 María Laura Raymond Eder
María Laura Raymond Eder Laura Fariña
Laura Fariña Francisco Carrau
Francisco Carrau Alberto Luis Rosa
Alberto Luis Rosa- 1Laboratorio de Genética y Biología Celular y Molecular, Departamento de Farmacología Otto Orsingher, Facultad de Ciencias Químicas, Universidad Nacional de Córdoba, Córdoba, Argentina
- 2Departamento de Agroalimentos, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba, Córdoba, Argentina
- 3Área Enología y Biotecnología de Fermentaciones, Facultad de Química, Universidad de la República, Montevideo, Uruguay
- 4Laboratorio de Biotecnología de Aromas, Facultad de Química, Universidad de la República, Montevideo, Uruguay
- 5Instituto de Farmacología Experimental de Córdoba (IFEC-CONICET), Córdoba, Argentina
Spontaneously fermenting grape juices represent complex ecosystems resulting from the dynamic interaction between the unique characteristics of a grape varietal and its indigenous associated microbiota. The extent to which specific grape variety volatile compounds versus microbially derived ones shape wine identity remains incompletely understood. In this work, we explored this issue by characterizing the volatile compound profiles at early stages of fermentation of the highly aromatic Isabella (V. labrusca L.) grape juice, conducted by native microbial communities prepared from either Isabella (homologous fermentation) or Malbec (V. vinifera L., heterologous fermentation) grapes. Results revealed that microbial starters derived from V. labrusca L. and V. vinifera L. markedly influenced the volatile profiles of the resulting fermented Isabella grape juices. Joint analysis of volatile profiles from Malbec and Isabella juices fermented with the same set of Vitis-specific microbial communities showed that, despite the strong influence of the microbial consortia, the fermented juices retained traits consistent with their original grape varietal identity. Characterization and identification of cultivable yeast species in these homologous and heterologous fermentations of Isabella grape juice showed H. uvarum, H. opuntiae, and S. bacillaris as dominant species in Malbec and Isabella microbial ecosystems. Our results highlight the potential of this innovative experimental approach to examine the relative roles of microbial communities and grape varietals in shaping wine identity. Moreover, they show that different Vitis-specific microbiota can distinctly influence the volatile profiles of a fermenting grape juice without altering its varietal identity.
1 Introduction
Wine fermentation is a complex process where the interplay between grape juice and its associated microbial populations, derived from both the vineyard and winery environments, shapes the chemical and sensory profiles of the resulting wine (Belda et al., 2017; de Celis et al., 2022; García-Izquierdo et al., 2024). Yeasts are particularly important in alcoholic fermentation, where they convert sugars into alcohol and carbon dioxide, while also producing various secondary metabolites that contribute to the wine’s flavor and aroma (Maicas and Mateo, 2023; Wang et al., 2023). The indigenous yeast communities that develop during spontaneous grape must fermentations are shaped by annual environmental conditions in the vineyard, agricultural practices, intrinsic factors of the grapevine, —including the physicochemical properties of the grape must, — and winemaking techniques (Varela and Borneman, 2016; Griggs et al., 2021; Liszkowska and Berlowska, 2021; de Celis et al., 2022; Maicas and Mateo, 2023). A consistent pattern of species emerges in the yeast population: non-Saccharomyces yeasts dominate the initial phase of fermentation, while Saccharomyces cerevisiae becomes the predominant species as fermentation progresses (Bezerra-Bussoli et al., 2013; Drumonde-Neves et al., 2016; Cilião Filho et al., 2017; Raymond Eder et al., 2017, 2018; Borren and Tian, 2021; de Celis et al., 2022; Torres-Guardado et al., 2022; Maicas and Mateo, 2023). Non-Saccharomyces yeasts, which are predominant on grape skins, are introduced into the must during grape crushing and play a significant role in the early fermentation stages, producing various secondary metabolites that strongly impact the organoleptic characteristics of wine (Kamilari et al., 2021; Morata et al., 2021; Tufariello et al., 2021; Romano et al., 2022; Torres-Guardado et al., 2022; Maicas and Mateo, 2023; Wang et al., 2023).
Understanding the dynamics of indigenous microbial communities of fermenting grape juices and musts is a central focus in enology, given its significance for both scientific research and industrial applications (Padilla et al., 2016; Varela, 2016; Griggs et al., 2021; de Celis et al., 2022; Maicas and Mateo, 2023; García-Izquierdo et al., 2024). An increasing number of studies point to a relationship between the grape microbiome and terroir, which includes factors such as soil composition, climate, and annual precipitation, suggesting that specific microbial populations may contribute to the regional identity of wines (Pretorius, 2017, 2020; Liu et al., 2020; Griggs et al., 2021; Kamilari et al., 2021; Zhang et al., 2023a). Moreover, it has been suggested that particular grape varieties harbor microbiomes involving specific yeast strains or species, influencing the unique characteristics of fermented grape juices (Baffi et al., 2011; Bezerra-Bussoli et al., 2013; Lederer et al., 2013; Raymond Eder et al., 2017, 2018, 2025; Raymond Eder and Rosa, 2019). We have recently suggested that Vitis-specific microbial communities play a critical role in shaping the identity of grape juice fermentations (Raymond Eder et al., 2025). However, it remains unclear how strongly a grape varietal–specific indigenous microbial community influences the final organoleptic properties of a given wine, how terroir-associated microbial signatures contribute to the wine’s identity and profile, and to what extent alternative microbiotas—such as those from different vintages or experimentally introduced from other Vitis species (Raymond Eder et al., 2025)—can alter the fermentation profile of a given grape varietal.
Although the OIV (International Organization of Vine and Wine) primarily recognizes Vitis vinifera L. as the species designated for vinification, other Vitis species and their hybrids are also used in winemaking, particularly in non-European countries (De la Fuente Lloreda, 2018). These non-vinifera Vitis species represent underexplored microbial ecosystems in enology and may serve as a potential source of yeasts with unique fermentative properties of both academic and industrial relevance (Drumonde-Neves et al., 2016; Raymond Eder et al., 2017, 2018; Raymond Eder and Rosa, 2019). Moreover, we have recently proposed that they can serve as powerful tools for studying the impact of indigenous microbiota on the aromatic profiles of fermenting grape juices (Raymond Eder et al., 2025). Building on the use of this innovative experimental system, in this study we investigate the impact of the indigenous microbiota of Malbec grapes—a conventional V. vinifera L. varietal used in winemaking—versus the native microbiota of Isabella grapes (V. labrusca L.), on the volatile profile at early stages of fermentation of Isabella grape juices.
2 Materials and methods
2.1 Grape juice fermentations
Grapes from Isabella (Vitis labrusca L.) and Malbec (Vitis vinifera L.) were harvested from two closely located, small vineyards (i.e., ~1.5 Ha each), with intermixed rows of both varietals (Raymond Eder et al., 2018), in Colonia Caroya, Argentina, during the 2021 vintage. Isabella and Malbec grape juice supernatants (~3.5 l each), collected after centrifugation of filtered musts (5000 rpm for 10 min at 20°C), were pasteurized (60°C, 30 min) (Malletroit et al., 1991; Englezos et al., 2018; Raymond Eder et al., 2025). The corresponding Isabella (I) and Malbec (M) sedimented fractions, which contain their native microbial communities (i.e., Imc and Mmc, respectively) were each suspended into 400 ml of pasteurized Isabella grape juice (Igj) to reconstitute homologous (Igj/Imc) or heterologous (Igj/Mmc) fermenting ecosystems (Raymond Eder et al., 2025). Fermentations were performed in triplicate for each condition (Igj/Imc and Igj/Mmc) in 500 ml Erlenmeyer flasks, sealed with air locks, without agitation, at 25°C. Aliquots were collected from 0 to 96 hours (T0 to T96) for volatile, physicochemical, and microbial analyses, as previously described (Raymond Eder et al., 2025). Figure 1 presents a schematic overview of the experimental design.

Figure 1. Experimental design flow-chart. 10 kg of Isabella and Malbec grapes were crushed, filtered, and the juices were centrifuged (5000 rpm, 10 min). The supernatant Isabella grape juice (Igj) was divided into 380 ml aliquots and pasteurized (60°C, 30 min). The Isabella (Imc) and Malbec (Mmc) sediments were resuspended in pasteurized Isabella juice and used to inoculate the pasteurized Igj. Triplicate fermentations were conducted for each Igj/Imc and Igj/Mmc at 25°C without agitation in Erlenmeyer flasks with air locks. Samples were taken every 24h from T0-T144. Appropriate dilutions of T0, T48, and T96 samples were plated and after incubation for 4 days at 25°C, yeast colonies were counted, isolated and identified. At T0 and T96 volatile compound levels (µg/l) were assessed by GC-MS.
2.2 Volatile compounds identification and quantification
Volatile compounds were extracted by HS-SPME-GC-MS using an automatic injector AOAC-6000 Shimadzu, according to the methodology exposed by Garde-Cerdán et al. (2018). GC-MS analyses were conducted using a Shimadzu GC-20 plus gas chromatograph coupled to a Shimadzu QP 2020 mass spectrometer with a DW-Wax 30 (Agilent Technologies J&W, Santa Clara, CA, USA) bonded fused silica capillary column, coated with poly(ethylene glycol). The experimental conditions were performed according to Dellacassa et al. (2017) as follows: column temperature, 40°C for 8 min, rising to 180°C at 3°C/min, then to 230°C at 20°C/min; injector temperature, 250°C; detector temperature, 250°C; injection mode: splitless (2 min); carrier gas, hydrogen, 30 kPa. GC-MS instrumental procedures, using an internal standard (1‐heptanol), were applied for quantitative purposes (Dellacassa et al., 2017). Volatile compounds were identified by comparison of Kovats indices (KI, Table 1; Supplementary Table S1).

Table 1. Volatile compounds in Igj at T0 and in Igj/Imc and Igj/Mmc at T96.
An ANOVA was conducted on the chemical and volatile compounds data obtained for the different treatments and replicas. Principal Component Analysis (PCA) was performed using InfoStat (InfoStat, FCA, Universidad Nacional de Córdoba, Argentina) to differentiate between samples and identify contributing compounds. A Hierarchical Cluster Analysis (HCA) was performed using Euclidean distances calculated from the average volatile profiles of each fermenting condition. Clustering was performed using the complete linkage method. The resulting dendrogram was constructed using base R functions (dist and hclust) and visualized with the dendextend package for enhanced customization.
2.3 Yeast identification from Isabella fermentations
Yeasts were isolated from Isabella homologous and heterologous fermentations at different time points (T0, T48, and T96 h) by plating pooled samples on YPD-Cm [yeast extract 1.0% (w/v), peptone 2.0% (w/v), glucose 2.0% (w/v), agar 2.0% (w/v), chloramphenicol 10 µg/ml] and YPD-Cm-Cx agar [yeast extract 1% (w/v), peptone 2% (w/v), glucose 2% (w/v), agar 2% (w/v), chloramphenicol 10 µg/ml, cycloheximide 0.5 µg/ml] to estimate total and non-Saccharomyces populations, respectively (Raymond Eder et al., 2025). Predominant yeast species were randomly selected from high-dilution YPD-Cm plates using a grid-based method to ensure unbiased isolation (Raymond Eder et al., 2017). Colonies were purified, stored in glycerol stocks, and subsequently identified by PCR-RFLP and/or sequencing of the 5.8S-ITS rDNA region. Restriction profiles were generated using Hinf I and CfoI, and representative isolates were confirmed by Sanger sequencing, with species assignment based on ≥99% identity to reference sequences in NCBI BLAST. Identified yeast strains were deposited in GenBank (NCBI) under the accession numbers OP584257, OP584258, OP584259, OP584260, OP584261, OP584262, OP584263, and OP584265.
3 Results
3.1 Physicochemical and volatile analyses of Isabella grape juice
High nitrogen compounds levels, such as α Amino (171.1 ± 4.3 mg/l), NH4+ (77.8 ± 4.2 mg/l), and yeast assimilable nitrogen-YAN- (235.5 ± 7.8 mg/l) were found in the Igj, as well as a total acidity value of 2.9 ± 0.0 g/l, a density of 1.083 ± 0.001 g/ml, and 19.1 ± 0.4°Brix. Volatile compounds in the Igj at T0 were analyzed using GC-MS (Table 1; Supplementary Table S1). The detected compounds exclude bound volatiles and potential artifacts arising from enzymatic treatments used for their release, thus accurately reflecting the aroma-active profile of a fresh grape juice (Raymond Eder et al., 2025). The recognized profile of Igj is distinguished by the presence of the two correlated aroma terpenes α-terpineol/linalool, a characteristic marker of the Isabella varietal, along with a high proportion of alcohols, particularly hexanol, phenethyl alcohol, isoamyl alcohol, and 1-nonanol, followed by some ethyl esters of medium chain fatty acids (Table 1; Supplementary Table S1). Ethyl 3-hydroxybutyrate and methyl anthranilate, previously identified as free volatile aroma compounds in Isabella grape samples (Ghaste et al., 2015), were also recognized in the analyzed grape juices. In particular, methyl anthranilate is associated with the perception of foxiness in V. labrusca L. grapes (Raymond Eder and Rosa, 2019).
3.2 Isabella grape juice fermentations
Indigenous microbial communities from Isabella (Imc) and Malbec (Mmc) grapes were prepared as indicated (Raymond Eder et al., 2025) (see also Figure 1). As recently reported, the centrifugation process enabled effective collection of the cultivable yeast community; additionally, the pasteurized grape juices showed no detectable presence of cultivable yeasts (i.e., <10 CFU/ml) (Raymond Eder et al., 2025). The mild pasteurization applied (i.e., 60°C for 30 minutes) is much less intense than other processes studied (i.e., 81.5 ± 0.5°C for 450 min) (Malletroit et al., 1991), preserving the sensory profile of the grape juice (Malletroit et al., 1991; Raymond Eder et al., 2025).
The pasteurized Igj was inoculated with Imc and Mmc and their effect on the volatile profiles of fermenting Igj were evaluated from both homologous (Igj/Imc) and heterologous (Igj/Mmc) fermentations. After four days of fermentation (i.e., T96), a total of forty-three compounds, including acetates, alcohols, esters, and terpenes, were identified and quantified in Igj (Table 1; Supplementary Table S1). Significant differences in the overall volatile compound concentrations were observed between Igj/Imc T0 and T96 fermentations (Figure 2; Table 1; Supplementary Table S1). The varietal compounds (E)-3-hexenyl acetate, heptyl acetate, ethyl 2-butenoate, ethyl 2-hexenoate, identified in Igj T0 samples, were found in lower concentrations at T96. At this fermenting time, the most abundant compounds in both conditions were isoamyl alcohol, phenethyl alcohol, isoamyl acetate, phenethyl acetate, ethyl octanoate, ethyl decanoate, and ethyl dodecanoate (Table 1). The compounds ethyl lactate, ethyl hexadecanoate, 2-nonanol, (E,E)-farnesol, 2-heptanone, and acetoin were identified only at T96. The terpenic and C6 compounds identified in Isabella grape juices showed no significant differences between fermenting times (i.e., T0 and T96; Figure 2). Significant differences were found for ethyl octanoate, ethyl decanoate, phenethyl alcohol, phenethyl acetate, and ethyl phenylacetate, which were quantified in higher concentrations in the homologous fermentations (Table 1; Supplementary Table S1; Figure 2).

Figure 2. Concentration of volatile compounds (µg/l) determined by GC-MS in Isabella grape juice (T0) and after 96 h of fermentation using Imc (T96 Imc) and Mmc (T96 Mmc) as inocula. Total ethyl ester (A), acetate ester (B), higher alcohol (C), and volatile (D) compound concentrations (see Table 1). Values are means ± SD (Table 1; Supplementary Table S1). Significant differences (p <0.05) are marked with an asterisk *.
The volatile compound profiles of fermented Igj using the alternative Imc and Mmc starters were further analyzed through Principal Component Analysis (PCA). Figure 3 illustrates that PC1 accounted for 49.0% of the variance, while PC2 accounted for 27.4%. This analysis demonstrated a distinct separation between fermented (i.e., T96) and non-fermented (i.e., T0) grape juices. Moreover, among the fermented grape juices, the analysis revealed clustering based on the starter used for fermentation (Figure 3). In Figure 3, Igj/Mmc fermentations are located in the negative values of PC1, whereas Igj/Imc samples cluster together in the positive values of PC1, primarily associated with ethyl esters, which are volatile aroma compounds found in significantly higher quantities under this fermenting condition.

Figure 3. Principal Component Analysis (PCA) performed on the concentration of the volatile compounds identified in the Isabella grape juice at T0 and the Igj/Imc and Igj/Mmc fermenting conditions at T96 (Table 1). Representation of the volatile compounds, the Isabella grape juice at T0 (i.e., IgjT0), and three replicas (i.e., A, B, C) of each fermenting condition using Imc (i.e., Igj/ImcA, Igj/ImcB, Igj/ImcC) or Mmc (i.e., Igj/MmcA, Igj/MmcB, Igj/MmcC) at T96 in the two first components (PC1 and PC2).
3.3 Impact of homologous and heterologous microbial starters on the volatile profiles of Malbec and Isabella grape juice
To evaluate the relative influence of the fermentation inoculum and the fermenting grape juice on volatile profiles, we conducted a PCA integrating the volatile profile data obtained in this study from Igj/Imc and Igj/Mmc fermentations at T96 with previously published data from Malbec grape juices (Mgj) (Raymond Eder et al., 2025), fermented using the identical Imc and Mmc inocula as in the present work. All Isabella and Malbec samples were prepared from grapes harvested in the same vintage and processed and fermented in parallel, which enables proper comparison of previous and current datasets (see Materials and Methods section) (Raymond Eder et al., 2025). Figure 4 shows that PC1 counted for 46.2%, while PC2 for the 29.9% of the variance. The PC1 showed a separation between the grape juice used for fermentation, with Igj (this work) and Mgj (Raymond Eder et al., 2025) locating at positive and negative values of the graph, respectively. An ANOVA performed on the miscellaneous varietal compounds (i.e., acetophenone, methyl salicylate, 3,5-dimethyl benzaldehyde, (z)-methyl cinnamate, and methyl anthranilate; Supplementary Table S2), revealed significant differences among the homologous fermentations (i.e., Igj/Imc and Mgj/Mmc). However, similar concentrations of these compounds were found in Isabella and Malbec fermentations performed with the heterologous microbial communities (i.e., Igj/Mmc and Mgj/Imc; Supplementary Table S2), suggesting that these communities reduce the varietal variability in these fermentations. Regarding the impact of the microbial communities in the volatile profiles, a differentiation among the microbial communities, although to a limited extent in the Igj fermentations, is observed in the PC2. In general, the Imc replicas were positioned in positive values, while Mmc were positioned in negative values in the analysis. To further analyze these results, a hierarchical cluster analysis (HCA) based on Euclidean distances between the average volatile profiles of each fermenting condition was performed and plotted (Supplementary Table S3; Supplementary Figure S1). The resulting dendrogram shows that the volatile profiles of fermenting grape juices cluster primarily according to the microbial community used as inoculum. Specifically, samples inoculated with the Malbec microbial community (Mmc) cluster together (Mgj/Mmc and Igj/Mmc), regardless of grape variety, indicating a strong shaping effect of this community. In contrast, the two samples inoculated with the Isabella microbial community (Imc) are more distant from each other, with Mgj/Imc appearing as the most distinct condition overall. These results suggest that the microbial community has a dominant influence on the volatile profile, but the interaction with grape varieties also contributes to the observed variability.
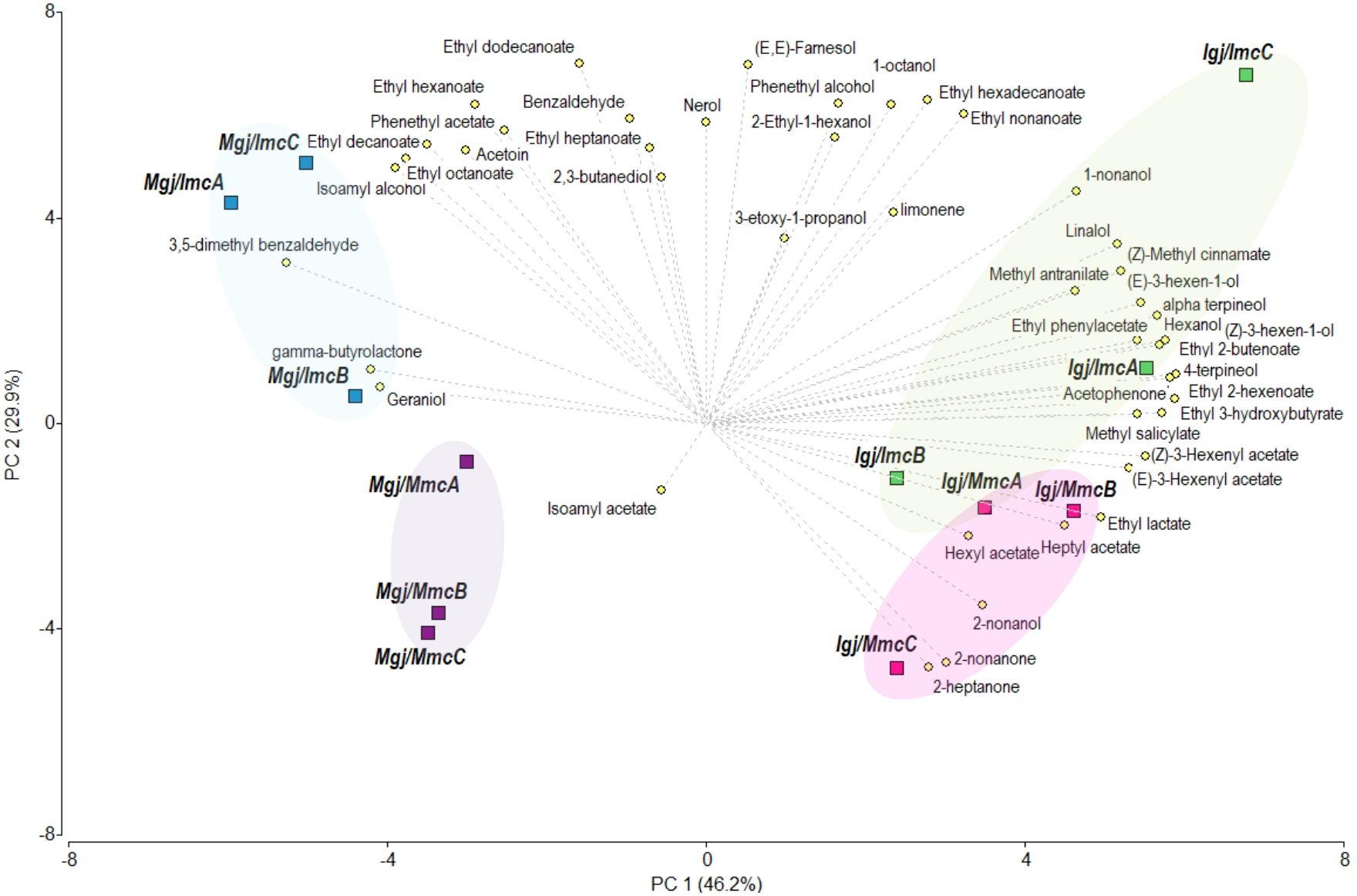
Figure 4. Principal Component Analysis (PCA) performed on the concentration of the volatile compounds identified in the Igj/Imc and Igj/Mmc fermenting conditions at T96 and the Mgj/Mmc and Mgj/Imc fermenting conditions at T96 reported in (Raymond Eder et al., 2025) (Supplementary Table S2). Representation of the volatile compounds and the three replicas (i.e., A, B, C) of each fermenting condition at T96 in the two first components (PC1 and PC2). Compounds that were not identified at T96 and compounds that did not increase their concentration from T0 to T96 were not considered in this analysis.
A factorial ANOVA was performed in order to evaluate the impact of the analyzed factors (i.e., grape juice -either Igj or Mgj- and inoculum -either Imc or Mmc-) and their interactions in the volatile profile variabilities. This analysis revealed that there were no significant effects from the interactions between the two factors for any of the volatile compounds identified (Supplementary Table S2). The grape juice and inoculum, however, significantly impacted 30 and 16 volatile compounds, respectively. The compounds phenethyl alcohol, phenethyl acetate, ethyl nonanoate, ethyl decanoate, ethyl hexadecanoate, and (E,E)-farnesol were the only compounds significantly impacted only by the inoculum. Interestingly, phenethyl acetate, ethyl decanoate, and ethyl hexadecanoate were present in significantly higher concentrations in fermentations performed using Imc (Supplementary Table S2).
These analyses reveal that alternative microbial communities (i.e., Imc or Mmc) can shape the profile of volatile compounds in ways that allow differentiation between fermentation conditions, while the fermenting grape juices preserve the aromatic typicity and varietal identity of each cultivar (Figure 4).
3.4 Yeast communities in fermenting Isabella grape juices
Differential CFU/ml counts obtained from YPD-Cm and YPD-Cm-Cx agar plates (Raymond Eder et al., 2017, 2018) were used to assess the contributions of total yeasts and non-Saccharomyces yeasts, respectively. Results from this analysis revealed that both conditions (i.e., Igj/Imc and Igj/Mmc) started with similar counts (~1–2 x 106 CFU/ml), with non-Saccharomyces species predominating during the early stages of fermentation (T0 to T96) (Figure 5; Supplementary Figure S2). S. cerevisiae started to become dominant at middle stages of fermentation (i.e., T144; not shown). Based on these results, we focused our analyses of non-Saccharomyces species present at early stages of fermentations (i.e., T0 to T96). A total of 140 isolates from Igj/Imc and Igj/Mmc were isolated and identified (Supplementary Figure S2). Hanseniaspora opuntiae and Hanseniaspora uvarum were the most common species at early stages of Igj/Imc and Igj/Mmc fermentations (Supplementary Figure S2), followed by Starmerella bacillaris. Similar results have been obtained when analyzing grape-associated indigenous yeast communities from this ecosystem (Raymond Eder et al., 2017, 2018, 2025; Raymond Eder and Rosa, 2019). Even if similarities were found among the main recognized yeast species in the homologous and heterologous fermenting samples (i.e., Igj/Imc and Mgj/Imc, respectively), differences in the relative proportion of these yeasts were observed at the analyzed times (Figure 3). Also, Igj/Imc fermentations at T96 revealed the presence of Torulaspora delbrueckii, Hanseniaspora vineae, and Pichia terricola as representative isolates (Figure 6). This greater yeast biodiversity, as well as changes in the relative contribution of the main yeast species identified in the fermentations, could help explain the variations observed in the evolving volatile profiles of Igj/Imc replicates (Figure 3).
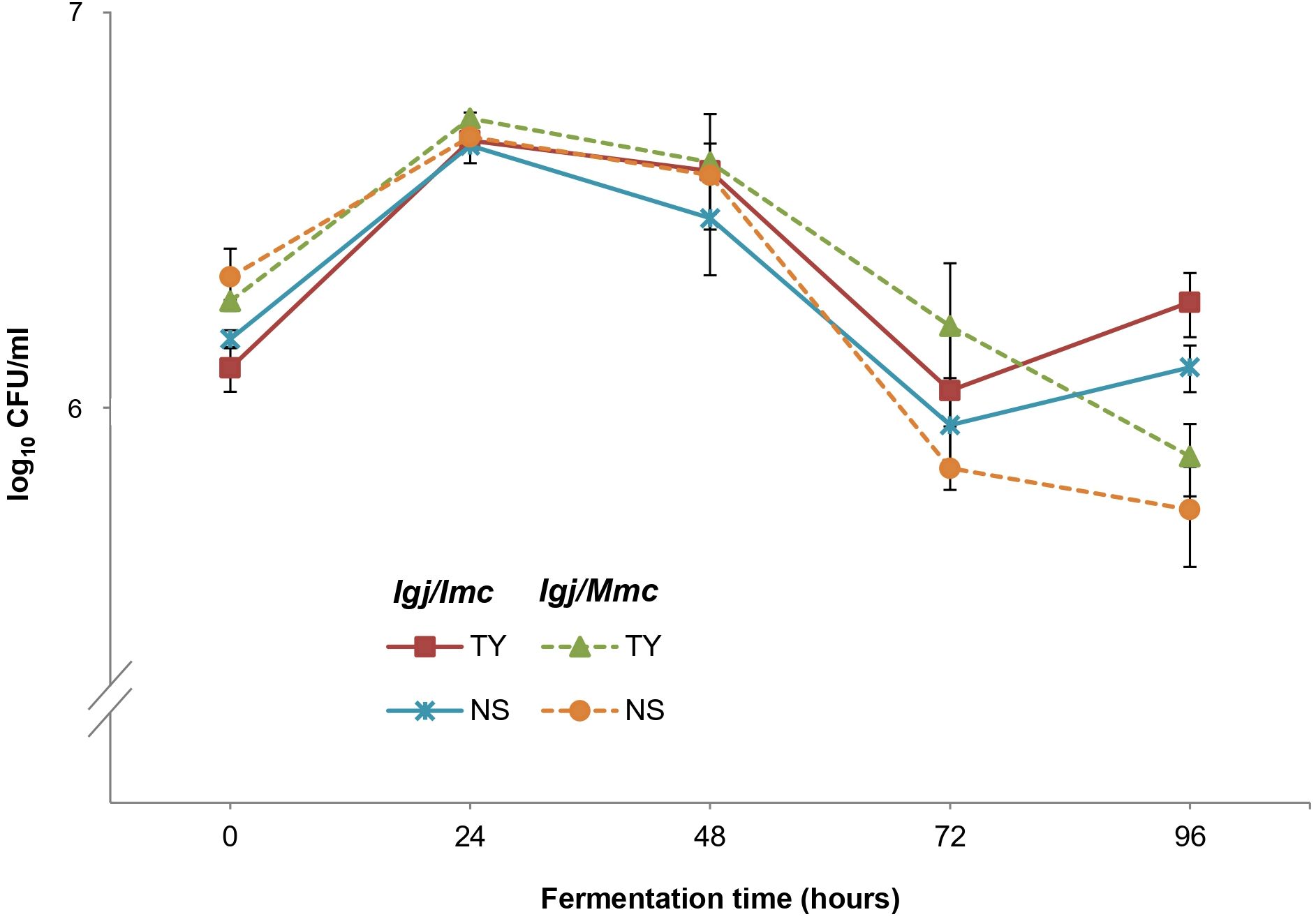
Figure 5. Population dynamics of non-Saccharomyces (NS) and total yeasts (TY) at initial stages of fermentations (0 to 96 h) of Isabella grape juice (Igj) using Isabella (Imc) or Malbec (Mmc) microbial communities as inocula. Each point represents log10 CFU/ml ± SD.

Figure 6. Main represented yeast species at T96 of the three replicas from each fermenting condition. Appropriate dilutions of replicas A, B, and C from Imc (left; columns 2–4) and Mmc (right; columns 6–8) conditions were plated individually, and percentages represent the relative contribution of the indicated yeast species.
4 Discussion
Varietal or primary wine flavors originate from the grape variety itself. In addition to the volatile compounds naturally present in grape juice, secondary flavors are developed through alcoholic fermentation via yeast metabolism, which produces higher alcohols, esters, and fatty acids (Carrau et al., 2020; Borren and Tian, 2021; Zilelidou and Nisiotou, 2021; Romano et al., 2022; Liang et al., 2023; Wang et al., 2023; Zhang et al., 2023a). In spontaneous fermentations, the complex native microbial community associated to grapes contributes to the final volatile profiles of wines (Padilla et al., 2016; Belda et al., 2017). Indigenous grape-associated yeasts species are particularly relevant contributors to the volatile profiles of fermenting juices at early times of fermentation (Jolly et al., 2014; Pretorius, 2017; Raymond Eder et al., 2025). In addition to yeast, lactic-acid and acetic-acid bacteria, even when present at low concentrations at early times of fermentations, can also contribute to the final volatile compositions of spontaneous fermentations (Barata et al., 2012; Andorrà et al., 2019; Shimizu et al., 2023; Raymond Eder et al., 2025).
We have recently demonstrated that microbial communities associated to different Vitis species condition the volatile profiles of early-stage Malbec grape juice fermentations (Raymond Eder et al., 2025). In this work, we have extended these studies to analyze the volatile profiles of Isabella grape juices fermented using homologous (Isabella; V. labrusca L.) or heterologous (i.e., Malbec; V. vinifera L.) native microbial communities. Our analyses consider the profiles of free volatile compounds in grape juices, excluding glycoside-bound volatiles (Raymond Eder et al., 2025). Although bound volatiles contribute to the full aromatic potential of a grape juice —upon enzymatic or fermentative release— the analyzed free fraction offers a representative expression of varietal identity at the juice stage. Moreover, this approach avoids potential artifacts introduced by exogenous enzymes, yields reproducible data under our standardized conditions, and reveals the aromatic identity of fresh varietal grape juice. This experimental strategy allowed us to perform a joint analysis of the volatile compound datasets obtained from homologous and heterologous fermentations of Malbec (Raymond Eder et al., 2025) and Isabella (this work) grape juices, conducted with the same starter microbial communities and grape juices samples prepared from grapes harvested in vintage of year 2021 (Raymond Eder et al., 2025). Taken together, the results presented in this work show that the microbiota shapes the volatile profiles of the fermented Malbec and Isabella juices, which nonetheless retain the identity of their original grape varietal.
It has been reported that Isabella, a varietal from the American-originated Vitis labrusca L., is rich in aroma compounds both qualitatively and quantitatively (Ghaste et al., 2015). Key alcohols and terpenes, including methyl anthranilate, β-phenyl ethanol, ethyl-3-hydroxybutyrate, ethyl-β-hydroxy hexanoate, furaneol, phenylacetaldehyde, tryptophol, and 2-hexenol, have been detected in Isabella grape juices and crushed grapes originating from Brazil and Italy, respectively (Ghaste et al., 2015; Gomez et al., 2024). Our analysis revealed that Isabella juice from grapes harvested in the geographic region of Córdoba (Argentina) has a substantial proportion of alcohols, esters, and terpenes, including the previously reported methyl anthranilate, β-phenyl ethanol, ethyl-3-hydroxybutyrate, and methyl salicylate (Ghaste et al., 2015; Gomez et al., 2024). Although different techniques of extraction of volatile compounds were used, Isabella juice from Córdoba was found to contain eleven ethyl esters, whereas only two (i.e., ethyl-butanoate and ethyl-metacrylate) and four (i.e., methyl salicylate, methyl anthranilate, methyl-β-hydroxybutyrate, ethyl-3-hydroxyhexanoate) ethyl esters were detected in the Brazilian and Italian studies, respectively (Ghaste et al., 2015; Gomez et al., 2024). Previous research on the volatile profiles of Isabella and Ives commercial wines also emphasized the prominent role of ethyl acetate and esters, contributing to their characteristic fruity aroma descriptors (Arcanjo et al., 2018). In the present study, isoamyl alcohol, phenethyl alcohol, isoamyl acetate, phenethyl acetate, ethyl octanoate, ethyl decanoate, and ethyl dodecanoate emerged as the dominant volatile compounds during the initial stages of fermentation. Interestingly, as it was observed in our recent study of homologous and heterologous fermentation of Malbec grape juice (Raymond Eder et al., 2025), differences were observed in the volatile profiles of the Isabella fermenting grape juice when using the alternative either Malbec or Isabella associated microbial communities as starters. Homologous fermentations (Igj/Imc) show higher concentrations of ethyl octanoate, ethyl decanoate, phenethyl alcohol, phenethyl acetate, and ethyl phenylacetate than the heterologous conditions (Igj/Mmc) (Raymond Eder et al., 2025).
The evolution of culturable yeast populations in homologous and heterologous Isabella grape juice fermentations was studied using culture-dependent methods. Although his strategy limits our study to the identification of only cultivable yeast species from the overall fungal, yeast, and bacterial biodiversity in the samples, it is known that cultivable non-Saccharomyces yeast species significantly contribute to aromatic profiles in early fermentation stages (Andorrà et al., 2010; Borren and Tian, 2021; Zilelidou and Nisiotou, 2021; Romano et al., 2022; de Celis et al., 2024). At T0, Hanseniaspora spp. (H. opuntiae and H. uvarum) and, to a lesser extent, S. bacillaris, predominated in the fermented juices, consistent with previous studies on yeast communities from this region’s spontaneously fermenting musts and reconstituted grape juices (Raymond Eder et al., 2017, 2018, 2025). As fermentation progressed, from T0 to T96, Hanseniaspora spp. dominated the fermentations in Imc, while S. bacillaris increased its contribution in Mmc. It has been recently reported that the dominating yeast species in a fermentation defines its performance and metabolite profile of the resulting wines (de Celis et al., 2024). In this sense, members of the Hanseniaspora genus have been reported to play a significant role in the production of volatile compounds in wine, particularly acetate esters, in a strain-dependent manner (Jolly et al., 2014; Martin et al., 2018). Specifically, H. uvarum significantly increased ethyl and isoamyl esters and reduced acetate ester concentrations in co-fermentations with S. cerevisiae in a strain-specific way (Tristezza et al., 2016; Hu et al., 2018b, 2018a; Zhang et al., 2023b). H. opuntiae has been shown to positively influence volatile profiles and sensory characteristics of fermentations, significantly increasing acetate esters, mainly 2-phenylethyl acetate, and ethyl esters levels (Hu et al., 2020; del Fresno et al., 2022, 2023; Filippousi et al., 2024). Moreover, fermentation studies where Hanseniaspora spp. were the dominating yeasts, were characterized by higher fusel alcohol acetates production (de Celis et al., 2024). (i.e., phenethyl alcohol). On the other hand, S. bacillaris has been reported to overproduce ethyl and other acetate esters, as well as terpenes (Andorrà et al., 2010; Englezos et al., 2015; Raymond Eder and Rosa, 2021). However, elevated levels of these compounds were not detected in the Igj/Mmc fermentation, where this yeast was dominant at T96. Four other non-Saccharomyces species previously recognized in this terroir (i.e., Candida diversa, T. delbrueckii, H. vineae, and P. terricola) were also isolated at these fermentations. Proposed signature V. labrusca L. non-Saccharomyces species (i.e., Candida azymoides, Pichia cecembensis, and Candida californica) (Drumonde-Neves et al., 2016; Raymond Eder et al., 2017, 2018) were not identified among the limited number of isolates characterized in this study. In addition to the contribution to the volatile profiles of the main yeast species, increased yeast diversity has been linked to greater flavor complexity (Carrau and Henschke, 2021). In this context, the minor yeast species present at lower abundances in Igj/Imc may have also contributed to the elevated concentrations of certain volatile compounds. T. delbrueckii, for instance, enhances succinic acid, linalool, acetate esters, medium chain fatty acids, and terpenes in aromatic grapes (Jolly et al., 2014; Varela and Borneman, 2016). Similarly, P. terricola has been reported to release precursors from grape juice, boosting free monoterpenes and norisoprenoids (Jolly et al., 2014). The apiculate yeast H. vineae has been reported to produce terpenes, sesquiterpenes, and high amounts of acetate esters, such as 2-phenylethyl acetate and ethyl acetate, which influence fermentations by producing flavors and increasing sensory complexity (Martin et al., 2018).
Our results show that microbial communities assembled in different Vitis species influence the volatile profiles of early-stage fermentations of different grape juices. Additional data from fermentations using other microbial communities and grape juice varietals would strengthen the conclusion that alternative yeast populations shape the volatile profiles of fermented juice while preserving grape varietal identity. Also, grape fermentations using pure cultures of selected non-Saccharomyces strains, and/or pools of selected strains that resemble native microbial communities, could further clarify the specific contributions of individual yeast species and strains to the final volatile profiles of fermented grape juices. Emerging evidence suggest that non-conventional Vitis ecosystems may carry unique yeast species or strains not found in V. vinifera L., presenting opportunities for the isolation of valuable Saccharomyces and non-Saccharomyces strains of potential relevance for the winemaking industry.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: NCBI GenBank, accession OP584257, OP584258, OP584259, OP584260, OP584261, OP584262, OP584263, and OP584265.
Author contributions
MR: Investigation, Writing – original draft, Conceptualization, Writing – review & editing, Methodology, Validation, Formal analysis, Project administration, Supervision. LF: Software, Investigation, Writing – review & editing, Formal Analysis. FC: Supervision, Writing – review & editing, Conceptualization. AR: Formal analysis, Validation, Methodology, Project administration, Resources, Supervision, Investigation, Writing – review & editing, Conceptualization, Funding acquisition, Writing – original draft.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
We thank S. Lauret (Colonia Caroya, Argentina) for assisting us in obtaining Malbec and Isabella grape samples, and H.E. Martínez Beladelli for providing technical assistance during the initial stages of this project. M.L.R.E. held a postdoctoral fellowship of the Consejo Nacional de Investigaciones Científicas y Técnicas de Argentina (CONICET). A.L.R. is Principal Investigator of CONICET.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffunb.2025.1643880/full#supplementary-material
Supplementary Figure 1 | Dendrogram resulting from hierarchical cluster analysis based on Euclidean distances calculated from the average profiles of volatile compounds in each fermenting condition (i.e., Igj/Imc, Igj/Mmc, Mgj/Mmc, and Mgj/Imc) at 96 hours (Supplementary Table S3). Each point of the dendrogram represents the centroid of three biological replicates performed in the study.
Supplementary Figure 2 | Main represented yeast species at the initial stages of fermentations (0 to 96 h) of Isabella grape juice using Isabella (Imc) or Malbec (Mmc) microbial communities as inocula. Percentages represent the relative contribution of the indicated yeast species among the colonies obtained at the indicated times of fermentation.
References
Andorrà I., Berradre M., Rozès N., Mas A., Guillamón J. M., and Esteve-Zarzoso B. (2010). Effect of pure and mixed cultures of the main wine yeast species on grape must fermentations. Eur. Food Res. Technol. 231, 215–224. doi: 10.1007/s00217-010-1272-0
Andorrà I., Miró G., Espligares E., Mislata A. M., Puxeu M., and Ferrer-Gallego R. (2019). “Wild Yeast and Lactic Acid Bacteria of Wine,” in Yeasts in Biotechnology. Ed. Peixoto Basso T. (London, England: IntechOpen).
Arcanjo N. M. D. O., Oliveira M. E. D. S., Araújo Í.B.D.S., da Silva F. L. H., and Madruga M. S. (2018). Red wine produced from the Isabella and Ives cultivar (Vitis Labrusca): Profile of volatiles and aroma descriptors. Food Sci. Technol. (Brazil) 38, 271–279. doi: 10.1590/1678-457X.04717
Baffi M. A., Dos Santos Bezerra C., Arévalo-Villena M., Isabel Briones-Pérez A., Gomes E., and Da -Silva R. (2011). Isolation and molecular identification of wine yeasts from a Brazilian vineyard. Ann. Microbiol. 61, 75–78. doi: 10.1007/s13213-010-0099-z
Barata A., Malfeito-Ferreira M., and Loureiro V. (2012). The microbial ecology of wine grape berries. Int. J. Food Microbiol. 153, 243–259. doi: 10.1016/j.ijfoodmicro.2011.11.025
Belda I., Ruiz J., Esteban-fernández A., Navascués E., Marquina D., Santos A., et al. (2017). Microbial contribution to wine aroma and its intended use for wine quality improvement. Molecules 22 (2), 189. doi: 10.3390/molecules22020189
Bezerra-Bussoli C., Baffi M. A., Gomes E., and Da-Silva R. (2013). Yeast diversity isolated from grape musts during spontaneous fermentation from a Brazilian winery. Curr. Microbiol. 67, 356–361. doi: 10.1007/s00284-013-0375-9
Borren E. and Tian B. (2021). The important contribution of non-Saccharomyces yeasts to the aroma complexity of wine: A review. Foods 10(1), 13. doi: 10.3390/foods10010013
Carrau F., Boido E., and Ramey D. (2020). Yeasts for low input winemaking: Microbial terroir and flavor differentiation. 1st Edn (Amsterdam, Netherlands: Elsevier Inc). doi: 10.1016/bs.aambs.2020.02.001
Carrau F. and Henschke P. A. (2021). Hanseniaspora vineae and the concept of friendly yeasts to increase autochthonous wine flavor diversity. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.702093
Cilião Filho M., Bertéli M. B. D., Valle J. S., Paccola-Meirelles L. D., Linde G. A., Barcellos F. G., et al. (2017). Genetic diversity and pectinolytic activity of epiphytic yeasts from grape carposphere. Genet. Mol. Res. 16, 1–13. doi: 10.4238/gmr16029698
de Celis M., Ruiz J., Benitez-Dominguez B., Vicente J., Tomasi S., Izquierdo-Gea S., et al. (2024). Multi-omics framework to reveal the molecular determinants of fermentation performance in wine yeast populations. Microbiome 12, 203. doi: 10.1186/s40168-024-01930-w
de Celis M., Ruiz J., Vicente J., Acedo A., Marquina D., Santos A., et al. (2022). Expectable diversity patterns in wine yeast communities. FEMS Yeast Res. 22, foac034. doi: 10.1093/femsyr/foac034
De la Fuente Lloreda M. (2018). Use of hybrids in viticulture. A challenge for the OIV. OENO One 52, 231–234. doi: 10.20870/oeno-one.oeno-one.2018.52.3.2312
del Fresno J. M., Carrau F., Escott C., Vaquero C., González C., and Morata A. (2023). Use of Hanseniaspora spp. in sequential fermentation with Saccharomyces cerevisiae to improve the aromatic complexity of Albillo Mayor white wines. Bio Web Conferences. doi: 10.1051/bioconf/20236802029
del Fresno J. M., Escott C., Carrau F., Herbert-Pucheta J. E., Vaquero C., González C., et al. (2022). Improving aroma complexity with hanseniaspora spp.: terpenes, acetate esters, and safranal. Fermentation 8, 654. doi: 10.3390/fermentation8110654
Dellacassa E., Trenchs O., Fariña L., Debernardis F., Perez G., Boido E., et al. (2017). Pineapple (Ananas comosus L. Merr.) wine production in Angola: Characterisation of volatile aroma compounds and yeast native flora. Int. J. Food Microbiol. 241, 161–167. doi: 10.1016/j.ijfoodmicro.2016.10.014
Drumonde-Neves J., Franco-Duarte R., Lima T., Schuller D., and Pais C. (2016). Yeast biodiversity in vineyard environments is increased by human intervention. PloS One 11, 1–13. doi: 10.1371/journal.pone.0160579
Englezos V., Rantsiou K., Cravero F., Torchio F., Giacosa S., Ortiz-Julien A., et al. (2018). Volatile profiles and chromatic characteristics of red wines produced with Starmerella bacillaris and Saccharomyces cerevisiae. Food Res. Int. 109, 298–309. doi: 10.1016/j.foodres.2018.04.027
Englezos V., Rantsiou K., Torchio F., Rolle L., Gerbi V., and Cocolin L. (2015). Exploitation of the non-Saccharomyces yeast Starmerella bacillaris (synonym Candida zemplinina) in wine fermentation: Physiological and molecular characterizations. Int. J. Food Microbiol. 199, 33–40. doi: 10.1016/j.ijfoodmicro.2015.01.009
Filippousi M. E., Chalvantzi I., Mallouchos A., Marmaras I., Banilas G., and Nisiotou A. (2024). The use of hanseniaspora opuntiae to improve ‘Sideritis’ Wine quality, a late-ripening greek grape variety. Foods 13, 1061. doi: 10.3390/foods13071061
García-Izquierdo I., Colino-Rabanal V. J., Tamame M., and Rodríguez-López F. (2024). Microbiota ecosystem services in vineyards and wine: A review. Agronomy 14, 131. doi: 10.3390/agronomy14010131
Garde-Cerdán T., Gutiérrez-Gamboa G., Baroja E., Rubio-Bretón P., and Pérez-Álvarez E. P. (2018). Influence of methyl jasmonate foliar application to vineyard on grape volatile composition over three consecutive vintages. Food Res. Int. 112, 274–283. doi: 10.1016/j.foodres.2018.06.048
Ghaste M., Narduzzi L., Carlin S., Vrhovsek U., Shulaev V., and Mattivi F. (2015). Chemical composition of volatile aroma metabolites and their glycosylated precursors that can uniquely differentiate individual grape cultivars. Food Chem. 188, 309–319. doi: 10.1016/j.foodchem.2015.04.056
Gomez H. A. G., Niederauer G. F., Minatel I. O., Antunes E. R. M., Carneiro M. J., Sawaya A. C. H. F., et al. (2024). Metabolite profiling reveals the influence of grapevine genetic distance on the chemical signature of juices. J. Sci. Food Agric. 104 (4), 2383–2397. doi: 10.1002/jsfa.13124
Griggs R. G., Steenwerth K. L., Mills D. A., Cantu D., and Bokulich N. A. (2021). Sources and assembly of microbial communities in vineyards as a functional component of winegrowing. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.673810
Hu K., Jin G. J., Mei W. C., Li T., and Tao Y. S. (2018a). Increase of medium-chain fatty acid ethyl ester content in mixed H. uvarum/S. cerevisiae fermentation leads to wine fruity aroma enhancement. Food Chem. 239, 495–501. doi: 10.1016/j.foodchem.2017.06.151
Hu K., Jin G. J., Xu Y. H., and Tao Y. S. (2018b). Wine aroma response to different participation of selected Hanseniaspora uvarum in mixed fermentation with Saccharomyces cerevisiae. Food Res. Int. 108, 119–127. doi: 10.1016/j.foodres.2018.03.037
Hu L., Liu R., Wang X., and Zhang X. (2020). The sensory quality improvement of citrus wine through co-fermentations with selected non-saccharomyces yeast strains and saccharomyces cerevisiae. Microorganisms 8, 323. doi: 10.3390/microorganisms8030323
Jolly N. P., Varela C., and Pretorius I. S. (2014). Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 14, 215–237. doi: 10.1111/1567-1364.12111
Kamilari E., Mina M., Karallis C., and Tsaltas D. (2021). Metataxonomic analysis of grape microbiota during wine fermentation reveals the distinction of Cyprus regional terroirs. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.726483
Lederer M. A., Nielsen D., Toldam-Andersen T. B., Herrmann J. V., and Arneborg N. (2013). Yeast species associated with different wine grape varieties in Denmark. Acta Agric. Scand. B Soil Plant Sci. 63, 89–96. doi: 10.1080/09064710.2012.723738
Liang L., Ma Y., Jiang Z., Eudes F., and Peng S. (2023). Dynamic analysis of microbial communities and flavor properties in Merlot wines produced from inoculation and spontaneous fermentation. Food Res. Int. 164, 112379. doi: 10.1016/j.foodres.2022.112379
Liszkowska W. and Berlowska J. (2021). Yeast fermentation at low temperatures: Adaptation to changing environmental conditions and formation of volatile compounds. Molecules 26, 1035. doi: 10.3390/molecules26041035
Liu D., Chen Q., Zhang P., Chen D., and Howell K. S. (2020). The fungal microbiome is an important component of vineyard ecosystems and correlates with regional distinctiveness of wine. mSphere 5, 1–15. doi: 10.1128/msphere.00534-20
Maicas S. and Mateo J. J. (2023). The life of saccharomyces and non-saccharomyces yeasts in drinking wine. Microorganisms 11, 1178. doi: 10.3390/microorganisms11051178
Malletroit V., Guinard J.-X.-X., Kunkee R. E., and Lewis M. J. (1991). Effect of pasteurization on microbiological and sensory quality of white grape juice and wine. J. Food Process Preserv 15, 19–29. doi: 10.1111/j.1745-4549.1991.tb00151.x
Martin V., Jose Valera M., Medina K., Boido E., and Carrau F. (2018). Oenological impact of the Hanseniaspora/Kloeckera yeast genus on wines — A review. Fermentation 4, 76. doi: 10.3390/fermentation4030076
Morata A., Loira I., González C., and Escott C. (2021). Non-saccharomyces as biotools to control the production of off-flavors in wines. Molecules 26, 4571. doi: 10.3390/molecules26154571
Padilla B., Gil J. V., and Manzanares P. (2016). Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 7. doi: 10.3389/fmicb.2016.00411
Pretorius I. S. (2017). Solving yeast jigsaw puzzles over a glass of wine. EMBO Rep. 18, 1875–1884. doi: 10.15252/embr.201745231
Pretorius I. S. (2020). Tasting the terroir of wine yeast innovation. FEMS Yeast Res. 20(1), foz084. doi: 10.1093/femsyr/foz084
Raymond Eder M. L., Conti F., and Rosa A. L. (2018). Differences between indigenous yeast populations in spontaneously fermenting musts from V. vinifera L. and V. labrusca L. Grapes harvested in the same geographic location. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.01320
Raymond Eder M. L., Fariña L., Carrau F., and Rosa A. L. (2025). Grape-specific native microbial communities influence the volatile compound profiles in fermenting grape juices. Food Chem. 466, 142155. doi: 10.1016/j.foodchem.2024.142155
Raymond Eder M. L., Reynoso C., Lauret S. C., and Rosa A. L. (2017). Isolation and identification of the indigenous yeast population during spontaneous fermentation of Isabella (Vitis labrusca L.) grape must. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.00532
Raymond Eder M. L. and Rosa A. L. (2019). Yeast diversity in Vitis non-vinifera ecosystems. Rev. Argent Microbiol. 51, 278–283. doi: 10.1016/j.ram.2018.09.004
Raymond Eder M. L. and Rosa A. L. (2021). Genetic, physiological, and industrial aspects of the fructophilic non-saccharomyces yeast species, starmerella bacillaris. Fermentation 7, 87. doi: 10.3390/fermentation7020087
Romano P., Braschi G., Siesto G., Patrignani F., and Lanciotti R. (2022). Role of yeasts on the sensory component of wines. Foods 11, 1921. doi: 10.3390/foods11131921
Shimizu H., Kamada A., Koyama K., Iwashita K., and Goto-Yamamoto N. (2023). Yeast diversity during the spontaneous fermentation of wine with only the microbiota on grapes cultivated in Japan. J. Biosci. Bioeng 136, 35–43. doi: 10.1016/j.jbiosc.2023.03.013
Torres-Guardado R., Esteve-Zarzoso B., Reguant C., and Bordons A. (2022). Microbial interactions in alcoholic beverages. Int. Microbiol. 25, 1–15. doi: 10.1007/s10123-021-00200-1
Tristezza M., Tufariello M., Capozzi V., Spano G., Mita G., and Grieco F. (2016). The oenological potential of hanseniaspora uvarum in simultaneous and sequential co-fermentation with Saccharomyces cerevisiae for industrial wine production. Front. Microbiol. 7. doi: 10.3389/fmicb.2016.00670
Tufariello M., Fragasso M., Pico J., Panighel A., Castellarin S. D., Flamini R., et al. (2021). Influence of non-saccharomyces on wine chemistry: A focus on aroma-related compounds. Molecules 26, 644. doi: 10.3390/molecules26030644
Varela C. (2016). The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 100, 9861–9874. doi: 10.1007/s00253-016-7941-6
Varela C. and Borneman A. R. (2016). Yeasts found in vineyards and wineries. Yeast. 34 (3), 111–128. doi: 10.1002/yea.3219
Wang X., Fan G., Peng Y., Xu N., Xie Y., Zhou H., et al. (2023). Mechanisms and effects of non-Saccharomyces yeast fermentation on the aromatic profile of wine. J. Food Composition Anal. 124, 1–20. doi: 10.1016/j.jfca.2023.105660
Zhang L., Liu Q., Li Y., Liu S., Tu Q., and Yuan C. (2023a). Characterization of wine volatile compounds from different regions and varieties by HS-SPME/GC-MS coupled with chemometrics. Curr. Res. Food Sci. 6, 100418. doi: 10.1016/j.crfs.2022.100418
Zhang Z., Wang H., Xia H., Sun L., Zhang Q., Yang H., et al. (2023b). Wine aroma modification by Hanseniaspora uvarum: A multiple-step strategy for screening potential mixed starters. Food Chem. X 20, 100930. doi: 10.1016/j.fochx.2023.100930
Keywords: terroir, non-Saccharomyces, volatile profile, fermentation, Isabella
Citation: Raymond Eder ML, Fariña L, Carrau F and Rosa AL (2025) Microbial communities from distinct Vitis species shape volatile profiles of fermenting juices while preserving varietal typicity. Front. Fungal Biol. 6:1643880. doi: 10.3389/ffunb.2025.1643880
Received: 09 June 2025; Accepted: 28 August 2025;
Published: 18 September 2025.
Edited by:
Athanasia Koliadima, University of Patras, GreeceReviewed by:
Angela Capece, University of Basilicata, ItalyLaura Pulcini, Council for Agricultural Research and Economics – Research Centre for Viticulture and Enology CREA-VE, Italy
Copyright © 2025 Raymond Eder, Fariña, Carrau and Rosa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alberto Luis Rosa, YWxyb3NhQHVuYy5lZHUuYXI=; María Laura Raymond Eder, bWxyYXltb25kZWRlckB1bmMuZWR1LmFy