 Youngshim Choi
Youngshim Choi Yinyan Ma
Yinyan Ma Samson Tom
Samson Tom Alla Danilkovitch
Alla Danilkovitch Liqing Yu
Liqing Yu- 1Division of Endocrinology, Diabetes and Nutrition, Department of Medicine, University of Maryland School of Medicine, Baltimore, MD, United States
- 2Britecyte, Inc., Fredrick, MD, United States
by Choi Y, Ma Y, Tom S, Danilkovitch A and Yu L (2025). Front. Bioeng. Biotechnol. 13:1579062. doi: 10.3389/fbioe.2025.1579062
In the published article, there was an error in Figure 7B as published. The graph in Figure 7B was identical to Figure 7A, and has been replaced with the correct version. The corrected Figure 7B and its caption “FIGURE 7: In vivo immunogenicity of xenogeneic hAF and allogeneic rAF in rats. Detection of (A) anti-hAF or (B) anti-rAF antibodies by ELISA coated with hAF or rAF extracts, respectively. Control: serum from rats treated with PBS. Background: no serum. Data are presented as mean ± SD by OD at 450 nm for each serum concentration (n = 6/group).” appear below.
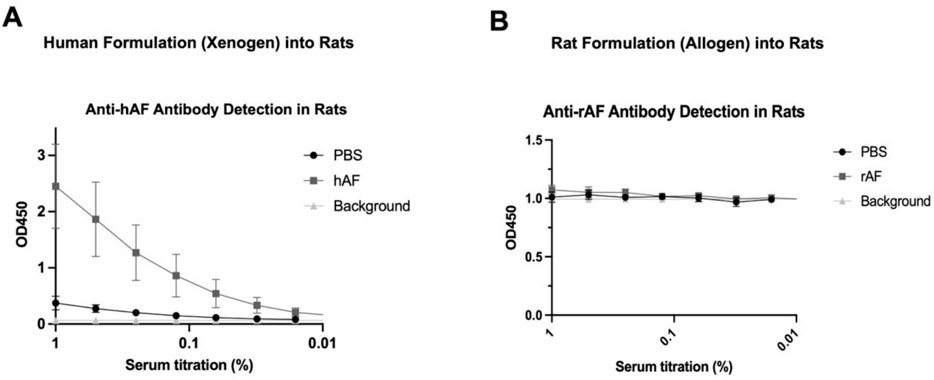
Figure 7. In vivo immunogenicity of xenogeneic hAF and allogeneic rAF in rats. Detection of (A) anti-hAF or (B) anti-rAF antibodies by ELISA coated with hAF or rAF extracts, respectively. Control: serum from rats treated with PBS. Background: no serum. Data are presented as mean ± SD by OD at 450 nm for each serum concentration (n = 6/group).
The original version of this article has been updated.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Keywords: adipose tissue, adipose stem cells, tissue engineering, MASLD, fibrosis, inflammation
Citation: Choi Y, Ma Y, Tom S, Danilkovitch A and Yu L (2025) Correction: An engineered adipose formulation decreases hepatic inflammation and fibrosis in a rodent model of metabolic dysfunction-associated steatotic liver disease. Front. Bioeng. Biotechnol. 13:1642526. doi: 10.3389/fbioe.2025.1642526
Received: 06 June 2025; Accepted: 18 June 2025;
Published: 26 June 2025.
Edited and reviewed by:
Luca Urbani, Foundation for Liver Research, United KingdomCopyright © 2025 Choi, Ma, Tom, Danilkovitch and Yu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alla Danilkovitch, YWRhbmlsa292aXRjaEBicml0ZWN5dGUuY29t; Liqing Yu, bHl1QHNvbS51bWFyeWxhbmQuZWR1
†These authors have contributed equally to this work