 Claudia Speciale1,2*
Claudia Speciale1,2* Marialetizia Carra3Fabio Fiori3Vito Giuseppe Prillo4Ethel Allué1,2Maurizio Cattani5
Marialetizia Carra3Fabio Fiori3Vito Giuseppe Prillo4Ethel Allué1,2Maurizio Cattani5- 1IPHES-CERCA, Catalan Institute of Human Palaeoecology and Social Evolution, Tarragona, Spain
- 2Departament d'Història i Història de l'Art, Universitat Rovira i Virgili (URV), Tarragona, Spain
- 3ArcheoLaBio, Bioarchaeological Research Centre, Department of History and Culture, University of Bologna, Bologna, Italy
- 4Department of Cultural Heritage, University of Padua, Padua, Italy
- 5Department of History and Cultures, University of Bologna, Bologna, Italy
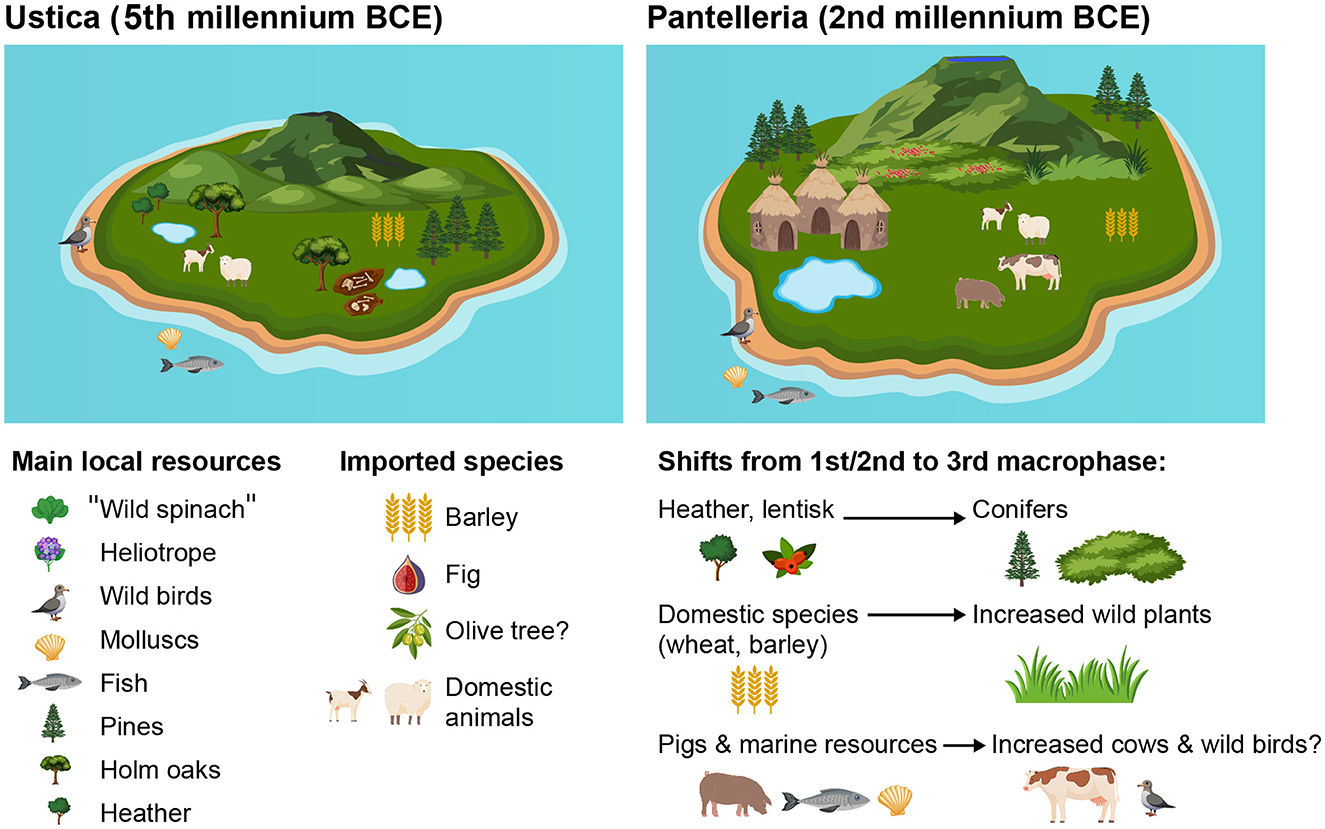
This study investigates prehistoric human adaptation to small Mediterranean island environments through the archaeobotanical and zooarchaeological analysis of two key sites: Piano dei Cardoni (Ustica, Neolithic) and Mursia (Pantelleria, Bronze Age). Both volcanic islands, differing in size, landscape features, and chronology of occupation, offer an ideal framework to explore how early human communities managed limited insular resources, navigated ecological constraints, and established sustainable subsistence systems. On Ustica, permanent settlement during the Middle Neolithic (c. 4800–4300 BCE) is reflected in a diversified exploitation of local vegetation and faunal resources. Archaeobotanical evidence suggests the co-occurrence of plant species linked to natural and anthropogenic habitats, including barley, fig, olive, and lentisk, possibly indicating deliberate introduction of tree crops for long-term occupation. Shrubland and woodland species were utilized without evident overexploitation, and faunal remains show a subsistence economy based on ovicaprines, complemented by wild birds and marine resources. The absence of large mammals and the reduced size of domesticates highlight adaptive strategies in a resource-limited setting. In contrast, Bronze Age Pantelleria (c. 1800–1500 BCE) shows a more structured subsistence model within a larger, ecologically more complex island. The Mursia settlement reveals a shift in plant use across its occupational phases, from cereal- and pulse-based agriculture to increased exploitation of wild plants like purslane and crop trees like fig. Wood charcoal indicates probably a technological selection of plant species, with a prevalence of pine, juniper, and heather. Zooarchaeological data reveals a dominant use of ovicaprines, marine fish, and mollusks, with a possible reorganization of livestock strategies over time. Comparative analysis reveals both continuity and divergence in insular adaptation. While both sites demonstrate human resilience through mixed subsistence strategies combining agriculture, foraging, and marine exploitation, local environmental and cultural factors drove distinct responses. The findings underscore the importance of interdisciplinary approaches in understanding human-environment interactions and the role of islands as dynamic laboratories of ecological and cultural experimentation during prehistory.
Graphical Abstract
1 Introduction
The historical landscape heterogeneity of the Mediterranean region exemplifies how humans can, through short-term decision-making, “create productive ecosystems in otherwise marginal environments” (Braje et al., 2017). The negative effects of human impact have resulted in thriving human-made ecosystems that generally possess resilience to withstand fluctuations. The creation of complex ecosystemic interactions was paradoxically triggered by the advent of humans, as shown by an initial increase in species richness (Lomolino and Van der Geer, 2023). This process ultimately gave life to a less biodiverse landscape, yet maintaining most of the time the same structure since later prehistory.
Beyond the general trend in biodiversity loss, resources on small islands (< 100 km2), regardless of geomorphology, altitude or other geographical features, are inherently limited (Keegan et al., 2008). This limitation arises from the intrinsic trophic constraints of islands, which are dependent on their area (Brose et al., 2004). However, the concept of “adaptiveness” in human colonization is shaped not only by natural factors, rather by the choices that humans make and how they respond to the challenges of the new insular environment (Giovas, 2016).
The management of resources, notably of subsistence resources, is then of crucial importance to achieve a successful long-term colonization and settlement by the newcomers, especially if they are on the top of the trophic chain like humans (Newsom and Wing, 2004) and significantly exposed to stochastic perturbations (Cherry and Leppard, 2018). Colonization by hunter-gatherer-fisher groups generally resulted in less drastic environmental changes compared to those by agricultural societies, which often introduced a wider range of external (both cultivated and wild) species of plants and animals, whether on purpose or unintentionally (Hofman and Rick, 2018; Plekhov et al., 2021).
Unless there are natural sedimentary sequences to compare with—and this happens mostly on the bigger islands—the effects of human arrival on islands are visible almost exclusively through the archaeobotanical and zooarchaeological datasets (Pasta et al., 2022), then a reconstruction of the pre-human environment is not always easily conceivable. In these cases, archaeological data represent the almost unique paleoenvironmental and paleovegetational proxies available for the researchers (Pasta and Speciale, 2021).
Among the others, the introduction of ovicaprines is a key factor in the transformation of the pre-human vegetation on small islands, particularly due to the natural absence of large wild herbivores. Goats' high tolerance for a wide range of climatic conditions, allows introduced populations to grow rapidly if not kept under control (Leppard and Pilaar Birch, 2019). This dynamic has recently drawn attention on the island of Alicudi (Aeolian Archipelago, Messina, Sicily), where feral goats have thrived in the context of widespread landscape abandonment (Figure 1). Their presence could quickly lead to overgrazing, especially on an island of 5 km2. Moreover, the introduction of other species that can affect local vegetation like rodents and lagomorphs or small carnivores could be directly or indirectly human induced (Masseti, 2003), sometimes since the early Neolithic (Trantalidou, 2008).

Figure 1. One of the feral goats on the top of Alicudi island (September 2023).
The reliance of farmer communities on a broad and diverse suite of plant and animal species contributed to overcome the inherent trophic limitations of the Mediterranean islands, making even very small and marginal island environments viable for human settlements (e.g., McLaughlin et al., 2018; Scerri et al., 2025). The prevalence of more aridity-tolerant ovicaprines over cattle and pigs at sites on smaller, ecologically marginal islands provides supporting evidence (Ramis, 2014). The same applies to crops, with a significant reliance primarily on hulled barley (Speciale, 2021; Speciale et al., 2023, 2024a).
This study addresses the following research questions in the two case studies, Ustica and Pantelleria (Figure 2):
- How did prehistoric human communities adapt their subsistence and settlement strategies to the ecological constraints of small volcanic central Mediterranean islands?
- What archaeobotanical and zooarchaeological indicators can be interpreted as signals of human adaptation?
- Do these indicators vary through time, particularly between the Middle Neolithic (mid-5th millennium BCE) and the Middle Bronze Age (1800–1500 BCE)?
Figure 2. Geographic position of the two case studies: Ustica and Pantelleria.
2 Geographic framework
The island of Ustica is an extinct volcano located 60 km off the northwestern Sicilian coast, west of the Aeolian islands. The emerged part of Ustica covers an area of less than 9 km2 and reaches an elevation of 248 m a.s.l., in Mt. Guardia dei Turchi (Figure 3A). Ustica is made from volcanic rock, and to a lesser extent from marine and continental sedimentary deposits (de Vita and Foresta Martin, 2017). The island is quite suitable for the human settlement, compared to other volcanic islands, thanks to the presence of two extensive former marine abrasion platforms on both its northern and southern sides, which offer ideal conditions for habitation and cultivation. Furthermore, the almost total absence of slopes and the geochemical nature of the soils allow the creation of small seasonal ponds (gorghi in the local dialect) for the natural collection of rainwater (Speciale et al., 2023). Piano dei Cardoni, a Neolithic site, is located in the southeastern part of the island, not far from the natural harbor of Cala Santa Maria, well sunlit and protected by the strong mistral winds.

Figure 3. (A) Ustica and the position of Piano dei Cardoni site; (B) Pantelleria and the position of Mursia site.
Ustica's modern vegetation is strongly influenced by its volcanic origin and Mediterranean climate. According to the most recent census (2009) the island still hosts approximately 400 vascular plant taxa (Speciale et al., 2023). The coastline hosts halophilous plants such as Crithmum maritimum and Limonium species. Inland, Mediterranean maquis dominates, with dense stands of Pistacia lentiscus, Phillyrea latifolia, and wild olive (Olea europaea var. sylvestris). On rocky slopes, garigue vegetation appears, composed of low shrubs like Cistus and Euphorbia dendroides. Aromatic herbs, including thyme (Thymus capitatus) and rosemary (Salvia rosmarinus), are widespread. Prickly pear (Opuntia ficus-indica) has naturalized, although it is introduced and capers (Capparis spinosa) grow spontaneously among walls and cliffs. Higher and more sheltered areas contain patches of Aleppo pine (Pinus halepensis).
Concerning the fauna present on the island, it is modest on land, with reptiles like the Italian wall lizard (Podarcis sicula) and a few small mammals, but its coastal and marine environments are particularly rich. Along the seashores, rocky habitats host numerous invertebrates such as crabs, mollusks, and echinoderms adapted to tidal conditions. The cliffs provide nesting sites for seabirds including yellow-legged gulls (Larus michahellis), shearwaters (Calonectris diomedea), and European shags (Phalacrocorax aristotelis). The island also lies on a migratory route, attracting raptors and passerines that rest along coastal areas. In the surrounding waters, groupers, barracudas, and pelagic species are commonly found, while sea turtles and dolphins are occasionally observed.
The island of Pantelleria, of volcanic origin as well, is in the middle of the Mediterranean Sea, between Sicily and Tunisia; it is quite close to the African coast, located approximately 70 km away (Cape Mustafà near Kelibia, Tunisia), and a bit more far from the Sicilian coast (around 100 km); the highest peak of Montagna Grande reaches an altitude of 836 m a.s.l. Due to the high permeability of the soils and the local microclimatic conditions, the island has several but limited water sources, called buvire in the island dialect and easily accessible for exploitation. The surface of the island (83 km2), especially in the northern part, is characterized by flat parts or gentle slopes, suitable for agricultural use or livestock farming. The Mursia site is located in the North-West of the island (Figure 3B), it is enclosed by high cliff above the sea and a massive perimetral wall of volcanic stones.
Pantelleria exhibits a highly diversified flora (around 570 vascular taxa) shaped by its volcanic substrate, rugged topography, and marked ecological gradients. The island's vegetation ranges from coastal scrub to well-developed woodlands, reflecting both altitudinal variation and historical land use. At lower elevations, the landscape is dominated by thermophilous shrublands, where P. lentiscus, O. europaea var. sylvestris, and E. dendroides prevail. Progressively, these communities give way to more structured formations, particularly in less disturbed areas. Woodland vegetation reaches its best expression in the higher and more humid central zones, especially around the Montagna Grande. Here, extensive stands of Aleppo pine (P. halepensis) and maritime pine (Pinus pinaster) are present. More significant, however, are the remnants of natural holm oak (Quercus ilex) forests, often mixed with Arbutus unedo, Erica arborea, and P. latifolia, which constitute the climax vegetation of the island. On exposed ridges and drier slopes, woodland cover becomes fragmented, replaced by garrigue and broomfields (Genista aspalathoides) and xerophytic plant species adapted to chronic water scarcity, such as T. capitatus, S. rosmarinus, and Thymelaea spp.
Pantelleria hosts a notable diversity of reptiles, birds, and invertebrates. Reptilian species include the Italian wall lizard (P. sicula) and the European leaf-toed gecko (Euleptes europaea), while small mammals such as the house mouse (Mus musculus) are widespread. The island lies along key migratory routes, attracting raptors and numerous passerine species during seasonal movements. Coastal and marine habitats are particularly rich, with seabird colonies including yellow-legged gulls (Larus michahellis) and European shags (P. aristotelis). The surrounding waters harbor diverse fish populations, occasional dolphins, and sea turtles, reflecting the ecological significance of the island's marine ecosystems (https://www.parconazionalepantelleria.it/).
3 Archaeological framework
The low altitude (and consequently limited visibility), the near-total absence of freshwater springs, the lack of valuable geological resources such as obsidian, and the considerable distance from the north-western coast of Sicily make Ustica relatively marginal in the exchange dynamics of the southern Tyrrhenian Sea during prehistory. However, permanent human occupation began at least by the end of the Middle Neolithic period (first half the 5th millennium BCE), as evidenced by recent excavations at Piano dei Cardoni (Speciale et al., 2020, 2021a,b, 2022, 2024b). These findings suggest the arrival of Neolithic groups in certain areas of the island, coinciding with a general demographic increase in Sicily (Mannino, 1998; Speciale, 2024; Speciale et al., 2024a). These groups may have used the island as a stepping stone in the obsidian trade network and the exchanges between the Aeolian islands and western Sicily (Speciale et al., 2021b; Forgia et al., 2024). According to previous research, the island during the Middle Neolithic period, permanently occupied for the first time, was invested by a Neolithic economic subsistence system, with the exploitation of domestic and wild animals and plants and the processing of crops by local stone tools (Mantia et al., 2021; Speciale et al., 2023). Despite the presence of imported obsidian and chert, all the rest of the resources were probably local. The island was as a matter of facts most probably independent in terms of staple food and husbandry, with local production of pottery as well (Speciale et al., 2021a,b, 2023; Magrı̀ et al., 2021; Prillo et al., 2024, 2025).
Following the distribution of pottery and obsidian, the Neolithic site of Piano dei Cardoni occupied an area of approximately 2 ha; the AMS dates (Speciale et al., 2024b) confirm the chronology hypothesized through the material culture (4,800–4,300 cal BCE). The megalithic funerary complex was lying in the southern coast of the island (“Mezzogiorno”), not far from the modern town of Ustica. It was probably covered by an earthen mound; it was excavated from 2019 to 2022 and revealed the death customs of this insular community, recording a long process of manipulation of the human bones and some rituals connected to agricultural activities and the sun (Speciale et al., 2021a, 2023; Mantia et al., 2021; Magrı̀ et al., 2021; Montana et al., in review) (Figure 4). The sampled stratigraphic units, here presented, belong to the phases of use of the megalithic structure, in particular the filling of the pit, the group of bones around and on the slab stone, and the supposed earthen mound above them. Being very close to the surface, the data coming from potentially contaminated layers were not included.

Figure 4. View of the funerary structure of Piano dei Cardoni (Ustica) before the removal of the big covering slab stone.
The prehistoric settlement of Mursia (Pantelleria) and its associated monumental necropolis, “I Sesi,” are among the most significant and best-preserved archaeological complexes in the central Mediterranean. The settlement, covering approximately 1 hectare, is distinguished by its monumental defensive wall, and the large burial mounds offer clear evidence of a complex society that warrants special attention in archaeological research. The life span of this site is of about three centuries and it is composed by 3 macro-phases, going along 300–350 years from half 18th c. to half 15th c. BCE (Cattani and Peinetti, 2023). The sediment samples analyzed from the Mursia site come from the sectors B and E, each indicating distinct residential features from different phases in the use of the village (Cattani, 2015; Debandi and Magrı̀, 2021). Sector B is identified as a residential area, with huts arranged in parallel rows - evidence of an intentional and organized layout of the settlement. The sector is located on a rocky promontory at the northern edge of the lava flow, a location likely chosen for its strategic position and proximity to the coast.
The site was chosen for its favorable geographical features: its dominant position over the Mursia plain, access to two potential landing points, and the presence of water from the coastal buvire. The promontory's topography, partially spared by the lava flows, was ideal for establishing the village. It is likely that the entire area, enclosed by the defensive wall, was intended for the settlement from the very beginning. The samples from dwelling B14 correspond to the sixth occupational phase, which aligns with the first and second macro-phases of the settlement (Debandi, 2015). These samples were collected from the fireside stone structure and its ash layers, while the samples of the dwelling E1 and E2 fall within the third macro-phase of the village, which is the last phase before it was abandoned. From this area, samples from the pottery and clay fireside with andirons were collected, but also from the outside space between two huts (Figure 5).

Figure 5. Mursia (Pantelleria, TP), E Area: zenithal photograph with huts E1 (right), E2 (middle) (Debandi et al., 2019).
The village of Mursia shows how Pantelleria was in the middle of exchange networks for precious materials and objects that connected the island to the Eastern Mediterranean, the northern coast of Africa and even Northern Europe (Cattani et al., 2024). So far, no clear indicators of imported staple sources have been found, despite the analyses highlighted the presence of imported pottery, and the isotopic signal of some faunal remains could indicate a potential provenance from outside the island (Dawson et al., 2024; Fiori et al., 2024).
4 Materials and methods
4.1 Piano dei Cardoni, Ustica
Some results are already published in Mantia et al. (2021), Speciale et al. (2021a, 2023), Prillo et al. (2024, 2025). The first 14C dates, published in Speciale et al. (2024b), are obtained on faunal remains and macrobotanical remains. The soils were floated during the excavation of Area 2 (years 2019–2020) of the site of Piano dei Cardoni, when samples were systematically collected in accordance with standard procedure (Pearsall, 2015) of around 20 liters per layer. Some of the samples were contaminated by modern seeds or charcoal and therefore they were not considered in this paper. In 2022, the sampling amount was increased, and the team used a hand-pump flotation machine, allowing to process a total of more than 1,250 liters of sediment. Unfortunately, the density of macroremains is generally very low, probably due to the nature of the site (being a funerary structure, the presence of botanical materials is very limited) (Supplementary material, Table 1). Furthermore, most of the plant material proved to be badly preserved and/or with a limited size in case of wood charcoal: the specimens mostly range from 0.5 to 2 mm in width, and they are not always well visible on the three cuts. Remains were selected by hand with a magnifying lens (4×) and then examined with a metallographic trinocular microscope Optika B-383MET (up to 500×). Carpological remains were observed with a binocular Euromex (up to 50×).
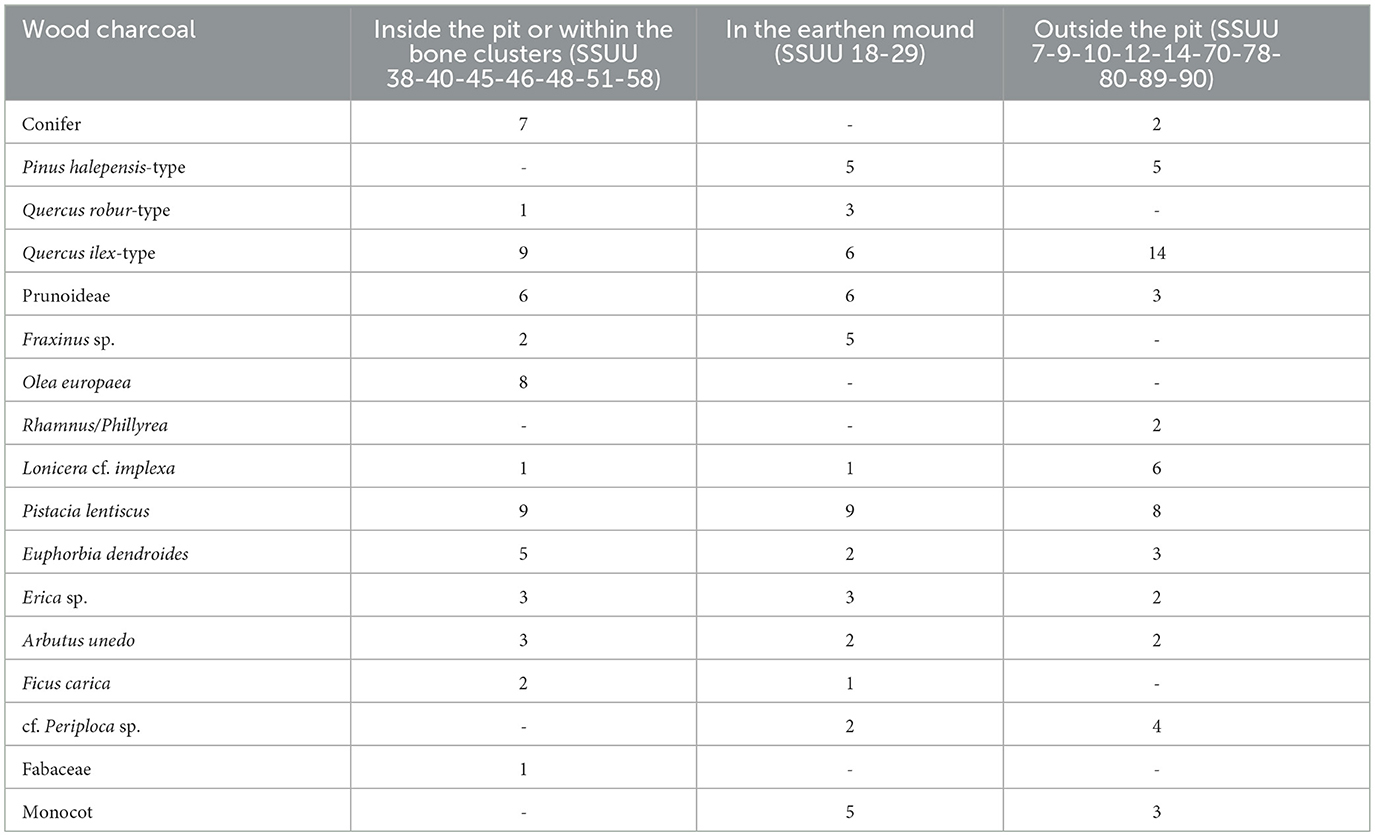
Table 1. Anthracological remains from Piano dei Cardoni (Ustica).
For the identification of the woody species, reference atlases (Cambini, 1967; Schweingruber, 1990), scientific literature (i.e., Asouti et al., 2015), and online tools—InsideWood, https://insidewood.lib.ncsu.edu and Microscopic Wood Anatomy, http://www.woodanatomy.ch/ -were used. For the identification of fruits and seeds, reference atlases were used (Neef et al., 2012; Sabato and Peña-Chocarro, 2021). In addition, the analysis was supported by direct comparison with the reference collection at the Unit of Archaeobotany, IPHES-CERCA when necessary. Due to their very low density in the soils, all the wood charcoal and seeds were observed. The nomenclature of all the plant species quoted in the text follows Pignatti et al. (2017–2019).
Faunal remains were quantified by determining the number of identified specimens (NISP) for each taxon, while the minimum number of individuals (MNI) was calculated only for the most common ones following Bökönyi (1970) for most species and Girod (2015) for molluscs. Several osteological atlases were used for the taxonomic identification: Schmid (2022) for mammals, Cohen and Serjeanston (1996) for the avifauna, Giannuzzi-Savelli et al. (1999) for marine molluscs, while fish bones identification was conducted using online resources (Archaeological Fish Resource, University of Nottingham, n.d.; Tercerie et al., 2022).
To identify some of the bone remains, it was necessary to use a reference collection, therefore some materials were moved to the Laboratory of ArchaeoZoology of the University of Salento (LAZUS, Lecce, Italy). The scientific nomenclature of domestic animals refers to Gentry et al. (2004). The distinction between sheep and goat was attempted using the criteria described in Boessneck et al. (1964), Boessneck (1969), Payne (1985), Halstead et al. (2002), and Zeder and Lapham (2010).
Data from the epiphyseal fusion of long bones were registered using the works of Silver (1969) for cattle, Bullock and Rackham (1982) for ovicaprines, and Bull and Payne (1982) for pigs. Wear teeth stages were recorded following Grant (1982) for cattle, Grant (1982) and Bull and Payne (1982) for pigs, and Payne (1973) for ovicaprines.
Ovicaprines' size and morphology were investigated using log standard index (LSI) values (Meadow, 1999). LSI values for post-cranial bones were calculated using established caprine standards (Davis, 1996), while also examining changes in sex ratio (Davis, 2000).
4.2 Mursia, Pantelleria
The 14C dates of Mursia were obtained on some hut contexts from the same phases of the huts here presented (Cattani, 2015), while new 14C analyses to directly date the samples of this paper are ongoing. The bioarchaeological data of Mursia come from different excavation contexts, and they have a partial sampling, because the climatic condition of the island, very arid, forces to preserve freshwater as much as possible. Therefore, the flotation of the soil samples has been limited for a long time, until the archaeologists started an alternative protocol with the use of sea water during the last excavation seasons. This hand flotation was made with two sieves with a mesh of 2 and 0.5 mm, and only the floating residue was filtered with the smaller one. This kind of flotation was followed by a final rinse with freshwater within the same day, exclusively for the small residue. This last action is essential to avoid the crystallization of the sea salt and the potential degradation of the remains.
Every soil sample was around 5 liters, and the bioarchaeological remains were compared by their volume. This method allowed to overcome the representing issue between the different categories, as well as the high fragmentation of the bioarchaeological remains (Supplementary material, Figure 1).
Furthermore, this analysis involved a double sifting action by a zooarchaeologist and an archaeobotanist in two separate steps, and the volume of the remains was measured by using three types of graduated containers of 0.5 ml, 2 ml, and 16 ml.
The preservation conditions for the archaeobotanical remains of Mursia are quite good, with low levels of vitrification and cracklings for the wood charcoal, despite the size is always very limited.
The differences in the distribution and typology of bioarchaeological remains help to clarify the use of the spaces within the various housing units, for example: the analysis of the floor layers, the dumping waste area, the area with cooking pottery.
The soil samples discussed in this contribution derive from sectors B and E of the Mursia site, each reflecting distinct habitation features associated to different phases in the life of the village (Cattani, 2015; Debandi and Magrı̀, 2021). The samples from hut B14 pertain to the sixth habitation phase, which corresponds to the first and second macro-phases of the settlement (Debandi, 2015). The first two samples (B1001; B1002) were taken from the upper and lower fills of the lithic cist SU 1097, a structure that supported the hut's hearth. The remaining samples from B14 (B1003; B1004) originate from an ash disposal area (SU 1119), also associated to this hearth's area. Samples from huts E1 and E2 correspond to the third macro-phase of the village and relate to the final phases of the site's occupation. The first two samples from structure E1 (E1501; E1504) were recovered from SU 2011, interpreted as the occupation layer for the activities inside the hut. Along the eastern wall of this structure, a concentration of ceramic items connected to food preparation was identified, including two andirons and a cooking bowl. The other samples were collected from the fills of a bowl-dish (E1507, Rep.E15036), the interior of a small olla (E1509, Rep.E15024), and the sediment surrounding the andiron (Rep.E15016; Debandi and Magrı̀, 2021). The samples from structure E2 relate to a beaten floor surface (SU 2067) and the soil surrounding the andiron (Rep.E16028; Debandi and Magrı̀, 2021), as well as a sample from the outdoor space between huts E2 and E3 (SU 1201).
The sorting of the carpological remains was conducted under a binocular stereoscopic microscope (10×). The carpological remains were identified by comparison with atlases (Cappers and Neef, 2016; Neef et al., 2012) and the reference collection of the Bioarchaeological Research Center ArcheoLaBio (Department of History and Cultures of the University of Bologna). The identifications were made using Leika and Konus stereomicroscopes (magnification 2×-7×). Pignatti et al. (2017–2019), the databases Plants of the World Online (https://powo.science.kew.org/) and The World Checklist of Vascular Plants (https://www.gbif.org/) were used for the scientific nomenclature. All the carpological remains found in the samples were examined.
The protocols for analyzing wood charcoal and faunal remains followed those described in Section 4.1.
4.3 Compared framework
The prehistoric human ecosystems of Ustica and Pantelleria are compared. The two islands show marked differences in terms of size, distance from the coast, and ecological features. Moreover, the two archaeological sites differ in chronology and function. Therefore, they offer a valuable opportunity for comparison to assess potential adaptations to the insular environment. The combined analysis of carpological, anthracological, and faunal remains provides a complete picture of how past communities interacted with their environment and what choices they made for their diets. Carpological data contributes understanding which plants were cultivated or gathered, while charcoal remains reflect woodland use and local vegetation. Faunal evidence, in turn, provides insight into subsistence practices and animal management.
This multidisciplinary approach is mostly due to the fact that the management of animals and vegetation is inextricably linked and that, as mentioned above, in general the impact of animal grazing is particularly dangerous on limited geographic areas like small islands (Anderson, 2002). Small Mediterranean islands likely required specific and deliberate adaptation strategies by the prehistoric human groups that settled them (Ramis, 2014).
Over time, as human groups became bigger, anthropogenic pressure increased and environmental conditions changed, these adaptive choices may have become more specialized, reflecting both ecological constraints and the need to manage limited resources sustainably. Signals of human adaptation include shifts in species composition toward more resilient taxa, evidence of landscape management (e.g., agroforestry, terraced landscapes, Bevan and Conolly, 2011), changes in herd composition (Ramis, 2017), and indicators of long-term resource sustainability (e.g., absence of overgrazing-related taxa) (Caballero et al., 2009).
5 Results
5.1 Piano dei Cardoni, Ustica
5.1.1 Anthracological data
The first analysis of the anthracological data from Piano dei Cardoni is published in Speciale et al. (2023). The new data include the stratigraphic units excavated in 2020 and 2022 (Table 1, Supplementary Table S1). The first results, very limited in terms of number of specimens, were enriched with new species not detected before. Here the new charcoal fragments analyzed are presented, mostly of very small size. That is the main reason why 25% of the record could not be identified, despite a very low rate of vitrification and cracks. Taxa are fairly evenly distributed all over the contexts, with the notable exception of olive trees, which are present exclusively inside the burial pit and here identified for the first time (Figure 6.1). Most of the record is represented by local shrubs like lentisk (P. lentiscus), heather (Erica sp.)—Figure 6.2—and spurge (E. dendroides), associated with an oak-pine woodland (Aleppo pine and holm oak), that includes ash trees (Fraxinus sp.) and wild plums (Prunoideae), confirming the mesophilous association as previously reconstructed (Speciale et al., 2023). Noteworthy also the likely first recovery of periploca (cf. Periploca sp.) and deciduous oaks (Quercus robur-type), today absent from the flora of Ustica, and of the fig tree (Ficus carica), maybe imported from the mainland (Figure 6.3).

Figure 6. Anthracological and carpological remains from Piano dei Cardoni (1–7) and Mursia (8–14). (6.1) Transversal section of Olea europaea charcoal; (6.2) Transversal section of Erica arborea-type charcoal; (6.3) Transversal section of Ficus carica charcoal; (6.4) Hordeum vulgare subsp. vulgare caryopsis; (6.5) Chenopodiastrum murale seed; (6.6) Heliotropium europaeaum fruitlet; (6.7) Althea hirsuta/Malva setigera fruitlet; (6.8) Transversal section of Erica arborea-type charcoal; (6.9) Radial section of Pinus cf. halepensis-type; (6.10) Radial section of Pinus cf. pinaster-type; (6.11) Triticum sp. glume; (6.12) Ficus carica achene; (6.13) Portulaca oleracea seed; (6.14) Chenopodium sp. fruit.
5.1.2 Carpological data
The carpological assemblage from Piano dei Cardoni is quite limited, due to the nature of the site. A total of 353 specimens has been identified, that adds to the 13 previously recovered (Speciale et al., 2023) (Table 2, Supplementary Table S1). 14% of charred fragments were unidentified (including lumps and carpological tissues with a volume of more than 1 mm3). The preservation conditions are in general not very good, especially for the remains recovered from the earthen mound. The only cereals identified are hulled barley (Hordeum vulgare subsp. vulgare, Figure 6.4); other Poaceae were found and some of them identified to the genus level (Poa, Lolium) and they are spread in all the contexts. Pulses are limited to the family level and absent from the earthen mound, while goosefoot (cf. Chenopodiastrum murale, Figure 6.5) is definitely the most represented taxon in all the contexts in terms of number of specimens, followed by heliotrope (Heliotropium europaeum, Figure 6.6). Some plants have a very low representation, like hairy mallow (Althaea hirsuta, Figure 6.7), black nightshade (Solanum nigrum) and black bindweed (Fallopia convolvulus). Recovered, in almost every context, parts of lentisk fruits and likely acorns. In general, the lowest diversity is within the pit, where species are mostly limited to Poaceae, Fabaceae, C. murale and H. europaeum.
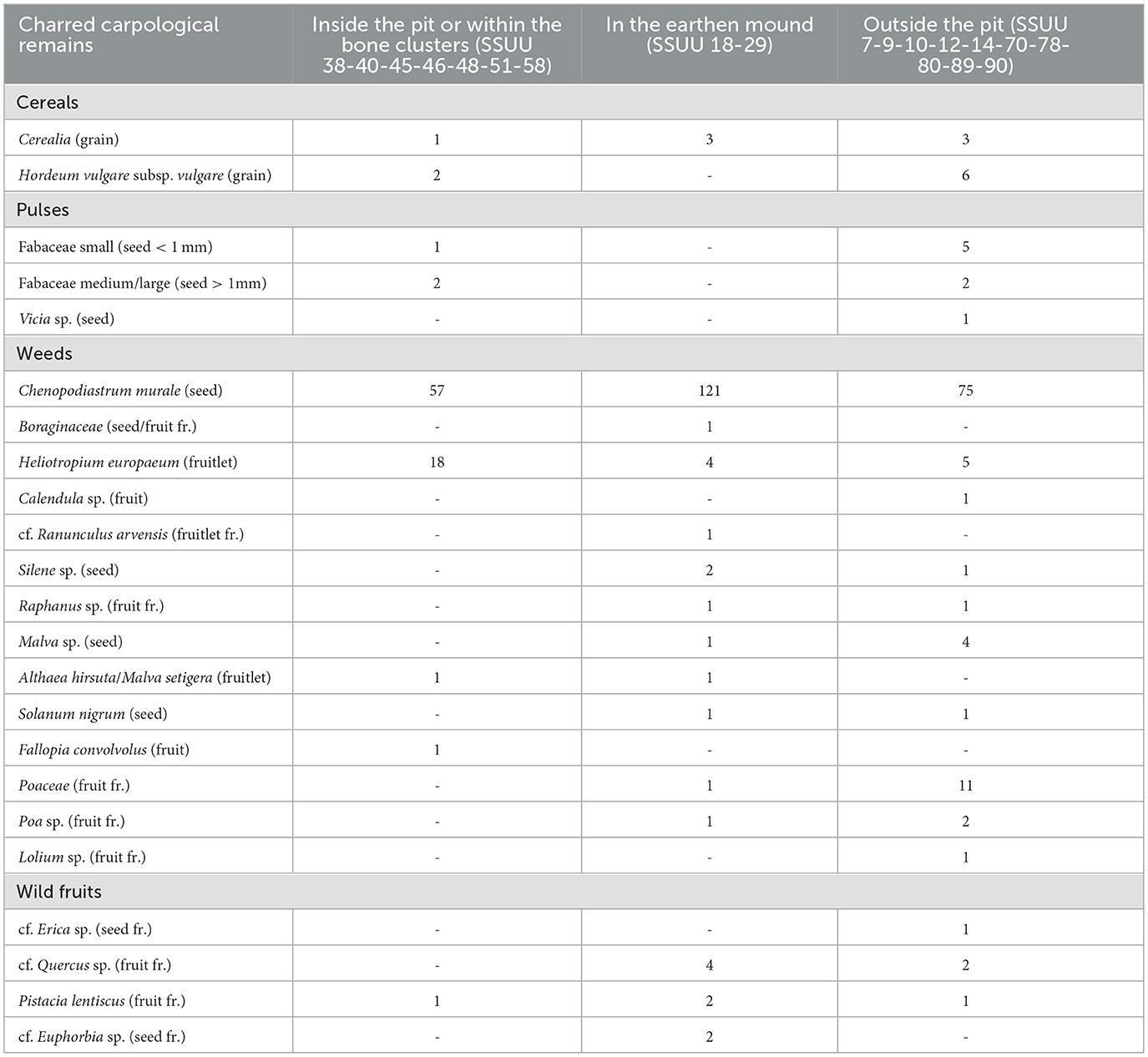
Table 2. Carpological remains from Piano dei Cardoni (Ustica).
5.1.3 Zooarchaeological data
The data presented here are an updated version of those published in previous studies (Prillo et al., 2024, 2025), now also including materials found during the 2022 excavation campaign. Furthermore, zooarchaeological data are presented by considering each specific contexts (Tables 3, 4), as done with carpological and anthracological data, thereby excluding faunal remains from unreliable or less significant stratigraphic units.
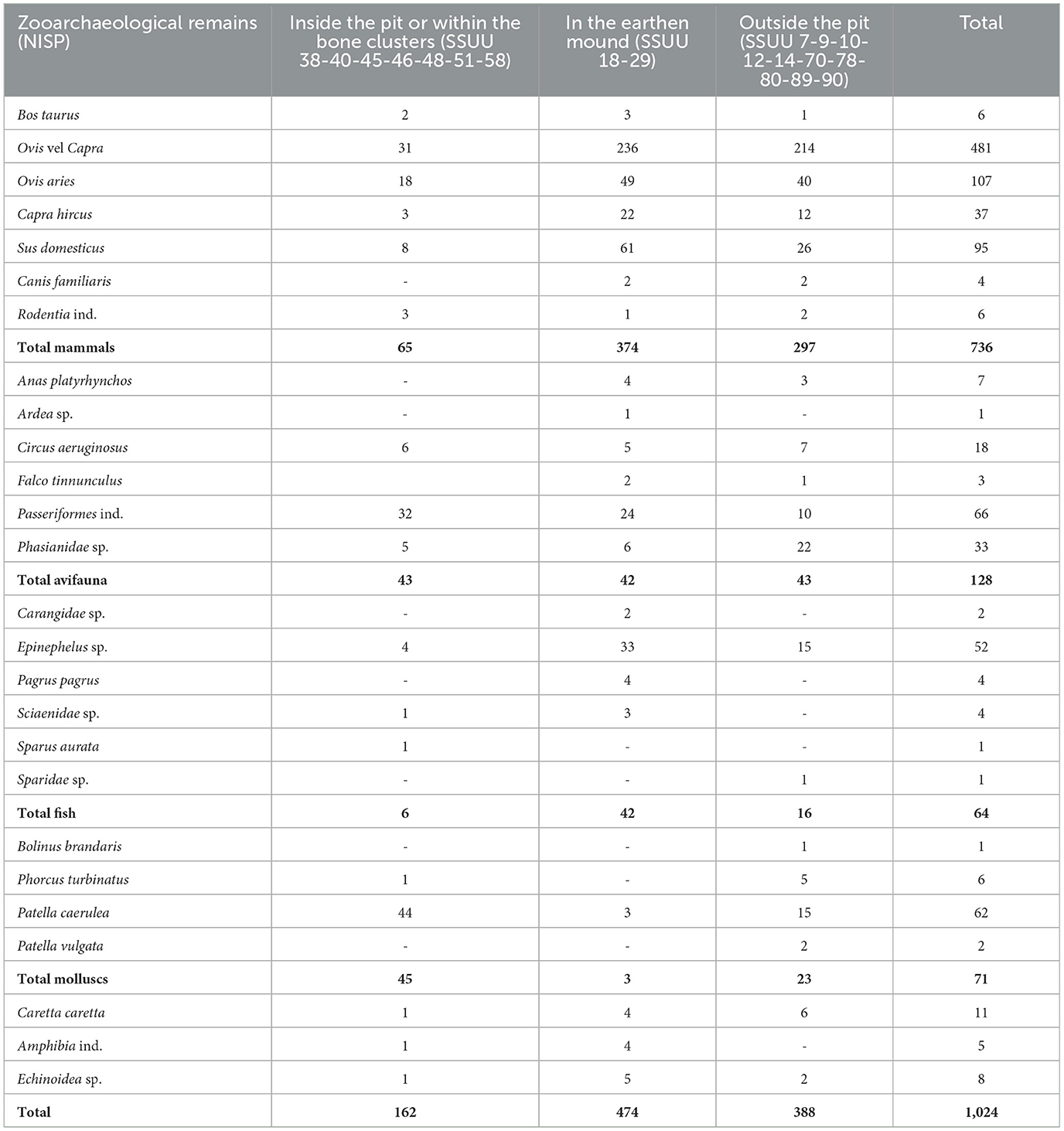
Table 3. Zooarchaeological remains from Piano dei Cardoni (Ustica) quantified by their NISP.
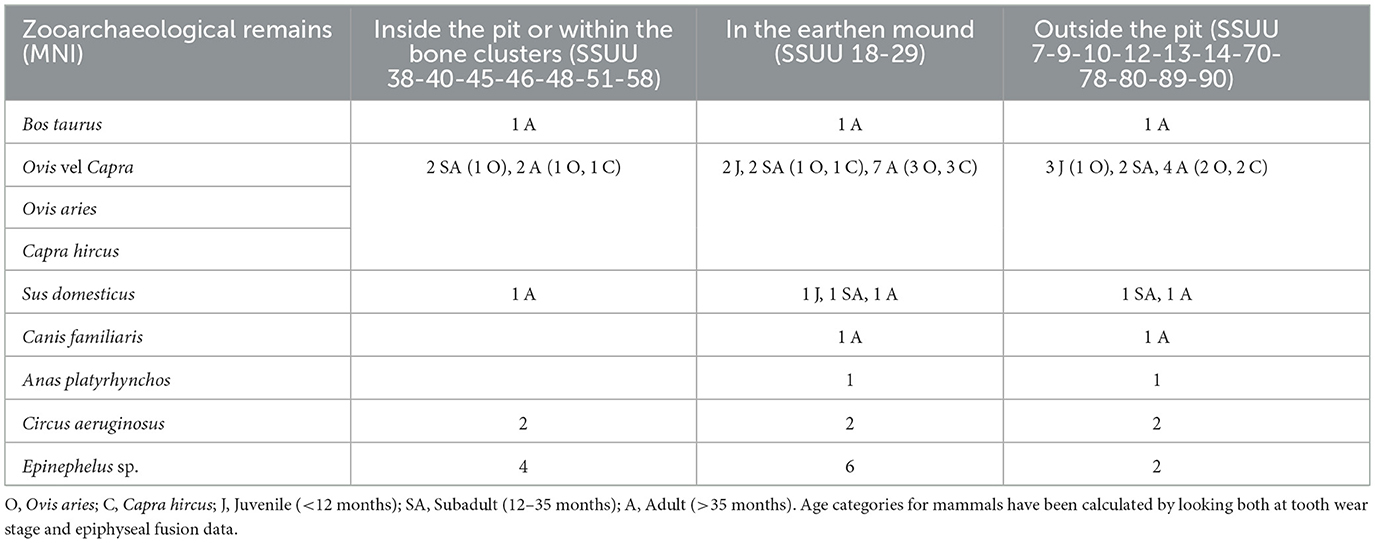
Table 4. Minimum number of individuals (MNI) of the most frequent animal species identified from Piano dei Cardoni (Ustica).
The faunal sample is mostly composed of mammal bones, with a clear dominance of ovicaprines. Sheep (Ovis aries) consistently outnumber goats (Capra hircus), although the bones classified within the general category of ovicaprines (Ovis vel Capra) are the most abundant. Pig (Sus domesticus) remains are also present in modest quantities, while cattle (Bos taurus) remains are nearly absent. Mortality profiles for domestic species indicate a prevalence of mature individuals, suggesting an interest in maximizing meat yield by allowing animals to reach their optimal size. Nevertheless, the presence of some piglets and young ovicaprines indicates that younger animals were also occasionally consumed.
No wild mammal species were identified in the archaeological contexts considered in this study, although the presence of rare hare (Lepus sp.) and fox (Vulpes vulpes) remains has been recovered in most superficial layers (Prillo et al., 2024, 2025). Hunting focused exclusively on avian species, which were intensively exploited. However, the most frequently hunted birds were small- to medium-sized species, namely marsh harrier (Circus aeruginosus), common kestrel (Falco tinnunculus) and members of the Phasianidae family and the Passeriformes order. Together with the presence of mallard (Anas platyrhynchos) and heroon (Ardea sp.), they suggest the presence of water bodies on the island during the Neolithic period.
Marine resources also played a significant role in the diet of Ustica's Neolithic inhabitants, as confirmed by the presence of numerous fish remains—mostly groupers (Epinephelus sp.) and mollusc shells—primarily Patellidae. Due to their fragmented state, no biometric data are available for the fish species; however, the presence of groupers and other fish families (Carangidae, Scianidae, Sparidae) suggest the exploitation of medium- to large-sized prey. Finally, other exploited marine species included sea turtles (Caretta caretta) and sea urchins (Echinoidea sp.).
Lastly, preliminary biometric data suggest that ovicaprines were smaller in size compared to those from other contemporary contexts in Sicily and southern Italy. This issue will be explored in detail to determine whether insular factors contributed to this size variation.
5.2 Mursia, Pantelleria
5.2.1 Anthracological data
A total of 300 wood charcoal specimens were recovered and analyzed (Figure 7, Supplementary Table S2). Approximately 10% of the wood charcoal fragments were not identified, mainly due to the small size. The preservation rate is quite good, with minimal evidence of distortion or fungal attacks. In the B14 hut assemblage, tree heather (E. arborea-type) dominates (almost 70%, Figure 6.8). Other shrubby species are present in lower proportions, including strawberry tree (A. unedo), alatern/mock privet (Rhamnus/Phillyrea), rockrose (Cistus sp.), lentisk and terebinth (Pistacia cf. terebinthus). Fragments of Q. ilex-type are very scarce.
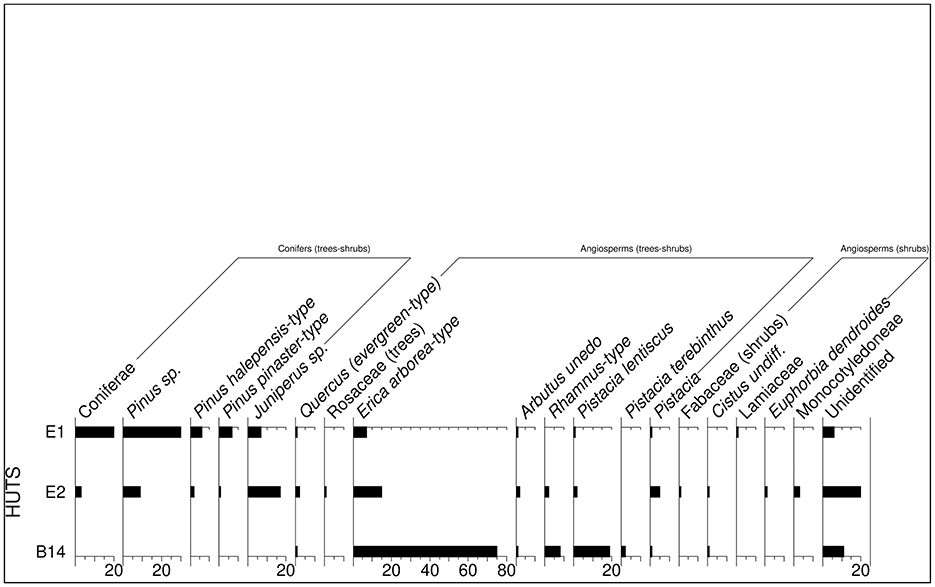
Figure 7. Percentage and table with the number of specimens of wood charcoal from huts B14, E1 and E2.
Wood gathering and usage appear to shift in the following phase, represented by huts E1 and E2. Conifers become the main exploited arboreal species (75% in E1, 50% in E2). Pines (Pinus sp., P. halepensis-type and maritime pine, Pinus pinaster-type, Figures 6.9, 6.10) are dominating E1 (50% of the total) and Juniperus sp. is bit more represented in E2 (almost 30% of the total). In E1, the remaining 25% includes shrubby species, along with a small proportion (2%) of Q. ilex-type. In E2, beyond a 3% of Q. ilex-type, cf. E. dendroides, Cistus sp., Rhamnus/Phillyrea, Fabaceae and monocots are also present.
5.2.2 Carpological data
The carpological analysis shows a high number of charred seeds and fruits in the dwelling B14, E1 and E2, whereas there are very few mineralized remains (Table 5, Supplementary Table S2).
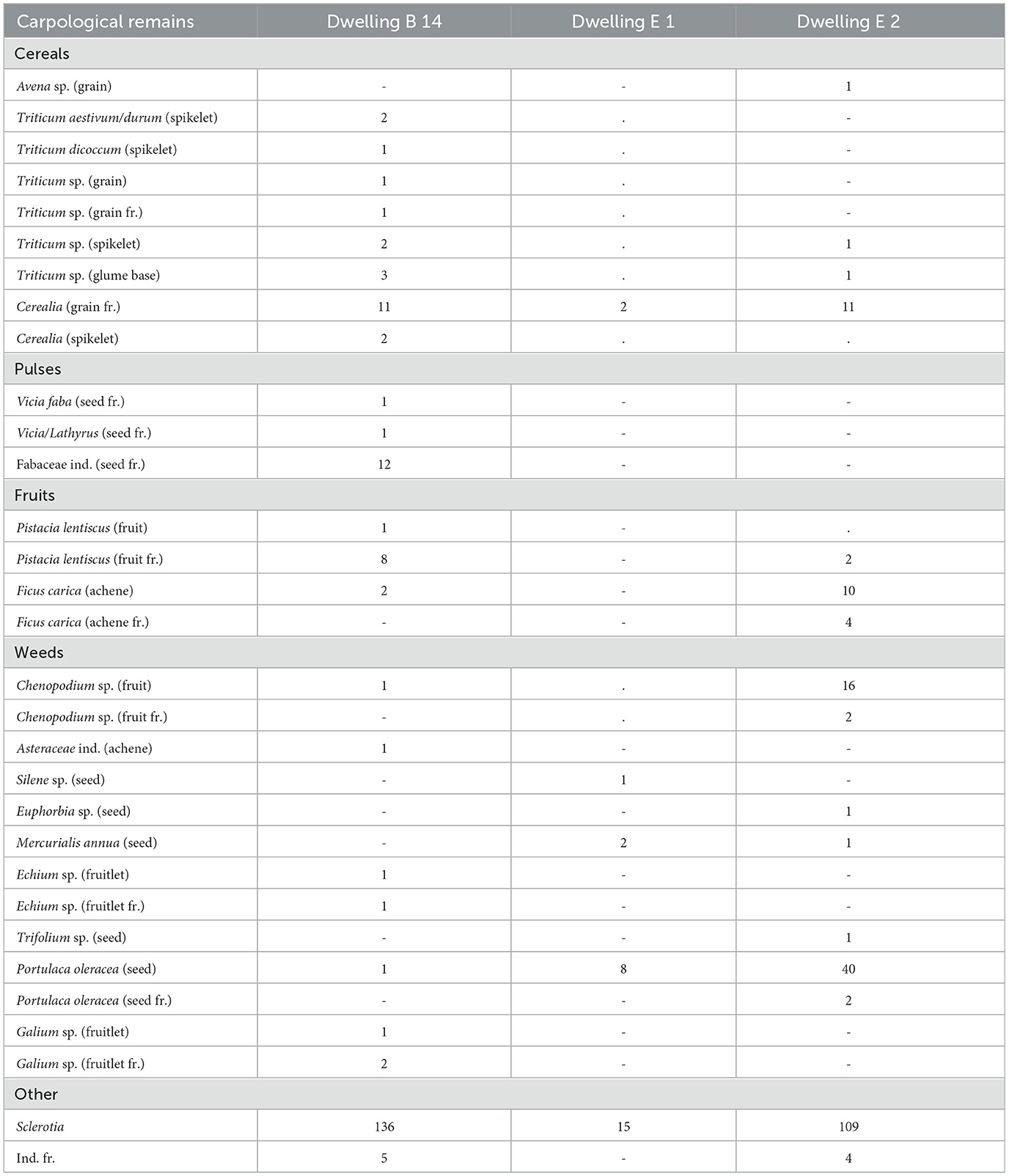
Table 5. Carpological remains from Mursia.
The soil samples revealed the presence of domestic plants, including free-threshing wheat (Triticum aestivum/durum), emmer wheat (Triticum dicoccum) (Figure 6.11), faba beans (Vicia faba) and other pulses (Vicia/Lathyrus). Notably, concentrations of emmer wheat were found specifically within the fireside stone structure of the dwelling B14, while a single oat grain (Avena sp.) was recovered from the outdoor space between dwellings E1 and E2. In these same samples, wild plants were identified as well: fig (Figure 6.12) and lentisk were present. F. carica was more common in the samples from the third macro-phase, while P. lentiscus was abundant in the first/second macro-phases of the site. In addition, various herbaceous species found in the samples could be used for many purposes. For example, purslane (Portulaca oleracea) (Figure 6.13), edible plant, and goosefoot (Chenopodium sp.) (Figure 6.14), potentially edible depending on the species, were mainly found in the samples of dwelling E2. Annual mercury (Mercurialis annua) and some species of bedstraw (Galium sp.) can be used to dye the textiles (Guarrera, 2006; Prigioniero et al., 2020); the first one was attested in the third macro-phase and the second one only in the first/second macro-phases. Other Mediterranean plants were identified in the site, such as the viper's bugloss (Echium sp.), the spurge, the catchfly (Silene sp.) and the clover (Trifolium sp.), which were maybe used by the inhabitants of the village for a pharmaceutic purpose (e.g., Sheydaei et al., 2025; Kolodziejczyk-Czepas, 2016).
In the same soil samples, a high number of sclerotia were found (fungal hypha masses), which depend on the organic properties of the layers. As expected, their number is not regular in the samples, but there was a high concentration inside the fireside stone structure on the dwelling B14.
5.2.3 Zooarchaeological data
The zooarchaeological analysis of the Mursia site primarily focuses on the first and second macro-phases of the village, based on the faunal remains recovered from dwelling B14 (Figure 8). In contrast, data from the third macro-phase are limited and derive solely from the analysis of soil samples associated with dwellings E1 and E2, as presented here. The samples from B14, second macrophase, includes 38,051 osteological remains. Of these, approximately 23% were taxonomically identified and anatomically described. The data show a subsistence economy centered on animal husbandry, with a predominance of goat (C. hircus) and sheep (O. aries), followed by pig (S. domesticus) and cattle (B. taurus). On this island, hunting on wild big mammals was not practiced because wild herbivores were absent. However, the exploitation of marine resources and the capture of wild birds played a significant role. As expected, fishing was particularly important and focused on medium- to large- sized species caught near the cliff, including dusky groupers (Epinephelus marginatus), wrasses (Labrus sp.), common seabream (Pagrus pagrus) and parrotfish (Sparisoma cretense). Additionally, the inhabitants of the village collected various mollusc families (Patellidae, Trochidae, Muricidae), sea urchin (Echinoidea), crabs (Eriphia sp.), as well as cuttlefish (Sepia sp.), sea turtle (Caretta caretta) and even monk seal (Monachus monachus). Migratory birds also provided a valuable seasonal meat resource during migration, as evidenced by remains of wild goose—Anser sp.—and yelkouan shearwater—Puffinus yelkouan.
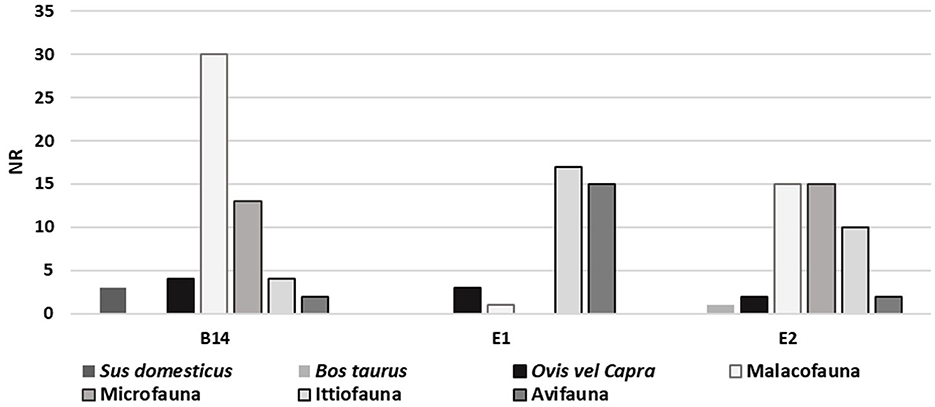
Figure 8. Faunal remains from Mursia. NR = NMI.
In contrast, preliminary zooarchaeological data from the third macro-phase indicate an increase in cattle and a decrease in pigs within the livestock assemblage. This trend appears to be supported by previous studies (Wilkens, 1987), although additional data are needed to confirm this hypothesis (Fiori, 2025).
6 Discussion
6.1 Ustica
6.1.1. Wood resources and fruit trees
The assemblage is dominated by shrubs such as lentisk, heather, and euphorbia, associated with a mixed oak-pine woodland, with the presence of ash trees and wild plums. Additionally, periploca, deciduous oaks, and fig were also identified for the first time on the island. One of the most relevant findings is the presence of olive trees exclusively inside the pit, where it was recorded for the first time in the site. The high biodiversity observed in such a small record suggests an absence of targeted selection, with the gathering activity likely encompassing species from various areas across the island.
This broad-spectrum exploitation may reflect several factors: 1. the island's small size, which makes it easily traversable within a day; 2. its naturally low biodiversity, influenced by the limited area and the low altitude and 3. the low number of different habitats due to environmental homogeneity. Additionally, 4. the apparent lack of preference for any particular tree species may have led to a more balanced exploitation of resources, distributing human impact evenly across shrubland and woodland areas, and thereby reducing the risks of overexploitation.
Both evergreen and deciduous oaks were present on the island, with the latter identified here for the first time. Deciduous oaks are absent from the island's current flora. Their presence on a small and relatively flat island like Ustica serves as a clear indicator of rather humid conditions—consistent with broader Mediterranean climatic trends during the mid-5th millennium BCE (Speciale et al., 2024a). Such conditions likely contributed to a favorable context for human habitation on the island as well. The new data also confirms the presence of holm oaks, ash trees and Aleppo pines in the island's natural vegetation, in line with earlier findings (Speciale et al., 2023). On the other hand, the potential presence of periploca (probably Periploca angustifolia, also occurring in the Egadi Islands, Pantelleria and Pelagian islands and in the Maltese Archipelago) is recorded for the first time for the Tyrrhenian insular context. This summer-deciduous taxon is adapted to extremely arid conditions; it is a shrub species with several uses in phytopharmacy (Huang et al., 2019).
The fig tree is here recorded for the first time. On mainland Sicily, F. carica increases in most of the lacustrine pollen sequences since mid-6th millennium BCE (e.g., Calò et al., 2012). Its spread likely parallels that of domestic cereals, even if the only presence in an archaeobotanical record as fruit so far is from mid-6th millennium in Grotta dell'Uzzo (Speciale, 2024). It remains uncertain whether the fig tree was naturally present on the island prior to human arrival; however, its introduction by humans during the Neolithic appears more plausible. The analysis of the wood cannot help in disentangling the exploitation of its fruits, and so far, no fig achenes have been found in the carpological record; its exploitation is then still an open question mark for neolithic Ustica. The framework is the opposite of Mursia, where fig achenes are present but fig tree charcoal is absent.
Olive trees are present both in the Sicilian lacustrine pollen record and in the Mesolithic layers of Grotta dell'Uzzo, where it has been identified as wild olive (Speciale, 2024), and exploited up to the Madonie mountains at least since the Bronze Age (Forgia and Oll?, 2023). Its presence in the Neolithic phase of Ustica was unsure from the previous record and it is now confirmed. It is believed that the cultivated olive tree (O. europaea L. var. europaea) originated from the domestication of its wild form (O. europaea var. sylvestris (Mill.) Lehr), which grows naturally throughout the Mediterranean basin. The distribution of the olive tree mostly aligns with that of its wild ancestor, one of the most representative taxon of the Mediterranean sclerophyllous vegetation (Carrión et al., 2010; Gianguzzi and Bazan, 2020).
Undoubtedly, from the Bronze Age onward, the marked increase in olive remains is closely linked to the production of olive oil (Caracuta, 2020; Palli et al., 2025; Schicchi et al., 2021). This process is also evident in the Middle Bronze Age record from Ustica (Speciale et al., 2023), and may indicate localized domestication dynamics, particularly in Southern Italy (D'Auria et al., 2017). Exploitation of olive trees on small Mediterranean islands is frequent after the fourth millennium BCE. On another small island, Kephalonia, from the Chalcolithic to the Middle Bronze period, the rising presence of Arbutus sp.—indicative of increasingly open landscapes, correlates with the increased presence of O. europaea, prominent also in the diagram of the Heraion of Samos (Ntinou and Stratouli, 2012; Mavromati, 2022).
In other geographic areas, the introduction of tree crops to islands suggests the gradual evolution of forest management toward agro-forestry (Dotte-Sarout, 2017). Arboriculture became a significant component of production systems of the Marquesas Islands (Huebert and Allen, 2016, 2020), the Society Islands (Lepofsky, 1994), and Tikopia (Kirch and Yen, 1982). Agroforestry represents usually the latest step in the development process of management of the landscape, commonly starting with the clearance of vegetation on the coast (Huebert and Allen, 2016). On a small Mediterranean island like Ustica, though, it seems that the occupation happened all in the same phase and the exploitation of wood resources was somehow managed from the very beginning.
Finally, O. europaea charcoal in Piano dei Cardoni was recovered only inside the funerary pit. The presence of olive trees in funerary contexts across other areas of Southern Italy is particularly significant. The earliest evidence of olive stones, dating to the Middle Neolithic, was recovered from a funerary context in Carpignano Salentino (Lecce) (Ingravallo and Tiberi, 2008). The use of olive wood persisted into the Copper Age, when it continued to hold ritual value, as evidenced by its employment in funerary practices at the stone mound in Macchia Don Cesare (Salve, Lecce) (Aprile and Fiorentino, 2018).
6.1.2. Crops and weeds
Carpological remains recovered from Piano dei Cardoni are relatively scarce, likely due to unfavorable preservation conditions. Despite this, the increased volume of floated soil samples significantly enhanced the quantity of the recovered remains in the latest campaign.
Barley is the predominant cereal, while other Poaceae, such as Poa and Lolium, are also present and evenly distributed across the different contexts. Their presence may indicate human use or their role as fodder. Many of the identified weeds could have grown in arable fields, entering the site through crop processing or other agricultural activities. Some potentially edible plants are present, such as the leaves of goosefoot (C. murale), that can also be sown into fields for grazing animals (Behre, 2008). Whether any of these plants were deliberately collected for medicine or food, for example, it is difficult to determine from the generally small quantities recovered, as well as poor preservation restricting identifications.
The diversity of pulses at Piano dei Cardoni is low, with no remains recovered from the earthen mound. Small-seeded legumes may have grown in arable fields, including stubble, or in pastures. Following C. murale, H. europaeum is the most frequent taxon. While its presence might be associated with agricultural activities, the relatively high quantities suggest it may have had a specific use– as a matter of fact, it is traditionally used as a medicinal plant for its analgesic properties. A few fragments of acorns and lentisk nuts were found in most contexts, indicating the exploitation of the local wild resources. The pit context exhibits the lowest taxonomic diversity, dominated by Poaceae, Fabaceae, C. murale, and H. europaeum.
Among the cereals, barley stands out for its drought resistance because the grains fill before the drier Mediterranean phase in May. This feature likely explains its general preference, particularly in island environments (Pérez-Jordà et al., 2018; Stika et al., 2024). As an example, during the loss of crop diversity in the Canary Islands, barley was the cereal that most clearly “resisted” until the arrival of the Spaniards, probably due to its lower demand for genetic enrichment and greater adaptability in case of isolation (Morales et al., 2023; Hagenblad and Morales, 2020).
The exclusive presence of barley at Piano dei Cardoni aligns with patterns observed at the earliest neolithic sites in Calabria. Wheat becomes more common starting in the Middle Neolithic, with overall crop diversity increasing during this period. In Apulia, for instance, Triticum spp. are dominant (Costantini and Stancanelli, 1994; Natali and Tiné, 2002), although the distribution of species varies significantly across regions (Natali et al., 2021). In Sicily, however, archaeobotanical data for this phase remain limited (Speciale et al., 2024a). It is therefore impossible, at this stage of research, to determine whether the predominance of barley at Piano dei Cardoni indicates a cultural choice or an adaptation to local environmental features—though it is worth noting that barley was preferred among other cereals on the Sicilian small volcanic islands until recent times (Speciale, 2021).
6.1.3. Animal breeding, hunting, and fishing
The faunal assemblage from the site is dominated by mammal remains, with a clear predominance of ovicaprines (O. aries more abundant than C. hircus). Pig (S. domesticus) remains are present in moderate quantities, while cattle (B. taurus) are almost absent—as expected on an island with limited water sources. Mortality profiles suggest that animals were raised to maturity to maximize meat yield, although young individuals, especially piglets and juvenile ovicaprines were also consumed.
The introduction of wild animal species to Mediterranean islands is not unusual and is observed in other insular contexts (Masseti, 2006; Hofman and Rick, 2018). “Niche enhancement” may occur when human groups take steps to boost the population and availability of their prey species, for instance by introducing them to islands to establish populations that can be hunted. One of the earliest recorded examples is the introduction of the cuscus, a marsupial, from New Guinea to New Ireland approximately 23,500 years ago. In Ustica, the introduction of hare and fox remains uncertain (Prillo et al., 2024).
Hunting activities primarily targeted birds. The most frequently identified species include mallard (A. platyrhynchos) and heron (Ardea sp.), suggesting the existence of small water reserves on the island during the Neolithic—possibly corresponding to the modern gorghi (Speciale et al., 2023). However, the most frequently hunted birds belong to predatory, pheasant-like and sparrow-like taxa. Notably, the presence of birds of prey, such as marsh harriers, in secondary deposition burials—alongside more commonly consumed species—suggests intentional consumption rather than an accidental presence. If these birds were intrusive, more complete skeletons and a broader range of anatomical elements would be recorded. Instead, the concentration of wing bones (e.g., ulna and carpometacarpus) points to selective inclusion, possibly linked to ritual practices (Prillo et al., 2024). Moreover, the consumption of birds of prey in insular contexts is not unexpected. Subsistence strategies on islands often relied on the full range of available animal resources. Exploitation of wild fauna—especially birds and small mammals—is typically more intense on islands than on the mainland, and in some cases, contributed to local extinctions (Rick et al., 2013; Bover et al., 2016; Médail and Pasta, 2024). The association of bird taxa linked to wetlands suggests the persistence of freshwater areas during the Neolithic.
Marine resources played an important role in the inhabitants' diet, as indicated by the numerous fish remains, and mollusks. The potential presence of medium- to large-sized fish could indicate the use of relatively advanced fishing techniques, a hypothesis further supported by the presence of bone hooks and stone weights (Mantia et al., 2021).
It is increasingly recognized that the introduction of (over)grazing livestock and other animals—such as rats—has often had dramatic effects on island ecosystems. These changes may have significantly altered the original landscape, making islands appear more marginal or depauperate than they were prior to historic times (Fitzpatrick et al., 2015). Ovicaprines from Ustica were smaller in size compared to those from other contemporary sites in Sicily and southern Italy. Although further investigation is needed, this size reduction may reflect human management strategies shaped by the island's limited resources. A similar trend of reduced body size under environmental pressure is for example recorded in the Bronze Age in the Balearic islands (Valenzuela, 2023; Valenzuela-Suau et al., 2023) and in the modern times in cattle from Pantelleria (La Mantia, 2018). As noted by Schüle (1993), large mammals on small islands are usually naturally absent, due to their potential to rapidly deplete local vegetation.
6.2. Pantelleria
6.2.1. Wood resources and fruit trees
The use of wood in Pantelleria during the Bronze Age is marked by the exploitation of the forest and shrub species, mostly pines (both Aleppo and maritime), evergreen oaks, lentisk and heather. This is the first time that the archaeobotanical analysis allows to describe the co-occurrence of two different species of pines on the island, dating back at least to the 2nd millennium BCE—likely reflecting their natural occurrence before human arrival, a presence that has continued to the present day. Heather and other Ericaceae, such as strawberry trees, are known to reach several meters in height, especially on islands, and it is therefore highly probable that the exploited individuals were arboreal rather than shrubby. A small quantity of juniper was also recorded, particularly in the layers rich of pines, suggesting their shared presence within the same vegetation formation. Significant the absence of deciduous oaks, that are recorded in the pollen sequence of Lago di Venere many centuries later (Calò et al., 2013). It is uncertain why the Bronze Age inhabitants of Mursia excluded deciduous oaks from any kind of wood procurement. The presence of oaks is so far limited to holm oaks, recorded in the three huts, while heather and lentisk are prevalent in the hut of the sector B and pines and juniper in the sector E.
The absence of olive trees stands out as unusual when compared to the general trend in Sicily, especially considering their documented presence in the record of Pantelleria during the Punic and Roman periods (Speciale et al., unpublished data) and in the Lago di Venere sequence (Calò et al., 2013). At the current state of research, it remains unclear whether olive trees were absent from the natural vegetation and only introduced by humans at a later stage, or whether their absence in these layers in these huts—absence recorded also in the carpological record—is simply a matter of chance. What is certain is that olive trees are quite spread in the Central Mediterranean in the Middle Bronze Age (see Section 6.1.1).
Compared to the rather diversified wood exploitation in Ustica, evidence from Pantelleria suggests a more selective exploitation of arboreal plants, possibly highlighting changes over time. However, the dataset is too limited to determine if the variation in species representation between the first/second and the third macro-phase of the village reflects a real shift in economic strategies and wood use. For sure, a general trend toward increased aridity is documented at around 1550 BCE (Speciale et al., 2016, 2024a). Despite this climatic drying, the archaeological record points to a marked increase in population—an inverse relationship with climate conditions. This apparent resilience may be attributed to improved methods for managing drought and food stress, as well as the development of extensive trade networks and logistical infrastructures typical of more complex societies (Palmisano et al., 2021).
When compared with data from the Aeolian Islands during the Bronze Age, Ericaceae emerge as the dominant plant family—used both for construction and as fuel—particularly on the island of Filicudi, and are also vastly represented in Lipari. In this latter island, the greater diversity of charcoal remains shows the island's heterogenous shrub and tree vegetation, including pines, holm and deciduous oaks, poplars, Fabaceae, and olive trees (Speciale, 2021).
In the Balearic Islands, at the site of Seis Passes, a dynamic exploitation of forest resources is evident, shown by the inversely proportional trends in the use of pines and lentisk (Picornell-Gelabert and Carrión Marco, 2017). A similar inverse relationship is observed between the huts in sectors B and E huts at Mursia.
Notably, often arboriculture is an important source on islands (as showed for fig and olive trees in neolithic Ustica but more in general as well, see e.g. Huebert, 2014). In Mursia, wild species represent an important subsistence source probably during the third macro-phase, but the presence of fig achenes, even if F. carica is absent from the charcoal record, indicates a potential import of the arboreal species, as seen for Ustica, from the mainland, or an exploitation of imported syconia, mostly during the third macro-phase.
6.2.2. Crops and weeds
As observed in the shift in wood gathering practices, the exploitation of economic plants appears to change as well across the occupational sequence. In the first and second macro-phases, domestic plant species are predominant. Hulled and naked wheats likely represent the exclusive cereals, and pulses also play an important role. The significant presence of lentisk fruits during these phases corresponds with its high representation in the charcoal record, suggesting a broader use of this plant resource. In contrast, the third macro-phase is characterized by a higher representation of weeds, particularly purslane and goosefoot. Despite this shift, cereals remain part of the assemblage, although they appear in more degraded conditions, which may reflect taphonomic factors affecting preservation. The presence of these remains within the huts strongly suggests human consumption. However, it is not clear whether the wild herbaceous taxa in the carpological assemblage of Mursia represent the exploitation of native plants, unintentional introductions as weeds alongside cultivated plants, or deliberate introductions for their nutritional and/or medicinal properties.
In the Aeolian Islands during the Bronze Age, crop exploitation patterns show notable variability. At Filo Braccio (Filicudi), there is a strong representation of hulled barley and pulses, particularly lentils and fava beans. In contrast, data from Lipari pertain only to the final phase of the Bronze Age and reveal a more heterogenous use of cultivated plants (Speciale, 2021).
Shifting cultivation appears to have been among the earliest agricultural strategies adopted on many islands (e.g., McCoy, 2006). Typically, island agricultural systems begin with the introduction of a limited number of plants, followed over time by increasing specialization—a dynamic described in several contexts (Fitzpatrick and Keegan, 2007; Fitzpatrick, 2015). In Polynesia, Kirch (1984) outlined this trajectory as one of adaptation, expansion, and intensification. Defining clear developmental stages in the Mediterranean is challenging. This complexity arises from the frequent cycles of occupation and abandonment, the close connections between many islands and the mainland, and the strategic importance of islands such as Pantelleria in wider exchange networks (Dawson, 2014). Nevertheless, the significant exploitation of wild plant species during the Bronze Age may be interpreted as a strategy of adaptation to the local resources and/or a response to broader climatic changes, such as those occurring at the mid-16th century BCE (Speciale et al., 2024a, see also Section 6.2.1).
6.2.3. Animal breeding, hunting, and fishing
Terrestrial animal resources at Mursia are not highly represented in the hut contexts here presented, whereas marine resources (fish and mollusks) appear to be more prominent, along with birds of various sizes, similar to the pattern observed in Ustica. Ovicaprines dominate along all the macro-phases, while pigs and bovids are recorded in lower quantities. Significant the absence of dogs; this looks in contrast with the increase of bovids and decrease of pigs in the latest phase—despite the cattle being driven on the mainland by herding dogs. However, the absence of wild predators and the limited extent of the territory likely enabled local strategies that made cattle husbandry possible without the use of dogs. Data are still too preliminary, but a change in husbandry strategy along the occupation of the village cannot be ruled out (Fiori, 2025), taking also into consideration the general increase in aridity during this phase, in contrast with the higher demand of water for cattle compared to other livestock. Finally, recent isotopic analyses on faunal remains belonging to the first and second macrophases opened the possibility of import of animals from the mainland (Dawson et al., 2024).
Marine resources played a significant role in the diet throughout all the phases, as did avifauna. This suggests that wild marine fauna and birds, easily available on the island, continued to form an important part of the subsistence economy even during the Middle Bronze Age—a pattern that contrasts somewhat with the broader trend in Southern Italy and Sicily, where a decline in wild resource exploitation is generally observed (De Grossi Mazzorin et al., 2004; Arena et al., 2020; Speciale et al., 2024a). The presence of marine seals, recovered among marine animals, was very common in Sicily until recent times (La Mantia and Pasta, 2008). The substantial reliance on meat-based resources at Mursia aligns with dietary trends observed in the Italian peninsula (Varalli et al., 2022), although only stable isotopes analysis of human remains can clarify the extent to which plant-based foods contributed to the Bronze Age diet at Mursia.
In the Aeolian islands, the faunal assemblage from the small island of Filicudi during the Early and Middle Bronze Age is dominated by ovicaprines, although both pigs and bovids are present. The low representation of marine resources may be attributed to excavation methods, while terrestrial wild animals are absent. At the multilayered site of Lipari Acropolis, a clear diachronic trend is observable: from the Neolithic to the end of the Bronze Age, the representation of bovids and pigs increases, while ovicaprines remain consistently present. This trend is accentuated with the most significant socio-cultural transformation, occurring between the Middle and Late Bronze Age, when the arrival of people from southern Italy may have led to a marked shift in dietary practices (Speciale, 2021; Villari, 1996).
When compared to other small Mediterranean islands, the pattern slightly diverges. During the Neolithic, smaller islands in the Aegean and Adriatic Sea, seem to prefer the exploitation of marine resources (Pilaar Birch, 2017), a trend also observed on larger islands in case of demographic pressure (Arikan, 2023). However, during the Bronze Age, dietary strategies on coastal areas seem to shift (Nuttall, 2021). In the western Mediterranean, Formentera (Balearic archipelago) stands out from larger neighboring islands, likely due to its geographic features, with a relatively higher proportion of wild animal remains in the faunal record (Ramis, 2017).
6.3. Adapting to insular features in Ustica and Pantelleria
Strategic voyaging and the transplantation of plants and animals were key to successful colonization, facilitating both subsistence and cultural continuity (Anderson, 2009). The suite of animals and crops provided not only a buffer against potential resource shortfalls but also a symbolic marker of identity, and a psychological anchor in unfamiliar environments (LeFebvre and Giovas, 2009), though adapted to the new insular context.
Taken together, the three datasets in Piano dei Cardoni reveal how Neolithic communities adapted to the insular environment through:
- A strong reliance on a diversified economy in which wild plants, cultivated crops and domesticates from the mainland and marine resources were combined.
- The heterogenous exploitation of woody species from all the potential series present on the island, probably due to the low plant diversity on such a small and flat insular environment and a choice of general low impact on local vegetation.
- The preference for barley as potential adaptation to local rather arid conditions or to some specific dietary choice, while the reliance on wild plants is quite common during Middle Neolithic also on non-insular environments.
- The domestic animal species, reflecting those on the mainland, though adapted to the island's constraints particularly in terms of water and fodder availability—a strong dependence on ovicaprine husbandry of limited size and very few pigs and cattle (Prillo et al., 2024, 2025).
- The whole economic system, relying on limited agriculture, but also significant exploitation of wild faunal resources: targeted bird hunting—typical insular preys, and marine resources.
For the settlement of Mursia during the Bronze Age, the indicators of insular adaptation are less evident: Pantelleria is more than ten times larger than Ustica and with a significant altitude gradient. Moreover, Bronze Age communities rely on a more structured and complex society. Compared with other islands—Ustica, Lipari, and Filicudi—Pantelleria shows parallels but also unique local responses, probably for being the largest and most heterogenous in terms of habitat variety, but also the most isolated among the case studies. Nevertheless, the potential signals are:
- The wood assemblage, revealing an integrated use of local forest and maquis species, including the co-occurrence of Aleppo and maritime pines, evergreen oaks, lentisk, heather, and juniper. Human groups relied more on maquis species like lentisk and heather tree before and then started exploiting more intensively holm oaks, pines and junipers. This change in fuel and wood management should be further investigated, including the absence of deciduous oaks.
- The absence of olive trees, in contrast with their known prevalence elsewhere in the Central Mediterranean, raising questions about local environmental conditions or cultural preferences, notably the production of lentisk instead of olive oil. Shifts between the first/second and third macro-phases—evident in wood use, cultivated and wild plant consumption, and animal resource management—point to a potential change in exploitation strategies, maybe in response to climatic stress around 1550 BCE and increasing societal complexity.
- The wide exploitation of wild species of plants during the Middle Bronze Age, such as lentisk and purslane, alongside cereals and pulses, and a crop tree like fig, highlighting a flexible subsistence system and an exploitation of plants that are aridity-tolerant.
- Marine and avifaunal remains dominating the faunal record, indicating a strong reliance on easily accessible wild resources, in contrast to some mainland trends during the 2nd millennium BCE.
- The potential import of cattle from the mainland, being extremely water-requiring and then probably unadapt to an island, especially in a phase of increased aridity like the one at the mid of the 16th century BCE.
7 Conclusions
Similarly to other island contexts, also in the prehistoric Mediterranean “individuals and collectives within societies are neither exclusively environmental stewards nor the agents of detrimental ecological changes” (Lepofsky and Kahn, 2011: p. 330), but act under a range of socio-economic and cultural motivations as well as environmental constraints and opportunities, each continuously evolving. Nevertheless, the management of resources remains one of the most crucial aspects of human settling on small islands.
This work shows how multidisciplinarity represents the only meaningful approach for the analysis of resources on islands and more in general in the archaeological contexts (Table 6). The carpological, anthracological and faunal analysis of the site of Piano dei Cardoni provides significant insights into the plant and animal resources used by the inhabitants of Ustica during the Neolithic, so after the first permanent occupation of the island. The presence of plant species now absent on the island, such as deciduous oaks, heather and strawberry trees, suggests climatic or environmental variations compared to the present day. The choices that the human settlers make in the colonization process, probably bringing tree crops like figs and olive trees, and not just herbal crops, tend to point to a clear intent of sedentary occupation of the island in the long term.
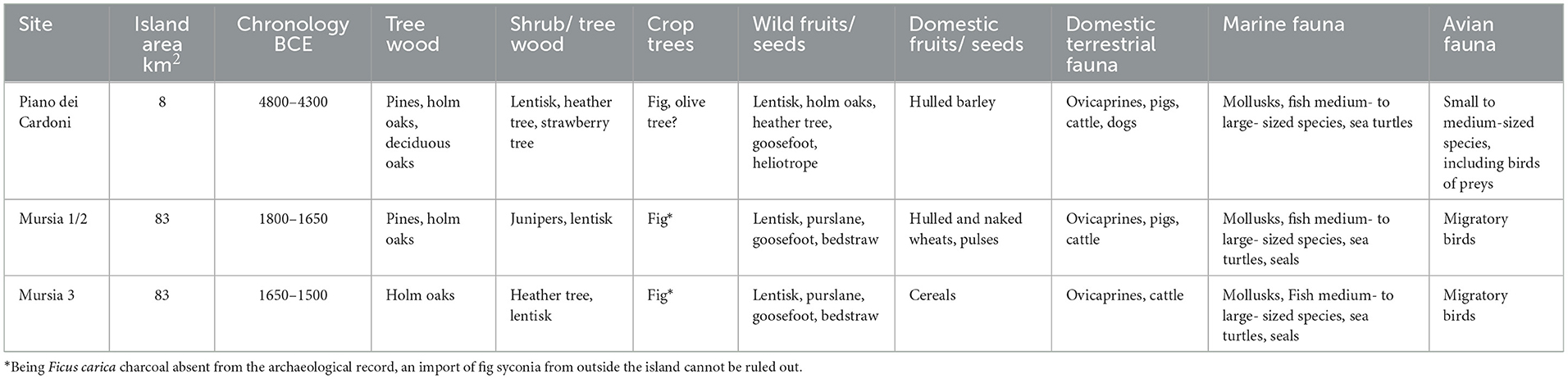
Table 6. Comparative table with the main plant and animal species from the two case studies.
For Pantelleria island, so far, no data is available for its Neolithic occupation. Therefore, data from the Bronze Age site of Mursia show human adaptation to a landscape that could have been already affected by 3,000 years of former human occupation. Nevertheless, Pantelleria is naturally more ecologically diverse and resource-rich than Ustica, and it remains unclear if Neolithic people were permanently occupying the island. The archaeobotanical and zooarchaeological data from Mursia offer a first multidisciplinary view of Bronze Age resource exploitation on Pantelleria, highlighting both continuity and transformation in subsistence strategies. Ultimately, the human Bronze Age community of Mursia appears leveraging diverse local ecological niches while embedded in broader Mediterranean networks of exchange and innovation.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
CS: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Validation, Writing – original draft, Writing – review & editing. MCar: Data curation, Investigation, Methodology, Writing – review & editing. FF: Data curation, Investigation, Writing – review & editing. VP: Data curation, Investigation, Writing – original draft, Writing – review & editing. EA: Data curation, Writing – review & editing. MCat: Funding acquisition, Methodology, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Postdoctoral contract to CS for the project “SILVA – Sicilian small IsLands Vegetation under the effect of the human Arrival” is part of the European Union's Horizon 2020 research and innovation programme MSCA-COFUND R2STAIR (GA 101034349). The IPHES-CERCA has received financial support through the “María de Maeztu” program for Units of Excellence (CEX2024-01485-M/funded by MICIU/AEI/10.13039/501100011033) and 2023: URV-PFR-1237; 2024: URV-PFR-1237, SGR 2021-1237.
Acknowledgments
Soprintendenza dei Beni Culturali di Palermo, Giuseppina Battaglia; Valeria Giuliana and Fondazione Mirella Vitale for the economic support of the investigations on Ustica; Istituto Nazionale di Geofisica e Vulcanologia, Osservatorio Vesuviano, Sandro de Vita and Mauro Di Vito. For the SEM pictures, we thank Mercè Moncusí from the Servei de Recursos Cientifico Tècnics at the Unversitat Rovira i Virgili (Tarragona). We sincerely thank the editors and the reviewers for helping us in improving significantly the manuscript. Salvatore Pasta gave us as well his notes and comments and we are thankful for his constant support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fearc.2025.1621064/full#supplementary-material
References
Anderson, A. (2002). Faunal collapse, landscape change and settlement history in Remote Oceania. World Archaeol. 33, 375–390. doi: 10.1080/00438240120107431
Anderson, A. (2009). The rat and the octopus: initial human colonization and the prehistoric introduction of domestic animals to Remote Oceania. Biol Invasions 11, 1503–1519. doi: 10.1007/s10530-008-9403-2
Aprile, G., and Fiorentino, G. (2018). “L'analisi antracologica dei residui di rogo per la ricostruzione delle fasi meno visibili del rituale incineratorio,” in I Tumuli Della Necropoli di Salve. Architetture e rituali nell'ideologia funeraria dell'età del Rame (Santo Spirito: Edipuglia), 63–69.
Arena, F., Gualdi-Russo, E., Olsen, J., Philippsen, B., and Mannino, M. A. (2020). New data on agro-pastoral diets in southern Italy from the Neolithic to the Bronze Age. Archaeol. Anthropol. Sci. 12:245. doi: 10.1007/s12520-020-01209-9
Arikan, B. (2023). Simulating land use dynamics and the role of marine resources in diets during the Early Bronze Age-II (ca. 4900–4700 cal. BP) at Gökçeada (ancient Imbros) in the northern Aegean. J. Archaeol. Sci.: Rep. 49:104002. doi: 10.1016/j.jasrep.2023.104002
Asouti, E., Kabukcu, C., White, C. E., Kuijt, I., Finlayson, B., Makarewicz, C., et al. (2015). Early Holocene woodland vegetation and human impacts in the arid zone of the southern Levant. Holocene 25, 1565–1580. doi: 10.1177/0959683615580199
Behre, K. E. (2008). Collected seeds and fruits from herbs as prehistoric food. Veget Hist. Archaeobot. 17, 65–73. doi: 10.1007/s00334-007-0106-x
Bevan, A., and Conolly, J. (2011). Terraced fields and Mediterranean landscape structure: an analytical case study from Antikythera, Greece. Ecol. Modell. 222, 1303–1314. doi: 10.1016/j.ecolmodel.2010.12.016
Boessneck, J. (1969). “Osteological differences between sheep (Ovis aries Linné) and goat (Capra hircus Linné),” in Science in Archaeology. A Comprehensive Survey of Progress and Research, eds. D. Brothwell and E. Higgs (London: Thames and Hudson), 331–358.
Boessneck, J., Müller, H. H., and Teichert, M. (1964). Osteologische Unterscheidungsmerkmale zwischen Schaf (Ovis aries) und Ziege (Capra hircus). Kühn Archiv. 78, 1–129.
Bökönyi, S. (1970). A new method for the determination of the number of individuals in animal bone material. Am. J. Archaeol. 74, 291–292. doi: 10.2307/503105
Bover, P., Valenzuela, A., Torres, E., Cooper, A., Pons, J., Alcover, J. A., et al. (2016). Closing the gap: new data on the last documented Myotragus and the first human evidence on Mallorca (Balearic Islands, Western Mediterranean Sea). Holocene 26, 1887–1891. doi: 10.1177/0959683616645945
Braje, T., Leppard, T., Fitzpatrick, S., and Erlandson, J. (2017). Archaeology, historical ecology and anthropogenic island ecosystems. Environ. Conserv. 44, 286–297. doi: 10.1017/S0376892917000261
Brose, U., Ostling, A., Harrison, K., and Martinez, N. D. (2004). Unified spatial scaling of species and their trophic interactions. Nature 428, 167–171. doi: 10.1038/nature02297
Bull, G., and Payne, S. (1982). “Tooth eruption and epiphysial fusion in pigs and wild boar,” in Ageing and Sexing Animal Bones from Archaeological Sites, eds. B. Wilson, C. Grigson, and S. Payne (Oxford: BAR Publishing), 55–72.
Bullock, D., and Rackham, J. (1982). “Epiphysial fusion and tooth eruption of feral goats from moffatdale, dumfories and galloway,” in Ageing and Sexing Animal Bones from Archaeological Sites, eds. B. Wilson, C. Grigson, and S. Payne (Oxford: BAR Publishing), 73–80.
Caballero, R., Fernandez-Gonzalez, F., Badia, R. P., Molle, G., Roggero, P. P., Bagella, S., et al. (2009). Grazing systems and biodiversity in Mediterranean areas: Spain, Italy and Greece. Pastos 39, 9–154.
Calò, C., Henne, P. D., Curry, B., Magny, M., Vescovi, E., La Mantia, T., et al. (2012). Spatio-temporal patterns of Holocene environmental change in southern Sicily. Palaeogeogr. Palaeoclimatol. Palaeoecol. 323, 110–122. doi: 10.1016/j.palaeo.2012.01.038
Calò, C., Henne, P. D., Eugster, P., van Leeuwen, J., Gilli, A., Hamann, Y., et al. (2013). 1200 years of decadal-scale variability of Mediterranean vegetation and climate at Pantelleria Island, Italy. Holocene 23, 1477–1486. doi: 10.1177/0959683613493935
Cambini, A. (1967). Riconoscimento microscopico del legno delle querce italiane. In Contributi scientifico-pratici per una migliore conoscenza del legno, 10, ed. G. Giordano (Roma: C.N.R.), 51–80.
Cappers, R. T. J., and Neef, R. (2016). Digital Atlas of Traditional Agricultural Practises and Food Processing. Groningen: Barkhuis and Groningen University Library.
Caracuta, V. (2020). Olive growing in Puglia (southeastern Italy): a review of the evidence from the Mesolithic to the Middle Ages. Veget. Hist. Archaeobot. 29, 595–620. doi: 10.1007/s00334-019-00765-y
Carrión, Y., Ntinou, M., and Badal, E. (2010). Olea europaea L. in the north Mediterranean Basin during the Pleniglacial and the Early–Middle Holocene. Quat. Sci. Rev. 29, 952–968. doi: 10.1016/j.quascirev.2009.12.015
Cattani, M. (2015). Gli scavi nel settore B dell'abitato dell'età del Bronzo di Mursia (Pantelleria). IpoTESI di Preistoria 7, 1–16. doi: 10.6092/issn.1974-7985/5965
Cattani, M., Barchiesi, F., Conti, A. M., Debandi, F., Fiori, F., Magrì, A., et al. (2024). Pantelleria come polo di attrazione e connettività nel Mediterraneo centrale: dalla circolazione dell'ossidiana agli scambi nella media età del Bronzo. Riv. Sci. Preistoriche 74, 79–89.
Cattani, M., and Peinetti, A. (2023). La durata di vita degli abitati. Attività e fasi costruttive tra analisi stratigrafica e cronologia. Spazi domestici nell'età del Bronzo. Memorie del Museo Civico di Storia Naturale 2, 45–64.
Cherry, J. F., and Leppard, T. P. (2018). Patterning and its causation in the pre-Neolithic colonization of the Mediterranean islands (Late Pleistocene to Early Holocene). J. Island Coast. Archaeol. 13, 191–205. doi: 10.1080/15564894.2016.1276489
Cohen, A., and Serjeanston, D. (1996). A Manual for the Identification of Bird Bones from Archaeological Sites. Revised edition. London: Archetype Publications.
Costantini, L., and Stancanelli, M. (1994). La preistoria agricola dell'Italia centro-meridionale: il contributo delle indagini archeobotaniche. Origini 18, 149–244.
D'Auria, A., Buonincontri, M. P., Allevato, E., Saracino, A., Jung, R., Pacciarelli, M. G., et al. (2017). Evidence of a short-lived episode of olive (Olea europaea L.) cultivation during the Early Bronze Age in western Mediterranean (southern Italy). Holocene 27, 605–612. doi: 10.1177/0959683616670218
Davis, S. J. M. (1996). Measurements of a group of adult female Shetland sheep skeletons from a single flock: a baseline for zoo-archaeologists. J. Archaeol. Sci. 23, 593–612. doi: 10.1006/jasc.1996.0056
Davis, S. J. M. (2000). The effect of castration and age on the development of the shetland sheep skeleton and a metric comparison between bones of males, females and castrates. J. Archaeol. Sci. 27, 373–390. doi: 10.1006/jasc.1999.0452
Dawson, H. (2014). Mediterranean Voyages: The Archaeology of Island Colonisation and Abandonment. New York, NY: Routledge.
Dawson, H., Cattani, M., Spatafora, F., Curci, A., Fiori, F., Mannino, M. A., et al. (2024). Dinamiche di sussistenza e resilienza nelle isole minori della Sicilia: indagini isotopiche preliminari a confronto sulle faune domestiche di Pantelleria ed Ustica durante l'età del Bronzo. Riv. Sci. Preistoriche 74, 243–255.
De Grossi Mazzorin, J., Riedel, A., and Tagliacozzo, A. (2004). “L'evoluzione delle popolazioni animali e dell'economia nell'età del Bronzo recente,” in L'età del Bronzo Recente in Italia: Atti del Congresso Nazionale di Lido di Camaiore, 26-28th October, 2000, ed. D. Cocchi Genick (Viareggio: Baroni), 227–232.
de Vita, S., and Foresta Martin, F. (2017). The palaeogeographic setting and the local environmental impact of the 130 ka Falconiera tuff-cone eruption (Ustica island, Italy). Ann. Geophys. 60, S0224–S0224. doi: 10.4401/ag-7113
Debandi, F. (2015). La capanna B14 dell'abitato dell'età del Bronzo di Mursia (Pantelleria). IpoTesi di Preistoria 7, 71–136. doi: 10.6092/issn.1974-7985/5966
Debandi, F., Cattani, M., and Peinetti, A. (2019). Focolari e piastre di cottura nell'abitato dell'età del Bronzo di Mursia (Pantelleria TP), IpoTESI di Preistoria 12, 111–142.
Debandi, F., and Magrì, A. (2021). Gli alari nell'abitato dell'età del Bronzo di Mursia (Pantelleria): analisi dei modelli e comparazioni tra Sicilia e Mediterraneo orientale. Riv. Sci. Preistoriche 71, 1–66.
Dotte-Sarout, E. (2017). Evidence of forest management and arboriculture from wood charcoal data: an anthracological case study from two New Caledonia Kanak pre-colonial sites. Veget. Hist. Archaeobot. 26, 195–211. doi: 10.1007/s00334-016-0580-0
Fiori, F. (2025). L'archeozoologia e lo studio delle razze autoctone: contesti insulari a confronto con l'Italia centrale (Ph.D. thesis, Scienze Storiche e Archeologiche. Memoria, Civilta' e Patrimonio). University of Bologna.
Fiori, F., Carra, M., Curci, A., Debandi, F. I., Iacono, P., Peinetti, A., et al. (2024). Il ruolo della bioarcheologia nella ricostruzione paleoeconomica e paleoambientale dell'abitato protostorico di Mursia (Pantelleria). Riv. Sci. Preistoriche 74, 459–469.
Fitzpatrick, S. M. (2015). The Pre-Columbian Caribbean: colonization, population dispersal, and island adaptations. Paleo America 1, 305–331. doi: 10.1179/2055557115Y.0000000010
Fitzpatrick, S. M., and Keegan, W. F. (2007). Human impacts and adaptations in the Caribbean Islands: an historical ecology approach. Earth Environ. Sci. Trans. R. Soc. Edinburgh 98, 29–45. doi: 10.1017/S1755691007000096
Fitzpatrick, S. M., Rick, T. C., and Erlandson, J. M. (2015). Recent progress, trends, and developments in island and coastal archaeology. J. Island Coast. Archaeol. 10, 3–27. doi: 10.1080/15564894.2015.1013647
Forgia, V., and Oll?, A. (2023). Il riparo di Vallone Inferno (Madonie, Sicilia). Oxford: BAR Publishing. doi: 10.30861/9781407361130
Forgia, V., Speciale, C., and Dawson, H. (2024). Il mare dentro l'Isola: un approccio territoriale allo studio delle risorse legate al mare nel Neolitico e nell'età del Bronzo in Sicilia. Riv. Sci. Preistoriche S4, 363–383.
Gentry, A., Clutton-Brock, J., and Groves, C. P. (2004). The naming of wild animal species and their domestic derivatives. J. Archaeol. Sci. 31, 645–651. doi: 10.1016/j.jas.2003.10.006
Gianguzzi, L., and Bazan, G. (2020). A phytosociological analysis of the Olea europaea L. var. sylvestris (Mill.) Lehr. forests in Sicily. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 154, 705–725. doi: 10.1080/11263504.2019.1681532
Giannuzzi-Savelli, R., Pusateri, F., Palmeri, A., and Ebreo, C. (1999). Atlante delle conchiglie marine del Mediterraneo. Rome: Evolver srl.
Giovas, C. M. (2016). Though she be but little: resource resilience, amerindian foraging, and long-term adaptive strategies in the Grenadines, West Indies. J. Island Coast. Archaeol. 11, 238–263. doi: 10.1080/15564894.2016.1193572
Girod, A. (2015). “Potenziale informativo dei molluschi marini, d'acqua dolce e terrestri in archeologia,” in Appunti di Archeomalacologia, ed. A. Girod (Florence: All'Insegna del Giglio), 11–128.
Grant, A. (1982). “The use of tooth wear as a guide to the age of domestic ungulates,” in Ageing and Sexing Animal Bones from Archaeological Sites, eds. B. Wilson, C. Grigson, and S. Payne (Oxford: BAR Publishing), 91–108.
Guarrera, P. M. (2006). Household dyeing plants and traditional uses in some areas of Italy. J. Ethnobiol. Ethnomed. 2:9. doi: 10.1186/1746-4269-2-9
Hagenblad, J., and Morales, J. (2020). An evolutionary approach to the history of Barley (Hordeum vulgare) cultivation in the Canary Islands. Afr. Archaeol. Rev. 37, 579–595. doi: 10.1007/s10437-020-09415-5
Halstead, P., Collins, P., and Isaakidou, V. (2002). Sorting the sheep from the goats: morphological distinction between the mandibles and the mandibular teeth of adult ovis and capra. J. Archaeol. Sci. 29, 543–545. doi: 10.1006/jasc.2001.0777
Hofman, C. A., and Rick, T. C. (2018). Ancient biological invasions and island ecosystems: tracking translocations of wild plants and animals. J. Archaeol. Res. 26, 65–115. doi: 10.1007/s10814-017-9105-3
Huang, M., Shen, S., Luo, C., and Ren, Y. (2019). Genus Periploca (Apocynaceae): a review of its classification, phytochemistry, biological activities and toxicology. Molecules 24:2749. doi: 10.3390/molecules24152749
Huebert, J. M. (2014). The Role of Arboriculture in Landscape Domestication and Agronomic Development: A Case Study from the Marquesas Islands, East Polynesia (Ph.D. thesis). University of Auckland, New Zealand.
Huebert, J. M., and Allen, M. S. (2016). Six centuries of anthropogenic forest change on a Polynesian high island: archaeological charcoal records from the Marquesas Islands. Quat. Sci. Rev. 137, 79–96. doi: 10.1016/j.quascirev.2016.01.017
Huebert, J. M., and Allen, M. S. (2020). Anthropogenic forests, arboriculture, and niche construction in the Marquesas Islands (Polynesia). J. Anthropol. Archaeol. 57:101122. doi: 10.1016/j.jaa.2019.101122
Ingravallo, E., and Tiberi, I. (2008). Il neolitico salentino nel circuito internazionale di prestiti e scambi. Studi di Antichità 12, 71–95.
Keegan, W. F., Fitzpatrick, S. M., Sullivan Sealey, K., LeFebvre, M. J., and Sinelli, P. T. (2008). The role of small islands in marine subsistence strategies: case studies from the caribbean. Hum. Ecol. 36, 635–654. doi: 10.1007/s10745-008-9188-z
Kirch, P. V. (1984). The Polynesian outliers: continuity, change, and replacement. J. Pac. Hist. 19, 224–238. doi: 10.1080/00223348408572496
Kirch, P. V., and Yen, D. E. (1982). Tikopia. The prehistory and ecology of a Polynesian outlier. Bernice P. Bishop Museum Bull. Honolulu 238, 1–396.
Kolodziejczyk-Czepas, J. (2016). Trifolium species – the latest findings on chemical profile, ethnomedicinal use and pharmacological properties. J. Pharm. Pharmacol. 68, 7, 845–861. doi: 10.1111/jphp.12568
La Mantia, T. (2018). La diversità degli animali domestici nelle isole circumsiciliane: un patrimonio scomparso prima di essere conosciuto. Naturalista Sicil. Ser. 4, 137–147.
La Mantia, T., and Pasta, S. (2008). “Distribuzione pregressa ed estinzione della Foca monaca (Monachus monachus) in Sicilia (Carnivora Phocidae),” in AAVV (a cura di), Atlante della biodiversità della Sicilia. Vertebrati terrestri (Palermo: ARPA Sicilia), 109–112.
LeFebvre, M. J., and Giovas, C. M. (2009). The zooarchaeology of Islands: towards synergy and synthesis. J. Island Coastal Archaeol. 4, 141–150. doi: 10.1080/15564890903149532
Lepofsky, D., and Kahn, J. (2011). Cultivating an ecological and social balance: elite demands and commoner knowledge in ancient Ma ‘ohi agriculture, society islands. Am. Anthropol. 113, 319–335. doi: 10.1111/j.1548-1433.2011.01333.x
Lepofsky, D. S. (1994). Prehistoric Agricultural Intensification in the Society Islands, French Polynesia. Berkeley, CA: University of California.
Leppard, T. P., and Pilaar Birch, S. E. (2019). “The insular ecology and palaeoenvironmental impacts of the domestic goat (Capra hircus) in Mediterranean Neolithization,” in Géoarchéologie des îles de la Méditerranée, ed. M. Ghilardi (Paris: CNRS Éditions via OpenEdition), 47–56.
Lomolino, M. V., and Van der Geer, A. A. (2023). The paradox of anthropogenic enrichment: homogenization and downsizing of insular mammals. Front. Biogeogr. 15:9967. doi: 10.21425/F5FBG59967
Magrì, A., Speciale, C., Montana, G., Mentesana, R., and Battaglia, G. (2021). “Piano dei Cardoni (Ustica, Palermo): primi risultati dell'analisi sulle ceramiche del Neolitico Medio-Recente,” in La ceramica in Sicilia dalla Preistoria all'Età contemporanea, Atti del II convegno internazionale (Quasar: Roma).
Mannino, G. (1998). Il neolitico nel palermitano e la nuova scoperta nell'isola di Ustica, Quaderni del Museo Archeologico Regionale. Antonio Salinas 4, 56–57.
Mantia, F., Larosa, N., Battaglia, G., and Speciale, C. (2021). “Piano dei Cardoni, Ustica (Neolitico Medio/Recente): per una tipologia dell'industria litica pesante,” in La Sicilia Preistorica. Dinamiche interne e relazioni esterne. Atti del Convegno Internazionale, Catania-Siracusa 7-9 ottobre 2021, eds. P. Militello, F. Nicoletti, and C. Panvini (Palermo: Regione Siciliana, Assessorato dei Beni Culturali e dell'Identità Siciliana).
Masseti, M. (2003). Holocene endemic and non endemic mammals of the Aegean islands. Br. Schl. Athens Stud. 9, 53–63. Available online at: https://www.jstor.org/stable/40960332
Masseti, M. (2006). Domestic fauna and anthropochorous fauna. Hum. Evol. 21, 85–93. doi: 10.1007/s11598-006-9009-6
Mavromati, A. (2022). Wood charcoal macroremains from the Heraion on Samos: firewood and tree management during the Early-Middle Bronze and Roman periods. Archaeol. Anthropol. Sci. 14:231. doi: 10.1007/s12520-022-01700-5
McCoy, M. D. (2006). Landscape, Social Memory, and Society: An Ethnohistoric-Archaeological Study of Three Hawaiian Communities (Ph. D. dissertation). Berkeley, CA: University of California, Berkeley.
McLaughlin, T. R., Stoddart, S., and Malone, C. (2018). Island risks and the resilience of a prehistoric civilization. World Archaeol. 50, 570–600. doi: 10.1080/00438243.2018.1515656
Meadow, R. (1999). “The use of size index scaling techniques for research on archaeozoological collections from the Middle East,” in Historia Animalium ex Ossibus. Festschrift für Angela von den Driesch, eds. C. Becker, H. Manhart, J. Peters, and J. Schibler (Espelkamp: Verlag Marie Leidorf GmbH, Rahden/Westf), 285–300.
Médail, F., and Pasta, S. (2024). The intermittent history of exploitation of terrestrial biotic resources on the small islands of the Western Mediterranean Basin. Hum. Ecol. 52, 397–408. doi: 10.1007/s10745-024-00503-7
Montana, G., Speciale, C., Battaglia, G., Randazzo, L., Carnevale, G., and Antonopoulou, M. V. (in review). The Neolithic site of Piano dei Cardoni in the island of Ustica (Italy): minero-petrographic chemical characterization of the ceramic finds clayey raw materials.
Morales, J., Speciale, C., Rodríguez-Rodríguez, A., Henríquez-Valido, P., Marrero-Salas, E., Hernández-Marrero, J. C., et al. (2023). Agriculture and crop dispersal in the western periphery of the Old World: the Amazigh/Berber settling of the Canary Islands (ca. 2nd−15th centuries CE). Veget. Hist. Archaeobot. doi: 10.1007/s00334-023-00920-6
Natali, E., Agrostelli, M., Feriozzi, F., Forgia, V., and Micheli, R. (2021). “I livelli neolitici della Grotta San Michele di Saracena (CS),” in Dal Pollino all'Orsomarso. Ricerche archeologiche fra Ionio e Tirreno (Roma: Edizioni Quasar), 33–59.
Natali, E., and Tiné, V. (2002). “Favella. Calabria,” in Le ceramiche impresse del Neolitico antico. Italia e Mediterraneo, eds. M. A. Fugazzola Delpino, A. Pessina, V. Tiné (Roma: Bullettino di Paletnologia Italiana, Studi di Paletnologia I), 832.
Neef, R., Cappers, R. T., and Bekker, R. M. (2012). Digital Atlas of Economic Plants in Archaeology, Vol. 17. Groningen: Barkhuis. doi: 10.2307/j.ctt20p56d7
Newsom, L. A., and Wing, E. S. (2004). On Land and Sea: Native American Uses of Biological Resources in the West Indies. Tuscaloosa: The University of Alabama Press.
Ntinou, M., and Stratouli, G. (2012). Charcoal analysis at Drakaina Cave, Kephalonia, Ionian islands, Greece. A case study of a specialized Late Neolithic and Chalcolithic site. Wood and charcoal. Evid. Nat. Hum. Hist. 13, 43–52.
Nuttall, C. (2021). Seascape Dialogues: Human-sea interaction in the Aegean from Late Neolithic to Late Bronze Age (Doctoral dissertation). Department of Archaeology and Ancient History, Uppsala University).
Palli, J., Fiolna, S., Bini, M., et al. (2025). The human-driven ecological success of olive trees over the last 3700 years in the Central Mediterranean. Quat. Sci. Rev. 356:109313. doi: 10.1016/j.quascirev.2025.109313
Palmisano, A., Bevan, A., Kabelindde, A., Roberts, N., and Shennan, S. (2021). Long-term demographic trends in prehistoric italy: climate impacts and regionalised socio-ecological trajectories. J. World Prehist. 34, 381–432. doi: 10.1007/s10963-021-09159-3
Pasta, S., D'Amore, G., Di Maggio, C., et al. (2022). “The impact of climate, resource availability, natural disturbances and human subsistence strategies on the sicilian landscape dynamics during the Holocene,” in From Hydrology to Hydroarchaeology in the Ancient Mediterranean, eds. G. Polizzi, V. Ollivier, S. Bouffier (Oxford: Archaeopress), 8–60. doi: 10.2307/j.ctv30pntvk.4
Pasta, S., and Speciale, C. (2021). “Comunità umane e piante in Sicilia: una lunga (prei)storia,” in La Sicilia Preistorica. Dinamiche interne e relazioni esterne, Proceedings of the International Conference, eds. P. Militello, F. Nicoletti, C. Panvini (Palermo: Regione Siciliana), 31–41.
Payne, S. (1973). Kill-off patterns in sheep and goats: the mandibles from Asvan Kale. Anatolian Stud. 33, 281–303. doi: 10.2307/3642547
Payne, S. (1985). Morphological distinctions between the mandibular teeth of young sheep, ovis, and, goats, capra. J. Archaeol. Sci. 12, 139–147. doi: 10.1016/0305-4403(85)90058-5
Pearsall, D. M. (2015). Paleoethnobotany. A Handbook of Procedures. Walnut Creek, CA: Left Coast Press.
Pérez-Jordà, G., Peña-Chocarro, L., Picornell-Gelabert, L., and Marco, Y. C. (2018). Agriculture between the third and first millennium BC in the Balearic Islands: the archaeobotanical data. Veget. Hist. Archaeobot. 27, 253–265. doi: 10.1007/s00334-017-0618-y
Picornell-Gelabert, L., and Carrión Marco, Y. (2017). Landscape and firewood procurement at the prehistoric and protohistoric site of Ses Païsses (island of Mallorca, Western Mediterranean). Quat. Int. 458, 56–74. doi: 10.1016/j.quaint.2017.03.018
Pignatti, S., Guarino, R., and La Rosa, M. (2017–2019). Flora d'Italia, 2nd Edn. Bologna: Edagricole di New Business Media.
Pilaar Birch, S. E. (2017). From the Aegean to the adriatic: exploring the earliest neolithic Island Fauna. J. Island Coast. Archaeol. 13, 256–268. doi: 10.1080/15564894.2017.1310774
Plekhov, D., Leppard, T. P., and Cherry, J. F. (2021). Island colonization and environmental sustainability in the postglacial Mediterranean. Sustainability 13:3383. doi: 10.3390/su13063383
Prigioniero, A., Geraci, A., Schicchi, R., Tartaglia, M., Zuzolo, D., Scarano, P., et al. (2020). Ethnobotany of dye plants in Southern Italy, Mediterranean Basin: floristic catalog and two centuries of analysis of traditional botanical knowledge heritage. J. Ethnobiol. Ethnomed. 16:31. doi: 10.1186/s13002-020-00384-2
Prillo, V. G., Speciale, C., and Miccichè, R. (2024). “Insularity and singularity: exploitation of animal resources in Sicilian insular and inland territories during Prehistoric times,” in Case Studies in European Zooarchaeology, eds. T. Mladenovic and D. Markovic (Oxford: BAR Publishing), 27–43.
Prillo, V. G., Speciale, C., Miccichè, R., Arienzo, I., and Carandente, A. (2025). The faunal assemblage of Piano dei Cardoni: a Neolithic context on the island of Ustica (Italy). Archaeofauna 34, 205–212. doi: 10.15366/archaeofauna2025.34.1.020
Ramis, D. (2014). “Early island exploitations: productive and subsistence strategies on the prehistoric Balearic Islands,” in The Cambridge Prehistory of the Bronze and Iron Age Mediterranean, ed. A. B. Knapp and P. van Dommelen (Cambridge: Cambridge University Press), 44–56. doi: 10.1017/CHO9781139028387.006
Ramis, D. (2017). Animal exploitation in the early prehistory of the Balearic Islands. J. Island Coast. Archaeol. 13, 269–282. doi: 10.1080/15564894.2017.1334721
Rick, T. C., Kirch, P. V., Erlandson, J. M., and Fitzpatrick, S. M. (2013). Archeology, deep history, and the human transformation of island ecosystems. Anthropocene 4, 33–45. doi: 10.1016/j.ancene.2013.08.002
Sabato, D., and Peña-Chocarro, L. (2021). Maris Nostri Novus Atlas. Seeds and fruits from the Mediterranean Basin. Aranjuez: Ediciones Doce Calles.
Scerri, E. M. L., Blinkhorn, J., Groucutt, H. S., et al. (2025). Hunter-gatherer sea voyages extended to remotest Mediterranean islands. Nature 641, 137–143. doi: 10.1038/s41586-025-08780-y
Schicchi, R., Speciale, C., Amato, F., Bazan, G., Di Noto, G., Marino, P., et al. (2021). The monumental olive trees as biocultural heritage of Mediterranean landscapes: the case study of Sicily. Sustainability 13:6767. doi: 10.3390/su13126767
Schmid, E. (2022). Atlas of Animal Bones. For Prehistorians, Archaeologists and Quaternary Geologists. Amsterdam: Elsevier.
Schüle, W. (1993). Mammals, vegetation and the initial human settlement of the Mediterranean islands: a palaeoecological approach. J. Biogeogr. 20, 399–411. doi: 10.2307/2845588
Schweingruber, F. H. (1990). Anatomy of European Woods, 800. Swiss Federal Institute for Forest, Snow and Landscape Research, Birmensdorf.
Sheydaei, P., Amaral, M. E., and Duarte, A. P. (2025). Genus Echium L.: phytochemical characterization and bioactivity evaluation for drug discovery. Plants 14:2548. doi: 10.3390/plants14162548
Silver, I. A. (1969). “The ageing of domestic animals,” in Science in Archaeology, eds. D. Brothwell and E. S. Higgs (London: Thames and Hudson), 283–302.
Speciale, C. (2021). Human-Environment Dynamics in the Aeolian Islands during the Bronze Age: A Paleodemographic Model. Oxford: BAR International Series. doi: 10.30861/9781407357980
Speciale, C. (2024). Sicily and the process of Neolithisation: a review of the archaeobotanical data. Veget. Hist. Archaeobot. 33, 185–194. doi: 10.1007/s00334-023-00963-9
Speciale, C., Battaglia, G., Larosa, N., and Magrì, A. (2022). Piano dei Cardoni, Ustica (Palermo). Notiziario di Preistoria e Protostoria 8, 79–81.
Speciale, C., De Angelis, F., Magrì, A., Larosa, N., Micciché, R., Rickards, O., et al. (2024b). The Neolithic site of Piano dei Cardoni (Ustica, Palermo) and the chronology of Sicily between the mid-6th and the end of the 5th millennium BCE. Riv. Sci. Preistoriche 74:S4.
Speciale, C., D'Oronzo, C., Stellati, A., and Fiorentino, G. (2016). Ubi Minor… deinde summa?: archaeobotanical data from the prehistoric village of Filo Braccio (Filicudi, Aeolian Archipelago): spatial analysis, crop production and paleoclimate reconstruction. Scienze dell'Antichità 22, 281–296.
Speciale, C., Freund, K. P., de Vita, S., Larosa, N., Forgia, V., Battaglia, G., et al. (2021b). Obsidian from the site of Piano dei Cardoni, Ustica (Palermo, Italy): preliminary results on the first occupation of the Island. Open Archaeol. 7, 273–290. doi: 10.1515/opar-2020-0140
Speciale, C., Giannitrapani, E., Mercuri, A. M., Florenzano, A., Sadori, L., and Combourieu-Nebout, N. (2024a). The establishment of the agricultural landscape of central sicily between the middle neolithic and the beginning of the iron age. Hum. Ecol. 52, 229–253. doi: 10.1007/s10745-024-00496-3
Speciale, C., Larosa, N., Battaglia, G., and Vassallo, S. (2020). Piano dei Cardoni, Ustica (Palermo). Notiziario Istituto di Preistoria e Protostoria 6, 62–64.
Speciale, C., Larosa, N., Spatafora, F., Calascibetta, A. M. G., Di Sansebastiano, G. P., Battaglia, G., et al. (2023). Archaeobotanical and historical insights on some steps of forest cover disruption at Ustica Island (Sicily, Italy) from prehistory until present day. Environ. Archaeol. 28, 312–327. doi: 10.1080/14614103.2021.1962578
Speciale, C., Mentesana, R. B., Montana, G., Forgia, V., Mantia, F., Battaglia, G., et al. (2021a). Materials and tools across volcanoes: exploitation of georesources in piano dei cardoni (Ustica, Italy) during prehistory. Ann. Geophys. 64:VO552. doi: 10.4401/ag-8684
Stika, H. P., Neugebauer, A., Rihuete-Herrada, C., et al. (2024). Crop cultivation in the Talayotic settlement of Son Fornés (Mallorca, Spain): agricultural practices on the western Mediterranean islands in the first millennium BCE. Veget. Hist. Archaeobot. 33, 91–101. doi: 10.1007/s00334-023-00957-7
Tercerie, S., Bearez, P., Pruvost, P., Bailly, N., and Vignes-Lebbe, R. (2022). Osteobase. World Wide Web electronic publication. osteobase.mnhn.fr.
Trantalidou, K. (2008). “Glimpses of Aegean island communities during the Mesolithic and Neolithic periods: the zooarchaeological point of view,” in A Colloquium on the Prehistory of the Cyclades (Cambridge: McDonald Institute Monographs), 19–27.
Valenzuela, A. (2023). The gymnesic islands at the crossroads: livestock body size as marker of social interactions during the first millennium BC. Quat. Int. 662, 130–142. doi: 10.1016/j.quaint.2022.08.006
Valenzuela-Suau, L., Bernad, D. R., and Valenzuela-Lamas, S. (2023). New contributions to understand animal size fluctuations in the western Mediterranean: the bronze age Balearic Islands. Quat. Int. 662, 117–129. doi: 10.1016/j.quaint.2023.04.009
Varalli, A., Moggi-Cecchi, J., and Goude, G. (2022). A multi-proxy bioarchaeological approach reveals new trends in Bronze Age diet in Italy. Sci. Rep. 12:12203. doi: 10.1038/s41598-022-15581-0
Villari, P. (1996). Le faune della tarda preistoria della Sicilia orientale. Siracusa: Ente fauna siciliana.
Wilkens, B. (1987). La Fauna dell'età del Bronzo di Mursia. Nota preliminare Atti Società Toscoscana Scienze Naurali. Mem. Serie A 94, 215–224.
Keywords: Neolithic, Bronze Age, Sicily, volcanic islands, subsistence system
Citation: Speciale C, Carra M, Fiori F, Prillo VG, Allué E and Cattani M (2025) Adapting to Mediterranean island environments: prehistoric human interaction with plants and animals at Piano dei Cardoni (Ustica) and Mursia (Pantelleria). Front. Environ. Archaeol. 4:1621064. doi: 10.3389/fearc.2025.1621064
Received: 30 April 2025; Accepted: 16 October 2025;
Published: 20 November 2025.
Edited by:
Yaowu Hu, Fudan University, ChinaReviewed by:
Jacob Morales, University of Las Palmas de Gran Canaria, SpainSuembikya Frumin, Bar-Ilan University, Israel
Copyright © 2025 Speciale, Carra, Fiori, Prillo, Allué and Cattani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Claudia Speciale, Y2xhdWRpYXNwZWNpYWxlQGdtYWlsLmNvbQ==