygen-limited conditions. Microb. Cell Factories 10 (1), 13. doi:10.1186/1475-2859-10-13
Ma, L., Tan, Y., Chen, X., Ran, Y., Tong, Q., Tang, L., et al. (2022). Injectable oxidized alginate/carboxylmethyl chitosan hydrogels functionalized with nanoparticles for wound repair. Carbohydr. Polym. 293, 119733. doi:10.1016/j.carbpol.2022.119733
Ma, S., Selvaraj, U., Ohman, D. E., Quarless, R., Hassett, D. J., and Wozniak, D. J. (1998). Phosphorylation-independent activity of the response regulators AlgB and AlgR in promoting alginate biosynthesis in mucoid Pseudomonas aeruginosa. J. Bacteriol. 180 (4), 956–968. doi:10.1128/jb.180.4.956-968.1998
Maier, U., and Büchs, J. (2001). Characterisation of the gas–liquid mass transfer in shaking bioreactors. Biochem. Eng. J. 7 (2), 99–106. doi:10.1016/S1369-703X(00)00107-8
Maleki, S., Mærk, M., Hrudikova, R., Valla, S., and Ertesvåg, H. (2017). New insights into Pseudomonas fluorescens alginate biosynthesis relevant for the establishment of an efficient production process for microbial alginates. New Biotechnol. 37, 2–8. doi:10.1016/j.nbt.2016.08.005
Mallakpour, S., Azadi, E., and Hussain, C. M. (2021). State-of-the-art of 3D printing technology of alginate-based hydrogels—An emerging technique for industrial applications. Adv. Colloid Interface Sci. 293, 102436. doi:10.1016/j.cis.2021.102436
Mallakpour, S., and Mohammadi, N. (2022). Development of sodium alginate-pectin/TiO2 nanocomposites: antibacterial and bioactivity investigations. Carbohydr. Polym. 285, 119226. doi:10.1016/j.carbpol.2022.119226
Manzo, J., Cocotl-Yañez, M., Tzontecomani, T., Martínez, V. M., Bustillos, R., Velásquez, C., et al. (2012). Post-transcriptional regulation of the alginate biosynthetic gene algD by the Gac/Rsm system in Azotobacter vinelandii. J. Mol. Microbiol. Biotechnol. 21 (3-4), 147–159. doi:10.1159/000334244
Martău, G. A., Mihai, M., and Vodnar, D. C. (2019). The use of Chitosan, alginate, and Pectin in the biomedical and food sector—biocompatibility, bioadhesiveness, and biodegradability. Polymers 11 (11), 1837. doi:10.3390/polym11111837
Martínez-Ortiz, I. C., Ahumada-Manuel, C. L., Hsueh, B. Y., Guzmán, J., Moreno, S., Cocotl-Yañez, M., et al. (2020). Cyclic di-GMP-Mediated regulation of extracellular mannuronan C-5 epimerases is essential for cyst formation in Azotobacter vinelandii. J. Bacteriol. 202 (24), e00135-20. doi:10.1128/jb.00135-20
Martínez-Salazar, J. M., Moreno, S., Nájera, R., Boucher, J. C., Espín, G., Soberón-Chávez, G., et al. (1996). Characterization of the genes coding for the putative sigma factor AlgU and its regulators MucA, MucB, MucC, and MucD in Azotobacter vinelandii and evaluation of their roles in alginate biosynthesis. J. Bacteriol. 178 (7), 1800–1808. doi:10.1128/jb.178.7.1800-1808.1996
Mathee, K., McPherson, C. J., and Ohman, D. E. (1997). Posttranslational control of the algT (algU)-encoded sigma22 for expression of the alginate regulon in Pseudomonas aeruginosa and localization of its antagonist proteins MucA and MucB (AlgN). J. Bacteriol. 179 (11), 3711–3720. doi:10.1128/jb.179.11.3711-3720.1997
Medina, A., Castillo, T., Flores, C., Núñez, C., Galindo, E., and Peña, C. (2023). Production of alginates with high viscosifying power and molecular weight by using the AT9 strain of Azotobacter vinelandii in batch cultures under different oxygen transfer conditions. J. Chem. Technol. Biotechnol. 98 (2), 537–543. doi:10.1002/jctb.7267
Mejı́a-Ruı́z, H., Guzmán, J., Moreno, S., Soberón-Chávez, G., and Espı́n, G. (1997). The Azotobacter vinelandii alg8 and alg44 genes are essential for alginate synthesis and can be transcribed from an algD-independent promoter. Gene 199 (1), 271–277. doi:10.1016/S0378-1119(97)00380-6
Merighi, M., Lee, V. T., Hyodo, M., Hayakawa, Y., and Lory, S. (2007). The second messenger bis-(3′-5′)-cyclic-GMP and its PilZ domain-containing receptor Alg44 are required for alginate biosynthesis in Pseudomonas aeruginosa. Mol. Microbiol. 65 (4), 876–895. doi:10.1111/j.1365-2958.2007.05817.x
Mollah, M. Z. I., Zahid, H. M., Mahal, Z., Faruque, M. R. I., and Khandaker, M. U. (2021). The usages and potential uses of alginate for healthcare applications. Front. Mol. Biosci. 8, 719972. doi:10.3389/fmolb.2021.719972
Moradali, M. F., Donati, I., Sims, I. M., Ghods, S., and Rehm, B. H. A. (2015). Alginate polymerization and modification are linked in Pseudomonas aeruginosa. mBio 6 (3), e00453–e00415. doi:10.1128/mbio.00453-15
Moradali, M. F., Ghods, S., and Rehm, B. H. A. (2018). “Alginate biosynthesis and biotechnological production,” in Alginates and their biomedical applications. Editors En B. H. A. Rehm,, and M. F. Moradali (Singapore: Springer), 1–25. doi:10.1007/978-981-10-6910-9_1
Moradali, M. F., and Rehm, B. H. A. (2020). “The regulation of alginate biosynthesis via cyclic di-GMP signaling,” in Microbial cyclic Di-Nucleotide signaling. Editors En S.-H. Chou, N. Guiliani, V. T. Lee, and U. Römling (Springer International Publishing), 223–238. doi:10.1007/978-3-030-33308-9_14
Moreno, S., Ertesvåg, H., Valla, S., Núñez, C., Espin, G., and Cocotl-Yañez, M. (2018). RpoS controls the expression and the transport of the AlgE1-7 epimerases in Azotobacter vinelandii. FEMS Microbiol. Lett. 365 (19), fny210. doi:10.1093/femsle/fny210
Nivens, D. E., Ohman, D. E., Williams, J., and Franklin, M. J. (2001). Role of alginate and its O acetylation in formation of Pseudomonas aeruginosa microcolonies and biofilms. J. Bacteriol. 183 (3), 1047–1057. doi:10.1128/jb.183.3.1047-1057.2001
Noar, J. D., and Bruno-Bárcena, J. M. (2018). Azotobacter vinelandii: the source of 100 years of discoveries and many more to come. Microbiology 164 (4), 421–436. doi:10.1099/mic.0.000643
Núñez, C., López-Pliego, L., Ahumada-Manuel, C. L., and Castañeda, M. (2022). Genetic regulation of alginate production in Azotobacter vinelandii a bacterium of biotechnological interest: a mini-review. Front. Microbiol. 13, 845473. doi:10.3389/fmicb.2022.845473
Oelze, J. (2000). Respiratory protection of nitrogenase in Azotobacter species: is a widely held hypothesis unequivocally supported by experimental evidence? FEMS Microbiol. Rev. 24 (4), 321–333. doi:10.1111/j.1574-6976.2000.tb00545.x
Oglesby, L. L., Jain, S., and Ohman, D. E. (2008). Membrane topology and roles of Pseudomonas aeruginosa Alg8 and Alg44 in alginate polymerization. Microbiology 154 (6), 1605–1615. doi:10.1099/mic.0.2007/015305-0
Pacheco-Leyva, I., Guevara Pezoa, F., and Díaz-Barrera, A. (2016). Alginate biosynthesis in Azotobacter vinelandii: overview of molecular mechanisms in connection with the oxygen availability. Int. J. Polym. Sci. 2016 (1), 2062360. doi:10.1155/2016/2062360
Paredes-Juarez, G. A., De Haan, B. J., Faas, M. M., and De Vos, P. (2014). A technology platform to test the efficacy of purification of alginate. Materials 7 (3), 2087–2103. doi:10.3390/ma7032087
Peña, C., Campos, N., and Galindo, E. (1997). Changes in alginate molecular mass distributions, broth viscosity and morphology of Azotobacter vinelandii cultured in shake flasks. Appl. Microbiol. Biotechnol. 48 (4), 510–515. doi:10.1007/s002530051088
Peña, C., Galindo, E., and Büchs, J. (2011). The viscosifying power, degree of acetylation and molecular mass of the alginate produced by Azotobacter vinelandii in shake flasks are determined by the oxygen transfer rate. Process Biochem. 46 (1), 290–297. doi:10.1016/j.procbio.2010.08.025
Peña, C., Hernández, L., and Galindo, E. (2006). Manipulation of the acetylation degree of Azotobacter vinelandii alginate by supplementing the culture medium with 3-(N-morpholino)-propane-sulfonic acid. Lett. Appl. Microbiol. 43 (2), 200–204. doi:10.1111/j.1472-765X.2006.01925.x
Peña, C., Miranda, L., Segura, D., Núñez, C., Espín, G., and Galindo, E. (2002). Alginate production by Azotobacter vinelandii mutants altered in poly-β-hydroxybutyrate and alginate biosynthesis. J. Industrial Microbiol. Biotechnol. 29 (5), 209–213. doi:10.1038/sj.jim.7000310
Petersen, A. B., Tøndervik, A., Gaardløs, M., Ertesvåg, H., Sletta, H., and Aachmann, F. L. (2023). Mannuronate C-5 epimerases and their use in alginate modification. Essays Biochem. 67 (3), 615–627. doi:10.1042/EBC20220151
Ponce, B., Urtuvia, V., Maturana, N., Peña, C., and Díaz-Barrera, A. (2021). Aumento de la producción de alginato y de los niveles de transcripción de la alginato liasa (alyA1) mediante el control de la tasa de transferencia de oxígeno en cultivos de Azotobacter vinelandii en condiciones diazotróficas. Electron. J. Biotechnol. 52, 35–44. doi:10.1016/j.ejbt.2021.04.007
Pritchard, M. F., Jack, A. A., Powell, L. C., Sadh, H., Rye, P. D., Hill, K. E., et al. (2017). Alginate oligosaccharides modify hyphal infiltration of Candida albicans in an in vitro model of invasive human candidosis. J. Appl. Microbiol. 123 (3), 625–636. doi:10.1111/jam.13516
Qamar, S., Karim, S., Aslam, S., Jahangeer, M., Nelofer, R., Nadeem, A. A., et al. (2024). Alginate-based bio-nanohybrids with unique properties for biomedical applications. Starch - Stärke 76 (1-2), 2200100. doi:10.1002/star.202200100
Qi, Y., Lu, L., Zhou, C., and Luo, B. (2009). “Purification of alginate for tissue engineering,” in 2009 3rd international conference on bioinformatics and biomedical engineering. Beijing, China, 1–4. doi:10.1109/ICBBE.2009.5163263
Qiu, D., Eisinger, V. M., Head, N. E., Pier, G. B., and Yu, H. D. (2008). ClpXP proteases positively regulate alginate overexpression and mucoid conversion in Pseudomonas aeruginosa. Microbiology 154 (7), 2119–2130. doi:10.1099/mic.0.2008/017368-0
Qiu, D., Eisinger, V. M., Rowen, D. W., and Yu, H. D. (2007). Regulated proteolysis controls mucoid conversion in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U. S. A. 104 (19), 8107–8112. doi:10.1073/pnas.0702660104
Quiroz-Rocha, E., Bonilla-Badía, F., García-Aguilar, V., López-Pliego, L., Serrano-Román, J., Cocotl-Yañez, M., et al. (2017). Two-component system CbrA/CbrB controls alginate production in Azotobacter vinelandii. Microbiology 163 (7), 1105–1115. doi:10.1099/mic.0.000457
Ramos, P. E., Silva, P., Alario, M. M., Pastrana, L. M., Teixeira, J. A., Cerqueira, M. A., et al. (2018). Effect of alginate molecular weight and M/G ratio in beads properties foreseeing the protection of probiotics. Food Hydrocoll. 77, 8–16. doi:10.1016/j.foodhyd.2017.08.031
Rashidzadeh, B., Shokri, E., Mahdavinia, G. R., Moradi, R., Mohamadi-Aghdam, S., and Abdi, S. (2020). Preparation and characterization of antibacterial magnetic-/pH-sensitive alginate/Ag/Fe3O4 hydrogel beads for controlled drug release. Int. J. Biol. Macromol. 154, 134–141. doi:10.1016/j.ijbiomac.2020.03.028
Rehman, Z. U., Wang, Y., Moradali, M. F., Hay, I. D., and Rehm, B. H. A. (2013). Insights into the assembly of the alginate biosynthesis machinery in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 79 (10), 3264–3272. doi:10.1128/AEM.00460-13
Remminghorst, U., Hay, I. D., and Rehm, B. H. A. (2009). Molecular characterization of Alg8, a putative glycosyltransferase, involved in alginate polymerisation. J. Biotechnol. 140 (3), 176–183. doi:10.1016/j.jbiotec.2009.02.006
Remminghorst, U., and Rehm, B. H. A. (2006a). Bacterial alginates: from biosynthesis to applications. Biotechnol. Lett. 28 (21), 1701–1712. doi:10.1007/s10529-006-9156-x
Remminghorst, U., and Rehm, B. H. A. (2006b). In vitro alginate polymerization and the functional role of Alg8 in alginate production by Pseudomonas aeruginosa. Applied and environmental microbiology. Appl. Environ. Microbiol. 72 (1), 298–305. doi:10.1128/AEM.72.1.298-305.2006
Riley, L. M., Weadge, J. T., Baker, P., Robinson, H., Codée, J. D. C., Tipton, P. A., et al. (2013). Structural and functional characterization of Pseudomonas aeruginosa AlgX: role of AlgX in alginate acetylation. J. Biol. Chem. 288 (31), 22299–22314. doi:10.1074/jbc.M113.484931
Robles-Price, A., Wong, T. Y., Sletta, H., Valla, S., and Schiller, N. L. (2004). AlgX is a periplasmic protein required for alginate biosynthesis in Pseudomonas aeruginosa. J. Bacteriol. 186 (21), 7369–7377. doi:10.1128/jb.186.21.7369-7377.2004
Rodríguez-Martínez, K., Muriel-Millán, L. F., Ortíz-Vasco, C., Moreno, S., Soberón-Chávez, G., and Espín, G. (2023). Defining the regulatory mechanisms of sigma factor RpoS degradation in Azotobacter vinelandii and Pseudomonas aeruginosa. Mol. Microbiol. 120 (1), 91–102. doi:10.1111/mmi.15107
Romero, Y., Guzmán, J., Moreno, S., Cocotl-Yañez, M., Vences-Guzmán, M. Á., Castañeda, M., et al. (2016). The GacS/A-RsmA signal transduction pathway controls the synthesis of alkylresorcinol lipids that replace membrane phospholipids during encystment of Azotobacter vinelandii SW136. PLoS One 11 (4), e0153266. doi:10.1371/journal.pone.0153266
Römling, U., Galperin, M. Y., and Gomelsky, M. (2013). Cyclic di-GMP: the first 25 years of a universal bacterial second messenger. Microbiol. Mol. Biol. Rev. 77 (1), 1–52. doi:10.1128/mmbr.00043-12
Rosales-Cruz, A., Reyes-Nicolau, J., Minto-González, E., Meneses-Carbajal, A., Mondragón-Albarrán, C., López-Pliego, L., et al. (2024). A homolog of the histidine kinase RetS controls the synthesis of alginates, PHB, alkylresorcinols, and motility in Azotobacter vinelandii. Curr. Microbiol. 81 (10), 311. doi:10.1007/s00284-024-03835-1
Rowley, J. A., and Mooney, D. J. (2002). Alginate type and RGD density control myoblast phenotype. J. Biomed. Mater. Res. 60 (2), 217–223. doi:10.1002/jbm.1287
Sabra, W., Zeng, A.-P., Lünsdorf, H., and Deckwer, W.-D. (2000). Effect of oxygen on formation and structure of Azotobacter vinelandii alginate and its role in protecting nitrogenase. Appl. Environ. Microbiol. 66 (9), 4037–4044. doi:10.1128/AEM.66.9.4037-4044.2000
Saji, S., Hebden, A., Goswami, P., and Du, C. (2022). A brief review on the development of alginate extraction process and its sustainability. Sustainability 14 (9), 5181. doi:10.3390/su14095181
Sanchez-Ballester, N. M., Bataille, B., and Soulairol, I. (2021). Sodium alginate and alginic acid as pharmaceutical excipients for tablet formulation: structure-function relationship. Carbohydr. Polym. 270, 118399. doi:10.1016/j.carbpol.2021.118399
Schmid, W., and Picker-Freyer, K. M. (2009). Tableting and tablet properties of alginates: characterisation and potential for soft tableting. Eur. J. Pharm. Biopharm. 72 (1), 165–172. doi:10.1016/j.ejpb.2008.10.006
Setubal, J. C., Dos Santos, P., Goldman, B. S., Ertesvåg, H., Espin, G., Rubio, L. M., et al. (2009). Genome sequence of Azotobacter vinelandii, an obligate aerobe specialized to support diverse Anaerobic metabolic processes. J. Bacteriol. 191 (14), 4534–4545. doi:10.1128/JB.00504-09
Shinabarger, D., Berry, A., May, T. B., Rothmel, R., Fialho, A., and Chakrabarty, A. M. (1991). Purification and characterization of phosphomannose isomerase-guanosine diphospho-D-mannose pyrophosphorylase. A bifunctional enzyme in the alginate biosynthetic pathway of Pseudomonas aeruginosa. J. Biol. Chem. 266 (4), 2080–2088. doi:10.1016/S0021-9258(18)52212-2
Sondermeijer, H. P., Witkowski, P., Woodland, D., Seki, T., Aangenendt, F. J., van der Laarse, A., et al. (2016). Optimization of alginate purification using polyvinylidene difluoride membrane filtration: effects on immunogenicity and biocompatibility of three-dimensional alginate scaffolds. J. Biomaterials Appl. 31 (4), 510–520. doi:10.1177/0885328216645952
Sriamornsak, P., Thirawong, N., and Korkerd, K. (2007). Swelling, erosion and release behavior of alginate-based matrix tablets. Eur. J. Pharm. Biopharm. 66 (3), 435–450. doi:10.1016/j.ejpb.2006.12.003
Stanisci, A., Tøndervik, A., Gaardløs, M., Lervik, A., Skjåk-Bræk, G., Sletta, H., et al. (2020). Identification of a pivotal residue for determining the block structure-forming properties of alginate C-5 epimerases. ACS Omega 5 (8), 4352–4361. doi:10.1021/acsomega.9b04490
Szekalska, M., Puciłowska, A., Szymańska, E., Ciosek, P., and Winnicka, K. (2016). Alginate: current use and future perspectives in pharmaceutical and biomedical applications. Int. J. Polym. Sci. 2016 (1), 7697031–17. doi:10.1155/2016/7697031
Tan, J., Luo, Y., Guo, Y., Zhou, Y., Liao, X., Li, D., et al. (2023). Development of alginate-based hydrogels: crosslinking strategies and biomedical applications. Int. J. Biol. Macromol. 239, 124275. doi:10.1016/j.ijbiomac.2023.124275
Tao, L., Shi, C., Zi, Y., Zhang, H., Wang, X., and Zhong, J. (2024). A review on the chemical modification of alginates for food research: chemical nature, modification methods, product types, and application. Food Hydrocoll. 147, 109338. doi:10.1016/j.foodhyd.2023.109338
Tatnell, P. J., Russell, N. J., and Gacesa, P. (1994). GDP-mannose dehydrogenase is the key regulatory enzyme in alginate biosynthesis in Pseudomonas aeruginosa: evidence from metabolite studies. Microbiology 140 (7), 1745–1754. doi:10.1099/13500872-140-7-1745
Tavares, I. M., Leitão, J. H., Fialho, A. M., and Sá-Correia, I. (1999). Pattern of changes in the activity of enzymes of GDP-D-mannuronic acid synthesis and in the level of transcription of algA, algC and algD genes accompanying the loss and emergence of mucoidy in Pseudomonas aeruginosa. Res. Microbiol. 150 (2), 105–116. doi:10.1016/S0923-2508(99)80028-X
Tomić, S. L., Babić Radić, M. M., Vuković, J. S., Filipović, V. V., Nikodinovic-Runic, J., and Vukomanović, M. (2023). Alginate-based hydrogels and scaffolds for biomedical applications. Mar. Drugs 21 (3), 177. doi:10.3390/md21030177
Tøndervik, A., Aarstad, O. A., Aune, R., Maleki, S., Rye, P. D., Dessen, A., et al. (2020). Exploiting Mannuronan C-5 epimerases in commercial alginate production. Mar. Drugs 18 (11), 565. doi:10.3390/md18110565
Topuz, F., Henke, A., Richtering, W., and Groll, J. (2012). Magnesium ions and alginate do form hydrogels: a rheological study. Soft Matter 8 (18), 4877–4881. doi:10.1039/C2SM07465F
Torres, M. L., Fernandez, J. M., Dellatorre, F. G., Cortizo, A. M., and Oberti, T. G. (2019). Purification of alginate improves its biocompatibility and eliminates cytotoxicity in matrix for bone tissue engineering. Algal Res. 40, 101499. doi:10.1016/j.algal.2019.101499
Urtuvia, V., Maturana, N., Acevedo, F., Peña, C., and Díaz-Barrera, A. (2017). Bacterial alginate production: an overview of its biosynthesis and potential industrial production. World J. Microbiol. Biotechnol. 33 (11), 198–10. doi:10.1007/s11274-017-2363-x
Valentini, M., and Filloux, A. (2016). Biofilms and cyclic di-GMP (c-di-GMP) signaling: lessons from Pseudomonas aeruginosa and other bacteria. J. Biol. Chem. 291 (24), 12547–12555. doi:10.1074/jbc.R115.711507
Wang, J., Liu, S., Huang, J., Ren, K., Zhu, Y., and Yang, S. (2023). Alginate: microbial production, functionalization, and biomedical applications. Int. J. Biol. Macromol. 242, 125048. doi:10.1016/j.ijbiomac.2023.125048
Wang, Y., Hay, I. D., Rehman, Z. U., and Rehm, B. H. A. (2015). Membrane-anchored MucR mediates nitrate-dependent regulation of alginate production in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 99 (17), 7253–7265. doi:10.1007/s00253-015-6591-4
Whitney, J. C., Hay, I. D., Li, C., Eckford, P. D. W., Robinson, H., Amaya, M. F., et al. (2011). Structural basis for alginate secretion across the bacterial outer membrane. Proc. Natl. Acad. Sci. U. S. A. 108 (32), 13083–13088. doi:10.1073/pnas.1104984108
Whitney, J. C., Whitfield, G. B., Marmont, L. S., Yip, P., Neculai, A. M., Lobsanov, Y. D., et al. (2015). Dimeric c-di-GMP is required for post-translational regulation of alginate production in Pseudomonas aeruginosa. J. Biol. Chem. 290 (20), 12451–12462. doi:10.1074/jbc.M115.645051
Wolfram, F., Kitova, E. N., Robinson, H., Walvoort, M. T. C., Codée, J. D. C., Klassen, J. S., et al. (2014). Catalytic mechanism and mode of action of the periplasmic alginate epimerase AlgG. J. Biol. Chem. 289 (9), 6006–6019. doi:10.1074/jbc.M113.533158
Wood, L. F., and Ohman, D. E. (2006). Independent regulation of MucD, an HtrA-Like protease in Pseudomonas aeruginosa, and the role of its proteolytic motif in alginate gene regulation. J. Bacteriol. 188 (8), 3134–3137. doi:10.1128/jb.188.8.3134-3137.2006
Wu, G., Cruz-Ramos, H., Hill, S., Green, J., Sawers, G., and Poole, R. K. (2000). Regulation of cytochrome bd expression in the obligate aerobe Azotobacter vinelandii by CydR (Fnr): sensitivity to oxygen, reactive oxygen species, and nitric oxide. J. Biol. Chem. 275 (7), 4679–4686. doi:10.1074/jbc.275.7.4679
Wu, G., Moir, A. J. G., Sawers, G., Hill, S., and Poole, R. K. (2001). Biosynthesis of poly-β-hydroxybutyrate (PHB) is controlled by CydR (Fnr) in the obligate aerobe Azotobacter vinelandii. FEMS Microbiol. Lett. 194 (2), 215–220. doi:10.1111/j.1574-6968.2001.tb09472.x
Xing, M., Cao, Q., Wang, Y., Xiao, H., Zhao, J., Zhang, Q., et al. (2020). Advances in research on the bioactivity of alginate oligosaccharides. Mar. Drugs 18 (3), 144. doi:10.3390/md18030144
Yang, J.-S., Xie, Y.-J., and He, W. (2011). Research progress on chemical modification of alginate: a review. Carbohydr. Polym. 84 (1), 33–39. doi:10.1016/j.carbpol.2010.11.048
Ye, R. W., Zielinski, N. A., and Chakrabarty, A. M. (1994). Purification and characterization of phosphomannomutase/phosphoglucomutase from Pseudomonas aeruginosa involved in biosynthesis of both alginate and lipopolysaccharide. J. Bacteriol. 176 (16), 4851–4857. doi:10.1128/jb.176.16.4851-4857.1994
Zhang, P., Yuan, L., Zeng, J., Zou, K., Liu, B., Qing, T., et al. (2022). Alginate production of Pseudomonas strains and its application in preparation of alginate-biomass hydrogel for heavy metal adsorption. Int. J. Biol. Macromol. 222, 1511–1521. doi:10.1016/j.ijbiomac.2022.09.252
Keywords: alginate, Azotobacter vinelandii, biosynthesis, regulation, oxygen transfer rate
Citation: Ponce B, Zamora-Quiroz A, González E, Andler R and Díaz-Barrera A (2025) Advances in alginate biosynthesis: regulation and production in Azotobacter vinelandii. Front. Bioeng. Biotechnol. 13:1593893. doi: 10.3389/fbioe.2025.1593893
Received: 14 March 2025; Accepted: 21 July 2025;
Published: 31 July 2025.
Edited by:
Xupeng Cao, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Dão Neto, Federal Institute of Education, Science, and Technology of Paraná (IFPR), BrazilFabián Martínez, University of Santiago, Chile
Copyright © 2025 Ponce, Zamora-Quiroz, González, Andler and Díaz-Barrera. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alvaro Díaz-Barrera, YWx2YXJvLmRpYXpAcHVjdi5jbA==