Explore article hub
 Paula Fernández-Gómez1
Paula Fernández-Gómez1 Dara Leong1
Dara Leong1 Gabriele Berg2,3,4
Gabriele Berg2,3,4 Fiona Brennan5,6
Fiona Brennan5,6 Tancredi Caruso7
Tancredi Caruso7 Trevor C. Charles8
Trevor C. Charles8 Luca S. Cocolin9
Luca S. Cocolin9 Lene Lange10
Lene Lange10 Olivia McAuliffe1
Olivia McAuliffe1 Emmanuelle Maguin11
Emmanuelle Maguin11 Orla O’Sullivan1,6,12
Orla O’Sullivan1,6,12 Yolanda Sanz13
Yolanda Sanz13 Inga Sarand14
Inga Sarand14 Angela Sessitsch15
Angela Sessitsch15 Hauke Smidt16
Hauke Smidt16 Nicholas Brereton7
Nicholas Brereton7 Marco Candela17
Marco Candela17 John Kenny1,6,12
John Kenny1,6,12 Tanja Kostic15
Tanja Kostic15 Jennifer Mahony12,18
Jennifer Mahony12,18 Martin Wagner19
Martin Wagner19 Paul D. Cotter1,6,12*
Paul D. Cotter1,6,12*- 1Food Bioscience, Teagasc Food Research Centre, Moorepark, Ireland
- 2Institute of Environmental Biotechnology, Graz University of Technology, Graz, Austria
- 3Microbiome Biotechnology, Leibniz Institute for Agricultural Engineering and Bioeconomy (ATB), Potsdam, Germany
- 4Institute for Biochemistry and Biology, University of Potsdam, Potsdam, Germany
- 5Environment, Soils and Landuse, Teagasc Johnstown Castle Environmental Research Centre, Wexford, Ireland
- 6VistaMilk SFI Research Centre, Fermoy, Cork, Ireland
- 7School of Biology and Environmental Science, University College Dublin, Belfield, Dublin, Ireland
- 8Department of Biology, University of Waterloo, Waterloo, ON, Canada
- 9Department of Agricultural, Forest and Food Sciences, University of Turin, Grugliasco, Italy
- 10LL-BioEconomy, Research & Advisory, Copenhagen, Denmark
- 11Micalis Institute, Université Paris-Saclay-INRAE-AgroParisTech, Jouy-en-Josas, France
- 12APC Microbiome Ireland, Cork, Ireland
- 13Microbial Ecology, Nutrition and Health Research Unit, Institute of Agrochemistry and Food Technology, Spanish National Research Council (IATA-CSIC), Valencia, Spain
- 14Department of Chemistry and Biotechnology, Tallinn University of Technology, Tallinn, Estonia
- 15Bioresources Unit, Center for Health & Bioresources, AIT Austrian Institute of Technology GmbH, Tulln, Austria
- 16Laboratory of Microbiology, Wageningen University and Research, Wageningen, Netherlands
- 17Department of Pharmacy and Biotechnology, University of Bologna, Bologna, Italy
- 18School of Microbiology, University College Cork, Cork, Ireland
- 19Austrian Competence Centre for Feed and Food Quality, Safety and Innovation (FFoQSI GmbH), Tulln, Austria
Abstract
Food system microbiomes include complex microbial networks that range from soil and marine environments to primary agriculture, farming, food processing, and distribution, and which influence human and environmental health. Advances in “omics” technologies, such as metagenomics, metatranscriptomics, metaproteomics, metabolomics, and culturomics, and their integration have deepened our understanding of microbiome dynamics and interactions. This growing knowledge is being leveraged to develop microbiome-based solutions enabling more sustainable food systems. This review explores microbiome interconnections along the food system and how this and other knowledge relating to microbiomes can be harnessed to, among other things, enhance crop resilience and productivity, improve animal health and performance, refine management practices in fishing and aquaculture, or prolong shelf life and reduce food spoilage during distribution. The often-overlooked role of bacteriophages on shaping microbiomes is discussed, as is the impact of diet on the human gut microbiota and, in turn, health. Despite advances, knowledge remains incomplete in particular areas and targeted experimental approaches are necessary to fill these gaps—going beyond merely predicting microbiome functionality. Ultimately, the ideal development of microbiome-based innovations in food systems will require collaboration between stakeholders and regulators to ensure safety, efficacy, and widespread adoption, unlocking its full potential to improve the health of animals, humans and the environment globally.
Key points
- Food system microbiomes form a complex, dynamic, and interconnected network that can be traced across soil and marine environments to primary agriculture, farming, and food processing sectors, ultimately influencing human gastrointestinal niches. These microbiomes impact both human and environmental health.
- Meta-omic technologies are valuable tools that contribute to advancing our understanding of the composition, functionality, and interactions of food system microbiomes within and across ecosystems to fully leverage the potential of microbiomes.
- Advances in microbiome-characterization technologies have helped to highlight the deterioration of specific microbiome networks, including decreased microbial diversity and the spread of antimicrobial resistance, and also to design positive microbiome modulation interventions.
- Microbiome-based innovations and applications have the potential to improve the resilience and sustainability of agri-food systems, and the global health of animals, humans, and the environment.
- An integrated approach involving all relevant stakeholders is vital for protecting existing microbiomes and restoring damaged microbial networks.
Introduction
Agri-food systems are crucial for society. They consist of multiple integrated sectors that provide nourishment while also contributing significantly to employment, trade, and innovation (1, 2). In 2021, the global annual value of agriculture, forestry, and fishing reached US$3.7 trillion and the sector was estimated to provide a livelihood for around one billion people, accounting for 27% of the global workforce (3). In addition, agriculture can have a variety of positive and negative impacts on environmental health, including various aspects of nutrient cycling, water purification, landscape conservation, as well the emission of ammonia and methane.
The 21st century has brought a wide spectrum of climate-related challenges, including prolonged droughts, extreme flooding episodes, and biodiversity loss, which test the resilience of agri-food systems. Moreover, (intense) agriculture is one of the most important anthropogenic drivers of these crises, contributing to six of nine planetary boundaries already being crossed. The planetary boundaries concept defines safe limits for nine critical processes that maintain a stable and resilient Earth. The boundaries “climate change”, “biosphere integrity”, “biogeochemical flows”, “land-system change”, and “freshwater use” are those that are already outside what has been defined as a safe operating space for humanity (4).
An estimated one-fifth of food produced globally for human consumption is lost or wasted (5), which highlights the necessity to reduce food losses along production and supply chains as well as at retail and consumer levels. Indeed, the targets associated with United Nations (UN) 2030 Sustainable Development Goals (SDGs) include reducing by 50% food losses per capita along production and supply chains, doubling agricultural productivity, ensuring sustainable and resilient food production systems, increasing water-use efficiency, and reducing land degradation (6). Ultimately, innovation is critical to ensure food availability, accessibility and affordability while addressing the need to reduce greenhouse gas emissions, energy demand and waste, increase productivity, and preserve environments and biodiversity (7).
In recent decades, there has been a notable increase in research dedicated to understanding microbiomes along the food system. A microbiome is defined as a characteristic microbial community occupying a well-defined habitat that has distinct physio-chemical properties. This term refers not only to the microorganisms involved but also includes their theatre of activity (8). Microbiomes, including those in food systems, are dynamic and interactive micro-ecosystems that are susceptible to changes over time and scale, including processes of coalescence among microbiomes that come into contact (9).
Agri-food-related environments—soil, water, air, plant, food-processing industries, food products, and animals, including humans—contain microbiomes that are often linked to form complex interacting networks. Indeed, microbiomes can influence each other directly, by the inter-microbiome dispersion of microorganisms able to colonize different ecosystems, or indirectly, through the exchange of different molecular mediators (10). This trans-microbiome molecular crosstalk is mediated by the microbiome secretome, which includes proteins and metabolites, which can control, in many cases selectively, the growth of certain taxa. Other mechanisms of microbiome interaction depend on predatory dynamics, involving inter-microbiome exchanges of bacterial, protist, and/or viral predators, such as phages (11). The rhizosphere microbiome offers an example of such a complex trans-microbiome interaction dynamic. Closely connected to soil and aquatic ecosystems, this microbiome can regulate the content of complex polysaccharides or secondary metabolites (e.g., polyphenols or tannins) produced by edible plants; moreover, once ingested by herbivores or omnivores (including humans), these plant foods can shape the composition and functional layout of their respective gut microbiomes.
This concept of interconnected environments is encompassed within the One Health approach, which aims to balance and optimize the health of people, animals, and ecosystems (12), with microbiomes playing a pivotal role (13). More specifically, microbial communities along the food system influence and shape the nutritional, quality, safety, and sensory attributes of food products. Conversely, human activities can modify those microbiomes, with both positive and negative outcomes for global health (Figure 1).
Figure 1
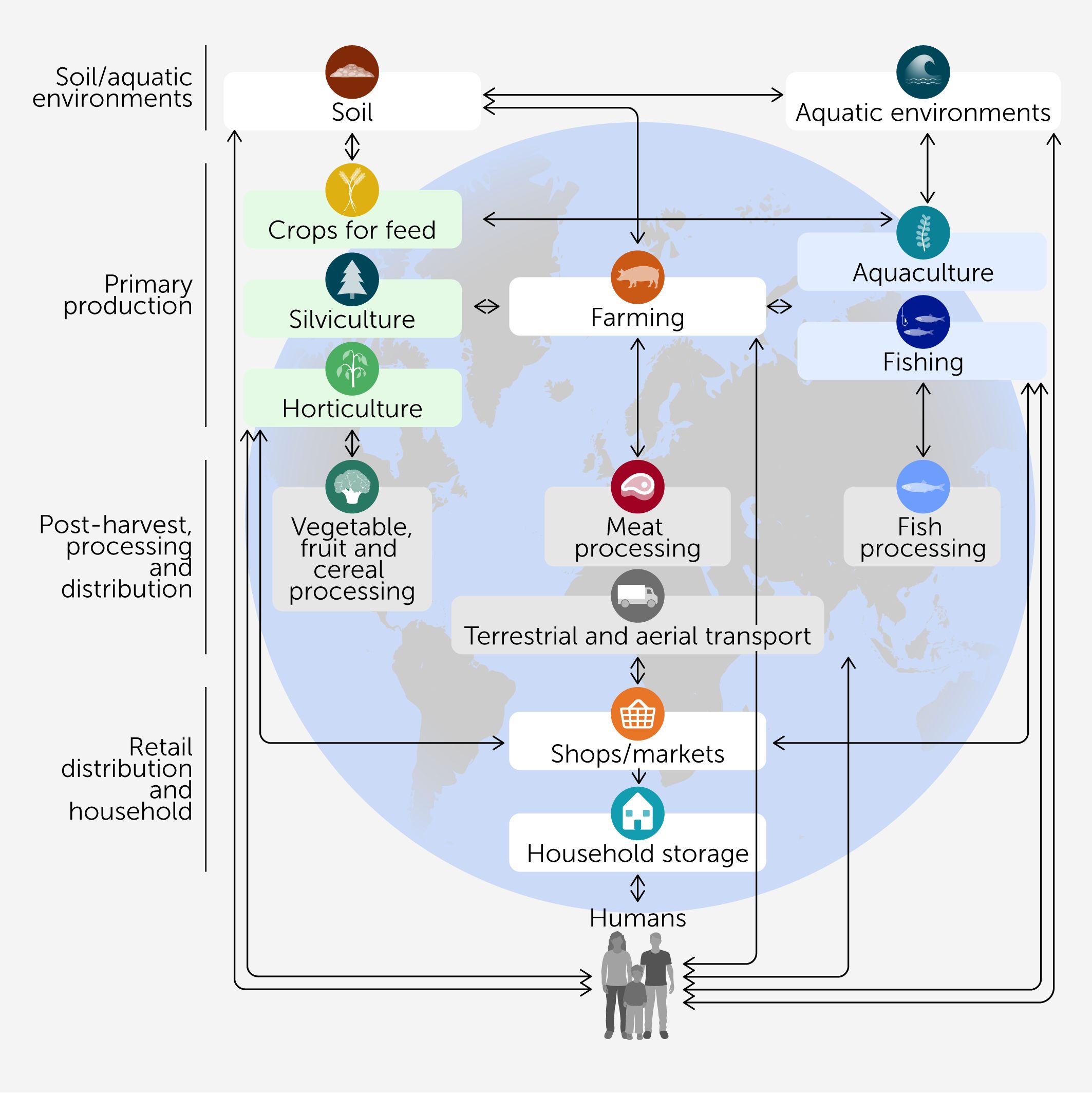
Figure 1. Microbiome connections. General microbiome interconnections map in agri-food systems and their potential to impact human health. The arrows represent the bidirectional influences between the most relevant food system environments, which have been grouped into broader categories (left-hand side labels) that correspond to the different sections of the review.
The rapid development of omics technologies has allowed us to analyze the structural and functional dynamics of these microbiomes at unprecedented spatiotemporal resolution, as well as the complex, interdependent networks that microbes and their host form in ecosystems. This has included the use of complementary approaches, such as metagenomics, metatranscriptomics, metaproteomics, metaproteomics, metabolomics, and culturomics, to provide valuable insights into the composition, functional capacity, and biochemical activity of microbial communities and specific components thereof (14, 15). It has also involved the development and use of large data modelling techniques, such as those from network science (16, 17). These advancements have led to several applications for sustainable food systems, as previously reviewed (18–23).
Figure 2 depicts the general pathway followed for the development of microbiome-based agri-food solutions. High-throughput DNA sequencing technologies have been particularly useful to help track the movement of specific strains into and through different ecological niches, thanks to the downstream analysis of these data through bioinformatic pipelines. Whole genome sequencing has been used for strain tracking during outbreak investigations, using well-defined thresholds of single nucleotide polymorphisms (SNPs) between genomes to differentiate strains, and allowing the identification of contamination routes in food processing facilities (24). Whole metagenome sequencing has also been used to characterize microbial networks and track strain movement through the reconstruction of metagenome-assembled genomes (MAGs), as in the case of the tracking of individual Bifidobacterium strains between mothers and infants (25, 26). These examples demonstrate how alterations in one microbiome can subsequently affect other microbiomes and how this knowledge can be harnessed to develop microbiome modulation strategies.
Figure 2
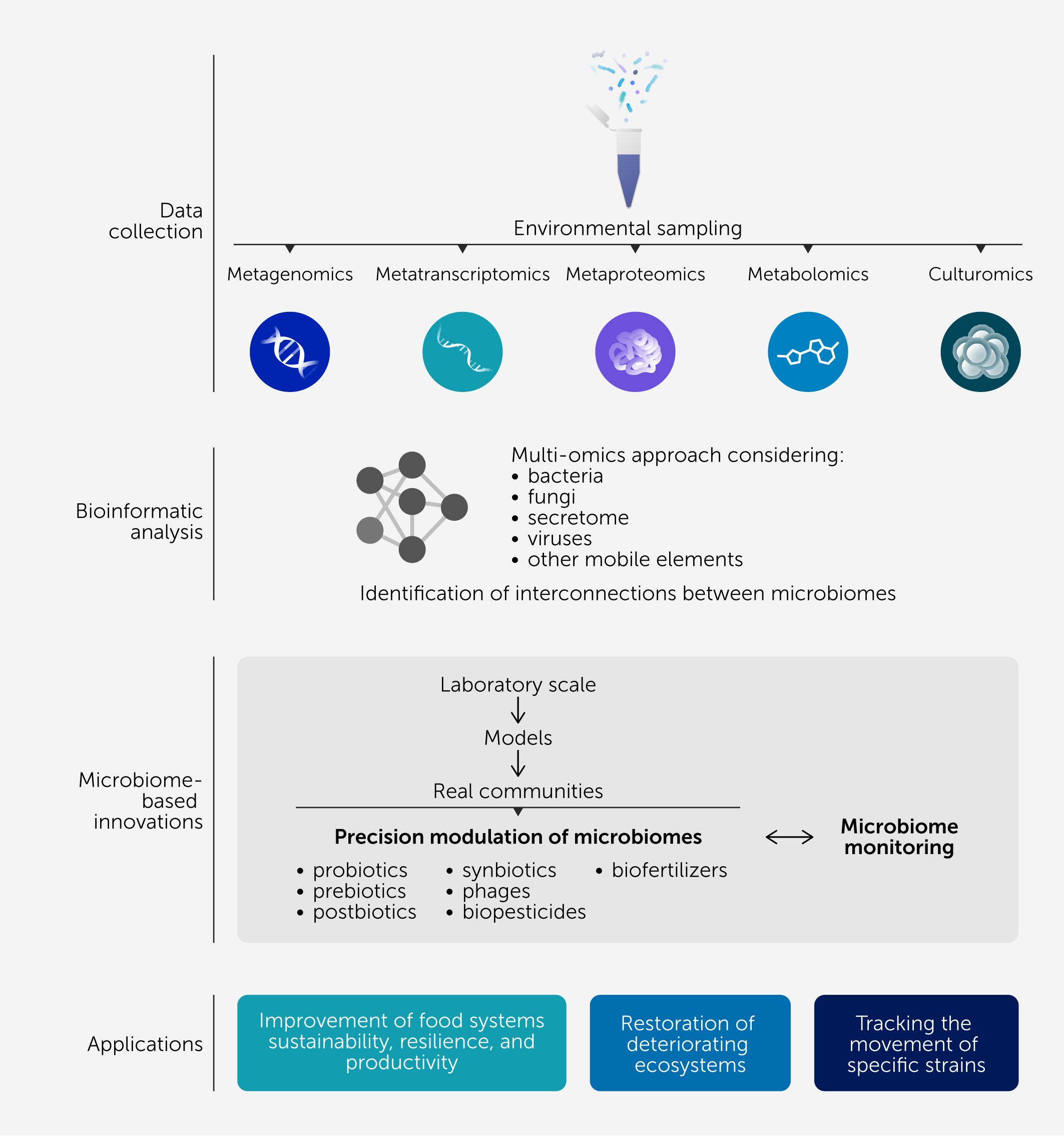
Figure 2. Multi-omics for microbiome interconnections research. Pathway for microbiome exploration using diverse omics methodologies and for the development of novel solutions leveraging microbiome interconnectedness. The importance of considering the different components of the microbiome and the functional capacity of the secretome (proteins and metabolites) is highlighted. The interconnections between microbiomes can be studied at different levels, from laboratory scale (increasing the systems complexity), followed by models of real environments (using ex vivo models, animal studies, greenhouses tests, etc.) and finally real-world communities (including field studies, studies with humans and livestock, and studies on rivers, oceans, soils or food processing facilities). Microbiome modification strategies can be designed by harnessing the knowledge relating to microbiome interconnectedness and information derived from microbiome monitoring.
Influencing microbial communities through intervention has the potential to alter human and environmental health and improve our understanding of microbial niches. Efforts to influence the microbiome in a persistent manner have taken many forms, including the use of probiotics, prebiotics, starter cultures, protective cultures and other biotics (27, 28). While pro- and prebiotics initially came to prominence as supplements for human health, they are now also widely used in agriculture, where they can also be referred to as biostimulants and biofertilizers, and aquaculture with a view to protecting farm animals and fish against disease or to enhance health. Some examples of microbes being used in agri- and aquaculture in this way include the application of bacterial communities to desert crops to protect against abiotic stress (29), the use of yeast to decrease postharvest disease in strawberry production (30, 31), bioprotective lactic acid bacteria treatments used to prevent spoilage in food production (30, 32), and targeted vaccination against the louse microbiome to reduce salmon infections (33). Microbial modulation approaches can also be employed to control a microbial population, and as such, decrease the use of antibiotics, which should be limited to strictly therapeutic purposes due to the potential spread of antibiotic resistance as well as collateral damage that can be inflicted on desirable microorganisms. Microbiome-targeted interventions also have the potential to restore deteriorating biodiversity and increase the resilience of entire (agricultural) ecosystems (34). While such interventions have proven to be successful in specific circumstances, there are also many examples of failures, with challenges including difficulties in translating research in laboratories or on a pilot scale to a real-world setting.
This review aims to highlight the importance of food system microbiomes in the context of environmental and societal health through a series of examples. More specifically, we highlight how the interconnectedness of microbiomes can be apparent through continuous links from soil and marine environments, to primary agriculture, farming and food processing sectors, and ultimately influencing the human gastrointestinal niches and, in the process, impacting both human and environmental health.
Microbiome interconnections and interventions throughout the food system
Primary production environments
Land used for agriculture
The implementation of agricultural practices that promote soil health is key in order to achieve the One Health objective, i.e., accomplishing the best health outcomes for people, animals, plants, and our environment (35, 36). The soil microbiome is closely interconnected with the microbial communities found within above- and below-ground plant microbiomes (Figure 3A). These short- and long-term associations are influenced by various abiotic and biotic factors. Ultimately, they play a crucial role in crop growth and health (37, 38), and can be harnessed to, for example, improve the yields, quality, and resilience of agricultural crops (39). In this regard, abiotic factors, such as soil type and salinity, water, and nutrient availability or photoperiod, have traditionally been the focus of strategies such as irrigation and fertilization.
Figure 3
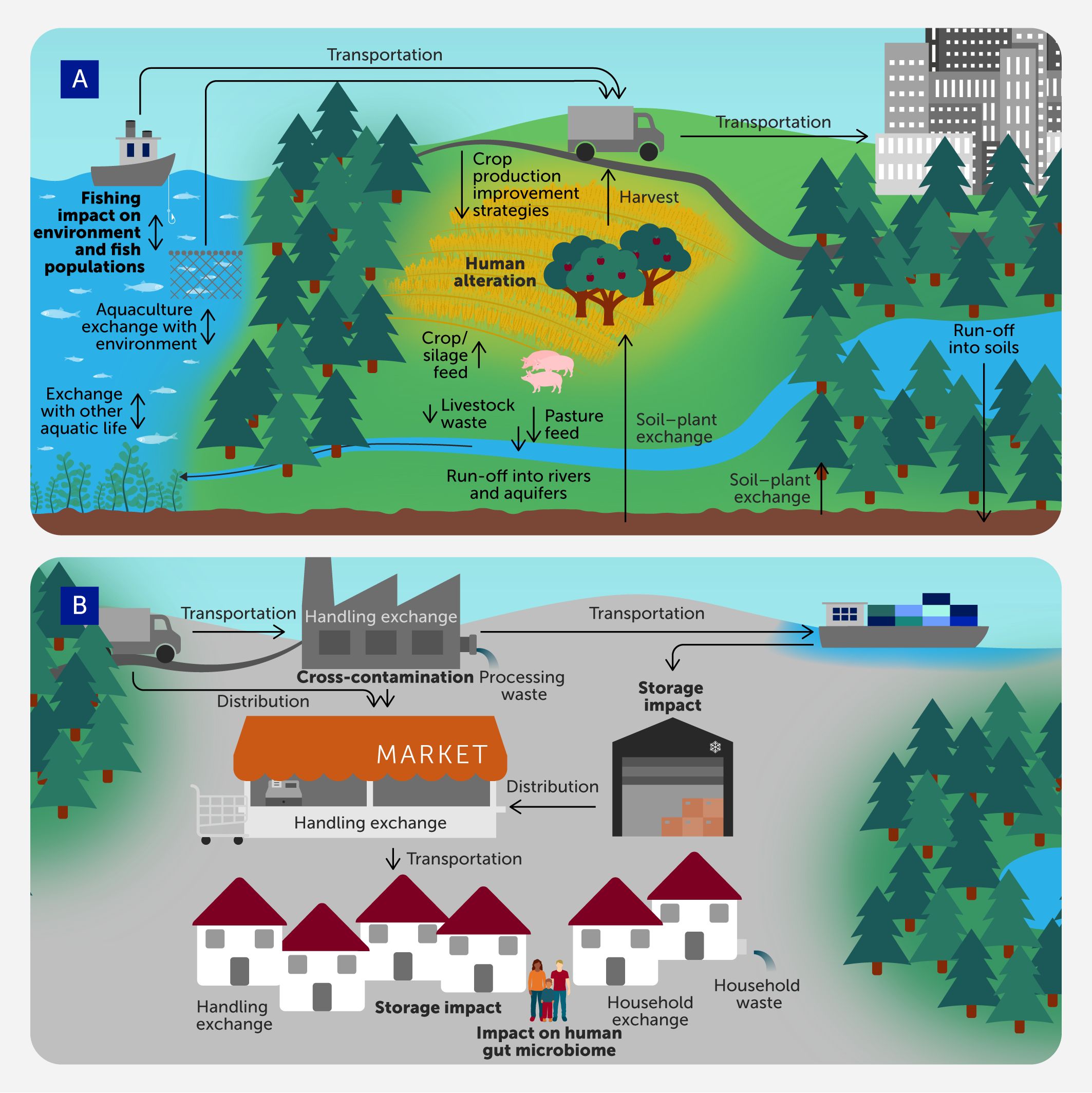
Figure 3. Microbiome interconnectedness in agri-food environments (A) in horticulture and silviculture cropping, livestock farming and aquatic environments, and (B) in distribution, processing, transport to retail, storage, markets, and household handling before reaching the consumer.
Some soil–plant interactions with potential applications in agriculture have been observed in non-managed field environments where plants are exposed to extreme conditions. The current knowledge regarding the role of plant-associated microbiomes on abiotic stress protection has been described in multiple review articles on drought stress (40), flooding stress (41), and salt tolerance (42, 43). Moreover, the influence of plant microbiomes on nitrogen fixation (44–46) and the mobilization of other minerals (47) have been well documented. Even host-associated microbiomes, such as those of soil nematodes, can be important for the decontamination of a polluted soil and the release of important macro- and micronutrients for soil fertilization (48). There are also studies that address the influence of microbiomes in mitigating the effects of other limiting abiotic factors on plant fitness, including cold stress (49), freezing stress (50), and other extreme conditions, such as the presence of heavy metals or acidic and alkaline soils (51). Additionally, co-cultivation and intercropping strategies have been widely adopted to increase production and quality by improving soil fertility. To this end, understanding the role of microbial communities in this response has greatly benefited from the application of omics-based technologies (52–54).
Extensive research has focused on how the composition and activities of soil microbiomes, the plant genome, and environmental characteristics combine to impact plant health (38, 55, 56). This approach is based on the so-called “microbiome-associated phenotype” models that consider the contributions of the host genotype, environmental factors, and microbiome to the final host phenotype (57). However, even more interesting is the possibility of intentionally modifying soil microbiomes to replicate similar benefits (58–63). Some successful applications include the engineering of soils with plant-associated bacteria consortia to protect different crops against abiotic stresses such as high salinity, to preserve stable wheat yields under drought, to improve phosphorus solubilization and nitrogen fixation, and to inhibit pathogen invasion (64–67). The last of these is particularly important as plant pathogens can result in substantial losses for producers, and these novel mitigation approaches might offer an alternative to the traditional use of phytosanitary products that can be toxic to soils, water bodies, and even consumers. However, more detailed information about the effectiveness of microbial consortia, including potentially synergistic and antagonistic/competitive interactions between strains, under operational conditions is essential for future implementation (68).
As noted above, the interaction between soil–plant microbiomes is not limited only to the below-ground niches. In fact, colonization occurs primarily at three distinct levels, i.e., the rhizosphere (zone of influence of root exudates), endosphere (internal plant compartments), and phyllosphere (aerial plant surfaces) (37, 69). Changes in climate and agricultural management also impact the phyllosphere microbiome, which is involved in diverse functions such as pathogen suppression, tolerance to extreme temperatures, and nitrogen fixation (70, 71). For example, water stress, fertilization, and arbuscular mycorrhizal colonization can affect the plant phyllosphere (72) and, a large proportion of leaf bacterial species are shared with those in the soil, thereby representing an example of microbiome transmission with implications for foodborne pathogen transmission (73). These findings provide evidence that below-ground microbial interactions and abiotic conditions can shape above-ground microbial communities and show the importance of understanding the whole plant microbiome in the context of developing sustainable agriculture solutions.
There are also strong connections between soil, food crops, and animal microbiomes (13, 74). Ruminant livestock are a very important component of agriculture due to their ability to feed on grass, crops, or fiber-rich crop residues not digestible by humans—often utilizing land not suitable for human food production (75). The common practice of companion cropping in pasture, such as the inclusion of clover, results in distinctive soil microbiome profiles and can increase yields by improving the efficiency of nitrogen fixation (76). As a result, a lower usage of fertilizers is needed, which in turn improves water quality in surrounding water bodies owing to reduced nitrogen runoff. Also, the re-inoculation of endophyte isolates can enhance drought tolerance and improve the growth of pasture and forage crops (77, 78). The application of animal manure instead of chemical fertilizers can also improve fodder crop yields and the nutrient efficiency of the system. However, this practice presents several risks that have led to increasingly restrictive regulations on its use. More specifically, while soil microbial communities decompose manure to release nutrients for plant and microbes, and this process temporarily alters the composition of these communities (79), the introduction of microorganisms through manure treatments on the grass phyllosphere and soil microbiome can have other implications of even greater concern, such as the spread of antimicrobial resistance between livestock and grassland. Manure application can introduce bacteria harboring antimicrobial resistance genes (ARGs) into soil and grass microbiomes, increasing also the risk of horizontal gene transfer within these microbial communities. These ARGs can ultimately be transmitted back to livestock through grazing or other interactions with the contaminated environment, highlighting the importance of understanding microbiome interconnections in order to mitigate biological risks (80, 81). Additionally, manure application can cause an excessive nitrogen input and, if not well managed, increased nutrient levels in run-off waters from the field that end up in the aquatic environment, seriously damaging biodiversity. Indeed, it has been estimated that, in 2018, manure contributed to around 55 million tonnes of nitrogen, either being dispersed in the air, mostly as ammonia gas, or leached in water bodies. Also of importance, nitrous oxide emissions from livestock manure left on pasture are the largest source of greenhouse gas emissions after methane from enteric fermentation (82).
A final example relates to green foliage crops that are commonly preserved as silage through fermentation for use as animal feed. Even though this is a traditional practice worldwide, there is potential to improve the nutritional and functional quality of silage using specific lactic acid bacteria, with consequent benefits for animal health and welfare (83). An important component within this process is the microbial community within the associated phyllospheres, which have been shown repeatedly to influence silage fermentation processes and final product characteristics (84, 85). Unsurprisingly, the introduction of silage into the diet of cattle has an effect on growth performance and the rumen microbiome (86). There remains considerable room for further optimization of these processes through the identification of effective strains, consortia, and/or modes of delivery that could provide persistent benefits across a range of crops and environments.
Aquatic environments
When looking at the effect that primary agricultural production can have on the environment, it is important to include potential impacts on both terrestrial and marine aquatic environments (Figure 3A). Recent technological advancements have allowed scientists to gain a better insight into bioprocesses performed by the marine microbiome that are essential to the health of the planet, such as oxygen production and global biogeochemical cycles (87). Moreover, host-mediated microbiome interactions can also be important for the restoration of degraded marine ecosystems. For example, the microbiomes associated with filter feeders, such as mussels, sponges or cnidarians, can nurture and contribute to the recovery (and protection) of damaged marine habitats (88).
As noted above, fertilizer use is particularly important to the effect of primary food production on the aquatic environment. Fertilizers can have a considerable effect on aquatic environments, especially in rivers and lakes where fertilizer run-off is more concentrated than at sea. The unintended effect of inappropriate fertilizer use can be severe disruption to the existing aquatic microbiome by the spread of pathogens (89, 90), as well as the dissemination of antimicrobial resistance genes (91). Fertilizer spread can also cause the overgrowth of particular species at the expense of others and can change nitrogen and phosphorus cycling, potentially causing severe alterations to aquatic microbiomes (92). Due to the risk of these damaging effects, both direct and indirect, fertilizer use must be monitored, well managed, and clearly regulated.
Aside from the study of unintended impacts on aquatic microbiomes, these communities have been the focus of ever-greater interest due to the growth of the aquaculture production industry. Notably, over the last 50 years, aquaculture-based production of fish and seafood has increased four-fold, overtaking the wild fish catch biomass, and so reducing the pressure on wild fish catches to support the growing population (93). This is important, as excessive fishing of wild fish can cause substantial harm to aquatic ecosystems by disrupting ecological balances. Regulations such as protected areas, types of fishing equipment allowed, fish catch limits, and size exclusion criteria are essential to ensuring population stability and biomass protection (94, 95). However, despite the benefits of aquaculture, associated practices can also have multiple noteworthy effects on marine waters and on the general microbiome of waters as a result of temperature and pH changes, nutrient availability, microplastics, or pest proliferation (96–98). This type of human activity has been shown to increase microbial load in water and lead to algal blooms that cause oxygen depletion and large-scale fish kills, thereby ruining entire ecosystems (99).
Other factors polluting aquatic environments include direct microbial contamination, resulting in an increase in microbial numbers (100, 101), and indirect contamination, such as by microplastics, pharmaceuticals, pesticides, and fertilizers—as next-generation pollutants (96, 102). The use of antimicrobials in agriculture, aquaculture, and clinical applications—especially when not necessarily needed—all contributes to an increase in the antimicrobial stress on aquatic system bacteria (103). Disruption of the aquatic microbiome can result in changes to the overall balance of aquatic species, including changes to plant-microbiome associations, which are essential for the general health of the aquatic ecosystem (104). One clear marker of aquatic disruption is coral decline. Although corals occupy a relatively small area of the ocean, they are a vital part of the marine ecosystem and their decline is of particular concern (105). The special sensitivity of corals to environmental changes is related to their nutritional dependency on a symbiotic association with populations of photosynthetic dinoflagellates. In this context, the identification of a possible pathway to recover damaged coral microbiome after the removal of algal stress shows the potential of microbiome-based studies in developing strategies to restore previously damaged ecosystems (106). In parallel, being able to monitor the bacterial communities of coral can allow a better understanding of the microbial shifts that have happened in response to human activity—including global temperature shifts, increasing ocean acidity, and higher aquatic microbial loads—and hopefully enable human intervention to help preserve these delicate bionetworks (107).
In this regard, the use of beneficial microorganisms is a promising strategy to help corals and sponges to respond to anthropogenic stressors (108).
Technological advances offer considerable opportunities to monitor aquatic microbiomes, including modelling shifts in fish communities (109) and using the skin and gill microbiota as indicators of fish gut health (110). One particularly interesting example is the application of novel sequencing technologies in the Continuous Plankton Recorder (CPR) survey, a major sampling programme implemented in 1931 and one of the oldest in the world (111). The use of molecular technologies for analysis in the CPR offers a significant opportunity to majorly advance our knowledge of global marine ecosystems. Indeed, pico- and nano-plankton, which could not be studied until recent decades, can be used as general indicators of marine health and provide a basis for the formation of marine policy (112). Although the insights from such surveys are not always immediately translatable to aquaculture settings, it is to be hoped that an ever-greater understanding of marine microbiomes will ultimately be valuable in a broad variety of ways. Thus, as nations worldwide prioritize sustainable environmental practices, integrating metagenome-based solutions into public policies regarding the health of aquatic ecosystems has the potential to improve the outcomes of conservation and restoration efforts.
Post-harvest, processing, and distribution environments
Fruit, vegetable, and cereal production
Continuing our overview of the interconnectedness between specific microbiomes in the food system (Figure 1), we now shift the focus from primary production to the interactions occurring during the distribution, transformation, and commercialization of produce.
Fruits and vegetables can have quite direct routes from the field to the consumer (Figure 3B). These types of food are often minimally processed and, if they are not subjected to microbial decontamination methods, can retain much of the microbiome present during harvest (113, 114). Indeed, the term “edible plant microbiome” has been used to describe these communities (115, 116). However, these microbiomes can be impacted by a variety of factors such as storage conditions, transportation, and/or packaging. These are particularly important in light of the increased demand from consumers for products that are out of season and/or not available locally due to the climate of a given region (117–119). Despite advances in the storage and distribution of these products, it has been estimated that globally between 25 and 50% of fruits and vegetables are lost post-harvest (120), emphasizing the importance of spoilage prevention to achieve a more sustainable food system. Improved strategies to monitor spoilage microbiota and food safety verification systems, such as strain-resolved metagenomics, have the ability to provide information regarding, for example, the presence of spoilage and pathogenic microbes, source attribution, temporal, and geographic distribution and cross contamination (121). This knowledge can then be harnessed to improve post-harvest storage practices. For example, using a biological control approach (122–124) technological advances in high-throughput sequencing methods can facilitate a more targeted identification of antagonistic bacteria and fungi for disease and spoilage postharvest control based on a deeper understanding of microbial interactions on epiphytic microbiomes (125, 126). Some specific recent examples have included the identification and successful application of biocontrol agents to decrease postharvest problems with strawberries and peaches (31, 127). This highlights the importance of collaborative efforts between academic research and industry stakeholders in order to translate metagenomic research into real-world applications. The practical experience and insights provided by the industries can help identify the main challenges in the sector and the best ways to apply the knowledge and cutting-edge technology necessary to successfully implement novel microbiome-based solutions.
Not all fruits and vegetables are consumed directly—some are prepared/processed in multiples ways, for example to produce juices, ready-to-eat salads, or fermented foods. Storage time and transportation between harvest and processing/fermentation can affect the final microbiome diversity and microbial load of the product, as shown for example in a tracking study developed on romaine lettuce (128). Often, and particularly for more complex produce distribution chains, a decontamination step is typically included to ensure the safety of the final product that arrives to the consumer. In this context, microbial distribution and interactions inside vegetable processing plants become relevant and the recent application of microbiome mapping as part of routine monitoring programs in food industries has shown potential as a way to decrease spoilage-related issues and ensure food safety (129). In the case of fermented products, the microbiota of the substrate and processing environment can become especially relevant if a spontaneous fermentation is involved (130).
Terrestrial farm animal production
It has been estimated that the terrestrial animal biomass (Figure 3) needs to almost double by 2050 in order to meet the global demand predicted by the UN Food and Agricultural Organisation (131). Farm animal microbiome research and its applications will be of key importance in the move towards more environmentally friendly intensification of animal husbandry on Earth. An increased understanding of microbial interactions in the digestive tract of production animals could allow for the development of diet-based modulation strategies to improve their metabolism, immune system, and overall feed efficiency (132). Notably, the application of omics technologies to the investigation of cattle rumen microbial composition and functionality patterns has greatly helped to unravel its role in the relationship between nutrition, nitrogen excretion, and methane emission (133–138), with a view to the development of new intervention products and strategies. For example, in beef cattle and lambs, diet has a direct effect on the rumen microbiome, with the abundance of certain genera of bacteria and archaea being a potential predictor of growth performance (139, 140). Interventions based on diet supplementation with prebiotics, probiotics, or a combination of both have been developed for different types of livestock, including dairy calves, bulls, sheep, and camels, generally showing positive effects on the rumen microbiome and improving general feed efficiency and other health markers (141–145). Diet can also affect the production of the greenhouse gas methane, a by-product of the fermentation of complex carbohydrates by methanogenic archaea in the rumen, with considerable implications for global warming (146–148). Notably, greenhouse gas emission intensities show geographic differences being lower in areas with a high level of specialization and productivity and higher in regions with lower quality of feed, reproduction efficiency, herd management practices, and animal genetics (149).
Microbiome interconnections are not unidirectional, and livestock grazing has been shown to influence the microbiome and productivity of soil. Grazing livestock species (cattle, sheep, swine, or multi-species) can impact soil microbial community structure and antibiotic resistance gene profiles in a variety of ways (150) and, over the long-term, intense grazing can detrimentally decrease soil carbon and nitrogen mineralization rates (151, 152).
As for plant-based foods, high-throughput sequencing technologies have also provided a better understanding of contamination routes in foods of animal origin (153–156). In the dairy sector, for example, microbiome-based studies have shown the influence of the grazing system on the microbiota of cow teat skin, which can be a reservoir of many microorganisms found in raw milk (157). Similarly, the microbial exchange between feed, bedding material, and cow teat skin was shown by the detection of live yeast provided as a supplement in a grass-diet system (158). Given the importance of the teat canal and the udder surface as a source of milk contamination and intramammary infections by mastitis pathogens, the effectiveness of different disinfection protocols has been extensively evaluated (159, 160). There is growing agreement that a better understanding of the mammary microbiota can lead to the identification of improved therapeutic products and disinfection protocols that favour beneficial microorganisms (161, 162). Importantly, while this review focuses on non-pathogenic microorganisms, microbial transfer is also critically important to the transmission of zoonotic pathogens such as Listeria monocytogenes, Salmonella, and Shiga-toxin producing Escherichia coli from plant or animal sources, as reviewed elsewhere (163).
Microorganisms can end up within the microbiome of food products via the raw ingredients or the food processing environment (Figure 3B). An example of the former relates to cheese production, whereby some non-starter lactic acid bacteria (lactic acid bacteria that do not form part of the added starter culture) originating from the milk became part of the cheese microbiome, being associated with positive or negative quality outcomes after cheese ripening (164). The pathways of microbial transfer along the cheese-making process have been described on several occasions, gaining special importance in the case of traditional raw milk cheeses where non-starter bacteria can shape the fermentation and ripening processes (165, 166).
Similarly, the routes of microbial transmission into and inside processing facilities have also been studied using next-generation sequencing approaches, for example, in the meat industry (167–169). In this way, facility-specific transmission maps can be created, helping to predict previously unidentified sources of bacterial contamination (170). Other successful applications of this approach include the longitudinal tracking of the divergence and spread of foodborne pathogens (171) and studies of the microbial colonization of newly open facilities identifying routes of entrance of pathogens and antibiotic-resistant microorganisms (172, 173). Despite these advances, investigations into the transmission of foodborne pathogen outbreaks continue to draw conclusions based on culture-dependent approaches combined with whole genome sequencing (24). Once again, academia–industry partnerships encourage knowledge exchange and contribute to research whose outcomes address critical issues in food production and are effectively integrated into industry practices, accelerating the adoption of metagenome-based innovations in the food system (174).
Aquaculture
In aquaculture (Figure 3), the microbiomes of fish can have a major impact on health and growth rates and should thus be of upmost concern for the associated production industry to prevent losses and maximise production. Modern sequencing technologies can greatly advance aquaculture microbiome monitoring (175), as the identification of a core microbiome can be essential in understanding the health of an aquaculture population and the bidirectional interactions between aquatic microbiomes and aquaculture (176). Importantly, while pathogens can be a natural part of an ecosystem they can also be extremely damaging to aquaculture and so need to be carefully observed, with interventions made when appropriate to prevent infection and production loss. Careful monitoring of multiple factors is vital to aquaculture, as the microbiota can be affected by numerous factors, including diet, disease, oxygenation, pH, and temperature (177, 178). The use of aquafeed needs to be carefully examined to ensure that it is providing the necessary nutrients for its intended recipients but is also free from contaminants, including fungal contamination and mycotoxins, which can spread beyond the aquaculture environment to the wild aquatic environment (179). Aquaculture can also damage aquatic environments via algal blooms derived from changes in water conditions and nutrient availability, and because intensive fish farms are vulnerable to infections that can spread into the wild (90). Probiotics are commonly used in aquaculture, especially particular strains of bacilli (180, 181), lactobacilli (182), and bifidobacteria (183). Certain strains have been shown to protect against disease and infection, and to improve the general health of the aquaculture fish and seafood (184). Antimicrobial use is sometimes necessary to prevent infections becoming pervasive but, as in other environments, their use can have unintended but severe consequences, including the increase of antimicrobial resistance in the environment (185–187).
Moving to fish processing, food safety and spoilage can be a major issue, especially considering the short shelf life of seafood. Although the shelf life of fish and shellfish has been significantly improved with advances in refrigeration and freezing technologies, fish losses are still estimated at 27% between landing and consumption (188). The addition of microorganisms can prevent spoilage through multiple mechanisms. Microorganisms can be added directly to act as a bioprotectants against foodborne pathogens (30), while bacteriocins (189) and bacteriophages can also be used to help control unwanted pathogens and spoilage organisms (190).
The improved understanding of aquaculture microbiomes is also being applied to develop novel monitoring and mitigation strategies. These include: investigations of the potential of gut microbiomes to mitigate disease in aquaculture (191); harnessing knowledge relating to the skin microbiome of fish to predict population collapse (192); using machine learning models to predict fish kills and toxic blooms in intensive aquaculture (193); and using seawater transfer to improve microbiome diversity (194) and the removal of ammonium and nitrite (195). While microbiome-informed practices and innovations can help to protect natural microbiomes, they can also offer advantages to the aquaculture industry itself. Indeed, such studies have shown that common practices in aquaculture, such as the use of bleaching powder to disinfect marine water, may not have the previously presumed effect as aquatic microbiomes treated in this way have been seen to recover quickly (196). Ideally, advances in our understanding of aquatic microbiomes can be leveraged towards a more efficient production system while, at the same time, reducing damage to the wider ecosystem.
The future growth of the aquaculture sector will be sustainable only if the industry transitions to more eco-friendly approaches that minimise environmental impacts, particularly in terms of reducing eutrophication in marine water. Integrated multi-trophic aquaculture (IMTA) provides one concrete solution, as a sustainable approach to intensive aquaculture. IMTA combines fish farming with the growth of lower-trophic species that can recycle/adsorb organic and inorganic nutrients to increase the eco-compatibility of the aquaculture practice. Crucial to its success is the selection of the appropriate combination of productive and nutrient-extractive species that complement each other from a trophic perspective. This enables novel eco-designed solutions that also integrate marine microbiomes as an innovation in the IMTA design. The selected species contribute to an overall holobiont solution to nutrient cycling. Indeed, many microbiomes—environmental and host-associated—can exert a primary role in the recirculation of organic matter in marine ecosystems, and their targeted integration at the metacommunity level in IMTA systems has the potential to lead to even more efficient reductions of ecological footprints (197).
Retail and household environments
Food uptake impacts all levels of physical and, in higher organisms, mental health. Access to a reliable food supply facilitates the development of societies and stabilizes political systems. The corollary is also true with disturbance of the food supply system frequently leading to major human crises, including, as an extreme, famine and mass starvation. Although enough food is produced on earth to feed the entire population, the biggest challenge in modern food production remains the logistics behind the food distribution system that largely determines whether food is consumed or discarded (198, 199). In developing countries, which lose up to 50% of their primary production due to spoilage and mismanagement, major distribution failures result from a lack of power and storage capacity during and after harvesting campaigns. In developed countries, food is discarded primarily due to shelf life issues and inappropriate buying patterns (200). At the retail level, food storage is the most essential economic parameter, especially for perishable food, and most of the costs of a supermarket or retail premise go into the energy supply for food cooling and presentation (Figure 3B). Understanding the microbial dynamics during cold storage of intermediate and final products is of utmost importance to lower the levels of foods that are discarded to an acceptable level. A special advantage for microbiome-based interventions lies in the enhanced capacity to detect fastidious growing spoilage-associated microbes that are usually difficult to detect by culture-based technology (201).
The issues raised above also pertain to households (Figure 3B). More and more households store food for an extended period of time due to the conditions of the modern working world. Knowledge relating to spoilage processes and control of the cooling processes in household refrigerators is still surprisingly scarce, even though a link between microbial communities found in the kitchen and the skin microbiome of humans has been described (202). Microbiome tracking strategies can also be used to detect routes of contamination in consumer households to, for example, show the importance of refrigerated storage of products that could limit the proliferation of opportunistic human pathogens (203). These approaches have also been taken even further to track the movement of lactic acid bacteria, such as Weissella and Leuconostoc species, from ready-to-eat salads to the human gut after their consumption (204, 205). Although microbiome tracking approaches continue to be applied predominantly for academic research, as costs and other barriers are reduced, it is expected that these approaches will be used more widely by industry and, eventually, consumers.
The role of bacteriophages in shaping microbial communities
While the bacterial and fungal components of food production systems and food products have been widely studied, the viral component of these ecosystems is less well defined. Bacteriophages (or phages) are bacterial viruses that typically infect strains of a given bacterial species. Phages may be considered beneficial in certain situations, for example, in the elimination or reduction of human or animal pathogens, or spoilage bacterial numbers (206). Conversely, phages that infect starter bacterial cultures are a major threat to food fermentations as they may compromise the acidification or flavor development of the product. In recent decades, studies have focused on defining the phage populations in food fermentation facilities relating to lactic acid bacteria such as Lactococcus lactis/cremoris and Streptococcus thermophilus (207–209). These studies initially used culture-based approaches to tease apart the interactions of phages and their hosts. In the context of phages, this requires the ability to perform plaque assays. More recently, high-throughput DNA sequencing has allowed a more unbiased examination of the viral component of the microbiome. The role of the virome, and more specifically the “phageome”, in influencing bacterial compositional changes in microbiomes has garnered increasing research attention (210–212). From an evolutionary perspective, the phages and their bacterial hosts have been in conflict for millennia, and recent studies have allowed us to question the mode of these interactions and their impact on microbiome maintenance, modulation, and functionality, including through horizontal gene transfer across food system niches (213–216).
One such example is the initial interaction between a phage and its host, dictated by the presence of a suitable receptor on the cell surface of the host bacterium, which may be a carbohydrate, protein, or (lipo)teichoic acid. These early interactions are exceptionally well studied in a small number of bacterial host systems, including specific spoilage and pathogenic species such as E. coli (217), L. monocytogenes (218, 219) and beneficial starter bacterial culture species such as Lactococcus lactis/cremoris (220) and Streptococcus thermophilus (221). These foundational studies have provided insights into generalized and specialized modes of host recognition and subsequent infection by phages of these species, and which may be applied to the development of tools to improve the consistency, sustainability, and safety along the food production chain. Combining these studies with developments in sequencing, computer-assisted prediction of the folding and functionality of structural proteins, and imaging is allowing further elucidation of these interactions and serves as a blueprint to studying the interactions of players within a microbiome.
The impact of microbiome interconnections and interventions on human health
One major area greatly enhanced by recent advances in multi-omics approaches is the study of the human gut microbiota and its contribution to human health. Many factors can influence the human gut microbiota, including genetics, geography, socioeconomic status, age, and health status. Notably with respect to this article, diet is a key modulator and diet–microbiome interactions are often at the forefront of microbiome research.
Having a diverse gut microbiome is frequently associated with health, potentially through protection against pathogens and disease (222, 223). However, this diversity can be affected by multiple factors. There is evidence that a modern, Western diet (high in simple carbohydrates, saturated fats and salt, but low in fiber, complex carbohydrates, and micronutrients) has a damaging effect on the gut microbiome and can contribute to multiple health problems including cancer, obesity, gastrointestinal and gut–brain-related issues (224, 225). Specific ultra-processed foods that are high in fat, sugar and/or salt are of particular note as an issue in a modern, Western diet. These foods often lack microorganisms, complex nutrients, prebiotics, and fiber, can contribute to decreased gut microbiome diversity and, in turn, various health issues (226).
There is growing evidence of the occurrence of food-to-gut microorganism transmission (116, 227). Although necessary to ensure food safety in large-scale production, many food processing techniques remove most or all naturally occurring microorganisms, thus contributing to an altered human gut microbiome and an increase in non-communicable diseases (228). Historically, food was sourced from the local area and was subject to seasonal variation. Some of this food would have been completely unprocessed and consumed quickly so microorganisms naturally present in the food were introduced to the consumer gut microbiota. Advances in processing, refrigeration, storage, and transport techniques have allowed for more food to be sourced from further afield and have allowed for foods not to be restricted to a particular season. However, these advantages in food availability are also contributing to the decrease in diversity in human gut microbiota as many of these foods now need to have a longer shelf life and many techniques to prevent spoilage and pathogenic activity also decrease or completely remove the indigenous microbiota of the foods. Similarly, the higher demand for convenience foods, for example, a pre-packaged, pre-washed salad in contrast to unprocessed salad, has further distanced the consumer from food’s naturally occurring microbiota. In any case, the safety of the food that reaches the consumer is of utmost importance. While the emphasis of this review is on the broader microbiome interconnections between food and the human gut, it is important to acknowledge that the food system also serves as a route of transmission of opportunistic human pathogens (229, 230).
In addition to live microorganisms, prebiotics and bioactive compounds are other components in food that can alter the gut microbiota. Prebiotics, such as certain fermentable fibers, polyphenols, and oligosaccharides, can promote positive effects on health through gut microbiota alteration (231–233). However, modern Western diets are often deficient in fiber and contain an overabundance of fat and sugar (234). The high proportion of meat in the modern Western diet has also been seen to decrease gut microbiota diversity, while plant-based diets have been shown to be advantageous to human health (235). Nevertheless, it is important not to overlook the possible nutrient inadequacies that can be associated with different dietary patterns, including vegan, vegetarian, and meat-based diets (236, 237). Bioactive compounds can contribute to human health, including via antioxidant and anti-inflammatory effects, and can come from a variety of sources, including marine algae-sourced bioactive compounds (238, 239). A relatively underutilized source of bioactive compounds are those resulting from food wastes and by-products, which could offer benefits without the need for further alterations to natural microbiomes through additional production streams (240).
Another diet-based approach that can beneficially impact gut microbiome diversity and function is through the consumption of fermented foods. Fermented foods have been widely shown to be advantageous to the gut microbiota through the promotion of a diverse microbiota and the potential to simultaneously provide prebiotics, probiotics, and bioactive compounds (241). Indeed, the European Food Safety Authority (EFSA) has approved the claim that live yoghurt cultures improve yoghurt lactose digestion in individuals with lactose maldigestion (242). Although some fermented foods have been produced over hundreds or thousands of years, many are not yet well characterized (243). Foods and feeds produced by spontaneous fermentation have been shown to typically result in products that contain very similar microbial taxa and applying ecological and evolutionary frameworks can be valuable with respect to understanding the assembly of fermented foods microbiomes (244, 245). Indeed, establishing a better understanding of such microbiomes, and the strains present therein, may offer advantages to human health that have not yet discovered. Some of these benefits may relate to the consumption of live microbes specifically (246–248). In addition, the consumption of specific probiotic strains, added to foods or taken as supplements, can contribute to reducing the incidence of particular human diseases (249–252).
Conclusions
Food system microbiomes form complicated and connected networks that can be very difficult to disentangle. Advances in technologies, especially in high-throughput sequencing technologies, have enhanced our understanding of these networks. Developments, especially in soil, plant, animal and marine microbiome research, have been instrumental in the recent progress of agriculture and aquaculture practices, including practical management strategies and interventions for more sustainable and resilient food systems (Table 1). However, understanding is only one step in the process of protecting and promoting the health and diversity of our microbiomes. Advances of knowledge, facilitated by new technologies, have illuminated many issues in microbiome networks, including reductions in microbial diversity and the spread of antimicrobial resistance.
Table 1
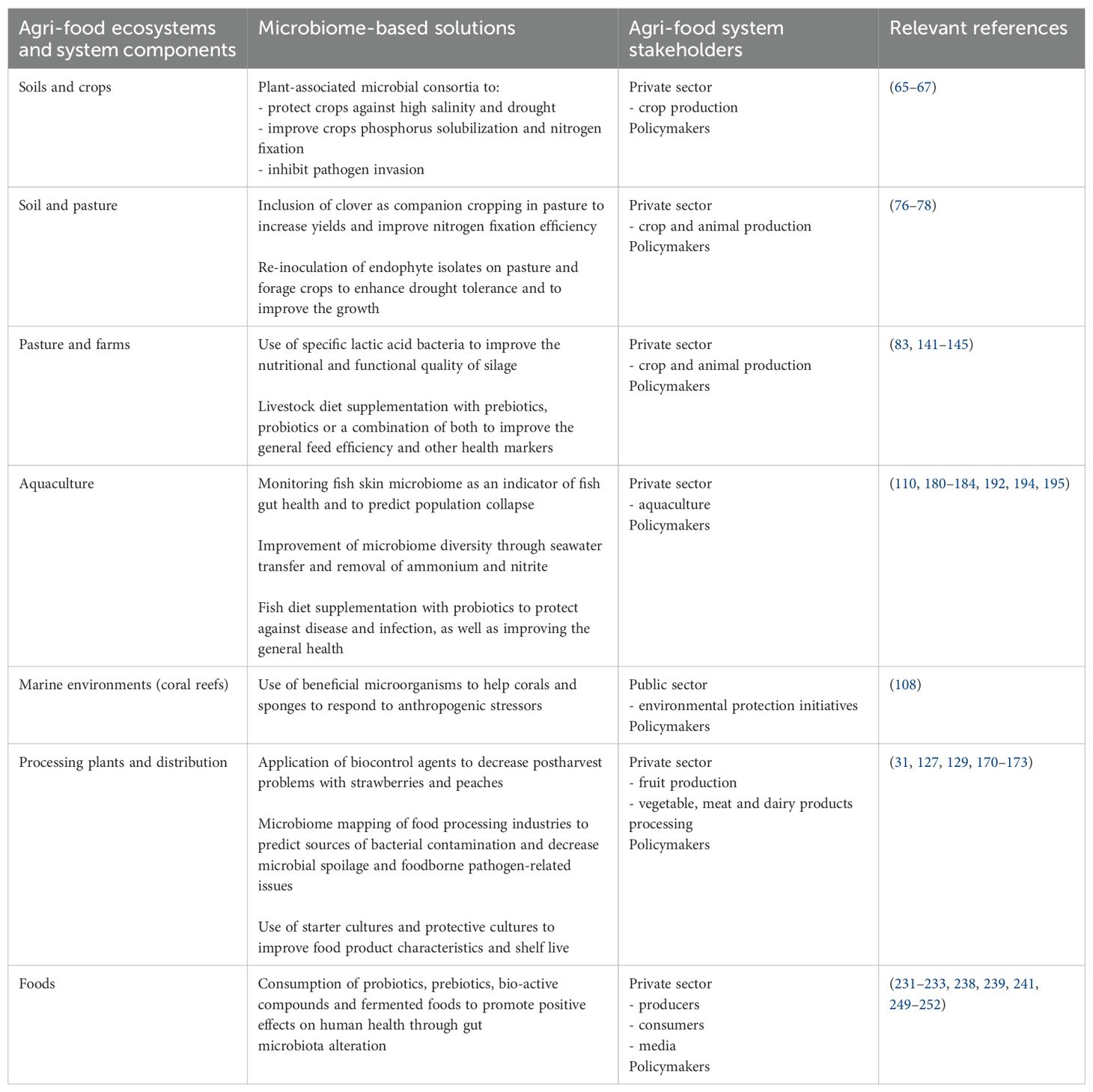
Table 1. Potential microbiome-based solutions for different sectors across the global agri-food system and the private and/or public stakeholders involved.
In addition to ensuring continuous advances in the understanding of food system microbiomes, developing a greater understanding of their composition, functionality, and interactions within and across ecosystems and the development of microbiome-based applications is necessary to fully leverage microbiome potential. In this regard, it is also essential to ensure that other framework conditions are met, e.g., societal awareness and understanding, and the establishment of a suitable regulatory landscape. Further promoting microbiology and microbiome education among different stakeholder groups would enable them to make well-informed decisions about microbiome-related products (21, 253). This is not only important for the end-users but also for the industry developing new microbiome-based products/applications and for the regulators who must assess them. The challenges faced by regulators are multifaceted. On the one hand, there is an absence of explicit legal requirements to account for microbiome-related effects when risks/benefits are considered (254). On the other hand, the roadmap for the approval of microbiome-based innovations is particularly complex because microbiome science has been generated in the absence of standards and validated methodologies (255). The implementation of a systemic approach involving all relevant stakeholders is vital to aid in protecting existing microbiomes and restoring damaged microbiome networks relating to the food system (256, 257).
While understanding microbiome composition is increasingly achievable, deciphering the functional roles of microbiome constituents remains a challenge. Culture-based approaches are crucial for translating this research into innovations and for understanding the causality behind correlations. Similarly, the generation and integration of omics data needs to be complemented with synthetic biology, high-throughput screening, and targeted experimental approaches in order to validate the functions of these microbiomes. These strategies can enable the translation of microbiome research into innovations within food systems, with the potential to improve human and animal health as well as the health of the planet at large.
Statements
Author contributions
PF-G: Conceptualization, Visualization, Writing – original draft, Writing – review & editing.
DL: Conceptualization, Visualization, Writing – original draft, Writing – review & editing.
GB: Writing – original draft, Writing – review & editing.
FB: Writing – original draft, Writing – review & editing.
TC: Writing – original draft, Writing – review & editing.
TCC: Writing – original draft, Writing – review & editing.
LC: Writing – original draft, Writing – review & editing.
LL: Writing – original draft, Writing – review & editing.
OM: Writing – original draft, Writing – review & editing.
EM: Writing – original draft, Writing – review & editing.
OO’S: Writing – original draft, Writing – review & editing.
YS: Writing – original draft, Writing – review & editing.
IS: Writing – original draft, Writing – review & editing.
AS: Writing – original draft, Writing – review & editing.
HS: Writing – original draft, Writing – review & editing.
NB: Writing – original draft, Writing – review & editing.
MC: Writing – original draft, Writing – review & editing.
JK: Writing – original draft, Writing – review & editing.
TK: Writing – original draft, Writing – review & editing.
JM: Writing – original draft, Writing – review & editing.
MW: Writing – original draft, Writing – review & editing.
PC: Conceptualization, Visualization, Writing – original draft, Writing – review & editing.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Funding
The authors declared financial support was received for this work. Research in the group of PC was funded through Science Foundation Ireland (SFI) under grant number (SFI/12/RC/2273) (APC Microbiome Ireland), and SFI together with the Irish Department of Agriculture, Food and the Marine (SFI/16/RC/3835) (VistaMilk). Research in the Cotter laboratory was funded Tby Friesland Campina, PrecisionBiotics Group, PepsiCo and Danone.
PC received support from Abbott, PepsiCo, Yakult, Lallemand and H&H to attend/present at scientific meetings/conferences.
PF-G received funding from the European Union’s Horizon 2020 Research and Innovation Programme under the INSPIRE COFUND Marie Skłodowska Curie grant agreement (No. 101034270).
Conflict of interest
FB and OOS were employed by VistaMilk Science Foundation Ireland (SFI) Research Centre, a research center which collaborates with industry partners as disclosed in their web page (https://www.vistamilk.ie/industry-partners-2/).
LL was employed by LL-BioEconomy, Research & Advisory.
AS was employed by AIT Austrian Institute of Technology GmbH, research centre funded by both the government of Austria (majority shareholder) and the Federation of Austrian as listed on their web page (https://www.ait.ac.at/en/about-the-ait).
MW was employed by FFoQSI GmbH - Austrian Competence Centre for Feed and Food Quality, Safety and Innovation, a research center which collaborates with industry partners as disclosed in their web page (https://ffoqsi.at/index.php/en-us/about-ffoqsi/partner-en).
Research in the Cotter laboratory was funded by Friesland Campina, PrecisionBiotics Group, PepsiCo and Danone.
PC received support from Abbott, PepsiCo, Yakult, Lallemand and H&H to attend/present at scientific meetings/conferences and is the Head of Microbiology and a co-founder of SeqBiome Ltd.
Neither the funders nor the non-funding companies were involved in the study design, collection, analysis, interpretation of data, the writing of this article, or the decision to submit it for publication.
The remaining authors declared that this work was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer MG declared a past co-authorship with the author PC to the handling editor.
The authors DL, GB, TC, TCC, LC, LL, EM, OO’S, HS, and TK declared that they were an editorial board member of Frontiers at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declared that no generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Davis B, Mane E, Gurbuzer LY, Caivano G, Piedrahita N, Schneider K, et al. Estimating global and country-level employment in agrifood systems. In: Food and Agriculture Organization of the United Nations. FAO statistics working paper series/ 23–34. Rome: FAO (2023). doi: 10.4060/cc4337en
2. Food and Agriculture Organization of the United Nations. The state of food and agriculture 2023 – revealing the true cost of food to transform agrifood systems. Rome: FAO (2023). doi: 10.4060/cc7724en
3. Food and Agriculture Organization of the United Nations. World food and agriculture – statistical yearbook 2023. Rome: FAO (2023). doi: 10.4060/cc8166en
4. Richardson K, Steffen W, Lucht W, Bendtsen J, Cornell SE, Donges JF, et al. Earth beyond six of nine planetary boundaries. Sci Adv (2023) 9(37):eadh2458. doi: 10.1126/sciadv.adh2458
5. United Nations Environment Programme. Food waste index report 2024. Think eat save: tracking progress to halve global food waste. Nairobi: UNEP (2024). Available at: https://www.unep.org/resources/publication/food-waste-index-report-2024
6. United Nations Environment Programme. Sustainable consumption and production indicators for the future SDGs. UNEP Discussion Paper - March 2015. Nairobi: UNEP (2015). Available at: https://www.unep.org/fr/node/10326.
7. Organization for Economic Co-operation and Development, Food and Agriculture Organization of the United Nations. OECD-FAO agricultural outlook 2023-2032. Paris: OECD (2023). doi: 10.1787/08801ab7-en
8. Berg G, Rybakova D, Fischer D, Cernava T, Vergès M-CC, Charles T, et al. Microbiome definition re-visited: old concepts and new challenges. Microbiome (2020) 8(1):103. doi: 10.1186/s40168-020-00875-0
9. Rillig MC, Antonovics J, Caruso T, Lehmann A, Powell JR, Veresoglou SD, et al. Interchange of entire communities: microbial community coalescence. Trends Ecol Evol (2015) 30(8):470–6. doi: 10.1016/j.tree.2015.06.004
10. Sessitsch A, Wakelin S, Schloter M, Maguin E, Cernava T, Champomier-Verges M-C, et al. Microbiome interconnectedness throughout environments with major consequences for healthy people and a healthy planet. Microbiol Mol Biol Rev (2023) 87(3):e00212–22. doi: 10.1128/mmbr.00212-22
11. Chevallereau A, Pons BJ, van Houte S, and Westra ER. Interactions between bacterial and phage communities in natural environments. Nat Rev Microbiol (2022) 20(1):49–62. doi: 10.1038/s41579-021-00602-y
12. World Health Organization. One health [online] (2023). Available at: https://www.who.int/health-topics/one-health
13. Law SR, Mathes F, Paten AM, Alexandre PA, Regmi R, Reid C, et al. Life at the borderlands: microbiomes of interfaces critical to One Health. FEMS Microbiol Rev (2024) 48(2):fuae008. doi: 10.1093/femsre/fuae008
14. Ferrocino I, Rantsiou K, McClure R, Kostic T, de Souza RSC, Lange L, et al. The need for an integrated multi-OMICs approach in microbiome science in the food system. Compr Rev Food Sci Food Saf (2023) 22(2):1082–103. doi: 10.1111/1541-4337.13103
15. Jagadeesan B, Gerner-Smidt P, Allard MW, Leuillet S, Winkler A, Xiao Y, et al. The use of next generation sequencing for improving food safety: translation into practice. Food Microbiol (2019) 79:96–115. doi: 10.1016/j.fm.2018.11.005
16. Jiang D, Armour CR, Hu C, Mei M, Tian C, Sharpton TJ, et al. Microbiome multi-omics network analysis: statistical considerations, limitations, and opportunities. Front Genet (2019) 10:995. doi: 10.3389/fgene.2019.00995
17. Layeghifard M, Hwang DM, and Guttman DS. Disentangling interactions in the microbiome: a network perspective. Trends Microbiol (2017) 25(3):217–28. doi: 10.1016/j.tim.2016.11.008
18. Ji C, Guo J, Ma Y, Xu X, Zang T, Liu S, et al. Application progress of culturomics in the isolated culture of rhizobacteria: a review. J Agric Food Chem (2024) 72(14):7586–95. doi: 10.1021/acs.jafc.3c08885
19. Lamas A, Regal P, Vázquez B, Miranda JM, Franco CM, and Cepeda A. Transcriptomics: a powerful tool to evaluate the behavior of foodborne pathogens in the food production chain. Food Res Int (2019) 125:108543. doi: 10.1016/j.foodres.2019.108543
20. Olmo R, Wetzels SU, Armanhi JSL, Arruda P, Berg G, Cernava T, et al. Microbiome research as an effective driver of success stories in agrifood systems – a selection of case studies. Front Microbiol (2022) 13:834622. doi: 10.3389/fmicb.2022.834622
21. Olmo R, Wetzels SU, Berg G, Cocolin L, Hartmann M, Hugas M, et al. Food systems microbiome-related educational needs. Microb Biotechnol (2023) 16(7):1412–22. doi: 10.1111/1751-7915.14263
22. Perkons I, Varunjikar MS, and Rasinger JD. Unveiling the potential of proteomics in addressing food and feed safety challenges. EFSA J (2023) 21(Suppl 1):e211013. doi: 10.2903/j.efsa.2023.e211013
23. Vishwanath PP, Bidaramali V, Lata S, and Yadav RK. Transcriptomics: illuminating the molecular landscape of vegetable crops: a review. J Plant Biochem Biotechnol (2024), 1–6. doi: 10.1007/s13562-023-00871-z
24. Pightling AW, Pettengill JB, Luo Y, Baugher JD, Rand H, and Strain E. Interpreting whole-genome sequence analyses of foodborne bacteria for regulatory applications and outbreak investigations. Front Microbiol (2018) 9:1482. doi: 10.3389/fmicb.2018.01482
25. Feehily C, O’Neill IJ, Walsh CJ, Moore RL, Killeen SL, Geraghty AA, et al. Detailed mapping of Bifidobacterium strain transmission from mother to infant via a dual culture-based and metagenomic approach. Nat Commun (2023) 14(1):3015. doi: 10.1038/s41467-023-38694-0
26. Kordy K, Gaufin T, Mwangi M, Li F, Cerini C, Lee DJ, et al. Contributions to human breast milk microbiome and enteromammary transfer of Bifidobacterium breve. PloS One (2020) 15(1):e0219633. doi: 10.1371/journal.pone.0219633
27. Gibson GR, Hutkins R, Sanders ME, Prescott SL, Reimer RA, Salminen SJ, et al. Expert consensus document: the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol (2017) 14(8):491–502. doi: 10.1038/nrgastro.2017.75
28. Hill C, Guarner F, Reid G, Gibson GR, Merenstein DJ, Pot B, et al. Expert consensus document. The International Scientific Association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol (2014) 11(8):506–14. doi: 10.1038/nrgastro.2014.66
29. Schmitz L, Yan Z, Schneijderberg M, de Roij M, Pijnenburg R, Zheng Q, et al. Synthetic bacterial community derived from a desert rhizosphere confers salt stress resilience to tomato in the presence of a soil microbiome. ISME J (2022) 16(8):1907–20. doi: 10.1038/s41396-022-01238-3
30. Saraoui T, Cornet J, Guillouet E, Pilet MF, Chevalier F, Joffraud J-J, et al. Improving simultaneously the quality and safety of cooked and peeled shrimp using a cocktail of bioprotective lactic acid bacteria. Int J Food Microbiol (2017) 241:69–77. doi: 10.1016/j.ijfoodmicro.2016.09.024
31. Zhimo VY, Kumar A, Biasi A, Salim S, Feygenberg O, Toamy MA, et al. Compositional shifts in the strawberry fruit microbiome in response to near-harvest application of Metschnikowia fructicola, a yeast biocontrol agent. Postharvest Biol Technol (2021) 175:111469. doi: 10.1016/j.postharvbio.2021.111469
32. Alessandria V, Ferrocino I, Carta V, Zuliani V, Seibert TM, Soeltoft-Jensen J, et al. Selection of food cultures with protective properties for cooked ham. Food Microbiol (2023) 112:104218. doi: 10.1016/j.fm.2023.104218
33. Valenzuela-Miranda D, Gonçalves AT, Valenzuela-Muñoz V, Nuñez-Acuña G, Liachko I, Nelson B, et al. Proximity ligation strategy for the genomic reconstruction of microbial communities associated with the ectoparasite Caligus rogercresseyi. Sci Rep (2022) 12(1):783. doi: 10.1038/s41598-021-04485-0
34. Peixoto RS, Voolstra CR, Sweet M, Duarte CM, Carvalho S, Villela H, et al. Harnessing the microbiome to prevent global biodiversity loss. Nat Microbiol (2022) 7(11):1726–35. doi: 10.1038/s41564-022-01173-1
35. Banerjee S and van der Heijden MGA. Soil microbiomes and one health. Nat Rev Microbiol (2023) 21(1):6–20. doi: 10.1038/s41579-022-00779-w
36. Montgomery DR, Rabinowitz P, Sipos Y, and Wheat EE. Soil health: a common focus for one health and planetary health interventions. One Health (2024) 18:100673. doi: 10.1016/j.onehlt.2023.100673
37. Santoyo G. How plants recruit their microbiome? New insights into beneficial interactions. J Adv Res (2022) 40:45–58. doi: 10.1016/j.jare.2021.11.020
38. Wei Z, Gu Y, Friman VP, Kowalchuk GA, Xu Y, Shen Q, et al. Initial soil microbiome composition and functioning predetermine future plant health. Sci Adv (2019) 5(9):eaaw0759. doi: 10.1126/sciadv.aaw0759
39. Song C, Jin K, and Raaijmakers JM. Designing a home for beneficial plant microbiomes. Curr Opin Plant Biol (2021) 62:102025. doi: 10.1016/j.pbi.2021.102025
40. Gu Z, Hu C, Gan Y, Zhou J, Tian G, and Gao L. Role of microbes in alleviating crop drought stress: a review. Plants (Basel) (2024) 13(3):384. doi: 10.3390/plants13030384
41. Martínez-Arias C, Witzell J, Solla A, Martin JA, and Rodríguez-Calcerrada J. Beneficial and pathogenic plant-microbe interactions during flooding stress. Plant Cell Environ (2022) 45(10):2875–97. doi: 10.1111/pce.14403
42. Pistelli L, Libik-Konieczny M, and Hasanuzzaman M. Editorial: plant-microbe interactions and their role in salinity tolerance. Front Plant Sci (2023) 14:1142563. doi: 10.3389/fpls.2023.1142563
43. Roy SA-O, Chakraborty AP, and Chakraborty RA-O. Understanding the potential of root microbiome influencing salt-tolerance in plants and mechanisms involved at the transcriptional and translational level. Physiol Plant (2021) 173(4):1657–81. doi: 10.1111/ppl.13570
44. Goyal RK and Habtewold JZ. Evaluation of legume-rhizobial symbiotic interactions beyond nitrogen fixation that help the host survival and diversification in hostile environments. Microorganisms (2023) 11(6):1454. doi: 10.3390/microorganisms11061454
45. Pang Z, Mao X, Zhou S, Yu S, Liu G, Lu C, et al. Microbiota-mediated nitrogen fixation and microhabitat homeostasis in aerial root-mucilage. Microbiome (2023) 11(1):85. doi: 10.1186/s40168-023-01525-x
46. Zhang L, Zhang M, Huang S, Li L, Gao Q, Wang Y, et al. A highly conserved core bacterial microbiota with nitrogen-fixation capacity inhabits the xylem sap in maize plants. Nat Commun (2022) 13(1):3361. doi: 10.1038/s41467-022-31113-w
47. Devi R, Kaur T, Kour D, Yadav A, Yadav AN, Suman A, et al. Minerals solubilizing and mobilizing microbiomes: a sustainable approach for managing minerals’ deficiency in agricultural soil. J Appl Microbiol (2022) 133(3):1245–72. doi: 10.1111/jam.15627
48. Jiang Y, Liu M, Zhang J, Chen Y, Chen X, Chen L, et al. Nematode grazing promotes bacterial community dynamics in soil at the aggregate level. ISME J (2017) 11(12):2705–17. doi: 10.1038/ismej.2017.120
49. Garcia Mendez S, Persyn A, De Tender C, Goormachtig S, and Willems A. Unravelling the bacterial community composition of Valerianella locusta, a cold-tolerant plant. Phytobiomes J (2023) 7(4):500–14. doi: 10.1094/PBIOMES-12-22-0106-R
50. Marian M, Antonielli L, Pertot I, and Perazzolli M. Amplicon sequencing and culture-dependent approaches reveal core bacterial endophytes aiding freezing stress tolerance in alpine Rosaceae plants. mBio. (2025) 16(4):e0141824. doi: 10.1128/mbio.01418-24
51. Tiwari P, Bose SK, Park K-I, Dufossé L, and Fouillaud M. Plant-microbe interactions under the extreme habitats and their potential applications. Microorganisms (2024) 12(3):448. doi: 10.3390/microorganisms12030448
52. He WX, Sun QF, Hashem A, Abd_Allah EF, Wu QS, and Xu YJ. Sod culture with vicia villosa alters the diversity of fungal communities in walnut orchards for sustainability development. Sustainability (2023) 15(13):10731. doi: 10.3390/su151310731
53. Song C, Sarpong CK, Zhang X, Wang W, Wang L, Gan Y, et al. Mycorrhizosphere bacteria and plant-plant interactions facilitate maize P acquisition in an intercropping system. J Cleaner Prod (2021) 314:127993. doi: 10.1016/j.jclepro.2021.127993
54. Xiao L, Lai S, Chen M, Long X, Fu X, and Yang H. Effects of grass cultivation on soil arbuscular mycorrhizal fungi community in a tangerine orchard. Rhizosphere (2022) 24:100583. doi: 10.1016/j.rhisph.2022.100583
55. Hartman K, van der Heijden MGA, Wittwer RA, Banerjee S, Walser JC, and Schlaeppi K. Cropping practices manipulate abundance patterns of root and soil microbiome members paving the way to smart farming. Microbiome (2018) 6(1):14. doi: 10.1186/s40168-017-0389-9
56. Wilhelm RC, Amsili JP, Kurtz KSM, van Es HM, and Buckley DH. Ecological insights into soil health according to the genomic traits and environment-wide associations of bacteria in agricultural soils. ISME Commun (2023) 3(1):1. doi: 10.1038/s43705-022-00209-1
57. Oyserman BO, Cordovez V, Flores SS, Leite MFA, Nijveen H, Medema MH, et al. Extracting the GEMs: genotype, environment, and microbiome interactions shaping host phenotypes. Front Microbiol (2020) 11:574053. doi: 10.3389/fmicb.2020.574053
58. Benmrid B, Ghoulam C, Zeroual Y, Kouisni L, and Bargaz A. Bioinoculants as a means of increasing crop tolerance to drought and phosphorus deficiency in legume-cereal intercropping systems. Commun Biol (2023) 6(1):1016. doi: 10.1038/s42003-023-05399-5
59. Cao M, Narayanan M, Shi X, Chen X, Li Z, and Ma Y. Optimistic contributions of plant growth-promoting bacteria for sustainable agriculture and climate stress alleviation. Environ Res (2023) 217:114924. doi: 10.1016/j.envres.2022.114924
60. Hayat R, Ali S, Amara U, Khalid R, and Ahmed I. Soil beneficial bacteria and their role in plant growth promotion: a review. Ann Microbiol (2010) 60(4):579–98. doi: 10.1007/s13213-010-0117-1
61. Rai S, Omar AF, Rehan M, Al-Turki A, Sagar A, Ilyas N, et al. Crop microbiome: their role and advances in molecular and omic techniques for the sustenance of agriculture. Planta (2022) 257(2):27. doi: 10.1007/s00425-022-04052-5
62. Vejan P, Abdullah R, Khadiran T, Ismail S, and Nasrulhaq Boyce A. Role of plant growth promoting rhizobacteria in agricultural sustainability-a review. Molecules (2016) 21(5):573. doi: 10.3390/molecules21050573
63. Wahab A, Muhammad M, Munir A, Abdi G, Zaman W, Ayaz A, et al. Role of arbuscular mycorrhizal fungi in regulating growth, enhancing productivity, and potentially influencing ecosystems under abiotic and biotic stresses. Plants (Basel) (2023) 12(17):3102. doi: 10.3390/plants12173102
64. Chapelle E, Mendes R, Bakker PAHM, and Raaijmakers JM. Fungal invasion of the rhizosphere microbiome. ISME J (2016) 10(1):265–8. doi: 10.1038/ismej.2015.82
65. Li H, La S, Zhang X, Gao L, and Tian Y. Salt-induced recruitment of specific root-associated bacterial consortium capable of enhancing plant adaptability to salt stress. ISME J (2021) 15(10):2865–82. doi: 10.1038/s41396-021-00974-2
66. Si J, Froussart E, Viaene T, Vázquez-Castellanos JF, Hamonts K, Tang L, et al. Interactions between soil compositions and the wheat root microbiome under drought stress: from an in silico to in planta perspective. Comput Struct Biotechnol J (2021) 19:4235–47. doi: 10.1016/j.csbj.2021.07.027
67. Wei D, Zhu D, Zhang Y, Yang Z, Hu Y, Song C, et al. Pseudomonas chlororaphis IRHB3 assemblies beneficial microbes and activates JA-mediated resistance to promote nutrient utilization and inhibit pathogen attack. Front Microbiol (2024) 15:1328863. doi: 10.3389/fmicb.2024.1328863
68. Nunes PSO, Lacerda-Junior GV, Mascarin GM, Guimarães RA, Medeiros FHV, Arthurs S, et al. Microbial consortia of biological products: do they have a future? Biol Control (2024) 188:105439. doi: 10.1016/j.biocontrol.2024.105439
69. Chalot M and Puschenreiter M. Editorial: exploring plant rhizosphere, phyllosphere and endosphere microbial communities to improve the management of polluted sites. Front Microbiol (2021) 12:763566. doi: 10.3389/fmicb.2021.763566
70. Perreault R and Laforest-Lapointe I. Plant-microbe interactions in the phyllosphere: facing challenges of the Anthropocene. ISME J (2022) 16(2):339–45. doi: 10.1038/s41396-021-01109-3
71. Zhu YG, Xiong C, Wei Z, Chen QL, Ma B, Zhou SYD, et al. Impacts of global change on the phyllosphere microbiome. New Phytol (2022) 234(6):1977–86. doi: 10.1111/nph.17928
72. Debray R, Socolar Y, Kaulbach G, Guzman A, Hernandez CA, Curley R, et al. Water stress and disruption of mycorrhizas induce parallel shifts in phyllosphere microbiome composition. New Phytol (2022) 234(6):2018–31. doi: 10.1111/nph.17817
73. Brandl MT, Mammel MK, Simko I, Richter TKS, Gebru ST, and Leonard SR. Weather factors, soil microbiome, and bacteria-fungi interactions as drivers of the epiphytic phyllosphere communities of romaine lettuce. Food Microbiol (2023) 113:104260. doi: 10.1016/j.fm.2023.104260
74. Attwood GT, Wakelin SA, Leahy SC, Rowe S, Clarke S, Chapman DF, et al. Applications of the soil, plant and rumen microbiomes in pastoral agriculture. Front Nutr (2019) 6:107. doi: 10.3389/fnut.2019.00107
75. Wang Y, Dang N, Feng K, Wang J, Jin X, Yao S, et al. Grass-microbial inter-domain ecological networks associated with alpine grassland productivity. Front Microbiol (2023) 14:1109128. doi: 10.3389/fmicb.2023.1109128
76. Chen L, Li D, Shao Y, Adni J, Wang H, Liu Y, et al. Comparative analysis of soil microbiome profiles in the companion planting of White clover and orchard grass using 16S rRNA gene sequencing data. Front Plant Sci (2020) 11:538311. doi: 10.3389/fpls.2020.538311
77. He A, Niu S, Yang D, Ren W, Zhao L, Sun Y, et al. Two PGPR strains from the rhizosphere of haloxylon ammodendron promoted growth and enhanced drought tolerance of ryegrass. Plant Physiol Biochem (2021) 161:74–85. doi: 10.1016/j.plaphy.2021.02.003
78. Zhang C, Cai K, Li M, Zheng J, and Han Y. Plant-growth-promoting potential of PGPE isolated from Dactylis glomerata L. Microorganisms (2022) 10(4):731. doi: 10.3390/microorganisms10040731
79. Sayre JM, Wang D, Lin JY, Danielson RE, Scow KM, and Mazza Rodrigues JL. Repeated manure inputs to a forage production soil increase microbial biomass and diversity and select for lower abundance genera. Agric Ecosyst Environ (2023) 354:108567. doi: 10.1016/j.agee.2023.108567
80. Do TT, Smyth C, Crispie F, Burgess C, Brennan F, and Walsh F. Comparison of soil and grass microbiomes and resistomes reveals grass as a greater antimicrobial resistance reservoir than soil. Sci Total Environ (2023) 857(Pt 1):159179. doi: 10.1016/j.scitotenv.2022.159179
81. Tyrrell C, Do TT, Leigh RJ, Burgess CM, Brennan FP, and Walsh F. Differential impact of swine, bovine and poultry manure on the microbiome and resistome of agricultural grassland. Sci Total Environ (2023) 886:163926. doi: 10.1016/j.scitotenv.2023.163926
82. Food and Agriculture Organization of the United Nations. Livestock and environment statistics: manure and greenhouse gas emissions Global, regional and country trends 1990–2018 [brief 14]. In: FAOSTAT analytical briefs. Rome: FAO (2020). Available at: https://openknowledge.fao.org/handle/20.500.14283/cb1922en.
83. Ridwan R, Abdelbagi M, Sofyan A, Fidriyanto R, Astuti WD, Fitri A, et al. A meta-analysis to observe silage microbiome differentiated by the use of inoculant and type of raw material. Front Microbiol (2023) 14:1063333. doi: 10.3389/fmicb.2023.1063333
84. Ali N, Wang S, Zhao J, Dong Z, Li J, Nazar M, et al. Using a high-throughput sequencing technology to evaluate the various forage source epiphytic microbiota and their effect on fermentation quality and microbial diversity of Napier grass. Arch Microbiol (2021) 203(8):4879–90. doi: 10.1007/s00203-021-02296-5
85. Yang X, Bao Y, Shao T, Wang W, Ma P, Wang W, et al. Altitudinal distribution patterns of phyllosphere microbial communities and their contribution to silage fermentation of Kobresia pygmaea along the elevation gradient on the Tibetan Plateau. Front Microbiol (2022) 13:874582. doi: 10.3389/fmicb.2022.874582
86. Zhang X, Wang H, and Guo X. Effects of total mixed ration with various silage on growth performance, serum parameters, ruminal fermentation, and bacteria community profile in beef cattle. Food Sci Nutr (2021) 9(11):5959–70. doi: 10.1002/fsn3.2498
87. Vezzulli L and Ventura M. Editorial overview: biotechnological insights of marine and terrestrial microbiome. Curr Opin Biotechnol (2022) 73:vii–ix. doi: 10.1016/j.copbio.2021.12.006
88. Corinaldesi C, Bianchelli S, Candela M, Dell’Anno A, Gambi C, Rastelli E, et al. Microbiome-assisted restoration of degraded marine habitats: a new nature-based solution? Front Mar Sci (2023) 10:1227560. doi: 10.3389/fmars.2023.1227560
89. Alegbeleye OO and Sant’Ana AS. Manure-borne pathogens as an important source of water contamination: an update on the dynamics of pathogen survival/transport as well as practical risk mitigation strategies. Int J Hyg Environ Health (2020) 227:113524. doi: 10.1016/j.ijheh.2020.113524
90. Jurelevicius D, Cotta SR, Montezzi LF, Dias ACF, Mason OU, Picão RC, et al. Enrichment of potential pathogens in marine microbiomes with different degrees of anthropogenic activity. Environ pollut (2021) 268(A):115757. doi: 10.1016/j.envpol.2020.115757
91. Zhang M-S, Li W, Zhang W-G, Li Y-T, Li J-Y, and Gao Y. Agricultural land-use change exacerbates the dissemination of antibiotic resistance genes via surface runoffs in Lake Tai Basin, China. Ecotoxicol Environ Saf (2021) 220:112328. doi: 10.1016/j.ecoenv.2021.112328
92. Knorr S, Weisener CG, and Phillips LA. Agricultural land management alters the biogeochemical cycling capacity of aquatic and sediment environments. Agric Ecosyst Environ (2023) 357:108661. doi: 10.1016/j.agee.2023.108661
93. Ritchie H. The world now produces more seafood from fish farms than wild catch. OurWorldInData.org [online] (2019). Available at: https://ourworldindata.org/rise-of-aquaculturearticle-citation
94. Dimarchopoulou D, Wibisono E, Saul S, Carvalho P, Nugraha A, Mous PJ, et al. Combining catch-based indicators suggests overexploitation and poor status of Indonesia’s deep demersal fish stocks. Fish Res (2023) 268:106854. doi: 10.1016/j.fishres.2023.106854
95. Dimitriadis C, Marampouti C, Calò A, Di Franco A, Giakoumi S, Di Franco E, et al. Evaluating the long term effectiveness of a Mediterranean marine protected area to tackle the effects of invasive and range expanding herbivorous fish on rocky reefs. Mar Environ Res (2024) 193:106293. doi: 10.1016/j.marenvres.2023.106293
96. Byeon E, Jeong H, Lee Y-J, Cho Y, Lee K-W, Lee E, et al. Effects of microplastics and phenanthrene on gut microbiome and metabolome alterations in the marine medaka Oryzias melastigma. J Hazard Mater (2024) 461:132620. doi: 10.1016/j.jhazmat.2023.132620
97. Minich JJ, Petrus S, Michael JD, Michael TP, Knight R, Allen EE, et al. Temporal, environmental, and biological drivers of the mucosal microbiome in a wild marine fish, Scomber japonicus. mSphere (2020) 5(3):e00401–20. doi: 10.1128/mSphere.00401-20
98. Sababadichetty L, Miltgen G, Vincent B, Guilhaumon F, Lenoble V, Thibault M, et al. Microplastics in the insular marine environment of the southwest Indian Ocean carry a microbiome including antimicrobial resistant (AMR) bacteria: a case study from Reunion Island. Mar pollut Bull (2024) 198:115911. doi: 10.1016/j.marpolbul.2023.115911
99. Mishra P, Naik S, Babu PV, Pradhan U, Begum M, Kaviarasan T, et al. Algal bloom, hypoxia, and mass fish kill events in the backwaters of Puducherry, Southeast coast of India. Oceanologia (2022) 64(2):396–403. doi: 10.1016/j.oceano.2021.11.003
100. Da Le N, Hoang TT, Nguyen TM, Rochelle-Newall E, Pham TM, Phung TX, et al. Microbial contamination in the coastal aquaculture zone of the Ba Lat river mouth, Vietnam. Mar pollut Bull (2023) 192:115078. doi: 10.1016/j.marpolbul.2023.115078
101. Newton K, Gonzalez E, Pitre FE, and Brereton NJB. Microbial community origin and fate through a rural wastewater treatment plant. Environ Microbiol (2022) 24(5):2516–42. doi: 10.1111/1462-2920.16025
102. Premke K, Wurzbacher C, Felsmann K, Fabian J, Taube R, Bodmer P, et al. Large-scale sampling of the freshwater microbiome suggests pollution-driven ecosystem changes. Environ pollut (2022) 308:119627. doi: 10.1016/j.envpol.2022.119627
103. Li J, Li J, Zhang Y, and Lu H. The responses of marine anammox bacteria-based microbiome to multi-antibiotic stress in mariculture wastewater treatment. Water Res (2022) 224:119050. doi: 10.1016/j.watres.2022.119050
104. Sun H, Wang T, Liu S, Tang X, Sun J, Liu X, et al. Novel insights into the rhizosphere and seawater microbiome of Zostera marina in diverse mariculture zones. Microbiome (2024) 12(1):27. doi: 10.1186/s40168-024-01759-3
105. Krishnaswamy VG, Mani K, Senthil Kumar P, Rangasamy G, Sridharan R, Rethnaraj C, et al. Prevalence of differential microbiome in healthy, diseased and nipped colonies of corals, Porites lutea in the Gulf of Kachchh, north-west coast of India. Environ Res (2023) 216(2):114622. doi: 10.1016/j.envres.2022.114622
106. van Duyl FC, van Bleijswijk JDL, Wuchter C, Witte HJ, Coolen MJL, Bak RPM, et al. Recovery patterns of the coral microbiome after relief of algal contact. J Sea Res (2023) 191:102309. doi: 10.1016/j.seares.2022.102309
107. Sun F, Yang H, Shi Q, and Wang G. Changes in coral bacterial communities during a natural bleaching event linked to el Niño in the South China Sea. Reg Stud Mar Sci (2022) 53:102383. doi: 10.1016/j.rsma.2022.102383
108. Ribeiro B, Garritano A N, Raimundo I, Delgadillo-Ordoñez N, Nappi J, Garcias-Bonet N, et al. Not only for corals: exploring the uptake of beneficial microorganisms for corals by sponges. NPJ Biofilms Microbiomes (2024) 10(1):125. doi: 10.1038/s41522-024-00584-8
109. Lewis DM, Thompson KA, MacDonald TC, and Cook GS. Understanding shifts in estuarine fish communities following disturbances using an ensemble modeling framework. Ecol Indic (2021) 126:107623. doi: 10.1016/j.ecolind.2021.107623
110. Legrand TPRA, Catalano SR, Wos-Oxley ML, Stephens F, Landos M, Bansemer MS, et al. The inner workings of the outer surface: skin and gill microbiota as indicators of changing gut health in yellowtail kingfish. Front Microbiol (2017) 8:2664. doi: 10.3389/fmicb.2017.02664
111. Vezzulli L, Martinez-Urtaza J, and Stern R. Continuous plankton recorder in the omics era: from marine microbiome to global ocean observations. Curr Opin Biotechnol (2022) 73:61–6. doi: 10.1016/j.copbio.2021.07.016
112. McQuatters-Gollop A, Stern RF, Atkinson A, Best M, Bresnan E, Creach V, et al. The silent majority: pico- and nanoplankton as ecosystem health indicators for marine policy. Ecol Indic (2024) 159:111650. doi: 10.1016/j.ecolind.2024.111650
113. Melo J and Quintas C. Minimally processed fruits as vehicles for foodborne pathogens. AIMS Microbiol (2023) 9(1):1–19. doi: 10.3934/microbiol.2023001
114. Santos MI, Grácio M, Silva MC, Pedroso L, and Lima A. One health perspectives on food safety in minimally processed vegetables and fruits: from farm to fork. Microorganisms (2023) 11(12):2990. doi: 10.3390/microorganisms11122990
115. Soto-Giron MJ, Kim JN, Schott E, Tahmin C, Ishoey T, Mincer TJ, et al. The edible plant microbiome represents a diverse genetic reservoir with functional potential in the human host. Sci Rep (2021) 11(1):24017. doi: 10.1038/s41598-021-03334-4
116. Wicaksono WA, Cernava T, Wassermann B, Abdelfattah A, Soto-Giron MJ, Toledo GV, et al. The edible plant microbiome: evidence for the occurrence of fruit and vegetable bacteria in the human gut. Gut Microbes (2023) 15(2):2258565. doi: 10.1080/19490976.2023.2258565
117. CBI Netherlands Ministry of Foreign Affairs. What is the demand for fresh fruit and vegetables on the European market? [online] (2024). Available at: https://www.cbi.eu/market-information/fresh-fruit-vegetables/what-demand
118. López L-A, Tobarra M-A, Cadarso M-Á, Gómez N, and Cazcarro I. Eating local and in-season fruits and vegetables: carbon-water-employment trade-offs and synergies. Ecol Econ (2022) 192:107270. doi: 10.1016/j.ecolecon.2021.107270
119. Vargas AM, de Moura AP, Deliza R, and Cunha LM. The role of local seasonal foods in enhancing sustainable food consumption: a systematic literature review. Foods (2021) 10(9):2206. doi: 10.3390/foods10092206
120. Bancal V and Ray RC. Overview of food loss and waste in fruits and vegetables: from issue to resources. In: Ray RC, editor. Fruits and vegetable wastes. Springer Nature, Singapore (2022) 3–29. doi: 10.1007/978-981-16-9527-8_1
121. Alegbeleye O, Odeyemi OA, Strateva M, and Stratev D. Microbial spoilage of vegetables, fruits and cereals. Appl Food Res (2022) 2(1):100122. doi: 10.1016/j.afres.2022.100122
122. Agirman B, Carsanba E, Settanni L, and Erten H. Exploring yeast-based microbial interactions: the next frontier in postharvest biocontrol. Yeast (2023) 40(10):457–75. doi: 10.1002/yea.3895
123. Hosseini A, Koushesh Saba M, and Watkins CB. Microbial antagonists to biologically control postharvest decay and preserve fruit quality. Crit Rev Food Sci Nutr (2023) 64(21):7330–42. doi: 10.1080/10408398.2023.2184323
124. Sharma RR, Singh D, and Singh R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: a review. Biol Control (2009) 50(3):205–21. doi: 10.1016/j.biocontrol.2009.05.001
125. Al Riachy R, Strub C, Durand N, Chochois V, Lopez-Lauri F, Fontana A, et al. The influence of long-term storage on the epiphytic microbiome of postharvest apples and on Penicillium expansum occurrence and patulin accumulation. Toxins (2024) 16(2):102. doi: 10.3390/toxins16020102
126. Kusstatscher P, Cernava T, Abdelfattah A, Gokul J, Korsten L, and Berg G. Microbiome approaches provide the key to biologically control postharvest pathogens and storability of fruits and vegetables. FEMS Microbiol Ecol (2020) 96(7):fiaa119. doi: 10.1093/femsec/fiaa119
127. Zheng R, Wang D, Li X, Yang M, Kong Q, and Ren X. Screening of core microorganisms in healthy and diseased peaches and effect evaluation of biocontrol bacteria (Burkholderia sp.). Food Microbiol (2024) 120:104465. doi: 10.1016/j.fm.2024.104465
128. Gu G, Ding Q, Redding M, Yang Y, O’Brien R, Gu T, et al. Differential microbiota shift on whole romaine lettuce subjected to source or forward processing and on fresh-cut products during cold storage. Int J Food Microbiol (2024) 416:110665. doi: 10.1016/j.ijfoodmicro.2024.110665
129. Valentino V, Sequino G, Cobo-Díaz JF, Álvarez-Ordóñez A, De Filippis F, and Ercolini D. Evidence of virulence and antibiotic resistance genes from the microbiome mapping in minimally processed vegetables producing facilities. Food Res Int (2022) 162(B):112202. doi: 10.1016/j.foodres.2022.112202
130. Einson JE, Rani A, You X, Rodriguez AA, Randell CL, Barnaba T, et al. A vegetable fermentation facility hosts distinct microbiomes reflecting the production environment. Appl Environ Microbiol (2018) 84(22):e01680–18. doi: 10.1128/AEM.01680-18
131. Food and Agriculture Organization of the United Nations. The future of food and agriculture – alternative pathways to 2050. Rome: FAO (2018).
132. Huaiquipán R, Quiñones J, Díaz R, Velásquez C, Sepúlveda G, Velázquez L, et al. Review: effect of experimental diets on the microbiome of productive animals. Microorganisms (2023) 11(9):2219. doi: 10.3390/microorganisms11092219
133. Du S, Bu Z, You S, Jiang Z, Su W, Wang T, et al. Integrated rumen microbiome and serum metabolome analysis responses to feed type that contribution to meat quality in lambs. Anim Microbiome (2023) 5(1):65. doi: 10.1186/s42523-023-00288-y
134. Fan Q, Xie K, Cui X, Zhang G, Zheng H, Chang S, et al. Microecosystem of yak rumen on the Qinghai-Tibetan Plateau is stable and is unaffected by soil or grass microbiota. Environ Microbiol (2022) 24(12):5760–73. doi: 10.1111/1462-2920.16236
135. Hess MK, Hodgkinson HE, Hess AS, Zetouni L, Budel JCC, Henry H, et al. Large-scale analysis of sheep rumen metagenome profiles captured by reduced representation sequencing reveals individual profiles are influenced by the environment and genetics of the host. BMC Genomics (2023) 24(1):551. doi: 10.1186/s12864-023-09660-3
136. Liu J, Bai Y, Liu F, Kohn RA, Tadesse DA, Sarria S, et al. Rumen microbial predictors for short-chain fatty acid levels and the grass-fed regimen in Angus Cattle. Anim (Basel) (2022) 12(21):2995. doi: 10.3390/ani12212995
137. Matthews C, Crispie F, Lewis E, Reid M, O’Toole PW, and Cotter PD. The rumen microbiome: a crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes (2019) 10(2):115–32. doi: 10.1080/19490976.2018.1505176
138. Wang D, Chen L, Tang G, Yu J, Chen J, Li Z, et al. Multi-omics revealed the long-term effect of ruminal keystone bacteria and the microbial metabolome on lactation performance in adult dairy goats. Microbiome (2023) 11(1):215. doi: 10.1186/s40168-023-01652-5
139. Omontese BO, Sharma AK, Davison S, Jacobson E, DiConstanzo A, Webb MJ, et al. Microbiome network traits in the rumen predict average daily gain in beef cattle under different backgrounding systems. Anim Microbiome (2022) 4(1):25. doi: 10.1186/s42523-022-00175-y
140. Liu T, Bu Z, Xiang K, Jia Y, and Du S. Effects of non-pelleted or pelleted low-native grass and pelleted high-native grass diets on meat quality by regulating the rumen microbiota in lambs. Microbiol Spectr (2024) 12(4):e0375823. doi: 10.1128/spectrum.03758-23
141. Chang M, Wang F, Ma F, Jin Y, and Sun P. Supplementation with galacto-oligosaccharides in early life persistently facilitates the microbial colonization of the rumen and promotes growth of preweaning Holstein dairy calves. Anim Nutr (2022) 10:223–33. doi: 10.1016/j.aninu.2022.04.009
142. Linde DA, Schokker D, du Toit CJL, Ramkilawon GD, and van Marle-Köster E. The effect of a Bacillus probiotic and essential oils compared to an ionophore on the rumen microbiome composition of feedlot cattle. Animals (Basel) (2023) 13(18):2927. doi: 10.3390/ani13182927
143. Michalak M, Wojnarowski K, Cholewińska P, Szeligowska N, Bawej M, and Pacoń J. Selected alternative feed additives used to manipulate the rumen microbiome. Animals (Basel) (2021) 11(6):1542. doi: 10.3390/ani11061542
144. Rabee AE, Younan BR, Kewan KZ, Sabra EA, and Lamara M. Modulation of rumen bacterial community and feed utilization in camel and sheep using combined supplementation of live yeast and microalgae. Sci Rep (2022) 12(1):12990. doi: 10.1038/s41598-022-16988-5
145. van Gastelen S, Burgers EEA, Dijkstra J, de Mol R, Muizelaar W, Walker N, et al. Long-term effects of 3-nitrooxypropanol on methane emission and milk production characteristics in Holstein-Friesian dairy cows. J Dairy Sci (2024) 107(8):5556–73. doi: 10.3168/jds.2023-24198
146. Barrett K, Lange L, Børsting CF, Olijhoek DW, Lund P, and Meyer AS. Changes in the metagenome-encoded CAZymes of the rumen microbiome are linked to feed-induced reductions in methane emission from Holstein cows. Front Microbiol (2022) 13:855590. doi: 10.3389/fmicb.2022.855590
147. Miller GA, Auffret MD, Roehe R, Nisbet H, and Martínez-Álvaro M. Different microbial genera drive methane emissions in beef cattle fed with two extreme diets. Front Microbiol (2023) 14:1102400. doi: 10.3389/fmicb.2023.1102400
148. Tseten T, Sanjorjo RA, Kwon M, and Kim SW. Strategies to mitigate enteric methane emissions from ruminant animals. J Microbiol Biotechnol (2022) 32(3):269–77. doi: 10.4014/jmb.2202.02019
149. Opio C, Gerber P, Mottet A, Falcucci A, Tempio G, MacLeod M, et al. Greenhouse gas emissions from ruminant supply chains – a global life cycle assessment. Rome: Food and Agriculture Organization of the United Nations (2013). Available at: https://www.fao.org/4/i3461e/i3461e00.htm.
150. Mhuireach GÁ, Dietz L, and Gillett T. One or many? Multi-species livestock grazing influences soil microbiome community structure and antibiotic resistance potential. Front Sustain Food Syst (2022) 6:926824. doi: 10.3389/fsufs.2022.926824
151. Wu Y, Chen D, Delgado-Baquerizo M, Liu S, Wang B, Wu J, et al. Long-term regional evidence of the effects of livestock grazing on soil microbial community structure and functions in surface and deep soil layers. Soil Biol Biochem (2022) 168:108629. doi: 10.1016/j.soilbio.2022.108629
152. Xun W, Yan R, Ren Y, Jin D, Xiong W, Zhang G, et al. Grazing-induced microbiome alterations drive soil organic carbon turnover and productivity in meadow steppe. Microbiome (2018) 6(1):170. doi: 10.1186/s40168-018-0544-y
153. Ferrocino I, Rantsiou K, and Cocolin L. Microbiome and -omics application in food industry. Int J Food Microbiol (2022) 377:109781. doi: 10.1016/j.ijfoodmicro.2022.109781
154. Imai K, Niwa R, Fujioka M, and Ito K. Understanding the quality and safety of food production through the lens of the microbiome of the built environment. Biosci Biotechnol Biochem (2024) 88(3):254–9. doi: 10.1093/bbb/zbad164
155. Sequino G, Valentino V, Villani F, and De Filippis F. Omics-based monitoring of microbial dynamics across the food chain for the improvement of food safety and quality. Food Res Int (2022) 157:111242. doi: 10.1016/j.foodres.2022.111242
156. Xu ZS, Ju T, Yang X, and Gänzle M. A meta-analysis of bacterial communities in food processing facilities: driving forces for assembly of core and accessory microbiomes across different food commodities. Microorganisms (2023) 11(6):1575. doi: 10.3390/microorganisms11061575
157. Frétin M, Martin B, Rifa E, Isabelle VM, Pomiès D, Ferlay A, et al. Bacterial community assembly from cow teat skin to ripened cheeses is influenced by grazing systems. Sci Rep (2018) 8(1):200. doi: 10.1038/s41598-017-18447-y
158. Verdier-Metz I, Delbès C, Bouchon M, Rifa E, Theil S, Chaucheyras-Durand F, et al. Dietary live yeast supplementation influence on Cow’s milk, teat and bedding microbiota in a grass-diet dairy system. Microorganisms (2023) 11(3):673. doi: 10.3390/microorganisms11030673
159. Ózsvári L and Ivanyos D. The use of teat disinfectants and milking machine cleaning products in commercial Holstein-Friesian farms. Front Vet Sci (2022) 9:956843. doi: 10.3389/fvets.2022.956843
160. Vargova M, Vyrostkova J, Lakticova KV, and Zigo F. Effectiveness of sanitation regime in a milking parlour to control microbial contamination of teats and surfaces teat cups’. Ann Agric Environ Med (2023) 30(1):55–60. doi: 10.26444/aaem/161037
161. Derakhshani H, Fehr KB, Sepehri S, Francoz D, De Buck J, Barkema HW, et al. Invited review: microbiota of the bovine udder: contributing factors and potential implications for udder health and mastitis susceptibility. J Dairy Sci (2018) 101(12):10605–25. doi: 10.3168/jds.2018-14860
162. Verdier-Metz I, Delbès C, Bouchon M, Pradel P, Theil S, Rifa E, et al. Influence of post-milking treatment on microbial diversity on the cow teat skin and in milk. Dairy (2022) 3(2):262–76. doi: 10.3390/dairy3020021
163. Mather AE, Gilmour MW, Reid SWJ, and French NP. Foodborne bacterial pathogens: genome-based approaches for enduring and emerging threats in a complex and changing world. Nat Rev Microbiol (2024) 22(9):543–55. doi: 10.1038/s41579-024-01051-z
164. Johnson J, Curtin C, and Waite-Cusic J. The cheese production facility microbiome exhibits temporal and spatial variability. Front Microbiol (2021) 12:644828. doi: 10.3389/fmicb.2021.644828
165. Lacorte GA, Cruvinel LA, de Paula Ávila M, Dias MF, de Abreu Pereira A, Nascimento AMA, et al. Investigating the influence of Food Safety Management Systems (FSMS) on microbial diversity of Canastra cheeses and their processing environments. Food Microbiol (2022) 105:104023. doi: 10.1016/j.fm.2022.104023
166. Sun L and D’Amico DJ. Composition, succession, and source tracking of microbial communities throughout the traditional production of a farmstead cheese. mSystems (2021) 6(5):e00830–21. doi: 10.1128/msystems.00830-21
167. Xu X, Rothrock MJ Jr, Mishra A, Kumar GD, and Mishra A. Relationship of the poultry microbiome to pathogen colonization, farm management, poultry production, and foodborne illness risk assessment. J Food Prot (2023) 86(11):100169. doi: 10.1016/j.jfp.2023.100169
168. Yang X, Narvaez-Bravo C, and Zhang P. Driving forces shaping the microbial ecology in meat packing plants. Front Microbiol (2023) 14:1333696. doi: 10.3389/fmicb.2023.1333696
169. Xu ZS, Pham VD, Yang X, and Gänzle MG. High-throughput analysis of microbiomes in a meat processing facility: are food processing facilities an establishment niche for persisting bacterial communities? Microbiome (2025) 13(1):25. doi: 10.1186/s40168-024-02026-1
170. Zwirzitz B, Wetzels SU, Dixon ED, Stessl B, Zaiser A, Rabanser I, et al. The sources and transmission routes of microbial populations throughout a meat processing facility. NPJ Biofilms Microbiomes (2020) 6(1):26. doi: 10.1038/s41522-020-0136-z
171. Fagerlund A, Langsrud S, and Møretrø T. In-depth longitudinal study of Listeria monocytogenes ST9 isolates from the meat processing industry: resolving diversity and transmission patterns using whole-genome sequencing. Appl Environ Microbiol (2020) 86(14):e00579–20. doi: 10.1128/AEM.00579-20
172. Belk AD, Frazier AN, Fuerniss LK, Delmore R, Belk K, Borlee B, et al. A pilot study: the development of a facility-associated microbiome and its association with the presence of Listeria spp. in one small meat processing facility. Microbiol Spectr (2022) 10(5):e0204522. doi: 10.1128/spectrum.02045-22
173. Cobo-Díaz JF, Alvarez-Molina A, Alexa EA, Walsh CJ, Mencía-Ares O, Puente-Gómez P, et al. Microbial colonization and resistome dynamics in food processing environments of a newly opened pork cutting industry during 1.5 years of activity. Microbiome (2021) 9(1):204. doi: 10.1186/s40168-021-01131-9
174. Struelens MJ, Ludden C, Werner G, Sintchenko V, Jokelainen P, and Ip M. Real-time genomic surveillance for enhanced control of infectious diseases and antimicrobial resistance. Front Sci (2024) 2:1298248. doi: 10.3389/fsci.2024.1298248
175. Chen C-Z, Li P, Liu L, and Li Z-H. Exploring the interactions between the gut microbiome and the shifting surrounding aquatic environment in fisheries and aquaculture: a review. Environ Res (2022) 214(4):114202. doi: 10.1016/j.envres.2022.114202
176. Najafpour B, Pinto PIS, Sanz EC, Martinez-Blanch JF, Canario AVM, Moutou KA, et al. Core microbiome profiles and their modification by environmental, biological, and rearing factors in aquaculture hatcheries. Mar pollut Bull (2023) 193:115218. doi: 10.1016/j.marpolbul.2023.115218
177. Neuman C, Hatje E, Zarkasi KZ, Smullen R, Bowman JP, and Katouli M. The effect of diet and environmental temperature on the faecal microbiota of farmed Tasmanian Atlantic Salmon (Salmo salar L.). Aquacult Res (2016) 47(2):660–72. doi: 10.1111/are.12522
178. Viver T, Ruiz A, Bertomeu E, Martorell-Barceló M, Urdiain M, Grau A, et al. Food determines ephemerous and non-stable gut microbiome communities in juvenile wild and farmed Mediterranean fish. Sci Total Environ (2023) 889:164080. doi: 10.1016/j.scitotenv.2023.164080
179. Bashorun A, Hassan ZU, Al-Yafei MA-A, and Jaoua S. Fungal contamination and mycotoxins in aquafeed and tissues of aquaculture fishes and their biological control. Aquaculture (2023) 576:739892. doi: 10.1016/j.aquaculture.2023.739892
180. James G, Das BC, Jose S, and VJ RK. Bacillus as an aquaculture friendly microbe. Aquacult Int (2021) 29(1):323–53. doi: 10.1007/s10499-020-00630-0
181. Ren W, Wu H, Guo C, Xue B, Long H, Zhang X, et al. Multi-strain tropical Bacillus spp. as a potential probiotic biocontrol agent for large-scale enhancement of mariculture water quality. Front Microbiol (2021) 12:699378. doi: 10.3389/fmicb.2021.699378
182. Huang J, Amenyogbe E, Ou G, Li Y, Wen Z, Jiang X, et al. Effects of Bacillus sp. and Lactobacillus sp. combination as a water additive on the culture pond water and growth performance of hybrid grouper (Epinephelus fuscoguttatus × Epinephelus polyphekadion). Front Mar Sci (2022) 9:1068997. doi: 10.3389/fmars.2022.1068997
183. Sahandi J, Jafaryan H, Soltani M, and Ebrahimi P. The use of two Bifidobacterium strains enhanced growth performance and nutrient utilization of rainbow trout (Oncorhynchus mykiss) fry. Probiot Antimicrob Proteins (2019) 11(3):966–72. doi: 10.1007/s12602-018-9455-2
184. Hoseinifar SH, Sun Y-Z, Wang A, and Zhou Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front Microbiol (2018) 9:2429. doi: 10.3389/fmicb.2018.02429
185. Jiang S, Shi B, Zhu D, Cheng X, Zhou Z, Xie J, et al. Cross-contamination and ecological risk assessment of antibiotics between rivers and surrounding open aquaculture ponds. Environ pollut (2024) 344:123404. doi: 10.1016/j.envpol.2024.123404
186. Šamanić I, Kalinić H, Fredotović Ž, Dželalija M, Bungur A-M, and Maravić A. Bacteria tolerant to colistin in coastal marine environment: detection, microbiome diversity and antibiotic resistance genes’ repertoire. Chemosphere (2021) 281:130945. doi: 10.1016/j.chemosphere.2021.130945
187. Zhang Z, Yang Q, Xu W, Tang R, Li L, and Li D. Short-term feeding of dietary florfenicol influences gut microbiome and growth performance of fast-growing Silurus meridionalis. Aquaculture (2023) 574:739645. doi: 10.1016/j.aquaculture.2023.739645
188. Food and Agriculture Organization of the United Nations. The state of world fisheries and aquaculture 2022. Towards blue transformation. Rome: FAO (2022). doi: 10.4060/cc0461en
189. Nisa M, Dar RA, Fomda BA, and Nazir R. Combating food spoilage and pathogenic microbes via bacteriocins: a natural and eco-friendly substitute to antibiotics. Food Control (2023) 149:109710. doi: 10.1016/j.foodcont.2023.109710
190. Harada LK, Silva EC, Campos WF, Del Fiol FS, Vila M, Dąbrowska K, et al. Biotechnological applications of bacteriophages: state of the art. Microbiol Res (2018) 212–213:38–58. doi: 10.1016/j.micres.2018.04.007
191. Diwan AD, Harke SN, and Panche AN. Studies on exploring the potentials of gut microbiomes to mitigate the bacterial and viral diseases of fish and shellfish in aquaculture farming. Microbe (2024) 2:100031. doi: 10.1016/j.microb.2023.100031
192. Al-Ashhab A, Alexander-Shani R, Avrahami Y, Ehrlich R, Strem RI, Meshner S, et al. Sparus aurata and Lates calcarifer skin microbiota under healthy and diseased conditions in UV and non-UV treated water. Anim Microbiome (2022) 4(1):42. doi: 10.1186/s42523-022-00191-y
193. Yñiguez AT and Ottong ZJ. Predicting fish kills and toxic blooms in an intensive mariculture site in the Philippines using a machine learning model. Sci Total Environ (2020) 707:136173. doi: 10.1016/j.scitotenv.2019.136173
194. Quezada-Rodriguez PR, Downes J, Egan F, Taylor RS, White S, Brenan A, et al. Assessment of gill microbiome of two strains of Atlantic salmon reared in flowthrough and recirculation hatcheries and following seawater transfer. Aquaculture (2024) 580:740322. doi: 10.1016/j.aquaculture.2023.740322
195. Neissi A, Rafiee G, Rahimi S, Farahmand H, Pandit S, and Mijakovic I. Enriched microbial communities for ammonium and nitrite removal from recirculating aquaculture systems. Chemosphere (2022) 295:133811. doi: 10.1016/j.chemosphere.2022.133811
196. Tang Y, Zhang H, Yan J, Luo N, Fu X, Wu X, et al. Assessing the efficacy of bleaching powder in disinfecting marine water: insights from the rapid recovery of microbiomes. Water Res (2023) 241:120136. doi: 10.1016/j.watres.2023.120136
197. Nissar S, Bakhtiyar Y, Arafat MY, Andrabi S, Mir ZA, Khan NA, et al. The evolution of integrated multi-trophic aquaculture in context of its design and components paving way to valorization via optimization and diversification. Aquaculture (2023) 565:739074. doi: 10.1016/j.aquaculture.2022.739074
198. Kummu M, de Moel H, Porkka M, Siebert S, Varis O, and Ward PJ. Lost food, wasted resources: global food supply chain losses and their impacts on freshwater, cropland, and fertiliser use. Sci Total Environ (2012) 438:477–89. doi: 10.1016/j.scitotenv.2012.08.092
199. United Nations. Global issues: food [online] (2024). Available at: https://www.un.org/en/global-issues/food
200. Ishangulyyev R, Kim S, and Lee SH. Understanding food loss and waste-why are we losing and wasting food? Foods (2019) 8(8):297. doi: 10.3390/foods8080297
201. Mobed A, Baradaran B, de la Guardia M, Agazadeh M, Hasanzadeh M, Rezaee MA, et al. Advances in detection of fastidious bacteria: from microscopic observation to molecular biosensors. TrAC Trends Analyt Chem (2019) 113:157–71. doi: 10.1016/j.trac.2019.02.012
202. Jeon YS, Chun J, and Kim BS. Identification of household bacterial community and analysis of species shared with human microbiome. Curr Microbiol (2013) 67(5):557–63. doi: 10.1007/s00284-013-0401-y
203. Tatsika S, Karamanoli K, Karayanni H, and Genitsaris S. Metagenomic characterization of bacterial communities on ready-to-eat vegetables and effects of household washing on their diversity and composition. Pathogens (2019) 8(1):37. doi: 10.3390/pathogens8010037
204. Mantegazza G, Gargari G, Duncan R, Consalez F, Taverniti V, Riso P, et al. Ready-to-eat rocket salads as potential reservoir of bacteria for the human microbiome. Microbiol Spec (2023) 11(1):e02970–22. doi: 10.1128/spectrum.02970-22
205. Mantegazza G, Duncan R, Telesca N, Gargari G, Perotti S, Riso P, et al. Lactic acid bacteria naturally associated with ready-to-eat rocket salad can survive the human gastrointestinal transit. Food Microbiol (2024) 118:104418. doi: 10.1016/j.fm.2023.104418
206. Tabla R, Gómez A, Rebollo JE, Molina F, and Roa I. Effectiveness of a bacteriophage cocktail in reducing cheese early blowing caused by Escherichia coli. LWT (2022) 153:112430. doi: 10.1016/j.lwt.2021.112430
207. Lavelle K, Murphy J, Fitzgerald B, Lugli GA, Zomer A, Neve H, et al. A decade of Streptococcus thermophilus phage evolution in an Irish dairy plant. Appl Environ Microbiol (2018) 84(10):e02855–17. doi: 10.1128/AEM.02855-17
208. Paillet T, Lossouarn J, Figueroa C, Midoux C, Rué O, Petit M-A, et al. Virulent phages isolated from a smear-ripened cheese are also detected in reservoirs of the cheese factory. Viruses (2022) 14(8):1620. doi: 10.3390/v14081620
209. Queiroz LL, Lacorte GA, Isidorio WR, Landgraf M, de Melo Franco BD, Pinto UM, et al. High level of interaction between phages and bacteria in an artisanal raw milk cheese microbial community. mSystems (2023) 8(1):e00564–22. doi: 10.1128/msystems.00564-22
210. Ledormand P, Desmasures N, Midoux C, Rué O, and Dalmasso M. Investigation of the phageome and prophages in French cider, a fermented beverage. Microorganisms (2022) 10(6):1203. doi: 10.3390/microorganisms10061203
211. McDonnell B, Parlindungan E, Vasiliauskaite E, Bottacini F, Coughlan K, Krishnaswami LP, et al. Viromic and metagenomic analyses of commercial spirulina fermentations reveal remarkable microbial diversity. Viruses (2024) 16(7):1039. doi: 10.3390/v16071039
212. Yu Z, Ma Y, Guan Y, Zhu Y, Wang K, Wang Y, et al. Metagenomics of virus diversities in solid-state brewing process of traditional Chinese vinegar. Foods (2022) 11(20):3296. doi: 10.3390/foods11203296
213. Costa MJ, Pastrana LM, Teixeira JA, Sillankorva SM, and Cerqueira MA. Bacteriophage delivery systems for food applications: opportunities and perspectives. Viruses (2023) 15(6):1271. doi: 10.3390/v15061271
214. Mäntynen S, Laanto E, Oksanen HM, Poranen MM, and Díaz-Muñoz SL. Black box of phage-bacterium interactions: exploring alternative phage infection strategies. Open Biol (2021) 11(9):210188. doi: 10.1098/rsob.210188
215. Ritz NL, Draper LA, Bastiaanssen TFS, Turkington CJR, Peterson VL, van de Wouw M, et al. The gut virome is associated with stress-induced changes in behaviour and immune responses in mice. Nat Microbiol (2024) 9(2):359–76. doi: 10.1038/s41564-023-01564-y
216. Strathdee SA, Hatfull GF, Mutalik VK, and Schooley RT. Phage therapy: from biological mechanisms to future directions. Cell (2023) 186(1):17–31. doi: 10.1016/j.cell.2022.11.017
217. Maffei E, Shaidullina A, Burkolter M, Heyer Y, Estermann F, Druelle V, et al. Systematic exploration of Escherichia coli phage–host interactions with the BASEL phage collection. PloS Biol (2021) 19(11):e3001424. doi: 10.1371/journal.pbio.3001424
218. Denes T, den Bakker HC, Tokman JI, Guldimann C, and Wiedmann M. Selection and characterization of phage-resistant mutant strains of listeria monocytogenes reveal host genes linked to phage adsorption. Appl Environ Microbiol (2015) 81(13):4295–305. doi: 10.1128/AEM.00087-15
219. Sumrall ET, Shen Y, Keller AP, Rismondo J, Pavlou M, Eugster MR, et al. Phage resistance at the cost of virulence: Listeria monocytogenes serovar 4b requires galactosylated teichoic acids for InlB-mediated invasion. PloS Pathog (2019) 15(10):e1008032. doi: 10.1371/journal.ppat.1008032
220. Goulet A, Mahony J, Cambillau C, and van Sinderen D. Exploring structural diversity among adhesion devices encoded by lactococcal P335 phages with AlphaFold2. Microorganisms (2022) 10(11):2278. doi: 10.3390/microorganisms10112278
221. Lavelle K, McDonnell B, Fitzgerald G, van Sinderen D, and Mahony J. Bacteriophage-host interactions in Streptococcus thermophilus and their impact on co-evolutionary processes. FEMS Microbiol Rev (2023) 47(4):fuad032. doi: 10.1093/femsre/fuad032
222. Agus A, Denizot J, Thévenot J, Massier S, Sauvanet P, Bonnet R, et al. Western diet induces a shift in microbiota composition enhancing susceptibility to adherent-invasive E. coli infection and intestinal inflammation. Gastroenterology (2016) 150(4):S23. doi: 10.1016/S0016-5085(16)30201-3
223. Akritidou T, Akkermans S, Smet C, Gaspari S, Sharma C, Matthews E, et al. Gut microbiota of the small intestine as an antimicrobial barrier against foodborne pathogens: impact of diet on the survival of S. Typhimurium and L. monocytogenes during in vitro digestion. Food Res Int (2023) 173(2):113292. doi: 10.1016/j.foodres.2023.113292
224. Kendig MD, Leigh S-J, and Morris MJ. Unravelling the impacts of western-style diets on brain, gut microbiota and cognition. Neurosci Biobehav Rev (2021) 128:233–43. doi: 10.1016/j.neubiorev.2021.05.031
225. Newsome R, Yang Y, and Jobin C. Western diet influences on microbiome and carcinogenesis. Semin Immunol (2023) 67:101756. doi: 10.1016/j.smim.2023.101756
226. Song Z, Song R, Liu Y, Wu Z, and Zhang X. Effects of ultra-processed foods on the microbiota-gut-brain axis: the bread-and-butter issue. Food Res Int (2023) 167:112730. doi: 10.1016/j.foodres.2023.112730
227. Carlino N, Blanco-Míguez A, Punčochář M, Mengoni C, Pinto F, Tatti A, et al. Unexplored microbial diversity from 2,500 food metagenomes and links with the human microbiome. Cell (2024) 187(20):5775–95. doi: 10.1016/j.cell.2024.07.039
228. Fernandes AE, Rosa PWL, Melo ME, Martins RCR, Santin FGO, Moura AMSH, et al. Differences in the gut microbiota of women according to ultra-processed food consumption. Nutr Metab Cardiovasc Dis (2023) 33(1):84–9. doi: 10.1016/j.numecd.2022.09.025
229. Focker M, van Asselt ED, Berendsen BJA, van de Schans MGM, van Leeuwen SPJ, Visser SM, et al. Review of food safety hazards in circular food systems in Europe. Food Res Int (2022) 158:111505. doi: 10.1016/j.foodres.2022.111505
230. Scarpa F and Casu M. Genomics and bioinformatics in one health: transdisciplinary approaches for health promotion and disease prevention. Int J Environ Res Public Health (2024) 21(10):1337. doi: 10.3390/ijerph21101337
231. Bedu-Ferrari C, Biscarrat P, Langella P, and Cherbuy C. Prebiotics and the human gut microbiota: from breakdown mechanisms to the impact on metabolic health. Nutrients (2022) 14(10):2096. doi: 10.3390/nu14102096
232. Sanders ME, Merenstein DJ, Reid G, Gibson GR, and Rastall RA. Probiotics and prebiotics in intestinal health and disease: from biology to the clinic. Nat Rev Gastroenterol Hepatol (2019) 16(10):605–16. doi: 10.1038/s41575-019-0173-3
233. Wang S, Xiao Y, Tian F, Zhao J, Zhang H, Zhai Q, et al. Rational use of prebiotics for gut microbiota alterations: specific bacterial phylotypes and related mechanisms. J Funct Foods (2020) 66:103838. doi: 10.1016/j.jff.2020.103838
234. Ye S, Shah BR, Li J, Liang H, Zhan F, Geng F, et al. A critical review on interplay between dietary fibers and gut microbiota. Trends Food Sci Technol (2022) 124:237–49. doi: 10.1016/j.tifs.2022.04.010
235. Shen X, Tilves C, Kim H, Tanaka T, Spira AP, Chia CW, et al. Plant-based diets and the gut microbiome: findings from the Baltimore longitudinal study of aging. Am J Clin Nutr (2024) 119(3):628–38. doi: 10.1016/j.ajcnut.2024.01.006
236. Boven L, Akkerman R, and de Vos P. Sustainable diets with plant-based proteins require considerations for prevention of proteolytic fermentation. Crit Rev Food Sci Nutr (2024) 65(14):2829–39. doi: 10.1080/10408398.2024.2352523
237. Neufingerl N and Eilander A. Nutrient intake and status in adults consuming plant-based diets compared to meat-eaters: a systematic review. Nutrients (2021) 14(1):29. doi: 10.3390/nu14010029
238. Ghallab DS, Ibrahim RS, Mohyeldin MM, and Shawky E. Marine algae: a treasure trove of bioactive anti-inflammatory compounds. Mar pollut Bull (2024) 199:116023. doi: 10.1016/j.marpolbul.2023.116023
239. Selvam J, Mal J, Singh S, Yadav A, Giri BS, Pandey A, et al. Bioprospecting marine microalgae as sustainable bio-factories for value-added compounds. Algal Res (2024) 79:103444. doi: 10.1016/j.algal.2024.103444
240. Sorrenti V, Burò I, Consoli V, and Vanella L. Recent advances in health benefits of bioactive compounds from food wastes and by-products: biochemical aspects. Int J Mol Sci (2023) 24(3):2019. doi: 10.3390/ijms24032019
241. Mukherjee A, Breselge S, Dimidi E, Marco ML, and Cotter PD. Fermented foods and gastrointestinal health: underlying mechanisms. Nat Rev Gastroenterol Hepatol (2024) 21(4):248–66. doi: 10.1038/s41575-023-00869-x
242. EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific opinion on the substantiation of health claims related to live yoghurt cultures and improved lactose digestion (ID 1143, 2976) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J (2010) 8(10):1763–81. doi: 10.2903/j.efsa.2010.1763
243. Ramos CL, Bressani APP, Batista NN, Martinez SJ, Dias DR, and Schwan RF. Indigenous fermented foods: nutritional and safety aspects. Curr Opin Food Sci (2023) 53:101075. doi: 10.1016/j.cofs.2023.101075
244. Louw NL, Lele K, Ye R, Edwards CB, and Wolfe BE. Microbiome assembly in fermented foods. Annu Rev Microbiol (2023) 77:381–402. doi: 10.1146/annurev-micro-032521-041956
245. Marco ML, Sanders ME, Gänzle M, Arrieta MC, Cotter PD, De Vuyst L, et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on fermented foods. Nat Rev Gastroenterol Hepatol (2021) 18(3):196–208. doi: 10.1038/s41575-020-00390-5
246. Hill C, Tancredi DJ, Cifelli CJ, Slavin JL, Gahche J, Marco ML, et al. Positive health outcomes associated with live microbe intake from foods, including fermented foods, assessed using the NHANES database. J Nutr (2023) 153(4):1143–9. doi: 10.1016/j.tjnut.2023.02.019
247. Iyer A, Mukherjee A, Gómez-Sala B, O’Connor EM, Kenny JG, and Cotter PD. The impact of live dietary microbes on health: a scoping review. J Food Sci (2024) 89(2):773–92. doi: 10.1111/1750-3841.16893
248. Marco ML, Hill C, Hutkins R, Slavin J, Tancredi DJ, Merenstein D, et al. Should there be a recommended daily intake of microbes? J Nutr (2020) 150(12):3061–7. doi: 10.1093/jn/nxaa323
249. Dhami M, Raj K, and Singh S. Relevance of gut microbiota to Alzheimer’s disease (AD): potential effects of probiotic in management of AD. Aging Health Res (2023) 3(1):100128. doi: 10.1016/j.ahr.2023.100128
250. Kim C-S, Jung S, Hwang G-S, and Shin D-M. Gut microbiota indole-3-propionic acid mediates neuroprotective effect of probiotic consumption in healthy elderly: a randomized, double-blind, placebo-controlled, multicenter trial and in vitro study. Clin Nutr (2023) 42(6):1025–33. doi: 10.1016/j.clnu.2023.04.001
251. Soheilian Khorzoghi M, Rostami-Nejad M, Yadegar A, Dabiri H, Hadadi A, and Rodrigo L. Impact of probiotics on gut microbiota composition and clinical symptoms of coeliac disease patients following gluten-free diet. Contemp Clin Trials Commun (2023) 35:101201. doi: 10.1016/j.conctc.2023.101201
252. Xia B, Liu X, Li Z, Ren J, and Liu X. The effects of microbiota-targeted approaches in inflammatory bowel disease: probiotics, probiotic foods, and prebiotics. Curr Opin Food Sci (2023) 49:100956. doi: 10.1016/j.cofs.2022.100956
253. Timmis K, Hallsworth JE, McGenity TJ, Armstrong R, Colom MF, Karahan ZC, et al. A concept for international societally relevant microbiology education and microbiology knowledge promulgation in society. Microb Biotechnol (2024) 17(5):e14456. doi: 10.1111/1751-7915.14456
254. Debode F, Caulier S, Demeter S, Dubois B, Gelhay V, Hulin J, et al. Roadmap for the integration of environmental microbiomes in risk assessments under EFSA’s remit. EFSA Support Publ (2024) 21(2):8602E. doi: 10.2903/sp.efsa.2024.EN-8602
255. Amos GCA, Logan A, Anwar S, Fritzsche M, Mate R, Bleazard T, et al. Developing standards for the microbiome field. Microbiome (2020) 8(1):98. doi: 10.1186/s40168-020-00856-3
256. D’Hondt K, Kostic T, McDowell R, Eudes F, Singh BK, Sarkar S, et al. Microbiome innovations for a sustainable future. Nat Microbiol (2021) 6(2):138–42. doi: 10.1038/s41564-020-00857-w
Keywords: meta-omic approaches, microbiome mapping, high-throughput sequencing, food-omics, microbiome interconnection, bacteriophages, agri-food system
Citation: Fernández-Gómez P, Leong D, Berg G, Brennan F, Caruso T, Charles TC, Cocolin LS, Lange L, McAuliffe O, Maguin E, O’Sullivan O, Sanz Y, Sarand I, Sessitsch A, Smidt H, Brereton N, Candela M, Kenny J, Kostic T, Mahony J, Wagner M and Cotter PD. Harnessing agri-food system microbiomes for sustainability and human health. Front Sci (2025) 3:1575468. doi: 10.3389/fsci.2025.1575468
Received: 12 February 2025; Accepted: 02 June 2025;
Published: 31 July 2025.
Edited by:
Francesca Turroni, University of Parma, ItalyReviewed by:
Michael Gänzle, University of Alberta, CanadaChristophe Lacroix, ETH Zürich, Switzerland
Copyright © 2025 Fernández-Gómez, Leong, Berg, Brennan, Caruso, Charles, Cocolin, Lange, McAuliffe, Maguin, O’Sullivan, Sanz, Sarand, Sessitsch, Smidt, Brereton, Candela, Kenny, Kostic, Mahony, Wagner and Cotter. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paul D. Cotter, UGF1bC5Db3R0ZXJAdGVhZ2FzYy5pZQ==