 Enrique Moreno1*
Enrique Moreno1* Celeste Samec2
Celeste Samec2 Luis Manuel del Papa3
Luis Manuel del Papa3 Constanza Taboada4
Constanza Taboada4 Pablo Mercolli5
Pablo Mercolli5 Rodrigo Nores6
Rodrigo Nores6 Stefanie Schirmer7
Stefanie Schirmer7 Noel Amano7*
Noel Amano7*- 1Instituto Regional de Estudios Socio-culturales (CONICET-UNCA), Escuela de Arqueología, Universidad Nacional de Catamarca, Catamarca, Argentina
- 2Instituto de Geocronología y Geología Isotópica (CONICET-UBA), Universidad de Buenos Aires, Buenos Aires, Argentina
- 3División Antropología, Museo de la Plata, Facultad de Ciencias Naturales y Museo (UNLP-CONICET), La Plata, Argentina
- 4Instituto Superior de Estudios Sociales (CONICET-UNT), Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Tucumán, Argentina
- 5Instituto Interdisciplinario Tilcara, Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires, Argentina
- 6Instituto de Antropología de Córdoba (CONICET-UNC), Departamento de Antropología, Facultad de Filosofía y Humanidades, Universidad Nacional de Córdoba, Córdoba, Argentina
- 7Max Planck Institute of Geoanthropology, Jena, Germany
Human-camelid interactions were an essential component in the everyday life of the societies that inhabited the South and Central Andes. While numerous studies have concentrated on this topic, they have primarily focused on the Puna plateaus, where camelids remain a key resource today. In contrast, the eastern Andean regions of Northwestern Argentina have been historically viewed as marginal and the presence of camelids in zooarchaeological assemblages has been often attributed to hunting or specialized pastoralist production for political and administrative centers located in the western valleys. Recent research in this area challenges and reshapes this view. This paper reassesses the presumed marginality of these regions by critically looking at zooarchaeological information including taxonomic, osteometric, age profiles and skeletal parts representation analyses from several key sites. We propose a preliminary model of camelid management in these environments, contributing to broader discussions of similar practices in non-high-Andean regions of South America.
1 Introduction
South American camelids (SAC), including the wild species Vicugna vicugna (vicuña) and Lama guanicoe (guanaco), and their domestic counterparts Lama pacos (alpaca) and Lama glama (llama), have been fundamental for human populations that inhabited the continent from the early Holocene to the present day (Bonavia, 2008; Mengoni Goñalons and Yacobaccio, 2006; Olivera and Elkin, 1994; Yacobaccio and Vilá, 2013). In the past, Andean societies hunted wild camelids (Custred, 1979; Dedenbach-Salazar Sáenz, 1990; Kent, 1982; Tomka, 1992; Wheeler, 1999; Aschero and Martínez, 2001; Grant, 2017; López, 2008; Moreno, 2011; Olivera, 1997) and herded domesticated ones, establishing technological, spatial, and symbolic ties with them (Grebe, 1984; Arnold et al., 1992; Van Kessel and Condori Cruz, 1992; Lecoq and Fidel, 2003; Yacobaccio, 2010; Núñez and Nielsen, 2011; Vilá, 2014; Gabelman, 2015; Berenguer and Martínez, 2016; Arnold and Yapita, 2018). Archaeological evidence from various regions of the Andes suggests that camelid domestication occurred approximately 6,000 to 4,000 years ago, laying the foundation for pastoral economies and the establishment of long-distance trade networks via llama caravans (Mengoni Goñalons and Yacobaccio, 2006). This domestication process brought about significant lifestyle changes across the Andean region (Yacobaccio and Vilá, 2013).
The European conquest and colonization that started in the 15th century caused substantial changes in Andean societies. Among these was the introduction of European livestock, including cattle, horses, goats, sheep, and pigs, which posed challenges largely unknown to the local populations as well as wild and domestic SAC. European populations settled mostly in the plains and valleys, showing less interest in the Andean highlands except for the exploitation of certain resources, such as minerals. As a consequence, SAC were confined to the highlands, where competition was less intense given the difficulties European livestock faced in adapting to high-altitude environmental conditions (Bonavia, 2008; Miller and Gill, 1990; Wheeler, 1991; Vilá, 2012). This fostered the persistence of traditional pastoral practices in high Andean landscapes involving both SAC domesticated species (Caracotche, 2001; Flores Ochoa, 1968, 1977; Göbel, 2002; Tomka, 1992).
This contextualization, albeit brief, shows how historical landmarks have influenced the main conceptualization of the intertwined history of humans and SAC. Traditional models of domestication and herding were developed based on ethnohistorical and ethnographic data, alongside archaeological evidence, primarily focused on high Andean landscapes (Flores Ochoa, 1968, 1977; Webster, 1973; Browman, 1974; Gundermann, 1984; Kuznar, 1991; Tomka, 1992; Nielsen, 1997-1998; Yacobaccio, 2007; Caracotche, 2001; Göbel, 2002; Berenguer, 2004; Wawrzyk and Vilá, 2013). These studies were largely inspired by Murra's (1972) model for the Central Andes, which posits that human subsistence and social reproduction relied on economic interactions between environments at different altitudes. According to this model, the arid highlands were primarily used for llama and alpaca herding, considered as the most suitable economic practice for the region. Additionally, these areas provided wild resources, such as guanacos and vicuñas, and became the focus of extensive research on hunting strategies and associated technologies (Custred, 1979; Dedenbach-Salazar Sáenz, 1990; Kent, 1982; Tomka, 1992; Aschero and Martínez, 2001; Moreno, 2011). SAC are uniquely adapted to these conditions, making their breeding and herding a fitting productive practice for these areas. In some instances, herding was seen as the only possible economic activity in such landscapes. The associated development of caravan exchange and the circulation of goods between different ecological zones were considered as crucial for social reproduction (Browman, 1980; Nielsen, 2006; Núñez and Dillehay, 1995).
Recent studies, however, have challenged this proposal, suggesting that herding practices may have also occurred in areas of greater biodiversity—regions where SAC are no longer present today due to the aforementioned factors (Bonavia, 2008; Moreno et al., 2022; Shimada and Shimada, 1985; Szpak et al., 2014). One example of this is the eastern border of the Andes in Argentina, which spans the eastern foothills of Northwestern Argentina (NWA) and extends westward to the semi-arid Chaco plains. This area is often seen as a frontier for complex pre-Hispanic cultural developments, primarily concentrated in the Intermontane valleys region (Figure 1).
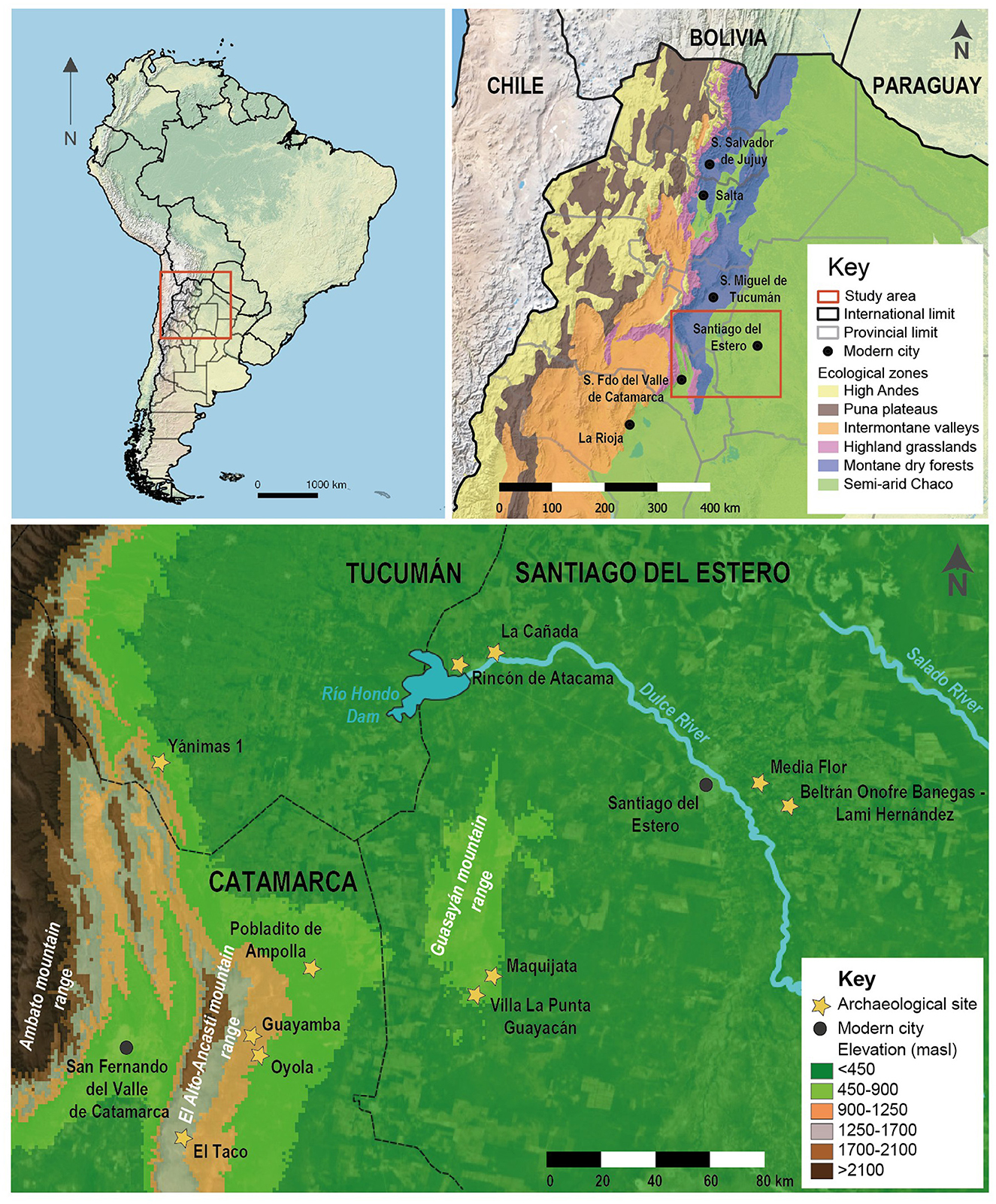
Figure 1. Location of the study area and main archaeological sites and microenvironments mentioned through the text.
This paper aims to explore the management models of domestic SAC outside traditional Andean environments in NWA, critically examining ecological models that assume productive practices are solely determined by local environmental characteristics. Drawing on several case studies, we will discuss analytical strategies for identifying domestic species and propose novel management models and herding production strategies for these regions. By integrating information from multiple lines of evidence and diverse zones within the study area, we will analyze similarities and differences to provide meaningful insights into this issue. Our focus will be on the first millennium of the Common Era (CE), a period during which most of the evidence from the case studies is concentrated, although we will make some references to periods both preceding and following this timeframe. This article's primary contribution lies in integrating data from several recently excavated archaeological sites, cross-referencing previously published information, and systematizing it within a unified framework. It also develops a theoretical interpretive approach to modeling camelid management within the study region. Additionally, it offers a new perspective on pastoralism, challenging preconceived notions about productive and economic systems in these environments, which have traditionally been viewed as peripheral to cultural developments in the pre-Hispanic Andes. Ultimately, this proposal is based on the concept of alternative management, where various production strategies are integrated in the same space, in which herding takes place on a small scale—i.e., small herds managed in spaces neighboring habitation areas—leveraging the micro-regional environmental diversity. This paper also aims to serve as a starting point for this proposal, comparing nearly all the information obtained from faunal assemblages in the region for the chronological period of interest and proposing an integrative approach that does not overlook local particularities. In the future, we aim to incorporate additional relevant data from other analytical techniques, some of which have been used only sparingly or are currently under development, such as stable isotope analysis (Moreno et al., 2022) or collagen peptide mass spectrometry fingerprinting (ZooMS) (Buckley et al., 2009; Pilaar Birch et al., 2019).
2 South American camelids and narratives about the past in Northwestern Argentina
There is a substantial body of archaeological, ethnohistorical, historical, and ethnographic literature on Andean pastoralism, particularly regarding llamas. The classic model typically emphasizes mobility between different altitudinal levels, over 3,000 meters above sea level (masl), following an annual cycle determined by the availability of pasture and water. However, literature on camelid pastoralism outside the high Andes is more limited. This underscores the importance of reconsidering proposals that emphasize the special adaptation of SAC to high Andean environments. While these animals are well-suited to the high-elevation Puna plateaus, they also thrive in other environments, as demonstrated by various studies across South America (Ayala Vargas, 2018; Browman, 1974; Marcoppido and Vila, 2013; Quispe Peña, 2011; Troll, 1958; Urquieta and Martinez, 1992).
In NWA, llamas and domesticated plants played a central role in shaping human settlement patterns over the last three millennia. Archaeological narratives typically refer to this period as the Formative Period (1,000 BCE to 1,000 CE approximately), which is marked by the development of diversified economic strategies—such as agriculture, herding, gathering, and hunting—and the creation of cultural landscapes where domestic structures were closely tied to productive territories (Korstanje et al., 2015). Subsequently, at the beginning of the second millennium, the Late Period was characterized by the consolidation of residential nuclei with greater architectural clustering than the one observed during the first millennium CE. Additionally, agricultural systems and productive practices were developed in response to population aggregation. These general characteristics, of course, exhibited local particularities shaped by social decisions and microenvironmental conditions. Later, in the 14th century, the process of Inka domination began in certain regions of Northwestern Argentina, lasting until the 16th century, when Spanish conquest and colonization fostered the disarticulation of Indigenous societies. The economic, social, and demographic impacts of this process were profound and catastrophic (Bethel, 1990; Todorov, 2007).
Beyond differences in settlement and production strategies, as well as micro-scale social processes, a key aspect is the importance of SAC for these populations. Camelid herding would have been a key strategy for producing food and other products, particularly in the Andean highlands and Puna plateaus. Studies of camelid management in NWA have largely followed this perspective, often replicating models from the broader South-Central Andes and focusing mainly on the western Puna highlands and the mesothermal valleys of Jujuy, Salta, Catamarca, and Tucumán provinces (Belloti López de Medina, 2015; Dantas et al., 2014; Göbel, 2002; Izeta, 2007; Izeta et al., 2009a; López, 2008; López et al., 2015; Mengoni Goñalons, 2013; Mercolli, 2010; Miyano, 2018; Olivera, 1991; Yacobaccio et al., 1997-1998). Outside these areas, research has primarily focused on guanaco hunting (Medina et al., 2014; Neme and Gil, 2008; Wolverton et al., 2015). The eastern slopes and plains of NWA have often been portrayed as a frontier that separated the agro-pastoral populations of the western Andes from the hunter-gatherer groups of the eastern lowlands (González, 1998). This narrative, largely shaped by both the region's environmental characteristics and the paucity of research, emphasized its role in providing wild resources and limiting the extension of Andean cultural and productive strategies. However, the eastern Andean slopes, which are the focus of this paper, present a promising case study. Our aim is to challenge these traditional narratives and explore the possibility of camelid herding in these so-called “marginal” regions (Ardissone, 1945; González, 1998; Kriscautzky, 1999; Pérez Gollán, 1991; Togo, 2005), which have been overlooked in the broader studies of the western Andean valleys and highlands.
This narrative has been challenged by recent systematic research conducted in the region, along with new theoretical and methodological perspectives that explore the ways people inhabited and produced in these territories. Multiple lines of evidence indicate the existence of established settlements with diversified economies and a significant investment of labor to modify the natural landscape, which span the last two millennia, at least until the Spanish conquest (Cruz, 2010; del Papa, 2020; Ortiz et al., 2017; Quesada et al., 2016; Taboada, 2017; Ventura, 2001; Zuccarelli Freire et al., 2022). While our primary aim is not to provide an extensive review of these proposals, we seek to contribute to the discussion by examining how local populations interacted with SAC, particularly in relation to the possibility of locally developed herding practices. In this regard, zooarchaeological studies provide valuable insights, highlighting the significance of SAC for past populations. In the eastern slope of the NWA Andes, in Jujuy province, Ortiz et al. (2017) identified camelid remains, osteometrically assigned to llamas and guanacos, at archaeological sites located in river terraces at altitudes below 1,000 masl and dated to the early CE. Unfortunately, the sample size was too small to determine whether these llamas were locally managed or acquired through exchange from high-altitude environments. In the eastern foothills of Tucumán province, the Yánimas 1 archaeological site reveals a high variability of faunal resources, including llamas, which are associated with herding activities. However, it remains unclear whether the herding occurred locally or if the animals were brought from elsewhere. Notably, wild species dominate the faunal assemblage from the site, with large mammals like deer and guanacos, as well as smaller species such as fish and reptiles (Nasif and Miguez, 2014). Eguía et al. (2016) propose that the scarce presence of camelid remains in archaeological sites at Guayamba in the El Alto-Ancasti mountain range (Catamarca province) could be explained by trade with populations from other areas, as this region may not have been favorable for SAC breeding. The presence and use of domestic SAC in the lowlands of Santiago del Estero province have sparked ongoing debate. Some researchers support their presence based on early colonial documents (e.g., Gramajo de Martínez Moreno, 1978; Palavecino, 1948; Raffino, 1975; Togo, 2004, 2005), while others argue that llamas were restricted to highland areas, suggesting that ecological and ethological constraints would have prevented its adaptation to lowland environments (Cione et al., 1979; Lorandi, 1978, 2015; Lorandi and Lovera, 1972).
One final aspect we wish to address is the frequent presence of rock art representations of SAC in the eastern slopes of the Andes. These images, often associated with anthropomorphic figures, highlight significant and longstanding human-camelid relationships in the region (Gheco, 2017, 2020; Gheco et al., 2019; Taboada and Rodríguez Curletto, 2022). The connection between rock art and the understanding of human-animal relationships requires further investigation and in-depth analysis. Nevertheless, it is crucial to acknowledge the importance of these representations in the region, especially the abundance, diversity, and complexity of SAC depictions, which illustrate a range of actions, designs, and interactions with other beings (Figure 2). These representations offer valuable insights into the central role of camelids in the lives of local populations.

Figure 2. Some examples of the relationships between humans and camelids in rock art from the eastern Andes. (A) digital reproduction of figures found at Ampolla archaeological site (Taboada and Rodríguez Curletto, 2022), (B–D) cases documented in the locality of Oyola (Gheco, 2017, 2020).
3 Environmental characteristics and archaeological cases of the eastern Andes in Northwestern Argentina
The case studies presented here encompass several locations, including the El Alto-Ancasti mountain range in the eastern region of Catamarca province, as well as the Guasayán mountain range and the Dulce River basin, situated respectively in the western and central areas of Santiago del Estero province, Argentina. These areas' unique environmental and cultural characteristics are essential for understanding human occupation patterns and the role of South American camelids in these contexts (see Figure 1).
The El Alto-Ancasti mountain range stretches approximately 140 km in a north-south direction, forming the eastern Andean foothills. Its topography is notable for a steep western slope, which descends toward the central valley of Catamarca, while the eastern slope is less steep, starting at around 2,000 masl in the highest areas and gradually descending to 500 masl at the western edge of the semi-arid Chaco plain. This topographical variation creates diverse microenvironments, which have been historically interpreted as supporting specific and specialized functions. In the higher elevations (up to about 1,500 masl), a highland grassland predominates, with sparse vegetation primarily consisting of grass species from the genera Stipa and Festuca. A second microenvironment develops between 1,200 and 700 masl, featuring a distinct vegetation profile, characterized by the presence of large trees. The cebil tree (Anadenanthera colubrina) is a prominent indicator species in this area, alongside other tree species such as algarrobo negro (Neltuma nigra), algarrobo blanco (Neltuma alba), and orco quebracho (Schinopsis haenkiana). Finally, around 500 masl, a dry forest microenvironment is present (Morlans, 1995). The most abundant species in this forest include quebracho blanco (Aspidosperma quebracho-blanco), mistol (Ziziphus mistol), algarrobo (Neltuma sp.), and quebracho colorado (Schinopsis balansae) (Zuccarelli Freire, 2020). In the El Alto-Ancasti mountain range, our research focuses on three case studies across three ecological zones: the high-altitude grasslands in El Taco, the montane dry forest in Oyola, and the semi-arid Chaco ecological zone in the northern part of the range, near Ampolla. Our case studies capture the environmental diversity of the region (Figure 1) and provide an opportunity to compare different zooarchaeological assemblages dated to the first millennium CE (Figure 3), which are relatively scarce for the region.

Figure 3. Chronological dates of the archaeological sites analyzed. The dates were calibrated with OxCal online software (version 4.4) using the Southern Hemisphere calibration curve SHCal20 (Hogg et al., 2020).
El Taco is situated within the highland grasslands and exhibits a settlement pattern that connects domestic compounds with agricultural terraces. This pattern has been identified in other environmentally similar regions within the study area (Dlugosz, 2005; Eguía et al., 2016; Gordillo et al., 2017; Quesada et al., 2012, 2016; Zuccarelli Freire, 2020; Zuccarelli Freire et al., 2022). Initially, this type of cultural landscape was conceptualized as primarily focused on herding, due to its environmental characteristics. However, it is now evident that this area was also intended for long-term habitation, requiring significant labor investments. Systematic surveys have uncovered numerous domestic compounds and agricultural terraces, suggesting the presence of integrated residential settlements with clearly defined domestic zones (Quesada et al., 2012; Zuccarelli Freire et al., 2022). Excavations at one of these compounds, El Taco 19 (ET19) (Figures 4A–C), have yielded faunal remains that offer valuable insights into animal consumption and use during the latter half of the first millennium CE (Egea and Moreno, 2021; Gastaldi et al., 2023; Moreno et al., 2022; Quesada et al., 2012). El Taco 19 has been interpreted as a large domestic compound consisting of 27 enclosures constructed with stones of varying shapes and sizes. Two of these enclosures were excavated, revealing walls that follow a particular construction sequence: the base features a double row of vertical stone slabs embedded into the sediment, which can reach a height of 1 meter. The sequence is completed by the placement of irregular blocks, though with the more regular sides selected to face the interior of the enclosure. The gaps between the slabs were filled with earth and rubble, while mortar was observed in the case of the blocks that complete the walls. Enclosure 1 measures approximately 5 meters on each side, with an opening facing south, while Enclosure 2 is larger and appears to have been open toward the south. It has been interpreted as a possible open space, likely unroofed or partially roofed (Gastaldi et al., 2023; Quesada et al., 2012).

Figure 4. Archaeological sites at El Alto-Ancasti mountain range: (A) Plan of El Taco 19 archaeological site, (B, C) Photographs of El Taco 19, (D) External view of Oyola 7 rockshelter, (E) Interior view during excavation of Oyola 7, (F) Plan of the Pobladito de Ampolla site showing the location of Ampolla 1, 8, 9, and 10 structures (A1, A8, A9, and A10) (modified from Salvatore, 2016). (G) Excavation of Ampolla 8 structure.
Oyola, located at the montane forest, is notable for its pre-Hispanic rock art (see Figures 2B–D), which features anthropomorphic, zoomorphic, and geometric figures typical of the region. These motifs have been linked to a possible ritual use, associated with the presence of cebil with hallucinogenic properties (Gheco, 2020). However, beyond the significant rock art records, intensive surveys and local knowledge of current residents led us to domestic compounds associated with agricultural terraces, evidence that suggests the configuration of a cultural landscape similar to that described for the highland grasslands (Quesada et al., 2012, 2016; Zuccarelli Freire, 2020; Zuccarelli Freire et al., 2022). To explore these occupations further, we excavated two domestic compounds (Oyola 50 and Oyola 51) and one rockshelter (Oyola 7) (Figures 4D, E). The compounds exhibit several similarities to ET19, not only in terms of chronology but also in the activities that took place there, which suggest similar forms of production and social reproduction despite microenvironmental differences (Gastaldi et al., 2023; Zuccarelli Freire et al., 2022). In contrast, the excavation of the rockshelter indicates an intermittent mode of occupation during the first millennium CE. Periods of greater intensity feature activities such as the use of fire and food consumption, while periods of lower occupation intensity are marked by occupational levels covered by sediments, possibly reflecting site disuse or abandonment (Gastaldi et al., 2016; Gheco, 2017; Gheco et al., 2019).
To the west of the previous area, in the semi-arid Chaco, the archaeological site of Pobladito de Ampolla (PDA) reveals itself as a place of residence and social reproduction, challenging the view of this area as merely transitional or peripheral (Taboada, 2011). The site comprises approximately twenty independent features, including various stone structures, isolated walls, small stone-covered mounds, larger earthen mounds, and 19 bedrock mortars (Figure 4F). These structures cover an area of about 8 hectares (400 × 200 meters), with features spaced 15 meters apart on average. This layout suggests a cohesive residential settlement with distinct domestic compounds, refuse disposal areas, grinding zones, and other spaces of undetermined function. Radiocarbon dates (see Figure 3) and associated ceramic materials indicate that PDA was occupied from the early first millennium to the end of that period or the beginning of the second millennium CE (Taboada, 2017; Taboada and Rodríguez Curletto, 2022). The information presented here derives from three excavated structures. Ampolla 8 is a semi-subterranean structure measuring 9 × 6 meters, primarily constructed from rock and earth. It shows possible signs of remodeling and partial roofing (Figure 4G). Evidence of domestic activities was found there, including combustion structures, burnt ceramic fragments, lithic artifacts (such as grinding and percussion tools), quartz debitage, beads, and faunal remains with cut marks and thermal alterations (Salvatore, 2016; Mercolli and Taboada, 2016). Ampolla 9 is a smaller stone structure featuring two intersecting walls built with unselected, rounded rocks. The limited material recovered—mostly ceramics, lithics, and faunal remains—and the absence of an identifiable occupation layer suggest it had a distinct, possibly non-domestic function. In contrast, Ampolla 10 appears to be a mound formed by the accumulation of cultural materials, including ceramic fragments, abundant faunal remains, lithics, beads, charcoal, and more. This structure is interpreted as a discard area.
Additionally, PDA includes both high- and medium-visibility rock art panels (Ampolla 1) dated to the site's period of use. These panels are located 300 meters southwest on a terrace containing 40 mortars, suggesting an integration of domestic and productive landscapes. The rock art depicts various animal motifs, including camelids, canids, and birds—species also represented in the archaeofaunal record. Camelid imagery exhibits signs of domestication, management, and/or care, such as depictions of harnesses and associations with human figures (see Figure 2A). These scenes likely reflect activities related to pastoralism (Taboada and Rodríguez Curletto, 2022).
In the case of Santiago del Estero province, its physiography consists of a vast loess sedimentary plain, interrupted at its southern, western, and northwestern edges by several mountain ranges, none of which reach significant elevations (Basualdo et al., 1985). This region lies within Argentina's semi-arid, continental subtropical zone, characterized by a scarcity of permanent water sources. Water is primarily concentrated in the province's two main rivers, the Dulce and Salado, as well as in springs and wells located near the mountain ranges. The Dulce and Salado rivers flow parallel to each other across the province from northwest to southeast, traversing low-gradient terrain. Their courses contribute to the formation of lagoons, marshes, swamps, and wetlands, particularly along the Salado River.
The Guasayán mountain range is a small, low elevation formation, approximately 80 km in length and 4 km in width at its widest point, with a maximum elevation of 790 masl (Roic and Villaverde, 1987). The eastern slope of the range is steeper than the western slope, resulting in higher rainfall on the eastern side and, consequently, denser vegetation. This vegetation is primarily composed of thorny bush clumps and cacti. For this area, we will consider the results obtained at two archaeological sites: Villa La Punta Guayacán (VP) and Maquijata (MJ). VP site is an open-air location lacking stone walls or artificial elevations, with human occupation evidence found in the natural profile of the creek (Figure 5A). Based on radiocarbon dates (Figure 3) and the homogeneity of the recovered materials, the site appears to reflect a short occupation period. Maquijata consists of a group of mounds that appear to have been reused over time and are closely associated with nearby water sources (Figure 5B). The mounds vary in height and size, forming a relatively extensive conglomerate. Radiocarbon dates place the occupation of the site at the transition between the end of the first millennium and the beginning of the second millennium CE.

Figure 5. Archaeological sites from Guasayán mountain range and Dulce River basin: (A) Excavation at Villa La Punta Guayacán (credits: José Togo), (B) General view of Maquijata (credits: José Togo), (C) General view of Rincón de Atacama, from one of the water spillways of Río Hondo dam (credits: José Togo), (D) General view of La Cañada (credits: José Togo), (E) Sketch of Beltrán Onofre Banegas-Lami Hernández archaeological site, (F) General view of a mound at Media Flor (credits: José Togo).
In contrast, the Santiago plain placed within the semi-arid Chaco, where the Dulce River flows, is characterized by a xerophytic and semi-deciduous forest. In the past, the forest canopy was predominantly composed of quebracho colorado santiagueño (Schinopsis lorentzii) and quebracho blanco (Aspidosperma quebracho-blanco) (Torrella and Adámoli, 2006). For this area, we will consider four case studies: Rincón de Atacama (RA), La Cañada (CA), Beltrán Onofre Banegas-Lami Hernández (BB), and Media Flor (MF). The RA site lies within the floodplain of the Termas de Río Hondo dam and is accessible only during drought periods. Although no visible structures reveal its full extent, the site was originally situated approximately 1,000 meters from the watercourse prior to the dam's construction in 1967 (Figure 5C). Evidence suggests that this area was occupied between 400 and 600 CE (Figure 3). CA is an archaeological site identified by the discovery of a large funerary urn, dated between 650 and 1,100 CE (Figure 5D). BB is characterized by the presence of mounds situated near a paleochannel of the Dulce River. At least four mounds have been identified, although there were likely more, as anthropogenic activities have leveled the terrain in recent years (Figure 5E). Occupations at this site have been dated to the Late Period (1,400–1,600 CE). MF site is located on the natural hillsides adjacent to a flood-prone area of the Dulce River (Figure 5F).
4 Materials and methods
Faunal remains from archaeological sites in the mentioned locations were analyzed using standard zooarchaeological methods. To assess the state of preservation of the faunal assemblages from the various archaeological sites, we took into account the different stages of weathering proposed by Behrensmeyer (1978). Taxonomic identification was carried out using reference samples and osteological manuals (Benavente et al., 1993; Olrog and Lucero, 1981; Pacheco Torres et al., 1979), taking into account factors such as skeletal part identification, laterality, and other relevant characteristics. For the quantification of bone specimens, species representation was estimated using NISP (Number of Identified Specimens per Taxon) (Grayson, 1984; Payne, 1975). Regarding the age profile, SAC were categorized into three groups based on bone fusion stages: early fusion (12–18 months), intermediate fusion (18–36 months), and late fusion (36–48 months). This classification follows the framework established by Mengoni Goñalons (2013), which builds on earlier works by Kaufmann (2009), Kent (1982), and Wheeler (1999). In this system, fused bone elements within each group indicate animals that survived beyond the specified age range (e.g., older than 12–18 months), but their exact age at death remains uncertain. Conversely, non-fused bone elements suggest that the animals died before reaching the particular age range (e.g., younger than 12–18 months). For the analysis of the identified skeletal parts, we considered the percentage value of Minimum Animal Units (%MAU) (Binford, 1984). This value is calculated by dividing the Minimum Number of Elements (MNE) by the number of times that element occurs in the skeleton. Since the objective is to analyze the relative importance of different anatomical parts, the values were standardized and expressed as percentages, with 100% assigned to the skeletal part with the highest MAU value (Mengoni Goñalons, 1988).
For interspecific identification of SAC, osteometric analysis was employed, a method widely used in the region to differentiate camelid species (Cartajena, 2009; Dantas, 2012; del Papa, 2020; Izeta et al., 2009b; Medina et al., 2014; Yacobaccio, 2010). This analysis involved measuring the maximum length and width of the proximal epiphysis of the 1st and 2nd phalanges recovered during excavation. The results were then compared with reference measurements for SAC (Izeta et al., 2009b; Mengoni Goñalons and Elkin, 2021; Mondini and Muñoz, 2017). It is important to note that, despite certain limitations, osteometry has proven to be a valuable tool for the taxonomic characterization of SAC, as discussed by del Papa (2020). Some of these limitations include dietary and environmental differences between reference samples and archaeological specimens (Cartajena et al., 2007), as well as the significant size variation observed within individuals of the same species, as demonstrated by Yacobaccio (2010) in the case of llamas. Nevertheless, osteometry remains a useful source of information, contributing to the integration of multiple lines of evidence.
It is important to highlight that some of the information presented here has been previously published by certain authors of this article. The composition of the samples from the various cases analyzed, along with the taxonomic and taphonomic analyses and osteometric data, can be examined in detail in those sources (Ahumada et al., 2015–2016; del Papa, 2012, 2020; del Papa et al., 2017; Mercolli and Taboada, 2016; Moreno and Ahumada, 2018; Moreno et al., 2022). In this article, this information has been systematized and consolidated through the standardization of criteria and the incorporation of additional analyses, such as the calculation of MAU% and osteometric comparisons, which introduce new data. Furthermore, previously scattered information—constituting virtually the only available zooarchaeological data for the region—has been integrated into a single dataset, facilitating new interpretations through comparative analysis. We consider this a foundational step toward advancing this line of research, with the future incorporation of methodologies we are currently exploring, such as ZooMS and stable isotope analyses, among others (Moreno et al., 2022).
5 South American camelids in the Argentinian eastern Andes
We will now present the zooarchaeological information obtained from the different case studies, with a particular emphasis on the importance of camelids in each case and, specifically, on the osteometric and interspecific identification data available.
5.1 Highland grasslands: El Taco
The faunal remains recovered from El Taco 19 archaeological site account for 1,029 skeletal fragments, which are generally very well-preserved (Figure 6), allowing for the taxonomic identification of 37% of the assemblage (Table 1). Notably, the highest percentages of identification correspond to the Camelidae family and the Artiodactyla order, together accounting for over 85% of the analyzed sample (Figure 7A) (Ahumada et al., 2015–2016; Moreno and Ahumada, 2018; Moreno et al., 2022). This underscores the significant role of large mammals, particularly SAC, within the subsistence strategies of the inhabitants of the site.
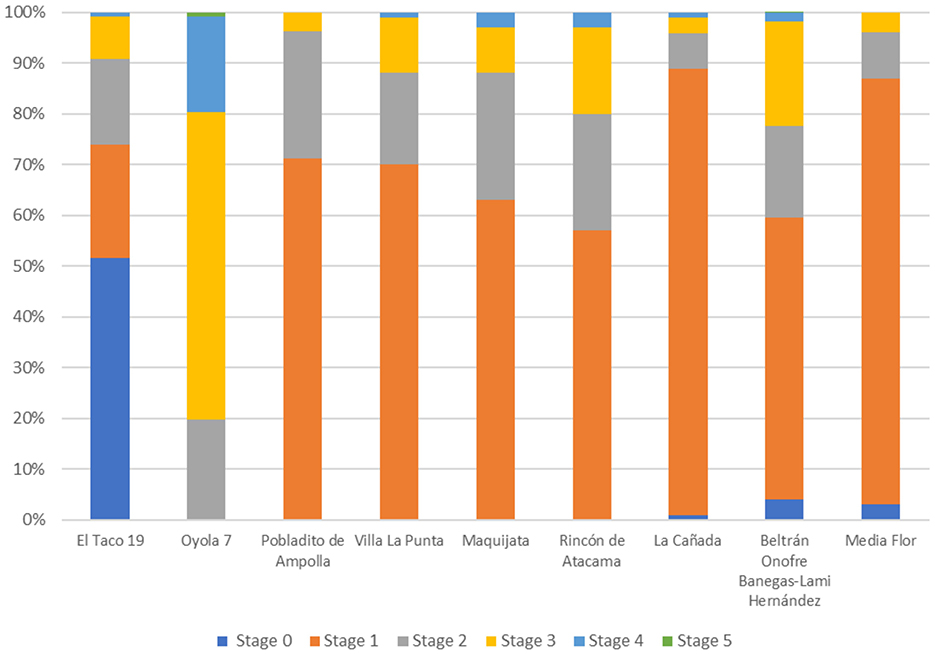
Figure 6. Weathering stages of the faunal remains of the archaeological sites considered, following Behrensmeyer (1978).
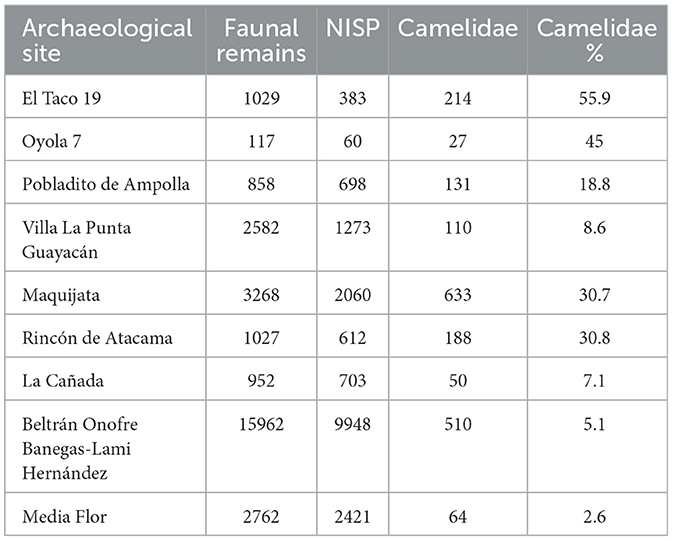
Table 1. Representation of the total number of recovered faunal remains, the number of identified specimens, and the percentage representation of South American camelids for each archaeological site considered here.
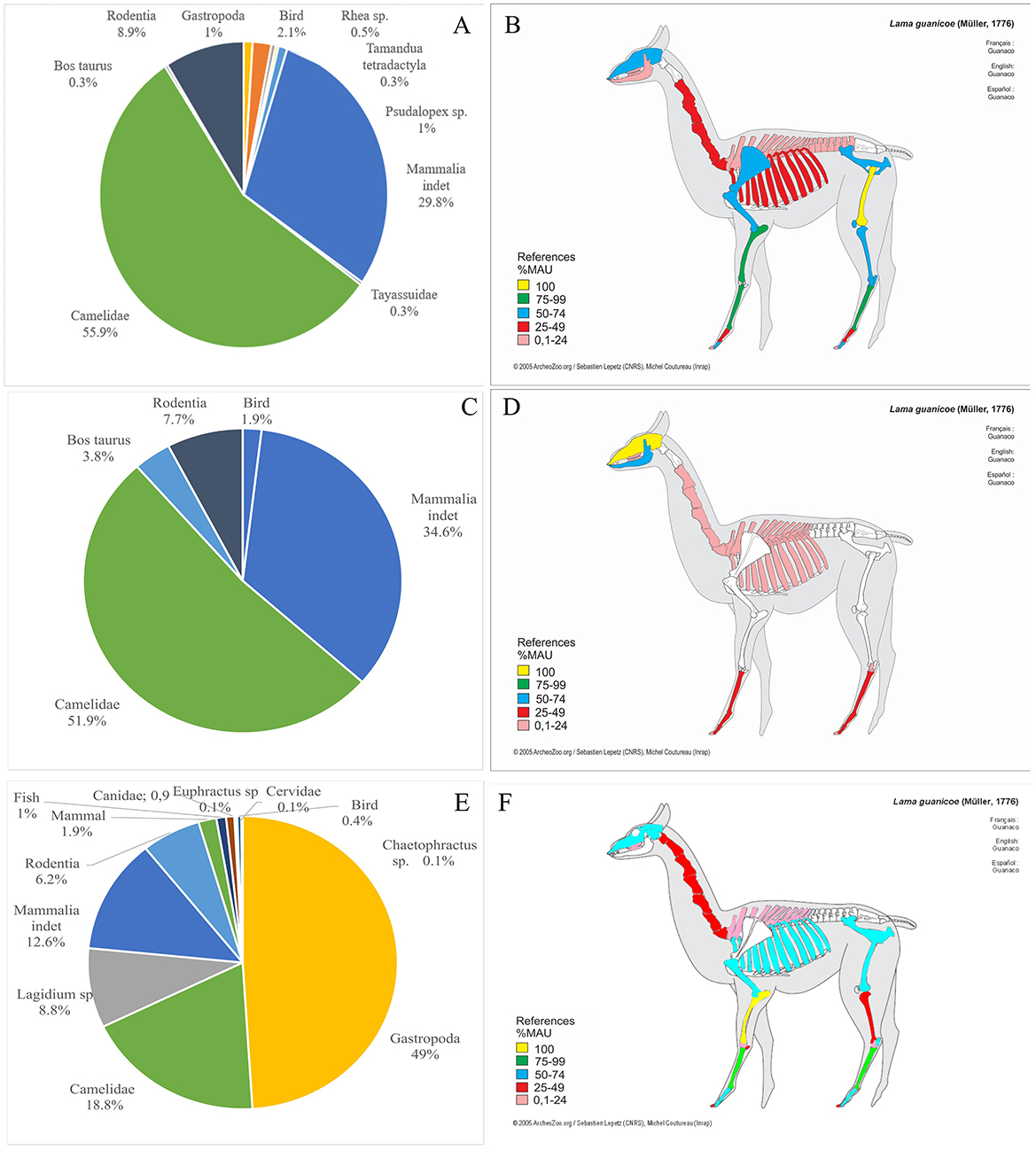
Figure 7. Left: Taxonomic representation identified in (A) El Taco 19, (C) Oyola 7, and (E) Pobladito de Ampolla faunal assemblages. Right: Representation of skeletal parts according to %MAU at (B) El Taco 19, (D) Oyola 7, and (F) Pobladito de Ampolla.
The results of the osteometric study of the first phalanges revealed three size groups, which likely correspond to small camelids (possibly vicuña), as well as larger groups that could be associated with llamas and guanacos (Moreno et al., 2022). Also, our analysis shows the presence of nearly all camelid skeletal parts, including both those with higher and lower economic yield (Figure 7B). This suggests that the slaughter of these animals likely occurred near ET19, as there would be no need to transport them, thus avoiding selective removal of skeletal parts (Binford, 1984; De Nigris and Catá, 2005; Mengoni Goñalons, 1999; Miller, 1979).
When evaluating the ages at death, we observe a relatively balanced distribution of newborns, young animals, and adults (Table 2). This pattern is typical of slaughter ages in herds managed for diverse purposes, suggesting the exploitation of both primary (such as meat) and secondary products (such as wool). Therefore, the information regarding the highland grasslands area aligns with the idea of a space where pastoral practices were carried out, not with a focus on specialization, but rather on a diversified economy. In this economy, agriculture, herding, gathering, hunting (including guanacos), and resource exchange with neighboring groups would have played key roles in supporting daily social reproduction in these spaces (Moreno et al., 2022; Quesada et al., 2012).
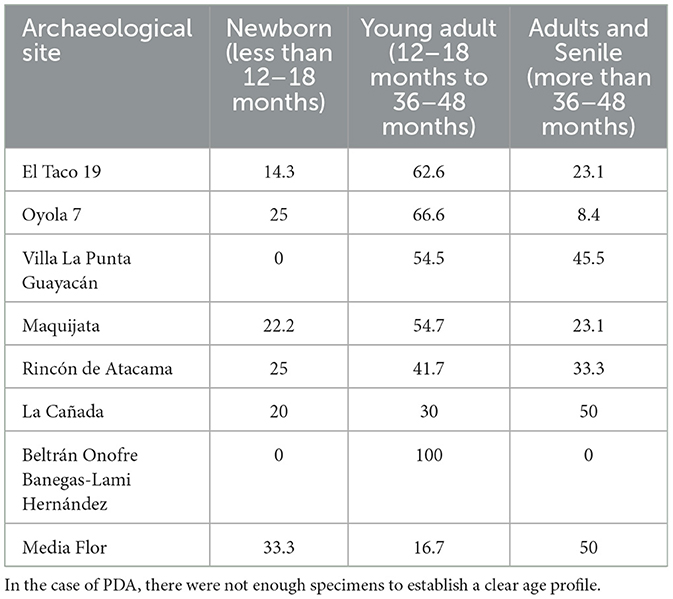
Table 2. Age profiles percentages representation for each archaeological site analyzed.
5.2 Montane dry forest: Oyola
The faunal assemblages recovered from the Oyola sites were very scarce and poorly preserved (Figure 6). However, some relevant data were obtained, particularly from the excavation of Oy7 (Moreno et al., 2022). The faunal remains consist of only 117 bone fragments, with camelids and other mammals predominating within the assemblage (Table 1, Figure 7C). Due to the absence of first and second phalanges, an osteometric analysis could not be conducted. However, analysis of the identified skeletal parts revealed the presence of skulls and the distal ends of both front and hind limbs—parts that are typically associated with a low economic yield (Figure 7D). While the zooarchaeological data from the Oyola sites may not provide as much detail as those from ET19, it still highlights the importance of SAC in the region (Moreno et al., 2022).
One aspect we were particularly interested in evaluating was the potential relationship between highland grasslands environments, which likely served as herding areas for SAC, and forest environments, which may have functioned as consumption areas. To explore this further, we conducted stable isotope analysis (δ13C and δ15N values) to compare the diets of SAC from ET19 and Oy7. The results of this analysis revealed differences in the diets of the animals consumed at each location. Based on these findings, it is likely that the breeding areas for the camelids at El Taco and Oyola were different, as would be expected if they were raised locally within their respective environments (Moreno et al., 2022).
5.3 Western semi-arid Chaco: Pobladito de Ampolla
The faunal remains analyzed were recovered from excavations in three structures at PDA: a sub-quadrangular structure (Ampolla 8), an indeterminate-shaped structure (Ampolla 9), and an earthen mound (Ampolla 10). These excavations were conducted in reduced areas of the three sectors of these archaeological sites, which explains the relatively small sample of faunal remains, as the overall state of preservation is good (Figure 6). The represented taxa show a predominance of gastropods; however, considering body size and dietary contribution, SAC are clearly the most important resource (Table 1, Figure 7E). Also, there is an important presence of viscachas, armadillos, canids, cervids, fish, birds, and undetermined eggshell fragments. Cervid remains were identified only in Ampolla 10, leading to the broader taxonomic category Artiodactyla being applied to bones that could not be definitively assigned to camelids or cervids. Osteometric analysis of five first phalanges allowed taxonomic differentiation of SAC, with four measurements indicative of llamas and one of guanaco. This identification is further supported by incisor morphology, which aligns with the anatomy of llamas and guanacos. Thus, both domestic and wild camelids appear to be represented, with osteometric data suggesting a higher prevalence of llamas. Despite the limited sample size, skeletal elements from both axial and appendicular regions are present (Mercolli and Taboada, 2016), indicating that inhabitants likely had access to whole animals rather than specific body parts (Figure 7F). This pattern suggests herding of domestic llamas, complemented by hunting of wild guanacos. Regarding the age at death estimates for camelids, it was not possible to establish a clear mortality profile due to the limited number of specimens wherein epiphyseal fusion could be observed. Additionally, a diverse range of faunal resources—including those from hunting, fishing, and gathering—was utilized. This evidence indicates that Pobaldito de Ampolla's inhabitants primarily relied on wild resources, supplemented by the use and consumption of domesticated camelids.
5.4 Guasayán mountain range
Villa La Punta Guayacán (VP) shows a faunal assemblage composed of 2,582 well preserved faunal remains (Figure 6), from which 1,273 were taxonomically assigned. A total of 110 specimens were identified as camelids, representing 8.6% of the specimens attributed to a specific taxon (Table 1), making camelids the third most abundant taxon after dolichotine (known as conejo de los palos) and other rodents (Figure 8A). Osteometric analysis at this site was limited to a single second phalanx, identified as Lama glama (del Papa, 2020). Camelid remains include various skeletal portions (Figure 8B) from both axial and appendicular regions, suggesting the processing of whole carcasses (del Papa, 2012). The age profile of the camelid remains shows a nearly balanced distribution between juvenile (54.5%) and adult (45.5%) animals, which may reflect a diversified subsistence strategy that relied on both primary and secondary products (Table 2).
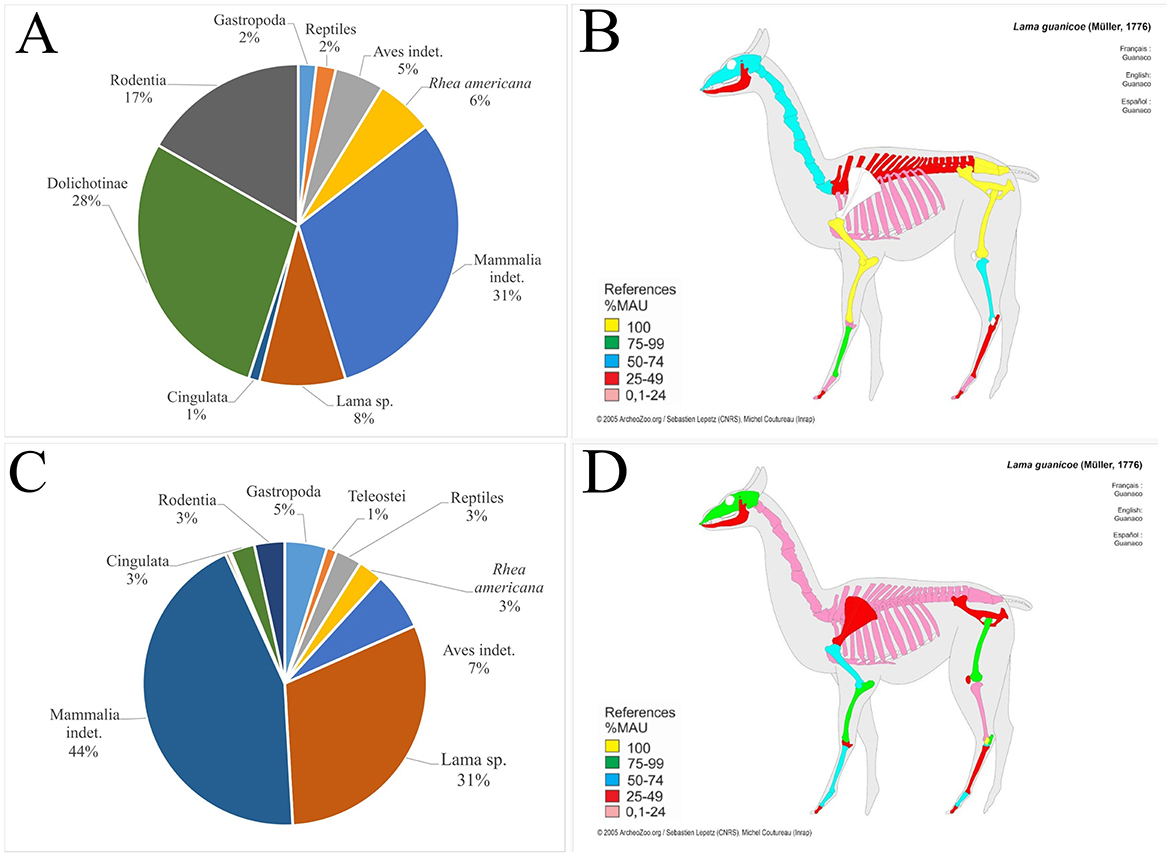
Figure 8. Left: Taxonomic representation identified in (A) Villa La Punta Guayacán and (C) Maquijata faunal assemblages. Right: Representation of skeletal parts according to %MAU at (B) Villa La Punta Guayacán and (D) Maquijata.
Analysis of the faunal remains obtained at Maquijata archaeological site revealed a well preserved assemblage (Figure 6), in which 30.7% of the taxonomically identified specimens belong to Camelidae (Table 1, Figure 8C). Osteometric analysis examined eight first phalanges and five second phalanges, identifying nine phalanges (both first and second) as Lama glama, one as Lama guanicoe, and three first phalanges as part of the llama-guanaco group (del Papa, 2015, 2020). Camelid remains at the site include a variety of skeletal elements from both axial and appendicular regions, suggesting the processing of whole carcasses (Figure 8D). The age profile is dominated by juvenile animals (Table 2), which may indicate a meat-sourcing strategy focused on young individuals or reflect calf mortality within herd management practices (del Papa, 2012). The predominance of camelid remains at the MJ site aligns with a more specialized subsistence strategy, likely reflecting an increased reliance on domesticated camelids during a period of favorable climatic conditions, with higher relative humidity and growing human populations (Drube, 2009).
5.5 Dulce River basin
In all the faunal assemblages of this region, the state of preservation is similar, with weathering stages 1 and 2 prevailing (Behrensmeyer, 1978), as shown in Figure 6.
In Rincón de Atacama, camelids account for 30.8% of taxonomically identifiable remains (Table 1, Figure 9A). Measurements of two first phalanges identified them as Lama glama (del Papa, 2020). The camelid remains include various skeletal elements, indicating whole-carcass processing (Figure 9B). The age profile is predominantly composed of sub-adult individuals (Table 2), which aligns with patterns observed at MJ, despite differences in the temporal contexts of their occupations.
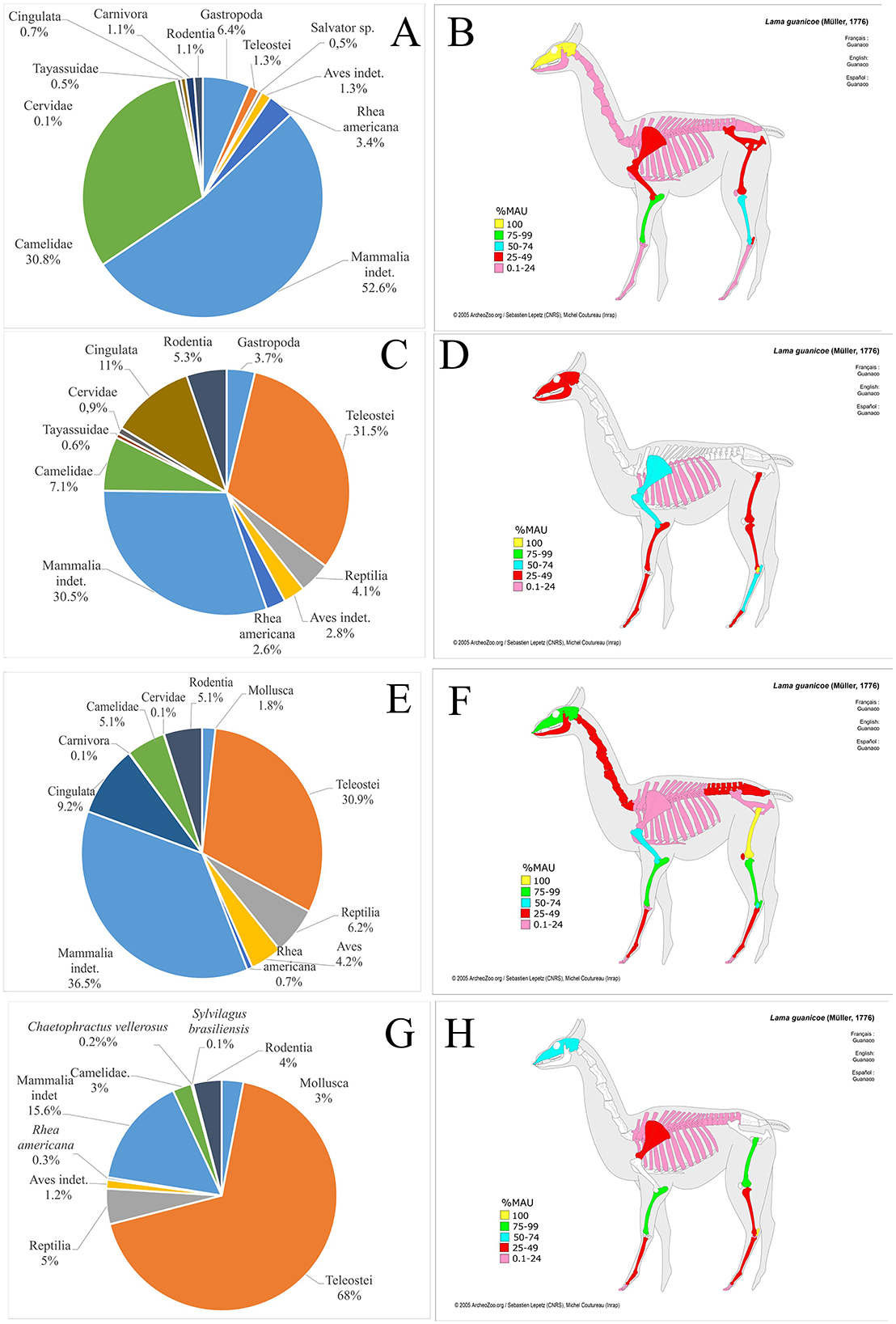
Figure 9. Left: Taxonomic representation identified in (A) Rincón de Atacama, (C) La Cañada, (E) Beltrán Onofre Banegas-Lami Hernández, and (G) Media Flor faunal assemblages. Right: Representation of skeletal parts according to %MAU at (B) Rincón de Atacama, (D) La Cañada, (F) Beltrán Onofre Banegas-Lami Hernández, and (H) Media Flor.
A total of 952 faunal remains were identified at the La Cañada archaeological site, with only 6.3% attributed to camelids (Table 1, Figure 9C). Fishes (Teleostei) were the most represented group. One second phalanx was measured and identified as Lama glama (del Papa, 2020). Camelid remains include various skeletal portions, indicating whole-carcass processing (Figure 9D). However, differential destruction—likely influenced by bone mineral density and potential fluvial transport—was observed (analysis in del Papa, 2012). The age profile is evenly split between young animals (including offspring and juveniles) and adults, suggesting a diversified strategy aimed at obtaining both primary and secondary products (Table 2).
At Beltrán Onofre Banegas-Lami Hernández, camelids account for only 5.1% of the identified faunal remains. This site is distinguished by the presence of a collection comprising 15,962 faunal remains, representing a significantly larger sample than those considered in other cases discussed here (Table 1, Figure 9E). Measurements of three first phalanges identified two as Vicugna vicugna and one as Lama guanicoe, while a second phalanx was assigned to Lama glama. The camelid remains include various skeletal portions, indicating whole-carcass processing (Figure 9F). The age profile is predominantly composed of sub-adult individuals (Table 2). The presence of vicuña underscores the complex social interaction networks established by the ancient inhabitants of Santiago del Estero during this period. This is supported by the fact that vicuñas are adapted to environments above 3,500 masl (Puna, Figure 1), but also by the increased presence of foreign material in later temporalities, such as marine shells and pottery (del Papa, 2012, 2020).
Finally, at Media Flor, camelids accounted for just 2.6% of the identified remains (del Papa, 2012) (Table 1, Figure 9G). Measurements of two first phalanges identified them as Lama guanicoe (del Papa, 2020). No llama remains were identified at this site; however, their absence may be attributed to the limited amount of osteological elements available for osteometric analysis. Camelid remains included various skeletal elements, suggesting whole-carcass processing (Figure 9H). The age profile is evenly split between young and adult animals, which may reflect a diversified strategy aimed at obtaining both primary and secondary products (Table 2).
The findings suggest that llamas were present throughout the analyzed sequence and across both areas of the semi-arid Chaco region. However, resource procurement through hunting and fishing played a significant role, with strategies adapting to the specific microenvironment studied. These strategies and their importance in social reproduction varied significantly, taking into account environmental particularities such as proximity to watercourses, as well as paleoenvironmental changes that could have influenced them (Cione and Tonni, 1981).
6 Discussion
The archaeological cases presented here offer new insights into the historical trajectories of human occupation and productive strategies in the regions studied, challenging traditional perspectives that viewed them as marginal areas in cultural development, solely specialized in the production of specific resources. Additionally, the new data discussed here allow us to revisit the relationship between humans and SAC, exploring the potential management of domestic SAC within this landscape. Camelid management appears to have been part of a diversified economy that incorporated agriculture, hunting, gathering, fishing, and other subsistence practices.
First, it is essential to address the marginal role traditionally attributed to environments located in the eastern foothills of the Andes. Recent studies are reevaluating the characteristics of the populations that inhabited this region, revealing greater complexity and a profound understanding of these environments. Notable examples include the Llanos de Moxos (Bolivia) and Amazonia (Brazil), among many others (Cruz, 2010; Jaimes Betancourt, 2012; Lombardo and Prümers, 2010; Neves, 2007; Ortiz, 2007; Rostain and Jaimes Betancourt, 2017; Valdez, 2013). For the specific case presented in this paper, evidence points toward a long term occupation (Figure 3). In some cases, long-lasting architecture was accompanied by the development of diversified economies that leveraged and enhanced local resources according to microenvironmental conditions and consistent efforts to modify the environment to particular productive strategies (del Papa, 2012, 2020; Quesada et al., 2012, 2016; Taboada, 2011; Taboada et al., 2013; Zuccarelli Freire et al., 2022). This challenges traditional narratives that emphasized the complementarity of different ecozones within mountain ranges while portraying the plains as exclusively inhabited by hunter-gatherer populations. Moreover, the concept of “periphery,” often used to describe pre-Hispanic human populations in South America, particularly in Northwestern Argentina, warrants reevaluation.
One noteworthy aspect is the relative synchrony of the occupations identified across all the cases discussed, which predominantly span the 1st millennium and part of the 2nd millennium CE. For the ET19 and Oy7 sites, the synchrony is almost exact, with dates ranging between the 5th and 10th centuries CE. PDA sites show slightly earlier occupations, beginning in the early 1st millennium for Ampolla 10 and somewhat later for Ampolla 8. In contrast, sites in Guasayán mountain range and Dulce River basin reveal a longer occupation period with notable differences, particularly in the use of faunal resources (see Figure 3). In Guasayán (e.g., MJ), the Late Period (1,200–1,400 CE) showed a predominance of camelids, a trend associated with population growth and the absence of water necessary for large-scale agriculture. During the Late Period in the Dulce River basin, population growth and proximity to the river likely encouraged the exploitation of other resources, such as fish, and possibly a shift toward agricultural predominance (Lorandi, 2015; Togo, 2005). Another significant aspect observed in this Late Period—both in PDA and the Dulce River basin—is an increase in the interregional circulation of objects, evidenced by the discovery of goods from distant regions, as well as indications of population growth (del Papa, 2012; Drube, 2009; Taboada, 2019; Taboada and Rodríguez Curletto, 2022).
The analyzed results reveal significant differences between the areas and sites studied, particularly in relation to the state of preservation of the various assemblages. One notable observation is the differential impact of environmental conditions on their preservation. In the case of the montane forest, faunal assemblages are more heavily affected by post-depositional processes, which significantly limits the potential for information recovery. Furthermore, substantial differences are observed in the taxa present and their relative relevance. The importance of camelids must be considered in relation to their body weight compared to other species. However, we observe that in some cases, particularly in the El Alto-Ancasti mountain range, camelids predominate considerably, whereas in other cases, such as BB or MF, other species are more prevalent. This discrepancy may be attributed to microenvironmental conditions and differential access to certain species. It may also be related to occupation patterns and historical changes.
Furthermore, these particularities underscore the importance of recognizing the diverse ways in which these populations inhabited the eastern Andean landscapes. Rather than adhering to a single model, their subsistence and settlement strategies were shaped by microenvironmental conditions and the diversification of social reproduction strategies. Focusing on the primary objective of this paper, SAC were clearly important for these populations, emerging as one of the primary taxa across the analyzed sites. In most cases, camelids were found alongside other species, including other large animals such as deer, peccaries, and foxes, as well as smaller animals like rodents, reptiles, armadillos, fish, and snails. This diversity highlights the wide range of animals utilized by these populations.
Common patterns are evident in the camelid remains, particularly in the representation of skeletal elements, which show no evidence of selective transport based on economic yield or other criteria indicative of intentional selection. The age profiles of camelids across most analyzed records reveal a balanced distribution of age classes, including juvenile, adult, and even senile individuals. These findings have been interpreted as evidence of domestic camelid herd management. This interpretation is supported by the expectation that, if these were wild animals, only the skeletal elements with higher economic value would typically have been transported from hunting sites to residential areas for consumption (Capriles, 2016; Miller, 1979). These findings provide evidence suggesting the presence of llamas in these assemblages. However, this does not preclude the presence of wild animals, i.e., guanacos, as indicated by osteometric data, particularly in cases where such analyses confirm the exclusive presence of guanacos, as observed at MF. This underscores the role of hunting in the subsistence strategies of populations with diversified economies. Guanaco hunting, along with the exploitation of other wild animals, appears to have been central to these populations. Given the challenges in clearly distinguishing llamas from guanacos using osteometric techniques—and the fact that none of the studies conducted allow for precise differentiation—the integration of multiple analytical approaches is essential for understanding the significance of domestic camelids in these assemblages. Future research in the region, combined with the application of new methodological tools, may further refine taxonomic identification among South American camelid species.
Moreover, the comprehensive processing of carcasses through a diversified strategy of primary and secondary resource utilization more closely aligns with patterns associated with domesticated animals. The balanced age profiles observed suggest herd management strategies aimed at obtaining both primary and secondary resources. Primary resources include meat, bones, and tendons, while secondary resources encompass wool and transport. However, in specific cases from the semi-arid Chaco region (e.g., MJ, RA, BB), sub-adults predominate (del Papa, 2012). This pattern has primarily been linked to a subsistence strategy focused on primary resources, as well as the constraints inherent in herd management practices.
Osteometry can shed light on the presence of SAC in this region through a comparative analysis of different sites. Figures 10, 11 present the measurements taken from the first and second phalanges, revealing three size groups. The smallest group likely represents vicuñas, which may have arrived in the region through trade or the circulation of goods, such as hides. The intermediate group corresponds primarily to indeterminate individuals considered as llama/guanaco, while the largest elements could indicate llamas, given that northern guanacos are generally smaller. Based on this analysis, we can propose the presence of both llamas and guanacos, which were likely consumed by the populations inhabiting these sites. However, the data raises an important question: were the domestic camelids breeded locally, or did they originate in other areas?
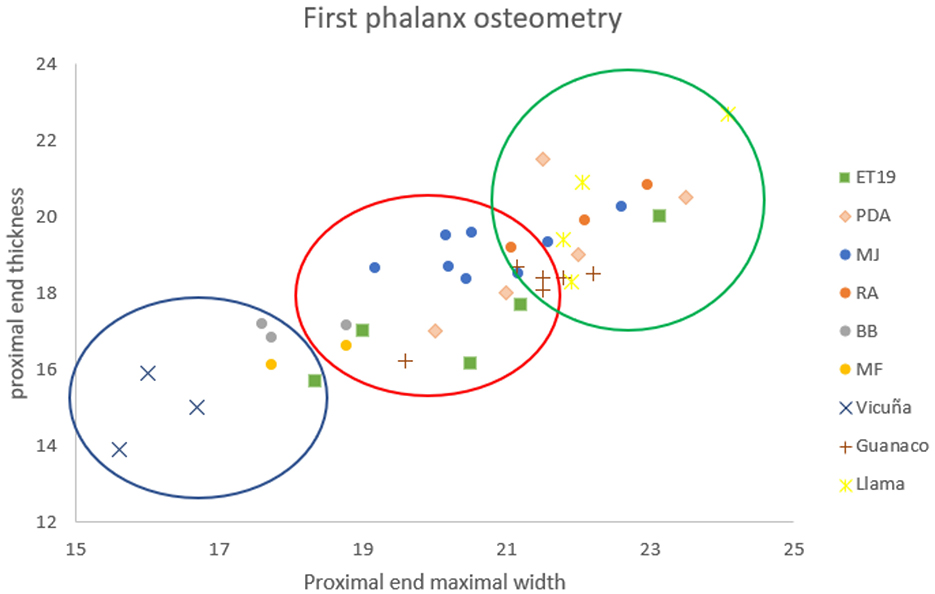
Figure 10. Comparative osteometry of first phalanges from various archaeological sites under consideration. Vicuña, guanaco, and llama measurements correspond to modern samples. All measurements are in mm.
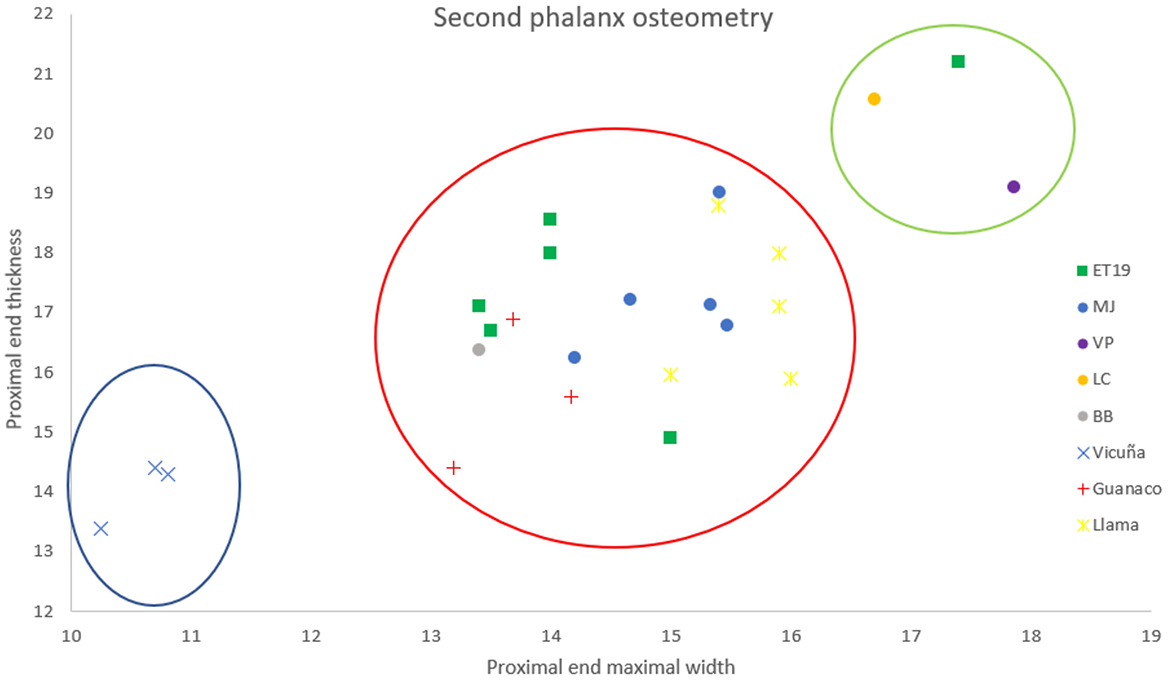
Figure 11. Comparative osteometry of second phalanges from various archaeological sites under consideration. Vicuña, guanaco, and llama measurements correspond to modern samples. All measurements are in mm.
Another indirect but significant source of information for understanding the interactions between SAC and humans in the eastern Andes is rock art, as highlighted in previous studies such as those carried out at Oyola and Pobladito de Ampolla (Gheco, 2017, 2020; Gheco et al., 2019; Taboada and Rodríguez Curletto, 2022). Many rock art sites depict not only anthropomorphic and camelid figures but also scenes of interaction, including the use of ropes or other binding tools. While this aspect requires further in-depth analysis, we include it here as an additional element to consider when exploring llama husbandry.
In this context, we propose the existence of local camelid breeding based on certain evidence, such as the osteometric information, the selection of skeletal parts, and the age profiles. Additionally, this hypothesis aligns with the idea of a diversified economy that incorporates various practices, one of which may have been herding. In this regard, one practice that likely played a significant role is hunting, as evidenced by the presence of various wild species, including vicuñas and possibly guanacos, as we noted previously (del Papa, 2012, 2020; Mercolli and Taboada, 2016; Moreno et al., 2022).
This brings us to the final subject we wish to address, that is herd management and specifically how this practice may have been carried out in the study area. To approach this question, we must first acknowledge the well-established general model for llama pastoralism. This model, supported by a wide range of archaeological, ethnohistorical, historical, and ethnographic sources, outlines the dynamics of herding practices, including mobility patterns, associated architectural features, labor organization, and ritual activities (Flores Ochoa, 1968, 1977; Webster, 1973; Browman, 1974; Gundermann, 1984; Kuznar, 1991; Tomka, 1992; Nielsen, 1997-1998; Yacobaccio, 2007; Caracotche, 2001; Göbel, 2002; Berenguer, 2004; Wawrzyk and Vilá, 2013). Here, we focus on two key aspects. The first one concerns the presence of structures associated with pastoralism, such as enclosures (corrals) and herding shelters (puestos) (Chaiavassa-Arias and Franco Salvi, 2024; Göbel, 2002; Yacobaccio, 2007; Yacobaccio et al., 1997-1998). In the study area, no stone corrals have been identified to date, though they may have been constructed using perishable materials. This possibility is supported by the abundant presence of bushes and trees in the region, which could have provided suitable building materials, as described in ethnohistorical references (Fernández, [1571]1914). Additionally, there is evidence of similar structures still in use in some parts of the area today. The second aspect relates to the herding dynamics specific to these environments. In the high Andean region, herding traditionally followed a mobility pattern linked to an annual cycle, dictated by the seasonal availability of pasture and water (Caracotche, 2001; Flores Ochoa, 1968, 1977; Göbel, 2002; Tomka, 1992). In contrast, the management of resources in the eastern Andes may have differed. It is possible that animals were allowed to roam freely across various spaces to graze on a mix of pastures, shrubs, and trees, reflecting an adaptation to the specific ecological conditions of this region.
Additionally, we believe it is important to consider the size of the herds that may have been managed by these populations. Commonly, large llama herds are associated with the specialization of pastoralism in the high Andes. This view reflects the environmental constraints of those regions, where herding is often seen as the primary adaptive strategy. In contrast, we propose that in the environments under study, characterized by the specific nature of the sites and their productive and reproductive dynamics, smaller herds were likely managed. These smaller herds would have been oriented toward self-sufficiency, meeting food and other resource needs on a more localized scale. This perspective considers the feasibility of managing smaller herds within a diversified economy, which would have included a variety of productive activities. For instance, strategies would have been necessary to prevent animals from grazing on cultivated areas, even if agricultural production in some cases might have been directed toward animal fodder.
A possible dynamic that may have occurred, particularly in the El Alto-Ancasti area, involves the exchange of animal parts among different domestic units. At El Taco, for example, a community-based settlement pattern has been proposed (Gastaldi et al., 2023; Quesada et al., 2012), where houses were situated within sight of one another and connected by marked paths. This spatial organization could have facilitated the exchange of resources. For instance, when an animal such as a llama was butchered, parts of it may have been shared or traded with neighboring households. This practice would have been especially practical in a warm and humid climate like this, where meat preservation poses serious challenges, unlike in the high Andes. Such exchange is a practice frequently carried out today: when an animal is slaughtered and butchered, neighbors are informed, and parts of the animal are sold.
Regarding the case studies presented here, we believe it is essential to expand the volume of data collected thus far and to generate more baseline information. Nevertheless, this work represents a significant initial step toward developing models that explore pre-Hispanic populations' interactions with their environments. These models not only provide insights into how different social groups in the past interacted, adapted, and formed communities but also shed light on how such processes persist in the present.
7 Final remarks
In this paper, we have explored the characteristics of pre-Hispanic SAC herding beyond the High Andean region. Our analysis focused on the eastern sector of the Andes, specifically the provinces of Catamarca and Santiago del Estero, in Argentina addressing two key discussion points. The first concerns the degree of determinism in how pre-Hispanic productive practices are understood in relation to local environmental characteristics. This perspective is closely tied to the integration of ecological zones and the adoption of similar productive strategies in comparable environments. The second discussion point, which builds on the first, examines the peripheral status often ascribed to regions like the eastern Andes within the broader narrative of pre-Hispanic cultural developments in the Andes.
Various studies, including our own, have challenged this perspective by highlighting the diverse strategies of social reproduction and economic production employed in these environments. These studies demonstrate how pre-Hispanic human populations adapted to these settings in ways that diverged significantly from previously proposed frameworks of domination and peripheral subordination to political-administrative centers (Cruz, 2010; del Papa, 2020; Ortiz et al., 2017; Quesada et al., 2016; Taboada, 2017; Ventura, 2001; Zuccarelli Freire et al., 2022).
Within this framework, the integration of diverse economic strategies and environmental interactions takes on particular significance, with camelid herding emerging as a key method for resource acquisition in these settings. Drawing on this theoretical and methodological approach, we have evaluated the available data to assess not only the presence of domesticated SAC in the study area but also their potential management practices. To this end, we synthesized information from various archaeological sites, where faunal remains underscore the importance of SAC. Osteometry is a crucial tool for interspecific differentiation among South American camelids. In the cases considered here, we have identified different size groups, which could primarily indicate the presence of llamas and guanacos. In addition to this analysis, zooarchaeological data—such as the representation of skeletal elements and age profiles—provided further evidence supporting the presence of domestic camelids in these assemblages, while also acknowledging the presence and significance of wild species.
Thus, the economic diversification of these populations is reinforced, as camelid husbandry, wild game hunting, gathering, agriculture, and other forms of production developed integrally during the first millennium and part of the second millennium CE.
In this context of diversified production, we propose a herding system centered on small herds with few animals, tailored to the needs of a family-based production scale. In this model, other productive activities would also contribute to securing the resources necessary for social reproduction. Notable differences emerge among the cases studied. In some areas, gathering or fishing may have held greater importance, while in others, agriculture likely played a more dominant role. These variations are further reflected in shifts in production and territoriality over time, particularly during the second millennium CE. Consequently, our proposal does not present a generalized model of land use and productive strategies for the eastern Andes. Instead, it highlights the need for detailed analyses and locally grounded perspectives to better understand the diverse ways of life of these populations.
Significant work remains to be done, particularly in the taxonomic differentiation of SAC species, which would enable more precise characterizations. Advancing new research approaches, such as stable isotope analysis, paleoproteomics, and paleogenetic studies, holds great potential for yielding valuable insights into the species represented in faunal assemblages. The integration of new evidence and innovative analytical techniques will be particularly relevant for taxonomic characterization and dietary analysis of the South American camelid populations under study. Moreover, perspectives such as those presented here—which aim to critically examine established narratives regarding productive strategies in different regions of Northwestern Argentina, particularly along the eastern Andean slopes—will contribute to the continued development of novel approaches to understanding human-animal relationships in these environments.
Data availability statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Author contributions
EM: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Validation, Visualization, Writing – original draft. CS: Conceptualization, Formal analysis, Investigation, Methodology, Resources, Supervision, Visualization, Writing – original draft. LdP: Formal analysis, Funding acquisition, Investigation, Methodology, Resources, Validation, Writing – original draft. CT: Formal analysis, Funding acquisition, Investigation, Resources, Validation, Writing – original draft. PM: Formal analysis, Investigation, Methodology, Writing – original draft, Validation. RN: Formal analysis, Investigation, Writing – original draft, Supervision, Validation. SS: Formal analysis, Writing – review & editing, Validation. NA: Methodology, Resources, Validation, Writing – review & editing, Investigation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was primarily funded on a bilateral cooperation program between CONICET (Argentina) and DAAD (Germany) that served as the framework for collaboration among some of the authors of this paper. The research was also funded by the Agencia Nacional de Promoción Científica y Tecnológica (Argentina) Projects PICT 2017-2589, PICT 2019-3043, PICT 2022-1021, PICT 2021-00034, and CONICET PIP 11220200100358CO and PIP 11/265.
Acknowledgments
We want to thank all members of the El Alto-Ancasti research group as well as the rest of our research teams. We are grateful to the local communities that supported our work and especially the residents and school of Ampolla. We also thank José Togo for the pictures used in this paper, and Patrick Roberts for the conceptualization and execution of the cooperation project between CONICET and DAAD.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahumada, M., and Moreno, E. (2015–2016) La escala doméstica y los animales. Tratamiento diferencial de partes esqueletarias y distribución diferencial intra-sitio en El Taco 19 (El Alto-Ancasti, Catamarca). Anal. Arqueol. Etnol. 70, 105–117. Available online at: http://hdl.handle.net/11336/107481
Ardissone, R. (1945). Las Pircas de Ancasti. Contribución al conocimiento de los restos de andenes en el noroeste de la Argentina. GAEA Anal. Soc. Argentina Estudios Geográficos 2, 383–416.
Arnold, D., Jiménez Aruquipa., D, and Yapita, J. (1992). Hacia un orden andino de las cosas. Tres pistas de los Andes meridionales. La Paz: Hisbol.
Arnold, D., and Yapita, J. (2018). Río de vellón, río de canto. Cantar a los animales, una poética andina de la creación. La Paz: Instituto de Lengua y Cultura Aymara.
Aschero, C., and Martínez, J. (2001). Técnicas de caza en Antofagasta de la Sierra, Puna meridional, Argentina. Relac de la Soc Argent de Antropol. XXVI, 215–241. Available online at: https://saantropologia.com.ar/wp-content/uploads/2015/01/Relaciones%2026/12-%20Aschero-Martinez%20O.pdf
Ayala Vargas, C. (2018). Los camélidos sudamericanos. Rev. Invest. Innov. Agropec. Recursos Natur. 5, 7–12. Available online at: http://www.scielo.org.bo/scielo.php?script=sci_arttext&pid=S2409-16182018000300003
Basualdo, M., Togo, J., and Urtubey, N. (1985). Aprovechamiento socioeconómico de la fauna autóctona de Santiago del Estero (inventario y uso popular más frecuente). Indoamérica 1. Laboratorio de Antropología, Facultad de Humanidades, Universidad Nacional de Santiago del Estero, Santiago del Estero, Argentina.
Behrensmeyer, A. F. (1978). Taphonomic and ecologic information from bone weathering. Paleobiology 4, 150–162. doi: 10.1017/S0094837300005820
Belloti López de Medina, C. (2015). Subsistence and economy at the Calchaquí valley (Salta, Argentina) during the Regional Developments Period (ca. 1000-1430 ad): zooarchaeology of Las Pailas locality. J. Archaeol. Sci. Rep. 4, 461–476. doi: 10.1016/j.jasrep.2015.10.016
Benavente, M. A., Adaro, L., Gecele, P., and Cunazza, C. (1993). Contribución a la determinación de especies animales en arqueología: familia Camelidae y taruca del norte. Santiago de Chile: Universidad de Chile, Vicerrectoría Académica y Estudiantil, Departamento Técnico de Investigación.
Berenguer, J. (2004). Caravanas, Interacción y Cambio en el Desierto de Atacama. Santiago de Chile: Sirawi Ediciones.
Berenguer, J., and Martínez, J. (2016). “Camelids in the Andes: rock art environment and myths,” in Animals into art, ed H. Morphy (Londres: Routledge), 390–416.
Bonavia, D. (2008). South American camelids. An expanded and corrected edition. Monograph 65. UCLA. Cotsen Institute of Archaeology Press.
Browman, D. (1974). Pastoral nomadism in the andes. Curr. Anthropol. 15, 188–196. doi: 10.1086/201455
Browman, D. (1980). Tiwanaku Expansion and altiplano Economic Patterns. Estudios Arqueol. 5, 107–120.
Buckley, M., Collins, M., Thomas-Oaies, J., and Wilson, J. C. (2009). Species identification by analysis of bone collagen using matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 23, 3843–3854. doi: 10.1002/rcm.4316
Capriles, J. (2016). “The subsistence economy of early Andean pastoralists: zooarchaeological insights from a Formative Period Wankarani House,” in The archaeology of Andean pastoralism, eds. J. Capriles, N. Tripcevich (Albuquerque: University of New Mexico Press), 39–54.
Caracotche, M. (2001). “The invisibility of time: an ethnoarchaeological study of the temporary sites of herders of the southern puna,” in Ethnoarchaeology of Andean South America: contributions to archaeological method and theory, ed L. Kuznar (Michigan: International Monographs in Prehistory), 97–115.
Cartajena, I. (2009). Explorando la variabilidad morfométrica del conjunto de camélidos pequeños durante el Arcaico Tardío y el Formativo Temprano en Quebrada Tulán, norte de Chile. Rev. Mus. Antrop. 2, 199–212. doi: 10.31048/1852.4826.v2.n1.5418
Cartajena, I., Núñez, L., and Grosjean, M. (2007). Camelid domestication on the western slope of the puna de atacama, northern Chile. Anthropozoologica 42, 155–173. Available online at: https://sciencepress.mnhn.fr/sites/default/files/articles/pdf/az2007n2a9.pdf
Chaiavassa-Arias, S., and Franco Salvi, V. (2024). El pastoreo en un paisaje aldeano del Valle de Tafi (Tucumán, Argentina). Arqueología 30:13395. doi: 10.34096/arqueologia.t30.n3.13395
Cione, A., Lorandi, A., and Tonni, E. (1979). Patrón de subsistencia y adaptación ecológica en la aldea prehispánica “El Veinte”, Santiago del Estero. Relac de la Soc. Arg. de Antropol. 13, 102–116.
Cione, A., and Tonni, E. (1981). First Record of some fishes and mammals from Santiago del Estero Province, Argentina. Stud. Neotrop. Fauna Environ. 16, 1–7. doi: 10.1080/01650528109360576
Cruz, P. (2010). Monte adentro. Aproximaciones sobre la ocupación prehispánica de la serranía de Calilegua (prov. de Jujuy). Intersecciones en Antropol. 11, 129–144. Available online at: https://www.ridaa.unicen.edu.ar/handle/123456789/1150
Custred, G. (1979). Hunting technologies in Andean culture. J. Soc. Am. LXVI, 7–19. doi: 10.3406/jsa.1979.2168
Dantas, M. (2012). Identificación interespecífica de camélidos en el Valle de Ambato (Catamarca, Argentina): una aproximación a la problemática desde distintas líneas de análisis. Rev. Mus. Antro. 5, 259–268. doi: 10.31048/1852.4826.v5.n1.9197
Dantas, M., Figueroa, G., and Laguens, A. (2014). Llamas in the cornfield. Prehispanic agropastoral system in the Southern Andes. Int. J. Osteoarchaeol. 24, 149–165. doi: 10.1002/oa.2351
De Nigris, M., and Catá, M. (2005). Cambios en los patrones de representación ósea del guanaco en Cerro de los Indios 1 (Lago Posadas, Santa Cruz). Intersecciones en Antropol. 6, 109–119. Available online at: https://www.scielo.org.ar/pdf/iant/n6/n6a09.pdf
Dedenbach-Salazar Sáenz, S. (1990). Inka Pachaq Llamanpa Willaynin. Uso y crianza de los camélidos en la época incaica. Estudio lingüístico y etnohistórico basado en las fuentes lexicográficas y textuales del primer siglo después de la Conquista. Bonn: Bonner Amerikanistische Studien, 16.
del Papa, L. (2012). Una aproximación al estudio de los sistemas de subsistencia a través del análisis arqueofaunístico en un sector de la cuenca del Río Dulce y cercanías a la Sierra de Guasayán. [doctoral thesis]. [La Plata, Buenos Aires]: Universidad Nacional de La Plata.
del Papa, L. (2015). First approach to study the presence of domesticated.camelids (lama glama) in the Chaco-Santiago region, a marginal zone of the South Central Andes. Int. J. Osteoarchaeol. 25, 45–60. doi: 10.1002/oa.2262
del Papa, L. (2020). Diferenciación de camélidos en la región Chaco-Santiagueña (Argentina) para el período Agroalfarero (350-1550 dC). Latin Am. Antiq. 2020, 1–17. doi: 10.1017/laq.2019.104
del Papa, L., de Santis, L., and Togo, J. (2017). “The fossorial faunal record at the Beltrán Onofre Banegas-Lami Hernández archaeological site (Santiago del Estero Province, Argentina): a taphonomic approach,” in Zooarchaeology in the Neotropics: Environmental Diversity and Human-Animal Interactions, eds. M. Mondini, S. Muñoz, P. Fernández (Cham: Springer), 137–156. doi: 10.1007/978-3-319-57328-1_9
Dlugosz, J. (2005). Prospecciones arqueológicas en los sitios Los Pedraza y Los Corpitos, Dpto. El Alto, Pcia. de Catamarca. [Degree thesis]. [San Miguel de Tucumán, Tucumán]. Universidad Nacional de Tucumán.
Drube, H. (2009). Las poblaciones aborígenes prehispánicas de Santiago del Estero: Evaluación de sus características bioantropológicas y de sus condiciones de salud, enfermedad y nutrición. [doctoral thesis]. [La Plata, Buenos Aires]: Universidad Nacional de La Plata.
Egea, D., and Moreno, E. (2021). Instrumentos líticos de cuarzo, prácticas sociales y vida campesina durante el 1° milenio de la era en el este de Catamarca, Argentina. Relac. de la Soc. Argent. de Antropol. XVL, 145–176. doi: 10.24215/18521479e005
Eguía, L., Prieto, C., and Gerola, I. (2016). Guayamba 2: abordando el espacio doméstico en los bosques orientales de Catamarca. Comechingonia 20, 43–72. doi: 10.37603/2250.7728.v20.n2.18044
Fernández, D. ([1571]1914). Primera parte de la historia del Perú. Libro 2. Madrid: Biblioteca Hispania.
Flores Ochoa, J. (1968). Los Pastores de Paratía: Una Introducción a su Estudio. México DF: Serie Antropología Social 10, Instituto Indigenista Interamericano.
Flores Ochoa, J. (1977). Pastores de Puna: Wywamichiq Punarunakuna. Lima: Estudios de la Sociedad Rural 5, Instituto de Estudios Peruanos.
Gabelman, O. (2015). Caminando con llamas. Caravanas actuales y analogías para el tráfico e intercambio prehispánico en Bolivia. Estudios Sociales del NOA 15, 33–58. Available online at: http://revistascientificas.filo.uba.ar/index.php/esnoa/article/view/1965
Gastaldi, M., Gheco, L., Moreno, E., Granizo, G., Ahumada, M., Egea, D., et al. (2016). Primeros resultados de las excavaciones estratigráficas en Oyola 7 (Sierra de El Alto-Ancasti, provincia de Catamarca, Argentina). Comechingonia 20, 73–104. doi: 10.37603/2250.7728.v20.n2.18045
Gastaldi, M., Quiroga Viñas, J., and Quesada, M. (2023). Devenir Casa. Temporalidades, Memorias e Historias de los Espacios Domésticos en la Sierra de El Alto-Ancasti (primer milenio d.C. Noroeste de Argentina). Relac de la Soc. Arg. de Antropol. 48, 288–315. doi: 10.24215/18521479e082
Gheco, L. (2017). El laberinto de las paredes pintadas. Una historia de los abrigos con arte rupestre de Oyola, Catamarca. [doctoral thesis]. [Córdoba]: Universidad Nacional de Córdoba.
Gheco, L. (2020). Una aproximación histórica al arte rupestre prehispánico de la sierra de El Alto-Ancasti (Provincia de Catamarca, Noroeste Argentino). Estud Atacameños 65, 263–290. doi: 10.22199/issn.0718-1043-2020-0030
Gheco, L., Gastaldi, M., Mastrangelo, N., Quesada, M., Marte, F., and Tascón, M. (2019). Entre humos, fuegos y pinturas: una metodología para conectar historias sobre una cueva con arte rupestre del sitio de Oyola (Catamarca, Argentina). Bol. Mus. Chil. De Arte Precolomb. 24, 131–152. doi: 10.4067/S0718-68942019000100131
Göbel, B. (2002). La arquitectura del pastoreo: Uso del espacio y sistema de asentamientos en la Puna de Atacama (Susques). Estud Atacameños 23, 53–79. doi: 10.4067/S0718-10432002002300005
Gordillo, I., Eguia, L., and Zuccarelli, V. (2017). “Las casas del sol naciente. Arqueología de la Vertiente Oriental del Alto-Ancasti,” in Arqueología de la Vertiente oriental Surandina. Interacción macro-regional, materialidades, economía y ritualidad, eds. V. Ventura, G. Ortiz, B. Cremonte (Buenos Aires: Sociedad Argentina de Antropología), 1–23.
Gramajo de Martínez Moreno, A. (1978). Evolución cultural en el territorio santiagueño a través de la arqueología. Santiago del Estero: Serie Monográfica N° 5. Publicación del Museo Arqueológico “Emilio Y Duncan Wagner”.
Grant, J. (2017). Of Hunting and Herding: Isotopic evidence in wild and domesticated camelids from the Southern Argentine Puna (2120–420 years BP). J. Archaeol. Sci. Rep. 11, 29–37. doi: 10.1016/j.jasrep.2016.11.009
Grayson, D. K. (1984). Quantitative Zooarchaeology: Topics in the analysis of Archaeological Faunas. Orlando, FL: Academic Press.
Grebe, V. (1984). Etnozoología andina: concepciones e interacciones del hombre andino con la fauna altiplánica. Estud Atacameños 7, 455–472. doi: 10.22199/S07181043.1984.0007.00032
Gundermann, H. (1984). Ganadería Aymara, ecología y forrajes: evaluación regional de una actividad productiva andina. Chungará 12, 99–124.
Hogg, A. G., Heaton, T. J., Hua, Q., Palmer, J. G., Turney, C. S. M., Southon, J., et al. (2020). SHCal20 southern hemisphere calibration, 0–55,000 years cal BP. Radiocarbon 62, 759–778. doi: 10.1017/RDC.2020.59
Izeta, A. (2007). Zooarqueología del sur de los valles Calchaquíes (Provincias de Catamarca y Tucumán, República Argentina): Análisis de conjuntos faunísticos del primer milenio A.D. Oxford: British Archaeological Reports. doi: 10.30861/9781407300542
Izeta, A., Laguens, A., Marconetto, M., and Scattolin, M. (2009a). Camelid handling in the Meridional Andes during the first millennium AD: a preliminary approach using stable isotopes. Int. J. Osteoarchaeol. 19, 204–219. doi: 10.1002/oa.1066
Izeta, A., Otaola, C., and Gasco, A. (2009b). Estándares métricos y variabilidad en falanges proximales de camélidos sudamericanos. Su importancia como conjunto comparativo para interpretaciones en arqueología. Rev. Mus. Antrop. 2, 169–180 doi: 10.31048/1852.4826.v2.n1.5416
Jaimes Betancourt, C. (2012). La cerámica de la Loma Salvatierra, Beni – Bolivia. La Paz: Ed. Plural.
Kaufmann, C. A. (2009). Estructura de Edad y Sexo en Lama guanicoe (Guanaco). Estudios actualísticos y arqueológicos en Pampa y Patagonia. Buenos Aires: Sociedad Argentina de Antropología, colección tesis doctorales.
Kent, J. (1982). The Domestication and exploitation of the South American camelids: methods of analysis and their application to circum-lacustrine archaeological sites in Bolivia and Peru. [Dissertation.]. [St. Louis, Missouri]: Washington University.
Korstanje, M. A., Lazzari, M., Basile, M., Bugliani, F., Lema, V., Pereyra Domingorena, L., et al. (2015). Crónicas Materiales Precolombinas. Arqueología de los Primeros Poblados del Noroeste Argentino. Buenos Aires: Sociedad Argentina de Antropología.
Kriscautzky, N. (1999). “Sociedades agropastoriles en la cadena del Ambato-Manchao, el caso de la quebrada de la Tala,” in Actas del XII Congreso Nacional de Arqueología Argentina, ed. C. Marín (La Plata: Editorial de la Universidad Nacional de La Plata), 89–91.
Kuznar, L. (1991). Herd composition in an aymara community of the peruvian altiplano: a linear programming problem. Hum. Ecol. 19, 369–387. doi: 10.1007/BF00888983
Lecoq, P., and Fidel, S. (2003). Prendas simbólicas de camélidos y ritos agro-pastorales en el sur de Bolivia. Textos Antropol. 14, 7–54.
Lombardo, U., and Prümers, H. (2010). Pre-Columbian human occupation patterns in the eastern plains of the Llanos de Moxos, Bolivian Amazonia. J. Archaeol. Sci. 37, 1875–1885. doi: 10.1016/j.jas.2010.02.011
López, G. (2008). Arqueología de cazadores y pastores en tierras altas. Ocupaciones humanas a lo largo del Holoceno en Pastos Grandes, Puna de Salta, Argentina. Oxford: BAR International series 1854. doi: 10.30861/9781407302317
López, P., Cartajena, I., Santander, B., Pavlovic, D., and Pascual, D. (2015). Camélidos domésticos en el Valle de Mauro (Norte Semiárido, Chile): múltiples análisis para un mismo problema. Intersecciones en Antropol. 16, 101–114. Available online at: https://www.ridaa.unicen.edu.ar/handle/123456789/1132
Lorandi, A. (1978). El desarrollo cultural prehispánico en Santiago del Estero, Argentina. J. la Soc. des Américanistes 65, 63–86. doi: 10.3406/jsa.1978.2156
Lorandi, A., and Lovera, D. (1972). Economía y patrón de asentamiento en la provincia de Santiago del Estero. Relac. de la Soc. Arg. de Antropol. 6, 173–191.
Lorandi, A. M. (2015). Tukuma-Tukuymanta: Los pueblos del búho: Santiago del Estero antes de la Conquista. Subsecretaría de Cultura de la Provincia de Santiago del Estero.
Marcoppido, G., and Vila, B. (2013). Manejo de llamas extra-andinas. Observaciones que contribuyen a su bienestar en un contexto no originario. Rev. Arg. de Producción Animal 33, 139–154. Available online at: https://aapa.org.ar/rapa/33/2/008-Fauna-Marcoppido%20y%20otros.pdf
Medina, M., Pastor, S., and Berberián, E. (2014). “Es Gente Fazil de Moverse de una Parte a Otra”. Diversidad en las Estrategias de Subsistencia y Movilidad Prehispánicas Tardías (Sierras de Córdoba, Argentina). Complutum 25, 73–88. doi: 10.5209/rev_CMPL.2014.v25.n1.45356
Mengoni Goñalons, G. (1988). Análisis de materiales faunísticos de sitios arqueológicos. Xama 1, 71–120.
Mengoni Goñalons, G. (1999). Cazadores de Guanacos de la estepa patagónica. Buenos Aires: Sociedad Argentina de Antropología, Colección Tesis Doctorales.
Mengoni Goñalons, G. (2013). “El aprovechamiento de la fauna en sociedades complejas: aspectos metodológicos y su aplicación en diferentes contextos arqueológicos del NOA,” in Al borde del imperio. Paisajes sociales, materialidad y memoria en áreas periféricas del noroeste argentino, eds. V. Williams, and M. B. Cremonte (Buenos Aires: Sociedad Argentina de Antropología), 311–396.
Mengoni Goñalons, G., and Elkin, D. (2021). Guanaco (Lama guanicoe) osteometric data from an individual from Northwest Argentina. Open Context. Available online at: http://openc ontext.org/projects/01860ee9-6fb6-4e71-8958-113fb852c850 (accessed May 31, 2022).
Mengoni Goñalons, G., and Yacobaccio, H. (2006). “The domestication of South American camelids: a view from the South-Central Andes,” in Documenting Domestication: New Genetic and Archaeological Paradigms, eds. M. Zeder, D. Bradley, E. Emshwiller, B. Smith (Los Angeles: University of California Press), 228–244.
Mercolli, P. (2010). “Estrategias de subsistencia en la Quebrada de Humahuaca Pcia. de Jujuy. Dos casos de estudio relacionados al manejo ganadero y la caza a través del tiempo en las sociedades humanas,” in Zooarqueología a principios del siglo XXI. Aportes teóricos, metodológicos y casos de estudio, eds. M. A. Gutiérrez, M. De Nigris, P. M. Fernández, M. Giardina, A. Gil, A. D. Izeta, G. Neme, H. Yacobaccio (Buenos Aires: Ediciones del Espinillo), 273–284.
Mercolli, P., and Taboada, C. (2016). Análisis de la fauna del sitio arqueológico El Pobladito de Ampolla (piedemonte catamarqueño, Argentina). Comechingonia 20, 127–152. doi: 10.37603/2250.7728.v20.n2.18047
Miller, G. (1979). An introduction to the ethnoarchaeology of andean camelids. [doctoral thesis dissertation]. [Los Angeles, California]: University of California, Berkeley.
Miller, G., and Gill, A. (1990). Zooarchaeology at Pirincay, a Formative Period Site in Highland Ecuador. J. Field Archaeol. 17, 49–68 doi: 10.1179/009346990791548510
Miyano, J. (2018). Humanos y animales en el oeste tinogasteño (Catamarca, Argentina): su relación a lo largo del tiempo desde el siglo IV a.C. hasta la actualidad. [doctoral thesis]. [Buenos Aires]: Universidad de Buenos Aires.
Mondini, M., and Muñoz, S. (2017). “Osteometrics of South-Central Andean Wild Camelids: New Standards,” in Zooarchaeology in the Neotropics: Environmental Diversity and Human-Animal Interactions, eds. M. Mondini, S. Muñoz, P. Fernández (Cham: Springer), 177–188. doi: 10.1007/978-3-319-57328-1_11
Moreno, E. (2011). La estructuración de paisaje cazador: Interrelaciones entre personas y vicuñas en el área del salar de Antofalla, Provincia de Catamarca, Argentina. Rev. Chilena de Antropol. 24, 63–96. Available online at: https://revistadeantropologia.uchile.cl/index.php/RCA/article/view/18160
Moreno, E., and Ahumada, M. (2018). Animales y humanos en las cumbres de Ancasti. Paisajes campesinos y recursos locales. Archaeofauna Int. J. Archaeozool. 27, 195–208. doi: 10.15366/archaeofauna2018.27.008
Moreno, E., Samec, C., and Ahumada, M. (2022). New insights into South American camelids management strategies at El Alto-Ancasti mountain range (Catamarca, Argentina) during the first millennium of the Common Era. Archaeol. Anthropol. Sci. 14:144. doi: 10.1007/s12520-022-01609-z
Morlans, M. (1995). Regiones naturales de Catamarca. Provincias geológicas y provincias fitogeográficas. Rev. Ciencia y Técnica 2, 1–36. Available online at: https://editorial.unca.edu.ar/Publicacione%20on%line/Ecologia/imagenes/pdf/006-fitogeografia-catamarca.pdf
Murra, J. (1972). “El control vertical de un máximo de pisos ecológicos en la economía de las sociedades andinas,” in Formaciones Económica y políticas del mundo andino, ed J. Murra. (Lima: Instituto de Estudios Peruanos), 59–115.
Nasif, N., and Miguez, G. (2014). La fauna relacionada a una comunidad prehispánica del piedemonte meridional de la Provincia de Tucumán (Argentina). Folia 22, 203–232. doi: 10.30972/fhn.02257
Neme, G., and Gil, A. (2008). Faunal exploitation and agricultural transitions in the South American agricultural limit. Int. J. Osteoarchaeol. 18, 293–306. doi: 10.1002/oa.944
Neves, E. (2007). El formativo que nunca terminó. La larga historia de estabilidad en las ocupaciones humanas de la Amazonía central. Boletín de Arqueol. PUCP 11, 117–142. doi: 10.18800/boletindearqueologiapucp.200701.005
Nielsen, A. (1997-1998) Tráfico de caravanas en el sur de Bolivia: observaciones etnográficas e implicancias arqueológicas. Relac. de la Soc. Arg. de Antropol. 22, 139–178.
Nielsen, A. (2006). “Estudios internodales e interacción interregional en los Andes circumpuneños: teoría, método y ejemplos de aplicación,” in Esferas de interacción prehistóricas y fronteras nacionales modernas: los Andes Sur Centrales, ed. H. Lechtman (Lima: Instituto de Estudios Peruanos), 29–62.
Núñez, L., and Dillehay, T. (1995). Movilidad giratoria, armonía social y desarrollo en los Andes Meridionales: patrones de tráfico e interacción económica. Antofagasta: Universidad Católica del Norte.
Núñez, L., and Nielsen, A. (2011). En ruta: arqueología, historia y etnografía del tráfico surandino. Córdoba: Encuentro Grupo Editor.
Olivera, D. (1991). “El Formativo en Antofagasta de la Sierra (Puna meridional Argentina). Análisis de sus posibles relaciones con contextos arqueológicos agro-alfareros tempranos del Noroeste Argentino y Norte de Chile,” in Actas de XI Congreso Nacional de Arqueología Chilena (Santiago de Chile: Museo Nacional de Historia Natural), Tomo II, 61–78.
Olivera, D. (1997). La importancia del recurso Camelidae en la Puna de Atacama entre los 10.000 y 500 años A. P. Estud Atacameños 14, 29–41. doi: 10.22199/S07181043.1997.0014.00004
Olivera, D., and Elkin, D. (1994). “De cazadores y pastores: El proceso de domesticación de Camélidos en la Puna Meridional Argentina,” in Zooarqueología de Camélidos. Perspectivas teóricas y metodológicas (First part), ed. Grupo de Zooarqueología de Camélidos (Buenos Aires: Grupo de Zooarqueología de Camélidos), 95–124.
Olrog, C., and Lucero, M. (1981). Guía de los mamíferos argentinos. San Miguel de Tucumán: Fundación Miguel Lillo.
Ortiz, G. (2007). La evolución del uso del espacio en las tierras bajas jujeñas (subárea del río San Francisco). [doctoral thesis]. [Córdoba, Argentina]: Universidad Nacional de Córdoba.
Ortiz, G., Mercolli, P., and Killian Galván, V. (2017). “Nuevas evidencias en el estudio de la economía y dieta en poblaciones tempranas de la cuenca del San Francisco (800 a.C−500 d.C.),” in Arqueología de la vertiente oriental Surandina: interacción macro-regional, materialidades, economía y ritualidad, eds. B. Ventura, G. Ortiz, M. Cremonte (Buenos Aires: Sociedad Argentina de Antropología), 51–78.
Pacheco Torres, V., Altamirano Enciso, R., and Guerra Porras, E. (1979). Guía Osteológica de Camélidos Sudamericanos. Lima: Universidad Nacional Mayor de San Marcos.
Payne, S. (1975). “Partial recovery and sample bias,” in Archaeozoological Studies, ed. A. T. Clason (Amsterdam: North Holland), 7–17.
Pérez Gollán, J. (1991). La cultura de La Aguada vista desde el valle de Ambato. Publicaciones Del CIFFyH 6, 157–173.
Pilaar Birch, S. E., Scheu, A., Buckley, M., et al. (2019). Combined osteomorphological, isotopic, aDNA, and ZooMS analyses of sheep and goat remains from Neolithic Ulucak, Turkey. Archaeol. Anthropol. Sci. 11, 1669–1681 doi: 10.1007/s12520-018-0624-8
Quesada, M., Gastaldi, M., and Granizo, G. (2012). Construcción de periferias y producción de lo local en las cumbres de El Alto-Ancasti. Relac. de la Soc. Argent de Antropol. XXXVII, 435–456. Available online at: https://www.saantropologia.com.ar/wp-content/uploads/2015/03/08-Quesada.pdf
Quesada, M., Zuccarelli, V., Gheco, L., Gastaldi, M., Boscatto, S., and Moreno, E. (2016). Paisaje y experiencia en Oyola a finales del Primer Milenio d.C. (Dpto. El Alto, Catamarca). Comechingonia 20, 13–42. doi: 10.37603/2250.7728.v20.n2.18043
Quispe Peña, E. (2011). Adaptaciones hematológicas de los camélidos sudamericanos que viven en zonas de elevadas altitudes. Rev. Complutense de Ciencias Veterinarias 5:0126. Available online at: https://www.produccion-animal.com.ar/produccion_de_camelidos/camelidos_general/131-altitudes.pdf
Raffino, R. (1975). Potencial ecológico y modelos económicos en el N.O. Argentino. Relac. de la Soc. Arg. de Antropol. 9, 21–43.
Roic, L., and Villaverde, A. (1987). “Antecedentes para una excursión botánica a la Sierra de Guasayán, provincia de Santiago del Estero,” in Libro de resúmenes de XXI Jornadas Argentinas de Botánica (Sociedad Argentina de Botánica, Santiago del Estero).
Rostain, S., and Jaimes Betancourt, C. (2017). Las siete maravillas de la Amazonía precolombina. La Paz: Plural editores.
Salvatore, B. (2016). La Historia Ocupacional de la Estructura Ampolla 8. Sitio Arqueológico El Pobladito de Ampolla en Tiempos Prehispánicos Tempranos. [degree thesis]. [Tucumán, Argentina]: Universidad Nacional de Tucumán.
Shimada, M., and Shimada, I. (1985). Prehistoric llama breeding and herding on the North Coast. Am. Antiq. 50, 3–26. doi: 10.2307/280631
Szpak, P., Millaire, J., White, C., and Longstaffe, F. (2014). Small scale camelid husbandry on the north coast of Peru (Virú Valley): insight from stable isotope analysis. J Anthropol. Archaeol. 36, 110–129. doi: 10.1016/j.jaa.2014.08.005
Taboada, C. (2011). Repensando la arqueología de Santiago del Estero. Construcción y análisis de una problemática. Relac. de la Soc. Arg. de Antropol. XXXVI, 197–219.
Taboada, C. (2017). “Espacio, cultura material y procesos sociales tardíos en la llanura santiagueña. Modelo para pensar las poblaciones de la región,” in Arqueología de la vertiente oriental Surandina: interacción macro-regional, materialidades, economía y ritualidad, eds. B. Ventura, G. Ortiz, M. Cremonte (Buenos Aires: Sociedad Argentina de Antropología), 237–266.
Taboada, C. (2019). Procesos sociales prehispánicos y pericoloniales en torno a los ríos Salado y Dulce (Santiago del Estero, Argentina). Rev. del Museo de La Plata 4, 511–540. doi: 10.24215/25456377e087
Taboada, C., Angiorama, C., Leiton, D., and López Campeny, S. (2013). En la llanura y los valles. Relaciones entre las poblaciones de las tierras bajas santiagueñas y el estado inca: materialidades, elecciones y repercusiones. Intersecciones Antropol. 14, 137–156. Available online at: https://www.scielo.org.ar/pdf/iant/v14n1/v14n1a08.pdf
Taboada, C., and Rodríguez Curletto, S. (2022). Entre el llano y las alturas. Estilo, secuencia y paisaje en el arte rupestre de Ampolla, Sierra de El Alto-Ancasti (Catamarca, Argentina). Chungara 54, 739–768. doi: 10.4067/S0717-73562022005001905
Togo, J. (2004). Arqueología santiagueña: Estado actual del conocimiento y evaluación de un sector de la cuenca del Río Dulce. [doctoral thesis]. [La Plata, Argentina]: Universidad Nacional de La Plata.
Togo, J. (2005). “Asentamientos humanos y aprovechamiento de los recursos en Santiago del Estero,” in Santiago del Estero: Una mirada ambiental, ed Amelia Giannuzzo y Myriam Ludueña, (Santiago del Estero: Universidad Nacional de Santiago), 91–120.
Tomka, S. (1992). Vicuñas and llamas: parallels in behavioral ecology and implications for the domestication of Andean camelids. Hum. Ecol. 20, 407–433. doi: 10.1007/BF00890428
Torrella, S., and Adámoli, J. (2006). “Situación Ambiental de la Ecorregión del Chaco Seco,” in La Situación Ambiental Argentina, eds. A. Brown, U. Martinez Ortiz, M. Acerbi, J. Corcuera (Buenos Aires: Fundación Vida Silvestre), 75–83.
Troll,. C. (1958). Las Culturas Superiores Andinas y el medio geográfico. Lima: Publicaciones del Instituto de Geografía, Facultad de Letras, Universidad Nacional Mayor de San Marcos.
Urquieta, B., and Martinez, P. (1992). Adaptación de mamíferos al ambiente altiplánico. Monografia de Medicina Veter. 4:1923.
Valdez, F. (2013). Arqueología amazónica. Las civilizaciones ocultas del bosque tropical. Marsella: IRD Éditions, Institut français d'études andines. doi: 10.4000/books.irdeditions.18758
Van Kessel, J., and Condori Cruz, D. (1992). Criar la vida. Trabajo y tecnología en el mundo andino. Santiago: Vivarium.
Ventura, B. (2001). “Los últimos mil años en la arqueología de las Yungas,” in Historia Argentina Prehispánica, eds. E. Berberian, A. Nielsen (Córdoba, Editorial Brujas), Tomo I, 447–492.
Vilá, B. (2014). Una aproximación a la etnozoología de los camélidos andinos. Etnoecológica 10, 1–16. Available online at: https://www.vicam.org.ar/publi/VilaEtnoecol2014.pdf
Wawrzyk, A., and Vilá, B. (2013). Dinámica de pastoreo en dos comunidades de la puna de Jujuy, Argentina: Lagunillas del Farallón y Suripujio. Chungará 45, 349–362. doi: 10.4067/S0717-73562013000200010
Webster, S. (1973). Native pastoralism in the south Andes. Ethnology 12, 115–133. doi: 10.2307/3773341
Wheeler, J. (1991). “Origen, evolución y status actual,” in Avances y perspectivas del conocimiento de los Camélidos Sudamericanos, ed S. Fernández- Baca (Santiago, Chile: FAO), 11–48.
Wheeler, J. (1999). Patrones prehistóricos de utilización de los camélidos sudamericanos. Bol. Arqueol. PUCP 3, 297–305. doi: 10.18800/boletindearqueologiapucp.199901.014
Wolverton, S., Otaola, C., Neme, G., Giardina, M., and Gil, A. (2015). Patch choice, landscape ecology, and foraging efficiency: the zooarchaeology of late Holocene foragers in western Argentina. J. Ethnobiol. 35, 499–518. doi: 10.2993/etbi-35-03-499-518.1
Yacobaccio, H. (2007). Andean camelid herding in the South Andes: ethnoarchaeological models for archaeozoological research. Anthropozoologica 42, 143–154. Available online at: https://sciencepress.mnhn.fr/fr/periodiques/anthropozoologica/42/2/l-elevage-des-camelides-dans-les-andes-du-sud-modeles-ethnoarcheologiques-appliques-l-archeozoologie
Yacobaccio, H. (2010). “Osteometría de llamas (Lama glama L.) y sus consecuencias arqueológicas,” in Zooarqueología a principios del siglo XX: aportes teóricos, metodológicos y casos de estudio, eds. M. Gutiérrez, M. De Nigris, M. Fernández, P. Giardina, M. Gil, A. Izeta et al. (Buenos Aires: Ediciones del Espinillo), 65–75.
Yacobaccio, H., Madero, C., Malmierca, M., and Reigadas, M. (1997-1998). Caza, domesticación y pastoreo de camélidos en la Puna Argentina. Relac. de la Soc. Arg. de Antropol. 22, 389–418.
Yacobaccio, H., and Vilá, B. (2013). La domesticación de los camélidos como proceso de interacción humana y animal. Intersecc. Antropol. 14, 227–238. Available online at: http://repositorio.filo.uba.ar/handle/filodigital/12654
Zuccarelli Freire, V. (2020). Desde las cumbres a las yungas: Las múltiples escalas de las prácticas agrarias prehispánicas en las Sierras de El Alto-Ancasti (Catamarca) durante el primer milenio A.D. [Doctoral thesis]. [Buenos Aires, Argentina]: Universidad de Buenos Aires.
Keywords: South American camelids, eastern Andes, herding, zooarchaeology, Northwestern Argentina
Citation: Moreno E, Samec C, del Papa LM, Taboada C, Mercolli P, Nores R, Schirmer S and Amano N (2025) Management of South American camelids in the eastern Andes of Northwestern Argentina: a comparative study of archaeological cases in the last two millennia. Front. Environ. Archaeol. 4:1552292. doi: 10.3389/fearc.2025.1552292
Received: 27 December 2024; Accepted: 20 March 2025;
Published: 14 April 2025.
Edited by:
Ramiro Barberena, Temuco Catholic University, ChileReviewed by:
Christian Mader, University of Bonn, GermanyMai Takigami, National Museum of Nature and Science, Japan
Jennifer Grant, National Scientific and Technical Research Council (CONICET), Argentina
Copyright © 2025 Moreno, Samec, del Papa, Taboada, Mercolli, Nores, Schirmer and Amano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Enrique Moreno, ZW5hbG1vckBnbWFpbC5jb20=; Noel Amano, YW1hbm9AZ2VhLm1wZy5kZQ==