 Nguyen Khoi Nghia1*
Nguyen Khoi Nghia1* Kovacs Emoke Dalma2
Kovacs Emoke Dalma2 Kovacs Melinda Haydee2Le Thi Xa3Lois Wright Morton4Hüseyin Barış Tecimen5Javad Robatjazi6
Kovacs Melinda Haydee2Le Thi Xa3Lois Wright Morton4Hüseyin Barış Tecimen5Javad Robatjazi6 Jegan Sekar7
Jegan Sekar7 Hendra Gonsalve W. Lasar6Trung Thanh Nguyen8Nguyen Minh Phuong1
Hendra Gonsalve W. Lasar6Trung Thanh Nguyen8Nguyen Minh Phuong1 Chau Thi Anh Thy1Do Thanh Luan1Nguyen Thi Kieu Oanh1
Chau Thi Anh Thy1Do Thanh Luan1Nguyen Thi Kieu Oanh1- 1Department of Soil Sciences, College of Agriculture, Can Tho University, Can Tho, Vietnam
- 2National Institute for Research and Development in Optoelectronics INOE 2000, Research Institute for Analytical Instrumentation Subsidiary, Cluj-Napoca, Romania
- 3Faculty of Basic, Soc Trang Community College, Soc Trang, Vietnam
- 4College of Agriculture and Life Sciences, Department of Sociology and Criminal Justice, Iowa State University, Ames, IA, United States
- 5Department of Environmental Sciences, College of Agriculture, Tennessee State University, Nashville, TN, United States
- 6Department of Soil and Crop Sciences, Texas A&M University, College Station, TX, United States
- 7M. S. Swaminathan Research Foundation, 3rd Cross, Institutional Area Taramani, Chennai, Tamil Nadu, India
- 8Center for Pharmaceutical Biotechnology, College of Medicine and Pharmacy, Duy Tan Univeristy, Danang, Vietnam
Introduction: Longan is a perennial crop and profitable export fruit well-suited to the Mekong Delta's climate and soils. Although soil deterioration in longan orchards has been studied, little research has addressed soil bacterial communities and their role in soil health. This study investigated the structure of soil bacterial communities and their associations with soil physicochemical properties in longan orchards.
Methods: Soil samples were collected from longan orchards cultivated for 15 (N15), 20 (N20) and 30 years (N30) in Vinh Long Province, Vietnam. High-throughput sequencing of 16S rRNA amplicons was employed to assess bacterial diversity and community composition, followed by correlation analyses with soil properties.
Results and Discussion: The dominant bacterial phyla identified across sites included Acidobacteria, Proteobacteria, Chloroflexi, Actinobacteria, and Bacteriodota. Bacterial abundance in these soils was positively correlated with sand, pH, NO3- and phosphate solubilizing microorganisms, and negatively correlated with moisture, silt, clay, NH4+, exchangeable potassium and available manganese. Longan orchard soils from all three time periods (N15, N20, N30) had more microbial species in common, than in unique ways. However, time under cultivation was an important factor, with species diversity decreasing with age. The youngest orchards (N15) exhibited the highest number of soil bacterial diversity, followed by decreasing diversity as the orchards aged (N20) and lowest at 30 years (N30). Soil properties exhibited complex and interlinked relationships with soil bacterial phyla. Overall, long-term cultivation of longan orchard soil negatively impacted microbial diversity, potentially altering key properties of soil, hence underscoring the necessity for sustainable soil management to preserve soil health and production.
Highlights
● Long-term cultivation of longan reduces soil health.
● Acidobacteria, Proteobacteria, Chloroflexi, and Actinobacteria were dominant in longan soils.
● Long-term longan cultivation soil decreased the richness and abundance of the soil bacterial community.
● Sand, pH, and NO3- strongly affect soil bacterial taxonomy in cultivated longan soils.
1 Introduction
The perennial fruit tree longan (Dimocarpus longan Lour) has high nutritional and commodity value and is extensively grown in the Vietnamese Mekong Delta providing livelihoods and household food security (1–3). In 2022, Vinh Long Province had approximately 5,853 ha of longan orchards, concentrated mainly in Long Ho District, which grows more than 4,000 ha (4). Longans are grown primarily in many types of soil including alluvial, sandy soils. They need highly fertilized soils for sustained growth and productivity of these plants (5). In the last few years, the fertility of longan orchard soils has declined over time because of intensive farming practices and high chemical use and long-term use of inorganic fertilizers to replace organic fertilizer has contributed to the loss of soil fertility (6). Furthermore, exchangeable cations are certainly detached from the topsoil when soil is moistened by rainwater and irrigation under tropical weather conditions, resulting in an increase in the process of soil acidity and compacting (7). Thus, orchard soil properties, especially soil microbial communities, are significantly altered after long-term cultivation (8–10).
Other studies have reported that soil exposure to several soil degradation and erosion processes, changing in soil fertility level, soil organic matter content, and salinization. These processes strongly affect soil health, fruit yield, and quality (11, 12). Soils cultivated with perennial fruit trees such as longan, and mango in the Vietnamese Mekong River Delta region are recently experiencing decreasing pH of the soil levels, low levels of nutrients and soil organic carbon, unbalanced nutrition, and declined activity of microbes as orchards grow old (3, 13). The soil microbial community diversity is one of the important soil health indicators (14). This is the principle of soil function (15, 16). In the disciplines of soil science, microbiology, and biodiversity, soil diversity of microbes is a freshly developed interdisciplinary topic that has garnered more attention lately. Research on the diversity of soil microbes is crucial for understanding natural biological processes, adapting to climate change, reducing different forms of pollution, preserving ecological service functions, and encouraging the sustainable use of soil. Numerous factors, which can be broadly categorized as natural and human causes, influence the variety and organization of soil microbes, particularly functional diversity. Natural factors contain vegetation type (17, 18), soil type (19), temperature and moisture (20). Human factors include management practices such as applied pesticides, fertilization (21) and soil tillage methods (22). Furthermore, microbial biomass, community metabolic diversity, and the quantity of microorganisms can all be considerably increased by leguminous plants (23). Microbial communities’ transition from rocky, bare soil to forest environments, and the diversity and abundance of microorganisms in forest ecosystems highlight the importance of plants (24). Another study found that cover cropping improves soil characteristics, microbial communities, and microbial functions while also reducing pesticide use, orchard maintenance expenses, and underground carbon storage (25). Microbial biomass carbon (MBC) showed a significant correlation with soil organic carbon (SOC) rising from 700 mg.kg−1 MBC at 25 g.kg−1 SOC to 2240 mg.kg−1 MBC at 90 g.kg−1. Most likely, SOC promotes MBC by releasing water-soluble organic carbon (26). A separate study indicated that soil age exerted a more significant influence on bacterial populations compared to fungal communities. This study indicated that soil characteristics involving pH, organic matter, total nitrogen, total phosphorus, and total potassium affected alterations in soil microbiota (27).
The physicochemical characteristics of soil, in addition to the kinds of vegetation communities, such as the size of the soil particles, the amount of water present, the pH, the activity of the enzymes, and the amount of organic matter, all factors affect on the diversity of soil microbial community structure. The prior research on the variety of soil microbes under various circumstances has mostly concentrated on soil characteristics and human influences. There haven’t been many studies on how the duration of raised garden beds impacts the physical and chemical properties of fruit plantation soil and microbial diversity. Through the examination of carbon dioxide and mineral composition, the correlations between the microbial community and environmental conditions were particularly interesting. There is a dearth of information on the variety and composition of the bacterial population in the soils used to grow longans in Vietnam’s Mekong Delta. To address this knowledge gap, the present study investigates the soil bacterial community structure and diversity, and explores the relationships between bacterial assemblages and soil properties in longan orchards of the Mekong Delta, with an emphasis on changes associated with orchard age.
2 Materials and methods
2.1 Study region
The study site of our research was conducted at An Binh commune, Long Ho district, Vinh Long Province (10°17′37.5″N 105°58′55.7″E) which is the famous place for longan cultivation. More information about this study site please refer to (3).
2.2 Soil sampling
Following the last longan harvest, nine soil samples from three distinct groups of longan trees were gathered from longan orchards in July 2023 in Vinh long province, Vietnam. Based on the age of the longan tree, the samples were divided into three groups: group 1 (N15), which was 15 years old; group 2 (N20), which was 20 years old; and group 3 (N30), which was 30 years old. Three different soil samples from three distinct longan tree ages within the same group were collected, corresponding to three different replicates for each group of longan tree age group, thus, in total there were 9 soil samples were collected in this study from nine different longan orchards. Using the Zig-Zag curve method, a coring soil auger with a diameter of 5 cm (Seymour 21306 AU-S6, USA) was used under the canopy of longan trees to ensure homogeneity. Refer to (3) for the method and procedures for soil sample collection and handling of the soil samples after collection. The selected sites of 9 different locations referred as 9 different soil samples of this study based on the criteria of the same local longan variety named as “Xuong com vang” using more or less the same cultivation techniques. Information regarding the age of the orchards and raised beds is detailly presented in Table 1.
Table 1. Information of the nine selected longan orchards in An Binh commune, Long Ho district, Vinh Long province, Vietnam used for this study.
2.3 Soil bacterial community structural analysis
The methods for total genomic DNA extraction, amplicon generation, library preparation, Illumina sequencing, and data analysis followed the same procedures as described by (28).
Amplicons targeting the V3–V4 regions of the 16S rRNA gene were generated using specific primers. The forward primer contains the sequence ‘CCTACGGRRBGCASCAGKVRVGAAT’ and the reverse primers contain the sequence ‘GGACTACNVGGGTWTCTAATCC’.
Next-generation sequencing was performed using the Illumina Miseq/Novaseq platform with paired-end sequencing. Sequences containing ambiguous ‘N’ characters were removed, retaining only those longer than 200 base pairs.
After qualifying by filtering and removing chimera, sequences were clustered into OTUs using VSEARCH (v1.9.6) at 97% similarity, with taxonomic assignment based on the Silva 138 database. The RDP classifier (Bayesian algorithm) was used to identify representative OTU sequences and analyze community composition at various taxonomic levels.
Alpha diversity metrics (e.g., Shannon, Chao1), rarefaction curves, and rank-abundance plots were used to assess species richness and evenness. Beta diversity was evaluated using (un)weighted UniFrac distances, and visualized through PCA, PCoA, and NMDS analyses. UPGMA clustering was performed using hierarchical clustering and average linkage methods.
Statistical tests such as ANOSIM, Metastats, LEfSE, Adonis, STAMP, and Wilcoxon rank-sum test were used to detect significant differences in microbial communities between groups. Environmental factors were integrated through heatmaps, RDA/CCA analyses, and network diagrams to explore their influence on microbial community composition and interactions. These approaches help identify dominant species and their roles in maintaining microbial community structure and functional stability.
The Venn graph shows the average of the three-sequence samples analyzed in each group. The averaging was performed by identifying the OTUs present in each sample within a group, then aggregating and selecting representative OTUs for the entire group. OTUs were considered “representative” if they appeared frequently in the majority (e.g., ≥2/3) of the samples within the same group. This approach allows the Venn diagram to more accurately reflect the stable and typical microbial diversity of each time-based group. Simply, we just used the data from three replicates (OTUs) for each soil group to make the average values for the intersecting (common) and non-intersecting (unique) parts of each soil group.
2.4 Soil physical, chemical and biological property analysis
In this study the soil physical, chemical and biological properties including soil porosity, soil texture, bulk density, moisture content, pHw (pH from soil extract by water), electrical conductivity (EC), total nitrogen (TN), soil organic matter (SOM), total phosphorus (TP), total potassium (TK), available nitrogen (NH4+, NO3-), available phosphorus (AP), exchangeable potassium (K+), exchangeable calcium (Ca2+), available silicate (SiO2), total boron (B), zinc (Zn), copper (Cu), manganese (Mn), silicate-solubilizing bacteria, calcium-solubilizing bacteria, nitrogen-fixing bacteria, phosphorpus-solubilizing bacteria, and potassium-solubilizing bacteria, urease, β-glucosidase, phytase, and phosphomonoesterase were measured. For all the information regarding the method of analysis for these above properties refer to (3). Additionally, we adopted data for the soil physical, chemical and biological properties of nine soil samples used in this study, namely L6, L13, L17 (group N15), L2, L3, L5 (group N20), L7, L9, L16 (group N30) from the previous published paper (3) of our working group. However, in our previously published paper (3) we grouped the soil properties according to the ages of the raised beds of longan orchards while in this study we grouped the soil properties according to the age of the longan trees, though to avoid the data repetition usage, we just used the data of the soil physical, chemical and biological properties of only nine selected soil samples according to the age of longan trees to assess the effect of longan tree’s age on bacterial community structure rather than to analyze the effect of the age of longan trees on soil attributes in depth and to avoid the overlong manuscript of this study, we did not present separately the soil physical, chemical and biological properties, but just did focused on the analysis of soil bacterial community structure and genetic diversity related to the longan tree’s age.
2.5 Statistical analysis
The data was analyzed using one-way analysis of variance (ANOVA). These statistical evaluations provided a thorough understanding of how different soil characteristics interact. Tukey’s test was computed at the 5% level to confirm significant differences, and the results are shown as means ± standard deviations (SDs). Minitab (version 19) was used to perform statistical tests. The “cor” function was used to perform Pearson’s correlation, and the “corrplot” package was used to create a correlation matrix.
3 Results and discussion
3.1 The influence of longan trees’ age on the bacterial diversity in longan orchard soil
3.1.1 Distribution of bacterial taxonomy
Figure 1 presents a detailed bar chart analysis of the bacterial taxonomic distribution in soil samples collected from longan orchards of varying ages. The data encompass multiple taxonomic levels, ranging from the broad classification of kingdom to the finer resolution at the species level. A consistent gradient of taxonomic richness is evident following the pattern: N15 > N20 > N30. Specifically, 15-year-old orchards (N15) presented significantly greater bacterial diversity than 20-year-old (N20) and 30-year-old (N30) orchards did. At lower hierarchical taxonomic ranks such as Order, Family, Genus, and Species, the reduction in bacterial counts was approximately 20% in N20 and 40% in N30 relative to those in the N15 group. Significant disparities among various age groups were discovered, as indicated by the various letters above the bars (Figure 1). Furthermore, the pronounced disparities at the genus and Species levels underscore the sensitivity of microbial community structures to orchard age. The inverse relationship between orchard age and bacterial taxonomic diversity observed in this study aligns with the finding of higher bacterial community abundance and elevated ascomycetes richness indices in younger orchard systems (29). This temporal pattern in microbial community composition suggests age-dependent modifications in the soil microbiota structure, potentially influenced by long-term changes in rhizosphere dynamics (30) and soil physicochemical properties (31). The greater bacterial diversity may enhance soil health through more robust nutrient cycling and a resilient biological network (32). Conversely, the decline in bacterial diversity with increasing orchard age may reflect changes in soil physicochemical properties and management practices over time (33, 34). The quantitative resolution and statistical robustness of taxonomic analysis facilitate comprehensive comparisons across hierarchical levels, providing valuable insights into the temporal dynamics of soil microbial communities under long-term orchard management (35).
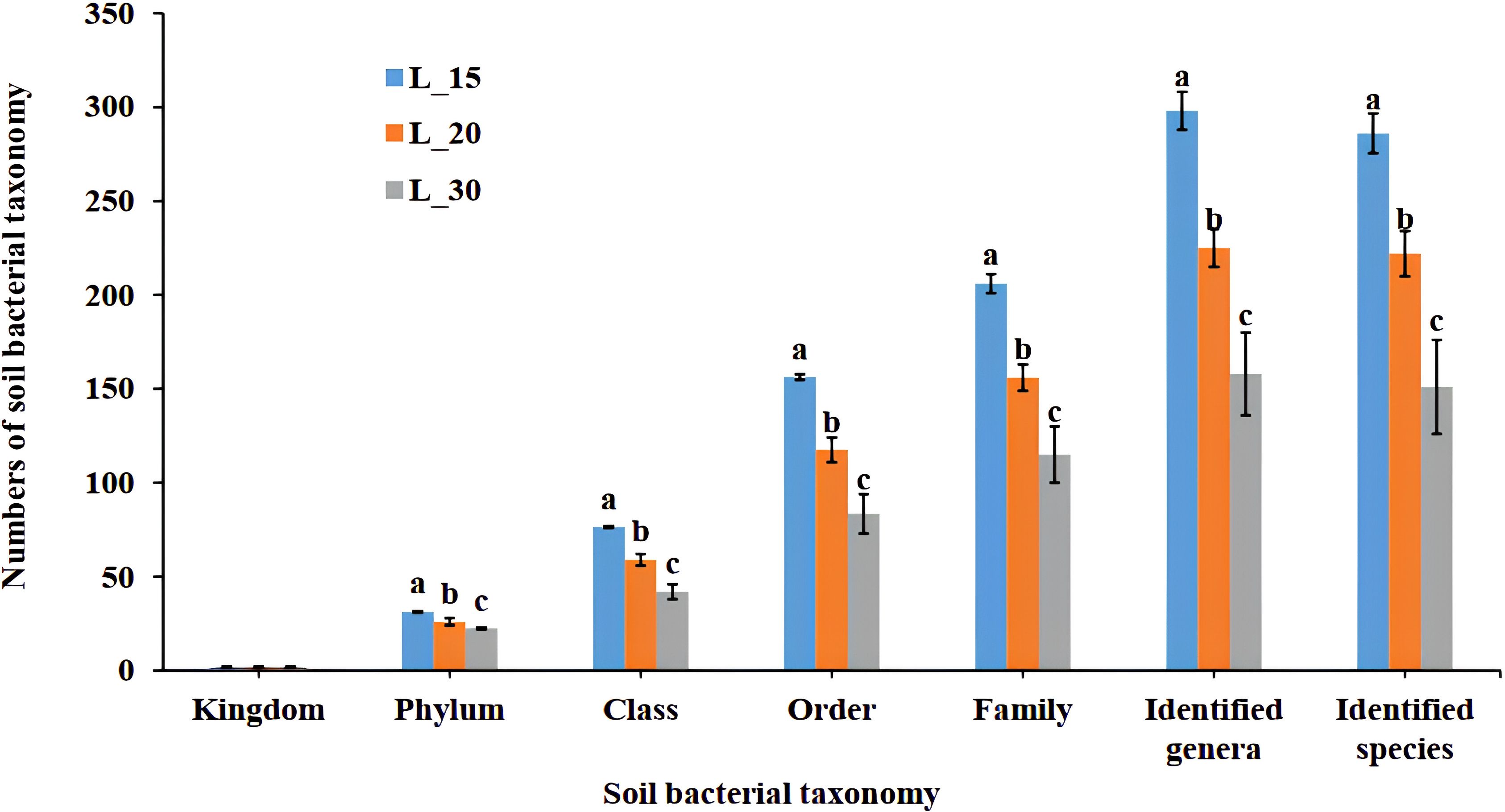
Figure 1. Statistics of taxonomic composition. N15, 15-year-old longan orchard; N20, 20-year-old longan orchard; N30, 30-year-old longan orchard. Means in the same group with different letters are significantly (p < 0:05) difference according to Tukey’s test.
3.1.2 Alpha diversity and beta diversity of the microbiota in longan orchard soil
Alpha diversity indices, specifically the Chao 1 and Shannon indices, were employed to quantitatively assess the microbial abundance and diversity of Operational Taxonomic Units (OTUs) in longan orchard soils of varying ages. The Chao index serves as a nonparametric estimator of species richness, with higher values indicating a greater number of OTUs within the sample; this infers that the soil supports a richer species composition. In parallel, the Shannon index integrates both species richness and evenness, providing an overview of community diversity. Elevated Shannon index values therefore reflect a more complex and uniformly distributed microbial community. Figure 2 presents these indices as box plots showing the variation in species abundance and diversity across the different orchard age cohorts. The 15-year-old orchard (N15) exhibits the highest median Chao1 values, suggesting that younger orchard soils harbor a more abundant microbial community (Figure 2A). The interquartile range for N15 was relatively narrow, indicating reduced variability in species richness compared with that in older orchard soils. Conversely, the 30-year-old orchard (N30) showed a substantially lower median and broader spread, reflecting both diminished richness and higher variability in the microbial community composition. The 20-year-old orchard (N20) presented intermediate values, supporting a progressive decline in species richness with orchard age. The Shannon index analysis (Figure 2B) corroborated these findings, revealing a comparable trend of higher diversity and an even distribution of the bacterial community in younger soils (N15) and reduced diversity in older ones (N30). The sequential reduction in the Shannon index values from N15 to N30 suggested that prolonged cultivation led to both species loss and increased dominance of specific bacterial groups, resulting in less balanced community structures. The consistent pattern observed across both diversity metrics strengthens the evidence that orchard age has a substantial influence on the soil bacterial community composition. The cumulative effects of management tackles may be one of the reasons contributing to this orchard-age-dependent drop in microbial diversity and community complexity (33), alterations in soil physicochemical properties (32), or evolutionary shifts in plant-soil interactions (36) over time. Such decreases in microbial diversity could be indicative of cumulative changes in soil properties, nutrient availability, or long-term management practices, all of which may cause stress in the microbial ecosystem (37).
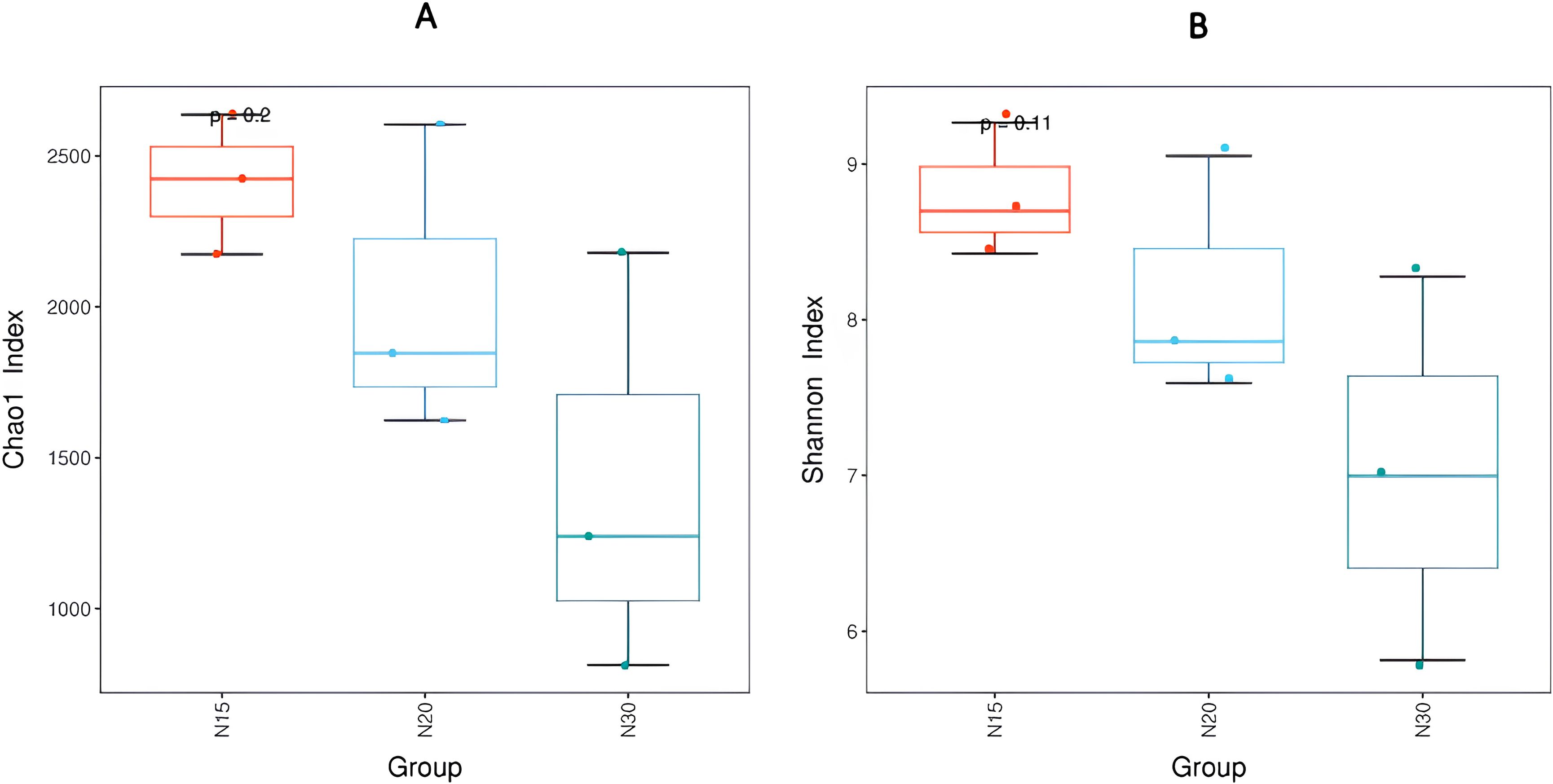
Figure 2. Alpha diversity boxplots of between-group species abundance and diversity comparison. (A) The left panel is the Chao1 index boxplot of each age group. X axis indicates the years cultivated of the groups and Y axis indicates the Chao 1 index. Each box diagram shows the minimum, first quartile, medium, third quartile and maximum values of the Chao1 index of the corresponding sample and the P value represents a significant difference. The right graph is the Shannon index boxplot of each group, revealing similar patterns of less diversity in bacterial community species as longan orchards age. (B) N15:15-year-old longan orchard; N20:20-year-old longan orchard; N30:30-year-old longan orchard.
Supplementary Table S1 presents indices of α-diversity to better understand and compare the species diversity of the three different orchard soils at 15 years, 20 years, and 30 years. The community richness of the three soil samples were significantly differed from the Ace diversity index of the bacterial communities for N15, N20 and N30, with values of 2,504, 1704 and 1,028, respectively. The Chao1 species richness index showed a similar pattern, with community richness decreasing as orchard soils aged. The Shannon index of community diversity significantly differed among the three different aged orchard soils, with a similar pattern of decreasing diversity in the older orchard soils. The Simpson index was not significant. This study explored the correlation between soil properties and alpha diversity in longan plantation soils in Vietnam’s Mekong River Delta, with a focus on understanding how soil characteristics influence microbial diversity (Supplementary Table S2). Distinct patterns emerged when the alpha diversity indices, including the Ace, Chao1, Shannon, and Observed_OTU indices, were analyzed in relation to various soil properties (Supplementary Table S2). Negative correlations were evident between Ace, Chao1, and several soil properties, notably moisture content, silt, clay, pHw (1:2.5), and exchangeable calcium. These findings suggest that higher levels of these soil properties correspond to reduced species richness, indicative of potential constraints on microbial diversity within longan orchard soils (38–40). Similarly, there were adverse correlations found among some soil characteristics and the Shannon diversity index, such as moisture content, silt, clay, pHw (1:2.5), exchangeable calcium, and available manganese. This implies that these soil characteristics may negatively impact species diversity within the microbial community inhabiting longan orchard soil (41, 42). The analysis of the observed OTUs also revealed negative correlations with various soil properties, including moisture content, silt, clay, pHw (1:2.5), exchangeable calcium, and available manganese. This suggests that higher values of these soil properties are associated with a lower number of observed OTUs, reflecting diminished microbial diversity within longan orchard soils (43). These findings underscore the potential limitations imposed by certain soil properties on microbial diversity within longan orchard soils. An elevated moisture content may suppress microbial diversity, whereas a relatively high clay content might restrict microbial movement and activity (44). Soil acidity, as indicated by pHw (1:2.5), may also constrain microbial diversity, along with nutrient availability, as reflected in the negative correlations with exchangeable calcium and available manganese (45, 46). Interestingly, the presence of phosphorous-solubilizing bacteria (PSB) did not consistently correlate with alpha diversity, suggesting a nuanced relationship between microbial community composition and specific functional groups within the soil microbiome (47). These results highlight the intricate interplay between soil properties and microbial diversity in agricultural soils, emphasizing the need for further research to elucidate the underlying mechanisms and develop strategies for enhancing microbial diversity and soil health within longan orchard systems (19).
3.1.3 Phylum and genus distributions
Analysis of the top 30 microbial phyla and their abundances (percent present) revealed the structure of the microbial communities in the longan orchard soil samples and their variation over time (Figure 3). The phylum Acidobacteriota was the most abundant phylum in N15 (50%), N20 and N30 (~35%) followed by Proteobacteria, Chloroflexi, and Actinobacteria for all 3 samples. Notably, the phyla Proteobacteria and Actinobacteria were most abundant in N30 (~35%), were less abundant in soil sample N20 and were least abundant in sample N15 (Figure 3A). The phylum-level taxonomic distribution indicated that the microbial community structure associated with the longan cultivation time, and it was connected with several of taxa, among which the dominant taxon was Acidobacteriota. However, the largest and most metabolically diverse group of soil bacteria is represented by Proteobacteria (48). A study reported Proteobacteria as the major phylum in rice (49). In addition to Acidobacteria, the most abundant taxa included Proteobacteria, Firmicutes, Alphaproteobacteria, Actinobacteria, Gammaproteobacteria, and Chloroflexi (50–52). Our findings are in agreement with these reports. In the three distinct longan farms, the bacteria of the phyla Acidobacteriota, Proteobacteria, Chloroflexi, Actinobacteriota, Bateroidota, Verrrucomicrobiota, Gemmatimonadota, and Myxococcota were the most frequently identified. The top four phyla that showed the most rises among soil samples in N15 were Acidobacteriota, Verrrucomicrobiota, Myxococcota, and Desulfobacterota, according to additional study. These four phyla of soil bacteria have been proposed to be advantageous to soil because they provide a variety of purposes for the ecology of soil (53). Acidobacteriota have been classified as oligotrophic bacteria due to how they primarily thrive and multiply in nutrient-poor and acidic soil settings (54). The phylum Verrucomicrobiota lives in soil. Methylacidiphilaceae is a family of Verrucomicrobiota that are methanotrophic, reducing the CH4 in the atmosphere (55), whereas the other family members function in the degradation of plant-derived organic matter to release nutrients (56). Myxococcota contains a notable family, Myxococcaceae, that plays a very important role in the soil carbon cycle (57, 58), bio-pesticide and bio-fertilizer for plants and soil (59). Desulfobacterota is a bacterial phylum comprising the anaerobic genus Desulfobacter, which plays an important role in the salinized field of rice. This genus is an early anaerobic prokaryotic microbe in the biosphere that gets exposed to sulfate-reducing bacteria. It has considerable promise for microbial remediation and is plays a role in the biogeochemical cycle of sulfur, organic matter degradation, metal ion reduction, and other processes. This mechanism can significantly decrease the levels of soluble inorganic salts in salinized soils through the swift and efficient adsorption and transformation of both positively and negatively charged ions (60).
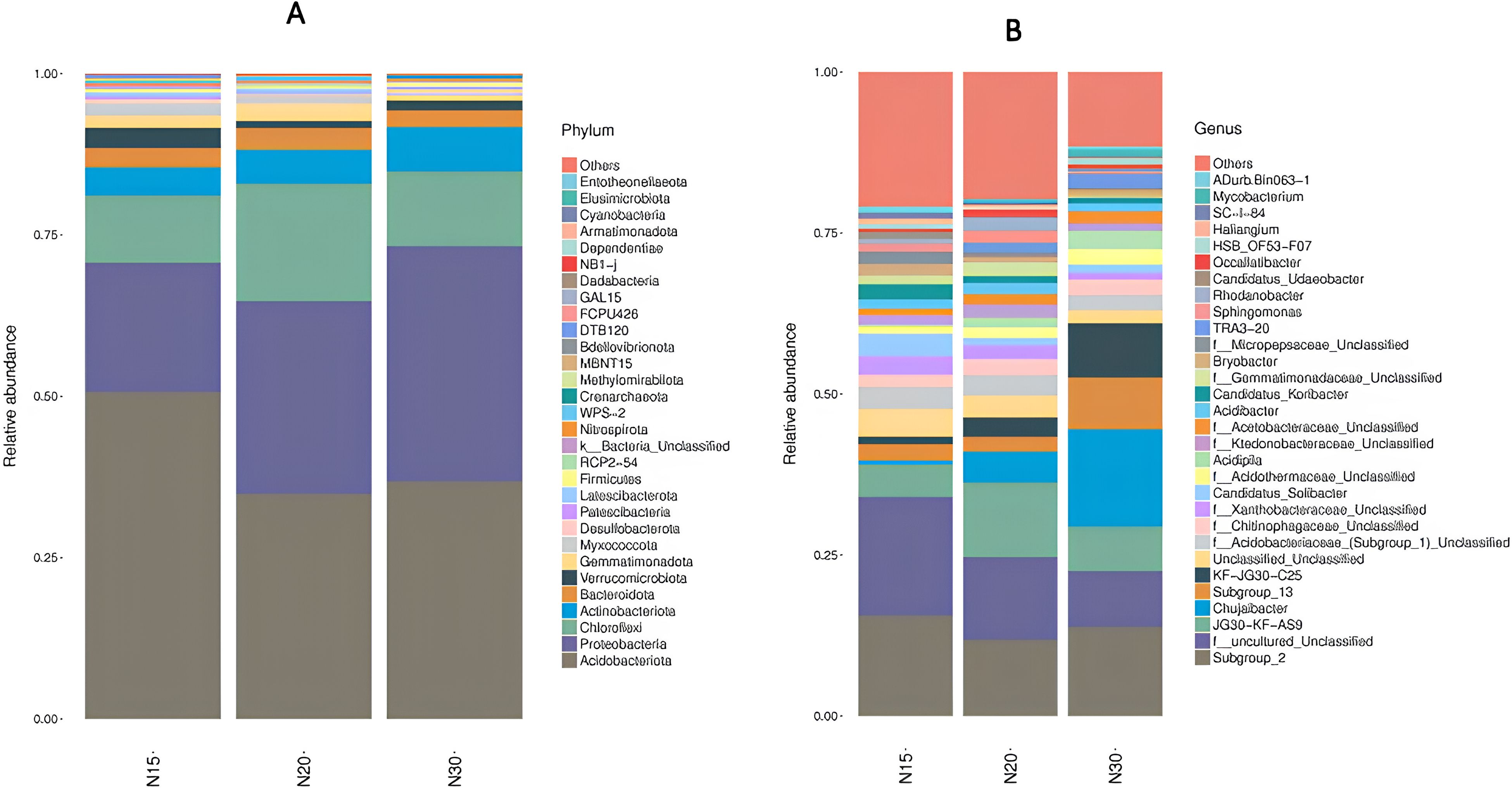
Figure 3. Stacked bar plot of phylum (A) and genus (B) distribution. X axis is the soils sampled in three age groups (15 years N15; 20 years N20; 30 years N30), and the Y axis is the relative abundance of different species. The legend colors correspond with the species taxonomic classification. 'Other' represents the relative abundance of all phylum/genus level classifications other than the top 30. N15:15-year-old longan orchard; N20:20-year-old longan orchard; N30:30-year-old longan orchard.
Conversely, the predominant phyla most significantly enriched in the N20 and N30 soil samples were Proteobacteria, Actinobacteriota, and Chloroflexi. Proteobacteria include Rhizobium sp. and Agrobacterium sp. which are advantageous to soil and plants (61). Actinobacteriota have specialized metabolisms that inhabit competitive and nutrient-limited environments with multifunctional roles in the soil (62). Numerous actinobacteria are involved in the mobilization and solubilization of nutrients, including iron and phosphates. They also operate as beneficial microorganisms in biologic nitrogen fixing and mycorrhizal symbiosis (63), whereas the Chloroflexi phylum predominates in the soil samples of N29 and N30, indicating a relationship between soil compaction in these two soil samples and chemical pesticide contamination since this phylum consists of a diverse group of organisms, including anoxygenic photoautotrophs, aerobic chemoheterotrophs, thermophilic organisms, and anaerobic organisms. They have been used to bioremediate soil pollution by reductive dehalogenation of organic chlorinated compounds as a means to obtain energy (64).
Comparing the top 30 genera identified from the soil samples collected at the longan orchard, Subgroup_2, f_uncultured_Unclassified and JG30-KF-AS9 were found to be abundant in all three soil samples (Figure 3B). Three other genera were also found to be abundant in the N30 soil samples, namely, subgroup_3, KF_JG30_C25 and Uncultured_Unclassified. In addition to the 30 analyzed genera, the study soil samples contained other genus groups. N15 had the greatest number of genera (~20%), followed by sample N20, and the lowest number of genera was detected in sample N30 (~10%). Thus, orchard soils that are 30 years or older have the lowest species richness. These findings suggest the soil microorganisms that dominate and subdominant microbiology in longan orchards change significantly over time, with reductions or increases in the special microflora. Moreover, N30 significantly stimulated the growth of the genus Chujaibacter, which is aerobic, Gram-negative, rod-shaped, and free of flagella. Phylogenetically, the genus belongs to the family Xanthomonadaceae of the order Xanthomonadales within the class Gammaproteobacteria. Chujabacter is a type of plant pathogenic bacterial genus (65). This finding suggests that the cultivation time of longan could lead to the form of a core bacterial group that causes plant pathogens under varying conditions.
3.1.4 Venn diagram analysis of the species
Venn diagrams are used to determine the number of common and unique species (such as OTUs) across multiple groups or samples, and they intuitively illustrate the similarities and overlaps of species in environmental samples. The quantity of common and unique species OTUs is shown in Figure 4, which are the average of the three-sequence samples analyzed in each group. The following pattern was detected for the total OTUs: N20 with 3371 (77.69%) > N15 with 3244 (74.74%) > N30 with 2423 (56.28%) in N30. The number of shared OTUs within N15, N20, and N30 was 1770 (40.8%). Unique individual species 479 (11.0%) were detected in N15, 712 (16.4%) were detected in N20, and 193 (4.45%) were detected in N30. The number of shared bacterial OTUs within N15 and N30 was 292 (6.745%), that within N20 and N30 was 186 (4.29%), and that within N15 and N20 was 703 (16.2%). Figure 4 illustrates that the total quantity of core OTUs was 1770, while the count of different OTUs reached 1384 across three sample groupings. This indicates that the species of bacteria in N15, N20, and N30 demonstrated both individuality and commonality, with the latter being more prevalent than the former. The N15 and N20 microbial species were very comparable, while the N30 microbial species significantly differed from the other two samples. The microbial species decreased with increasing orchard age. These results indicate that the soil microorganisms shared by longan farms N15, N20, and N30 were the most common and that there were fewer distinct microbial species associated with N30 than with the other farms. The microbial species found in longan orchards of varying ages clearly had similarities. These results imply that the species of microorganisms may be impacted by prolonged cultivation times, with long-term cultivation having a negative influence on the formation of microorganism species. The analysis of microbial communities in soils under longan cultivation of varying durations revealed that 479 microorganisms were significantly associated with the N15 group, 712 with N20, and 193 with N30. These results underscore the substantial influence of planting years on the structure and abundance of soil bacterial communities in longan orchards. Collectively, the findings indicate that prolonged continuous cropping alters both the composition and richness of soil microbiota, which may in turn affect soil quality and plant performance. This observation is consistent with previous studies reporting a decline in soil bacterial diversity with increasing years of tobacco monoculture (66–68). Such shifts are likely driven by environmental selective pressures, where long-term cultivation fosters the emergence of microbial communities better adapted to the altered soil environment (69, 70).
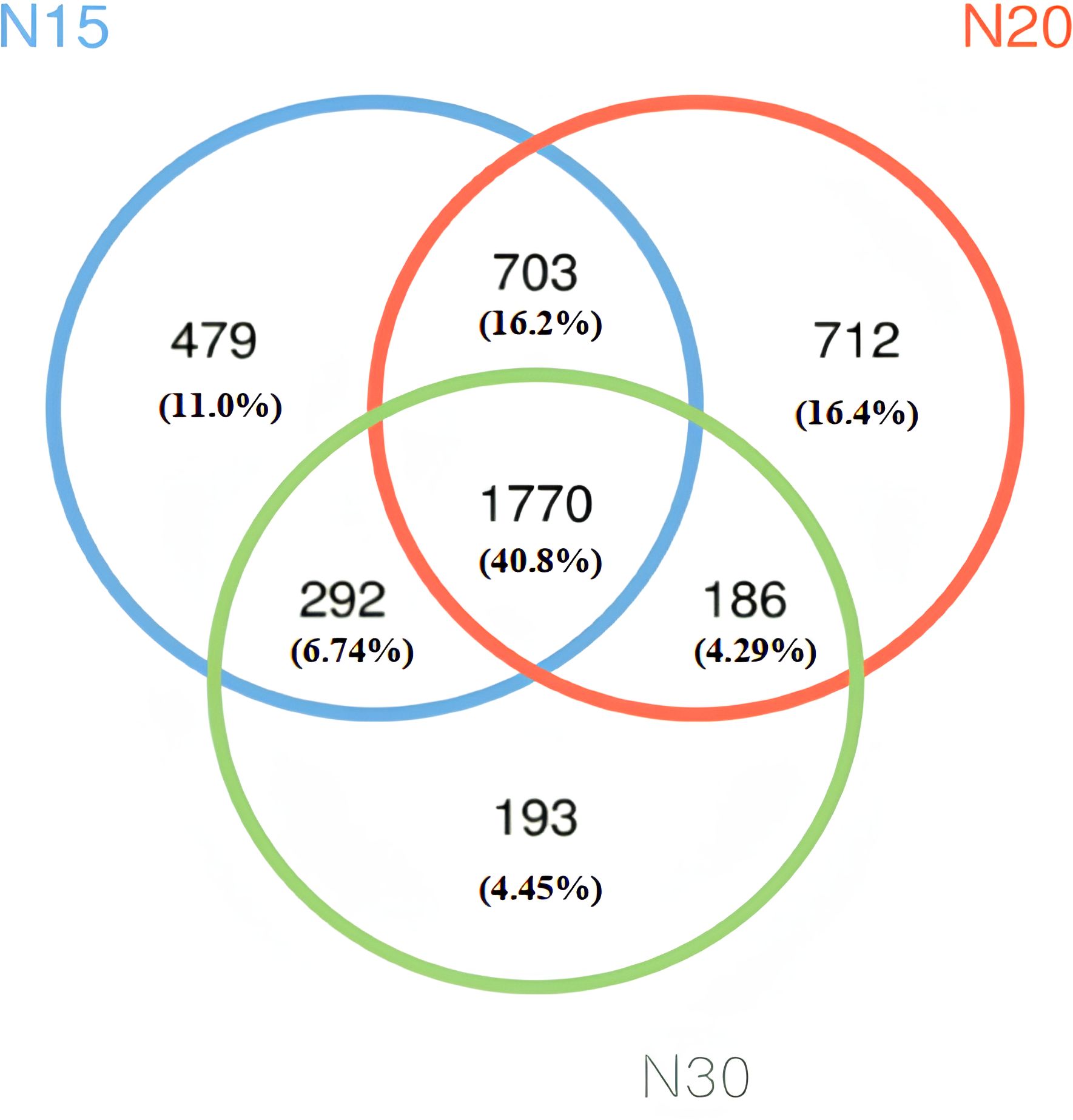
Figure 4. Venn diagram: Numbers of species (OTUs) common or unique to each group. The circles of different colors in the Venn diagram represent the three different time groups in the study based on years growing longan trees (N15, N20, N30). The numbers in the figure represent the numbers of species (OTUs) unique to each group or found in common among the groups. Areas within the circles where there is no overlap among groups represent the number of OTUs unique to that group; where all three circles overlap in the middle represent the number of OTUs shared by all groups; partial overlaps between two circles represent shared OTUs between those two-time groups. N15:15-year-old longan orchard; N20:20-year-old longan orchard; N30:30-year-old longan orchard.
3.1.5 Relative abundance analysis
Figure 5 presents the findings of the proportional abundance study of soil bacterial phyla for longan orchards across different cultivation ages. The analysis indicated that key bacterial groups, including Acidobacteriota, Proteobacteria, Chloroflexxi, Actinobateriota and Bacteridota exhibited relatively high overall abundances across all three groups of soil samples. Notably, for most phyla, there was an inverse relationship between abundance and orchard soil duration. An exception to this trend is observed for Proteobacteria, whose abundance does not decline as orchard age increases, suggesting that members of this phylum might possess adaptive traits that allow them to maintain their populations even under prolonged management or environmental stress conditions. It was also evident that longer cultivation of the orchard soil resulted in a lower abundance of bacterial groups, with the exception of the phylum Proteobacteria. In contrast, ten phyla, including Verrucomicrobiot, Gemmatimonadota, Myxococcota, Desulfobacterota, Latescibacterota, Firmicutes, an undefined group (k:Bacteria_Uncl), Nitrospirota, WPS-2, and Crenarchaeota, were consistently less abundant across all the soil samples. The relative scarcity of these phyla may reflect their dependence on specific ecological niches or substrate availability that are less prevalent or may indicate competitive exclusion by more dominant bacterial groups in the studied longan orchard soil. According to statistical analysis, the relative abundances of the majority of phyla varied significantly between the three age groups, underscoring that soil microbial communities are sensitive to the duration of cultivation. However, exceptions are noted for two phyla, Patescibacteria and RCP2-54, which do not show statistically significant differences between soils of different ages. These results suggest that these groups might be more resilient to changes induced by long-term cultivation (71) or that they fulfill functional roles that remain stable despite shifts in overall community structure (72).
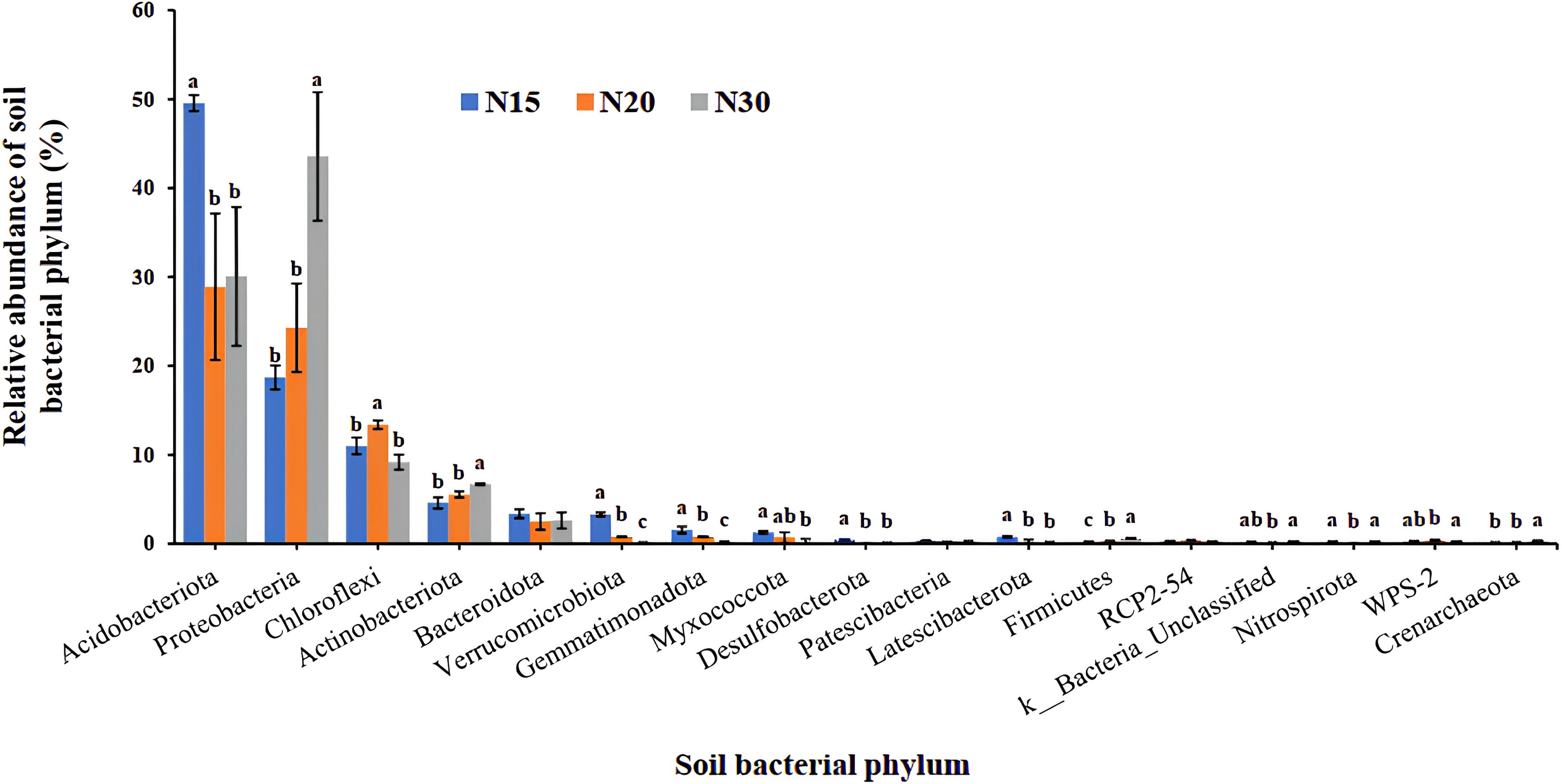
Figure 5. Relative abundance of soil bacterial phylum. N15:15-year-old longan orchard; N20:20-year-old longan orchard; N30:30-year-old longan orchard. Means in the same group with different letters are significantly (p < 0:05) difference according to Tukey’s test.
3.1.6 Correlation heatmap analysis
The heatmap (Figure 6) illustrates the Spearman correlation coefficients between the soil bacterial community composition at the phylum level and the ages of longan orchards. Hierarchical clustering analysis of the genetic correlations of 30 common phyla associated with the three ages of longan orchards revealed three distinct correlation groups. This finding demonstrated significant variations in the bacterial community structure across the different stages of the orchard. Species abundance differed across the three soil age groups. Group I encompasses less abundant phyla, such as Dadabacteria, NB1-j, and Dependentiae, which were weakly correlated across all orchard ages. The phyla in Group II included Acidobacteriota, Proteobacteria, Chloroflexi, Actinobateriota and the four phyla in Group III included Gemmatimonadota, Myxococcota, Bacteridota, and Verrucomicrobiota, which are most common in the soil (color ranging from 0.1 to 0.5). Fewer group III species were present (color<0) in all three soil samples from longan orchards. In comparison, the remaining phyla in Group 2 also had a decreasing presence in the soil samples in the order of N15>N20>30. The dendrogram topology reflects phylogenetic relationships among bacterial taxa, with closely related phyla displaying similar correlation patterns. The horizontal clustering demonstrated distinct community composition patterns across the orchard age groups, with N15 and N20 showing greater similarity than N30. This suggests a threshold effect on the community response to orchard aging (73), where both deterministic processes and temporal changes in soil conditions influence species distribution patterns (74).
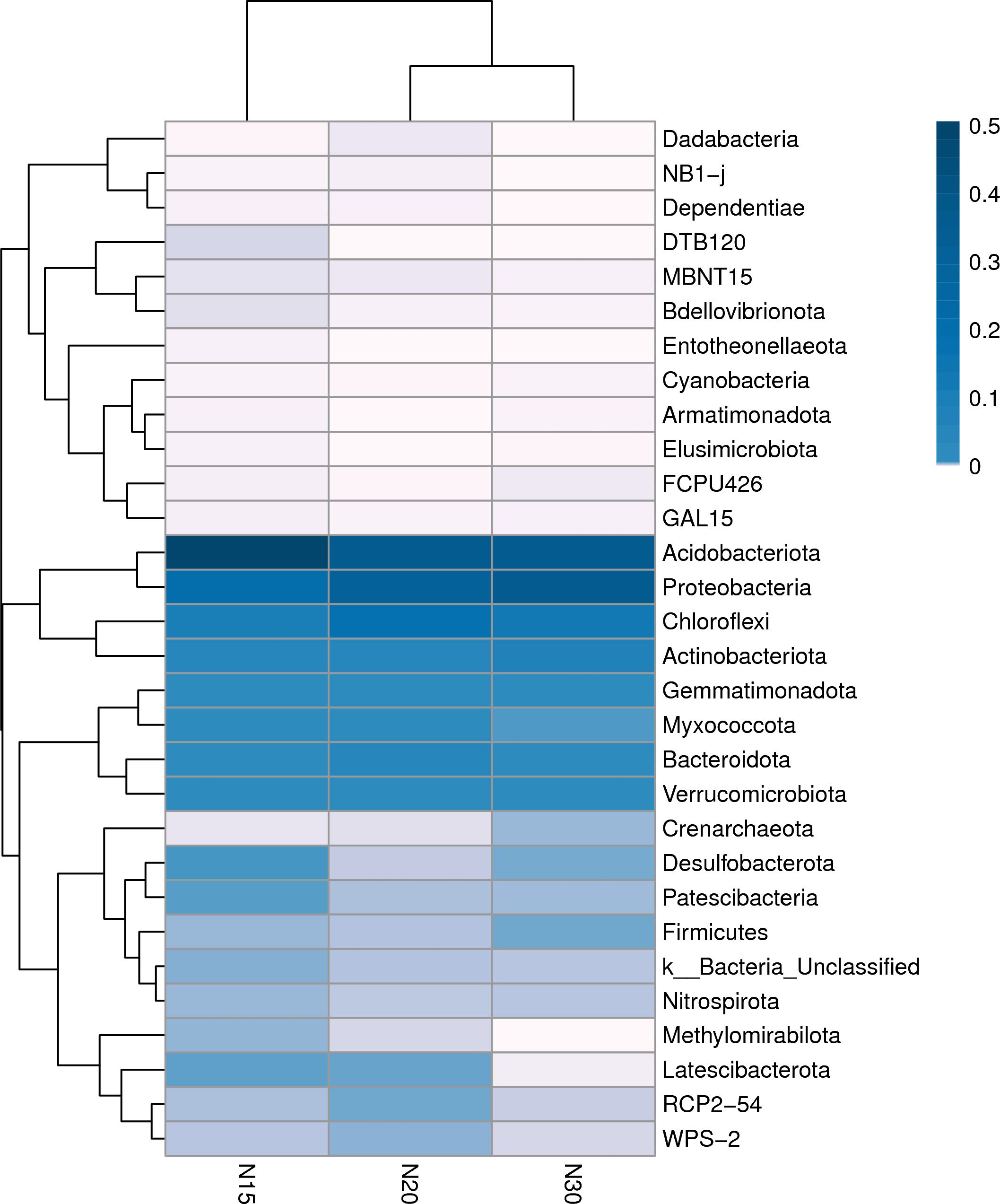
Figure 6. Species distribution heat map. The columns represent samples and/or groups and the rows represent species. The dendrogram above the heatmap is the cluster result of the samples and/or groups and the dendrogram to the left is the species cluster. The colors in the heat map represent the relative abundance of the corresponding species in the corresponding sample or group. I: 15-year-old longan orchard; II: 20-year-old longan orchard; III: 30-year-old longan orchard.
3.1.7 Principal coordinate analysis
3.1.7.1 PC1–PC2
The PCoA results indicated that samples within the same group were not tightly clustered but rather spatially dispersed, suggesting distinct microbial community structures among the groups (Figure 7). The first principal coordinate (PCoA1) accounted for 40.63% of the total variation, while the second (PCoA2) explained 21.05%, cumulatively capturing 61.68% of the variation in bacterial community composition. Notably, in the PCoA1 dimension, the bacterial communities in N15 and N20 soils were positioned in close proximity, indicating a degree of structural similarity.
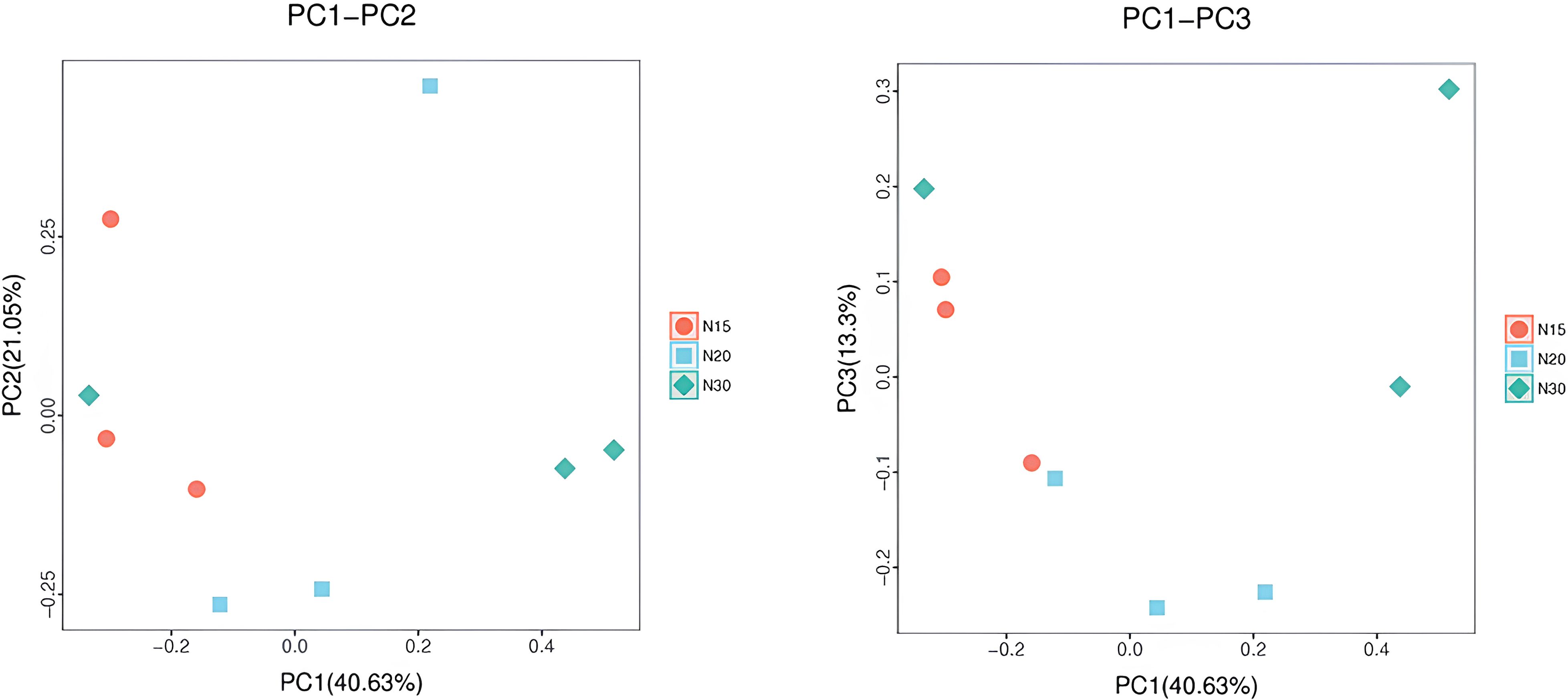
Figure 7. PCoA Plot. Samples of the same group are represented in the same color and shape. PC1_vs_PC2 is the PCoA plot obtained for the first and second principal coordinates; the X and Y axes represent the first and second principal coordinates, respectively. The value in percentage in the axis label represents the contribution of the corresponding coordinate to the sample variance and measures how much this principal is extracted from the original information. The distance between the sample points indicates the similarity of the microbial community in the sample. The closer the points, the higher the similarity. Samples clustered together are composed of similar microbial compositions. N15:15-year-old longan orchard; N20:20-year-old longan orchard; N30:30-year-old longan orchard.
3.1.7.2 PC1–PC3
PCoA1 explained 40.63% of the total variance, whereas PCoA3 contributed an additional 13.3%. Collectively, these components explained 53.93% of the variance within the bacterial community datasets, indicating that more than half of the variability in microbial composition can be attributed to these two principal coordinates. The percentage of variation in the PCoA plot demonstrated that samples from 15-year-old (N15) and 20-year-old (N20) orchards were similar to each other, suggesting that their microbial community compositions are highly similar. This close proximity implies that the transition in microbial community structure between these two age groups is minimal, potentially reflecting similar soil management practices or environmental conditions in these relatively younger orchards. Conversely, samples from older orchards likely exhibit distinct separation, indicating shifts in bacterial populations with increasing soil cultivation age. These findings are relevant for understanding the temporal evolution of the soil microbiota and can inform strategies for sustainable orchard soil management.
The PCoA analysis of PC1–PC2 and PC1–PC3 indicated significant regional differentiation across soil samples, suggesting that variations in microbial community composition were affected by the duration of longan cultivation which is directly related to soil property compositions. This study presents findings on environmental influences, including nutrient availability and soil physicochemical quality, which may have substantially influenced bacterial populations. The PC1–PC2 ordination, comprising PCoA1 and PCoA2, accounted for 61.68% of the overall variation, underscoring significant ecological gradients affecting microbial community composition. Samples from 15-year-old (N15) and 20-year-old (N20) orchards were closely aligned along the PCoA1 axis, suggesting a significant structural resemblance, potentially influenced by analogous soil management practices. Additional research has demonstrated that variables including pH, moisture, soil grain structure, and soil water retention capacity affect the development and proliferation of local microbial communities (75). The examination of PC1–PC3 corroborated these findings, with PCoA1 and PCoA3 together representing 53.93% of the overall variation. The close proximity of N15 and N20 samples in both ordinations indicates negligible alterations in the microbial population throughout the initial phase of orchard development. The increased separation noted among older orchard samples indicates that extended soil cultivation and aging processes result in substantial alterations in bacterial assemblages (3). The results align with earlier studies suggesting that prolonged land use exacerbates microbial divergence and diminishes microbial network stability (76). The findings underscore that temporal dynamics and management practices significantly influence soil microbial ecology, with crucial implications for sustainable orchard management.
3.1.8 Correlations between soil bacterial phyla in different longan orchard soils
Supplementary Table S3 presents a comprehensive correlation analysis of the bacterial phyla identified in the different longan orchard soils, revealing complex interrelationships within the microbial community structure. The Pearson correlation analysis revealed several significant correlations (either positive or negative) indicating potential ecological interactions or responses to shared environmental factors. Acidobateriota, Verrucomicrobiota, and Desulfobacterota showed a substantial beneficial correlation (p<0.01), while Actinobacteriota had a significant negative correlation (p<0.05). These relationships suggest potential synergistic interactions between Acidobacteriota and their positively correlated counterparts, possibly reflecting shared ecological niches or metabolic dependencies. The negative correlation with Actinobacteriota might indicate competitive relationships for similar resources or antagonistic interactions in the soil environment. There was a substantial positive correlation between Actinobacteriota and Proteobacteria (p<0.01), Firmicutes (p<0.001) and Crenarchaeota (p<0.01). Conversely, there was a significant negative correlation with Verrucomicrobiota (p<0.05) and Gemmatimonadota (p<0.01). This pattern suggests potential competitive relationships or opposing responses to environmental conditions between these groups. The strong positive correlations might indicate cooperative metabolic activities or similar responses to soil conditions, whereas negative correlations could reflect niche competition or differential responses to environmental stressors.
Chloroflexi was significantly negatively correlated with several phyla, including Nitrospirota and Crenarchaeota (p<0.05), but exhibited a high positive correlation with WPS_2 (p<0.001). These relationships might indicate niche differentiation or competitive exclusion processes within the soil ecosystem. These correlation patterns provide insights into the complex ecological networks within the soil bacterial communities of longan orchards. The presence of both positive and negative correlations suggests that these microbial communities are structured by a combination of cooperative and competitive interactions, as well as shared responses to environmental variables.
3.1.9 Correlation between the soil properties and the soil bacterial taxonomy
The correlation coefficients between various soil properties and soil bacterial taxonomy are detailed in Table 2 and Figure 1. These findings highlight systematic correlations between the physicochemical properties of soil and bacterial community structures across hierarchical taxonomic scales, from phylum down to species. A consistent trend was the strong negative correlation between the soil moisture content and bacterial abundance at all taxonomic ranks. This finding indicates that relatively high moisture levels are associated with reduced bacterial diversity and abundance, potentially due to the limiting effects of excessive water on oxygen availability and microbial metabolism in the soil matrix. Additionally, pH measurements (pHw,1:2.5) demonstrated negative correlations across all taxonomic levels. The consistently low correlation coefficients, particularly at more resolved taxonomic levels, suggest that more acidic or alkaline soil conditions may adversely affect bacterial proliferation, thereby potentially influencing enzymatic activities and nutrient availability with bacterial abundance across all taxonomic ranks. In contrast, a strong positive correlation existed between the sand content and bacterial abundance, with the correlation strength increasing at lower taxonomic ranks. This finding implies that sandy soils, which typically offer better aeration and drainage, provide microhabitats conducive to bacterial colonization and diversity. Similarly, the silt content also exhibited positive correlations, although the magnitude of these relationships varied across taxonomic levels, indicating a complex interaction between soil texture components and microbial habitat suitability. Conversely, the clay content was significantly negatively correlated with bacterial abundance. The high negative coefficients suggest that soils with relatively high clay fractions might impede microbial activity due to reduced pore space and limited oxygen diffusion, which can constrain bacterial metabolism and growth (77, 78). Our study also revealed that nitrogen forms exhibit divergent patterns: ammonium (N-NH4+) is negatively correlated with bacterial abundance, whereas nitrate (N-NO3-) is positively correlated with bacterial abundance. These opposing trends may reflect differences in nitrogen availability and transformation within the soil, with nitrate potentially serving as an electron acceptor under anaerobic conditions and supporting certain bacterial groups (79, 80). Furthermore, exchangeable calcium (Ca2+) and available manganese (Mn) are negatively associated with bacterial abundance, although the strength of these relationships varies slightly across taxonomic ranks. These findings suggest that elevated levels of these cations may exert inhibitory effects on certain microbial populations, possibly through competitive interactions or alterations in soil pH (81). The result indicates the increased exchanged cations (N-NH4+, Ca2+, Mn) reveal the higher H+ abundance in soil colloids and subsequently more acidic soil, which is unfavorable to bacteria. Finally, the presence of phosphorous-solubilizing bacteria (PSB) is positively correlated with overall bacterial abundance, underscoring their potential role in mediating soil phosphorous availability and promoting nutrient cycling processes within longan orchard ecosystems.
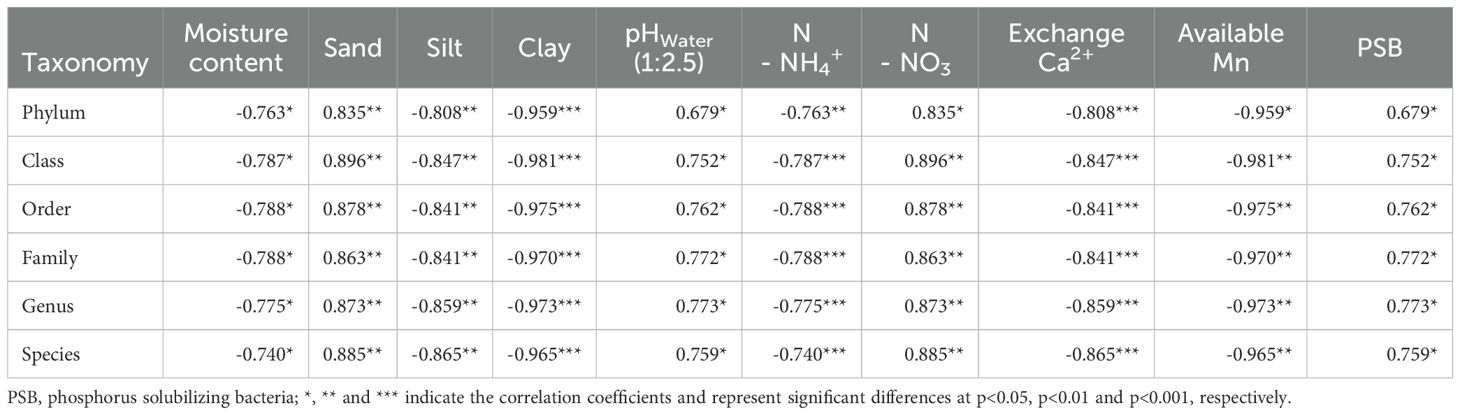
Table 2. Correlation between soil properties and soil bacterial taxonomy of different longan orchard soils in the Mekong River Delta of Vietnam.
The observed correlations between soil properties and soil bacterial taxonomy provide helpful knowledge into the environmental drivers shaping bacterial community composition in the soil of longan orchards in Vietnam’s Mekong River Delta. Soil chemical properties serve as critical indicators of microbial health (82). The consistent negative correlation between moisture content and bacterial abundance implies that higher soil moisture may restrict oxygen diffusion and limit bacterial growth (83). In contrast, the positive association with the sand content suggests that well-aerated, porous soils support greater bacterial populations (84). Although the negative correlation between clay content and bacterial abundance indicates potential physical constraints, some studies have reported that clay-rich soils can foster greater bacterial diversity than silt or sand fractions can (85, 86). Soil pH, measured as pHw (1:2.5), has emerged as a strong predictor of bacterial abundance, highlighting the pivotal role of pH in microbial community structuring (87, 88). Additionally, the relationships between nitrogen levels, exchangeable calcium, available manganese, and bacterial abundance highlight the influential role of soil nutrients on microbial dynamics. The positive correlation between the presence of PSB and bacterial abundance suggests a potential role of PSB in enhancing soil fertility and nutrient availability in longan orchard soils. These findings align with the work, which reported that the soil pH, total nitrogen, available nitrogen (NH4+ and NO3-), and clay content significantly affect the soil bacterial community (p < 0.05) (89).
3.2 Correlations between physical, chemical, and biological parameters in different longan orchard soils
3.2.1 Physical parameters and soil bacteria of different longan orchard soils
Supplementary Tables S4A, B present the correlations between the physical parameters and the soil bacterial phyla of the different longan orchards. There was a strong positive correlation between moisture content and Actinobacteriota (p<0.01). However, Acidobacteriota, Desulfobacterota, and Latescibacterota presented strong negative correlations with moisture content (p<0.01), whereas Verrucomicrobiota presented a very strong negative correlation (p<0.001). Among the bacterial phyla investigated, their correlations with the sand and clay contents differed. The abundances of Actinobacteriota, Firmicutes, and Crenarchaeota were strongly negatively correlated with the sand content and strongly positively correlated with the clay content. In contrast, Gemmatimonadota has a very strong positive correlation with sand, whereas Chloroflexi, Verrucomicrobiota, and the WPS-2 phylum have strong positive correlations with sand and negative correlations with clay content.
Soil moisture content is a crucial determinant of soil microbial biodiversity, as it influences oxygen diffusion and nutrient conductance within the soil environment. In longan orchards, bacterial growth and community structure are affected by varying soil compaction levels, creating aerated, saturated, and moist soil conditions as the orchard beds age. The relationship between soil moisture and soil particle size distribution explains the presence of different bacterial phyla. A study demonstrated that the abundance of Actinobacteriota was highest under saturated soil moisture conditions (90). Another study reported that the optimal abundance of actinobacteria occurred at 40% of the soil’s maximum moisture capacity, highlighting the complexity of the impact of moisture on the microbial community composition owing to inconsistencies in the soil moisture content and the immediate responses of microorganisms to these changes (91). However, in our study, a strong negative correlation between moisture and clay content significantly reduced the abundance of the Acidobacteriota, Desulfobacterota, Latescibacterota, and Verrucomicrobiota phyla. This suggests that coarse soil texture, which promotes aeration, enhances the growth and activity of these phyla, whereas clay content, which constrains aeration, has the opposite effect (92).
3.2.2 Chemical parameters and soil bacterial phyla of different longan orchard soils
The correlations between chemical parameters and soil bacterial phyla in different longan orchard soils are presented in Supplementary Tables S5A, B. Different phyla have different correlations with the chemical components that seem to influence the soil bacteria present. These findings suggest that orchard soil management, which alters soil chemical properties, could influence specific groups of bacteria or the whole bacterial community in the soil for longan growth. For pH and EC, the phyla Desulfobacterota, Latescibacterota and Acidobacteriota had a beneficial correlation (p<0.01), whereas Actinomicest had an inverse correlation. In addition, Proteobacteria, Firmicutes, RCP2-54, k:Bacteria_Uncl and Crenarchaeota were negatively correlated with the two pH and EC parameters, whereas the remaining groups were positively correlated. A study examining the impact of soil pH on the bacterial community composition in red soils grown with pomelo in Pinghe County, Fujian Province, China, demonstrated that the relative abundances of Proteobacteria, Actinobacteria, Crenarchaeota, and Firmicutes had a negative correlation with soil pH. Conversely, soil pH was shown a positive association with the relative abundances of Acidobacteria, Chloroflexi, Bacteroidetes, Planctomycetes, and Gemmatimonadetes (93). The other research showed that soil pH and electrical conductivity significantly influenced the overall microbial diversity in the soil. Elevating soil pH directly reduced the total Shannon diversity of the soil. Conversely, an increase in soil electrical conductivity directly enhanced the total Shannon diversity of the soil. Soil pH had a negative correlation with bacterial diversity in comparison to soil electrical conductivity in the structural equation models (94). A negative link exists between the relative abundance of Proteobacteria and soil pH (94), while a positive correlation is shown between Acidobacteria and soil pH (95).
For NH4+ and NO3- nitrogen, the strongest positive correlation was found for the phyla Actinobacteriota and Firmicutes, with high significance (p<0.001), whereas the strongest negative correlation was found for the phyla Verrucomicrobiot and Gemmatimonadota. Similarly, the phyla Proteobacteria and Crenarchaeota were positively correlated at p<0.01, whereas the phyla Myxococcota, Desulfobacterota, and Latescibacterota were negatively correlated at p<0.01. Notably, the phylum Acidobacteriota was negatively correlated with ammonium but positively correlated with nitrate. There is a lack of study about the available nitrogen content in relation to the relative abundances of various bacterial phyla; however, certain studies indicate that available nitrogen influences the bacterial community structure in soil (93, 96). Furthermore, research on a 20% reduction in nitrogen markedly enhanced the relative abundances of Proteobacteria, Bacteroidetes, Firmicutes, and Deinococcus-Thermus (97).
The available macronutrients P and K presented the strongest positive correlations with the phyla Nitrospirota and k:Bacteria_Uncl, followed by the phylum Crenarchaeota. Chloroflexi, RCP2–54 and WPS-2 were negatively correlated with available phosphorus and available potassium. The remaining nine phyla had positive correlations with these two elements. For exchangeable calcium, bacterial groups presented a more pronounced correlation, with five phyla having strong negative correlations with this factor, including the phyla Chloroflexi, Actinobacteriota, Firmicutes k:Bacteria_Uncl, and Crenarchaeota, with correlation coefficients ranging from (-0.947) to (-0.754). Six phyla had positive correlations with the exchangeable calcium factor, with correlation coefficients ranging from 0.608 - 0.842. Most bacterial groups were negatively correlated with silicon availability, with the phylum Nitrospirota showing the strongest correlation with this element (p<0.001). The phyla Chloroflexi, RCP2–54 and WPS-2 were positively correlated, with positive correlation coefficients ranging from 0.650-0.743. Significant positive correlations were detected between phyla and the availability of the minerals zinc (Zn) and manganese (Mn). Four phyla, Acidobacteriota, Verrucomicrobiot, Desulfobacterota, and Latescibacterota, were significantly positively correlated with Zn, and five phyla, Proteobacteria Firmicutes, k:Bacteria_Uncl, Nitrospirota, and Cenarchaeota, were strongly positively correlated with Mn.
The phylum Nitrospirota has been reported to increase significantly in the soil rhizosphere under low to moderate phosphorus levels, likely due to its role in nitrite oxidation and complete ammonia oxidation (comammox) processes (98). Similarly, a study conducted on red paddy soils in Vietnam demonstrated a positive correlation between acid phosphatase (AcP) activity and both total phosphorus (TP) and the relative abundance of several phyla including Chloroflexi, Proteobacteria, and Actinobacteria (99). These results align with our study, in which these same phyla showed positive associations with phosphorus availability. A study in Brazil found that six bacterial phyla were positively correlated with soil chemical properties such as pH, base saturation (V%), K+, Ca²+, Mg²+, cation exchange capacity (CEC), and total organic carbon (TOC). Among them, Bacteroidetes and Proteobacteria showed the strongest positive associations with pH, V%, Ca²+, Mg²+, CEC, and TOC. The phylum Chloroflexi was also significantly and positively correlated with all these variables except Ca²+. Particularly relevant to our findings, Nitrospira (now classified under Nitrospirota) and Candidatus Rokubacteria were the only phyla to exhibit positive and significant correlations with K+. In addition, Nitrospira was uniquely associated with phosphorus (P) and nitrogen content in pasture leaves (Ngrass), suggesting its role in nutrient cycling (100). These patterns further support the positive relationship observed in our study between available potassium and Nitrospirota.
3.2.3 Biological parameters and soil bacteria of different longan orchard soils
The correlations between the relative abundances of specific bacterial phyla and beneficial bacteria and the soil enzyme activities of longan orchards are presented in Supplementary Tables S6A–C. These relationships exhibit notable directionality and magnitude variations depending on the phylum. With respect to nitrogen-fixing bacteria, three phyla, Chloroflexi, RCP2-54, and WPS-2, demonstrated significant positive correlations. These findings suggest that these taxa may be functionally linked to nitrogen fixation processes, either through direct nitrogen-fixing capabilities or by fostering an environment conducive to these processes. In contrast, the remaining nine phyla exhibited significant negative correlations with nitrogen-fixing bacteria (p<0.01). This inverse relationship could indicate competitive interactions or different ecological niches where these groups suppress or are not favorably associated with nitrogen fixation. Four phyla, Proteobacteria, Actinobacteriota, Firmicutes, and Crenarchaeota, showed strong and positive correlations with phosphate solubilization, implying a role in increasing the availability of phosphorus to plants by solubilization mechanisms. In contrast, the other eleven phyla were negatively correlated with phosphorous-solubilizing bacteria (p<0.05), suggesting that they may either be less involved in phosphorous turnover or may be indirectly affected by competitive dynamics in the rhizosphere. Soil enzyme activities further delineated these functional associations. Chloroflexi, RCP2-54, and WPS-2 demonstrated robust and statistically significant positive correlations with key enzymes, including β-glucosidase, urease, phosphomonoester, and phytase. These enzymes are crucial for the decomposition of organic matter and the cycling of nutrients. The strong positive associations imply that these phyla may contribute directly to increased enzymatic activities, potentially facilitating carbon, nitrogen, and phosphorous cycling in longan orchard soils. In contrast, the remaining phyla either had negative correlations or weak positive correlations with these enzymatic activities, suggesting a lesser or inhibitory role in soil biochemical processes. Overall, these correlation patterns underscore the complexity of soil microbial community interactions, where specific phyla appear to be intimately linked to beneficial biochemical processes. A study indicated that the enzymatic activity of paddy soil was associated with both bacterial diversity and composition. The variations in soil enzyme activity were strongly associated with predominant optimum taxonomic units (OTUs), the majority of which were phylogenetically connected to enzyme producers from Planctomycetes, Proteobacteria, and Chloroflexi, exhibiting differing levels of 16S rRNA gene sequence similarity (101).
4 Conclusions
In conclusion, there was a decrease in the number of soil bacteria according to the soil cultivation time, revealing the following pattern: N15>N20>N30. Furthermore, the OTUs index demonstrated that their shared characteristics outnumbered the differences among them. The variety and abundance of the community of bacteria were high for N15, which was less impacted by the longan cultivation period, implying that cultivation time had an important effect on the microbiome. Acidobacteria, Proteobacteria, Chloroflexi, Actinobacteria, and Bacteriodota constituted the most common microbes in longan cultivation soils. The microbial community in longan growing soil was positively correlated with sand, pH, NO3- and phosphorous-solubilizing microorganisms, and negatively correlated with soil moisture, silt, clay, NH4+, exchange of calcium and available Mn. Physical, chemical or biological factors in the soil have complex relationships (positive or negative correlations) with different bacterial phyla. The results of this study are important for managing longan orchard soils over time to retain and increase tree productivity. Understanding the relationships among microbial communities and soil physical, chemical and biological properties; and recognizing that they change as the orchard ages is critical for improving orchard management. This suggests that the depletion of some soil nutrients over time as the orchard ages could affect microbial community species and their richness and abundance. Routine soil testing and management methods, including the incorporation of organic materials, micronutrients, cultivation of cover crops, and earthworms, can stimulate and supplement beneficial bacteria such as the nitrogen-fixing bacteria, phosphorous-solubilizing microbes, and potassium-dissolving bacteria, as well as the nutrient-related activity of enzymes. Future research should explore the relationships of specific microbial communities in longan orchard soils and these types of management practices to understand their impacts on tree health, fruit yields, and disease pressure.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
NN: Writing – original draft, Investigation, Funding acquisition, Project administration, Data curation, Conceptualization, Supervision, Methodology, Writing – review & editing. KD: Visualization, Validation, Data curation, Writing – review & editing. KH: Writing – review & editing, Validation, Visualization, Data curation, Formal analysis. LX: Software, Writing – original draft, Validation, Formal analysis, Data curation, Visualization. LM: Resources, Writing – review & editing, Conceptualization, Writing – original draft, Validation, Data curation, Visualization. HT: Investigation, Data curation, Resources, Writing – original draft, Visualization, Software, Validation. JR: Conceptualization, Validation, Writing – original draft, Visualization, Resources, Data curation, Methodology. JS: Visualization, Data curation, Software, Validation, Resources, Investigation, Writing – review & editing. HL: Visualization, Resources, Validation, Writing – review & editing, Data curation, Methodology, Software. TN: Software, Visualization, Writing – review & editing, Validation, Data curation. NP: Supervision, Investigation, Software, Funding acquisition, Writing – review & editing, Project administration, Writing – original draft, Validation, Data curation, Methodology, Resources. CT: Writing – original draft, Software, Validation, Supervision, Visualization, Data curation, Resources, Writing – review & editing. DT: Resources, Software, Writing – original draft, Writing – review & editing, Visualization, Methodology, Investigation. NT: Visualization, Resources, Software, Validation, Writing – review & editing, Methodology, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Ministry of Education and Training, Vietnam, Grant Number: B2023-TCT-14.
Acknowledgments
We are very grateful to all the colleagues who were responsible for the sample collection and data analyses. We would also like to extend our gratitude to the reviewers for their constructive feedback.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor HT declared a past co-authorship with the author(s) LM and HT.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsoil.2025.1610343/full#supplementary-material
References
1. General Statistics Office of Vietnam. Statistical yearbook of Vietnam 2020. Vietnam: Statistical Publishing House (2021).
2. Truc NTT and Thuc LV. Impacts of adopting specialized agricultural programs relying on “good practice”–Empirical evidence from fruit growers in Vietnam. Open Agric. (2022) 7:39–49. doi: 10.1515/opag-2022-0069
3. Nguyen NK, Nguyen PM, Chau ATT, Do LT, Nguyen THT, Tran DHV, et al. Long-term changes in soil biological activity and other properties of raised beds in Longan orchards. PeerJ. (2024) 12:e18396. doi: 10.7717/peerj.18396
4. Statistics Office of Vinh Long. Vinh Long province statistical yearbook 2022. Statistical Publishing House (2022).
5. Hau TV and Hieu TS. Induction of flowering in longan in the Mekong delta, Vietnam. VI Int Symposium Lychee Longan Other Sapindaceae. (2019), 1293. doi: 10.17660/ActaHortic.2020.1293.27
6. Van LD and Ngoc HN. Role of organic fertilizer application in improving soil chemistry characteristics and fruit yield of Nam Roi pomelo in Hau Giang province. Can Tho Univ J Science. (2020) 56:82. doi: 10.22144/ctu.jsi.2020.07
7. Agegnehu G, Amede T, Erkossa T, Yirga C, Henry C, Tyler R, et al. Extent and management of acid soils for sustainable crop production system in the tropical agroecosystems: a review. Acta Agriculturae Scandinavica Section B—Soil Plant Science. (2021) 71:852–69. doi: 10.1080/09064710.2021.1954239
8. Zhao Z, Zheng W, Ma Y, Wang X, Li Z, Zhai B, et al. Responses of soil water, nitrate and yield of apple orchard to integrated soil management in Loess Plateau, China. Agric Water Management. (2020) 240:106325. doi: 10.1016/j.agwat.2020.106325
9. Dang LV, Ngoc NP, and Hung NN. Soil quality and pomelo productivity as affected by chicken manure and cow dung. Sci World J. (2021) 2021:6289695. doi: 10.1155/2021/6289695
10. Dang LV, Ngoc NP, and Hung NN. Effects of biochar, lime, and compost applications on soil physicochemical properties and yield of pomelo (Citrus grandis Osbeck) in alluvial soil of the Mekong Delta. Appl Environ Soil Science. (2022) 2022(1):5747699. doi: 10.1155/2022/5747699
11. Chen S, Lin B, Li Y, and Zhou S. Spatial and temporal changes of soil properties and soil fertility evaluation in a large grain-production area of subtropical plain, China. Geoderma. (2020) 357:113937. doi: 10.1016/j.geoderma.2019.113937
12. Maleki S, Karim A, Zeraatpisheh M, Poozeshi R, and Feizi H. Long-term cultivation effects on soil properties variations in different landforms in an arid region of eastern Iran. Catena. (2021) 206:105465. doi: 10.1016/j.catena.2021.105465
13. Dang LV and Hung NN. Effects of the age of raised beds on the physicochemical characteristics of fruit orchardsoil in the Vietnamese Mekong Delta. PeerJ. (2023) 11:e16178. doi: 10.7717/peerj.16178
14. Zhao Y, Zhang F, Yang L, Wang D, and Wang W. Response of soil bacterial community structure to different reclamation years of abandoned salinized farmland in arid China. Arch Microbiol. (2019) 201:1219–32. doi: 10.1007/s00203-019-01689-x
15. Nielsen UN, Wall DH, and Six J. Soil biodiversity and the environment. Annu Rev Environ resources. (2015) 40:63–90. doi: 10.1146/annurev-environ-102014-021257
16. Lehmann J, Bossio DA, Kögel-Knabner I, and Rillig MC. The concept and future prospects of soil health. Nat Rev Earth Environment. (2020) 1:544–53. doi: 10.1038/s43017-020-0080-8
17. Han XM, Wang RQ, Wang MC, Zhou J, and Guo WH. Effects of vegetation type on soil microbial community structure and catabolic diversity assessed by polyphasic methods in North China. J Environ Sci. (2007) 19:1228–34. doi: 10.1016/S1001-0742(07)60200-9
18. Bi J, He D, Sha Y, and Huang Z. Functional diversity of soil microbial community under different types of vegetation in the desert grassland. Agric Res Arid Areas. (2009) 5:149–55. doi: 10.1016/S0038-0717(01)00052-9
19. Marschner P, Yang CH, Lieberei R, and Crowley DE. Soil and plant specific effects on bacterial community composition in the rhizosphere. Soil Biol Biochem. (2001) 33:1437–45. doi: 10.1016/S0038-0717(01)00052-9
20. Rinnan R, Stark S, and Tolvanen A. Responses of vegetation and soil microbial communities to warming and simulated herbivory in a subarctic heath. J Ecology. (2009) 97:788–800. doi: 10.1111/j.1365-2745.2009.01506.x
21. Yan J, Hang X, and Wang S. Variation of soil microoganism under different vegetation coverages and fertilization systems in black soil. Chin J Soil Sci. (2009) 40:240–4.
22. Zhang Y, Li T, Wu H, Bei S, Zhang J, and Li X. Effect of different fertilization practices on soil microbial community in a wheat–maize rotation system. Sustainability. (2019) 11(15):4088. doi: 10.3390/su11154088
23. Li JH, Jiao SM, Gao RQ, and Bardgett RD. Differential effects of legume species on the recovery of soil microbial communities, and carbon and nitrogen contents, in abandoned fields of the Loess Plateau, China. Environ management. (2012) 50:1193–203. doi: 10.1007/s00267-012-9958-7
24. Li Y, Liu X, Yin Z, Chen H, Cai X, Xie Y, et al. Changes in soil microbial communities from exposed rocks to arboreal rhizosphere during vegetation succession in a karst mountainous ecosystem. J Plant Interactions. (2021) 16:652493. doi: 10.3389/fpls.2021.652493
25. Ma W, Yang Z, Hou S, Ma Q, Liang L, Wang G, et al. Effects of living cover on the soil microbial communities and ecosystem functions of hazelnut orchards. Front Plant Science. (2021) 12:652493. doi: 10.3389/fpls.2021.652493
26. Liddle K, McGonigle T, and Koiter A. Microbe biomass in relation to organic carbon and clay in soil. Soil Systems. (2020) 4:41. doi: 10.3390/soilsystems4030041
27. He C, Wang R, Ding W, and Li Y. Effects of cultivation soils and ages on microbiome in the rhizosphere soil of Panax ginseng. Appl Soil Ecology. (2022) 174:104397. doi: 10.1016/j.apsoil.2022.104397
28. Nghia NK, Robatjazi J, Vy VDT, Tecimen HB, Lasar HGW, Lesueur D, et al. Effect of organic farming practices on soil health improvement of coconut farms. Environ Technol Innovation. (2025) 38:104067. doi: 10.1016/j.eti.2025.104067
29. Santolamazza-Carbone S, Iglesias-Bernabé L, Sinde-Stompel E, and Gallego PP. Soil microbiota impact on Boletus edulis mycelium in chestnut orchards of different ages. Appl Soil Ecology. (2023) 185:104790. doi: 10.1016/j.apsoil.2022.104790
30. Xie B, Chen Y, Cheng C, Ma R, Zhao D, Li Z, et al. Long-term soil management practices influence the rhizosphere microbial community structure and bacterial function of hilly apple orchard soil. Appl Soil Ecology. (2022) 180:104627. doi: 10.1016/j.apsoil.2022.104627
31. Baca-Patiño B, González-Rodríguez A, García-Oliva F, García A, Lara-De La Cruz I, Garibay-Orijel R, et al. Land-use change from native forest to avocado orchards: Effects on soil nutrient transformation and microbial communities. Appl Soil Ecology. (2025) 205:105748. doi: 10.1016/j.apsoil.2024.105748
32. Grassi A, Pagliarani I, Granata E, Avio L, Cristani C, Mascagni F, et al. Floor vegetation management affects the diversity and composition of mycorrhizal symbionts and soil microbial communities in a Mediterranean olive orchard. Pedosphere. (2025) 01:008. doi: 10.1016/j.pedsph.2025.01.008
33. Zawadzka K, Oszust K, Pylak M, Panek J, Gryta A, and Frąc M. Beneath the apple trees-Exploring soil microbial properties under Malus domestica concerning various land management practices. Appl Soil Ecology. (2024) 203:105642. doi: 10.1016/j.apsoil.2024.105642
34. Zhao J, Yuan X, Liu Z, Shi H, Zhai B, and Zhu Y. Divergent responses of soil physicochemical properties in 6-m profiles to long-term overfertilization in rainfed apple orchards on China’s Loess Plateau. Agriculture Ecosyst Environment. (2024) 361:108817. doi: 10.1016/j.agee.2023.108817
35. Chen M, Feng S, Wang J, Gao M, Liu M, Wang K, et al. The stability of soil aggregates in sweet cherry (Prunus avium L.) orchards of different ages and varieties. Heliyon. (2025) 11(3):e42189. doi: 10.1016/j.heliyon.2025.e42189
36. Liu Z, Ma X, He N, Zhang J, Wu J, and Liu C. Shifts in microbial communities and networks are correlated with the soil ionome in a kiwifruit orchard under different fertilization regimes. Appl Soil Ecology. (2020) 149:103517. doi: 10.1016/j.apsoil.2020.103517
37. Dixit M, Ghoshal D, Meena AL, Ghasal PC, Rai A, Choudhary J, et al. Changes in soil microbial diversity under present land degradation scenario. Total Environ Adv. (2024) 10:200104. doi: 10.1016/j.teadva.2024.200104
38. Shannon CE. A mathematical theory of communication. Bell system Tech J. (1948) 27:379–423. doi: 10.1002/j.1538-7305.1948.tb01338.x
39. Whittaker RH. Evolution and measurement of species diversity. Taxon. (1972) 21:213–51. doi: 10.2307/1218190
40. Chao A. Nonparametric estimation of the number of classes in a population. Scandinavian J Stat. (1984) 11(4):265–70. https://www.jstor.org/stable/4615964
41. Hillebrand H, Blasius B, Borer ET, Chase JM, Downing JA, Eriksson BK, et al. Biodiversity change is uncoupled from species richness trends: Consequences for conservation and monitoring. J Appl ecology. (2018) 55:169–84. doi: 10.1111/1365-2664.12959
42. Jiao S, Chen W, Wang J, Du N, Li Q, and Wei G. Soil microbiomes with distinct assemblies through vertical soil profiles drive the cycling of multiple nutrients in reforested ecosystems. Microbiome. (2018) 6:1–13. doi: 10.1186/s40168-018-0526-0
43. Fierer N and Jackson RB. The diversity and biogeography of soil bacterial communities. Proc Natl Acad Sci. (2006) 103:626–31. doi: 10.1073/pnas.0507535103
44. Nannipieri P, Ascher J, Ceccherini MT, Landi L, Pietramellara G, and Renella G. Microbial diversity and soil functions. Eur J Soil Sci. (2003) 54:655–70. doi: 10.1046/j.1351-0754.2003.0556.x
45. Rousk J, Bååth E, Brookes PC, Lauber CL, Lozupone C, Caporaso JG, et al. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. (2010) 4:1340–51. doi: 10.1038/ismej.2010.58
46. Pietri JA and Brookes PC. Relationships between soil pH and microbial properties in a UK arable soil. Soil Biol Biochem. (2008) 40:1856–61. doi: 10.1016/j.soilbio.2008.03.020
47. Khan MS, Zaidi A, and Wani PA. Role of phosphate-solubilizing microorganisms in sustainable agriculture—a review. Agron Sustain Dev. (2007) 27:29–43. doi: 10.1051/agro:2006011
48. Janssen PH. Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl Environ Microbiol. (2006) 72:1719–28. doi: 10.1128/AEM.72.3.1719-1728.2006
49. Arjun JK and Harikrishnan K. Metagenomic analysis of bacterial diversity in the rice rhizosphere soil microbiome. Biotechnol Bioinf. Bioeng. (2011) 1:361–7. http://rgcb.sciencecentral.in/id/eprint/584
50. Chowdhury SP, Schmid M, Hartmann A, and Tripathi AK. Diversity of 16S-rRNA and nifH genes derived from rhizosphere soil and roots of an endemic drought tolerant grass, Lasiurus sindicus. Eur J Soil Biol. (2009) 45:114–22. doi: 10.1016/j.ejsobi.2008.06.005
51. Dhondge HV, Barvkar VT, Paul D, Dastager SG, Pable AA, and Nadaf AB. Exploring the core microbiota in scented rice (Oryza sativa L.) rhizosphere through metagenomics approach. Microbiological Res. (2022) 263:127157. doi: 10.1016/j.micres.2022.127157
52. Pratibha G, Manjunath M, Raju BMK, Srinivas I, Rao KV, Shanker AK, et al. Soil bacterial community structure and functioning in a long-term conservation agriculture experiment under semi-arid rainfed production system. Front Microbiol. (2023) 14:1102682. doi: 10.3389/fmicb.2023.1102682
53. Kalam S, Basu A, Ahmad I, Sayyed RZ, El-Enshasy HA, Dailin DJ, et al. Recent understanding of soil acidobacteria and their ecological significance: a critical review. Front Microbiol. (2020) . 11:580024. doi: 10.3389/fmicb.2020.580024
54. Kielak AM, Barreto CC, Kowalchuk GA, Van Veen JA, and Kuramae EE. The ecology of Acidobacteria: moving beyond genes and genomes. Front Microbiol. (2016) 7:744. doi: 10.3389/fmicb.2016.00744
55. Schmitz RA, Peeters SH, Versantvoort W, Picone N, Pol A, Jetten MS, et al. Verrucomicrobial methanotrophs: ecophysiology of metabolically versatile acidophiles. FEMS Microbiol Rev. (2021) 45:fuab007. doi: 10.1093/femsre/fuab007
56. Rakitin AL, Kulichevskaya IS, Beletsky AV, Mardanov AV, Dedysh SN, and Ravin NV. Verrucomicrobia of the family chthoniobacteraceae participate in xylan degradation in boreal peat soils. Microorganisms. (2024) 12:2271. doi: 10.3390/microorganisms12112271
57. Lueders T, Kindler R, Miltner A, Friedrich MW, and Kaestner M. Identification of bacterial micropredators distinctively active in a soil microbial food web. Appl Environ Microbiol. (2006) 72:5342–8. doi: 10.1128/AEM.00400-06
58. Zhang L and Lueders T. Micropredator niche differentiation between bulk soil and rhizosphere of an agricultural soil depends on bacterial prey. FEMS Microbiol Ecology. (2017) 93:fix103. doi: 10.1093/femsec/fix103
59. Gross V, Reinhard A, Petters S, Pichler M, and Urich T. Adding complexity to soil food webs: Myxobacteria have broad predation spectra with bacteria, yeasts and filamentous fungi in vitro. Eur J Soil Biol. (2023) 117:103508. doi: 10.1016/j.ejsobi.2023.103508
60. Shan S, Wei Z, Cheng W, Du D, Zheng D, and Ma G. Biofertilizer based on halotolerant microorganisms promotes the growth of rice plants and alleviates the effects of saline stress. Front Microbiol. (2023) 14:1165631. doi: 10.3389/fmicb.2023.1165631
61. Rahimlou S, Bahram M, and Tedersoo L. Phylogenomics reveals the evolution of root nodulating alpha-and beta-Proteobacteria (rhizobia). Microbiological Res. (2021) 250:126788. doi: 10.1016/j.micres.2021.126788
62. Mitra D, Mondal R, Khoshru B, Senapati A, Radha TK, Mahakur B, et al. Actinobacteria-enhanced plant growth, nutrient acquisition, and crop protection: Advances in soil, plant, and microbial multifactorial interactions. Pedosphere. (2022) 32:149–70. doi: 10.1016/S1002-0160(21)60042-5
63. Boukhatem ZF, Merabet C, and Tsaki H. Plant growth promoting actinobacteria, the most promising candidates as bioinoculants? Front Agronomy. (2022) 4:849911. doi: 10.3389/fagro.2022.849911
64. Chau TAT. Syntrophic interaction in organochlorine bioremediation: A review. CTU J Innovation Sustain Dev. (2018) 10):70–80. doi: 10.22144/ctu.jen.2018.040
65. Kim SJ, Ahn JH, Weon HY, Hong SB, Seok SJ, Kim JS, et al. Chujaibacter soli gen. nov., sp. nov., isolated from soil. J Microbiol. (2015) 53:592–7. doi: 10.1007/s12275-015-5136-y
66. She S, Niu J, Zhang C, Xiao Y, Chen W, Dai L, et al. Significant relationship between soil bacterial community structure and incidence of bacterial wilt disease under continuous cropping system. Arch Microbiol. (2017) 199:267–75. doi: 10.1007/s00203-016-1301-x
67. Wang S, Cheng J, Li T, and Liao Y. Response of soil fungal communities to continuous cropping of flue-cured tobacco. Sci Rep. (2020) 10:19911. doi: 10.1038/s41598-020-77044-8
68. Yang Y, Ahmed W, Ye C, Yang L, Wu L, Dai Z, et al. Exploring the effect of different application rates of biochar on the accumulation of nutrients and growth of flue-cured tobacco (Nicotiana tabacum). Front Plant Science. (2024) 15:1225031. doi: 10.3389/fpls.2024.1225031
69. Meola M, Lazzaro A, and Zeyer J. Diversity, resistance and resilience of the bacterial communities at two alpine glacier forefields after a reciprocal soil transplantation. Environ Microbiol. (2014) 16:1918–34. doi: 10.1111/1462-2920.12435
70. Wang D, Ning Q, Deng Z, Zhang M, and You J. Role of environmental stresses in elevating resistance mutations in bacteria: Phenomena and mechanisms. Environ Pollution. (2022) 307:119603. doi: 10.1016/j.envpol.2022.119603
71. Pierantoni DC, Conti A, Corte L, Benincasa P, Cardinali G, and Guiducci M. Soil microbiota resilience in a two-decade long-term experiment comparing an organic and a conventional cropping system. Appl Soil Ecol. (2024) 201:105481. doi: 10.1016/j.apsoil.2024.105481
72. Singavarapu B, Du J, Beugnon R, Cesarz S, Eisenhauer N, Xue K, et al. Functional potential of soil microbial communities and their subcommunities varies with tree mycorrhizal type and tree diversity. Microbiol spectrum. (2023) 11:e04578–22. doi: 10.1128/spectrum.04578-22
73. Guo Y, Hou L, Zhang Z, Zhang J, Cheng J, Wei G, et al. Soil microbial diversity during 30 years of grassland restoration on the Loess Plateau, China: Tight linkages with plant diversity. Land Degradation Dev. (2019) 30:1172–82. doi: 10.1002/ldr.3300
74. Liu Y, Chen X, Liu J, Liu T, Cheng J, Wei G, et al. Temporal and spatial succession and dynamics of soil fungal communities in restored grassland on the Loess Plateau in China. Land Degradation Dev. (2019) 30:1273–87. doi: 10.1002/ldr.3289
75. Philippot L, Chenu C, Kappler A, Rillig MC, and Fierer N. The interplay between microbial communities and soil properties. Nat Rev Microbiol. (2024) 22:226–39. doi: 10.1038/s41579-023-00980-5
76. Xue R, Wang C, Zhao L, Sun B, and Wang B. Agricultural intensification weakens the soil health index and stability of microbial networks. Agriculture Ecosyst Environ. (2022) 339:108118. doi: 10.1016/j.agee.2022.108118
77. Fomina M and Skorochod I. Microbial interaction with clay minerals and its environmental and biotechnological implications. Minerals. (2020) 10:861. doi: 10.3390/min10100861
78. Wilber SA and Newman DK. The contrasting roles of nitric oxide drive microbial community organization as a function of oxygen presence. Curr Biol. (2022) 32:5221–34. doi: 10.1016/j.cub.2022.10.008
79. Xu M, Zhang Q, Xia C, Zhong Y, Sun G, Guo J, et al. Elevated nitrate enriches microbial functional genes for potential bioremediation of complexly contaminated sediments. ISME J. (2014) 8:1932–44. doi: 10.1038/ismej.2014.229
80. Zhang C, You Z, Li S, Zhang C, Zhao Z, and Zhou D. NO3– as an electron acceptor elevates antibiotic resistance gene and human bacterial pathogen risks in managed aquifer recharge (MAR): A comparison with O2. Environ Res. (2024) 248:118277. doi: 10.1016/j.envres.2024.118277
81. Macedo WV, Sakamoto IK, Azevedo EB, and Damianovic MHR. The effect of cations (Na+, Mg2+, and Ca2+) on the activity and structure of nitrifying and denitrifying bacterial communities. Sci Total Environment. (2019) 679:279–87. doi: 10.1016/j.scitotenv.2019.04.397
82. Fierer N, Bradford MA, and Jackson RB. Toward an ecological classification of soil bacteria. Ecology. (2007) 88:1354–64. doi: 10.1890/05-1839
83. Stark JM and Firestone MK. Mechanisms for soil moisture effects on activity of nitrifying bacteria. Appl Environ Microbiol. (1995) 61:218–21. doi: 10.1128/aem.61.1.218-221.1995
84. Carson JK, Gonzalez-Quinones V, Murphy DV, Hinz C, Shaw JA, and Gleeson DB. Low pore connectivity increases bacterial diversity in soil. Appl Environ Microbiol. (2010) 76:3936–42. doi: 10.1128/AEM.03085-09
85. Sessitsch A, Weilharter A, Gerzabek MH, Kirchmann H, and Kandeler E. Microbial population structures in soil particle size fractions of a long-term fertilizer field experiment. Appl Environ Microbiol. (2001) 67:4215–24. doi: 10.1128/AEM.67.9.4215-4224.2001
86. Mhete M, Eze PN, Rahube TO, and Akinyemi FO. Soil properties influence bacterial abundance and diversity under different land-use regimes in semi-arid environments. Sci African. (2020) 7:e00246. doi: 10.1016/j.sciaf.2019.e00246
87. Maestre FT, Delgado-Baquerizo M, Jeffries TC, Eldridge DJ, Ochoa V, Gozalo B, et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc Natl Acad Sci. (2015) 112:15684–9. doi: 10.1073/pnas.1516684112
88. Zheng W, Zhao Z, Lv F, Wang R, Wang Z, Zhao Z, et al. Assembly of abundant and rare bacterial and fungal sub-communities in different soil aggregate sizes in an apple orchard treated with cover crop and fertilizer. Soil Biol Biochem. (2021) 156:108222. doi: 10.1016/j.soilbio.2021.108222
89. Yang L, Han D, Jin D, Zhang J, Shan Y, Wan M, et al. Soil physiochemical properties and bacterial community changes under long-term polycyclic aromatic hydrocarbon stress in situ steel plant soils. Chemosphere. (2023) 334:138926. doi: 10.1016/j.chemosphere.2023.138926
90. Ansari J, Bardhan S, Eivazi F, Anderson SH, and Mendis SS. Bacterial community diversity for three selected land use systems as affected by soil moisture regime. Appl Soil Ecology. (2023) 192:105100. doi: 10.1016/j.apsoil.2023.105100
91. Borowik A and Wyszkowska J. Soil moisture as a factor affecting the microbiological and biochemical activity of soil. Plant Soil Environ. (2016) 62:2016. doi: 10.17221/158/2016-PSE
92. Chau JF, Bagtzoglou AC, and Willig MR. The effect of soil texture on richness and diversity of bacterial communities. Environ Forensics. (2011) 12:333–41. doi: 10.1080/15275922.2011.622348
93. Muneer MA, Hou W, Li J, Huang X, ur Rehman Kayani M, Cai Y, et al. Soil pH: a key edaphic factor regulating distribution and functions of bacterial community along vertical soil profiles in red soil of pomelo orchard. BMC Microbiol. (2022) . 22:38. doi: 10.1186/s12866-022-02452-x
94. Yang C, Wang X, Miao F, Li Z, Tang W, and Sun J. Assessing the effect of soil salinization on soil microbial respiration and diversities under incubation conditions. Appl Soil Ecol. (2020) 155:103671. doi: 10.1016/j.apsoil.2020.103671
95. Wang Q, Wang C, Yu WW, Turak A, Chen D, Huang Y, et al. Effects of nitrogen and phosphorus inputs on soil bacterial abundance, diversity, and community composition in chinese fir plantations. Front Microbiol. (2018) 9:1543. doi: 10.3389/fmicb.2018.01543
96. Tang Y, Liu J, Bao J, Chu G, and Peng F. Soil type influences rhizosphere bacterial community assemblies of pecan plantations, a case study of eastern China. Forests. (2022) . 13:363. doi: 10.3390/f13030363
97. Xie W, Shen X, Li W, Yan L, Li J, Ding B, et al. The effects of nitrogen reduction and sheep manure incorporation on the soil characteristics and microbial community of korla fragrant pear orchards. Agronomy. (2025) . 15:545. doi: 10.3390/agronomy15030545
98. Zhou B, Cheng S, Peng S, Li W, Li C, Wang Q, et al. Response of bacterial community structure to different phosphorus additions in a tobacco-growing soil. Front Plant Science. (2024) . 15:1344733. doi: 10.3389/fpls.2024.1344733
99. Hegyi A, Nguyen TBK, and Posta K. Metagenomic analysis of bacterial communities in agricultural soils from Vietnam with special attention to phosphate solubilizing bacteria. Microorganisms. (2021) 9:1796. doi: 10.3390/microorganisms9091796
100. Paes da Costa D, das Graças Espíndola da Silva T, Sérgio Ferreira Araujo A, Prudêncio de Araujo Pereira A, William Mendes L, dos Santos Borges W, et al. Soil fertility impact on recruitment and diversity of the soil microbiome in sub-humid tropical pastures in northeastern Brazil. Sci Rep. (2024) 14:3919. doi: 10.1038/s41598-024-54221-7
Keywords: alpha diversity, longan, metagenome, soil bacteria, soil microbial diversity
Citation: Nghia NK, Dalma KE, Haydee KM, Xa LT, Morton LW, Tecimen HB, Robatjazi J, Sekar J, Lasar HGW, Nguyen TT, Phuong NM, Thy CTA, Luan DT and Oanh NTK (2025) Bacterial diversity in longan orchard alluvial soil is influenced by cultivation time and soil properties. Front. Soil Sci. 5:1610343. doi: 10.3389/fsoil.2025.1610343
Received: 14 April 2025; Accepted: 23 June 2025;
Published: 14 July 2025.
Edited by:
Huu Tuan Tran, Văn Lang University, VietnamReviewed by:
Jialin Hu, Texas A and M University, United StatesTung Tra, Nguyen Tat Thanh University, Vietnam
Copyright © 2025 Nghia, Dalma, Haydee, Xa, Morton, Tecimen, Robatjazi, Sekar, Lasar, Nguyen, Phuong, Thy, Luan and Oanh. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nguyen Khoi Nghia, bmtuZ2hpYUBjdHUuZWR1LnZu