Carbon (C) neutrality is the urgent and ultimate goal of mitigating the anthropogenic greenhouse gas (GHG) emissions associated with global climate challenges. As the ocean is the largest active C reservoir on Earth, marine microorganisms play important roles in the global C cycle. For example, marine microorganisms can remove carbon dioxide (CO2) from the atmosphere by C sequestration processes, and reduce methane (CH4) emissions through their interactions with the environment, especially dissolved organic matter (DOM). However, how marine microorganisms interact with DOM to realize C sequestration remains unclear. In the marine ecosystem, the capture, transformation, and transport of C are mainly driven by the biological pump (BP) (Honjo et al., 2014), microbial carbon pump (MCP) (Jiao et al., 2010), and microbially induced carbonate precipitation (MICP) (Macreadie et al., 2017). The MCP mechanism emphasizes the contribution of marine microorganisms to recalcitrant dissolved organic carbon (RDOC) production through the degradation of particulate organic matter (POM), cell death, and interspecies interactions (Jiao et al., 2010). The BP, MCP and MICP altogether may work cooperatively to drive efficient C sequestration and storage in marine environments, thereby mitigating global climate change.
The Research Topic “Carbon Storage by Marine Microorganisms for Carbon Neutrality” presents five publications in Frontiers in Microbiology and Frontiers in Marine Science, providing new insights into our understanding of marine microbial community diversity and functions, biotic interactions (e.g., macrophyte-phytoplankton, algae-bacteria, bacteria-fungi), microbe-DOM interactions, and possible C storage mechanisms in marine ecosystems. Marine macrophytes (e.g., seagrasses, macroalgae) are important primary producers, and serve a variety of ecosystem functions such as biogeochemical cycling of C, nitrogen (N) and phosphorus (P) through macrophyte-microbe interactions (Egan et al., 2013; Human et al., 2015; Xie et al., 2017), and mitigation of global climate change through blue carbon sequestration (Duarte et al., 2018). In this Research Topic, Chen et al. analyzed the diversity and composition of epiphytic bacterial and fungal communities from 11 types of macroalgae and 2 types of seagrasses. The results indicate that macrophyte host phylogeny influences the bacterial community structure, while geographic differences mainly shape the fungal community structure. C metabolizing bacteria and fungi are enriched in the epiphytic environment of marine macrophytes. Also, phytoplankton are widely distributed in the ocean and contribute importantly to C sequestration via the BP (Honjo et al., 2014). Zhao et al. found distinct species-specific algae-bacteria relationships in different water layers, being more pronounced in the mesopelagic and bathypelagic zones than in the euphotic zone. A laboratory experiment further showed that phytoplankton reshaped the bacterioplankton community structure in different water layers. In a study of marine picophytoplankton (e.g., Prochlorococcus, Synechococcus, picoeucaryote) and their distribution patterns in the West Pacific, Prochlorococcus was found to dominate over Synechococcus and picoeucaryotes, and the abundance of Prochlorococcus was negatively correlated with nutrient concentrations, while temperature and salinity were closely correlated with the spatial variation in the picophytoplankton community, highlighting the importance of picophytoplankton in contributing to the C pool in the oligotrophic ocean (Wang et al.). To further understand bacterial transformation and processing of DOM in aquatic ecosystems, three bacteria (Roseobacteria sp., Marinobacter sp., and Bacillus sp.) were selected to investigate the processing of DOM and lysate organic matter (LOM) from the marine diatom Skeletonema dohrnii. The results showed that such epiphytic bacteria were able to process DOM and LOM, and convert them into more complex refractory dissolved organic matter (RDOM) (Liu et al.). In addition, in this Research Topic, Ma et al. reviewed current knowledge and future exploration of thermophilic microorganisms from submarine hydrothermal environments as a valuable source of thermostable tolerant carbonic anhydrases, highlighting their potential applications for rapid CO2 capture in the marine ecosystem.
Based on these studies as mentioned above and other existing literatures, we propose a conceptual framework to summarize biotic interactions on the DOM through the microbial loop and microbially-driven DOM degradation and transformation by the MCP mechanism (Figure 1). First, marine microorganisms interact with each other and with macroorganisms to form complex networks through food webs (e.g., the degradation of macroorganism-derived DOM). The necromass from those macroorganisms and microorganisms may sink to the seabed and C may, thereby, be sequestered from upper waters. Second, microbially-driven DOM degradation may result in diverse products, including labile dissolved organic carbon (LDOM), which is mainly used by microorganisms and produces CO2 and/or CH4. CO2 may be re-fixed by autotrophic or chemo-autotrophic microorganisms (Dyksma et al., 2016; Vasquez-Cardenas et al., 2020; Taubert et al., 2022), and more importantly, a recent study indicates that CH4 may be re-transformed into DOM (Yang et al., 2020). A part of such DOM may be used by marine microorganisms and further transformed into RDOC, or RDOM by the MCP mechanism for C sequestration. A comprehensive understanding of those mechanisms, especially microorganism-DOM interactions, is necessary for us to develop efficient technologies for increasing C storage in the marine ecosystem. In the future, several strategies may enhance C sequestration (Figure 1). These may include (1) the enhancement of RDOM formation by the selection of microbial taxa/genes for efficient transformation of DOM and LDOM into RDOM (McCarren et al., 2010; Osterholz et al., 2015; Lechtenfeld et al., 2015); (2) the reduction of CH4 emissions by inhibiting CH4 production (Huang et al., 2009) or strengthening CH4 oxidation (Sivan et al., 2014; Egger et al., 2015); and (3) the re-transformation of CO2 and CH4 into organic C (Dyksma et al., 2016; Yang et al., 2020). The implementation of such strategies for increasing C sequestration in marine ecosystems calls for the urgent development of innovative science and technology in microbial ecology, microbiome engineering and geoengineering.
Figure 1
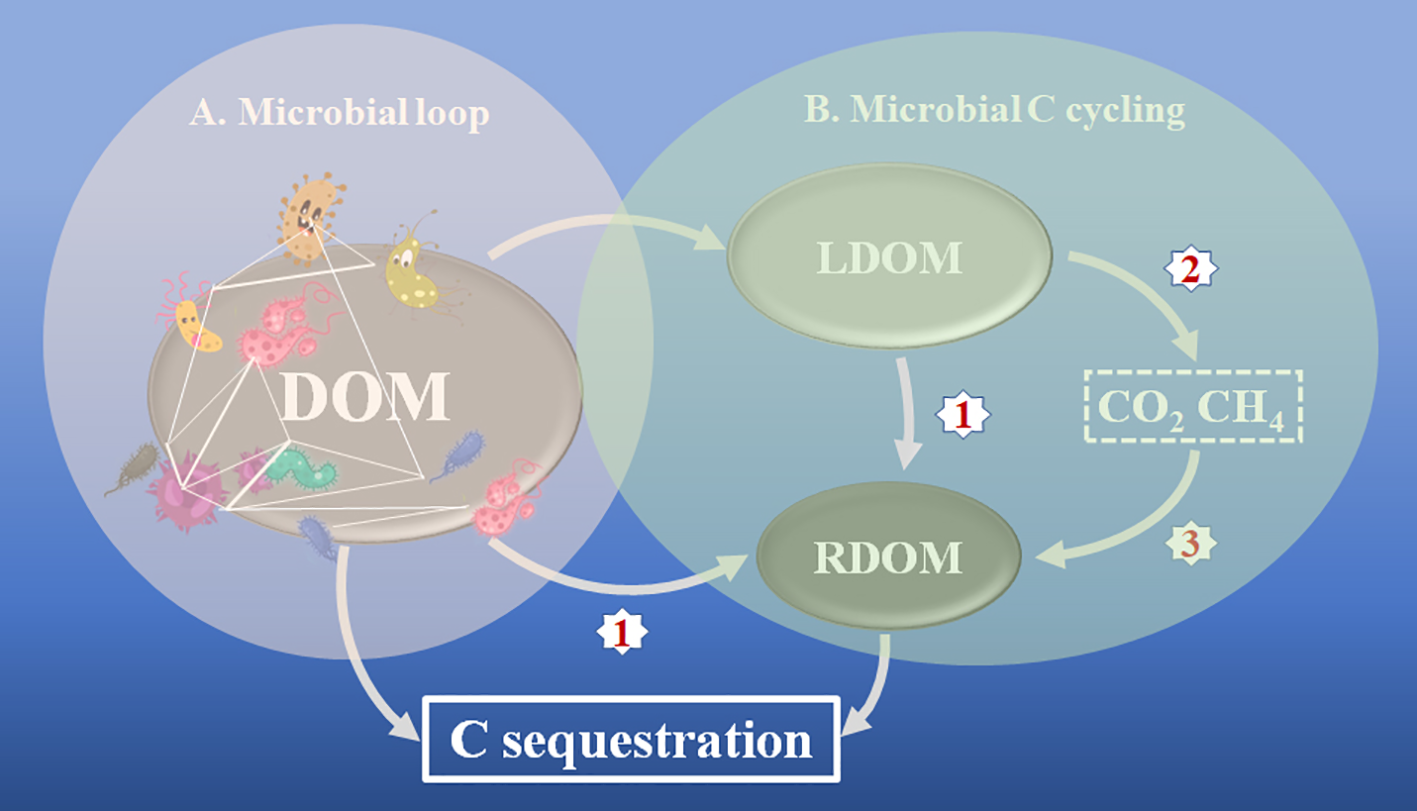
A conceptual framework for understanding C sequestration mechanisms in the marine ecosystem. (A) Biotic interactions are shown as complex networks between marine organisms (including macro-organisms and micro-organisms) affecting DOM dynamics. C sequestration is mainly driven by the biological pump (BP) mechanism. (B) Microbially-driven DOM degradation and transformation are largely explained by the microbial carbon pump (MCP) mechanism. The red numbers (1, 2 and 3) indicate possible future strategies for enhancing C sequestration in the marine ecosystem. DOM, dissolved organic matter; LDOM, labile dissolved organic matter; RDOM, refractory dissolved organic matter.
Funding
This work was supported by the Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai) (313022002, SML2020SP004 and 311021006), and by the National Natural Science Foundation of China (91951207 and 31770539).
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Statements
Author contributions
ZH drafted this manuscript; LL, XW, WQ and CZ edited it. All authors contributed to the article and approved the submitted version.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1
Duarte B. Martins I. Rosa R. Matos A. R. Roleda M. Y. Reusch T. B. H. et al . (2018). Climate change impacts on seagrass meadows and macroalgal forests: An integrative perspective on acclimation and adaptation potential. Front. Mar. Sci.5. doi: 10.3389/fmars.2018.00190
2
Dyksma S. Bischof K. Fuchs B. M. Hoffmann K. Meier D. Meyerdierks A. et al . (2016). Ubiquitous gammaproteobacteria dominate dark carbon fixation in coastal sediments. ISME J.10 (8), 1939–1953. doi: 10.1038/ismej.2015.257
3
Egan S. Harder T. Burke C. Steinberg P. Kjelleberg S. Thomas T. (2013). The seaweed holobiont: Understanding seaweed-bacteria interactions. FEMS Microbiol. Lett.37 (3), 462–476. doi: 10.1111/1574-6976.12011
4
Egger M. Rasigraf O. Sapart C. J. Jilbert T. Jetten M. S. Röckmann T. et al . (2015). Iron-mediated anaerobic oxidation of methane in brackish coastal sediments. Environ. Sci. Technol.49 (1), 277–283. doi: 10.1021/es503663z
5
Honjo S. Eglinton T. I. Taylor C. D. Ulmer K. M. Sievert S. M. Bracher A. et al . (2014). Understanding the role of the biological pump in the global carbon cycle: An imperative for ocean science. Oceanography27 (3), 10–16. doi: 10.5670/oceanog.2014.78
6
Huang B. Yu K. Gambrell R. P. (2009). Effects of ferric iron reduction and regeneration on nitrous oxide and methane emissions in a rice soil. Chemosphere74 (4), 481–486. doi: 10.1016/j.chemosphere.2008.10.015
7
Human L. R. D. Snow G. C. Adams J. B. Bate G. C. Yang S. C. (2015). The role of submerged macrophytes and macroalgae in nutrient cycling: A budget approach. Estuar. Coast. Shelf Sci.154, 169–178. doi: 10.1016/j.ecss.2015.01.001
8
Jiao N. Herndl G. J. Hansell D. A. Benner R. Kattner G. Wilhelm S. W. et al . (2010). Microbial production of recalcitrant dissolved organic matter: Long-term carbon storage in the global ocean. Nat. Rev. Microbiol.8 (8), 593–599. doi: 10.1038/nrmicro2386
9
Lechtenfeld O. J. Hertkorn N. Shen Y. Witt M. Benner R. (2015). Marine sequestration of carbon in bacterial metabolites. Nat. Commun.6, 6711. doi: 10.1038/ncomms7711
10
Macreadie P. I. Serrano O. Maher D. T. Duarte C. M. Beardall J. (2017). Addressing calcium carbonate cycling in blue carbon accounting. Limnol. Oceanogr. Lett.2 (6), 195–201. doi: 10.1002/lol2.10052
11
McCarren J. Becker J. W. Repeta D. J. Shi Y. Young C. R. Malmstrom R. R. et al . (2010). Microbial community transcriptomes reveal microbes and metabolic pathways associated with dissolved organic matter turnover in the sea. Proc. Natl. Acad. Sci. U.S.A.107 (38), 16420–16427. doi: 10.1073/pnas.1010732107
12
Osterholz H. Niggemann J. Giebel H.-A. Simon M. Dittmar T. (2015). Inefficient microbial production of refractory dissolved organic matter in the ocean. Nat. Commun.6 (1), 7422. doi: 10.1038/ncomms8422
13
Sivan O. Antler G. Turchyn A. V. Marlow J. J. Orphan V. J. (2014). Iron oxides stimulate sulfate-driven anaerobic methane oxidation in seeps. Proc. Natl. Acad. Sci. U.S.A.111 (40), E4139–E4147. doi: 10.1073/pnas.1412269111
14
Taubert M. Overholt W. A. Heinze B. M. Matanfack G. A. Houhou R. Jehmlich N. et al . (2022). Bolstering fitness via CO2 fixation and organic carbon uptake: mixotrophs in modern groundwater. ISME J.16 (4), 1153–1162. doi: 10.1038/s41396-021-01163-x
15
Vasquez-Cardenas D. Meysman F. J. R. Boschker H. T. S. (2020). A cross-system comparison of dark carbon fixation in coastal sediments. Glob. Biogeochem. Cycles34 (2), e2019GB006298. doi: 10.1029/2019GB006298
16
Xie X. F. He Z. L. Hu X. J. Yin H. Q. Liu X. D. Yang Y. F. (2017). Large-Scale seaweed cultivation diverges water and sediment microbial communities in the coast of nan’ao island, south China Sea. Sci. Total Environ.598, 97–108. doi: 10.1016/j.scitotenv.2017.03.233
17
Yang S. Lv Y. Liu X. Wang Y. Fan Q. Yang Z. et al . (2020). Genomic and enzymatic evidence of acetogenesis by anaerobic methanotrophic archaea. Nat. Commun.11 (1), 3941. doi: 10.1038/s41467-020-17860-8
Summary
Keywords
marine microorganism, carbon storage, carbon degradation, dissolved organic matter, greenhouse gas emission, algal-bacterial interaction, microbe-DOM-environment
Citation
He Z, Lin L, Wang X, Qin W and Zhang C (2022) Editorial: Carbon storage by marine microorganisms for carbon neutrality. Front. Mar. Sci. 9:1018397. doi: 10.3389/fmars.2022.1018397
Received
13 August 2022
Accepted
23 August 2022
Published
06 September 2022
Volume
9 - 2022
Edited and reviewed by
Lasse Riemann, University of Copenhagen, Denmark
Updates
Copyright
© 2022 He, Lin, Wang, Qin and Zhang.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhili He, hezhili@sml-zhuhai.cn
This article was submitted to Aquatic Microbiology, a section of the journal Frontiers in Marine Science
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.