 Li-Jian Zhou1†
Li-Jian Zhou1† Tong Huan
Tong Huan Yue Wu
Yue Wu Ye-Wang Zhang
Ye-Wang Zhang- 1The People’s Hospital of Danyang, Affiliated Danyang Hospital of Nantong University, Zhenjiang, China
- 2School of Medicine, Nantong University, Nantong, China
- 3School of Pharmacy, Jiangsu University, Zhenjiang, China
Nicotinamide adenine dinucleotide (phosphate) oxidases are the enzymes that catalyze the oxidation of NAD(P)H to produce NAD(P)+, which is the cofactor of many dehydrogenases. To reduce costs, cofactor regeneration of NAD(P)+ is essential for both enzymatic and whole-cell biotransformations. In the present review, the enzymatic or microbial production of rare sugars like L-tagatose, L-xylulose, L-gulose, and L-sorbose with cofactor regeneration was summarized. And the cofactor regeneration in some value-added chemicals, including acetoin, 1,3-dihydroacetone, vanillic acid, chlorolactone, acetophenone, and kinetic resolution of racemic phenylethanol by employing NADH/NADPH oxidase was also reviewed. The engineering of these enzymes by modifying the enzyme surface, reshaping the catalytic pocket, and mutating the substrate-binding domain of NADH oxidase to improve the catalytic performance for potential industrial applications was discussed in the future outlook.
1 Introduction
Nicotinamide adenine dinucleotide (NAD), as the most important coenzyme, could help other NAD-dependent enzymes to catalyze chemical reactions by switching between the reduced and oxidized form (NAD+/NADH). In eukaryotic cells, the energy could be produced using NADH in cellular metabolism. The process from NADH to NAD+ is generally catalyzed by NADH oxidase (NOX) and produces reactive oxygen species (ROS) in eukaryotic cells, and another main source of ROS in vivo is NADPH oxidase. These two categories of enzymes catalyze the NAD(P)H to produce NAD(P)+ for coupling with the dehydrogenases catalyzed oxidoreduction reactions for the production of enantiopure chemicals or pharmaceuticals. The increasing attention on these enzymes is due to their potential capacity to regenerate the expensive cofactor NAD(P)+ in industrial enzymatic preparation of value-added products with NAD(P)+-dependent dehydrogenases.
Most NOXs have a highly conserved catalytic residue -cysteine, located in the active site. They catalyze the oxidation reactions of NADH to NAD+ coupled with the four-electron transfer mode by consumption of O2 or the two-electron transfer mode by reducing H2O2 (Li et al., 2018). Compared with the H2O2-forming NOX, the H2O-forming NOX is more interesting because of its good compatibility in enzymatic reactions in aqueous solution. Similarly, the oxidized NADP+ could also be recycled by NADPH oxidase and follows a similar reaction mechanism.
Oxidoreductase is the largest class of enzymes, and NAD(P)+ is the most common cofactor of the oxidoreductase. Now, the chemical synthesis route in chemical, paper, cosmetic, and other industries is undergoing a change to biocatalytic, especially enzymatic methods. In this review, we summarized the potential industrial production of value-added chemicals with cofactor regeneration of NAD+/NADP+. In the enzymatic reactions by using coupled or combined enzymes, the strategy of protein engineering for remodeling the enzyme is also included to improve the catalytic efficiency.
2 Enzymatic production of rare sugars with dehydrogenase and NOX
Rare sugars are highly valued mono- or disaccharides with beneficial health effects. However, most synthetic chemical routes are both limited and economically unfeasible due to the expensive raw materials, harsh reaction conditions, and severe environmental pollution. Enzymatic transformation has become a powerful tool in the field. Many NAD-dependent enzymes are hydrogenases, which oxidize alcohols or sugars (Stolarczyk et al., 2020) to produce rare sugars. The cofactors are bound weakly to the enzyme and could be easily released from the protein. Thus, the regeneration of the cofactors is essential for the combined or cascade enzymatic reactions, which will significantly lower the cost. In the past decade, several L-form rare sugars have been successfully synthesized with dehydrogenase coupled with NAD(P)H oxidases as shown in Table 1.
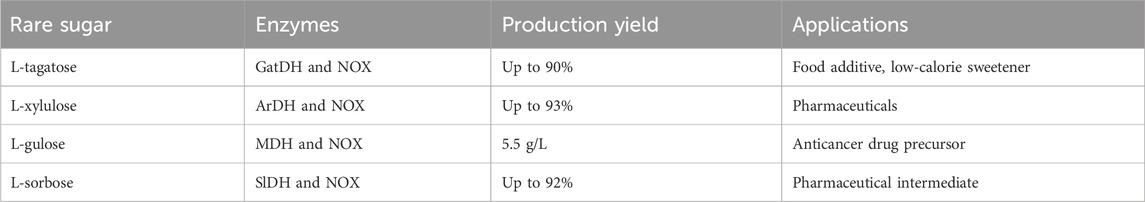
Table 1. Enzymatic synthesis of the rare sugars and their applications.
2.1 Production of L-tagatose with galactitol dehydrogenase
As a rare sugar, L-tagatose has great potential applications in the therapy of diabetes, food additives, and cosmetics because of its low-calorie value (Mahmood et al., 2021). Considering the increasing diabetes and obesity, low- or zero-caloric sweeteners are gaining attention in the food industry (Ravikumar et al., 2022; Ravikumar et al., 2021). The biological production method has drawn obvious achievements for L-tagatose production (Lu et al., 2024; Yuan et al., 2021). However, chemical synthesis of L-tagatose leads to low yield and by-products (Tang et al., 2022). Su et al. used galactitol dehydrogenase (GatDH) and H2O-forming NOX (SmNox) for efficient enzymatic synthesis of L-tagatose (Su et al., 2021). The yield of 90% after 12 h reaction was obtained when the substrate concentration of 100 mM and 3 mM NAD+ was supplied. There was no by-product formation and a high yield of L-tagatose with low costs using this enzymatic route (Figure 1). Later, they prepared the combined cross-linked enzyme aggregates containing GatDH and SmNox for the synthesis of L-tagatose (Li et al., 2022b). The combined cross-linked enzyme aggregates exhibited high thermal stability and industrial potential for the L-tagatose preparation.
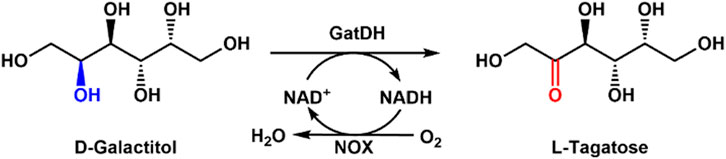
Figure 1. Enzymatic production of L-tagatose from D-galactitol with NOX and GatDH.
2.2 Enzymatic production of L-xylulose with arabinitol dehydrogenase
L-Xylulose, as another important rare sugar, is usually used as an anticancer and cardioprotective agent or as a precursor for the preparation of antiviral drugs. The chemical synthesis of the L-xylulose method has disadvantages of low yield, environmental pollution, and low purity because it is difficult to separate from other isomers. The substrate for the biological preparation of L-xylulose could be L-arabinitol or xylitol.
Gao et al. reported the biotransformation of L-arabinitol to L-xylulose with immobilized whole Escherichia coli cells expressed L-arabinitol dehydrogenase and NOX (Gao et al., 2015). By coupling two enzymes in the cells, a molar conversion of 96% from L-arabinitol to L-xylulose was achieved. It was reported later that a product titer of 48.45 g/L for L-xylulose was reached by co-expressing the two enzymes in E. coli with pETDuet-xdh-smnox (Tesfay et al., 2021). They immobilized the E. coli cells expressing the two enzymes with entrapment, and a yield of 96% was obtained when the substrate concentration was 150 mM (Gao et al., 2016a). The group then developed a cell-free system containing the two enzymes for the synthesis of L-xylulose from arabinitol, and a final yield of 78.4% was achieved when the substrate concentration was 250 mM (Gao et al., 2016b). Then they immobilized the two enzymes onto inorganic hybrid nanoflowers to enhance the L-xylulose production, and a maximum yield of 91% was observed, which was 2.9-fold higher than the free enzymes (Patel et al., 2017). They demonstrated a sequential co-immobilization of L-arabinitol dehydrogenase and NADH oxidase for the conversion of L-arabinitol to L-xylulose recently (Patel et al., 2024). The results indicated that the co-immobilized enzymes exhibited 6.5-fold higher activity than that of free enzymes, and the maximum conversion of 93.6% was obtained.
Generally, xylitol could be enzymatically converted into xylulose with xylitol dehydrogenase. Another possible enzymatic route is using arabinitol dehydrogenase, which is an NAD-dependent short-chain dehydrogenase that can convert xylitol to L-xylulose. Zhu et al. (2022) cloned arabinitol dehydrogenase (ArDH) from Aspergillus nidulans for the preparation of L-xylulose from xylitol and coupled NOX for the cofactor NAD+ regeneration (Figure 2). It was found that the high substrate concentration of xylitol inhibits the enzymatic reaction. A yield of 92.7% was achieved when the substrate concentration was 10 mM, but the conversion was only 18.4% when the substrate concentration was 80 mM.
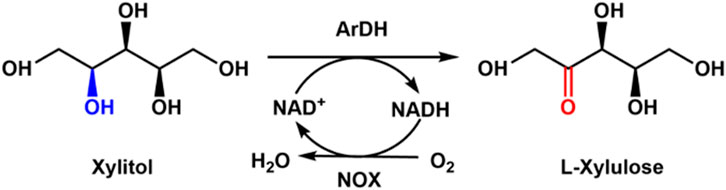
Figure 2. Enzymatic preparation of L-xylulose with ArDH coupled with NOX.
2.3 Enzymatic synthesis of L-gulose with mannitol dehydrogenase
L-Gulose is a building block for the preparation of the anticancer drug bleomycin and antiviral agents. The chemical synthesis routes require complex protections and deprotections of the hydroxyl groups, resulting in inconvenience, high cost, and inefficiency. Woodyer et al. constructed recombinant Escherichia coli harboring mannitol dehydrogenase for the production of L-gulose and L-tagatose (Woodyer et al., 2010). The productivities were only 4.6 and 0.9 g/L for L-tagatose and L-gulose, respectively. Later, mannitol dehydrogenase and NADH oxidase were co-expressed in E. coli by using the pACYDuet-1 vector, and the whole cell expressing mannitol dehydrogenase and NADH oxidase was employed to convert D-sorbitol to L-gulose (Figure 3). With the highly efficient cofactor regeneration, a volumetric product titer of 5.5 g/L for L-gulose was obtained after the optimization of the reaction conditions, including pH, temperature, substrate concentration, and metal ions.
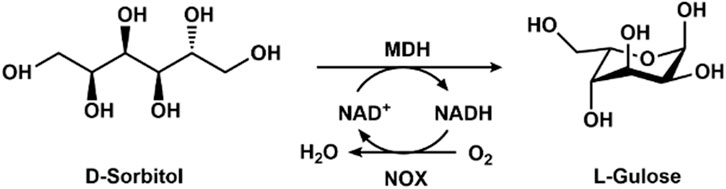
Figure 3. Enzymatic synthesis of L-gulose with mannitol dehydrogenase (MDH) and NOX.
2.4 Production of L-sorbose with sorbitol dehydrogenase
L-Sorbose is an intermediate for the synthesis of L-ascorbic acid, and could be used as the starting material to synthesize the glycosidase inhibitor 1-deoxygalactonojirim and other rare sugars like L--tagatose and L-iditol. Kim et al. (2016) cloned, characterized, and immobilized a highly efficient sorbitol dehydrogenase from Gluconobacter oxydans G624, which showed higher activity toward D-sorbitol. However, the sorbitol dehydrogenase was inhibited by NADPH; thus, it is necessary to lower the cofactor concentration in the reaction system. They then co-expressed sorbitol dehydrogenase and NADPH oxidase in E. coli for the production of L-sorbose (Kim et al., 2019). After optimization of the reaction conditions, a high yield of 92% was achieved by using the whole-cell catalysts (Figure 4). The L-sorbose production was 4.1 g/L, which was 20.5-fold higher than that of E. coli without NADPH oxidase for the cofactor regeneration. Later, they discovered a novel NADPH oxidase which could oxidize both NADH and NADPH (Gao et al., 2019), and the L-sorbose production could be improved 53-fold with this enzyme. One drawback of the sorbitol dehydrogenase is that the working pH range (9.0–10.5) is much higher than the optimal pH of NADPH oxidase.
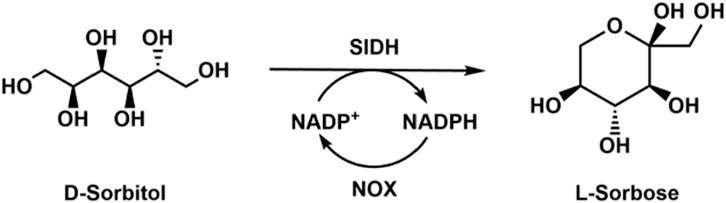
Figure 4. Enzymatic conversion of D-sorbitol to produce L-sorbose with sorbitol dehydrogenase (SlDH) and NOX.
3 Enzymatic production of ketone with dehydrogenase coupled cofactor regeneration
Alcohol dehydrogenase (ADH) is the main enzyme catalyzing the oxidation of alcohol substrate to ketone product. The typical dehydrogenation reaction involves the oxidation of alcohol with the oxidized NAD(P)+ as the electron acceptor. A practical challenge in applying the ADHs for the oxidation of alcohol is the consumption of NAD(P)+. Therefore, it is feasible to recycle the cofactor NAD(P)+ in the ADH-catalyzed oxidation of alcohol to the synthesis of ketone.
3.1 Enzymatic synthesis of acetoin with acetaldehyde lyase (FLS)
Acetoin has been used in the food and pharmaceutical industries as a flavor enhancer and precursor for alkaloid ligustrazine (Peng et al., 2020). It could be produced with chemical reduction of diacetyl, fermentation from microbial cells, or enzymatic production from ethanol (Li et al., 2022a). Although some strains, including Enterobacter, Klebsiella, Serratia, Saccharomyces cerevisiae, and Bacillus could be utilized for the production of acetoin (Li et al., 2022a), the yield was low because acetoin is an intermediate in the 2,3-butanediol metabolic pathway. By deleting the butanediol dehydrogenase encoding gene, which converts acetoin to 2,3-butanediol, and overexpressing NADH oxidase (NOX) to enhance the cofactor regeneration, the final yield of 100.1 g/L was achieved (Bae et al., 2016). Later, Guo et al. co-expressed butanediol dehydrogenase, hemoglobin, and NADH oxidase in E. coli for the production of acetoin, and a product titer of 86.74 g/L was obtained. To overcome the complex regulation of cell growth and purification of acetoin from the fermentation medium, Li et al. (2022a) developed an enzymatic cascade reaction for the production of acetoin. They immobilized ethanol dehydrogenase (EtDH), NOX, and formolase (FLS) onto epoxy-modified magnetic nanoparticles for the synthesis of acetoin (Figure 5). In the cascade enzymatic reactions, EtDH catalyzes the conversion of ethanol to acetaldehyde, which is subsequently transferred to acetoin with the catalysis of formolase. NADH oxidase is responsible for the regeneration of NAD+ in the first enzymatic step. A high conversion of 90% was achieved by using the three immobilized enzymes for one-pot biosynthesis of acetoin, suggesting that this is a promising way to produce value-added acetoin from ethanol.
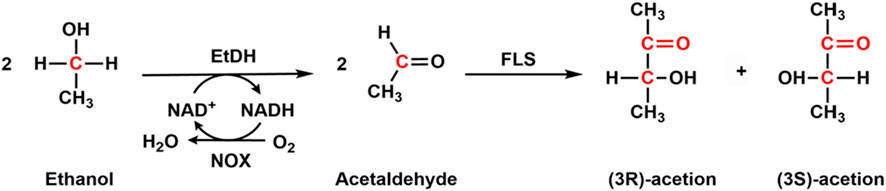
Figure 5. Cascade enzymatic synthesis of acetoin from ethanol with ethanol dehydrogenase (EtDH), NOX, and Formolase (FLS).
3.2 Synthesis of 1,3-dihydroacetone from glycerol with glycerol dehydrogenase
Glycerol is the byproduct of biodiesel and a cheap raw material, and it is economical to convert it to value-added chemicals (Fokum et al., 2021; Li et al., 2023). Fan et al. immobilized whole cells harboring glycerol dehydrogenase (GDH) and NADH oxidase genes through a click reaction (Fan et al., 2021). By employing the immobilized whole cell, glycerol could be converted into 1,3-dihydroxyacetone, around 400 times higher in price. The same group immobilized GDH and Nox for bioconversion of glycerol to 1,3-dihydroxyacetone, and the final product concentration of 3.5 mM was obtained, which was four times higher than that without cofactor regeneration (Figure 6). Then they combined immobilized GDH and NOX with cross-linking to form aggregates for efficient regeneration of NAD+; there was about a 1.5 times improvement compared with the free enzyme system (Xu et al., 2020). By using the combined cross-linked enzyme aggregate, about 4.6% glycerol was converted into 1,3-dihydroxyacetone with a final concentration of 9.4 mM. The combined cross-linked enzyme aggregate could regenerate NADH with a high speed of a turnover number of 2137 in the production of 1,3-dihydroxyacetone.
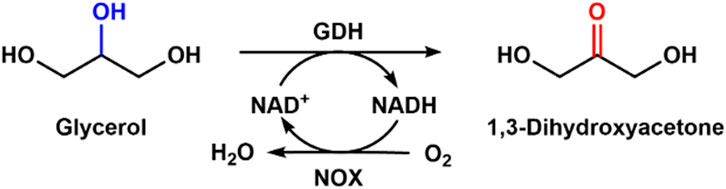
Figure 6. Bioconversion of glycerol with GDH and NOX mixed with NAD+ regeneration.
3.3 Enzymatic production of statin precursor with alcohol dehydrogenase and NADPH oxidase
Generally, an excess of reactive oxygen species will lead to plenty of devastating diseases (Panday et al., 2015). Among these diseases, cardiovascular diseases are the main leading cause of death worldwide. The treatments of the diseases are focused on using drugs such as statins to decrease low-density lipoprotein cholesterol. One attractive approach for the synthesis of statins is the enzymatic route, in which the oxidation of chlorolactol to chlorolactone is the key step. The whole cell co-expressing alcohol dehydrogenase and NADPH oxidase could help to produce chlorolactone (Figure 7), which was performed by co-expressing the two enzymes by Jiao et al. (2016). Another group immobilized alcohol dehydrogenase and NADPH oxidase onto Eupergit CM, Amino-agarose, and epoxy agarose-UAB, and used the immobilized enzymes to catalyze the conversion of chlorolactol, and a yield of 94.7% for chlorolactone was obtained (Garcia-Bofill et al., 2021).
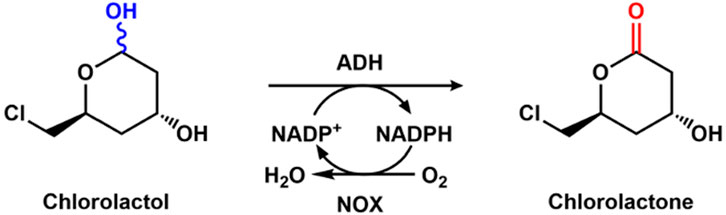
Figure 7. Enzymatic oxidation of chlorolactol to chlorolactone with alcohol dehydrogenase (ADH) and NOX.
3.4 Biosynthesis of 2,5-dimethylpyrazine with L-threonine dehydrogenase
L-Threonine is widely distributed in nature and could be prepared through microbial fermentation, making it a relatively cheap raw material. Thus, the conversion of L-threonine to a more valuable chemical is attractive and engaging for the chemical or pharmaceutical industry. Liu et al. (2023) reconfigured the L-threonine metabolic pathway in Escherichia coli by overexpressing NADH oxidase and L-threonine dehydrogenase (TDH) using the pACYCDuet-1 vector. The whole cell can effectively catalyze L-threonine to L-2-aminoacetoacetic acid, which could be further converted into 2,5-dimethylpyrazine. And the recombinant E. coli overexpressing the two enzymes can convert L-threonine to produce 2,5-dimethylpyrazine (Figure 8), which is a flavor compound and the precursor for some drugs, and a yield of 2.0 g/L was obtained with a molar conversion of 22.2%.

Figure 8. Synthesis of 2,5-dimethylpyrazine using L-threonine as a substrate with TDH.
4 Enzymatic synthesis of vanillic acid with vanillin dehydrogenase
Vanillic acid has been widely used in the pharmaceutical, food, cosmetic, and flavor industries because of its biological activities (Liaqat et al., 2023). Chemical synthesis of vanillic acid has some disadvantages, including low yield and environmental pollution (Ji et al., 2024). The alternative route for vanillic acid is the enzymatic transformation of lignin, which is a greener production method. Xu et al. (2024) cloned and expressed vanillin dehydrogenase (VDH) from Bacillus ligniniphilus L1 and NADH oxidase (NOX) from Streptococcus pyogenes for the biotransformation of vanillin into vanillic acid. They immobilized VDH and NOX to form cross-linked enzyme aggregates (Combi-CLVNAs). The Combi-CLVAs were used to produce vanillic acid (Figure 9). With fed-batch addition of vanillin to the reaction mixture, the production of 44.21 mM vanillic acid was obtained, which was 22.5-fold higher than without NOX in the presence of 2 mM NAD+. After 4 cycles, a total of 149.98 mM vanillic acid was harvested using Combi-CLVNAs. Finally, 38.69 mM vanillic acid was prepared from vanillin-rich extracts derived from lignin depolymerization with combi-CLVNAs. In the whole process using combi-CLVNAs, there were no by-products, which provides a green way for vanillic acid production.
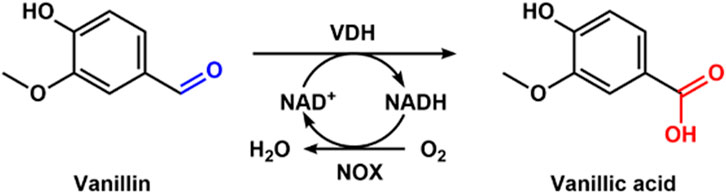
Figure 9. Enzymatic synthesis of vanillic acid with VDH and NOX.
5 Kinetic resolution of racemic phenylethanol with alcohol dehydrogenase
Optically active alcohols are important pharmaceutical or chemical intermediates and could be biosynthesized with alcohol dehydrogenase. However, these enzymes are highly dependent on the oxidized NAD cofactors, which are too expensive to be used stoichiometrically. So, scientists tried to use NADH oxidase to regenerate the cofactor. (R)-alcohol dehydrogenase from Lactobacillus brevis (LbADH) was used to enantioselectively oxidize racemic phenylethanol to produce acetophenone, and another product is (S)-phenylethanol, and NADH oxidase from Lactobacillus sanfranciscensis was employed to reduce O2 to H2O, coupling with the regeneration of cofactor NAD+ (Riebel et al., 2003). The mutant of LbADH G37D prefers NAD+ as the cofactor and is coupled with LsNox to yield 50% conversion of racemic phenylethanol to (S)-phenylethanol and acetophenone. It was found that more than 100 turnovers were obtained depending on the relative concentration of alcohol to cofactor. The regeneration system can also recycle NADP+ with the same NADH oxidase.
Wu et al. (2022) designed and constructed fusion proteins containing alcohol dehydrogenase (ADH) and NADH oxidase for the production of chiral alcohols (Figure 10). The NOX in the fusion proteins exhibited much higher specific activities than that of the individual enzyme, which could regenerate NAD+ in the enantioselective oxidation of racemic 1-phenylethanol. The fusion proteins linked NOX at the N-terminus showed higher turnover numbers than the individual enzyme, especially at low cofactor concentrations.
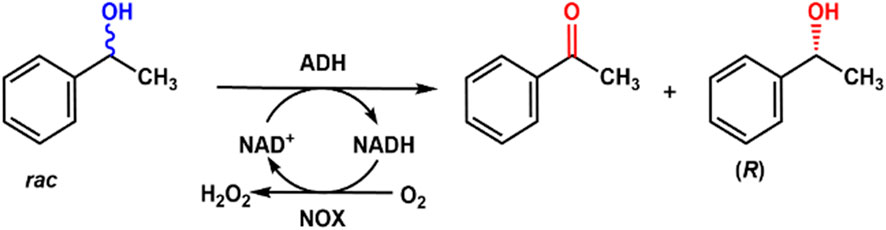
Figure 10. Enantioselective oxidation of racemic phenylethanol with alcohol dehydrogenase (ADH) and NOX.
To expand the application of alcohol dehydrogenase for alcohol oxidation, the cofactor must be recycled to make the process economical because of the stoichiometric consumption of the expensive NADP. Aalbers and Fraaije (2019) designed an artificial alcohol oxidase by fusion of alcohol dehydrogenase and NADPH oxidase for continuous oxidation of alcohol (Figure 11). In this system, a catalytic amount of NADP+ was required, and the purified enzymes could perform a good catalytic efficiency of 69%–99% conversion and 99% ee with racemic phenylethanol as the substrate. Compared with other fusion proteins, the NADH oxidase fusion partner has some advantages and could be a valuable tool for the development of robust catalysts. The NADH oxidase was immobilized for the regeneration of NAD+ in the kinetic resolution of racemic phenylethanol (Rocha-Martín et al., 2011).
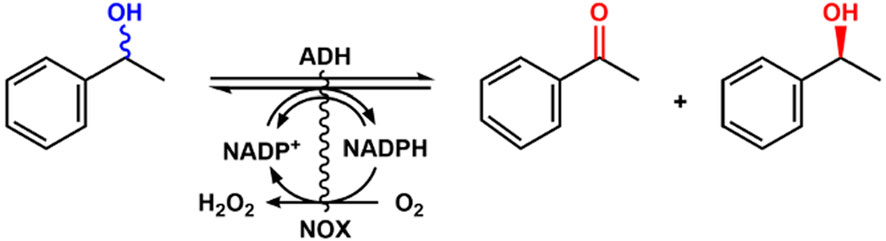
Figure 11. Kinetic resolution of racemic phenylethanol with fused NOX and ADH.
6 Conclusion and future outlook
Now, all the chemical synthesis and transformations are undergoing the shift from chemical methods to biological routes. With the advances of synthetic biology, important enzymes and whole cells are engineered and have great potential applications in pharmaceutical, food, cosmetic, petrochemical, and other industries. Oxidoreductases, as the largest enzyme class, have essential biological functions in life, and will have a huge future market in the industry. However, although the cofactor is expensive, most oxidoreductases need cofactors to transfer electrons, atoms, or chemical groups in a stoichiometric amount. From the industrial aspects, the regeneration of the cofactor will lower the cost and make the preparation of valuable chemicals economical. NAD(P)H oxidases have wide applications in biotechnology and are used as genetic tools to regulate physiological roles of compartment-specific changes in NAD(P)+/NAD(P)H equilibrium in living cells.
Much research work has been done on the discovery, catalytic mechanism, biochemical characterization, and deciphering the structure of NAD(P)H oxidase. It is easy to conclude that value-added chemicals could be prepared with NAD(P)-dependent dehydrogenases, which need the cofactor regeneration with NAD(P)H oxidases. However, there are some biocompatibility issues for the enzymes. It is known that most dehydrogenases have a pH optimum around 8.0–10.0, and some NAD(P)H oxidases are not active in the pH range. For example, the optimal pH of NOX from Lactobacillus rhamnosus is 5.5, which cannot work together with most dehydrogenases (Zhang et al., 2012). To overcome the pH incompatibility, the enzyme was engineered to shift the pH optimum and working pH range by mutation of Asp251 to Arg, and the activity was improved 1.44-fold than the wild-type at pH 7.5 (Zhou et al., 2021). NADPH is more difficult to recycle, and the price of NADPH is higher than that of NADH; So, the NAD(P)H oxidases that could regenerate both NAD+ and NADP+ are more interesting for industrial production. Generally, engineering the surface, catalytic pocket, and the substrate binding domain is a practical approach for enhancing the production of value-added products (Figure 12). These enzymes have achieved notable enhancements in catalytic efficiency, substrate specificity, and operational stability through site-directed evolution and rational design. The dual-substrate NOXs with the ability to switch substrate from NADH to NADPH open new avenues for their industrial applications in the biosynthesis of high-value chemicals. Furthermore, the adaptation of these enzymes to varying pH and temperature conditions is crucial for their integration into industrial processes, ensuring compatibility and efficiency. Discovering new enzymes with high specific activity on both substrates would benefit the applications (Gao et al., 2019; Li et al., 2018), or engineering the enzyme protein to switch the substrate specificity by mutation of Leu179 to Ser (Li et al., 2019). There was a report that engineered NADH oxidase for the regeneration of oxidized non-natural cofactor nicotinamide cytosine dinucleotide (NCD), which could expand the development of NCD-linked redox system (Wang et al., 2025). There is no doubt that more and more NAD(P)-dependent dehydrogenases will be applied in industry with the regeneration of NAD(P)+ by NAD(P)H oxidase. The synergy achieved by coupling engineered NADH oxidase with other enzymes in biocatalytic systems has demonstrated remarkable improvements in product yield and reaction efficiency.
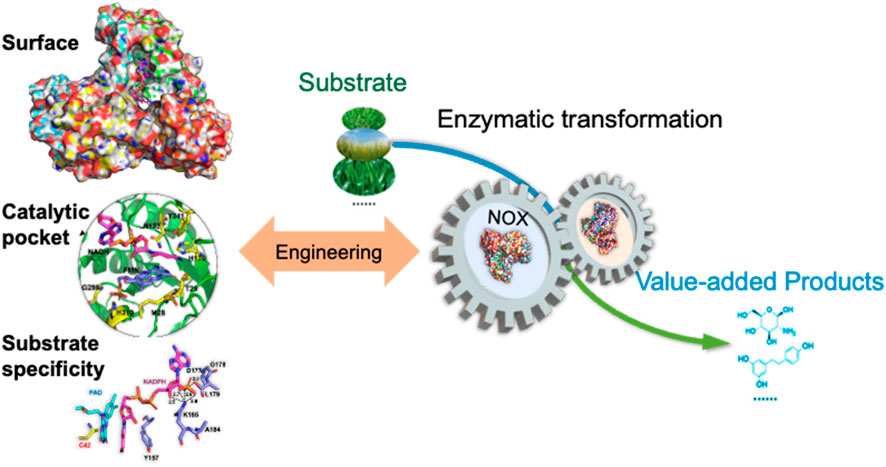
Figure 12. Protein engineering of the surface, catalytic pocket, and substrate binding domain to improve the catalytic activity of NOX to enhance the production of value-added chemicals.
NAD(P)H oxidases catalyze the oxidation of NAD(P)H to regenerate NAD(P)+, in which the product is the oxidized form of the chemicals. The contrary reaction for the production of reduced forms of chemicals to form a chiral center is attracting more attention because of the sharply increasing demand for enzymatic transformations. To regenerate the reduced form NAD(P)H, many enzymes, including glucose dehydrogenase (Zhu et al., 2019), formate dehydrogenase (Wu et al., 2025), and phosphite dehydrogenase (Zhang et al., 2022) have been employed. However, to drive the reaction to the product, a second substrate and product must be introduced for the regeneration enzyme, which will complicate the separation and purification of the products. Recycling or regenerating the reduced form NAD(P)H of the cofactor without introducing by-products is a challenge for enzymologists and chemists.
Author contributions
L-JZ: Writing – review and editing, Conceptualization. S-XL: Writing – original draft. TH: Writing – original draft. YW: Writing – original draft. Y-WZ: Writing – review and editing, Conceptualization, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors appreciated the financial support from the Elderly Health Research Project of Jiangsu Provincial Health Commission (No. LKM2022083).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
ADH, alcohol dehydrogenase; ArDH, arabinitol dehydrogenase; EtDH, ethanol dehydrogenase; FLS, formolase; GDH, glycerol dehydrogenase; GatDH, Galactitol dehydrogenase; MDH, mannitol dehydrogenase; SlDH, sorbitol dehydrogenase; TDH-L-threonine dehydrogenase; NAD, Nicotinamide adenine dinucleotide; NOX, NADH oxidase/NADPH oxidase; ROS, reactive oxygen species; VDH, vanillin dehydrogenase.
References
Aalbers, F. S., and Fraaije, M. W. (2019). Design of artificial alcohol oxidases: alcohol dehydrogenase-NADPH oxidase fusions for continuous oxidations. ChemBioChem 20, 51–56. doi:10.1002/cbic.201800421
Bae, S.-J., Kim, S., and Hahn, J.-S. (2016). Efficient production of acetoin in Saccharomyces cerevisiae by disruption of 2,3-butanediol dehydrogenase and expression of NADH oxidase. Sci. Rep. 6, 27667. doi:10.1038/srep27667
Fan, X.-M., Shen, J.-J., Xu, Y.-Y., Gao, J., and Zhang, Y.-W. (2021). Metabolic integration of azide functionalized glycan on Escherichia coli cell surface for specific covalent immobilization onto magnetic nanoparticles with click chemistry. Bioresour. Technol. 324, 124689. doi:10.1016/j.biortech.2021.124689
Fokum, E., Zabed, H. M., Ravikumar, Y., Elshobary, M. E., Chandankere, R., Zhang, Y., et al. (2021). Co-fermentation of glycerol and sugars by Clostridium beijerinckii: enhancing the biosynthesis of 1,3-propanediol. Food Biosci. 41, 101028. doi:10.1016/j.fbio.2021.101028
Gao, H., Kim, I.-W., Choi, J.-H., Khera, E., Wen, F., and Lee, J.-K. (2015). Repeated production of L-xylulose by an immobilized whole-cell biocatalyst harboring L-arabinitol dehydrogenase coupled with an NAD+ regeneration system. Biochem. Eng. J. 96, 23–28. doi:10.1016/j.bej.2014.12.017
Gao, H., Khera, E., Lee, J.-K., and Wen, F. (2016a). Immobilization of multi-biocatalysts in alginate beads for cofactor regeneration and improved reusability. J. Vis. Exp., e53944. doi:10.3791/53944
Gao, H., Kim, T.-S., Mardina, P., Zhou, P., Wen, F., and Lee, J.-K. (2016b). Rare sugar production by coupling of NADH oxidase and L-arabinitol dehydrogenase. RSC Adv. 6, 66609–66616. doi:10.1039/c6ra11614k
Gao, H., Li, J., Sivakumar, D., Kim, T.-S., Patel, S. K. S., Kalia, V. C., et al. (2019). NADH oxidase from Lactobacillus reuteri: a versatile enzyme for oxidized cofactor regeneration. Int. J. Biol. Macromol. 123, 629–636. doi:10.1016/j.ijbiomac.2018.11.096
Garcia-Bofill, M., Sutton, P. W., Guillen, M., and Alvaro, G. (2021). Enzymatic synthesis of a statin precursor by immobilised alcohol dehydrogenase with NADPH oxidase as cofactor regeneration system. Appl. Catal. A-Gen. 609, 117909. doi:10.1016/j.apcata.2020.117909
Ji, T., Liaqat, F., Khazi, M. I., Liaqat, N., Nawaz, M. Z., and Zhu, D. (2024). Lignin biotransformation: advances in enzymatic valorization and bioproduction strategies. Industrial Crops Prod. 216, 118759. doi:10.1016/j.indcrop.2024.118759
Jiao, X.-C., Zhang, Y.-J., Chen, Q., Pan, J., and Xu, J.-H. (2016). A green-by-design system for efficient bio-oxidation of an unnatural hexapyranose into chiral lactone for building statin side-chains. Catal. Sci. Technol. 6, 7094–7100. doi:10.1039/C6CY01085G
Kim, T.-S., Patel, S. K. S., Selvaraj, C., Jung, W.-S., Pan, C.-H., Kang, Y. C., et al. (2016). A highly efficient sorbitol dehydrogenase from Gluconobacter oxydans G624 and improvement of its stability through immobilization. Sci. Rep. 6, 33438. doi:10.1038/srep33438
Kim, T.-S., Gao, H., Li, J., Kalia, V. C., Muthusamy, K., Sohng, J. K., et al. (2019). Overcoming NADPH product inhibition improves D-sorbitol conversion to L-sorbose. Sci. Rep. 9, 815. doi:10.1038/s41598-018-37401-0
Li, F.-L., Shi, Y., Zhang, J.-X., Gao, J., and Zhang, Y.-W. (2018). Cloning, expression, characterization and homology modeling of a novel water-forming NADH oxidase from Streptococcus mutans ATCC 25175. Int. J. Biol. Macromol. 113, 1073–1079. doi:10.1016/j.ijbiomac.2018.03.016
Li, F.-L., Zhou, Q., Wei, W., Gao, J., and Zhang, Y.-W. (2019). Switching the substrate specificity from NADH to NADPH by a single mutation of NADH oxidase from Lactobacillus rhamnosus. Int. J. Biol. Macromol. 135, 328–336. doi:10.1016/j.ijbiomac.2019.05.146
Li, X.-Y., Huang, J.-Y., Zhou, Q., Xu, Y.-Y., Prabhu, P., and Zhang, Y.-W. (2022a). Immobilization of alcohol dehydrogenase, acetaldehyde lyase, and NADH oxidase for cascade enzymatic conversion of ethanol to acetoin. Energies 15, 4242. doi:10.3390/en15124242
Li, X.-Y., Xu, M.-Q., Liu, H., Zhou, Q., Gao, J., and Zhang, Y.-W. (2022b). Preparation of combined cross-linked enzyme aggregates containing galactitol dehydrogenase and NADH oxidase for L-tagatose synthesis via in situ cofactor regeneration. Bioprocess. Biosyst. Eng. 45, 353–364. doi:10.1007/s00449-021-02665-w
Li, L., Zhou, Y., Huang, C., Jian, L., Lin, Z., Lin, L., et al. (2023). Insight into the influence of plant oils on the composition of diacylglycerol fabricated by glycerolysis and esterification. Industrial Crops Prod. 204, 117324. doi:10.1016/j.indcrop.2023.117324
Liaqat, F., Xu, L., Khazi, M. I., Ali, S., Rahman, M. U., and Zhu, D. (2023). Extraction, purification, and applications of vanillin: a review of recent advances and challenges. Industrial Crops Prod. 204, 117372. doi:10.1016/j.indcrop.2023.117372
Liu, X.-X., Xi, X.-Q., Shen, T.-S., Shi, H.-L., Zhang, Y.-J., Kan, Y.-C., et al. (2023). Enhancing biosynthesis efficiency of 2,5-dimethylpyrazine by overexpressing L-threonine dehydrogenase and NADH oxidase in Escherichia coli. Mol. Catal. 550, 113550. doi:10.1016/j.mcat.2023.113550
Lu, C., Chen, Z., Ravikumar, Y., Zhang, G., Tang, X., Zhang, Y., et al. (2024). Improving catalytic efficiency of L-arabinose isomerase from Lactobacillus plantarum CY6 towards D-galactose by molecular modification. Foods 13, 1727. doi:10.3390/foods13111727
Mahmood, S., Iqbal, M. W., Zhang, W., and Mu, W. (2021). A review on L-ribose isomerases for the biocatalytic production of L-ribose and L-ribulose. Food Res. Int. 145, 110409. doi:10.1016/j.foodres.2021.110409
Panday, A., Sahoo, M. K., Osorio, D., and Batra, S. (2015). NADPH oxidases: an overview from structure to innate immunity-associated pathologies. Cell. Mol. Immunol. 12, 5–23. doi:10.1038/cmi.2014.89
Patel, S. K. S., Otari, S. V., Kang, Y. C., and Lee, J.-K. (2017). Protein-inorganic hybrid system for efficient his-tagged enzymes immobilization and its application in L-xylulose production. RSC Adv. 7, 3488–3494. doi:10.1039/c6ra24404a
Patel, S. K. S., Gupta, R. K., Karuppanan, K. K., Kim, I.-W., and Lee, J.-K. (2024). Sequential Co-immobilization of enzymes on magnetic nanoparticles for efficient L-xylulose production. Int. J. Mol. Sci. 25, 2746. doi:10.3390/ijms25052746
Peng, K., Guo, D., Lou, Q., Lu, X., Cheng, J., Qiao, J., et al. (2020). Synthesis of ligustrazine from acetaldehyde by a combined biological–chemical approach. ACS Synth. Biol. 9, 2902–2908. doi:10.1021/acssynbio.0c00113
Ravikumar, Y., Ponpandian, L. N., Zhang, G., Yun, J., and Qi, X. (2021). Harnessing -arabinose isomerase for biological production of -tagatose: recent advances and its applications. Trends Food Sci. and Technol. 107, 16–30. doi:10.1016/j.tifs.2020.11.020
Ravikumar, Y., Razack, S. A., Ponpandian, L. N., Zhang, G., Yun, J., Huang, J., et al. (2022). Microbial hosts for production of D-arabitol: current state-of-art and future prospects. Trends Food Sci. and Technol. 120, 100–110. doi:10.1016/j.tifs.2021.12.029
Riebel, B. R., Gibbs, P. R., Wellborn, W. B., and Bommarius, A. S. (2003). Cofactor regeneration of both NAD+ from NADH and NADP+ from NADPH:NADH oxidase from Lactobacillus sanfranciscensis. Adv. Synth. Catal. 345, 707–712. doi:10.1002/adsc.200303039
Rocha-Martín, J., Vega, D., Bolivar, J. M., Godoy, C. A., Hidalgo, A., Berenguer, J., et al. (2011). New biotechnological perspectives of a NADH oxidase variant from Thermus thermophilus HB27 as NAD+-recycling enzyme. BMC Biotechnol. 11, 101. doi:10.1186/1472-6750-11-101
Stolarczyk, K., Rogalski, J., and Bilewicz, R. (2020). NAD(P)-dependent glucose dehydrogenase: applications for biosensors, bioelectrodes, and biofuel cells. Bioelectrochemistry 135, 107574. doi:10.1016/j.bioelechem.2020.107574
Su, W.-B., Li, F.-L., Li, X.-Y., Fan, X.-M., Liu, R.-J., and Zhang, Y.-W. (2021). Using galactitol dehydrogenase coupled with water-forming NADH oxidase for efficient enzymatic synthesis of L-tagatose. New Biotech. 62, 18–25. doi:10.1016/j.nbt.2021.01.003
Tang, X., An, Y., Iqbal, M. W., Cong, H., Zhang, G., Zhang, Y., et al. (2022). The characterization of a novel D-allulose 3-epimerase from Blautia produca and its application in D-allulose production. Foods 11, 3225. doi:10.3390/foods11203225
Tesfay, M. A., Win, X., Lin, H., Liu, Y., Li, C., Lin, J., et al. (2021). Efficient L-xylulose production using whole-cell biocatalyst with NAD+ regeneration system through co-expression of xylitol dehydrogenase and NADH oxidase in Escherichia coli. Biochem. Eng. J. 175, 108137. doi:10.1016/j.bej.2021.108137
Wang, X., Liu, Y., Hu, Y., Huang, Y., Zhang, L., Xue, H., et al. (2025). Engineering nicotinamide adenine dinucleotide oxidase for regeneration of oxidized non-natural cofactor. ChemBioChem e202500254, 2500254. doi:10.1002/cbic.202500254
Woodyer, R. D., Christ, T. N., and Deweese, K. A. (2010). Single-step bioconversion for the preparation of L-gulose and L-galactose. Carbohydr. Res. 345, 363–368. doi:10.1016/j.carres.2009.11.023
Wu, X., Zhang, C., Xing, X.-H., Yun, Z., Zhao, L., and Wu, Q. (2022). Construction and characterization of novel bifunctional fusion proteins composed of alcohol dehydrogenase and NADH oxidase with efficient oxidized cofactor regeneration. Biotechnol. Appl. Biochem. 69, 1535–1544. doi:10.1002/bab.2225
Wu, J., Nuylert, A., Iwaki, M., Miki, S., and Asano, Y. (2025). Study on the asymmetric synthesis of chiral 3,3,3-trifluoro-2-hydroxypropanoic acids by lactate dehydrogenase. ChemBioChem 26, e202500047. doi:10.1002/cbic.202500047
Xu, M.-Q., Li, F.-L., Yu, W.-Q., Li, R.-F., and Zhang, Y.-W. (2020). Combined cross-linked enzyme aggregates of glycerol dehydrogenase and NADH oxidase for high efficiency in situ NAD+ regeneration. Int. J. Biol. Macromol. 144, 1013–1021. doi:10.1016/j.ijbiomac.2019.09.178
Xu, L., Sethupathy, S., Liang, Z., Zhuang, Z., Zhang, Y., Sun, J., et al. (2024). NAD+ regeneration-coupled enzymatic bioconversion of lignin-derived vanillin into vanillic acid: a cleaner production approach. Industrial Crops Prod. 222, 119921. doi:10.1016/j.indcrop.2024.119921
Yuan, J., Ravikumar, Y., Zhang, G., Yun, J., Zhang, Y., Zabed, H. M., et al. (2021). L-arabinose isomerase from Lactobacillus parabuchneri and its whole cell biocatalytic application in D-tagatose biosynthesis from D-galactose. Food Biosci. 41, 101034. doi:10.1016/j.fbio.2021.101034
Zhang, Y.-W., Tiwari, M. K., Gao, H., Dhiman, S. S., Jeya, M., and Lee, J.-K. (2012). Cloning and characterization of a thermostable H2O-forming NADH oxidase from Lactobacillus rhamnosus. Enzyme Microb. Technol. 50, 255–262. doi:10.1016/j.enzmictec.2012.01.009
Zhang, L., King, E., Black, W. B., Heckmann, C. M., Wolder, A., Cui, Y., et al. (2022). Directed evolution of phosphite dehydrogenase to cycle noncanonical redox cofactors via universal growth selection platform. Nat. Commun. 13, 5021. doi:10.1038/s41467-022-32727-w
Zhou, Q., Gao, J., and Zhang, Y.-W. (2021). Optimal pH shift of the NADH oxidase from Lactobacillus rhamnosus with a single mutation. Biotechnol. Lett. 43, 1413–1420. doi:10.1007/s10529-021-03129-7
Zhu, Y.-H., Liu, C.-Y., Cai, S., Guo, L.-B., Kim, I.-W., Kalia, V. C., et al. (2019). Cloning, expression and characterization of a highly active alcohol dehydrogenase for production of ethyl (S)-4-Chloro-3-Hydroxybutyrate. Indian J. Microbiol. 59, 225–233. doi:10.1007/s12088-019-00795-0
Keywords: NADH oxidase, cofactor regeneration, biocatalysis, cascade enzymatic reactions, pharmaceuticals
Citation: Zhou L-J, Long S-X, Huan T, Wu Y and Zhang Y-W (2025) Regeneration of cofactor NAD(P)+ with NAD(P)H oxidase for the production of value-added chemicals. Front. Bioeng. Biotechnol. 13:1650600. doi: 10.3389/fbioe.2025.1650600
Received: 20 June 2025; Accepted: 21 August 2025;
Published: 01 September 2025.
Edited by:
Jung-Kul Lee, Konkuk University, Republic of KoreaReviewed by:
Shulai Liu, Zhejiang University of Technology, ChinaJeya Marimuthu, National Institute of Ocean Technology, India
Mugesh Sankaranarayanan, Vel Tech Rangarajan Dr.Sagunthala R&D Institute of Science and Technology, India
Hao Dong, Ocean University of China, China
Copyright © 2025 Zhou, Long, Huan, Wu and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ye-Wang Zhang, emhhbmd5ZXdhbmdAdWpzLmVkdS5jbg==
†These authors have contributed equally to this work