 Luis Pezo-Lanfranco1,2,3*
Luis Pezo-Lanfranco1,2,3* Aldemar Crispin3
Aldemar Crispin3 Alonso Prado-Barragán3
Alonso Prado-Barragán3 Tatiana Abad3
Tatiana Abad3 Marco Machacuay3
Marco Machacuay3 Marco Yseki3Manuel Gorriti3
Marco Yseki3Manuel Gorriti3 Luis Miranda3José Apolín3Alice DiMuro1,2
Luis Miranda3José Apolín3Alice DiMuro1,2 Pedro Novoa3,4
Pedro Novoa3,4 André Carlo Colonese1,2†
André Carlo Colonese1,2† Ruth Shady3,4†
Ruth Shady3,4†- 1Institute of Environmental Science and Technology (ICTA), Universitat Autònoma de Barcelona, Bellaterra, Spain
- 2Department of Prehistory, Universitat Autònoma de Barcelona, Bellaterra, Spain
- 3Zona Arqueológica Caral, Ministerio de Cultura, Lima, Peru
- 4Facultad de Ciencias Sociales, Universidad Nacional Mayor de San Marcos, Lima, Peru
The balance between fishing and farming for the development of complex societies in ancient Central Andes is the subject of a long debate. Despite the availability of marine resources in one of the world's richer marine ecosystems and the extreme aridity of the landscape, current evidence suggests that farming was the prime mover of early Andean civilization. In this study, we employed stable isotope analysis to reconstruct the diets of 38 human individuals from two occupations at Vichama, a coastal settlement with monumental architecture located in the Huaura Valley, Early Formative−1 period (EF-1, 1800–1500 BCE) and the Late Intermediate Period (LIP, 1000–1300 CE). Results show a considerable contribution of C3 cultivated plants and marine protein during EF-1, which indicates the first importance of farming at that time, whereas marine consumption increased during the LIP, a period of higher marine productivity. Our findings shed light on the complex interplay between climate alternances and coastal/inland sites' hegemony in the Central Andes.
Introduction
The connection between subsistence systems, climate oscillations, and social complexity has been extensively studied and documented for the Central Andes. Studies have shown that ancient Andean populations developed adaptive strategies modulated by ecological constraints since they arrived at the region ~14,000 years ago until the beginning of the Late Holocene (Dillehay, 2017; León, 2013; Piperno and Dillehay, 2008). Domesticate evidence dates to as early as 8000 BCE, and available data suggest that irrigation technologies supported plant cultivation by ~4500 BCE (Dillehay et al., 2005, 2007). Since ~3000 BCE, farming was the most important economic activity (Grobman et al., 2012; Yseki et al., 2023).
Increasing settlement size and distribution, shifts in domestic architecture, and the spatiotemporal modeling of demographic dynamics for the Peruvian coast indicate an exponential population growth between 2000 and 1 BCE (Goldberg et al., 2016). The process has been attributed to a complex interplay between environmental changes and the expansion of agricultural regimes in the region (Contreras, 2010; Goldberg et al., 2016; Wilson et al., 2022).
In the last decades, a large volume of archaeological information has been produced for the Peruvian North-Central coast (PNCC), the epicenter of early complexity in the Central Andes (Creamer et al., 2013; Haas and Creamer, 2006; Makowski, 2023; Shady et al., 2001; Shady, 2018). Although pioneering evidence suggested that marine resources were the foundation of initial social complexity in this extremely challenging environment (Moseley, 1975), new evidence of plant consumption at coastal and middle valley monumental sites strongly indicates that farming was the subsistence system supporting the emergence of early civilization in the region (e.g., Caral civilization) and subsequent sociopolitical developments (Dillehay et al., 2007; Haas et al., 2013; Pezo-Lanfranco and Colonese, 2024; Shady, 2006a,b; Shoji et al., 2023; Tung et al., 2021).
In this area, studies suggest that coastal and inland societies would have been organized around symbiotic farming–fishing economies and littoral–inland trade networks since at least the Initial Formative (3000–1800 BCE), with coastal communities based on marine resources and some contribution of cultivated crops and inland groups relying in cultivated plants supplemented by marine products (Pezo-Lanfranco et al., 2022; Shady, 2018; Yseki et al., 2023).
This system, however, would have been influenced by the variability of sea surface temperatures (SSTs), upwelling dynamics, and productivity intensity in the Humboldt Current System (HCS), and El Niño Southern Oscillation (ENSO). The increase of SSTs produces a decrease in marine biomass, whereas in upper valleys, landslides increase as a consequence of intense rainfall, causing imponderable effects in farming production (Contreras, 2010). In the north and central coast of Peru, humid periods are linked to the prevalence of El Niño–like conditions, whereas La Niña–like conditions can be associated with dry and cold climate (Salvatteci et al., 2014, 2019; Leclerc, 2023).
Although the final causes and the complex mechanisms among the factors involved (e.g., ENSO teleconnections, Walker circulation intensity, meridional displacement of the Inter-Tropical Convergence Zone, and the South Pacific Subtropical High) are still in discussion (Mächtle and Eitel, 2013; Salvatteci et al., 2014, 2016; Scholz et al., 2014; Yseki et al., 2022), variations in SSTs, upwelling conditions, and productivity intensity in the HCS occurring on multiple timescales (Salvatteci et al., 2016, 2019; Yseki et al., 2022) likely had a significant impact on the availability and selection of marine resources utilized by past coastal communities and influenced the rise and decline of ancient Andean societies during the Holocene (Wilson et al., 2022, 2024). These effects occurred alongside persistent hazards, such as tectonic activity and geomorphological particularities (Orloff, 2022; Salvatteci et al., 2019; Sandweiss et al., 2009).
Export production refers to the fraction of organic carbon produced by photosynthetic organisms in the photic zone that is eventually deposited on the ocean floor; thus, it serves as a proxy for SST fluctuations in oceanography (Herbert, 2003; Scholz et al., 2014; Wefer et al., 2015). A comprehensive palaeoceanographic study of the HCS based on high-resolution data (i.e., alkenone and bromide-to-titanium [Br/Ti] ratio) has revealed centennial-scale changes in SSTs and, in general, an increase in exported productivity from ~3000 to 1100 BCE (Salvatteci et al., 2019).
Studies suggest that a cold period along the Peruvian coast between 1800 and 1200 BCE produced a decrease in SSTs and an increase of marine productivity (Salvatteci et al., 2016, 2019), leading to changes in the equilibrium of political power to the benefit of coastal populations. High marine productivity of the Peruvian sea has been detected between ~1700 and 1500 BCE (Salvatteci et al., 2019), coinciding with the flourishing of several ceremonial centers along the coastal area such as Vichama (Shady et al., 2015) and Bandurria (Chu, 2011), located, respectively, along the coast of the northern and southern interbasins of Huaura Valley, the more productive of the PNCC. However, the extent to which changes in marine productivity may have led to an increased reliance on marine resources at the expense of agricultural products, well known in the region since previous periods, remains untested. To investigate this issue, we integrate stable isotope data (δ13Ccollagen, δ15Ncollagen, δ13Capatite) and Bayesian stable isotope mixing models (BSIMMs) of human individuals from Vichama, an Early Formative (EF) coastal site with monumental architecture, “key” to understanding the timing of dietary changes and the nuances of the Formative economy. From a broader theoretical perspective, this article offers valuable insights into how climatic conditions influenced the rise and collapse of complex prehistoric societies (Kennett and Marwan, 2015).
Archaeological context: Vichama
Vichama is an archaeological site located in the modern city of Végueta, 130 km north of Lima, in the PNCC (UTM WGS84 N8779900/E212200). The site is at the right margin of the lower Huaura Valley (Huaura–Supe interbasin), at 1.5 km from the Pacific Ocean and 6.5 km northwest to the river mouth (Figure 1). The Huaura River is a permanent source of water that flows ~158 km from its origin in the Andean highlands (i.e., Sura Saca lagoon—Cordillera de Raura) to the sea and works as a natural corridor that links transversally the coast with important socioeconomic regions, such as the Mantaro basin and Eastern Andean lowlands (e.g., Alto Marañón, Alto Huallaga).
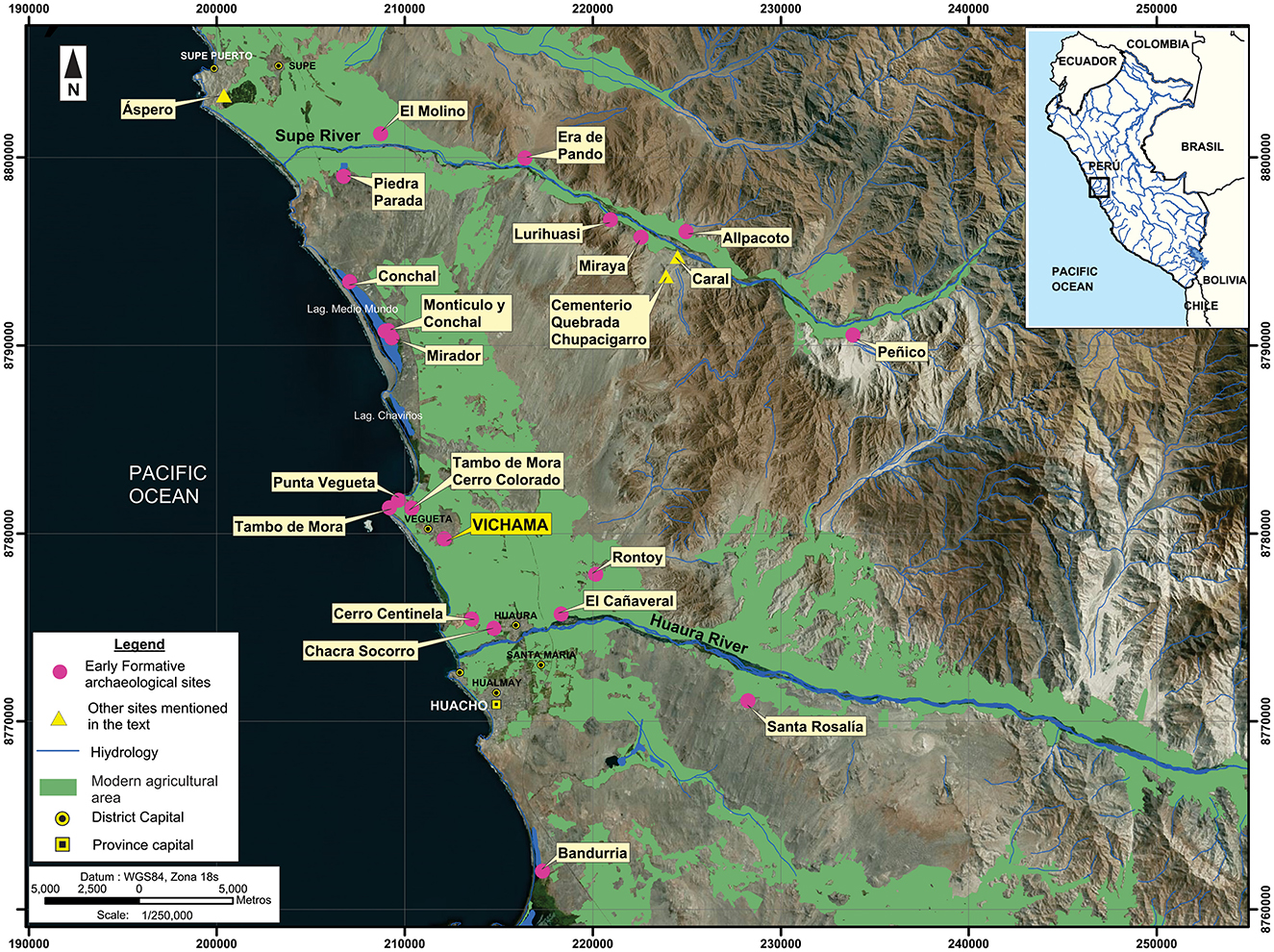
Figure 1. Location of Vichama during the Early Formative (1800–1500 BCE) and other major formative settlements of the Peruvian North-Central coast.
Although the lower Huaura Valley has been the object of archaeological prospections since the early 20th century (Engel, 1987; Tello, 2004), Vichama was not systematically studied until the 1970s (Aguilar, 2006; Cárdenas, 1977). Since 2007, Vichama has been intensively excavated and conserved by the Caral Project (Zona Arqueológica Caral-ZAC), sponsored by the Peruvian government and local institutions (Shady et al., 2008, 2015).
Archaeological evidence and radiocarbon dates indicate the site was occupied during two chrono-cultural periods: during the Early Formative−1 (EF-1, 1800–1500 BCE), when monumental architecture flourished through six architectonic periods (P1–P6), and during the Late Intermediate Period (LIP, 1000–1300 CE), when the site became a fishing village with decreased political power in this section of the valley during a period of high marine productivity (Figure 2).
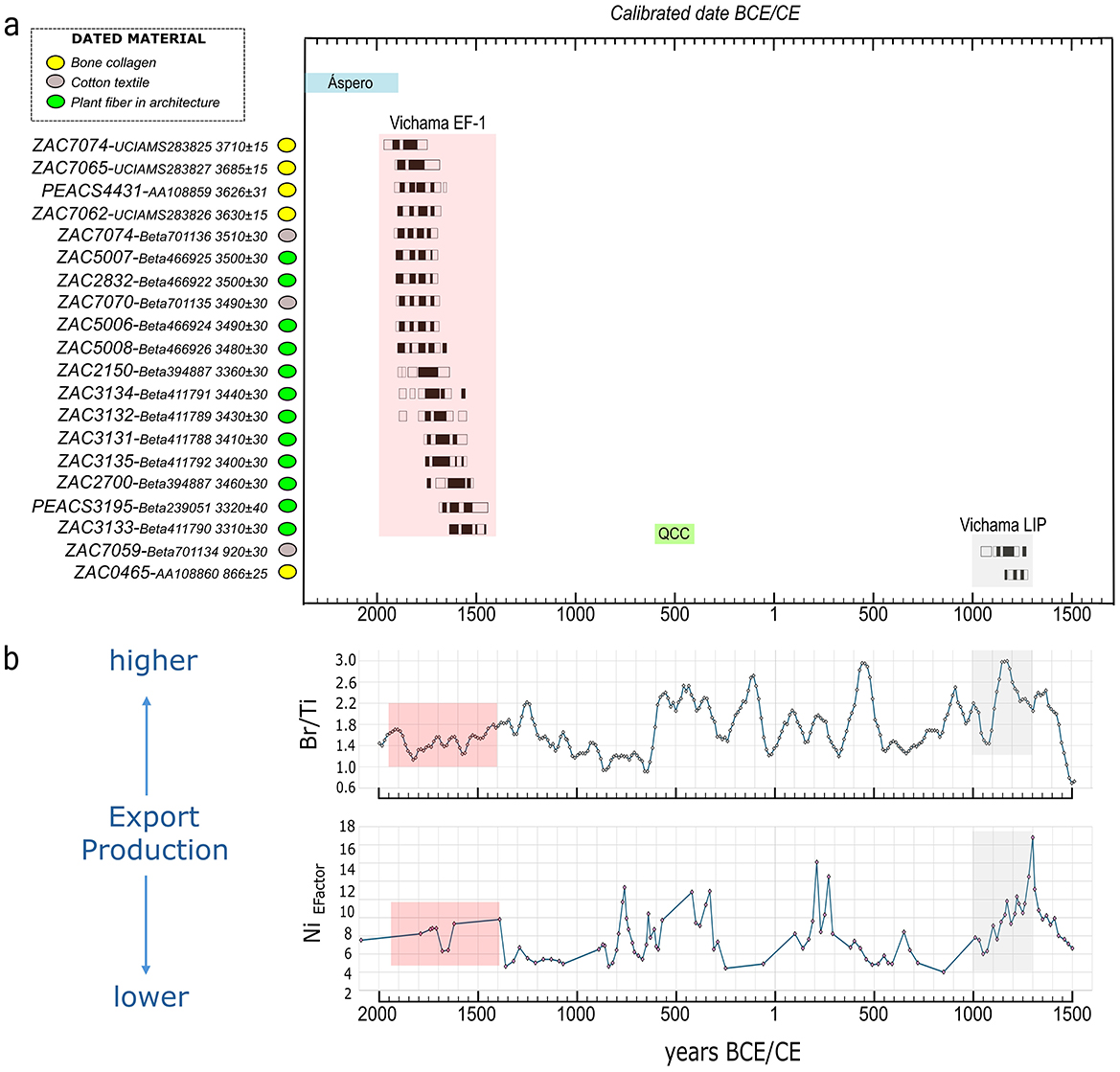
Figure 2. (a) Radiocarbon dates of Vichama and proxies for export productivity in the Peruvian coast during the Early Formative−1 (EF-1) and Late Intermediate (LIP) periods. See also the chronology of two sites mentioned in the article (i.e., Áspero and Quebrada Chupacigarro Cemetery—QCC). (b) Export productivity indexes: records of bromine-to-titanium ratio (Br/Ti, core M135-004-3, latitude 17°S) and authigenic nickel enrichment factor (EFactor; core G-10-GC-02, latitude 14°S). The bromine-to-titanium ratio is a semiquantitative method to estimate sedimentary total organic carbon, and the nickel (EFactor) indicates the organic sinking flux (Salvatteci et al., 2016). Colored areas represent the compared periods' classification (red for EF-1 and gray for LIP). Raw data available at: https://doi.pangaea.de/10.1594/PANGAEA.887109.
Vichama is a multicomponent complex that comprises nine sectors, including Formative monumental architecture (Vi-01) and shell middens (Vi-02, Vi-03), and architecture and cemeteries of later periods (Vi-04–Vi-09) scattered across 136 ha in the hills of Cerro Halconcillo, a rocky elevation that rises 114 m above the sea level in the middle of one desertic oasis of 305 ha (Figure 3a).
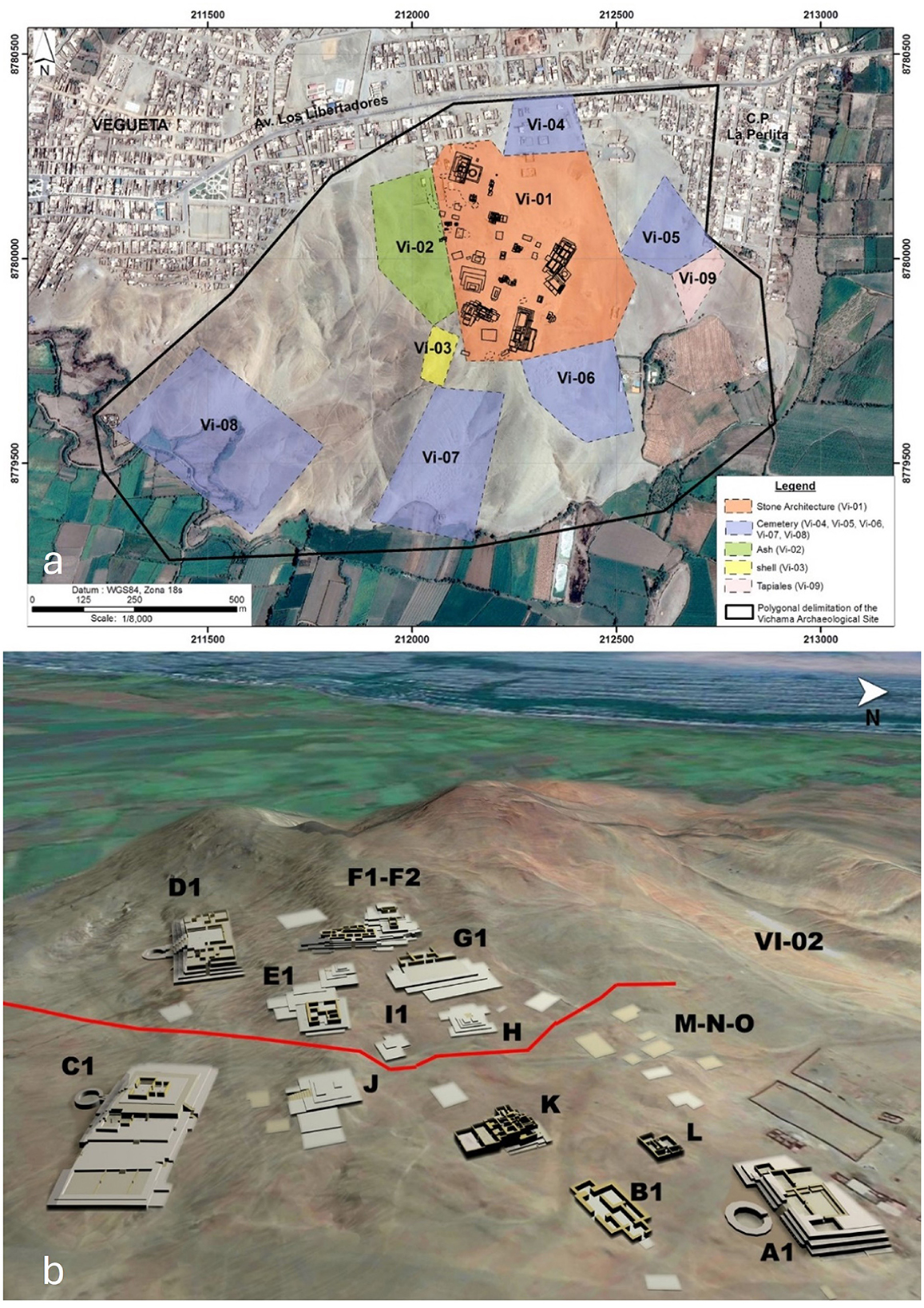
Figure 3. (a) Sectors of Vichama archaeological site. (b) Three-dimensional reconstruction of some relevant monumental buildings in the upper and lower halves of the site. Upper half: Edificio Público Mayor (Sector D1), Edificio Las Ofrendas (Sector F), Edificio Público Residencial (Sector E1), Edificio Público G1, Edificio Público Mediano (Sector H), Edificio Público Menor (Sector I1). Lower half: Edificio Principal (Sector A1), Edificio Residencial de Elite (Sector B1), Edificio Público Los Depósitos (Sector K), Edificio de las Chakanas (Sector C1), Edificio Público Menor (Sector L), Edificio Residencial de Elite (Sector J), Sector Residencial (Sector M-N-O), domestic and burial areas (VI-02). See Supplementary material 2 for detailed locations of funerary contexts and individuals evaluated in this work.
During the EF-1, Vichama was part of an emerging complex polity in the lower Huaura Valley, which would have been composed of no < 20 settlements with monumental architecture (Aguilar, 2006; Cárdenas, 1977). It is possible that Vichama represented the rise of a new sociopolitical center in the region following the decline of the Caral society. The nuclear area at that time comprised 29 buildings, including public (e.g., stepped platforms, sunken circular courts, squares, and walls with reliefs), and domestic architecture of orthogonal design spread across 25 ha (Shady et al., 2015). This area is divided into two halves, upper and lower, following the bipartite spatial division common in Andean settlements (Figure 3b). The ceremonial buildings exhibit various architectural features, including stone walls arranged in horizontal rows, architectural fills using mega-shicras, and walls adorned with high-relief decorations (Shady et al., 2008, 2015). Changes in ceremonial architecture and paraphernalia (i.e., polychrome sculptures of unfired clay and particular iconography) suggest ideological transformations.
The diet in Vichama, current evidence
The “visual basin” of Vichama encompasses an approximate area of 9.2 km2 that included diverse ecosystems that possibly were exploited for subsistence, among them, rocky and sandy beaches, swamps, lomas (fog oases), farming fields, the river, and the sea. The coast of Huaura Valley is one of the richest areas of the Peruvian sea, which currently cumulate ~20% of the total national production of fisheries (IMARPE, 2010, p. 64). Islands and islets (e.g., Anat, Azakan) located < 1 km from the shore, potentially provided dense clam banks, marine turtles, and a variety of seabirds.
The importance of fishing is undeniable. The zooarchaeological record includes a total of 91 species (23 fish, 61 mollusks, 3 marine mammals, and 4 marine birds), recovered from primary contexts (see Supplementary material 1: S1—Marine species).
In addition, the exploitation of coastal salt-banks (i.e., Punta de Atahuanca and Salinas de Huacho, among others) to produce dry-salted fish probably was an important economic activity in the region during the Initial Formative and EF periods (Engel, 1987; Tello, 2004). Modern fishermen of Végueta employ several traditional techniques such as redes de cerco, diving, chinchorro, and arte de pinta that could be used in the past to capture the wide array of reported species (IMARPE, 2010, p. 67).
According to the last report (Yseki et al., 2024), the archaeobotanical inventory of edible plants in Vichama has yielded more than 35,000 macro-remains representing 17 taxa derived from 123 primary contexts (see Supplementary material 1: S1—Edible plants species). The botanical record includes fruits (guava, lucuma, pacay, avocado and ciruela del fraile), annuals like chili peppers (e.g., Capsicum baccatum), beans (i.e., lima bean, common bean, and jack bean), squashes, and roots and tubers (i.e., achira, sweet potato, and potato). Based on their ubiquity, it seems that fruit trees, especially guava and lucuma, as well as chili peppers, were regularly consumed and used in ceremonial activities. Similarly, the high ubiquity in offering contexts of remains of lima bean, achira, sweet potato, and potato highlights the economic importance of these plants in Vichama (Yseki et al., 2024).
The absence of Zea mays (maize) macro-remains suggests that it was probably not grown, processed, or stored at Vichama. However, a recent study reported starch grains with potential fermentation-related damage in dental calculus of individuals from Áspero (Yseki et al., 2023), with a minimal presence of maize macro-remains (Shady and Cáceda, 2008). Thus, maize consumption at Vichama cannot be ruled out based solely on macro-remains analysis, as maize may have been consumed as a beverage or flour (Yseki et al., 2024).
The marine fauna and some differences between periods suggest fluctuations in sea productive conditions linked to the upwelling of rich-nutrient marine waters. For instance, during the EF period (1800–1200 BCE), there is a general trend to increasing exported productivity, especially during the EF-1 (1800–1500 BCE), with some fluctuations (i.e., lower productivity between 1800 and 1600 BCE, with the lowest values around 1700 BCE and higher productivity between 1600 and 1500 BCE) that should have influenced dietary composition, possibly with higher consumption of marine products; however, the plant record shows that agriculture was ongoing during the flourishment of monumental architecture.
These conditions, however, could be relatively different from those observed during the LIP (1000–1300 CE) when productivity reached even higher values and enhanced upwelling conditions, with its maximum at ~1000–1100 CE (Kennett and Marwan, 2015). During the LIP, the archaeological deposits are plenty of marine remains, such as fish bones and shells, fishing nets, and artifacts for marine exploitation.
Stable isotopes in diet reconstruction
While faunal and plant remains can provide insight into shifts in economic practices, stable isotope analysis is more effective for targeting the net consumption of specific food resources at an individual level. Stable carbon and nitrogen isotope analyses of collagen (δ13Ccoll and δ15N) and bioapatite (δ13Cap) extracted from human bones and teeth offer valuable insights into the diets of ancient populations (Schoeninger and Reitsema, 2023). The δ13Ccoll is a proxy of protein sources and trophic position of consumers in the food web, as well as the photosynthetic pathway of the plants they consumed. The δ13Cap instead reflects the entire dietary carbohydrates, lipids, and proteins, providing a useful means to assess the energetic constituents of the diet and photosynthetic pathways (Jim et al., 2004; Kellner and Schoeninger, 2007). In the Andean central coast, C3 plants, which use the Calvin–Benson photosynthetic pathway, are the most prevalent and typically exhibit mean δ13C values of −25.7 ± 2.1‰. In contrast, among C4, which follow the Hatch–Slack photosynthetic pathway, there are only two potentially edible domesticated plants (kiwicha—Amarantus sp., and maize—Zea mays), with typical values of approximately −11.3 ± 1.4‰. Additionally, crassulacean acid metabolism (CAM) plants have isotopic values (−10.8 ± 0.2‰) that overlap with those of C4 (Pezo-Lanfranco and Colonese, 2024).
Usually, C4 plant consumers show significantly higher δ13C values (Kellner and Schoeninger, 2007). In coastal areas, however, using δ13C to trace maize consumption can be challenging due to the presence of marine resources (i.e., fish and mollusks), which have overlapping δ13C values to C4 plants, approximately −11.9 ± 2.1‰ (Pezo-Lanfranco and Colonese, 2024). In terms of δ15N values, herbivores tend to have values 3–7‰ higher than the plants they consume, while carnivores exhibit δ15N values 3–5‰ higher than their prey. Because marine sources have higher isotopic values compared to terrestrial sources (Minagawa and Wada, 1984; Schoeninger and DeNiro, 1984), marine fauna displays higher δ15N values than terrestrial ones, linked to trophic fractionation within complex marine food webs (Chisholm et al., 1982; DeNiro and Epstein, 1981; Hedges and Reynard, 2007; Schoeninger, 2010; Schoeninger et al., 1983). Stable isotope analysis is particularly informative when combined with Bayesian mixing models for deriving quantitative estimates of dietary composition as probability distributions (Cheung and Szpak, 2021; Fernandes et al., 2014, 2015; Phillips, 2012).
Hypothesis and expectations
By employing a combination of stable isotope proxies from collagen (δ13Ccoll, δ15Ncoll, n = 36) and apatite (δ13Cap, n = 26) of 38 human individuals from Vichama, and BSIMM estimations of the caloric contribution of the potential food sources (i.e., marine and terrestrial fauna, as well as C3 and C4 plants; Fernandes et al., 2014, 2015) consumed by 21 Vichama individuals (see Material and Methods for model implementation details), we aim to elucidate the primary sources of energy for two occupational periods at Vichama: EF-1 (1800–1500 BCE) and the LIP (1000–1300 CE).
We tested the hypothesis that changes in marine productivity influenced the degree to which marine resources were incorporated into Andean subsistence economies (Kennett and Marwan, 2015; Wilson et al., 2024). According to paleoclimatic data, an increased marine productivity between 1800 and 1500 BCE was concomitant with the flourishing of Vichama (Shady et al., 2015). However, the economic importance of plants and fish and their relative contribution to diet is unknown. It is expected that the inhabitants of Vichama, particularly during EF-1, relied more strongly on marine products as a significant part of their diet.
For comparison, the LIP (1000–1300 CE) was a time of climate cooling and decreasing SSTs, when marine productivity reached its peak (Kennett and Marwan, 2015). Because the LIP has been recognized as the period of higher development of agriculture in the Central Andes, under increasing conditions of upwelling and marine productivity, a mixed diet with a higher contribution of marine sources is expected. This reliance should be reflected in paleodietary reconstructions.
Results
Dietary reconstruction
The isotopic values of 38 human individuals from Vichama were classified into the two occupations, EF period (1800–1500 BCE, n = 30), and the LIP (1500–1300 CE, n = 8), used here for comparative purposes (Table 1, Supplementary material 1: S2 for individuals' contexts and bioarchaeological details and S3 for radiocarbon dates). Only a fraction of the 59 elements sampled (36 bones and 23 teeth) provided intact collagen with reliable δ13Ccoll and δ15N values from bone (n = 21) or teeth (n = 15) according to accepted preservation criteria (DeNiro, 1985; Van Klinken, 1999). Attenuated total reflectance–Fourier transform infrared spectroscopy (ATR-FTIR) studies conducted in a subsample of 10 individuals consistently suggest that no diagenetic alteration affected the inorganic fraction of bones from Vichama contexts (France et al., 2020; see Supplementary material 1: S4 for the ATR-FTIR data). Thus, δ13Cap values (n = 26) were included in our analysis to complement the paleodietary information.
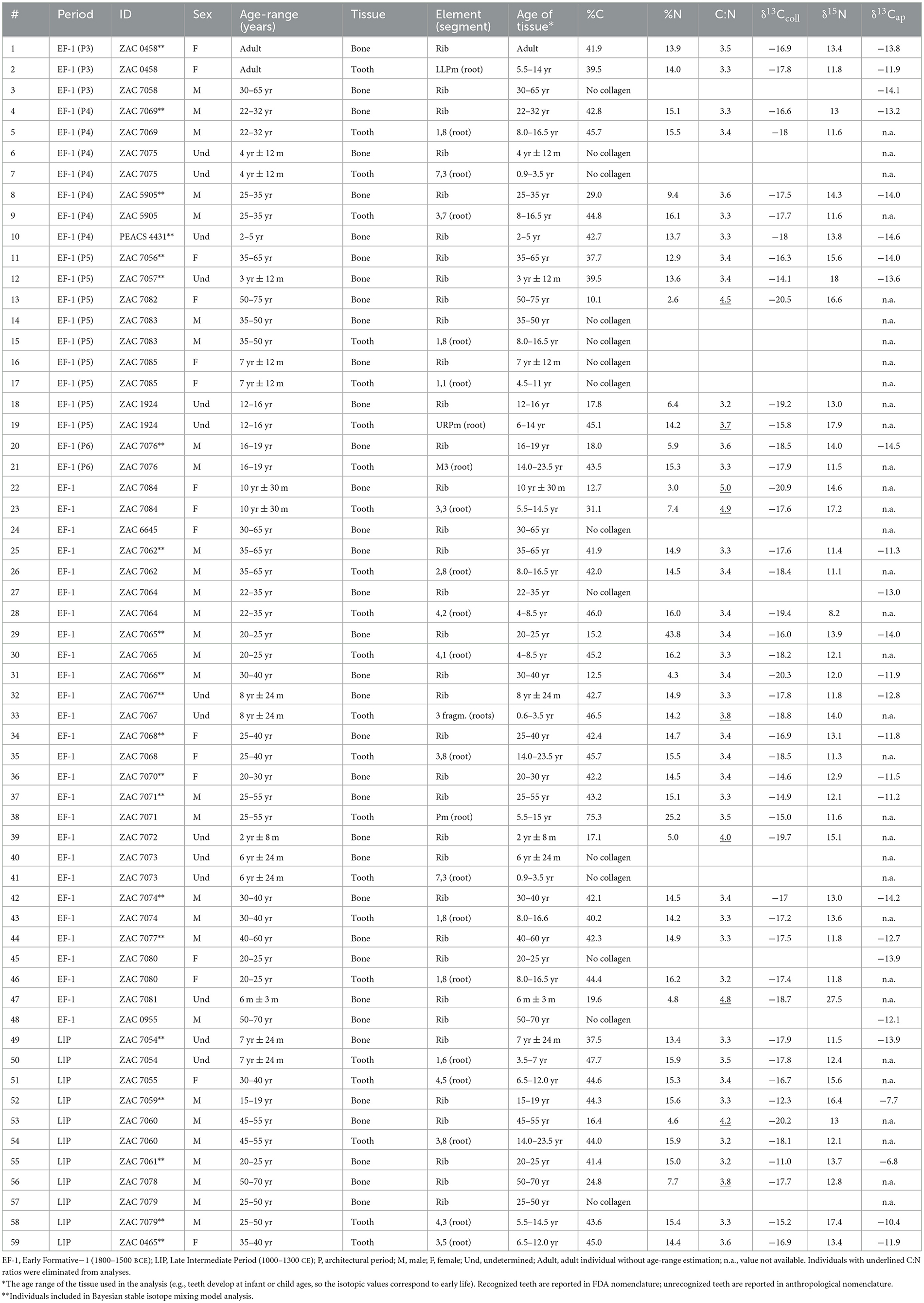
Table 1. Isotopic values from Vichama individuals.
The δ13Ccoll values from both bone and tooth dentin ranged from −20.3 to −11.0 ‰, δ15N values ranged from +8.2 to + 18.0 ‰, whereas δ13Cap values ranged from −14.6 to −6.8 ‰ (Supplementary material 1: S5).
Although the contribution of marine protein is evident in some male individuals (Figure 4a), the mean δ15N values are lower than expected for fishers and suggest the consumption of marine protein of a low trophic level, possibly little fishes and shellfish (with mean values of δ13Ccoll = −12.3 ‰, σ = 0.5, and δ15N = + 10.4 ‰, σ = 1.2) over the entire sequence. No significant differences were observed for δ13Ccoll, δ15N, and δ13Cap values between females and males or between preadults and adults for both EF-1 and LIP occupations (Supplementary material 1: S5). The significant difference observed between adults and preadults of EF-1 probably resulted from the weaning effect in a couple of infants (i.e., PEACS4431 and ZAC7057) or different protein sources (i.e., ZAC7067) in some preadults.
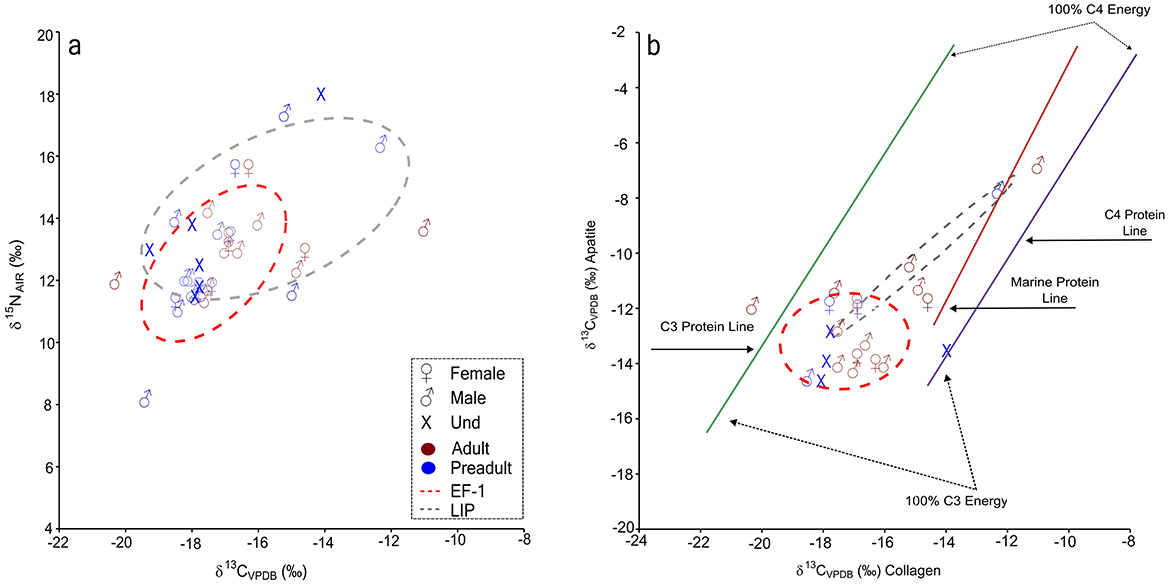
Figure 4. Scatterplot of stable isotope markers in Vichama individuals of Early Formative period and the Late Intermediate Period and overlapping standard ellipse areas (estimated for 68.5% of the population), for comparison. (a) δ13Ccoll and δ15N values. (b) δ13Ccoll and δ13Cap values in the workspace of Kellner and Schoeninger (2007). V-PDB, Vienna Pee Dee Belemnite; AIR, atmospheric air.
Inter-population comparisons revealed a significant difference only for δ13Cap (Mann–Whitney U = 18.50, p = 0027). In general, the δ13Ccoll, and δ13Cap indicate diets dominated by protein and energy sources from C3 ecosystems (Figure 4b; Kellner and Schoeninger, 2007). Nevertheless, individuals from the LIP exhibited a much broader range of δ13Ccoll and δ15N values, indicating relatively higher marine protein consumption, notably by some male individuals (e.g., ZAC7061, ZAC7059), compared to the EF-1 population.
The comparisons of protein sources of Vichama EF-1 and LIP phases with other populations from the PNCC show a trend to consume a diet intermediary between that of Áspero, the most representative site of fishermen in the Supe Valley during the Initial Formative (3000–1800 BCE), based on C3 plants and marine protein of higher trophic level, and the diet of later farmers from Quebrada Chupacigarro cemetery (middle Supe Valley, Late Formative, 500–400 BCE), virtually based on plants (Pezo-Lanfranco et al., 2022). LIP diets show an overlap area greater than EP-1 with Áspero diets (Figure 5).
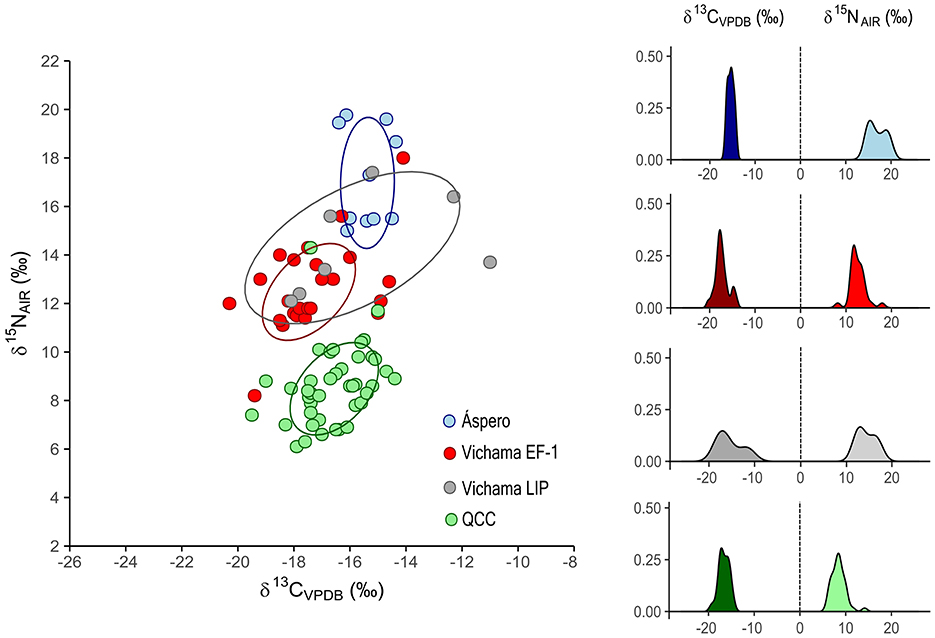
Figure 5. Scatterplot of δ13C and δ15N with standard ellipse areas (estimated for 68.5% of the population), and density kernel plots of δ13Ccoll and δ15N values of Vichama and other Formative populations from the Peruvian North-Central coast. EF-1, Early Formative−1 period; LIP, Late Intermediate Period; QCC, Quebrada Chupacigarro Cemetery.
In addition, we estimate the proportions of dietary components estimated with BSIMMs for 21 Vichama individuals with a set of complete isotopic data (Table 2, Figure 6; see also Supplementary material 1: S6 for the model's results). These results suggest that most calories consumed came from C3 plants (contribution ranging from 14% to 86%), followed by fish and mollusks (8% and 58%), C4 plants (3%−19%), and terrestrial fauna (3%−11%).
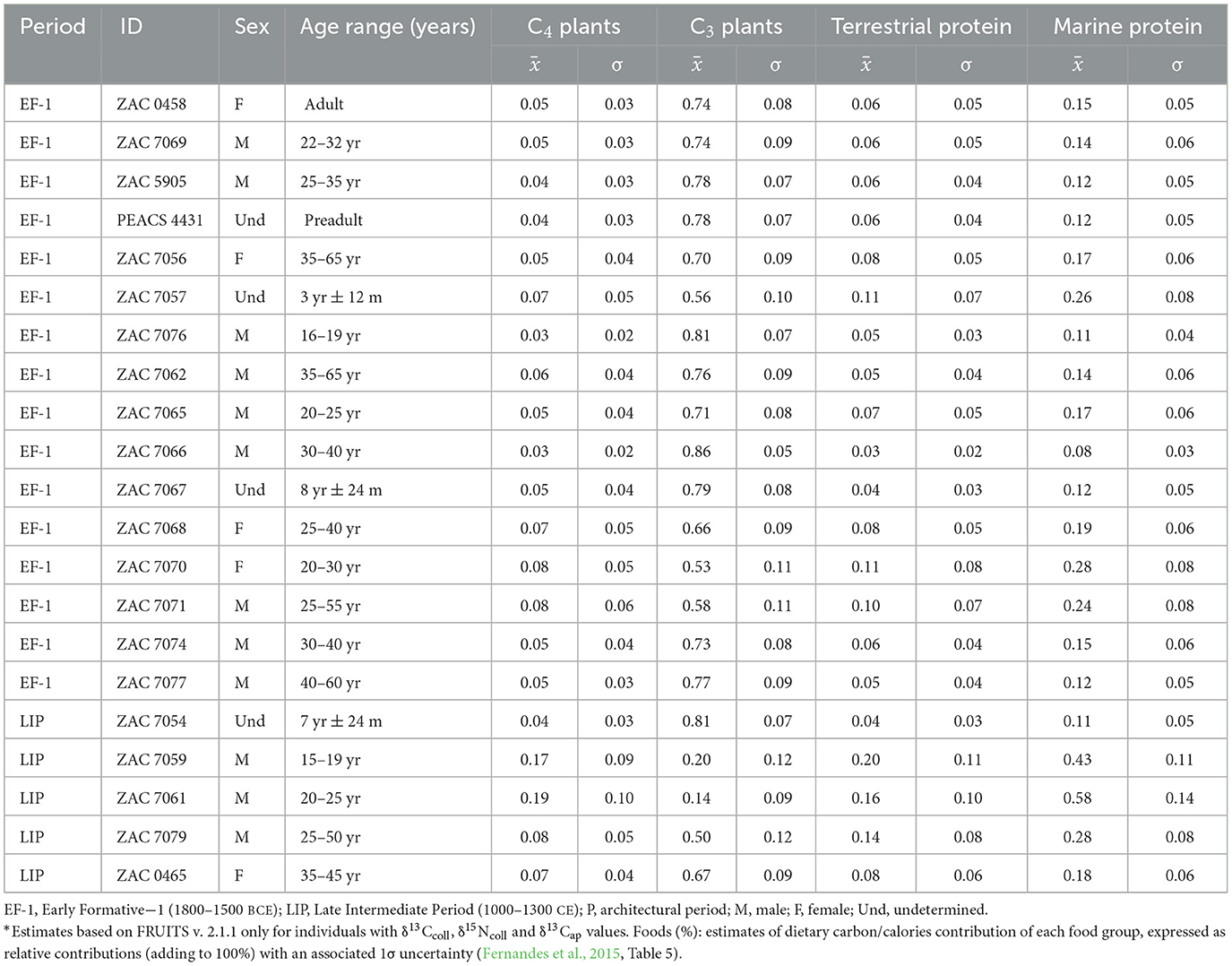
Table 2. Estimates of dietary carbon/calories contribution of each food group in Vichama individuals.*
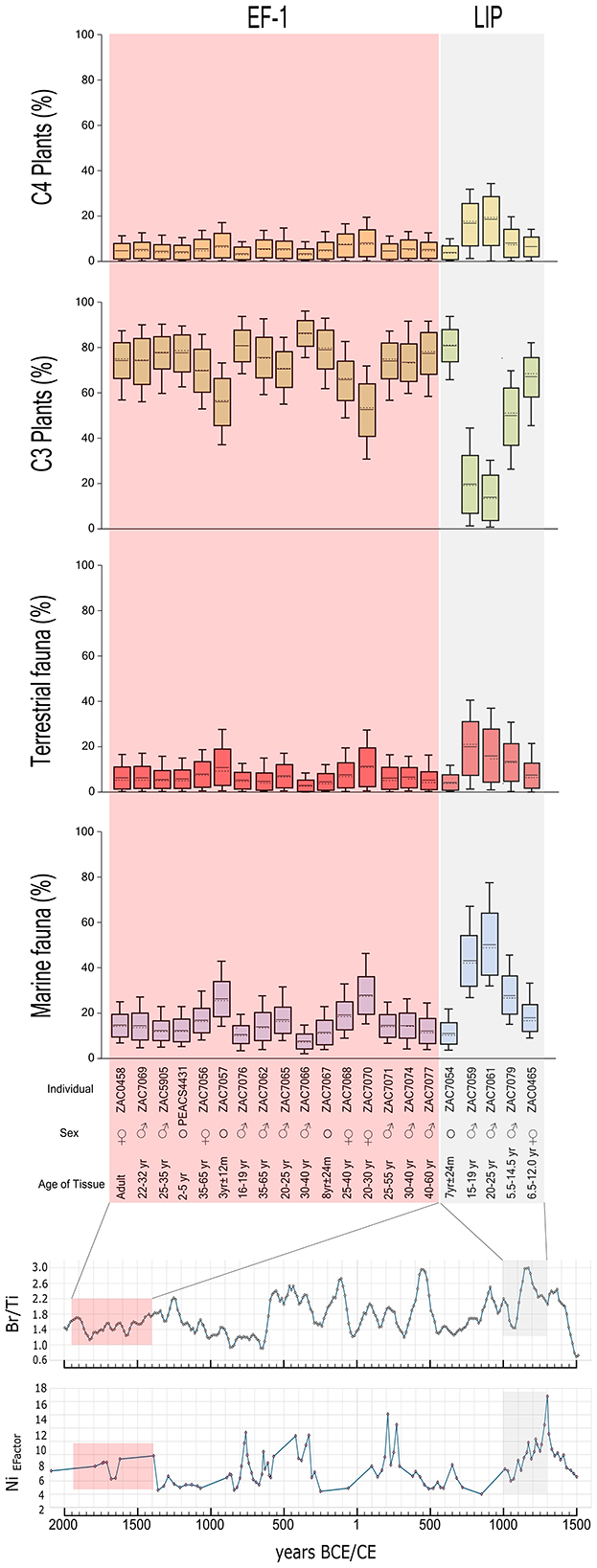
Figure 6. Dietary caloric estimations of food sources in Vichama individuals for the Early Formative−1 (EF-1, 1800–1500 bce) and the Late Formative Period (LIP, 1000–1300 ce), and their correspondence with exported production markers. The boxes represent a 68% confidence interval, and the whiskers, a 95% confidence interval. The horizontal discontinuous line indicates the median, and the continuous line indicates the mean. ♂, male; ♀, female; °, undetermined sex.
Thus, most individuals from the EF-1 occupation of Vichama show values compatible with a wide farming–fishing diet, with an unexpectedly high contribution of plants to people living near the sea. The comparison of diets between EF-1 and the LIP suggests a trend of increasing marine protein consumption in the later period, fewer C3 plants, and a discrete increase of C4 and terrestrial protein. The Kullback–Leibler divergence is 0.141, indicating a moderate difference in the average probability distributions among individuals of EF-1 and LIP, consistent with dietary change between periods with some overlap.
Discussion
The nature of diet at Vichama
Our study suggests that the two distinct occupational phases of Vichama were underpinned by different economic systems. During EF-1, dated to 1800–1500 BCE, although some individuals relied significantly on marine resources for calories (>25%), subsistence was based primarily on the cultivation of C3 plants. Plant remains, including several high-energy crops like tubers, beans, and fruits, were found mainly in funerary contexts associated with public buildings, yet our results reveal the significance of these plants at the household level beyond ritual use.
This agricultural production has likely supported Vichama's political prominence at the end of the Caral era. Cultivation seems to have been a plausible activity in Vichama during the EF-1. The agricultural area near Vichama extends approximately 970 ha, and the water sources are located < 2 km from Vichama. In the zone, the phreatic level is approximately 0.85 m depth (INRENA, 2005).
Evidence of botanical macro- and micro-remains shows the consumption of several domesticated crops in coastal settlements during the Initial Formative (3000–1800 BCE; Haas et al., 2013; Shady, 2006a,b,c; Vega-Centeno, 2005; Yseki et al., 2023, 2024; Zechenter, 1988). For example, plant micro-remains from human dental calculus at Áspero, in the same region, indicate significant sweet potato and moderate potato consumption during the Initial Formative period (Yseki et al., 2023), despite their scarcity in the macro-botanical record (Shady and Cáceda, 2008).
Vichama's location is appropriate for maize cultivation due to extensive arable lands and water availability. Interestingly, the isotopic evidence obtained by this work suggests a marginal contribution of C4 (lower than 10% in most cases) during EF-1 and agrees with the lack of maize evidence in the archaeological record. The near absence of maize in Vichama contrasts with its presence in some early sites from adjacent valleys of the PNCC (Haas et al., 2013; Shady, 2006b; Vega-Centeno, 2005; Zechenter, 1988). In Áspero, maize starch has been detected in dental calculus (Yseki et al., 2023), and in Pativilca Valley, maize pollen and starches appear as early as 2500 cal BCE (Haas et al., 2013). The presence of amaranth could account for the C4 signal detected at Vichama, although no macro-remains of this plant have been recorded at the site, unlike at Caral, located 30 km to the northeast, where amaranth has been documented in an earlier period (Shady, 2006c).
Approximately 3,000 years later, during the LIP (1300–1500 CE), when Vichama had long lost its regional political influence, the site's residents relied primarily on C3 plants, supplemented with a greater intake of marine resources. This increased focus on marine organisms is corroborated by the presence of fishing gear, such as nets and fishhooks made from mussel shells, as well as abundant fish and shell remains, all attesting to extensive fishing activities during the LIP. While maize may have been more consumed during the LIP, isotopic equifinality complicates distinguishing between C4 plants and marine carbon sources. Two young male individuals from the LIP (ZAC7059 and ZAC7061) exhibit values of C4 intake of approximately 20%; however, both consumed approximately 40%−60% of calories from marine sources and show auditory exostoses, an osteological marker associated with repetitive aquatic activity in cold waters (Pezo-Lanfranco et al., 2009), suggesting that they were divers or fishers rather than maize consumers. In contrast, the only individual from EF-1 with auditory exostosis (ZAC5905, a male) shows, counterintuitively, an unexpected low proportion of marine contribution to diet and δ15N values compatible with marine resources from the lower trophic level.
The overlap between maize and marine carbon values remains a potential source of uncertainty in isotopic analyses for this region. Regardless, if maize was consumed in Vichama during either EF-1 or the LIP, it likely did not constitute a staple or significant dietary component, contributing < 30% of total dietary calories (Ambrose and Norr, 1993).
During the EF-1, terrestrial mammal consumption appears to have been occasional, with contributions from these protein sources increasing in the LIP (20% of total calories), likely due to a greater availability of camelids during this period (León, 2013). The faunal record at Vichama includes only six taxa: three bird species, commensal rodents, frogs, and camelids, the latter being the only potentially edible domesticated taxon. However, archeozoological studies of inter-period consumption are still pending. The limited availability of domesticated species in the Andes, such as camelids, guinea pigs, and dogs, along with certain wild species from the lomas and desert areas, however, suggests that terrestrial protein remained a relatively minor dietary component during the Formative period, as observed in other studies (Pezo-Lanfranco et al., 2022; Pezo-Lanfranco and Colonese, 2024). As a potential limitation of this study, the presence of little fish species can lead to mimicking the signal of wild terrestrial protein from lomas, potentially C4 protein (Coutts et al., 2011; Cadwallader et al., 2012).
Although the archaeological record of EP-1 and the LIP support fishing activities for both periods, our results revealed that the reliance on marine products was higher during the LIP, which coincides with a time of high marine productivity, compared to EP-1. However, due to the limited LIP sample size, these interpretations should be considered provisional pending further evidence.
Finally, although some preadult individuals included in the BSIMMs could theoretically retain nitrogen isotopic signals from breastfeeding and weaning, leading to spurious interpretations, this is unlikely, as the analyzed tissues (tooth roots) correspond to older children, who typically consume an adult diet. Among the LIP individuals, two are adults and three are children or preadolescents, with sampled tissues ranging from 5.5 to 14.5 years. Previous research on weaning patterns in Andean populations places the completion of weaning between 1.5 and 3 years of age (see Pezo-Lanfranco et al., 2020 for a comprehensive review), suggesting that the isotopic values in the LIP are not influenced by the weaning process. In the case of EF-1, among two analyzed infants (PEACS 4431 and ZAC7057), only ZAC7057 shows an elevated nitrogen value, which likely indicates ongoing breastmilk consumption and could potentially mimic a high intake of marine protein in the model. All other individuals analyzed are adults.
Dietary evidence and climate oscillations in Vichama
The isotope data generated in this study suggests two distinct economic regimes that are likely mirroring changes in political organization at Vichama over time. Food production systems during the EF-1 period were likely coordinated by centralized political institutions, aimed at generating surplus through intensified agriculture. By contrast, the diversified diet observed during the LIP period reflects the absence of these centralized systems at Vichama. This shift supports the existence of a diversified economy focused on small-scale farming and fishing for household consumption, with a reduced emphasis on surplus generation. These economic systems may also have been shaped by climate oscillations and the prevailing ecological conditions.
Wet and dry alternances that fueled and inhibited marine productivity, fishing activity, and, possibly, the expansion and contraction of lomas and farming fields into the region. These phenomena are linked to complex ocean-atmosphere interactions (Salvatteci et al., 2019), including oscillations in the Southern Pacific Anticyclone, trade winds, and coastal winds with effects on marine currents (Salvatteci et al., 2016, 2019; Yseki et al., 2022).
Apart from dietary adaptation, those fluctuating climate conditions could also be reflected in sociopolitical resilience and ideological response to environmental constraints, as suggested by the presence of climate-linked deities in Vichama's iconography (e.g., frog-human deity), potentially reflecting ideological adaptations to environmental constraints (Shady et al., 2015).
Reliance on marine protein during the EF-1 period might have been limited by lower biomass and marine productivity along the Peruvian coast at that time. Between 2300 and 1300 BCE, SSTs showed reduced seasonality, with cooler summers compared to present conditions (Carré et al., 2014; Loubere et al., 2013). Additionally, multidecadal fluctuations in upwelling and sea surface temperature between 1800 and 1500 BCE suggest that the EF-1 period at Vichama unfolded amid highly variable oceanographic conditions (Salvatteci et al., 2016, 2019), potentially making marine resources less predictable. Interestingly, this period corresponds to a greater reliance on C3 plants.
As argued by previous research (Caramanica et al., 2020; Contreras, 2010; Dillehay and Kolata, 2004; Lima et al., 2023; Wilson et al., 2024), these oscillations coupled with ENSO alternances, could have been produced an opportunistic intensification of certain products (fisheries/crops) when technological level, and sociopolitical conditions allow it, producing corresponding diets with verifiable marine–farming fluctuations over the time. Theoretically, under El Niño–like conditions, the concurrence of drops in marine productivity with wetter conditions (raining) along the coast and lomas flourishing, despite landslides and disasters, would lead to increased land productivity in a sort of “high-risk farming”. By comparison, under La Niña–like conditions, sea temperatures cooling, increased upwelling, and marine productivity, concomitant to a dry climate (i.e., lower rainfall and lomas contraction), would have fueled a “low-risk farming” and greater marine production, probably with more marine fauna and more maize in the record, as we can observe during the LIP.
Climate conditions during EF-1 (1800–1500 BCE) at Vichama also included relatively wet periods in the highlands and dry conditions in the coastal valleys, marked by greater seasonality in river flows, with increased precipitation in the wet season and lower runoff during the dry season (Leclerc, 2023; Salvatteci et al., 2019). The development of irrigation systems could have enabled intensified agriculture during the wet season, allowing surplus production to sustain populations through the drier months. Conversely, during the LIP, cooling sea temperatures, enhanced upwelling, and increased marine productivity, coupled with drier climates marked by reduced rainfall and contraction of lomas vegetation, may have fostered a shift toward greater reliance on marine resource exploitation alongside small-scale farming (Carré et al., 2014; Leclerc, 2023; Loubere et al., 2013; Salvatteci et al., 2019).
This study offers new insights into how climate variability shaped subsistence economies in early Andean societies and expands our understanding of the trajectory of Andean civilization. In this sense, our assessment of subsistence patterns during EF-1, a period of emerging political complexity, aligns well with the idea that decreased marine productivity may have encouraged greater reliance on, and intensification of, terrestrial resources (Wilson et al., 2024). Conversely, our findings partially support the hypothesis that stable climatic conditions favor the development of agrarian societies, whereas climatic volatility contributes to political instability, decentralization, and fragmentation (Kennett and Marwan, 2015). Opportunism in the exploitation of available resources and resilience in the face of climatic challenges appear to have been key behaviors of these societies. Climate oscillations could have represented a challenging opportunity to develop adaptive buffer strategies, such as the intensification of farming or other economic activities. This may coincide with the hegemony of coastal, inland, or highland societies over different periods.
These results should be refined and tested by future research taking into consideration some limitations: (1) palaeoceanographic and climate data are fragmentary, ambiguous, and subject of interpretation and do not necessarily can be congruent with our conclusions and (2) equifinality, that is, varying combinations of food contributions that may produce the same isotopic values in a consumer, persists as a potential issue for interpretations.
Materials and methods
Individuals and samples
This is a multiproxy stable isotope study designed to reconstruct the composition of Vichama diet in quantitative terms. Our aim is to investigate the economy of Vichama to unravel the role of marine and vegetal resources in the processes of social complexity during the EF period.
A total of 59 samples (36 bones and 23 teeth) from 38 human individuals recovered from burial contexts of Vichama were sampled for the analysis. The skeletal elements recovered for isotope analyses were mainly ribs and long bones, carpals, or cranial bones when ribs were not available. Teeth were sampled when available, and isotopic values were obtained from roots, which makes a valid direct comparison of tooth and bone samples.
Individuals' sex and age were estimated using standardized methods (Buikstra and Ubelaker, 1994). To control the potential confusing factors of breastfeeding and weaning diets, for comparisons, individuals were classified in two categories (i.e., preadults and adults) according to the tissues (i.e., teeth or bones) that originate the values (AlQahtani et al., 2010).
Radiocarbon dates
The chronological framework of this work is based on 8 direct [Accelerator Mass Spectrometry (AMS) radiocarbon dates from individuals' collagen, and fabrics from funerary contexts], and 12 indirect radiocarbon dates from associated material or stratigraphy (R. Shady personal communication, June 2024). Radiocarbon dates of Vichama come from the AMS facilities of three different laboratories using in-house protocols through commercial services. Individuals' dates were calibrated (bce/ce) with Calib 8.1.0 package (Stuiver and Reimer, 1993), using the Mixed Marine SoHCal 20 calibration curve (Heaton et al., 2020; Hogg et al., 2020), a local marine radiocarbon reservoir correction value (ΔR = 200 ± 20) estimated from paired samples (i.e., bone collagen and cotton fabrics) of the burial context ZAC7074. For individuals directly dated, we included the mean relative contribution of marine carbon to collagen calculated using Food Reconstruction Using Isotopic Transferred Signals (FRUITS) for each phase. Calibrated dates were rounded to 10 (see Supplementary material 1: S3). Individuals who were not directly dated were assigned to each period according to archaeological methods (i.e., stratigraphy, funerary patterns, pottery styles, and textiles' decorations).
Laboratory procedures
First, samples were cleaned mechanically to remove surface contaminants. Then, samples were divided for collagen and apatite extractions. Collagen was extracted in the Institut de Ciencia i Tecnologia Ambientales of the Universitat Autònoma de Barcelona (Spain).
Samples were physically cleaned and immersed in 0.6 M HCl at 4°C for several days, for demineralization, then rinsed with ultrapure water and immersed in 0.05 M sodium hydroxide (NaOH) for 20 min at room temperature. The NaOH wash cycles were repeated as needed, typically 1 or 2 times, until no further color change occurred in the solution. Samples were rinsed three times with ultrapure water for 10 min each to ensure the complete removal of NaOH. Samples were gelatinized in 0.001 M HCl (pH3) at 80°C for 48 h. The supernatant containing collagen was filtered using polyethylene Ezee filters (9 mL, pore size 60–90 μm, Elkay Laboratories Ltd.), then frozen for at least 48 h at −20°C, and freeze-dried. Stable isotope analysis of successfully extracted samples was performed at SUERC, East Kilbride (UK) using a Delta V Advantage continuous-flow isotope ratio mass spectrometer coupled via a ConfloIV to an IsoLink elemental analyzer (Thermo Scientific, Bremen, Germany). Bone collagen (approximately 0.7 mg) was combusted in the presence of oxygen in a single reactor containing tungstic oxide and copper wires at 1,020°C to produce nitrogen (N2) and carbon dioxide (CO2). A magnesium perchlorate trap was used to eliminate water produced during the combustion process, and the gases were separated in a Gas chromatography (GC) column heated between 70°C and 240°C. Helium was used as a carrier gas throughout the procedure. N2 and CO2 entered the mass spectrometer via an open split arrangement within the ConfloIV and were analyzed against their corresponding reference gases. The International Atomic Energy Agency reference materials USGS40 (L-glutamic acid, δ13C Vienna Pee Dee Belemnite (V-PDB) = −26.39 ± 0.04‰, δ15N (AIR) = −4.52 ± 0.06‰) and USGS41a (L-glutamic acid, δ13C (V-PDB) = 36.55 ± 0.08‰, δ15N (AIR) = 47.55 ± 0.15‰) were used to normalize δ15N and δ13C values. Normalization was checked using the well-characterized Elemental Microanalysis IRMS fish gelatin standard B2215 (δ13C (V-PDB) = −22.92 ± 0.10‰, δ15N (AIR) = 4.26 ± 0.12‰) and/or USGS88 (marine collagen, δ13C (V-PDB) = −16.06 ± 0.07‰, δ15N (AIR) = 14.96 ± 0.14‰). Precision is ± 0.2‰ for δ15N and ± 0.1‰ for δ13C.
For bioapatite analysis, bone samples were grounded with an agate mortar and pestle and processed in the Environmental Isotope Laboratory from the University of Arizona (USA). In this case, bone samples were cleaned in distilled water with an ultrasonic cleaner and powdered using a mortar and pestle. Samples weighing ~10 mg each were soaked for 24–72 h in a 2% sodium hypochlorite solution to remove the organic material. Then, the samples were treated with 0.1 M acetic acid to remove exogenous carbonates. The samples were also allowed to react with dehydrated phosphoric acid and silver (silver removes the sulfur gases from the evolved CO2) at 70°C in the KIEL-III Sample Preparation system. The δ13C values of the resultant CO2 were measured using the Finnigan MAT252 Isotope Ratio Mass Spectrometer. The calibration of the isotopic ratios was based on repeated measurements of NBS-19 and NBS-18 with a precision of ±0.08‰ for δ13C (1σ).
Only individuals with valid isotope values of collagen were included in the analysis. The reliability of collagen was validated following current accepted conventional preservation criteria: C/N ratio between 2.9 and 3.6 (DeNiro, 1985) and wt%C of 26%−44% and wt%N of 11%−16% (Van Klinken, 1999). We used a subsample of 10 individuals to assess the preservation of bioapatite (i.e., mineralogy and crystallinity) with ATR-FTIR.
ATR-FTIR screening was conducted at the Laboratory of Molecular Spectroscopy, Servei de Quimica—Universitat Autònoma de Barcelona using a Bruker Alpha II FTIR compact spectrometer, coupled to an Eco-ATR. Spectra of 1 mg of bone powder were collected in absorbance (A) mode with a spectral range of 400 to 4,000 cm−1° ( ), for 100 scans and a resolution of 8 cm−1. Spectra generation and baseline corrections were performed with OPUS 7.8 software. To evaluate bioapatite diagenesis, we assessed five indexes (IRSF, Infrared Splitting Factor; C/P, Carbon/Phosphate; C/C, Carbon/Carbon; BPI, Type B Phosphate index; API, Type A Phophate index) relative to conventional values for well-preserved archaeological samples (France et al., 2020). See Supplementary material 1: S4 for ATR-FTIR raw data and indexes.
), for 100 scans and a resolution of 8 cm−1. Spectra generation and baseline corrections were performed with OPUS 7.8 software. To evaluate bioapatite diagenesis, we assessed five indexes (IRSF, Infrared Splitting Factor; C/P, Carbon/Phosphate; C/C, Carbon/Carbon; BPI, Type B Phosphate index; API, Type A Phophate index) relative to conventional values for well-preserved archaeological samples (France et al., 2020). See Supplementary material 1: S4 for ATR-FTIR raw data and indexes.
Isotopic analysis and BSIMM models
To identify trends of isotope values we used the “classic” scatterplots of isotopic values (i.e., δ13Ccoll vs. δ15N; δ13Ccoll vs. δ13Cap) and rKin -Standard Ellipse Areas (Albeke, 2017). After evaluation for normal distribution with Shapiro–Wilk test for normality (α = 0.05), descriptive statistics of isotope values and comparisons by sex (i.e., males, females and undetermined) and wide age-categories (i.e., preadults and adults) were performed using Kruskal–Wallis (α =0.05) and Mann–Whitney (α = 0.05) tests with SPSS v.29 (Microsoft®). Kernel Density analysis and graphics were performed with the function geom_density() of the ggplot2 package in R environment.
The BSIMM software FRUITS (v. 2.1.1 Beta program; Fernandes et al., 2014, 2015), was employed to estimate the proportional calorie contribution of different food sources to the diet using three dietary proxies δ13Ccoll, δ15N and δ13Cap (see Supplementary material 2 for the FRUITS model's implementation). In this research, we run individuals' estimations. The consumer data for individuals consist of their δ13Ccol, δ15Ncol, and δ13Cap isotopic values. For Vichama, as a group, we use the average of isotope values and the standard error of the mean.
We charged our model with four potential food sources consumed (Food groups), with their respective composition of macro-nutrients (Food fractions: bulk, protein, and energy): terrestrial fauna, providing proteins and lipids; marine fauna, providing proteins and lipids; and C3 and C4 plants, providing carbohydrates and proteins. The isotopic mean values of these food groups come from previous dietary reconstructions for the Andean Central Coast (Pezo-Lanfranco et al., 2022; see Supplementary material 1: S7).
In the Central Coast of Peru, C3 plants (including tubers, legumes, annuals, and fruits) have mean δ13C and δ15N values of −25.7 ± 2.1‰ and +5.3 ± 3.2‰, respectively. The C4 plants (restricted to Zea mays in such cases) have mean δ13C and δ15N values of −11.3 ± 1.4‰ and +7.3 ± 3.0‰. The CAM plants have δ13C mean values of −10.8 ± 0.2‰ that overlap with C4, and δ15N of +3.8 ± 0.4‰. Terrestrial faunal (e.g., cervids, rodents, and camelids) show δ13Ccoll and δ15N mean values ranging between −17.5 ± 3.2‰ and +9.0 ± 3.1‰. The δ13C and δ15N mean values for marine fauna (including fishes, little fishes such as anchovy and sardines, mollusks, and marine birds and mammals from modern and archaeological origin) are −11.9 ± 2.1‰ and +13.2 ± 3.0‰, respectively (Pezo-Lanfranco and Colonese, 2024). The δ13C values from modern fauna specimens were corrected for the “fossil fuel effect” adjusting the values by +1.5‰ (Marino and McElroy, 1991). To avoid issues with preservation (Szpak and Chiou, 2019), only values from modern plants were used to feed the model.
Isotope fractionation factors, derived from experimental studies, were established at +4.8 ± 0.5‰ between diet and collagen, +10.1 ± 0.5‰ between diet and apatite, +5.5 ± 0.5 ‰ for δ15N diet to collagen (Fernandes et al., 2014, 2015).
The weighted values of each fraction of macronutrients (lipids, carbohydrates, and proteins) followed published parameters (Fernandes et al., 2014, 2015; Pezo-Lanfranco et al., 2022; Pezo-Lanfranco and Colonese, 2024). The values of δ13Cap represent the total carbon mix in the diet. Therefore, we use the same bulk value for each food group. Terrestrial and marine fauna δ13C bulk values were estimated as a weighted mean of lipid and protein δ13C values (Fernandes et al., 2014, 2015). For collagen, the carbon of the protein and energy routed to the total collagen was established at 74 ± 4% and 26%, respectively (Fernandes et al., 2012). We assumed that nitrogen was derived exclusively from proteins (100%). Lipids and carbohydrates were added to the model as “energy”. To integrate this combinatorial effect, we applied a “concentration-dependent” model (Fernandes et al., 2014, 2015).
Following previous reconstructions using FRUITS (Fernandes et al., 2014, 2015; Pezo-Lanfranco et al., 2022) the isotopic composition of each nutritional fraction (protein, carbohydrates, and lipids) was obtained from the mean values of δ13Ccoll and δ15Ncoll using the following fractionation factors: −2‰ (Δ13Cprotein − collagen), −8‰ (Δ13Clípids−collagen), and +2‰ (Δ15Nprotein − collagen) for terrestrial mammals and −1‰ (Δ13Cprotein − collagen), −7‰ (Δ13Clípids−collagen), and +2‰ (Δ15Nprotein − collagen) for marine animals. For plants, the offsets were −2‰ (Δ13Cbulk − protein) and +0.5‰ (Δ13Cbulk − lipids), while for the δ15N value of plant protein, the known value of δ15N recorded for the plant was assumed. The carbon weight (concentrations) of each food fraction (protein and energy) from each food group was calculated according to its macronutrient composition (Fernandes et al., 2014, 2015; Pezo-Lanfranco et al., 2022; see details in Supplementary material 2). A physiological, conservative, and acceptable range of protein consumption stipulated between 5% and 45% of the total calories was charged as “prior” (Fernandes et al., 2014). The estimates of FRUITS models reflect carbon content or equivalent calorie contributions expressed as relative contributions (adding up to 1 or 100%) with an associated 1σ uncertainty (Fernandes et al., 2015).
Finally, a Kullback–Leibler divergence test (Lyman, 2008) was performed to estimate the statistical difference between the probability distributions of the four food groups between periods (EF-1 vs. LIP). This test was executed with the Entropy package of the R environment (Supplementary material 2).
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Author contributions
LP-L: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AC: Conceptualization, Data curation, Investigation, Project administration, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AP-B: Data curation, Investigation, Validation, Visualization, Writing – original draft, Writing – review & editing. TA: Data curation, Investigation, Validation, Visualization, Writing – original draft, Writing – review & editing. MM: Investigation, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing. MY: Data curation, Investigation, Validation, Writing – original draft, Writing – review & editing. MG: Data curation, Investigation, Validation, Writing – original draft, Writing – review & editing. LM: Data curation, Investigation, Validation, Writing – original draft, Writing – review & editing. JA: Data curation, Investigation, Validation, Writing – original draft, Writing – review & editing. AD: Data curation, Formal analysis, Software, Visualization, Writing – original draft, Writing – review & editing. PN: Conceptualization, Data curation, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. ACC: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Software, Supervision, Writing – original draft, Writing – review & editing. RS: Conceptualization, Data curation, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The Vichama project was funded by the Zona Arqueológica Caral Supe (UE 003, Ministry of Culture of Peru). The research has also received funding from the European Union's Horizon 2020 research and innovation program under the Marie Sklodowska-Curie Action project PACHAMAMA, Grant Agreement No. 101062179 (LP-L) and the ERC Consolidator project TRADITION, Grant Agreement No. 817911 (ACC). This work contributes to the “ICTA-UAB María de Maeztu” Program for Units of Excellence of the Spanish Ministry of Science and Innovation (CEX 2019-000940-M). This work also contributes to EarlyFoods (Evolution and impact of early food production systems), funded by the Agència de Gestió d'Ajuts Universitaris i de Recerca de Catalunya (SGR-Cat-2021, 00527). The funders had no role in the study design, data collection and analysis, the decision to publish, or the preparation of the manuscript.
Acknowledgments
The authors are grateful to Sonia Lopez and her field team for their valuable collaboration during the sampling work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fearc.2025.1611071/full#supplementary-material
References
Aguilar, M. A. (2006). “Surgimiento de las sociedades complejas en los andes centrales,” in Una perspectiva desde el valle de Huaura, Perú. Licentiate Monography (Lima: Universidad Nacional Federico Villarreal).
Albeke, S. E. (2017). “rKIN: (kernel) isotope niche estimation,” in R Package Version 0.1. Available online at: https://cran.r-project.org/web/packages/rKIN/rKIN.pdf
AlQahtani, S. J., Hector, M. P., and Liversidge, H. M. (2010). Brief communication: the London atlas of human tooth development and eruption. Am. J. Phys. Anthropol. 142, 481–490. doi: 10.1002/ajpa.21258
Ambrose, S. H., and Norr, L. (1993). “Experimental evidence for the relationship of carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate,” in Prehistoric Human Bone: Archaeology at the Molecular Level (Berlin: Springer), 1–37.
Buikstra, J., and Ubelaker, D. (1994). Standards for Data Collection from Human Skeletal Remains. Fayetteville: Arkansas Archeological Survey Research Series, No.44.
Cadwallader, L., Beresford-Jones, D. G., Whaley, O., and O'Connell, T. (2012). The sign of maize? A reconsideration of what 13C values say about Paleodiet in the Andean Region. Hum. Ecol. 40, 487–509. doi: 10.1007/s10745-012-9509-0
Caramanica, A., Huaman Mesia, L., Morales, C. R., Huckleberry, G., Castillo, L. J., and Quilter, J. (2020). El Niño resilience farming on the north coast of Peru. Proc. Natl. Acad. Sci. U.S.A. 117:24127. doi: 10.1073/pnas.2006519117
Cárdenas, M. (1977). Informe Preliminar del Trabajo de Campo en el Valle de Huaura (Departamento de Lima). Lima: Instituto Riva Agüero, Seminario de Arqueología, PUCP.
Carré, M., Sachs, J. P., Purca, S., Schauer, A. J., Braconnot, P., Angeles-Falcón, R., et al. (2014). Holocene history of ENSO variance and asymmetry in the eastern tropical Pacific. Science 345, 1045–1048. doi: 10.1126/science.1252220
Cheung, C., and Szpak, P. (2021). Interpreting past human diets using stable isotope mixing models. J. Archaeol. Method Theory 28, 1106–1142. doi: 10.1007/s10816-020-09492-5
Chisholm, B. S., Nelson, D. E., and Schwarcz, H. P. (1982). Stable-carbon isotope ratios as a measure of marine versus terrestrial protein in ancient diets. Science 216, 1131–1132. doi: 10.1126/science.216.4550.1131
Chu, A. (2011). Household organization and social inequality at Bandurria, a Late Preceramic village in Huaura, Peru (PhD. Dissertation). University of Pittsburgh, Pittsburgh, PA, United States.
Contreras, D. A. (2010). Landscape and environment: Insights from the Prehispanic Central Andes. J. Arch. Res. 18, 241–288. doi: 10.1007/s10814-010-9038-6
Coutts, K., Chu, A., and Krigbaum, J. (2011). Paleodiet in Late Preceramic Peru: Preliminary Isotopic data from Bandurria. J. Island Coast. Archeol. 6, 196–210. doi: 10.1080/15564894.2011.586086
Creamer, W., Ruiz, A., Perales, M., and Haas, J. (2013). The Fortaleza Valley, Peru: Archaeological investigation of late archaic sites (3000–1800 BC). Field. Anthropol. 44, 1–108 doi: 10.3158/0071-4739-44.0.1
DeNiro, M. J. (1985). Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to paleodietary reconstruction. Nature 317, 806–809. doi: 10.1038/317806a0
DeNiro, M. J., and Epstein, S. (1981). Influence of diet on the distribution of nitrogen isotopes in animals. Geochim. Cosmochim. Acta 45 341–351. doi: 10.1016/0016-7037(81)90244-1
Dillehay, T. D. (2017). Where the Land Meets the Sea: Fourteen Millennia of Human History at Huaca Prieta, Peru. Austin: University of Texas Press.
Dillehay, T. D., Elling, H., and Rossen, J. (2005). Preceramic irrigation canals in the Peruvian Andes. Proc. Natl. Acad. Sci. U.S.A. 102, 17241–17244. doi: 10.1073/pnas.0508583102
Dillehay, T. D., and Kolata, A. L. (2004). Long-term human response to uncertain environmental conditions in the Andes. Proc. Natl. Acad. Sci. U.S.A. 101, 4325–4330. doi: 10.1073/pnas.0400538101
Dillehay, T. D., Rossen, J., Andres, T. C., and Williams, D. E. (2007). Preceramic adoption of peanut, squash, and cotton in northern Peru. Science 316, 1890–1893. doi: 10.1126/science.1141395
Engel, F. A. (1987). “De las Begonias al Maíz,” in Vida y Producción en el Perú Antiguo (Lima: Centro de Investigaciones de Zonas Áridas CIZA, Universidad Nacional Agraria La Molina).
Fernandes, R., Grootes, P. M., Nadeau, M. J., and Nehlich, O. (2015). Quantitative diet reconstruction of a neolithic population using a Bayesian mixing model (FRUITS): The case study of Ostorf (Germany). Am. J. Phys. Anthropol. 158, 325–340 doi: 10.1002/ajpa.22788
Fernandes, R., Millard, A. R., Brabec, M., Nadeau, M. J., and Grootes, P. M. (2014). Food reconstruction using isotopic transferred signals (FRUITS): a Bayesian model for diet reconstruction. PLoS ONE 9:e87436. doi: 10.1371/journal.pone.0087436
Fernandes, R., Nadeau, M. J., and Grootes, P. M. (2012). Macronutrient-based model for dietary carbon routing in bone collagen and bio-apatite. Archaeol. Anthropol. Sci. 4, 291–301. doi: 10.1007/s12520-012-0102-7
France, C. A. M., Sugiyama, N., and Aguayo, E. (2020). Establishing a preservation index for bone, dentin, and enamel bioapatite mineral using ATR-FTIR. J. Arch. Sci. Rep. 33, 102551. doi: 10.1016/j.jasrep.2020.102551
Goldberg, A., Mychajliw, A. M., and Hadly, E. A. (2016). Post-invasion demography of prehistoric humans in South America. Nature 532, 232–235. doi: 10.1038/nature17176
Grobman, A., Bonavia, D., Dillehay, T. D., Piperno, D. R., Iriarte, J., and Holst, I. (2012). Preceramic maize from Paredones and Huaca Prieta Peru. Proc. Natl. Acad. Sci. U. S. A. 109, 1755–1759. doi: 10.1073/pnas.1120270109
Haas, J., and Creamer, W. (2006). Crucible of Andean Civilization: The Peruvian coast from 3000 to 1800 BC. Curr. Anthropol. 47, 745–775. doi: 10.1086/506281
Haas, J., Creamer, W., Huamán, L., Goldstein, D., Reinhard, K., and Vergel, C. (2013). Evidence for maize (Zea mays) in the Late Archaic (3000–1800 B.C.) in the Norte Chico region of Peru. Proc. Natl. Acad. Sci. U.S.A. 110, 4945–4949 doi: 10.1073/pnas.1219425110
Heaton, T., Köhler, P., Butzin, M., Bard, E., Reimer, R., Austin, W., et al. (2020). Marine20 - the marine radiocarbon age calibration curve (0–55,000 cal BP). Radiocarbon 62, 779–820. doi: 10.1017/RDC.2020.68
Hedges, R. E. M., and Reynard, L. M. (2007). Nitrogen isotopes and the trophic level of humans in archaeology. J. Archaeol. Sci. 34, 1240–1251. doi: 10.1016/j.jas.2006.10.015
Herbert, T. (2003). “Alkenone paleotemperature determinations,” in Treatise on Geochemistry: The Oceans and Marine Geochemistry, ed. H. Elderfeld (New York: Elsevier), 391–432.
Hogg, A., Heaton, T., Hua, Q., Palmer, J., Turney, C., Southon, J., et al. (2020). SHCal20 southern hemisphere calibration, 0–55,000 years cal BP. Radiocarbon 62, 759–778. doi: 10.1017/RDC.2020.59
IMARPE (2010). Informe: Delimitación y caracterización de bancos naturales de invertebrados bentónicos comerciales y áreas de pesca artesanal en la región Lima entre Punta Litera-Playa Grande, Barranca y Huaura de la Region Lima. Lima: Instituto del Mar del Perú.
INRENA (2005). Inventario y monitoreo de las aguas subterráneas en el valle Huaura: Anexos (Lima: Autoridad Nacional del Agua). Available online at: https://hdl.handle.net/20.500.12543/2037 (Accessed July 7, 2023).
Jim, S., Ambrose, S., and Evershed, R. P. (2004). Stable carbon isotopic evidence for differences in the dietary origin of bone cholesterol, collagen, and apatite: implications for their use in paleodietary reconstruction. Geochim. Cosmochim. Acta 68, 61–72. doi: 10.1016/S0016-7037(03)00216-3
Kellner, C. M., and Schoeninger, M. J. (2007). A simple carbon isotope model for reconstructing human diet. Am. J. Phys. Anthropol. 133, 1112–1127. doi: 10.1002/ajpa.20618
Kennett, D. J., and Marwan, N. (2015). Climatic volatility, agricultural uncertainty, and the formation, consolidation and breakdown of preindustrial agrarian states. Phylos. T. R. Soc. A 373:20140458. doi: 10.1098/rsta.2014.0458
Leclerc, E. L. (2023). Coast and highland paleoclimate of the north-central Peruvian Andes and its influence on coastal water availability and cultural development, 6000 to 2000 cal BP. Quat. Sci. Rev. 314:108209. doi: 10.1016/j.quascirev.2023.108209
Lima, M., Gayó, E. M., Gurruchaga, A., Estay, S. A., and Santoro, C. M. (2023). 1000 years of population, warfare, and climate change in pre-Columbian societies of the Central Andes. PLoS ONE 18:e0278730. doi: 10.1371/journal.pone.0278730
Loubere, P., Creamer, W., and Haas, J. (2013). Evolution of the El Nino-Southern Oscillation in the late Holocene and insolation driven change in the tropical annual SST cycle. Glob. Planet. Change 100:129e144. doi: 10.1016/j.gloplacha.2012.10.007
Mächtle, B., and Eitel, B. (2013). Fragile landscapes, fragile civilizations - how climate determined societies in the pre-Columbian south Peruvian Andes. Catena 103, 62–73 doi: 10.1016/j.catena.2012.01.012
Makowski, K. (2023). Pre-Hispanic andean urbanism and its ‘anti-urban' peculiarities. J. Urban Archaeol. 8, 165–196. doi: 10.1484/J.JUA.5.135664
Marino, B. D., and McElroy, M. B. (1991). Isotopic composition of atmospheric CO2 inferred from carbon in C4 plant cellulose. Nature 349, 127–131. doi: 10.1038/349127a0
Minagawa, M., and Wada, E. (1984). Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochim. Cosmochim. Acta 48, 1135–1140. doi: 10.1016/0016-7037(84)90204-7
Moseley, M. E. (1975). The Maritime Foundations of Andean Civilization. Menlo Park, CA: Cummings Publishing Company.
Orloff, C. R. (2022). Caral, South America's Oldest City (2600–1600 BC): ENSO environmental changes influencing the late archaic period site on the North Central Coast of Peru. Water 14:1403. doi: 10.3390/w14091403
Pezo-Lanfranco, L., and Colonese, A. C. (2024). The role of farming and fishing in the rise of social complexity in the Central Andes: a stable isotope perspective. Sci. Rep. 14:4582. doi: 10.1038/s41598-024-55436-4
Pezo-Lanfranco, L., Crispin, A., Machacuay, M., Novoa, P., and Shady, R. (2020). Isotopic evidence of weaning behaviors from farming communities of the Peruvian North-Central Coast. Insights into the demography of the transition from the middle to the late formative periods (500-400 B.C.). Bioarchaeol. Int. 3–4, 191–216. doi: 10.5744/bi.2020.3019
Pezo-Lanfranco, L., Machacuay, M., Novoa, P., Peralta, R., Mayer, E., Eggers, S., et al. (2022). The diet at the onset of the Andean Civilization: New stable isotope data from Caral and Áspero, North-Central Coast of Peru. Am. J. Biol. Anthropol. 177, 402–424. doi: 10.1002/ajpa.24445
Pezo-Lanfranco, L., Pezo-Lanfranco, S., and Eggers, S. (2009). Exostosis auditiva como marcador osteológico de actividad acuática en poblaciones formativas de la Costa Norte del Perú. Paleopatología 6, 1−18. Available online at: https://asociacionpaleopatologia.es/revista-paleopatologia/#andISSN1579-0606
Phillips, D. L. (2012). Converting isotope values to diet composition: the use of mixing models. J. Mammal. 93, 342–352. doi: 10.1644/11-MAMM-S-158.1
Piperno, D. R., and Dillehay, T. D. (2008). Starch grains on human teeth reveal early broad crop diet in northern Peru. Proc. Natl. Acad. Sci. U.S.A. 105, 19622–19627. doi: 10.1073/pnas.0808752105
Salvatteci, R., Gutiérrez, D., Field, D., Sifeddine, A., Ortlieb, L., Bouloubassi, I., et al. (2014). The response of the Peruvian Upwelling Ecosystem to centennial-scale global change during the last two millennia. Clim. Past 10, 715–731. doi: 10.5194/cp-10-715-2014
Salvatteci, R., Gutierrez, D., Sifeddine, A., Ortlieb, L., Druffel, E., Boussafir, M., et al. (2016). Centennial to millennial-scale changes in oxygenation and productivity in the eastern tropical South Pacific during the last 25000 years. Quat. Sci. Rev. 131, 102–117. doi: 10.1016/j.quascirev.2015.10.044
Salvatteci, R., Schneider, R. R., Blanz, T., and Mollier-Vogel, E. (2019). Deglacial to Holocene ocean temperatures in the humboldt current system as indicated by alkenone paleothermometry. Geophys. Res. Lett. 46, 281–292. doi: 10.1029/2018GL080634
Sandweiss, D. H., Shady, R., Moseley, M., Keefer, D. K., and Orloff, C. R. (2009). Environmental change and economic development in coastal Peru between 5,800 and 3,600 years ago. Proc. Natl. Acad. Sci. USA 106, 1359–1363. doi: 10.1073/pnas.0812645106
Schoeninger, M. J. (2010). Diet reconstruction and ecology using stable isotope ratios, in A companion to Biological Anthropology (Hoboken, NJ: Willey-Blackwell), 445–464.
Schoeninger, M. J., and DeNiro, M. J. (1984). Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim. Cosmochim. Acta 48, 625–639. doi: 10.1016/0016-7037(84)90091-7
Schoeninger, M. J., DeNiro, M. J., and Tauber, H. (1983). Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220, 1381–1383. doi: 10.1126/science.6344217
Schoeninger, M. J., and Reitsema, L. J. (2023). “Diet reconstruction and ecology,” in A Companion to Biological Anthropology, ed. C. S. Larsen (Malden: Willey-Blackwell), 510–526.
Scholz, F., McManus, J., Mix, A., Hensen, C., and Schneider, R. (2014). The impact of ocean deoxygenation on the ocean's iron supply. Nat. Geosci. 7, 433–437. doi: 10.1038/ngeo2162
Shady, R. (2006a). La civilización Caral: sistema social y manejo del territorio y sus recursos. Su trascendencia en el proceso cultural andino. Bol. Arqueol. PUCP 10, 59–89 doi: 10.18800/boletindearqueologiapucp.200601.004
Shady, R. (2006b). “America's first city? The case of Late Archaic Caral,” in Andean Archaeology III, eds. D. W. Isbell, and H. Silverman (Boston: Springer), 28–66.
Shady, R. (2006c). “Caral-Supe and the North- Central area of Peru: The history of maize in the land where Civilization came into being,” in Histories of Maize: Multidisciplinary Approaches to the Prehistory, Linguistics, Biogeography, Domestication, and Evolution of Maize, eds. J. Staller, R. Tykot, and B. Benz (Cambridge, MA: Academic Press), 381–402.
Shady, R. (2018). El sistema social de Caral y su trascendencia: El manejo transversal del territorio, la complementariedad social y política y la interacción intercultural (Lima: Nayra Kunan Pacha, Revista de Arqueología Social, Proyecto Especial Arqueológico Caral-Supe), 19–90.
Shady, R., and Cáceda, D. (2008). Áspero, la ciudad pesquera de la civilización Caral - Supe, recuperamos su historia para vincularla con nuestro presente Proyecto Especial Arqueológico Caral – Supe. Lima: Instituto Nacional de Cultura.
Shady, R., Crispín, A., and Quispe, E. (2008). “Vichama de Végueta comparte con Caral 5000 años de Identidad Cultural,” in Proyecto Especial Arqueológico Caral-Supe (Lima: Instituto Nacional de Cultura).
Shady, R., Haas, J., and Creamer, W. (2001). Dating Caral, a Preceramic urban center in the Supe Valley on the central coast of Peru. Science 292, 723–726. doi: 10.1126/science.1059519
Shady, R., Machacuay, M., Quispe, E., Novoa, P., and Leyva, C. (2015). Vichama, historia social de la Civilización en Végueta. 3800 años de memoria colectiva de nuestros ancestros de Vichama ante el cambio climático. Lima: Biblioteca Nacional del Perú.
Shoji, K., Vásquez, V. F., and Rosales, T. E. (2023). Starch grains on human teeth as evidence for 4000 BCE potato consumption at the Cruz Verde site, northern coast of Peru. J. Archaeol. Sci. Rep. 51:104152. doi: 10.1016/j.jasrep.2023.104152
Stuiver, M., and Reimer, P. J. (1993). CALIB rev. 8. Radiocarbon 35, 215–230. doi: 10.1017/S0033822200013904
Szpak, P., and Chiou, K. L. (2019). A comparison of nitrogen isotope compositions of charred and desiccated botanical remains from northern Peru. Veg. Hist. Archaeobot. 29, 527–538. doi: 10.1007/s00334-019-00761-2
Tello, J. C. (2004). La Muralla de Santa. Arqueología y Sociedad 15, 11–32. doi: 10.15381/arqueolsoc.2004n15.e12731
Tung, T. A., Dillehay, T. D., Feranec, R., and DeSantis, L. R. G. (2021). Early specialized maritime and maize economies on the north coast of Peru. Proc. Natl. Acad. Sci. U. S. A. 117, 32308–32319. doi: 10.1073/pnas.2009121117
Van Klinken, G. J. (1999). Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J. Arch. Sci. 26, 687–695. doi: 10.1006/jasc.1998.0385
Vega-Centeno, R. (2005). Ritual and Architecture in a Context of Emergent Complexity: A Perspective from Cerro Lampay, a Late Archaic Site in the Central Andes (PhD. Dissertation). The University of Arizona, Tucson, AZ, United States.
Wefer, G., Fischer, G., and Iversen, M. (2015). “Export production,” in Encyclopedia of Marine Geosciences, eds. J. Harff, M. Meschede, S. Petersen, and J. Thiede (Dordrecht: Springer).
Wilson, K. M., McCool, W. C., Brewer, S. C., Zamora-Wilson, N., Schryver, P. J., Lamson, R. L. F., et al. (2022). Climate and demography drive 7000 years of dietary change in the Central Andes. Sci. Rep. 12:2026. doi: 10.1038/s41598-022-05774-y
Wilson, K. M., McCool, W. C., and Coltrain, J. B. (2024). Climate and oceanic condition changes influence subsistence economic adaptation through intensification on the Central Andean coasts. Quat. Int. 689, 79–92. doi: 10.1016/j.quaint.2023.07.002
Yseki, M., Crispín, A., Abad, T., Prado, A., Novoa, P., and Shady, R. (2024). First comprehensive macroremains analysis of edible plants in Vichama site, Peru during the Early Formative Period 1(1800-1500 BCE). Veg. Hist. Archaeobot. 34, 91–100. doi: 10.1007/s00334-024-01004-9
Yseki, M., Pezo-Lanfranco, L., Machacuay, M., Novoa, P., and Shady, R. (2023). Analysis of starch grains trapped in human dental calculus in Áspero, Peru during the Initial Formative Period (3000–1800 BCE). Sci. Rep. 13:14143. doi: 10.1038/s41598-023-41015-6
Yseki, M., Turcq, B., Caquineau, S., Salvatteci, R., Solis, J., Skilbeck, C. G., et al. (2022). Millennial variability of terrigenous transport to the central–southern Peruvian margin during the last deglaciation (18–13 kyr BP). Clim. Past. 18, 2255–2269. doi: 10.5194/cp-18-2255-2022
Keywords: Andean formative period, dietary reconstruction, Bayesian stable isotope mixing models, climate change, Peruvian North-Central coast
Citation: Pezo-Lanfranco L, Crispin A, Prado-Barragán A, Abad T, Machacuay M, Yseki M, Gorriti M, Miranda L, Apolín J, DiMuro A, Novoa P, Colonese AC and Shady R (2025) Refining dietary shifts linked to climate oscillations in the Central Andes: stable isotope evidence from Vichama (1800–1500 BCE). Front. Environ. Archaeol. 4:1611071. doi: 10.3389/fearc.2025.1611071
Received: 13 April 2025; Accepted: 30 June 2025;
Published: 18 July 2025.
Edited by:
Michelle S. Eusebio, University of the Philippines Diliman, PhilippinesReviewed by:
Kurt Wilson, Lawrence University, United StatesWeston McCool, University of Utah Department of Anthropology, United States
Copyright © 2025 Pezo-Lanfranco, Crispin, Prado-Barragán, Abad, Machacuay, Yseki, Gorriti, Miranda, Apolín, DiMuro, Novoa, Colonese and Shady. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luis Pezo-Lanfranco, THVpcy5QZXpvQHVhYi5jYXQ=
†These authors share senior authorship