 Benjamin Irvine1,2*
Benjamin Irvine1,2*- 1British Institute at Ankara (BIAA), Ankara, Türkiye
- 2Department of Archaeology, Faculty of Humanities and Letters, Bilkent University, Ankara, Türkiye
This study provides a broad overview of human dietary habits and subsistence practices across time in the Aegean World (defined in the East by the coastal littoral of western Anatolia, Crete to the South, and eastern mainland Greece in the West). This was done principally through the collation and examination of previously published stable carbon and nitrogen isotope ratios (δ13C and δ15N values) from human bulk bone collagen. The sites/populations examined in this study date from the Neolithic to the Late Byzantine periods; ca. 6000 BC to the early 16th century AD. Broadly speaking, the stable isotope values demonstrate general consistency diachronically, although a slight and gradual increase in δ13C values over time is observed. The δ15N values are also broadly similar diachronically, with the exception of the Classical (and to a lesser extent Hellenistic and Byzantine) periods which have noticeably higher δ15N values than the preceding and following periods. Interestingly, and perhaps unexpectedly, there are no clearly observable differences between the sub-regions of the Aegean World (i.e., East Aegean, Crete, West Aegean). The observed findings would, therefore, suggest broadly similar dietary habits and subsistence practices in the Aegean World from the Neolithic period onwards, perhaps pointing toward dietary habits and subsistence practices being a further facet of connectivity in the region.
1 Introduction
Stable isotope analysis in bioarchaeology has become an increasingly utilized tool with which to enable the exploration of dietary variability within regional and temporal frameworks. δ13C and δ15N values are the two most commonly employed, and published, stable isotope ratios for investigating dietary habits. The use of stable carbon and nitrogen isotope meta-analysis to examine large-scale regional and pan-regional diachronic changes has, in recent times, become an accepted and effective method with which to examine palaeodietary and subsistence patterns, and changes, on these scales. Stable carbon isotope analysis can be particularly useful for investigating the importance of marine versus terrestrial protein inputs in human diets and the importance of C4 plants/crops (e.g., millet), whilst stable nitrogen isotope analysis provides information about the consumption of marine foods (which tend to have much higher δ15N values than terrestrial foods) as well as the importance of plant vs. animal consumption in human diets (Papathanasiou and Richards, 2015). By combining δ13C and δ15N values with more traditional analyses of dietary practices derived from the same context as the samples (e.g., archaeobotanical and zooarchaeological data), it is possible to evaluate dietary habits, subsistence practices, and agricultural practices interregionally, diachronically, and comparatively. In the past two decades palaeodiet reconstruction and examination incorporating stable isotope analysis at a diverse range (geographically and chronologically) of sites in the eastern Mediterranean has demonstrated its importance and validity as a method of investigation.
Whilst there are several studies examining dietary habits at singular sites or in micro-regions (see Table 1) and some studies have examined dietary habits on a larger regional basis around the Aegean and its geographical sub-regions (e.g., see Bourbou and Garvie-Lok, 2015 and Bourbou et al., 2011 for Byzantine period Greece and Crete; Irvine, 2022 for the greater Near East; Irvine et al., 2019 for Bronze Age Anatolia; Papathanasiou and Richards, 2015 for Greece across time), there have been no studies examining dietary habits diachronically in the Aegean World as a distinct whole. There also have not been any previous studies that have combined the stable isotope values from each corner of the Aegean World in an attempt to compare dietary habits and subsistence practices from the different sub-regions of the Aegean World (i.e., East, West, and Crete). Therefore, it would be an important effort to compare these different regions of the Aegean World. Thus, this study aims to examine human dietary habits and related subsistence practices and agricultural strategies diachronically in the Aegean World, through an examination of previously published δ13C and δ15N values from human bulk bone collagen. In particular, investigating any possible patterns chronologically, as well as interregionally for the macro-region of the Aegean World. Dietary habits and subsistence practices, and variations in them, can also provide information about potential residential mobility and the movement of people (for example, people moving from regions or settlements with different isotopically visible dietary habits and/or subsistence practices toward the end of their life) and food resources. They can also provide information about connectivity on a larger scale, in terms of shared/similar dietary habits, subsistence practices, and agricultural strategies across macro-regions as well as vice-versa (i.e., heterogeneity as a result of differences in these aspects of food acquisition, preparation, and consumption). At EBA II period Bademaǧaci in south west Central Anatolia, for example, a female adult individual was observed to have slightly different δ13C and δ15N values than the rest of the sampled population. It was considered that this could be the result of different dietary habits, either due to intra-population differences, or potentially coming from a different location. Her non-locality in origin was confirmed by δ34S analysis, with her δ34S values being different to the other individuals in the sample population, and significantly outside of the assumed local range (Irvine et al., 2019). Something similar was found at Roman period Ephesus (coastal western Anatolia), with non-local origin being confirmed in a couple of individuals by δ34S values following an observation of differences in their δ13C and δ15N values, relative to the rest of the sampled population (Lösch et al., 2014). At Bronze Age Kaymakçi, inland western Anatolia, more positive δ13C values in some cattle has been suggested to indicate that these animals were managed in a different way, likely coming from further away from the site, and not originally local to the site (Irvine et al., 2025). Further afield, in eastern Syria at Bronze Age Tell Ashara-Terqa, isotopic values (δ15N values in conjunction with 87Sr/86Sr values) have suggested that rather than individuals being non-local, the food they consumed was imported from elsewhere (Sołtysiak, 2019). These few examples, hopefully, help to demonstrate how examining dietary habits in individuals can also, in some instances, provide information about residential mobility and the movement of people and food resources.
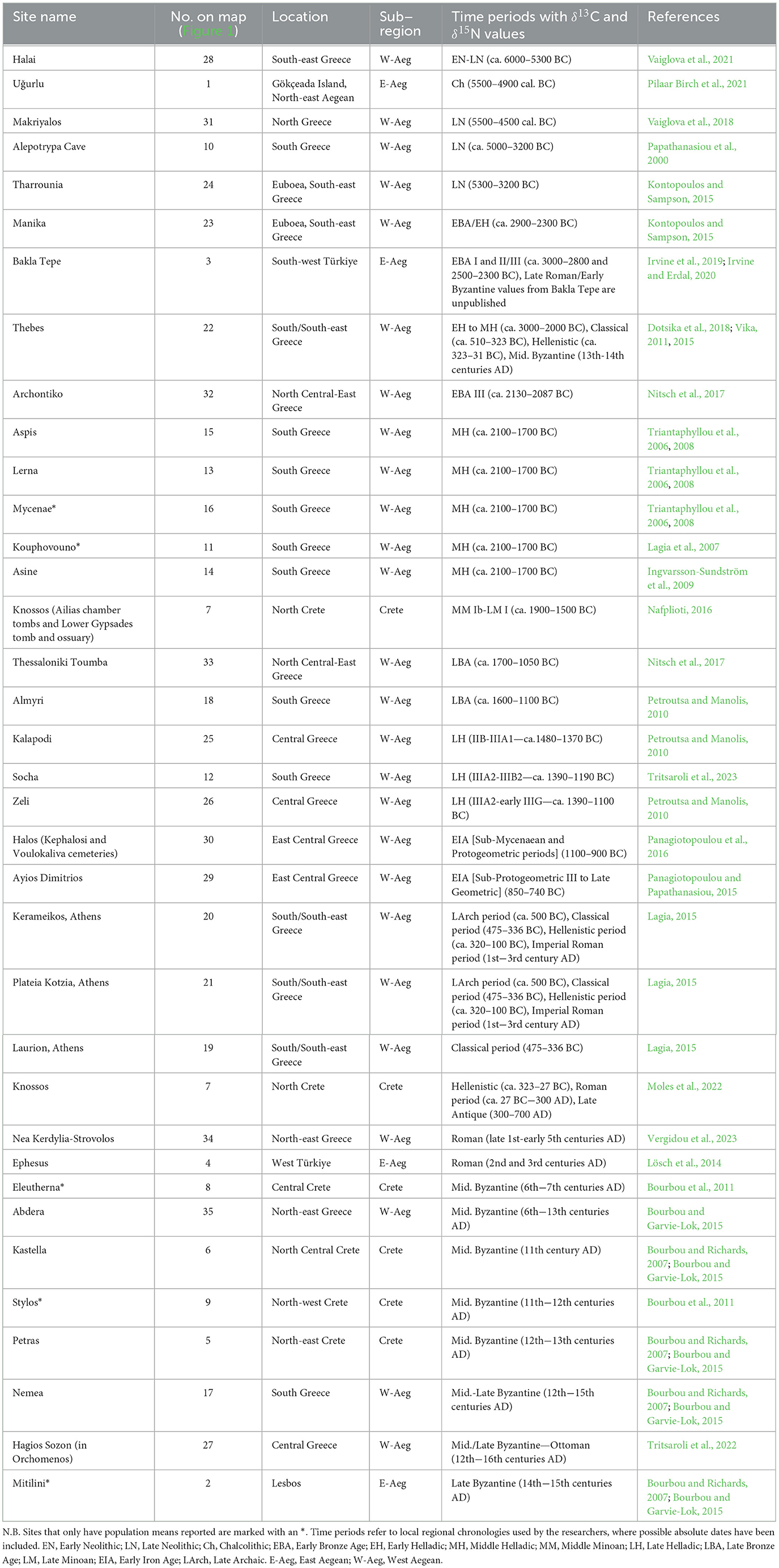
Table 1. Summary overview, in chronological order, of sites with human stable isotope data (δ13C and δ15N values) from the Aegean World.
An important first step is to outline what is meant by the Aegean World, and define its geographical limits—thereby determining where the examined sites come from (see Figure 1). In this study, the Aegean World is defined as including the coastal littoral of western Anatolia, southern Thrace, Crete, the Aegean and Cycladic Islands, and broadly speaking eastern mainland Greece (i.e., Aegean facing)—demarcated as Euboea, Attica, Boeotia, coastal Thessaly, nothing West of Corinth, and everything South and East of Sparta in the Peloponnese. Mycenae may be an exception to this rule, but as it is “culturally” speaking Aegean facing/focused it was included. It has been well established that the Aegean World has been a connected region for millennia—both physically (trade and exchange networks, migrations, etc.) and metaphysically (ideas, technology, cultural and social aspects, etc.). We have evidence for the movement of people (Nafplioti, 2008; Nafplioti et al., 2021; Richards et al., 2022; Skourtanioti et al., 2023), writing (the presence of Linear B script at Late Minoan III Knossos and Armenoi on Crete—Palaima, 2010; cf. Richards et al., 2022), material culture (Rahmstorf, 2015; Şahoǧlu, 2004, 2019; Ünlü, 2016; Ünlüsoy, 2016), and the transference of technological knowledge—including subsistence practices/agro-pastoral strategies (Krauß et al., 2018; Rahmstorf, 2015; Şahoǧlu, 2019) as well as the circulation of raw resources such as Melian obsidian, for example, from as early on as the Mesolithic and Neolithic periods, as well as metals and textiles in later periods (Coleman et al., 2017; Milić, 2014; Rahmstorf, 2015; Şahoǧlu, 2019; Ünlüsoy, 2016). There is also later prehistoric and historic period cultural connectivity and the presence and control in the region of city states/proto-empires/kingdoms as well as the cultural and imperial hegemony of the Roman and Byzantine Empires (Parkinson and Galaty, 2007). Connectivity is also witnessed through the distribution of architectural features (Şahoǧlu, 2004) and socio-cultural phenomena such as burial habits and mortuary practices (Massa and Şahoǧlu, 2011; Şahoǧlu, 2024). Dietary connectivity has previously been proposed for certain periods of the Greek world of the region (Greek mainland, islands, and Crete), with researchers using the phrases “Neolithic diet” and “Byzantine diet” to refer to the exploitation of similar foodstuffs, and broadly tightly clustered human stable carbon and nitrogen ratio values (Papathanasiou and Richards, 2015).
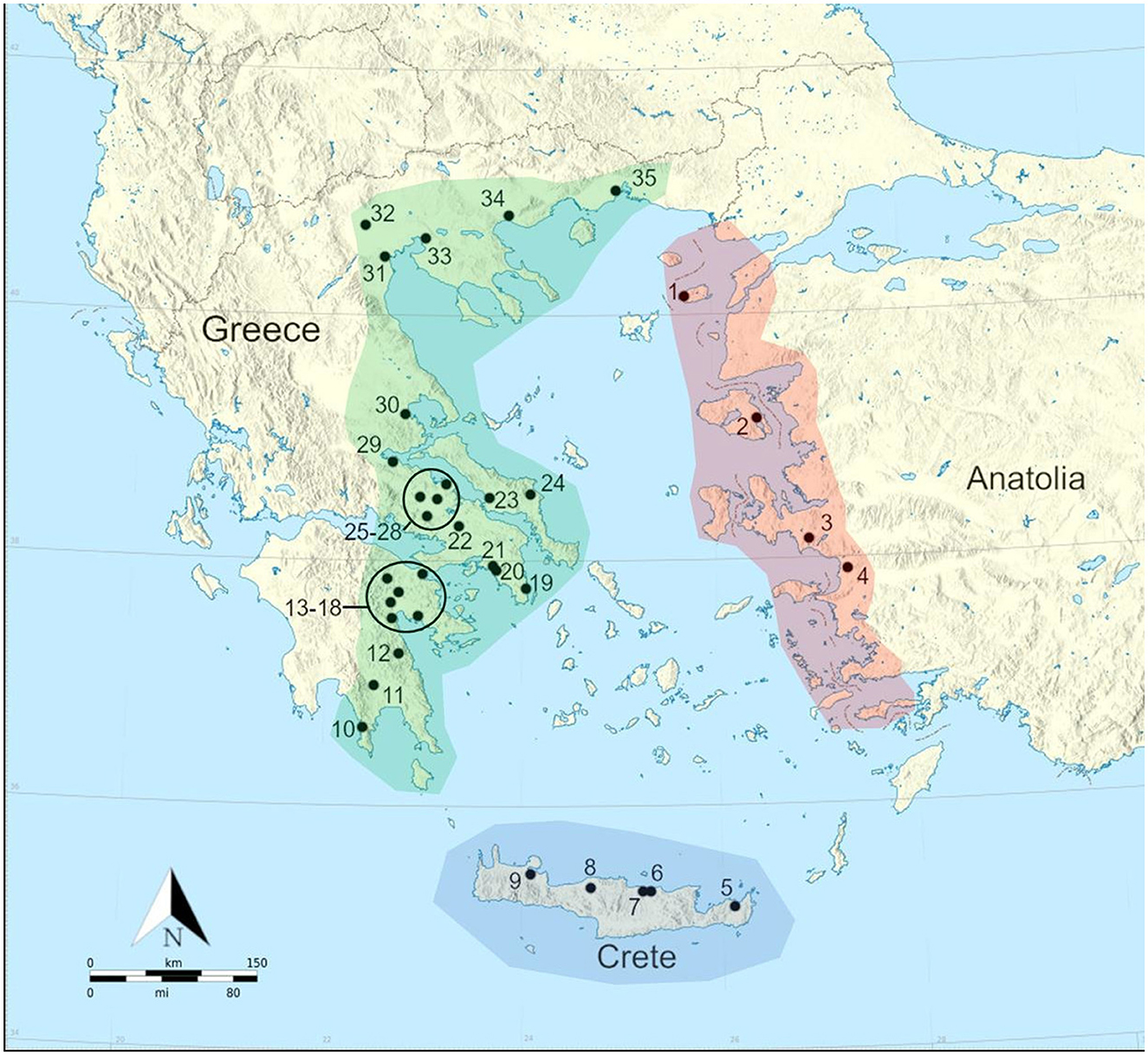
Figure 1. Map of sites with δ13C and δ15N values from bulk bone collagen of adult humans examined in this paper. 1: Uǧurlu; 2: Mitilini; 3: Bakla Tepe; 4: Ephesus: 5: Petras; 6: Kastella; 7: Knossos; 8: Eleutherna; 9: Stylos; 10: Alepotrypa Cave; 11: Kouphovouno; 12: Socha; 13: Lerna; 14: Asine; 15: Aspis; 16: Mycenae; 17: Nemea; 18: Almyri; 19: Laurion; 20: Kerameikos; 21: Plateia Kotzia; 22: Thebes; 23: Manika; 24: Tharrounia; 25: Kalapodi; 26: Zeli; 27: Hagios Sozon; 28; Halai; 29: Ayios Dimitrios; 30: Halos; 31: Makriyalos; 32: Archontiko; 33: Thessaloniki Toumba; 34: Nea Kerdylia-Strovolos; 35: Abdera. The polygons approximately demarcate the different sub-regions defined in this paper: red = East Aegean; blue = Crete; green = West Aegean.
Whilst archaeobotanical and zooarchaeological evidence are vital components to consider when examining dietary habits and subsistence practices, they provide only pieces of the story regarding land use and subsistence (Halstead and Isaakidou, 2020; Papathanasiou, 2015). Archaeological plant and animal remains are considered in this study but only in a basic overview manner, to provide some dietary context (of potentially exploited food resources) for the human isotope values, which this study focuses on. There is an array of archaeobotanical and zooarchaeological reports and publications for sites of the region, as well as some regional summaries. For example, for an excellent summary of farming, land-use, and subsistence in Neolithic Greece, see Halstead and Isaakidou (2020). Papathanasiou (2015) provides a great summary for Neolithic and Bronze Age Greece. Bourbou and Garvie-Lok (2015) also provide a wonderful summary and overview of possible food resources consumed during the Byzantine periods. From the more traditional sources of dietary and subsistence data (i.e., archaeobotanical and zooarchaeological and also documentary and textual sources for the historical periods), it can be said, very generally, that from the Neolithic period onwards the dietary habits of the region appear to have been primarily based on C3 terrestrial resources, cereals and legumes and domesticated animals, particularly sheep/goat (cf. Vika, 2011). It is also understood that not examining stable isotope data from plants and animals in any real depth to provide contextual dietary and subsistence and environmental information is a limitation of this study. However, this kind of data doesn't currently exist on a large-scale (geographical or chronologically) for the region as a whole, or large parts of the sub-regions; something highlighted as a limitation for interpreting human dietary habits in Greece by Papathanasiou and Richards (2015), for example. Although, see Bourbou and Garvie-Lok (2015) for a summary of possible δ13C and δ15N values for Byzantine Greece and Vaiglova et al. (2021) for Neolithic Greece. Furthermore, this paper is intended only to provide a general and broad overview of the larger picture of dietary habits and subsistence practices over time in Aegean World. Additionally, human osteoarchaeological analyses are also important components of understanding past human dietary habits and subsistence practices, but not all osteoarchaeological studies in the region have focused on reconstructing dietary habits and there is not space and time to include these within the scope of this paper, and therefore, they are also not included.
2 Materials and methods
Sites and populations with published human δ13C and δ15N values from the Aegean World, as defined previously in the introduction, were collated and analyzed to examine dietary habits and related subsistence practices across time in past Aegean populations. Namely, sites in the western Anatolian coastal littoral (East Aegean), Crete, and the eastern and southern regions of Greece (West Aegean) (see Figure 1). In terms of chronological periods, the δ13C and δ15N values come from populations with a range of dates from the Neolithic to the Late Byzantine periods; ca. 6000 BC to the early 16th century AD (see Table 1 for an overview of the sites and populations examined in this paper). To ensure inter-population comparability only isotopic values obtained from bulk bone collagen of adult individuals was collated and examined. Only isotopic values from adults were examined to avoid potential issues and biases in the data due to the isotopic effects of breastfeeding, the weaning process, and possible sub-adult diet (which may be different to that of the adults). Furthermore, only δ13C and δ15N values which are within the accepted ranges for collagen quality control (C:N atomic ratio of 2.9–3.6, %C of 10–47%, and %N of 5–17%) were collated for this study to ensure a control of the quality of the examined data (Ambrose, 1990; DeNiro, 1985; van Klinken, 1999).
The isotopic values from the collated and examined populations were also grouped by chronological periods to enable diachronic examination and visualization (Figures 2, 3). These chronological groupings are as follows: Neolithic-Chalcolithic (ca. 6000–3200 BC); Early Bronze Age/Early Helladic (ca. 3000–2000 BC); Middle Bronze Age/Middle Helladic/Middle Minoan (ca. 2000–1700 BC); Late Bronze Age/Late Helladic (ca. 1700–1050 BC); Early Iron Age (ca. 1100–700 BC); Late Archaic-Classical (ca. 500-323 BC); Hellenistic (ca. 323–27 BC); Roman (ca. 27 BC–AD 400); Byzantine/medieval (ca. AD 300–1600). Subsequently these chronological groups were then defined regionally between the East Aegean, Crete, and West Aegean. The mean values (and standard deviations), ranges in values, and minimum and maximum values were then calculated for these chronological groups for δ13C and δ15N, and box and whisker plots were also generated for the chronological groups—for the Aegean World as a whole and by sub-region. The box and whisker plots were created in Excel, and show the median, the mean (the “X”), the upper and lower quartiles (the upper and lower lines of the box which equal the 75th and 25th percentiles, respectively). The whiskers show the 5th and 95th percentiles; outliers are defined as values below the 5th or above the 95th percentiles. However, this was done with some caveats. For example, there are some sites where only the mean values were reported and published (those marked with an * in Table 1), and these were not included when calculating chronological group means or visualized in the plots. However, they will be discussed in relation to the examined data for the relevant chronological periods. Additionally, for examining East Aegean vs. Crete vs. West Aegean, even though some chronological periods for each sub-region are only represented by a single site/population, this was done to allow for regional comparison – with the caveat that this is understood as a potential limitation. Additionally, Early Bronze Age III Archontiko, Late Archaic and Classical Kerameikos, Late Archaic Plateia Kotzia, and Byzantine Abdera were not included in the δ13C and δ15N values box and whisker plots for all sites (Figure 4) as there is only one or two values/individuals, but these were integrated into the data examination and analysis and visualization by chronological and regional groups (Figures 5, 6). There are also δ13C and δ15N values from several Late Neolithic to Late Bronze Age (Late Minoan III) populations on Crete (Richards et al., 2022). However, these were not included in the collated and examined data of this study as in many cases there is no defined chronology or demographic information for distinct individuals due to the comingled nature of the burial contexts where the sampled human skeletal remains came from. However, these values will be incorporated in the discussion.
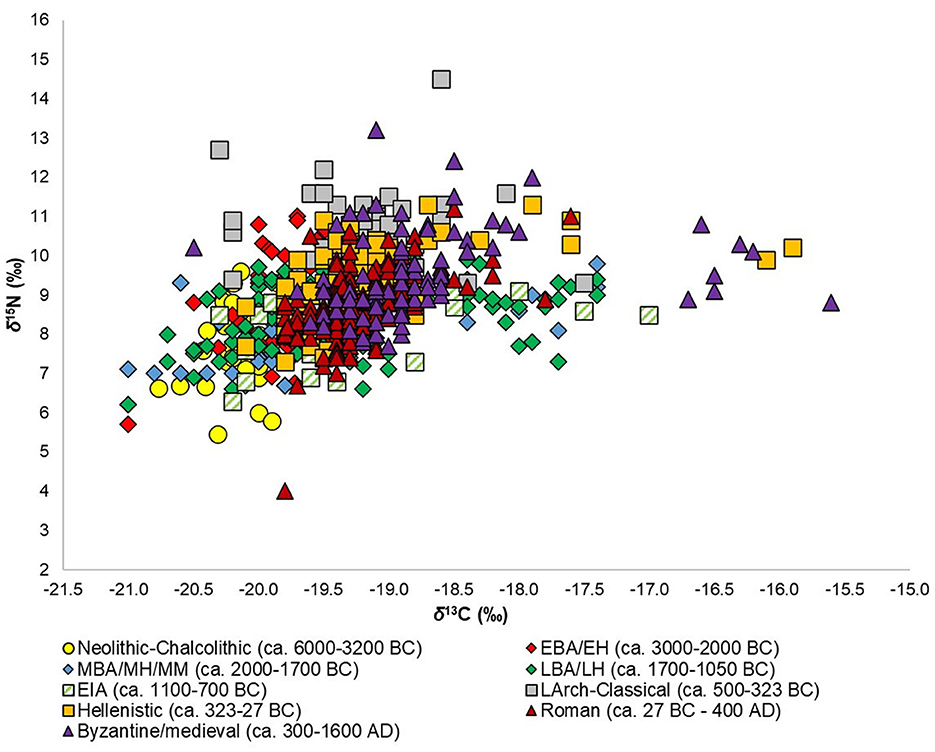
Figure 2. Individual δ13C and δ15N values by chronological group for the entire Aegean World (i.e., from sites and populations examined in this study).
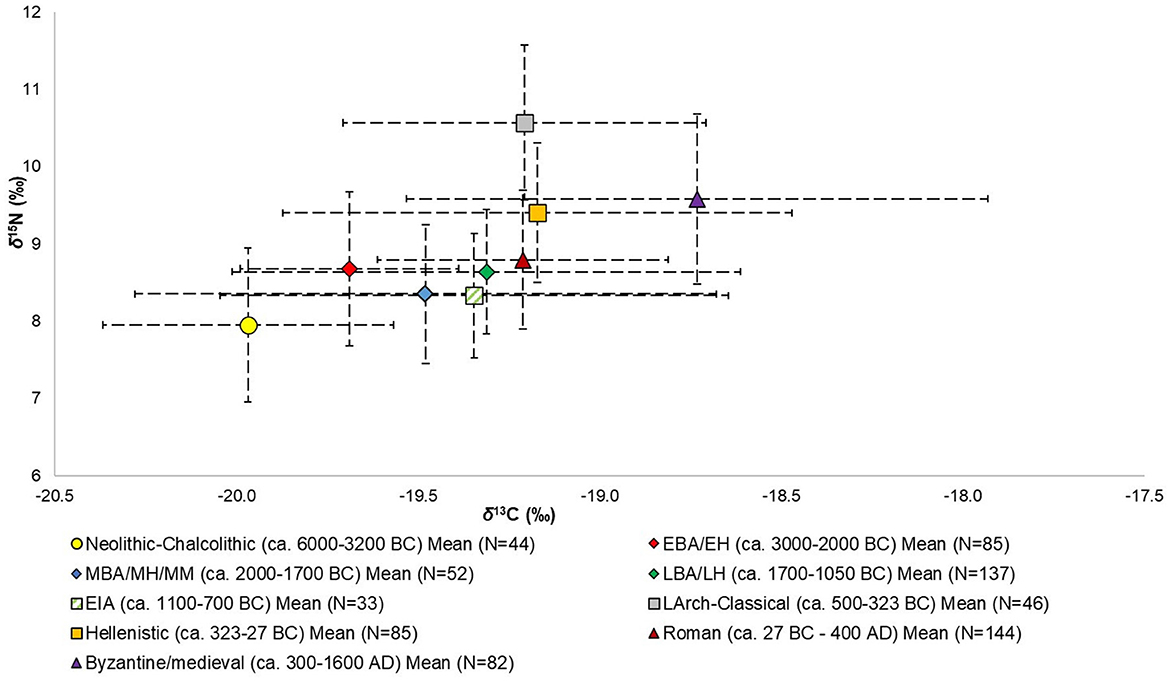
Figure 3. Mean δ13C and δ15N values and standard deviations by chronological group (see Table 2 for values) for the entire Aegean World (i.e., from sites and populations examined in this study).
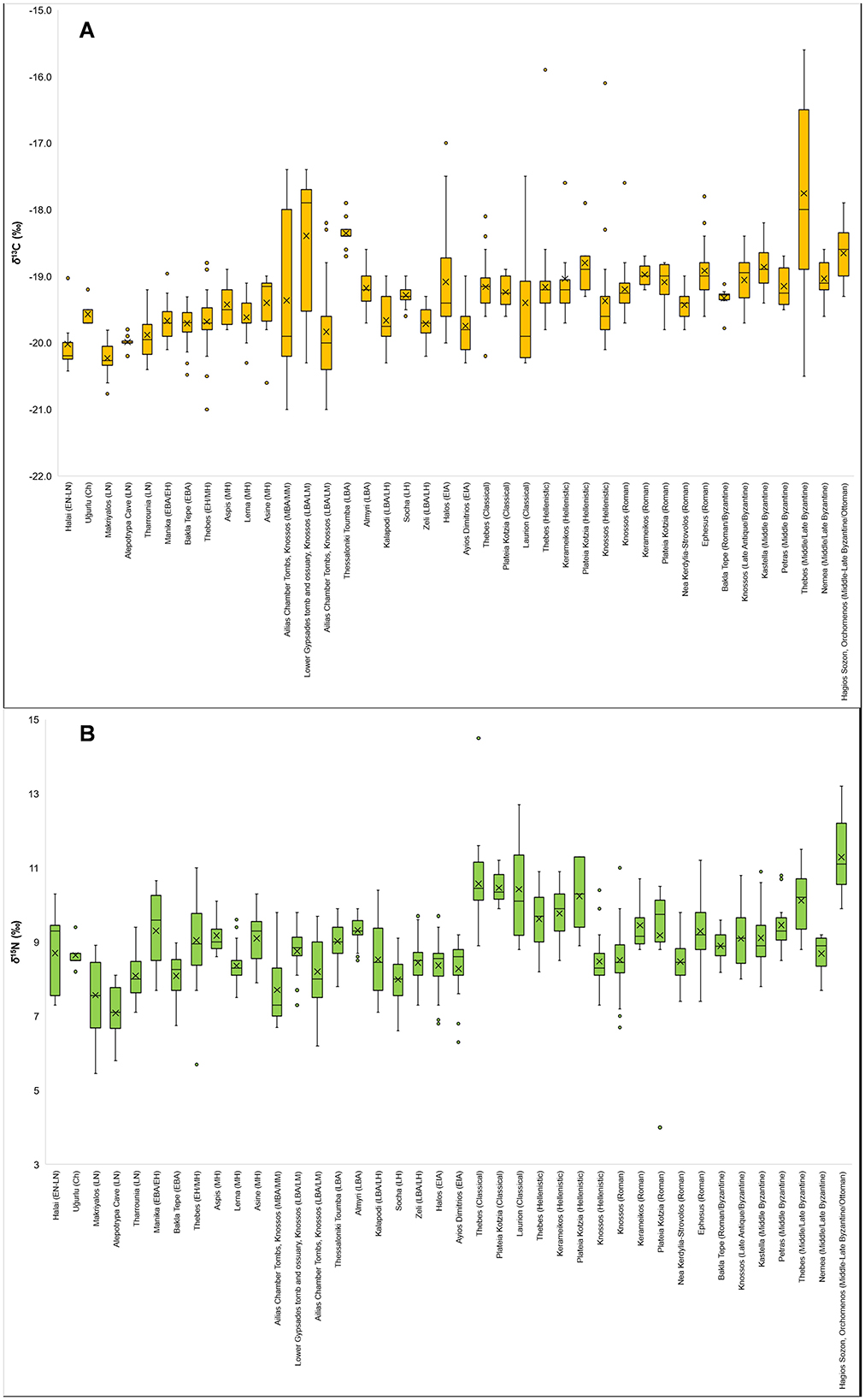
Figure 4. Box and whisker plots for δ13C (A) and δ15N (B) values of all sites examined in this study.
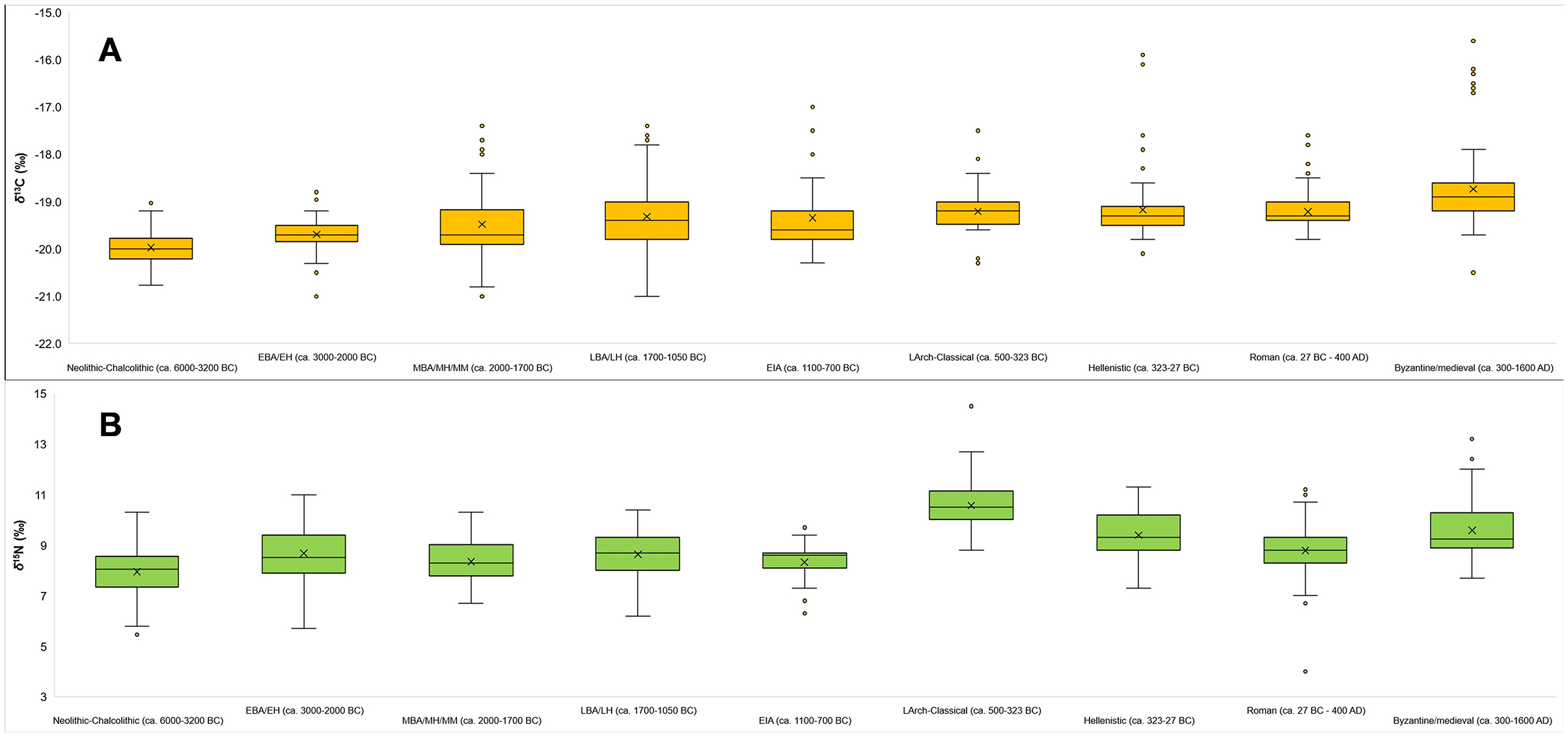
Figure 5. Box and whisker plots for δ13C (A) and δ15N (B) values by chronological group for the entire Aegean World (i.e., from sites and populations examined in this study).
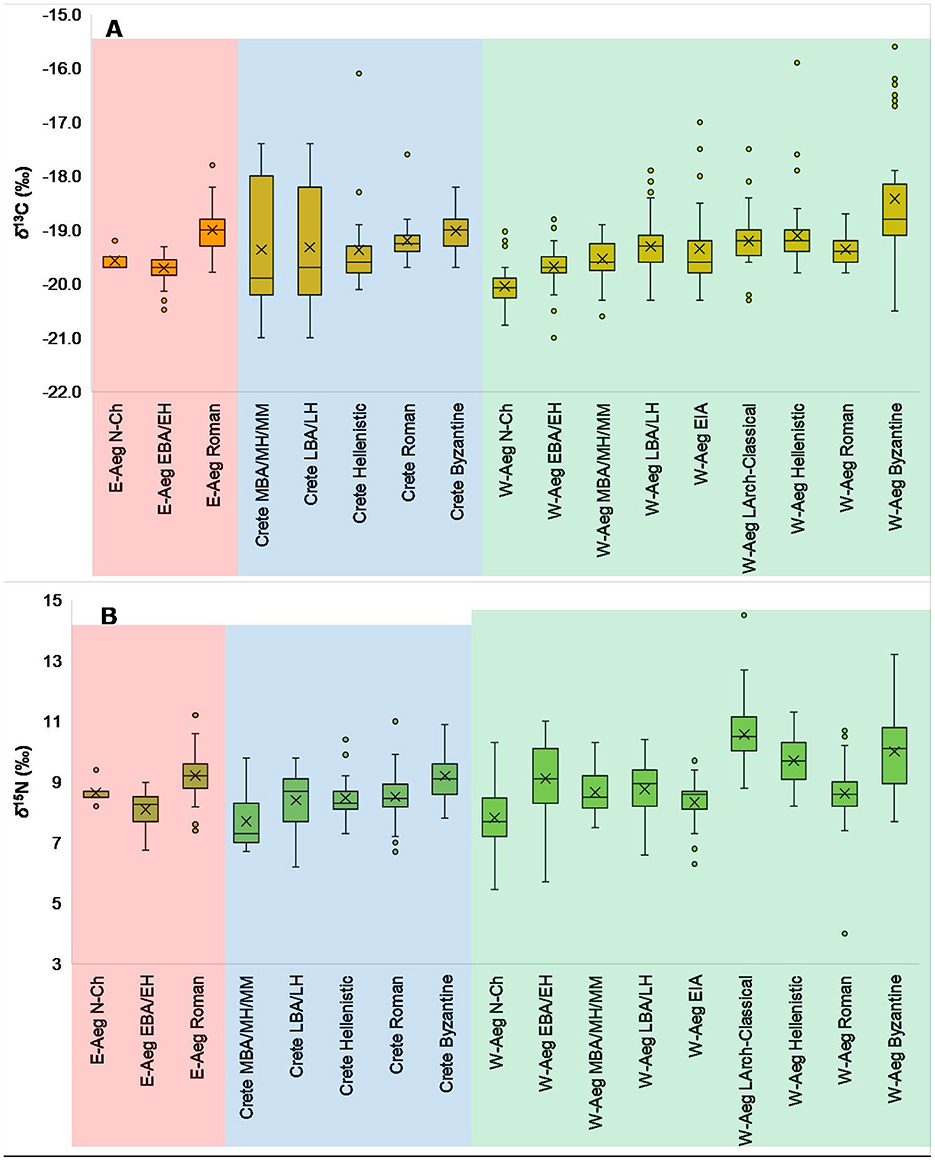
Figure 6. Box and whisker plots for δ13C (A) and δ15N (B) values by sub-region and chronological groupings.
3 Results
About 700 δ13C and δ15N values were examined (corresponding to roughly the same number of individuals), with the highest number belonging to the Roman (N = 144) and Late Bronze Age/Late Helladic (N = 137) periods, and the lowest numbers to the Early Iron Age (EIA) (N = 33), Neolithic-Chalcolithic (N = 44), and Late Archaic-Classical (N = 46) periods. Most of the isotopic values also come from West Aegean sites and populations (72.2%), which needs to be considered when evaluating the data, especially when considering potential patterns and differences between sub-regions (see below). The number of δ13C and δ15N values/individuals as well as the mean values and standard deviations, ranges of values, and minimum and maximum values for δ13C and δ15N by chronological grouping are given in Table 2. The individual δ13C and δ15N values by chronological grouping are visualized in Figure 2 and the means and standard deviations for the chronological groups are visualized in Figure 3.
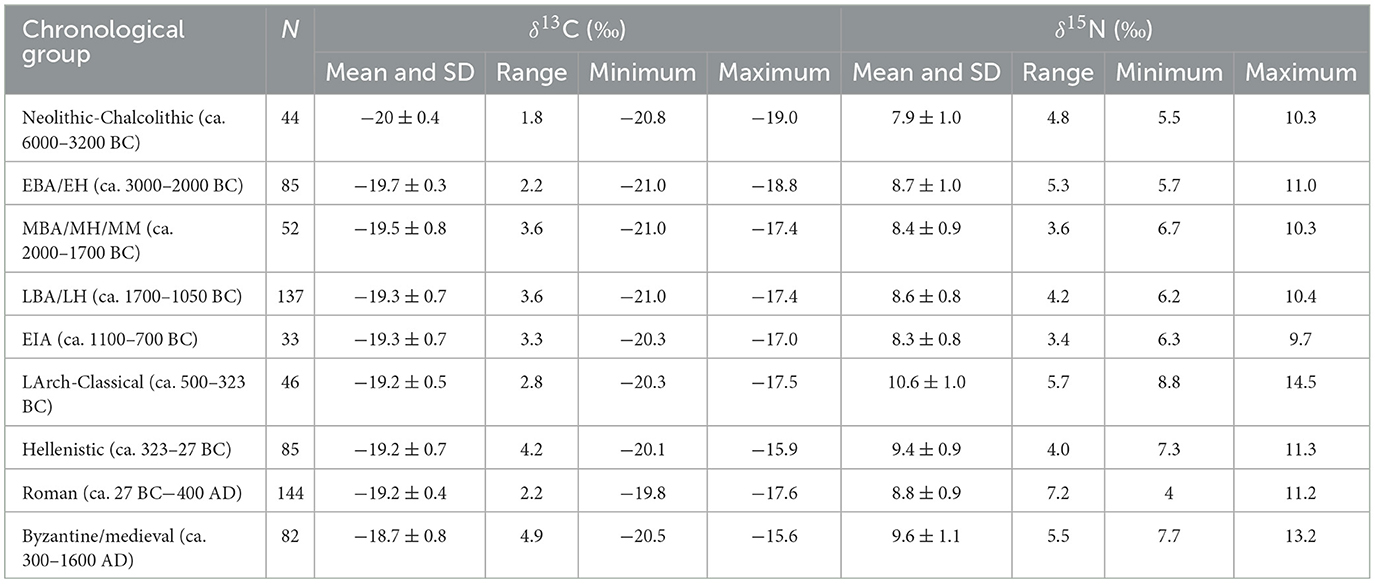
Table 2. Means, standard deviations (SD), ranges, and minimum and maximum δ13C and δ15N values by chronological grouping.
From the data and their visualizations in Figures 2, 3 we can observe that there is a general increase in the mean δ13C values over time from −20.0 ± 0.4‰ in the Neolithic-Chalcolithic to −18.7 ± 0.8‰ in the Byzantine/medieval period. A general increase in the range of δ13C values over time is also observed, with the exception of the Late Archaic-Classical and Roman periods (Table 2). For δ15N values there is more variability and no clear observable patterns over time, although we can perhaps say that after the EIA δ15N values tend to be around or above 9‰ (Table 2; Figures 3, 5). Interestingly, in the Roman period, whilst it has one of the lowest ranges for δ13C values (2.2‰, from −19.8‰ to −17.6‰ —Table 2), it has the greatest range in δ15N values (7.2‰, from 4.0‰ to 11.2‰ — Table 2).
To enable a more nuanced examination of these initial observations, box and whisker plots for δ13C and δ15N values were generated for all sites (Figure 4) and by the chronological groupings (Figure 5)—although see description of exceptions in Section 2. Figure 5 also demonstrates a general gradual increase in δ13C values over time, although actually the majority of δ13C values are generally quite consistent over time and it is rather an increase in diversity from the MBA onwards (i.e., an increase in the range and presence of outlying values) in the values that increases over time. This is particularly observable in Figure 4 when looking at the data on a site-by-site basis. Whilst Figures 4, 5 for δ15N values also demonstrate the aforementioned lack of clear observable patterns and being more irregular, the overall and general picture (Figure 5) for the δ15N values is, again, generally quite consistent over time, with many values being within ca. 3‰ of each other – i.e., one trophic level. The exceptions being the Classical, and, also, to a lesser extent Hellenistic and Byzantine, periods which, as a group, demonstrate a marked overall increase in δ15N values relative to other chronological periods.
Box and whisker plots were also generated for δ13C and δ15N values for the chronological groups, but further organized by sub-region: East Aegean, Crete, and West Aegean (Figure 6). To enable easier visualization and comparative examination these charts are organized from left to right by chronological period and then sub-region. As highlighted earlier, it is understood that there are some limitations here as some sub-regions do not have δ13C and δ15N values for particular chronological periods, and some chronological periods in sub-regions are only represented by single sites. As stated above, another limitation is that the vast majority (72.2%) are from the West Aegean which creates an obvious bias when attempting to compare these sub-regions. However, ultimately, this is the data that is available. It was, therefore, decided to examine the data in this way (whilst understanding and accepting the aforementioned limitations) to see if there any general observable patterns by region and whether there are differences between the regions, or if any observable patterns for the overall Aegean World can be elaborated upon by sub-regional nuances and characteristics. Similar patterns in δ13C values, as observed for the overall macro-region are also observed by sub-region, with a general consistency in (mean) values over time (almost all being around ca. −19.0‰ to −19.5‰, with only a few not—the Neolithic to EBA populations, these having mean values between −19.5‰ and −20.0‰), but with a steady and gradual increase. Furthermore, overall, there appears to be little difference between the regions within chronological periods where comparisons are possible (i.e., at least two of the sub-regions represented per chronological group). An exception to this is obviously Crete in the Middle and Late Bronze Ages where an increase in the ranges and diversity of δ13C values is observed. Additionally, the greater presence of outliers with more positive δ13C values in later periods would possibly imply more diverse variations in these periods. As previously observed, there is, relatively, more irregularity in (mean) δ15N values. But, as also previously noted, with the exception of the Late Archaic-Classical group with markedly greater δ15N values, and also possibly a greater presence of δ15N value outliers in the Roman periods, there is actually an overall and broad consistency in δ15N values for all sub-regions over time.
4 Discussion
4.1 Overall and chronological patterns in the Aegean World
The overall shift in mean δ13C values to more positive values in the later chronological periods, relative to earlier periods, is something that has been observed for the East Mediterranean and Near East as a whole (Irvine, 2022). The cause for this shift in δ13C values has been hypothesized to be related to the deliberate introduction and exploitation of C4 plants in the arable agricultural systems (particularly in post-Middle/Late Bronze Age populations) (Irvine, 2022). Although, this shift in δ13C values over time may also be related to climatic changes affecting the isotopic values of food resources at the base of food webs (i.e., plants) and a general overall pattern of decreasing water availability in the macro-region from the Neolithic period onwards (Araus et al., 2014; Irvine, 2022). Environmental factors, including anthropogenically affected environments, may also be considered, as more positive δ13C values at the base of the food web (i.e., in plants) may be indicative of a less forested and more open environment (Vika, 2011; van Klinken et al., 2000). The same causes are likely to, therefore, also be true for this observed trend in the examined Aegean World populations. The general increase in the range of δ13C values and the presence of a greater number of outliers with more positive δ13C values (Figure 5), most noticeable from the later Bronze Age periods onwards, would suggest, however, that this is perhaps more likely to be related to changing subsistence practices and agricultural strategies. Perhaps as a result of agriculture becoming more extensive (cf. Vaiglova et al., 2020—wheat was less intensively cultivated in the Late Bronze Age than in the Neolithic at Knossos, for example) and an increase in the environmental range of consumed plant and animal resources, as well as the deliberate introduction of C4 crops into the arable repertoire (Vika, 2011)—i.e., there was an increase in the diversity of exploited locales and practices of plant (crop) cultivation and animal management and rearing. It has been suggested, however, that even though there was a presence of C4 plants in the region in later periods, its importance was likely to have been relatively minor—seen as an inferior food, mainly for the poor or in times of desperation—acting more as a supplement to the primary staples of C3 crops (Bourbou and Richards, 2007; Bourbou and Garvie-Lok, 2015; Bourbou et al., 2011). Consequently, we could consider the reasons for this increase in δ13C values over time to be multi-factorial; including pan-regional/large-scale climatic changes, environmental changes and anthropogenically affected environments, the deliberate introduction and exploitation of C4 crops, and also possibly (in certain later periods as will be discussed further below) an increased input of marine resources in dietary habits.
Interestingly, there is a notable decrease in the range of δ13C values in the Roman period, which may suggest a greater standardization and homogeneity of subsistence practices and agricultural strategies, perhaps due to the Imperial hegemony of the region. In the Roman period, zooarchaeological evidence has noted that there are, generally, more pigs in Anatolian assemblages and less diverse husbandry strategies (Slim and Çakirlar, 2023). Concerning animal husbandry practices, at least, this could provide some evidence for less range in the δ13C values—pigs tend not to be herded or managed over large distances and diverse environments, especially if the husbandry strategies were less diverse. However, the Roman period exhibits the greatest range in δ15N values (also observed for all sub-regions of the period, see Figure 6) which may suggest that whilst there may have been a greater homogeneity in subsistence practices and agricultural strategies, dietary habits (particularly related to the relative inputs and consumption of animal protein) were diverse and variable in this period. Whilst there is also a decrease in the range of δ13C values in the Late Archaic-Classical period, it is difficult to accurately assess this as the isotopic values from this period come from sites around Athens and also Thebes, and therefore may reflect more of a local micro-regional pattern. The same may also be true when considering the δ15N values for this chronological period—see also further discussion below about these sites.
The box and whisker plots for each site (Figure 4) and by chronological groupings (Figure 5), however, suggest that whilst there is a gradual increase in δ13C values over time, there is actually a general and broad overall homogeneity. To clarify, there is “noise” and variation around the means, but the mean values at each site and in each time period remain relatively consistent over time. Individual sites (Figure 4) do demonstrate more variability, which would point to variability in isotopic values, and thereby subsistence practices and agricultural strategies, being more pronounced at local micro-regional levels. Generally, a similar pattern is observed for δ15N values, with the Classical, and to a lesser extent Hellenistic and Byzantine, sites and periods providing an exception to this with relatively greater δ15N values—the Classical populations have the greatest δ15N values of all the examined sites and chronological periods (see Figures 4, 5, and also Lagia, 2015). This would suggest that in these periods that people had a greater input of animal protein in their diets. However, the caveat to this is that the sites demonstrating this marked increase in δ15N values in the Classical and Hellenistic periods are the sites around Athens and also Thebes, which may point toward a more local, micro-regional pattern in dietary habits and subsistence practices. Furthermore, at these sites not only do we see an increase in δ15N values, but also a relatively greater range and presence of outliers (at the site level—see Figure 4) of δ13C values. These two combined factors may suggest that in the Classical and Hellenistic periods of this region that there may have been an input of marine resources in the dietary habits of the populations. Indeed, Lagia (2015), who analyzed the Athenian isotopic values suggested that the people likely had a significant input of terrestrial animal protein as well as a contribution of low trophic level fish such as sardines, anchovies, sea-bream, and garum in their diets. Furthermore, a positive correlation between δ13C and δ15N values, further suggesting a contribution of marine resources, was also observed (Lagia, 2015)—as in this study. The high δ15N values at Classical Thebes, alternatively, have been suggested to be the result of freshwater consumption, or possibly more intensive crop cultivation and increased levels of manuring (Vika et al., 2009; Vika, 2011). An interesting aside is that the δ13C and δ15N values of individuals sampled from Grave Circle A at Mycenae demonstrate evidence of some marine resource consumption and a greater input of animal protein in their dietary habits (Triantaphyllou et al., 2006, 2008). Although it should be noted that Grave Circle A is likely an elite burial and, therefore, dietary habits of individuals interred here may not be representative of the general population. However, this last point may serve to remind us that as well as local micro-regional patterns, intra- and inter-site/population social, political, status, and cultural differences and effects may also have played some role in dietary habits. A Boeotian local micro-regional character is also emphasized by the fact that a relatively similar pattern in the δ13C and δ15N values is also observed for Byzantine Thebes (Figure 4). At Byzantine Thebes, though, there is significant diversity in individual isotopic values which may be skewing the mean values and ranges (Dotsika et al., 2018). But, it has also been suggested for this population that there could be a significant input of C4 crops/C4-fed animals as well as the possible consumption of small and low trophic level marine fish such as sardines or anchovies (Dotsika et al., 2018). However, although we may suggest a local micro-regional phenomenon, the fact that the Bronze Age population from Thebes and the other relatively nearby Bronze Age Boeotian populations of Kalapodi and Zeli do not show this pattern (Figure 4) means that it could also be considered as a temporal pattern for the micro-region; i.e., only in post-Bronze Age periods.
The consumption of marine and freshwater resources in the Aegean World has long been a point of contention in the literature concerning dietary habits (for example, see reviews of this in Garvie-Lok, 2001; Vika and Theodoropoulou, 2012; Vika et al., 2009), and needs to be discussed here. Whilst many isotopic studies into the dietary habits of populations of the Aegean World (especially the Greek part—Crete and West Aegean) have suggested that (marine or freshwater) fish were likely a minor dietary input, there is a plethora of archaeological (e.g., fish hooks and net weights, as well as fish bones) and also textual (from later, historical, periods) evidence to suggest that the sea and its resources were exploited from prehistoric to Classical times in the Aegean World (cf. Garvie-Lok, 2001; Vika and Theodoropoulou, 2012). A study by Vika and Theodoropoulou (2012) into the δ13C and δ15N values of marine and freshwater fish (from bulk bone collagen) of the Aegean World found that there was an overlap in the values between marine and freshwater species and, furthermore, that δ15N values were not as enriched as expected. Garvie-Lok (2001) has also suggested that it can be difficult to distinguish between Aegean and Mediterranean low trophic level marine fish and domesticated terrestrial animals by δ13C and δ15N values alone – especially when considering modest dietary inputs of fish. These findings have been used to suggest that the input of fish into human dietary habits in the Aegean World should be reconsidered. This means that whilst the majority of previous isotopic studies (for some key examples, see Irvine and Erdal, 2020; Lösch et al., 2014; Kontopoulos and Sampson, 2015; Nitsch et al., 2017; Panagiotopoulou et al., 2016; Papathanasiou and Richards, 2015; Papathanasiou et al., 2000; Petroutsa and Manolis, 2010; Pilaar Birch et al., 2021; Vaiglova et al., 2021) into human dietary habits in the Aegean World have suggested predominantly terrestrial based diets with variations in δ13C values likely being a result of C4 plant input and variations in δ15N values being a result of varying inputs of terrestrial animal protein, or agricultural strategies such as manuring, we may need to consider that variations in these isotopic values may be the result of fish consumption—even if relatively minor/modest and subtle. Whilst marine or freshwater resource consumption does not seem to have been of major importance or a primary staple of human dietary habits (this study, and also the majority of other isotopic investigations into dietary habits in the Aegean World; for some key examples, see Irvine and Erdal, 2020; Lösch et al., 2014; Kontopoulos and Sampson, 2015; Nitsch et al., 2017; Panagiotopoulou et al., 2016; Papathanasiou and Richards, 2015; Papathanasiou et al., 2000; Petroutsa and Manolis, 2010; Pilaar Birch et al., 2021; Vaiglova et al., 2021), further research into this is needed. Specifically, the obtaining of isotopic values from fish bones at an intra-site and local regional level, to more precisely clarify specific (including regional, temporal, and ethnic/cultural) human dietary habits, would be an important direction for future research.
Whilst we see a relative (to other Bronze Age Aegean populations) increase in δ13C values, and their range, in some Middle and Late Bronze Age (and to a certain extent Hellenistic) Knossos populations, a corresponding increase and increased range in δ15N values is not observed (Figure 4)—a similar pattern is also observed at EIA Halos (Figure 4). This lack of correlation seems to, on first examination, not clearly suggest an input of marine resources. This relative, and overall, pattern is also very clearly observable when examining the chronological groups by sub-region (Figure 6)—although this is unsurprising as these periods on Crete are mainly represented by these same sites. However, with that being said, Nafplioti (2016) has stated that the δ13C and δ15N values indicate that a third of the overall sampled population from Bronze Age Knossos likely had marine resource inputs in their dietary habits. For the Ailias chamber tombs population at Knossos, Nafplioti (2016) suggested that the more positive δ13C values (and their mean carbon human-animal isotopic offset), as well as the high δ15N values, of a group of nine individuals (Group A, Figure 3 in Nafplioti, 2016) points to marine resource consumption – in combination with terrestrial C3 plant and animal proteins. For the sampled population from the Lower Gypsades tomb at Knossos, Nafplioti (2016) has suggested that the majority of them, according to their δ13C and δ15N values, had some dietary input from marine resources. Furthermore, whilst a corresponding increase in δ15N values is not immediately clear in Figure 4, an increase in δ15N values, especially given the relative offset of the δ15N values to those of the sampled terrestrial fauna, an increase in δ15N values is observable at the site/population level. An isotopic study of dietary habits in later periods at Knossos by Moles et al. (2022) suggested that dietary habits in the Hellenistic period were likely terrestrial mixed C3-based, but in the Hellenistic, Roman and, Late Antique periods there was a small to modest input of marine food resources (or low trophic level marine foods) in the dietary habits of a few individuals. This is something that may actually be observable in this study, with these later periods having relatively (for Knossos) more positive δ13C values and greater δ15N values, as well as a greater number of outliers above the 95th percentile (Figure 4). Also, in the isotopic values from the actual study into these time periods at Knossos by Moles et al. (2022), there are a few individuals with δ13C values > −19‰ and corresponding δ15N values >9.5‰, which may indicate some marine consumption. These examples provide an important reminder that whilst an overall and broad pattern may be visible, there may be slight and more nuanced intra-population differences that are not visible when examining dietary habits at a large-scale, such as in this study, as well as highlighting the importance of establishing robust isotopic baselines for local regions and food webs.
At Byzantine/medieval Hagios Sozon, Orchomenos the human δ15N values are relatively high, but a corresponding increase in δ13C values is not observed, and therefore this would suggest a greater input of animal protein into their dietary habits (perhaps of omnivorous species such as pigs), a heavy reliance on dairy foods, or perhaps more intensive agricultural practices with well-manured crops (Figure 4). The δ15N values for animals from the site are also relatively high, with some pigs, sheep, goat, and cattle exhibiting values >9‰ (Tritsaroli et al., 2022). Although, following an examination of the trophic enrichment, Tritsaroli et al. (2022) suggested that the humans had a low animal protein intake and that the high δ15N values in the animals (and thereby humans) were a result of an arid environment or agro-pastoral practices. This may further point toward a local micro-regional characteristic for the region; either climatic, environmental, or in terms of agricultural practices and strategies.
Research has demonstrated that in the Neolithic periods of Greece dietary habits were likely grain-centric with modest inputs of animal protein, and exploited resources were dominated by domesticated species with very little input from wild resources (Halstead and Isaakidou, 2020; Papathanasiou, 2015). There is a dominance of farming, and the general picture of farming in Neolithic Greece is one of small-scale, with intensive crop cultivation and small-scale intensive livestock husbandry with a focus on mainly primary consumption (i.e., meat) and the animals' diet consisting of plants from manured cultivation plots (Halstead and Isaakidou, 2020). The human δ13C and δ15N values for the West Aegean from the Neolithic period appear to possibly support this, with a relatively narrow range in δ13C values, but a greater range in δ15N values (Figure 6). This range may be due to a combination of intensive and extensive cultivation practices, with some crops being substantially manured, as has been observed through δ15N values of charred grains from several Greek sites of the Neolithic period (Halstead and Isaakidou, 2020; Vaiglova et al., 2020, 2021). Free-threshing wheat, in particular, appears to have been selectively more intensively cultivated at some sites than other wheat species and barley in the Neolithic periods of Greece and Crete (Vaiglova et al., 2020, 2021). Therefore, on a site-by-site basis, the consumption of resources from a combination of small-scale animal husbandry (i.e., animals not herded far from the settlement, and indeed penned and foddered in some instances—cf. Vaiglova et al., 2020) with a dominance of domesticated crops (C3 cereals like wheat and barley and legumes like lentils) also cultivated close to the settlement would hypothetically result in a narrow range of δ13C values in human consumers (assuming similar crops grown in relatively similar areas/field locations under similar humidity conditions). Variations in manuring practices would result in a greater range of plant δ15N values, and therefore also in foddered animals and ultimately human consumers of the crops and animals. For example, at Neolithic Halai and Makriyalos the animals exhibit a narrow range in δ13C values but a wide range in their δ15N values, possibly as a result of the wide range in δ15N values of the crops, and, furthermore, the human δ15N values (and their range) suggest that the bulk of their protein input came from plants rather than animals (Vaiglova et al., 2021).
Post-Neolithic prehistoric (i.e., Bronze Age) dietary habits are predominantly C3 and terrestrial-based, as has also been noted previously for sites in the region (Irvine, 2022; Irvine et al., 2019; Papathanasiou, 2015; Richards et al., 2022; Vika, 2011, 2015). Although, the later Bronze Age periods do exhibit a wider range in δ13C values than the Early Bronze Age (Figure 5) which may be an indication of more extensive and diverse agro-pastoral practices and the introduction of C4 crops such as millet. In the Bronze Age, archaeobotanical assemblages in Greece are dominated by wheat (emmer, einkorn, bread, and possibly spelt) and barley (two- and six-row), possibly millet, and legumes of lentils, peas, bitter vetch, grass pea, and fava beans (Papathanasiou, 2015). A similar picture is also found in Anatolia during this period (cf. Irvine, 2022; Kavak and Çakan, 2022). Zooarchaeological assemblages in the Aegean region during this period are dominated by terrestrial domesticated animals, especially sheep/goat, cattle, and pigs (cf. Irvine, 2022; Papathanasiou, 2015).
With the exception of Thebes (discussed above), the mean δ13C values of the Byzantine period populations are relatively similar (roughly around −19.0‰ —Figure 4). Something also observed in the mean δ13C values of Eleutherna (−18.9‰ ± 0.6‰) and Stylos (−18.8‰ ± 0.8‰) (Bourbou et al., 2011). The mean δ15N values of Eleutherna (8.2‰ ± 1.4‰) and Stylos (9.4‰ ± 1.7‰) (Bourbou et al., 2011) are also in a similar general range as the other Byzantine sites of ca. 8.5 to 9.5‰ (Figure 4). However, Eleutherna is slightly lower than the rest of the Byzantine populations, whilst Thebes and Hagios Sozon in particular have distinctly higher δ15N values (Figure 4). The ranges and clustering of mean δ13C and δ15N values for the humans agree quite well with dietary habits based on the approximated ranges for the δ13C and δ15N values of domesticated terrestrial animals of ca. −21.5‰ to −19.5‰ for δ13C and ca. 3.5‰ to 6.5‰ for δ15N for Byzantine period Greece and Crete given in Bourbou and Garvie-Lok (2015). The general tight clustering of the mean isotopic values of the Byzantine populations, and its unexpectedness considering the diverse range of available food resources has previously been noted (Bourbou and Garvie-Lok, 2015; Bourbou et al., 2011). Even to the extent that is has been suggested that the broadly similar isotopic values may indicate the existence of a general “Byzantine diet” of C3 crops (wheat and barley) and domesticated animals feeding on C3 plants; in agreement with textual and documentary sources which point toward a diet of grains, bread, oil, and wine supplemented with other food resources (Bourbou and Garvie-Lok, 2015 and references within; Bourbou et al., 2011; Dotsika et al., 2018). Although Byzantine period Thebes stands out as an obvious outlier from this, and to a certain extent Hagios Sozon due to its overall high δ15N values, a similar observation can be made from this study of general similarity in dietary habits in the Byzantine period. It should be stressed, however, that this similarity is an overall similarity, but there are cases of intra-population variability. For example, some individuals have lower δ15N values, more positive δ13C values; implying greater direct/indirect C4 input into human dietary habits (cf. Bourbou and Richards, 2007; Bourbou and Garvie-Lok, 2015; Bourbou et al., 2011; Dotsika et al., 2018). There are also other individuals, at an intra-site level, who have isotopic values suggesting significant marine resources inputs/dependence (cf. Bourbou and Richards, 2007; Bourbou and Garvie-Lok, 2015; Bourbou et al., 2011; Dotsika et al., 2018). Also, at medieval Petras on Crete and Gatteliusi on Mitilini (both island populations) Garvie-Lok (2001) found some evidence through correlation of the human δ13C and δ15N values that terrestrial dietary habits were supplemented with marine resources.
4.2 Sub-region patterns: East Aegean vs. Crete vs. West Aegean
It was important to consider patterns in the isotopic values by sub-region of the Aegean World—East Aegean, Crete, and West Aegean (Figure 6), due to topographical and geographical differences. For one, Crete is an island in a border zone between the southern extent of the Aegean into the East Mediterranean, and as such has different geographical and also human connectivity aspects compared to the East and West Aegean. The topography of the East Aegean region is mostly composed of broad and fertile lowland valley plains, whilst in the West Aegean the southern part is characterized by more rugged topography, and in its northern part (north of Boeotia) there are large broad plains interspersed with more rugged hilly areas. The topography of Crete, beyond being a large(r) island located further from a mainland than the other smaller Aegean islands, is also more rugged, with several high mountain ranges, multiple hilly areas, and some small lowland plains and valleys. The modern climates of Crete, and the East and West Aegean are relatively similar, classified according to the Köppen climate classification as hot-summer Mediterranean (Csa), although the areas to the north of the Aegean Sea are classified as cold semi-arid (BSk) (Rubel and Kottek, 2011). The past climatic conditions in the Aegean World are thought to have been, overall and generally, quite similar to those of today (mild, wet winters and relatively warm to hot, dry/drier summers), although with some chronological variation (for example, see Moody, 2014, 2016; Psomiadis et al., 2018). Additionally, the Aegean region as a whole is at the center of the Eastern Mediterranean vegetation zone (Runemark, 1971), and, generally speaking, the overall modern biome for the East and West Aegean regions are considered to be Mediterranean forests, woodlands, and scrub (see One Earth —https://www.oneearth.org/ecoregions/mediterranean-woodlands-and-forests/), and the ecoregion is classified as Aegean and Western Turkey sclerophyllous and mixed forests [see Digital Observatory for Protected Areas (DOPA) Explorer—https://dopa-explorer.jrc.ec.europa.eu/ecoregion/81201; and One Earth—https://www.oneearth.org/ecoregions/aegean-and-western-turkey-sclerophyllous-and-mixed-forests/]. Whilst there are environmental and climatic similarities between the regions there are some differences and it would, therefore, be expected that these differences would have potentially influenced and affected subsistence practices and agricultural strategies (in terms of available cultivatable land and arable field agriculture and livestock rearing and management). Concurrently, it would also be expected that the three sub-regions would have different and varying isoscape values which would influence δ13C and δ15N values in plants, animals, and thereby humans. However, what is interesting is that, with a few exceptions, and the expected micro-regional site-by-site variability, the δ13C and δ15N values are broadly consistent with relatively little overall variation, not only between the sub-regions, but also across chronological periods. This would, therefore, imply, that across the Aegean World, that similar crops were being cultivated in similar ways and that similar animals were being reared and managed as well as exploited in similar ways. It may even, very tentatively, suggest that chronological variations in climate (i.e., drier, or colder, or wetter periods) did not have dramatic and significant effects on subsistence practices and dietary habits. Although, this should be investigated and tested further, and more thoroughly. That is not to say that there are no differences or variation, of course there are (as discussed above), but the general overall “big picture” pattern is one of relative consistency. Yet, we need to acknowledge this overall observation in this study, of relative consistency between regions and across time, with some important caveats and recognition of limitations. By looking at a “big picture” and overall patterns we do lose, and perhaps even obscure, micro-regional and intra-population and intra-chronological period nuances and subtleties in variations and diversity in dietary habits and associated subsistence practices and agricultural strategies – it has the effect of “smoothing out” the data to a certain extent [see also the latter part of the discussion in Papathanasiou and Richards (2015)]. Nafplioti (2017) has also highlighted that large-scale studies such as this may result in the exaggeration of patterns, and that interpretations may be compromised by small sample sizes, the lack of robust isotopic baseline values, and the paucity of isotopic values from regions or time periods. The available and published human δ13C and δ15N values for the Aegean World is heavily biased toward the western region (i.e., West Aegean), which has the obvious potential to create prejudices in the obtained and analyzed data and how we interpret it. And lastly, many of the sub-regions are only represented by single sites/populations which also has the potential to result in biases in the data and its interpretation. Ultimately, these final two limitations mean that we are only seeing part of the picture whilst applying it on a large-scale. However, with all that being said, and acknowledged, there is arguably enough data and information to at the very least put forward these thoughts as a starting point of consideration. As always, these kinds of thoughts and observations will be further clarified (enforced or dismissed) by additional isotopic values and related research in the region. Furthermore, whilst initially being surprising, it, perhaps, should not be completely implausible. The similarity between the East and West Aegean in terms of human δ13C and δ15N values, and thereby implied dietary habits and subsistence practices, has previously been observed for the Early-Middle Bronze Age/Early-Middle Helladic period (Irvine et al., 2019). Although not plotted in the Figures of this paper (see explanation in Section 2), the δ13C and δ15N values from Middle Helladic Kouphovouno suggest a C3-based terrestrial diet that is generally quite similar to other populations of a similar chronological period, although with perhaps a very slightly greater reliance on crops and lower input of animal protein: δ13C mean value of −19.8‰ ± 0.3‰; δ15N mean value of 7.9‰ ± 0.4‰ (Lagia et al., 2007). Furthermore, researchers examining dietary habits in the Greek world (Greek mainland and islands, and Crete) using stable isotope analyses have been using the terms “Neolithic diet” and “Byzantine diet” for more than a decade to explain the general homogeneous patterns observed for potential exploited food resources and overall and general tight clustering of the δ13C and δ15N values (Bourbou and Garvie-Lok, 2015; Bourbou et al., 2011; Dotsika et al., 2018; Papathanasiou and Richards, 2015). And, as outlined in the introduction, decades of archaeological research, including recent research applying quantifiable scientific and biomolecular methods such as isotopic and aDNA analyses, have consistently pointed toward high levels of connectivity and mobility in the Aegean World from at least the Neolithic period onwards, and indeed even earlier. This has involved physical connectivity through trade and exchange networks, the movement of people and resources—including agricultural resources, and more metaphysical connectivity such as the movement of ideas and material culture “fashions,” burial habits and mortuary practices, and cultural and socio-cultural connectivity. For long periods of its history the Aegean World (or at least significant parts of it) was also under the control of singular empires and/or relatively culturally and socially similar city states/proto-empires/kingdoms, further providing a singular connective aspect to the region. Dietary habits and subsistence practices may simply be one more window through which macro-regional connectivity in the Aegean World can be observed.
5 Conclusion
Whilst it can be said that the δ13C values are actually quite consistent over time in the Aegean World, there is a clearly observable general and overall, if small, increase in δ13C values over time. Greater variability in δ13C values is observed on a site-by-site basis, which is to be largely expected due to varying local and micro-regional environmental and ecological conditions that are likely to have influenced subsistence practices and agricultural strategies, and thus δ13C values. A similar general and overall relative consistency (albeit relatively more irregular) is also observed in the δ15N values over time. This is with the exception of the Classical, and to a lesser extent the Hellenistic and Byzantine, sites and periods. This is particularly true of the Classical and Hellenistic sites and populations around Athens and also at Thebes, which have markedly greater δ15N values. The majority of the Aegean World sites and populations (both regionally and diachronically) have dietary habits that are predominantly terrestrial and C3-based (with a heavy reliance on domesticated plant and animal food resources with marginal wild input), with likely varying (direct or indirect) inputs of C4 plants in periods after later Bronze Age periods (see also Papathanasiou and Richards, 2015). The sites around Athens, and also at Thebes, may have had a greater input of freshwater and/or low trophic level marine resources in their dietary habits (relative to earlier periods and even later periods where marine consumption has been suggested to be more common), and is something which would constitute a local micro-regional flavor and characteristic. Modest marine resource consumption may also be true for Bronze Age, Roman, and Late Antique Knossos. However, it should be noted that previous research has suggested that identifying marine and freshwater resource consumption in the Aegean World by δ13C and δ15N values alone may be tricky, and that stable isotope analysis into dietary habits may be underestimating the contribution of marine and freshwater resources in human dietary habits in this region. As a result, further research into the isotopic values of potentially exploited and consumed marine and freshwater species is needed to further clarify this issue.
One further initial, and perhaps surprising but interesting, observation is that this relative and broad consistency in δ13C and δ15N values for the overall Aegean World is also noted when examining the different sub-regions (East Aegean, Crete, West Aegean). Furthermore, expected differences, with perhaps overall distinct sub-regional patterns, between the three sub-regions of the Aegean World are not observed. This may point toward dietary habits, subsistence practices, and agricultural strategies being a definable aspect of connectivity in the region. However, in concluding, it should again be emphasized that (due to time and space constraints in this study) these are broad and general observable patterns in the human isotopic values and, furthermore, that with the data currently available this remains only a tentative observation and “jumping-off” point of consideration that should be explored further. For example, future research should also endeavor to more precisely incorporate archaeobotanical, zooarchaeological, palaeoenvironmental, and palaeoclimatic datasets (including isotopic analyses applied to these materials and proxies), and chronological variations in them. This would help to provide a more nuanced examination of dietary habits and subsistence practices over time in the region.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
BI: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
Thank you to Argyro Nafplioti, Yilmaz Selim Erdal, and Christina Papageorgopoulou for putting together this research topic and for inviting me to contribute. Thank you very much as well to the two reviewers and the editor for their comments and suggestions which helped to improve this paper.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ambrose, S. (1990). Preparation and characterization of bone and tooth collagen for isotopic analysis. J. Archaeol. Sci. 17, 431–451. doi: 10.1016/0305-4403(90)90007-R
Araus, J. L., Ferrio, J. P., Voltas, J., Aguilera, M., and Buxó, R. (2014). Agronomic conditions and crop evolution in ancient Near East agriculture. Nat. Commun. 5:3953. doi: 10.1038/ncomms4953
Bourbou, C., Fuller, B. T., Garvie-Lok, S. J., and Richards, M. P. (2011). Reconstructing the diets of Greek Byzantine populations (6th-15th centuries AD) using carbon and nitrogen stable isotope ratios. Am. J. Phys. Anthropol. 146, 569–581. doi: 10.1002/ajpa.21601
Bourbou, C., and Garvie-Lok, S. (2015). “Bread, Oil, Wine, and Milk: Feeding Adults and Infants in Byzantine Greece,” in Archaeodiet in the Greek World: Dietary Reconstruction from Stable Isotope Analysis, eds. A. Papathanasiou, M. P. Richards, and S. C. Fox (Princeton, NJ: The American School of Classical Studies at Athens) 171–194.
Bourbou, C., and Richards, M. P. (2007). The Middle Byzantine menu: palaeodietary information from isotopic analysis of humans and fauna from Kastella, Crete. Int. J. Osteoarchaeol. 17, 63–72. doi: 10.1002/oa.855
Coleman, J. E., Karimali, E., Karali, L., Fillios, M., Diffey, C., Vaiglova, P., et al. (2017). “The Environment and Interactions of Neolithic Halai,” in Communities, Landscapes and Interaction in Neolithic Greece: Proceedings of the International Conference, Rethymno, eds. A. Sarris, E. Kalogiropoulou, T. Kalayci, and L. Karimali (Ann Arbor: International Monographs in Prehistory) 96–124. doi: 10.2307/j.ctvw049k3.13
DeNiro, M. J. (1985). Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317, 806–809. doi: 10.1038/317806a0
Dotsika, E., Michael, D. E., Iliadis, E., Karalis, P., and Diamantopoulos, G. (2018). Isotopic reconstruction of diet in Medieval Thebes (Greece). J. Archaeol. Sci. Rep. 22, 482–491. doi: 10.1016/j.jasrep.2018.08.019
Garvie-Lok, S. J. (2001). Loaves and fishes: a stable isotope reconstruction of diet in medieval Greece. [dissertation]. [Calgary, Alberta]: University of Calgary.
Halstead, P., and Isaakidou, V. (2020). “Pioneer farming in earlier Neolithic Greece,” in Farmers at the Frontier: A Pan-European Perspective on Neolithisation, eds. K. J. Gron, L. Sørensen, and P. Rowley-Conwy (Oxford: Oxbow) 77–100. doi: 10.2307/j.ctv13gvh1g.7
Ingvarsson-Sundström, A., Richards, M. P., and Voutsaki, S. (2009). Stable isotope analysis of the Middle Helladic population from two cemeteries at Asine: Barbouna and the East Cemetery. Mediterr. Archaeol. Archaeom. 9, 1–14. Available online at: https://www.maajournal.com/index.php/maa/article/view/287
Irvine, B. (2022). Investigating a subsistence model of staple finance for the late 4th to early 2nd Millennium BCE of the Greater Near East. Arkeoloji Bilimleri Dergisi/Turk. J. Archaeol. Sci. 2, 52–121.
Irvine, B., and Erdal, Y. S. (2020). Multi-isotopic analysis of dietary habits and mobility at third millennium BC Bakla Tepe, West Anatolia. Archaeol. Anthropol. Sci. 12:111. doi: 10.1007/s12520-020-01078-2
Irvine, B., Erdal, Y. S., and Richards, M. P. (2019). Dietary habits in the Early Bronze Age (3rd millennium BC) of Anatolia: a multi-isotopic approach. J. Archaeol. Sci. Rep. 24, 253–263. doi: 10.1016/j.jasrep.2019.01.015
Irvine, B., Luke, C., Çakirlar, C., Kaner, T., Özbal, R., Findiklar, S., et al. (2025). Stable isotope analysis of faunal remains from Bronze Age Kaymakçi, Western Anatolia. Holocene 35, 223–233. doi: 10.1177/09596836241297077
Kavak, S., and Çakan, H. (2022). Archaeobotanical evidence of agricultural products in the southern Mediterranean part of Turkey during the Bronze Ages from Tatarli Höyük. Turk. J. Bot. 46, 379–395. doi: 10.55730/1300-008X.2716
Kontopoulos, I., and Sampson, A. (2015). Prehistoric diet on the island of Euboea, Greece: an isotopic investigation. Mediterr. Archaeol. Archaeom. 15, 97–111. doi: 10.5281/zenodo.18361
Krauß, R., Marinova, E., De Brue, H., and Weninger, B. (2018). The rapid spread of early farming from the Aegean into the Balkans via the Sub-Mediterranean-Aegean Vegetation Zone. Quat. Int. 496, 24–41. doi: 10.1016/j.quaint.2017.01.019
Lagia, A. (2015). “Diet and the polis: an isotopic study of diet in Athens and Laurion during the Classical, Hellenistic, and Imperial Roman periods,” in Archaeodiet in the Greek World: Dietary Reconstruction from Stable Isotope Analysis, eds. A. Papathanasiou, M. P. Richards, and S. C. Fox (Princeton, NJ: The American School of Classical Studies at Athens) 119–145.
Lagia, A., Petroutsa, E., and Manolis, S. (2007). “Health and diet during the Middle Bronze Age in the Peloponnese: the site of Kouphovouno,” in Cooking up the Past: Food and Culinary Practices in the Neolithic and Bronze Age, eds. C. Mee and J. Renard (Oxford: Oxbow) 313–328.
Lösch, S., Moghaddam, N., Grossschmidt, K., Risser, D. U., and Kanz, F. (2014). Stable isotope and trace element studies on gladiators and contemporary Romans from Ephesus (Turkey, 2nd and 3rd Ct. AD) - implications for differences in diet. PLoS One 9:e110489. doi: 10.1371/journal.pone.0110489
Massa, M., and Şahoǧlu, V. (2011). “Western Anatolian burial customs during the Early Bronze Age,” in Across the Cyclades and western Anatolia during the 3rd millennium BC, eds. V. Şahoǧlu and P. Sotirakopoulou (Istanbul: Çagatay Anadol) 164–171.
Milić, M. (2014). PXRF characterisation of obsidian from central Anatolia, the Aegean and central Europe. J. Archaeol. Sci 41:285e296. doi: 10.1016/j.jas.2013.08.002
Moles, A. C., Reade, H., Jourdan, A.-L., and Stevens, R. E. (2022). Stable isotopes reveal dietary shifts associated with social change in Hellenistic, Roman and Late Antique Knossos. J. Archaeol. Sci. Rep. 45:103609. doi: 10.1016/j.jasrep.2022.103609
Moody, J. (2014). “Prehistoric and historic climate and weather in (East) Crete” in A Cretan landscape through time: Priniatikos Pygros and environs: BAR International Series 2634, eds. B. P. C. Molloy and C. N. Duckworth (Oxford: Archaeopress) 23–164.
Moody, J. (2016). “The Roman climate in the southwest Aegean: was it really different?” in Roman Crete new perspectives, eds. J. E. Francis and A. Kouremenos (Oxford: Oxbow Books) 59–82. doi: 10.2307/j.ctvh1drr7.10
Nafplioti, A. (2008). “Mycenaean” political domination of Knossos following the Late Minoan IB destructions on Crete: negative evidence from strontium isotope ratio analysis (87Sr/86Sr). J. Archaeol. Sci. 35, 2307–2317. doi: 10.1016/j.jas.2008.03.006
Nafplioti, A. (2016). Eating in prosperity: first stable isotope evidence of diet from Palatial Knossos. J. Archaeol. Sci. Rep. 6, 42–52. doi: 10.1016/j.jasrep.2016.01.017
Nafplioti, A. (2017). Archaeodiet in the Greek World: Dietary Reconstruction from Stable Isotope Analysis Edited by Anastasia Papathanasiou, Michael P. Richards, and Sherry C. Fox. Am. J. Archaeol. 121. doi: 10.3764/ajaonline1214.nafplioti
Nafplioti, A., Driessen, J., Schmitt, A., and Crevecoeur, I. (2021). Mobile (after-) lifeways: People at pre-and protopalatial Sissi (Crete). J.Archaeol. Sci. Rep. 35:102718. doi: 10.1016/j.jasrep.2020.102718
Nitsch, E., Andreou, S., Creuzieux, A., Gardeisen, A., Halstead, P., Isaakidou, V., et al. (2017). A bottom-up view of food surplus: using stable carbon and nitrogen isotope analysis to investigate agricultural strategies and diet at Bronze Age Archontiko and Thessaloniki Toumba, northern Greece. World Archaeol. 49, 105–137. doi: 10.1080/00438243.2016.1271745
Palaima, T. G. (2010). “Linear B,” in The Oxford Handbook of the Bronze Age Aegean, ed. E. Cline (Oxford: Oxford University Press) 356–372. doi: 10.1093/oxfordhb/9780199873609.013.0027
Panagiotopoulou, E., and Papathanasiou, A. (2015). “Dietary reconstruction at the Geometric-Period burial site of Ayios Dimitrios.” in Archaeodiet in the Greek World: Dietary Reconstruction from Stable Isotope Analysis, eds. A. Papathanasiou, M. P. and Richards, S. C. Fox (Princeton, NJ: The American School of Classical Studies at Athens) 105–117.
Panagiotopoulou, E., van der Plicht, J., Papathanasiou, A., Voutsaki, S., Nikolaou, E., and Tsiouka, F. (2016). Isotopic (13C, 15N) investigation of diet and social structure in Early Iron Age Halos, Greece. J. Archaeol. Sci. Rep. 10, 212–220. doi: 10.1016/j.jasrep.2016.09.020
Papathanasiou, A. (2015). “Stable isotope analyses in Neolithic and Bronze Age Greece: An overview,” in Archaeodiet in the Greek World: Dietary Reconstruction from Stable Isotope Analysis, eds. A. Papathanasiou, M. P. Richards, and S. C. Fox (Princeton, NJ: The American School of Classical Studies at Athens) 25–55.
Papathanasiou, A., Larsen, C. S., and Norr, L. (2000). Bioarchaeological inferences from a Neolithic ossuary from Alepotrypa Cave, Diros, Greece. Int. J. Osteoarchaeol. 10, 210–228. doi: 10.1002/1099-1212(200005/06)10:3<210::AID-OA523>3.0.CO;2-2
Papathanasiou, A., and Richards, M. P. (2015). “Summary: Patterns in the carbon and nitrogen isotope data through time,” in Archaeodiet in the Greek World: Dietary Reconstruction from Stable Isotope Analysis, eds. A. Papathanasiou, M. P. Richards, and S.C. Fox (Princeton, NJ: The American School of Classical Studies at Athens) 195–203.
Parkinson, W. A., and Galaty, M. L. (2007). Secondary states in perspective: an integrated approach to state formation in the prehistoric Aegean. Am. Anthropol. 109, 113–129. doi: 10.1525/aa.2007.109.1.113
Petroutsa, E. I., and Manolis, S. K. (2010). Reconstructing late bronze age diet in mainland Greece using stable isotope analysis. J. Archaeol. Sci. 37, 614–620. doi: 10.1016/j.jas.2009.10.026
Pilaar Birch, S. E., Boz, B., and Erdoǧu, B. (2021). Isotope paleodietary analysis of humans from Chalcolithic Uǧurlu, Gökçeada Island, Turkey. J. Archaeol. Sci. Rep. 39:103136. doi: 10.1016/j.jasrep.2021.103136
Psomiadis, D., Dotsika, E., Albanakis, K., Ghaleb, B., and Hillaire-Marcel, C. (2018). Speleothem record of climatic changes in the northern Aegean region (Greece) from the Bronze Age to the collapse of the Roman Empire. Palaeogeogr. Palaeoclimatol. Palaeoecol. 489, 272–283. doi: 10.1016/j.palaeo.2017.10.021
Rahmstorf, L. (2015). “The Aegean before and after c. 2200 BC between Europe and Asia: Trade as a prime mover of cultural change,” in 2200 BC – Ein Klimasturz als Ursache für den Verfall der Alten Welt. 2200 BC – A climatic breakdown as a cause for the collapse of the old world. 7. Mitteldeutscher Archäologentag vom 23. bis 26. Oktober 2014 in Halle (Saale). 7th Archaeological Conference of Central Germany October 23-26, 2014 in Halle (Saale), eds. H. Meller, H. W. Arz, R. Jung, and R. Risch (Halle (Saale): Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt, Landesmuseum für Vorgeschichte) 149–180.
Richards, M., Smith, C., Nehlich, O., Grimes, V., Weston, D., Mittnik, A., et al. (2022). Finding mycenaeans in minoan crete? Isotope and DNA analysis of human mobility in Bronze Age Crete. PLoS One 17:e0272144. doi: 10.1371/journal.pone.0272144
Rubel, F., and Kottek, M. (2011). Comments on: “The thermal zones of the Earth” by Wladimir Köppen (1884). Meteorol. Z. 20, 361–365. doi: 10.1127/0941-2948/2011/0285
Runemark, H. (1971). “Distribution patterns in the Aegen,” in Plant life of South-West Asia, eds. P. H. Davis, P. C. Harper, and I. C. Hedge (Edinburgh: The Botanical Society of Edinburgh) 3–12.
Şahoǧlu, V. (2004). Interregional contacts around the Aegean during the Early Bronze Age: new evidence from the Izmir region. Anadolu 27, 97–120. doi: 10.1501/Andl_0000000310
Şahoǧlu, V. (2019). “The Early Bronze Age Anatolian trade network and its role on the transformation of the Anatolian and Aegean communities,” in Connecting Cultures, eds. V. Şahoǧlu, M. Şevketoǧlu, and Y. H. Erbil (Ankara: Ankara University Press) 115–131.
Şahoǧlu, V. (2024). The Early Bronze Age cemetery at Çeşme–Boyalik in coastal western Anatolia. Annu. Br. Sch. Athens.. 119, 1–30. doi: 10.1017/S0068245423000102
Skourtanioti, E., Ringbauer, H., Ruscone, G. A. G., Bianco, R. A., Burri, M., Freund, C., et al. (2023). Ancient DNA reveals admixture history and endogamy in the prehistoric Aegean. Nat. Ecol. Evol. 7, 290–303. doi: 10.1038/s41559-022-01952-3
Slim, F. G., and Çakirlar, C. (2023). Pigs and polities in Iron Age and Roman Anatolia: an interregional zooarchaeological analysis. Quat. Int. 662–663, 47–62. doi: 10.1016/j.quaint.2022.05.013
Sołtysiak, A. (2019). Strontium and nitrogen isotopic evidence of food import to tell Ashara–Terqa, a Bronze age city on the Euphrates, Syria. Int. J. Osteoarchael. 29, 127–133. doi: 10.1002/oa.2724
Triantaphyllou, S., Richards, M. P., Touchais, G., Philippa-Touchais, A., and Voutsaki, S. (2006). Analyses of Middle Helladic skeleton material from Aspis, Argos: stable isotope analysis of human remains. École Française d'Athènes: Bulletin de correspondance Hellénique 130, 627–637. doi: 10.3406/bch.2006.7430
Triantaphyllou, S., Richards, M. P., Zerner, C., and Voutsaki, S. (2008). Isotopic dietary reconstruction of humans from Middle Bronze Age Lerna, Argolid, Greece. J. Archaeol. Sci. 35, 3028–3034. doi: 10.1016/j.jas.2008.06.018
Tritsaroli, P., Grigorakakis, G., and Richards, M. (2023). Bioarchaeological insights into the Late Helladic communities of South Kynouria, Peloponnese: the case of the LH IIIA2-IIIB2 burial cluster of Socha. Int. J. Osteoarchaeol. doi: 10.1002/oa.3268
Tritsaroli, P., Mion, L., Herrscher, E., Andr,é, G., and Vaxevanis, G. (2022). Health, diet, and mortuary practices in the countryside of Byzantine and post-Byzantine Boeotia: the case of Hagios Sozon in Orchomenos. Int. Osteoarchaeol. 32, 1238–1252. doi: 10.1002/oa.3159
Ünlü, E. (2016). The handle wagging the cup. Formal aspects of alcohol consumption in the transfer of ideology: Anatolia and the Aegean towards the end of the third millennium BC. Oxford J. Archaeol. 35, 345–358. doi: 10.1111/ojoa.12093
Ünlüsoy, S. (2016). “Troy and the Aegean during the third millennium BC,” in Early Bronze Age Troy: Chronology, Cultural Development, and Interregional Contacts, eds. E. Pernicka, S. Ünlüsoy, and S. W. E. Blum (Bonn: Dr. Rudolf Habelt GmbH) 397–405.
Vaiglova, P., Coleman, J., Diffey, C., Tzevelekidi, V., Fillios, M., Pappa, M., et al. (2021). Exploring diversity in Neolithic agropastoral management in mainland Greece using stable isotope analysis. Environ. Archaeol. 28, 62–85. doi: 10.1080/14614103.2020.1867292
Vaiglova, P., Gardeisen, A., Buckley, M., Cavanagh, W., Renard, J., Lee-Thorp, J., et al. (2020). Further insight into Neolithic agricultural management at Kouphovouno, southern Greece: expanding the isotopic approach. Archaeol. Anthropol. Sci. 12:43. doi: 10.1007/s12520-019-00960-y
Vaiglova, P., Halstead, P., Pappa, M., Triantaphyllou, S., Valamoti, S. M., Evans, J., et al. (2018). Of cattle and feasts: multi-isotope investigation of animal husbandry and communal feasting at Neolithic Makriyalos, northern Greece. PLoS One 13:e0194474. doi: 10.1371/journal.pone.0194474
van Klinken, G. J. (1999). Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J. Archaeol. Sci. 26, 687–695. doi: 10.1006/jasc.1998.0385
van Klinken, G. J., Richards, M. P., and Hedges, R. E. M. (2000). “An overview of causes for stable isotopic variations in past European human populations: environmental, ecophysiological and cultural effects,” in Biogeochemical Approaches to Paleodietary Analysis, eds. S.H., Ambrose and M.A. Katzenberg (New York/London: Kluwer Academic/Plenum Publishers) 39–63. doi: 10.1007/0-306-47194-9_3
Vergidou, C., Karamitrou-Mentessidi, G., Malama, P., Darakis, K., Dee, M. W., Voutsaki, S., et al. (2023). Exploring dietary differentiation in the Roman province of Macedonia: isotopic evidence from Pontokomi-Vrysi and Nea Kerdylia-Strovolos. Environ. Archaeol. 1–23. doi: 10.1080/14614103.2023.2248774
Vika, E. (2011). Diachronic dietary reconstructions in ancient Thebes, Greece: results from stable isotope analyses. J. Archaeol. Sci. 38, 1157–1163. doi: 10.1016/j.jas.2010.12.019
Vika, E. (2015). “Variations in diet in prehistoric Thebes: the case of the Bronze Age mass burial,” in Archaeodiet in the Greek World: Dietary Reconstruction from Stable Isotope Analysis, eds. A. Papathanasiou, M.P. Richards, and S. C. Fox (Princeton, NJ: The American School of Classical Studies at Athens) 77–88.
Vika, E., Aravantinos, V., and Richards, M. P. (2009). Aristophanes and stable isotopes: a taste for freshwater fish in Classical Thebes, (Greece)? Antiquity 83, 1076–1083. doi: 10.1017/S0003598X00099361
Keywords: western Anatolia, Greece, Crete, Aegean, stable isotopes, palaeodiet
Citation: Irvine B (2025) An isotopic overview of dietary habits and subsistence practices in the Aegean World. Front. Environ. Archaeol. 4:1525822. doi: 10.3389/fearc.2025.1525822
Received: 10 November 2024; Accepted: 16 May 2025;
Published: 13 June 2025.
Edited by:
Argyro Nafplioti, Foundation for Research and Technology Hellas (FORTH), GreeceReviewed by:
Yu Itahashi, University of Tsukuba, JapanMonica Margarit, Valahia University of Târgovişte, Romania
Copyright © 2025 Irvine. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Benjamin Irvine, YmVuamFtaW4uaXJ2aW5lQGJpYWEuYWMudWs=