 Dominik Appel1†
Dominik Appel1† Regina Hummel1†Martin Weidemeier1
Regina Hummel1†Martin Weidemeier1 Kristina Endres2,3Christina Gölz1
Kristina Endres2,3Christina Gölz1 Michael K. E. Schäfer1,2,4*
Michael K. E. Schäfer1,2,4*- 1Department of Anesthesiology, University Medical Center, Johannes Gutenberg-University, Mainz, Germany
- 2Focus Program Translational Neurosciences (FTN) of the Johannes Gutenberg-University, Mainz, Germany
- 3Department of Psychiatry and Psychotherapy, University Medical Center, Johannes Gutenberg-University, Mainz, Germany
- 4Research Center for Immunotherapy (FZI), Johannes Gutenberg-University, Mainz, Germany
The α-secretase A disintegrin and metalloprotease 10 (ADAM10) regulates various physiological and pathophysiological processes. Despite its broad functional implications during development, plasticity, and disease, no pharmacological approaches to inhibit ADAM10 in acute brain injury have been reported. Here, we examined the effects of the ADAM10 inhibitor GI254023X on the neurological and histopathological outcome after experimental traumatic brain injury (TBI). C57BL/6N mice were subjected to the controlled cortical impact (CCI) model of TBI or sham procedure and received GI254023X or vehicle during the acute phase of injury (n = 40, 100 mg/kg, 25% DMSO, 0.1 M Na2CO3, intraperitoneal, 30 min and 24 h after TBI). GI254023X treatment did not improve neurological deficits from 1 to 7 days post-injury (dpi) but animals treated with GI254023X exhibited smaller brain lesions compared to vehicle treatment. Determination of brain mRNA expression by quantitative PCR showed that TBI-induced up-regulation of Adam10 and Adam17 was not influenced by GI254023X but the up-regulation of the matrix metalloproteinase genes Mmp2 and Mmp9 was attenuated. GI254023X treatment further increased the T cell marker Cd247 but did not affect blood brain barrier integrity, as assessed by Occludin mRNA expression and IgG brain extravasation. However, in agreement with neuroprotective effects of ADAM10 inhibition, GI254023X treatment attenuated axonal injury, as indicated by decreased generation of spectrin breakdown products (SBDPs) and decreased immunostaining using anti-non-phosphorylated neurofilament (SMI-32). Interestingly, reduced axonal injury in GI254023X-treated animals coincided with subtle mRNA dysregulation in the glutamate receptor subunit genes Gria1 and Grin2b. Quantitative PCR also revealed that GI254023X mitigated up-regulation of the pro-inflammatory markers Il6, Tnfa, and Lcn2 but not the up-regulation of the pan-microglia marker Aif1, the M2 microglia marker Arg1 and the reactive astrocyte marker Gfap. Taken together, the ADAM10 inhibitor GI254023X attenuates brain tissue loss, axonal injury and pro-inflammatory gene expression in the CCI model of TBI. These results suggest that ADAM10 may represent a therapeutic target in the acute phase of TBI.
Introduction
Traumatic brain injury (TBI) is a leading cause of death and disability and 69 million individuals worldwide are estimated to sustain a TBI each year (Dewan et al., 2018). The initial primary injury leads to irreversible tissue damage and triggers a plethora of secondary processes, such as defective cerebral perfusion and autoregulation, breakdown of the BBB, early formation of brain edema, mitochondrial dysfunction, oxidative stress, and other processes cumulating in the induction of neuronal cell death (Werner and Engelhard, 2007; Schäfer et al., 2014; Abdul-Muneer et al., 2015; Akamatsu and Hanafy, 2020). Subsequently, the activation of brain resident glia and infiltration of peripheral immune cells toward injury sites generates a highly inflammatory environment and oxidative stress, which is unfavorable for neuronal survival (Schäfer and Tegeder, 2018; Morganti-Kossmann et al., 2019).
Unfortunately, the pathophysiology of TBI is still incompletely understood and decades of animal research providing pharmacological targets for early therapeutic interventions have not been yet translated to the clinic. Given the complexity of TBI, the targeting of multiple secondary injury pathways has been proposed to better support neuronal survival and brain repair than targeting single factors (Loane and Faden, 2010; Hummel et al., 2020). However, also the targeting of single factors can have profound implications when they are at the forefront of regulatory processes such as enzymes with a variety of substrates executing different biological effects.
One such potential target is the metalloproteinase ADAM10, which can cleave almost 100 different substrates, including Notch family members, amyloid precursor protein (APP), cell adhesion molecules such as cadherin and members of the L1 family of cell adhesion molecules, the IL-6 cytokine receptor and membrane-bound chemokines (Wetzel et al., 2017; Hsia et al., 2019). ADAM10 has been best characterized in the context of early neural development (Jorissen et al., 2010), synapse function (Prox et al., 2013) and the non-amyloid pathway which prevents accumulation of neurotoxic Aβ peptides—hallmarks of Alzheimer’s disease (AD)—and generates potentially neuroprotective APP α-fragments (Thornton et al., 2006; Claasen et al., 2009; Gralle et al., 2009). Furthermore, the proteolytic activity of ADAM10 plays an important role in immunity and several inflammatory pathologies (Dreymueller and Ludwig, 2017; Lambrecht et al., 2018) and its dysregulation is linked to various types of cancer such as breast cancer (Tsang et al., 2018), colon cancer (Gavert et al., 2007), and malignant pleural mesothelioma (Sépult et al., 2019).
In contrast, few studies have investigated the role of ADAM10 in acute brain injury. Using unilateral entorhinal cortex lesion (ECL) in rodents, ADAM10 mRNA and protein expression up-regulation was observed in the denervated dentate gyrus, presumably in astrocytes (Warren et al., 2012; Del Turco et al., 2014). ADAM10 increased within 7 days post-injury—during reactive astrogliosis, axonal sprouting, and reactive synaptogenesis—and evidence was provided that ADAM10-mediated cleavage of N-cadherin prevents synaptic reorganization and thereby interferes with functional recovery (Warren et al., 2012). Another study reported that suppression of ADAM10 expression by miR-144 increased Aβ peptide accumulation and aggravated cognitive impairments within 1 week following TBI (Sun et al., 2017). Along this line, ADAM10 up-regulation has been implicated in decreased Aβ production and synapse rescue mediated by the PKC activator bryostatin 1 in a model of mild TBI (Zohar et al., 2011). However, also deleterious effects of ADAM10 were inferred from experiments using transgenic mice, expressing a dominant-negative ADAM10 mutant (ADAM10-dn). These mice showed a decreased seizure score and less hippocampal neuronal damage following kainate administration (Clement et al., 2008). More recently, layer-specific upregulation of ADAM10 was demonstrated in cortical neurons and microglia after whisker lesioning. In this model of cortical synapse remodeling, GI254023X, a selective inhibitor of ADAM10 (Ludwig et al., 2005), prevented ADAM10-mediated cleavage of the fraktalkine receptor CX3CR1 and thereby microglia-mediated synapse elimination (Gunner et al., 2019). Together, these studies imply a role for ADAM10 in various models of acute brain injury.
In this study, using controlled cortical impact (CCI) as a clinically relevant model of TBI, we examined the effects of GI254023X administration during the acute stage, at 30 min and 24 h post trauma, with respect to the neurological outcome and brain pathology at 7 days after injury (dpi). Combining behavioral assessment (immuno-) histology, immunohistochemistry, immunoblot, and mRNA expression analyses of various markers, we tested the hypothesis that pharmacological inhibition of ADAM10 has beneficial effects after experimental TBI.
Materials and Methods
Animals
All animal experiments were performed after approval by the animal Care and Ethics Committee of the Landesuntersuchungsamt Rheinland-Pfalz (protocol 23177-07/G13-01-018) observing the institutional guidelines of the Johannes Gutenberg University, Mainz, Germany. Adult male C57Bl/6N mice (8 weeks old) from Janvier (Germany) were investigated, weighing approximately 25 g. Animals were housed in a temperature-controlled environment (23 ± 1°C, 55 ± 5% relative humidity) with a 12 h light-dark circle and food and water ad libitum. A total of 40 mice was used in this study and randomly allocated to four experimental groups (sham: vehicle n = 9, GI254023X n = 7; CCI: vehicle n = 12, GI254023X n = 12). All experimenters, one performing the sham or CCI procedure and another one performing drug application, behavioral tests and tissue preparations, were blind to the treatment groups.
Experimental Traumatic Brain Injury, Physiological Parameters, and Behavioral Analysis
The CCI model was used to induce an experimental brain injury as described before (Hummel et al., 2020). Briefly, after induction of anesthesia (isoflurane 4 Vol.%, maintenance 2.1 Vol.%) mice were attached to a stereotactic apparatus (Kopf Instruments) and a right parietal craniotomy was performed. CCI was induced according to the following parameters: impactor tip diameter: 3 mm, impact velocity: 6 m/s, impact duration: 200 ms, displacement: 1.5 mm). After the procedure, skull and skin were carefully closed and the animals transferred to a neonatal incubator (IC8000, Draeger, Luebeck, Germany) for approximately 1 h with controlled air temperature and ambient humidity. Sham animals were handled identically in terms of anesthesia and skin incision. Since craniotomy already leads to brain damage in our TBI model (Cole et al., 2011) only slight drilling on the exposed skull surface instead of craniotomy was performed. Body temperature was controlled by a feedback heating device and adjusted to 37°C intraoperatively. Physiological and technical parameters, including body weight, rectal and pericranial temperature as well as anesthesia time and duration of operation, were monitored before and/or during the operation (Supplementary Table 1). Behavioral tests were performed in a blinded and unbiased fashion 1 day before and 1 and 7 days after CCI using a neurological severity score (NSS). The NSS was modified from Tsenter et al. (2008) and assesses the severity of neurological impairments ranging from 0 (no impairment) to 12, as described (Hummel et al., 2020).
Drug Treatment
The pharmacological ADAM10 inhibitor GI254023X was administered via intraperitoneal injection (100 mg/kg body weight, 30 min and 24 h after CCI). GI254023X was purchased from Aobious (CAS No.: 260264-93-5), and dissolved in 25% DMSO in 0.1 M Na2CO3. Control mice were handled identically but received vehicle solution (25% DMSO in 0.1 M Na2CO3). The anti-proteolytic activity of GI254023X was confirmed in vitro prior to vivo administration (Supplementary Figure 1).
Brain Sectioning and Histology
After the observation period of 7 days, mice were sacrificed by cervical dislocation in deep anesthesia (4 vol% isoflurane). Brains were collected and frozen in dry powdered ice and stored at −80°C. Serial cryosections of 12 μm were prepared using a cryostat (HM 560 Cryo-Star, Thermo Fisher Scientific, Walldorf, Germany) and collected in 500 μm intervals on glass slides (Superfrost plus). Sections were collected from Bregma +3.14 to −4.36 mm. Tissues between the intervals were processed for protein and RNA extractions as described (Menzel et al., 2017; Krämer et al., 2019). Cresyl violet staining was performed with 16 consecutive sections per mouse brain, images were taken using a stereo microscope (Stemi 305, Zeiss, Oberkochen, Germany) and the brain lesion areas were determined using ZEN imaging software tools (Zeiss, RRID:SCR_013672) by investigators blind to the treatment groups. Brain lesion volume was calculated by summation of areas multiplied by the distance between sections. Data were expressed relative to the volume of the ipsilesional hemisphere.
Immunohistochemistry and Immunoblotting
Analysis of immunohistochemical images for non-phosphorylated neurofilament was performed essentially as described (Menzel et al., 2017). Briefly, cryosections were fixed in 4% paraformaldehyde (PFA) in phosphate buffered saline (PBS) for 10 min, washed in PBS and incubated in blocking solution (5% goat serum, 0.5% bovine serum albumin, and 0.1% Triton-X-100, in PBS) for 1 h at room temperature. Primary antibodies were applied in blocking solution (Covance, mouse anti-non-phosphorylated neurofilament heavy chain, NF-H, clone SMI-32, 1:1,000, RRID:AB_509998) overnight at 4°C. Sections were then washed in PBS and incubated with an Alexa488-conjugated secondary anti-mouse antibodies (Life Technologies, Grand Island, NY, United States, 1:500, RRID:AB_2534069). Images were acquired using a confocal scanning microscope (LSM510, Zeiss) and identical filter and acquisition parameters. Analysis for non-phosphorylated neurofilament was performed using the ImageJ software (ImageJ, RRID:SCR_003070), appropriate threshold settings and the “Analyze Particle” plugin essentially as described (Menzel et al., 2017). All investigators were blinded to the treatment groups.
For immunoblotting, brain samples were collected during cryosectioning and homogenized in RIPA buffer and protein concentrations were determined by Lowry assay (BioRad). Immunoblotting and dot blot analysis were performed essentially as described (Hummel et al., 2020). Briefly, 30 μg of protein for each sample was resolved by 10% SDS-PAGE. Nitrocellulose membranes were washed in TBS containing 0.1% Tween, blocked with 2.5% non-fat milk and incubated with mouse anti-α-fodrin/αII-spectrin (Enzo Life Sciences, 1:750, RRID:AB_10554860), mouse anti-GAPDH (Acris Antibodies GmbH, 1:1,000, RRID:AB_1616730), and secondary infrared (IR) dye-conjugated antibodies (LI-COR; goat anti-mouse IgG IRDye 800, RRID:AB_10793856). For dot blot analysis, 10 μg of brain tissue protein (5 μg/μl, lysed in RIPA buffer) were dotted onto a nitrocellulose membrane, washed and incubated with goat anti-mouse IgG IRDye 800 (LI-COR, 1:10,000) for 1 h at room temperature. Protein band or protein dot densities were digitalized and quantified using the Odyssey SA imaging system and Odyssey CLx software (LI-COR biotechnology, RRID:SCR_014579).
Gene Expression Analysis
Gene expression was quantified as outlined before (Hummel et al., 2020). Briefly, RNA was extracted from brain tissue and reverse-transcribed into cDNA using RNeasy Kit and QuantiScript Reverse Transcription Kit (Qiagen), respectively. Equal amounts of cDNA were amplified by qPCR using a LightCycler (Hoffmann-La Roche AG). All values were normalized to the reference gene (PPIA) and absolute quantification was performed using a target specific standard curve and LightCycler Software (Hoffmann-La Roche AG, RRID:SCR_012155) and obtained data were expressed relative to the corresponding sham condition. Primer sequences, annealing temperature, and amplification products lengths are listed in Table 1.
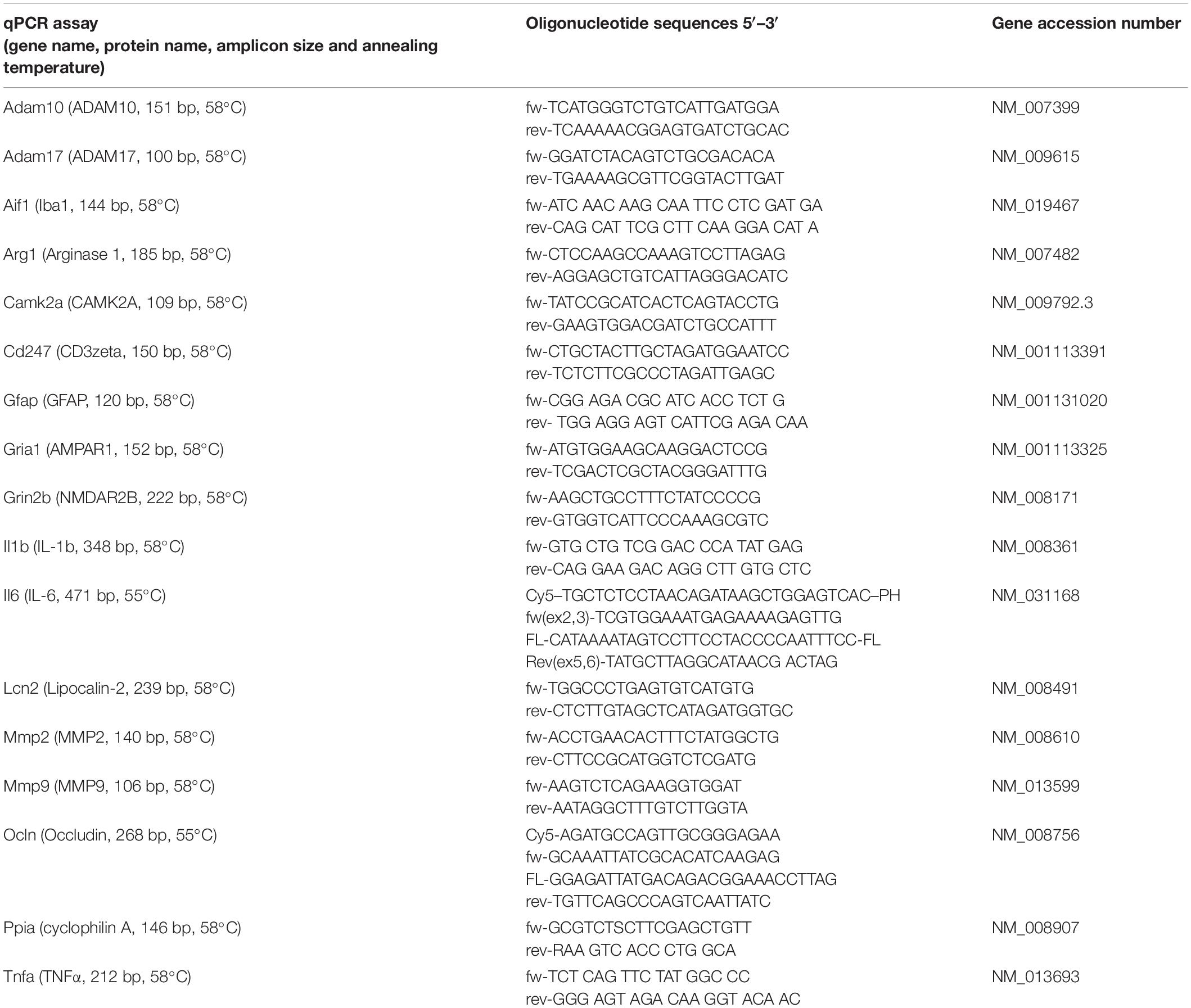
Table 1. Oligonucleotide primers.
Statistical Analysis
All analyses were performed using GraphPad Prism® (RRID:SCR_002798). Data distribution was analyzed using the Shapiro-Wilk-test. Dependent on data distribution, differences in pairwise comparisons were calculated by the Student’s t-test or the Mann-Whitney-U test. Multiple groups over different time points (body weight) were analyzed by two-way ANOVA, whereas non-parametrically distributed groups (NSS) were analyzed via Kruskal-Wallis test separately for each time point followed by Dunn’s multiple comparison test. If not otherwise stated, all other data were expressed relative to the corresponding sham condition. If applicable, data outliers were identified using ROUT’s test and removed as outlined in figure legends. Values from individual mice are shown as mean ± SEM, ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001, ****p < 0.0001.
Results
GI254023X Does Not Attenuate Neurological Deficits but Brain Tissue Loss After TBI
To study the influence of ADAM10 inhibition in acute brain injury, we used the CCI model of TBI which allows induction of highly reproducible and region-specific cortical injury (Smith et al., 1995). Mice were subjected to CCI or sham procedure. Body weight, rectal and pericranial temperature, as well as anesthesia and surgery duration of the procedure were within the expected range, before and during the procedure, respectively, and did not differ between the groups (Supplementary Table 1). GI254023X (100 mg/kg, i.p.) or vehicle were administered during the acute phase of TBI, at 30 min and 24 h after trauma. TBI led to a minor weight loss within the first 24 h after induction, which was almost restored at 7 dpi both in vehicle and GI254023X treated mice (Figure 1A). To assess neurological deficits and recovery after TBI, we used a modified 12-point neurological severity score (NSS) comprising motor ability, alertness, balance, and general behavior. Pronounced neurological deficits were evident at 1 dpi in CCI compared to sham groups (Figure 1B). At 7 dpi, the neurological deficits were decreased in the CCI groups compared to 1 dpi. However, the NSS did not differ between GI254023X- and vehicle-treated animals at 1 or 7 dpi (Figure 1B). To determine the structural brain damage at 7 dpi, brains were cut to coronal slices starting at Bregma + 3.14 mm and were stained with cresyl violet (Figure 1C). Next, the histopathological brain damage was determined by lesion volumetry (Figure 1D). This analysis revealed significantly smaller brain lesions in GI254023X-treated as compared to vehicle-treated mice. Hence, the posttraumatic administration of the ADAM10 inhibitor GI254023X does not attenuate neurological deficits but reduces brain tissue loss after TBI.
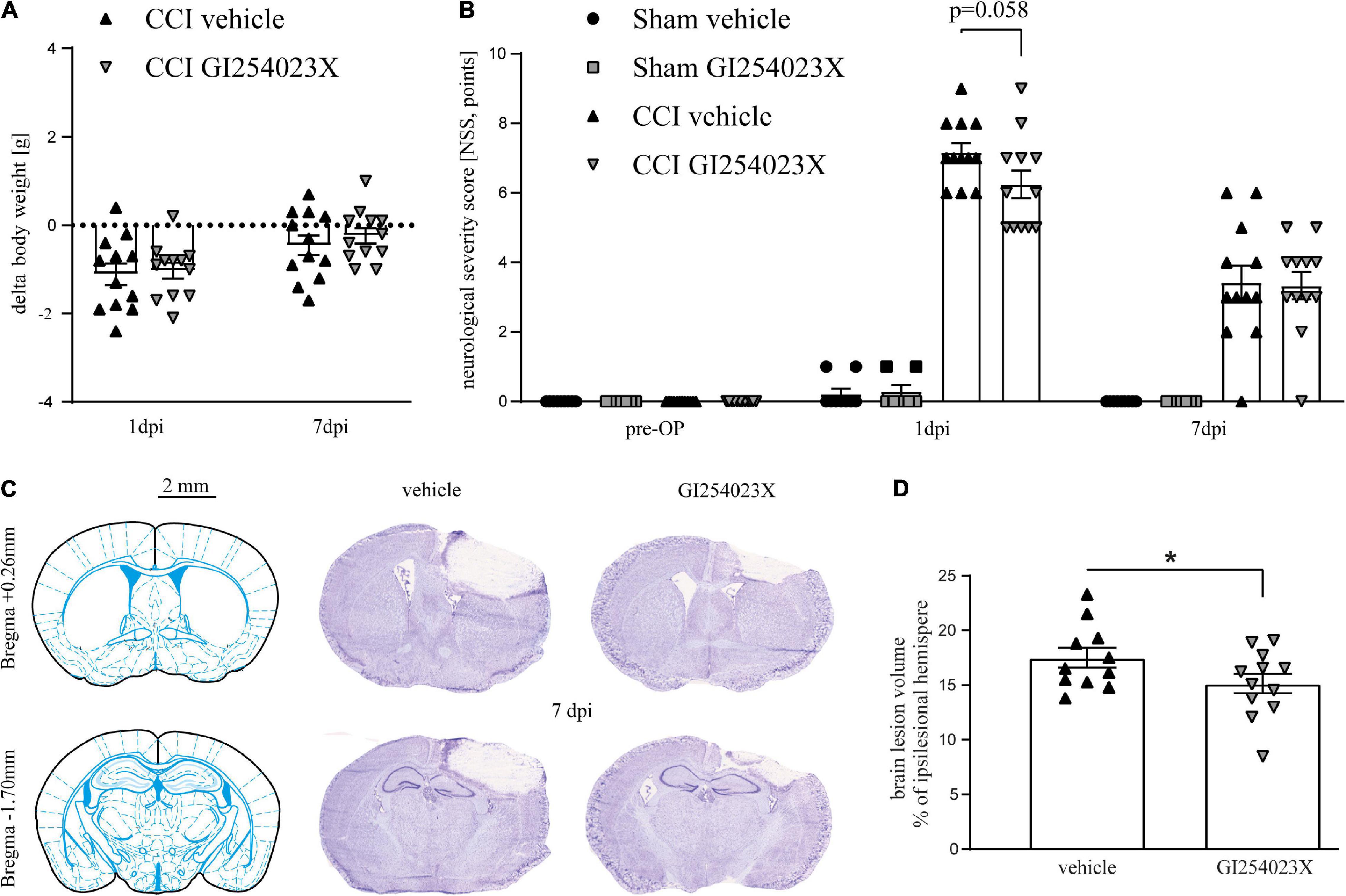
Figure 1. GI254023X does not attenuate neurological deficits but brain tissue loss after TBI. (A) Loss of body weight at 1 and 7 dpi compared to 1 day pre-OP in vehicle- or GI254023X-treated CCI mice. The dotted line refers to the pre-operation body weight. No differences were observed between vehicle and GI254023X treated mice as tested by two-way ANOVA. (B) Evaluation of neurological deficits using a NSS (0 = no impairment, 12 = maximal impairment). Animals were tested at the day before surgery (pre-OP), at 1 and 7 dpi. No differences were observed between vehicle and GI254023X treated mice as tested by Kruskal-Wallis test for each time point. (C) Schemes showing Bregma levels at +0.26 and −1.7 mm (Franklin and Paxinos Mouse Atlas 3rd edition) and cresyl violet stained coronal brain cryosections. Scale bar: 2 mm. (D) Brain lesion volumetry calculated from 16 consecutive sections at 7 dpi showing smaller lesions in GI254023X compared to vehicle treated mice as tested by Student’s t-test (*p < 0.05). Data are expressed as lesion volume in% of the ipsilesional hemisphere. One outlier was removed after identification by ROUT’s test in (D). Values from individual animals and mean ± SEM are shown.
GI254023X Influences Matrix Metalloproteinase Gene Expression but Not Overall BBB Integrity After TBI
To elucidate the effect of the ADAM10 inhibitor on the gene expression of metalloproteases, ipsilesional brain samples were investigated by qPCR at 7 dpi. Initially, mRNA expression levels of ADAM10 and its close homolog ADAM17 were examined. Both genes appeared up-regulated after TBI but no differences were observed between vehicle and GI254023X treatment (Figures 2A,B). Next, we examined expression regulation of the matrix metalloproteinase genes Mmp2 and Mmp9, which have been associated with BBB damage and neuronal cell death after TBI (Sifringer et al., 2007; Vajtr et al., 2009). Both genes appeared up-regulated in response to TBI but their expressions were attenuated by GI254023X treatment (Figures 2C,D). Previous findings on the role of ADAM10 on T cell migration (Schulz et al., 2008) prompted us to determine gene expression of the CD3 zeta subunit (Cd247) as a pan-T cell marker. Interestingly, the up-regulation of this gene was more pronounced in GI254023X-treated as compared to vehicle-treated animals (Figure 2E). Data on the mRNA expression of MMP2, MMP9, and Cd247 suggested alterations in BBB integrity. Therefore, we assessed BBB integrity by determining mRNA levels of the tight junction marker Occludin (Figure 2F) and the extravasation of Immunoglobulin G (IgG) as a proxy of BBB permeability (Figures 2G,H). However, we did not find differences for these markers suggesting that GI254023X did not influence overall BBB integrity after TBI.
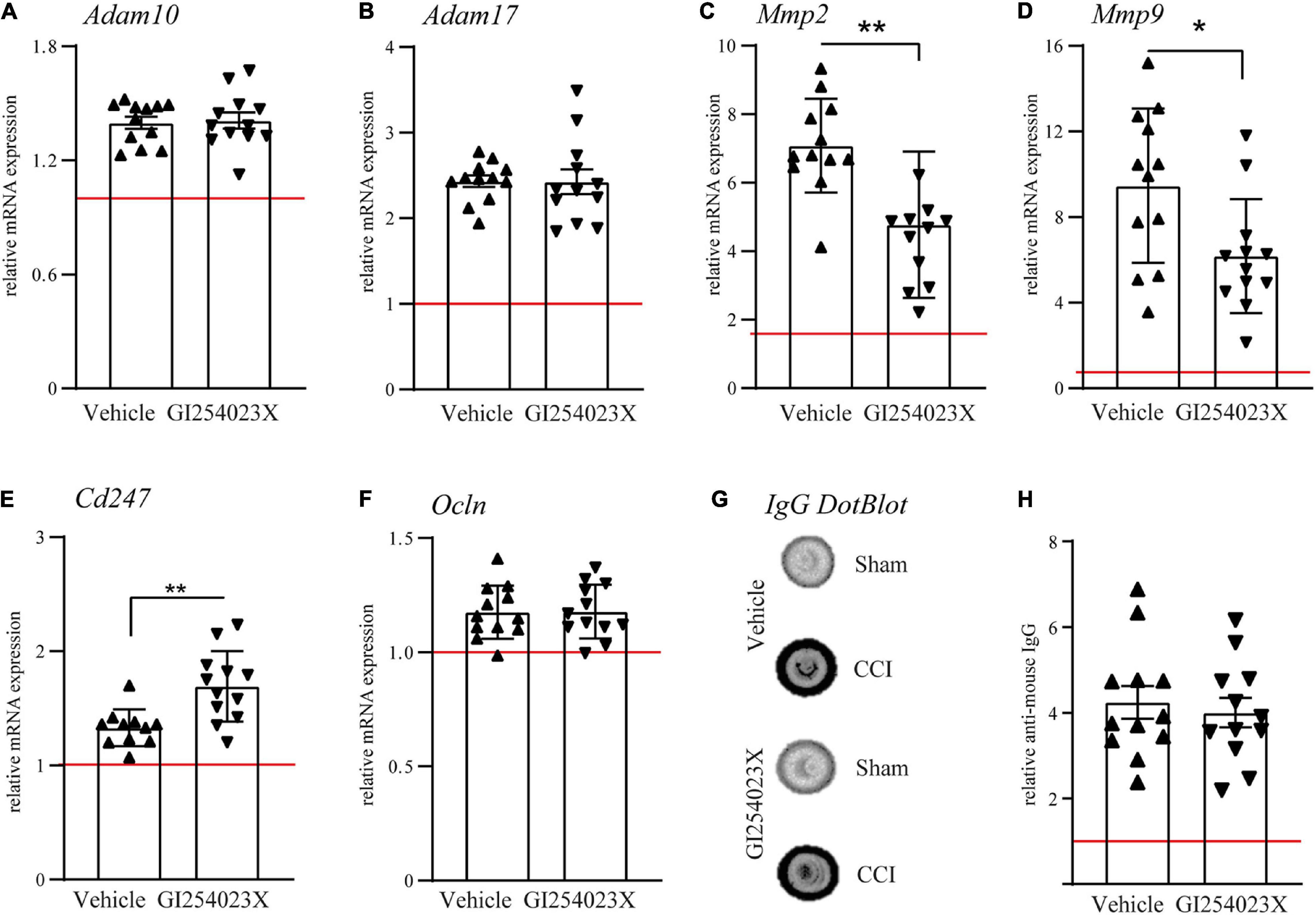
Figure 2. GI254023X influences matrix metalloproteinase gene expression but not overall BBB integrity after TBI. (A–F) Histograms showing relative mRNA expression of ADAM10, ADAM17, MMP2, MMP9, CD3 CD3zeta (Cd247), and Occludin (Ocln) at 7 dpi in ipsilesional brain tissue samples. (G) Representative anti-IgG Dot-blot using 7 dpi protein lysates from ipsilesional brain tissue. (H) Densitometric quantification of IgG dot blots as a proxy of BBB damage. mRNA expression values were normalized to the reference gene Ppia (cyclophilin A) and data are shown relative to the corresponding sham group (red line = sham). P-values were calculated by Student’s t-test or Mann-Whitney test (*p < 0.05, **p < 0.01). One outlier was removed after identification by ROUT’s test in (E). Values from individual animals and mean ± SEM are shown.
GI254023X Reduces Axonal Injury and Alters the Gene Expression of Glutamate Receptor Subunits After TBI
Our results showing smaller brain lesions in GI254023X-treated mice after TBI, prompted us to examine issues of brain homeostasis at 7 dpi. Therefore, we analyzed αII-spectrin breakdown products (SBDPs), which serve as indicators for disturbed neuronal Ca2+ homeostasis, axonal injury and degeneration after TBI (Gölz et al., 2019). Immunoblot analysis of protein lysates from ipsilesional brain tissues and corresponding samples from sham mice revealed a marked increase of the 145/150 kDa SBDPs (Figure 3A), known to result from increased caspase and calpain activity (Zhang et al., 2009). There was a 4.5 fold increase of SBDPs in brain lysates from vehicle treated mice but only a 1.7 fold increase in the corresponding brain lysates from GI254023X treated mice (Figure 3B). To further assess trauma-induced axonal injury at perilesional sites, we immunostained brain cryosections with the SMI-32 antibody clone specific to non-phosphorylated neurofilament (Figure 3C). The number of SMI-32 immunoreactive particles was lower in mice treated with GI254023X than in vehicle-treated mice (Figure 3D). As glutamate receptors play an important role in regulating intracellular calcium levels (Krishnamurthy and Laskowitz, 2016) and trigger neurotoxicity after TBI (Carvajal et al., 2016), we performed mRNA expression analyses of ipsilesonal brain tissue by quantitative PCR for the glutamate ionotropic receptor AMPA type subunit 1 (Gria1), the glutamate ionotropic receptor NMDA subunit 2B (Grin2b), and the Calcium/calmodulin-dependent protein kinase type II alpha chain (Camk2a) (Figures 3E–G). We found that GI254023X treatment increased the expression of Gria1 but lowers the expression of Grin2b compared to vehicle-treated animals at 7 dpi (Figures 3E,F). No effects of GI254023X were observed on the expression of Camk2a (Figure 3G). Together, these results show that GI254023X reduces axonal injury and alters the gene expression of glutamate receptor subunits after TBI.
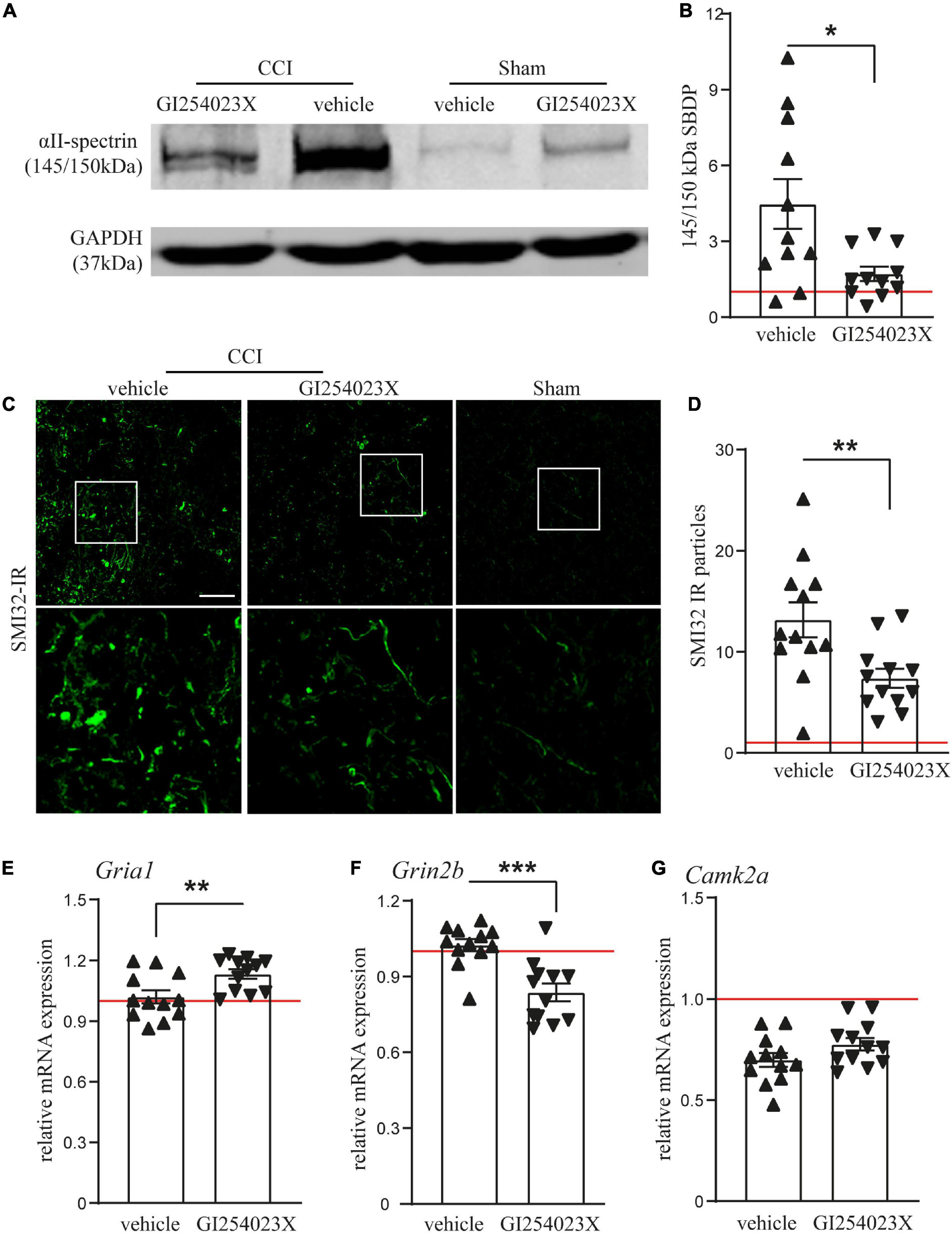
Figure 3. GI254023X reduces axonal injury and alters the gene expression of glutamate receptor subunits after TBI. (A) Representative western blot showing TBI-induced generation of spectrin breakdown products (SBDPs, 145–150 kDa) at 7 dpi in ipsilesional brain tissue samples. (B) Semi-quantitative densitometry of protein bands revealed reduced levels of SBDPs in GI254023X-treated animals compared to vehicle treatment (n = 11 each). (C) Immunofluorescence images of cryosections for non-phosphorylated Neurofilament-H (SMI-32) at 7 dpi at perilesional sites indicate increased axonal injury at 7 dpi in CCI animals compared to sham. (D) Quantitative assessment of SMI-32 immunoreactive (IR) particles at 7 dpi showing decreased numbers of SMI-32-IR particles in GI254023X- compared to vehicle-treated animals (n = 12 each). (E–G) Histograms showing relative mRNA expression of AMPAR1 (Gria1), NMDAR2B (Grin2b), and CamK2A (Camk2a) at 7 dpi in ipsilesional brain tissue samples. Expression values were normalized to the reference Ppia (cyclophilin A) and data from CCI animals (n = 12 each group) are shown relative to sham (red line = sham). Two outliers were removed after identification by ROUT’s test in (B). Values from individual animals and mean ± SEM are shown. Differences between GI254023X- and vehicle-treated animals were calculated by Student’s t-test (*p < 0.05, **p < 0.01, ***p < 0.001).
GI254023X Attenuates Pro-inflammatory Gene Expression After TBI
As ADAM10 activity regulates a variety of inflammatory processes, we examined mRNA expressions of the pro-inflammatory cytokines TNFα and IL-6 and the acute phase factor LCN2 along with markers for microglia/macrophages and/or reactive astrocytes (Figures 4A–F). TNFα and IL-6 mRNA expression levels were increased in response to TBI at 7 dpi but their up-regulation was attenuated in GI254023X-treated animals (Figures 4A,B). Similarly, LCN2 mRNA expression, which has been associated with astrocyte-mediated neurotoxicity (Bi et al., 2013), was induced by TBI but GI254023X attenuated its up-regulation (Figure 4C). However, no differences were found in the mRNA expression of the pan-microglia marker Aif1 or the M2 microglia marker Arg1 (Figures 4D,E). Also, the reactive astrocyte marker GFAP was induced by TBI but not different between GI254023X and vehicle-treated animals (Figure 4F). Together, these results show that GI254023X attenuates pro-inflammatory gene expression after TBI.
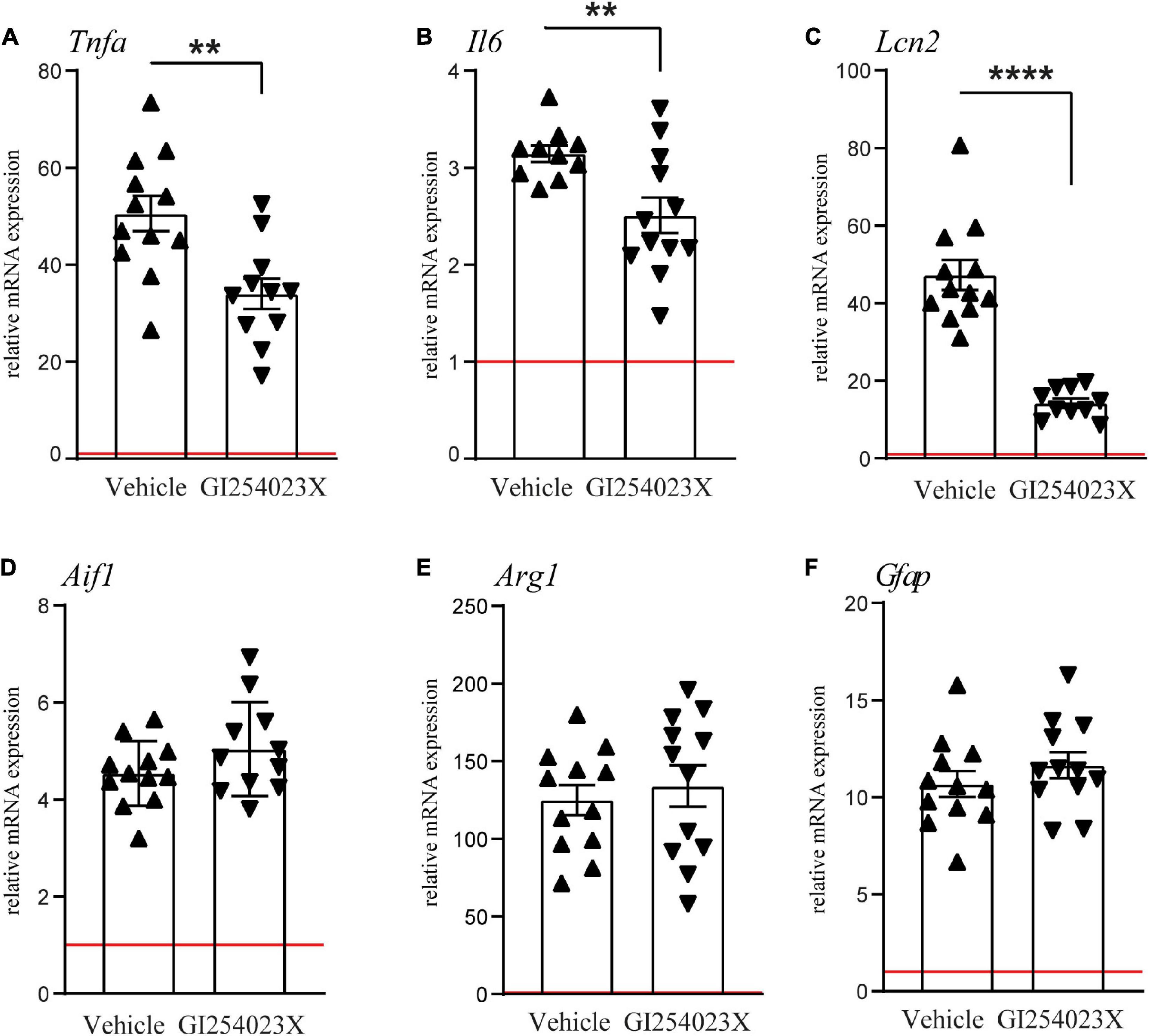
Figure 4. GI254023X attenuates pro-inflammatory gene expression after TBI. (A–F) Histograms showing relative mRNA expression of TNFa (Tnfa), IL-6 (Il6), LCN2 (Lcn2), Iba1 (Aif1), Arginase1 (Arg1), and GFAP (Gfap) at 7 dpi in ipsilesional brain tissue samples. Expression values were normalized to the reference gene Ppia and data are shown relative to sham (red line = sham). Outliers were removed from GI254023X- or vehicle-treated CCI groups after identification by ROUT’s test. Values from individual animals and mean ± SEM are shown. P-values were calculated by Student’s t-test (A–C) (**p < 0.01, ****p < 0.0001).
Discussion
We here report that the ADAM10 inhibitor GI254023X attenuates brain tissue loss, axonal injury and pro-inflammatory gene expression in the murine CCI model of TBI. GI254023X is a selective and potent inhibitor of ADAM10 that chelates the Zn2+ of the protease active site (Ludwig et al., 2005). A 100-fold selectivity of GI254023X was reported for the α-secretase ADAM10 over ADAM17 and several studies confirmed its selectivity (Pruessmeyer et al., 2014; Brummer et al., 2018; Vezzoli et al., 2019). GI254023X has been applied in various animal disease models. Beneficial effects were found in a mouse model of muscle injury and regeneration in which both genetic inhibition of ADAM10 and its pharmacological inhibition by GI254023X accelerated regeneration (Mizuno et al., 2018). In a bacterial sepsis model, GI254023X reduced vascular injury, vascular endothelial (VE) cadherin cleavage and concomitant loss of endothelial barrier function (Powers et al., 2012). Other studies using administration of GI254023X focused on mouse models of chronic neurodegenerative diseases such as Huntington disease (Vezzoli et al., 2019). It has been concluded that the inhibition of the proteolytic activity of ADAM10 by GI254023X has neuroprotective potential in this model. Interestingly, even in the context of AD, where beneficial effects of ADAM10 or its activation were shown by many laboratories in AD animal models (Postina et al., 2004; Tippmann et al., 2009; Zhang et al., 2013; Corbett et al., 2015; Meng et al., 2017) administration of GI254023X showed neuroprotective potential (Shackleton et al., 2016). Altogether, there is consensus on beneficial effects of GI254023X in various disease conditions and our study extends these findings in an animal model of TBI.
Up to date, ADAM10 inhibition by GI254023X has not been reported in the context of TBI or stroke, which share many pathophysiological features (Bramlett and Dietrich, 2004; Schäfer et al., 2014). In the present study we applied GI254023X at 30 min and 24 h after TBI which corresponds to the acute phase (Toshkezi et al., 2018). Previous studies reported upregulation of ADAM10 from 2 to 15 days after ECL (Warren et al., 2012) or 7 days after ECL (Del Turco et al., 2014). Our mRNA expression data suggest that ADAM10 expression is slightly up-regulated at 7 dpi, the time-point of histopathological examination. It has been proposed that ADAM10 plays a plasticity-enhancing and neuroprotective role during the first phase following injury. It shapes the extracellular environment for sprouting fibers, clears synaptic sites, and liberates neuroprotective APP fragments (Endres and Deller, 2017). Our results showing smaller lesions and reduced axonal injury in GI254023X-treated mice at 7 dpi contradict earlier assumptions that ADAM10 plays a neuroprotective role during the first phase following cerebral injury (Endres and Deller, 2017). It is possible, however, that the inherent differences between the ECL model and the CCI model of TBI used in this study, particularly in terms of the induction and localization of injury, may have an impact on whether inhibition or activation of ADAM10 exerts neuroprotective effects. Our results showing reduced αII-spectrin cleavage and less SMI32-immunoreactivity in the injured hemisphere indicate neuroprotective effects of GI254023X. These effects coincided with downregulation of Gria1 and upregulation of Grin2b, coding for the AMPA receptor subunit 1 (AMPAR1) and the NMDA receptor subunit 2B (NR2B), respectively. The downregulation of Grin2b in GI254023X-treated animals is intriguing as glutamate binding to extrasynaptic NR2B subunit triggers neurotoxicity (Carvajal et al., 2016). At the present, we can only speculate that expression changes of Gria1 and Grin2b contribute to the beneficial effects of ADAM10 inhibition by GI254023X. However, the neuroprotective effects of GI254023X observed in our study are in agreement with previous work demonstrating that transgenic mice expressing a dominant-negative ADAM10 mutant (ADAM10-dn) showed less neuronal cell death and neuroinflammation after kainate injection than wild-type mice, which indicates beneficial effects of ADAM10 inhibition in context with neurodegeneration (Clement et al., 2008).
Interestingly, ADAM10 has been ascribed a pro-inflammatory role in the early phase of acute lung inflammation through improved alveolar recruitment of leukocytes (Pruessmeyer et al., 2014). ADAM10 also enhances transendothelial migration of T cells (Schulz et al., 2008) which was inhibited by GI254023X. In addition, GI254023X increased T cells’ killing capacity and increased the number of T cells undergoing activation-induced cell death (Schulte et al., 2007). These findings suggest that GI254023X may have anti-inflammatory effects via attenuation of T cells infiltration into the lesioned brain. As compared to vehicle-treated mice, we here found in GI254023X treated mice an increased Cd247 gene expression (coding for CD3zeta) at 7 dpi in the lesioned brain, which we have earlier shown to correlate with increased T cell infiltration (Krämer et al., 2019). However, the previous observations did not support a critical role of T cells in brain damage progression probably due to their low numbers in the brain parenchyma (Krämer et al., 2019). It appears therefore unlikely that T cells counteract or contribute to the ameliorative effects of GI254023X treatment on brain tissue loss and axonal injury in the present study.
An important finding of our study is that GI254023X attenuated the mRNA expression of the pro-inflammatory cytokines TNFα and IL6 and the acute phase factor LCN2. These factors are associated with worse outcomes in TBI and have been considered as therapeutic targets (Tuttolomondo et al., 2014; Rodney et al., 2018). TNFα and IL6 are important mediators of inflammation and cause neuronal cell death and neurological dysfunction but this may also trigger reparative processes (Lenzlinger et al., 2001). LCN2 gene expression is increased early after TBI in mice (Huang et al., 2016) and represents a potent neurotoxic factor (Bi et al., 2013). To the best of our knowledge, there are no published data on a possible connection between LCN2 and ADAM10 but several studies support a regulatory role of ADAM10 in TNFα and IL-6 mediated actions. ADAM10 cleaves and thereby mediates the release of TNFα (Arduise et al., 2008) and has also been linked to the regulation of neural IL-6 levels after activation by aromatic retinoid acetritin (Dos Santos Guilherme et al., 2019). It has been further reported that TNFα-induced apoptosis can be regulated via an ADAM10-NF-κB feedback loop in a B cell lymphoma cell line (Yang et al., 2019). A similar feedback loop was shown for the ADAM family member ADAMTS-7, which upregulates TNF-α and conversely, TNF-α induced ADAMTS-7 through NF-κB signaling (Lai et al., 2014). It is therefore possible that the neuroprotective and anti-inflammatory effects of GI254023X treatment found in the present study are partially due to the interference with feedback loops involving ADAM10, TNFα, and NF-κB signaling. As mentioned before, neuronal cell death and neuroinflammation were also reduced in ADAM10-dn mice compared to wild-type mice following kainate-induced acute brain injury (Clement et al., 2008). Hence, future studies are needed to uncover the existence of feedback loops between pro-inflammatory cytokines and ADAM10 in TBI and other types of acute brain injuries.
To conclude, we here studied for the first time pharmacological inhibition of ADAM10 in experimental TBI. Our results showing beneficial effects on brain histopathology and inflammatory response to TBI suggest that ADAM10 may represent a therapeutic target in the acute phase of TBI. Further studies in the CCI model and other models of TBI are required to optimize treatment regimen and to investigate whether the beneficial effects of ADAM10 inhibition are restricted to the acute phase or can lead to sustained brain protection after TBI.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The animal study was reviewed and approved by the Animal Care and Ethics Committee of the Landesuntersuchungsamt Rheinland-Pfalz.
Author Contributions
DA, RH, MW, and CG performed the experiments and collected the data. MS and KE conceived the study. MS, KE, and RH designed the experiments and data analysis. MS, DA, and RH performed the data analysis and wrote the manuscript. All authors approved the final manuscript.
Funding
This study was supported by the Mainz Research School of Translational Biomedicine (TransMed) of the University Medical Center, Johannes Gutenberg-University Mainz, Germany and by the Deutsche Forschungsgemeinschaft (SCHA1261/4-3 to MS).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We gratefully acknowledge the excellent technical assistance of Wiesia Bobkiewicz, Tobias Hirnet, Frida Kornes, and Dana Pieter (Department of Anesthesiology, Mainz, Germany). We are grateful to Ralf Postina for providing the pcDNA3-ADAM10-HA plasmid. Part of the data presented in this study is submitted as a doctoral thesis by MW and as a professional dissertation (Habilitation) by RH to the Medical Faculty of Johannes Gutenberg-University, Mainz.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2021.661462/full#supplementary-material
References
Abdul-Muneer, P. M., Chandra, N., and Haorah, J. (2015). Interactions of oxidative stress and neurovascular inflammation in the pathogenesis of traumatic brain injury. Mol. Neurobiol. 51, 966–979. doi: 10.1007/s12035-014-8752-3
Akamatsu, Y., and Hanafy, K. A. (2020). Cell death and recovery in traumatic brain injury. Neurotherapeutics 17, 446–456. doi: 10.1007/s13311-020-00840-7
Arduise, C., Abache, T., Li, L., Billard, M., Chabanon, A., Ludwig, A., et al. (2008). Tetraspanins regulate ADAM10-mediated cleavage of TNF-alpha and epidermal growth factor. J. Immunol. (Baltimore Md. 1950) 181, 7002–7013. doi: 10.4049/jimmunol.181.10.7002
Bi, F., Huang, C., Tong, J., Qiu, G., Huang, B., Wu, Q., et al. (2013). Reactive astrocytes secrete lcn2 to promote neuron death. Proc. Natl. Acad. Sci. U.S.A. 110, 4069–4074. doi: 10.1073/pnas.1218497110
Bramlett, H. M., and Dietrich, W. D. (2004). Pathophysiology of cerebral ischemia and brain trauma: similarities and differences. J. Cereb. Blood Flow Metab. 24, 133–150. doi: 10.1097/01.wcb.0000111614.19196.04
Brummer, T., Pigoni, M., Rossello, A., Wang, H., Noy, P. J., Tomlinson, M. G., et al. (2018). The metalloprotease ADAM10 (a disintegrin and metalloprotease 10) undergoes rapid, postlysis autocatalytic degradation. FASEB J. 32, 3560–3573. doi: 10.1096/fj.201700823rr
Carvajal, F. J., Mattison, H. A., and Cerpa, W. (2016). Role of NMDA receptor-mediated glutamatergic signaling in chronic and acute neuropathologies. Neural Plast. 2016:2701526.
Claasen, A. M., Guevremont, D., Mason-Parker, S. E., Bourne, K., Tate, W. P., Abraham, W. C., et al. (2009). Secreted amyloid precursor protein-alpha upregulates synaptic protein synthesis by a protein kinase G-dependent mechanism. Neurosci. Lett. 460, 92–96. doi: 10.1016/j.neulet.2009.05.040
Clement, A. B., Hanstein, R., Schroder, A., Nagel, H., Endres, K., Fahrenholz, F., et al. (2008). Effects of neuron-specific ADAM10 modulation in an in vivo model of acute excitotoxic stress. Neuroscience 152, 459–468. doi: 10.1016/j.neuroscience.2007.10.060
Cole, J. T., Yarnell, A., Kean, W. S., Gold, E., Lewis, B., Ren, M., et al. (2011). Craniotomy: true sham for traumatic brain injury, or a sham of a sham? J. Neurotrauma 28, 359–369. doi: 10.1089/neu.2010.1427
Corbett, G. T., Gonzalez, F. J., and Pahan, K. (2015). Activation of peroxisome proliferator-activated receptor α stimulates ADAM10-mediated proteolysis of APP. Proc. Natl. Acad. Sci. U.S.A. 112, 8445–8450. doi: 10.1073/pnas.1504890112
Del Turco, D., Schlaudraff, J., Bonin, M., and Deller, T. (2014). Upregulation of APP, ADAM10 and ADAM17 in the denervated mouse dentate gyrus. PLoS One 9:e84962. doi: 10.1371/journal.pone.0084962
Dewan, M. C., Rattani, A., Gupta, S., Baticulon, R. E., Hung, Y. C., Punchak, M., et al. (2018). Estimating the global incidence of traumatic brain injury. J. Neurosurg. 130, 1039–1408.
Dos Santos Guilherme, M., Stoye, N. M., Rose-John, S., Garbers, C., Fellgiebel, A., and Endres, K. (2019). The synthetic retinoid acitretin increases IL-6 in the central nervous system of Alzheimer disease model mice and human patients. Front. Aging Neurosci. 11:182. doi: 10.3389/fnagi.2019.00182
Dreymueller, D., and Ludwig, A. (2017). Considerations on inhibition approaches for proinflammatory functions of ADAM proteases. Platelets 28, 354–361. doi: 10.1080/09537104.2016.1203396
Endres, K., and Deller, T. (2017). Regulation of alpha-secretase ADAM10 in vitro and in vivo: genetic, epigenetic, and protein-based mechanisms. Front. Mol. Neurosci. 10:56. doi: 10.3389/fnmol.2017.00056
Gavert, N., Sheffer, M., Raveh, S., Spaderna, S., Shtutman, M., Brabletz, T., et al. (2007). Expression of L1-CAM and ADAM10 in human colon cancer cells induces metastasis. Cancer Res. 67, 7703–7712. doi: 10.1158/0008-5472.can-07-0991
Gölz, C., Kirchhoff, F. P., Westerhorstmann, J., Schmidt, M., Hirnet, T., Rune, G. M., et al. (2019). Sex hormones modulate pathogenic processes in experimental traumatic brain injury. J. Neurochem. 150, 173–187. doi: 10.1111/jnc.14678
Gralle, M., Botelho, M. G., and Wouters, F. S. (2009). Neuroprotective secreted amyloid precursor protein acts by disrupting amyloid precursor protein dimers. J. Biol. Chem. 284, 15016–15025. doi: 10.1074/jbc.m808755200
Gunner, G., Cheadle, L., Johnson, K. M., Ayata, P., Badimon, A., Mondo, E., et al. (2019). Sensory lesioning induces microglial synapse elimination via ADAM10 and fractalkine signaling. Nat. Neurosci. 22, 1075–1088. doi: 10.1038/s41593-019-0419-y
Hsia, H.-E., Tüshaus, J., Brummer, T., Zheng, Y., Scilabra, S. D., and Lichtenthaler, S. F. (2019). Functions of ‘A disintegrin and metalloproteases (ADAMs)’ in the mammalian nervous system. Cell. Mol. Life Sci. 76, 3055–3081. doi: 10.1007/s00018-019-03173-7
Huang, C., Sakry, D., Menzel, L., Dangel, L., Sebastiani, A., Krämer, T., et al. (2016). Lack of NG2 exacerbates neurological outcome and modulates glial responses after traumatic brain injury. Glia 64, 507–523. doi: 10.1002/glia.22944
Hummel, R., Ulbrich, S., Appel, D., Li, S., Hirnet, T., Zander, S., et al. (2020). Administration of all-trans retinoic acid after experimental traumatic brain injury is brain protective. Br. J. Pharmacol. 177, 5208–5223. doi: 10.1111/bph.15259
Jorissen, E., Prox, J., Bernreuther, C., Weber, S., Schwanbeck, R., Serneels, L., et al. (2010). The disintegrin/metalloproteinase ADAM10 is essential for the establishment of the brain cortex. J. Neurosci. 30, 4833–4844. doi: 10.1523/jneurosci.5221-09.2010
Krämer, T. J., Hack, N., Brühl, T. J., Menzel, L., Hummel, R., Griemert, E. V., et al. (2019). Depletion of regulatory T cells increases T cell brain infiltration, reactive astrogliosis, and interferon-γ gene expression in acute experimental traumatic brain injury. J. Neuroinflamm. 16:163.
Krishnamurthy, K., and Laskowitz, D. T. (2016). “Cellular and molecular mechanisms of secondary neuronal injury following traumatic brain injury,” in Translational Research in Traumatic Brain Injury, eds D. Laskowitz and G. Grant (Boca Raton FL: CRC Press).
Lai, Y., Bai, X., Zhao, Y., Tian, Q., Liu, B., Lin, E. A., et al. (2014). ADAMTS-7 forms a positive feedback loop with TNF-α in the pathogenesis of osteoarthritis. Ann. Rheum. Dis. 73, 1575–1584. doi: 10.1136/annrheumdis-2013-203561
Lambrecht, B. N., Vanderkerken, M., and Hammad, H. (2018). The emerging role of ADAM metalloproteinases in immunity. Nat. Rev. Immunol. 18, 745–758. doi: 10.1038/s41577-018-0068-5
Lenzlinger, P. M., Morganti-Kossmann, M. C., Laurer, H. L., and McIntosh, T. K. (2001). The duality of the inflammatory response to traumatic brain injury. Mol. Neurobiol. 24, 169–181. doi: 10.1385/mn:24:1-3:169
Loane, D. J., and Faden, A. I. (2010). Neuroprotection for traumatic brain injury: translational challenges and emerging therapeutic strategies. Trends Pharmacol. Sci. 31, 596–604. doi: 10.1016/j.tips.2010.09.005
Ludwig, A., Hundhausen, C., Lambert, M. H., Broadway, N., Andrews, R. C., Bickett, D. M., et al. (2005). Metalloproteinase inhibitors for the disintegrin-like metalloproteinases ADAM10 and ADAM17 that differentially block constitutive and phorbol ester-inducible shedding of cell surface molecules. Comb. Chem. High Throughput Screen. 8, 161–171. doi: 10.2174/1386207053258488
Meng, T., Cao, Q., Lei, P., Bush, A. I., Xiang, Q., Su, Z., et al. (2017). Tat-haFGF(14-154) upregulates ADAM10 to attenuate the Alzheimer phenotype of APP/PS1 mice through the PI3K-CREB-IRE1α/XBP1 pathway. Mol. Ther. Nucleic Acids 7, 439–452. doi: 10.1016/j.omtn.2017.05.004
Menzel, L., Kleber, L., Friedrich, C., Hummel, R., Dangel, L., Winter, J., et al. (2017). Progranulin protects against exaggerated axonal injury and astrogliosis following traumatic brain injury. Glia 65, 278–292. doi: 10.1002/glia.23091
Mizuno, S., Yoda, M., Shimoda, M., Chiba, K., Nakamura, M., and Horiuchi, K. (2018). Inhibition of ADAM10 in satellite cells accelerates muscle regeneration following muscle injury. J. Orthop. Res. 36, 2259–2265. doi: 10.1002/jor.23878
Morganti-Kossmann, M. C., Semple, B. D., Hellewell, S. C., Bye, N., and Ziebell, J. M. (2019). The complexity of neuroinflammation consequent to traumatic brain injury: from research evidence to potential treatments. Acta Neuropathol. 137, 731–755. doi: 10.1007/s00401-018-1944-6
Postina, R., Schroeder, A., Dewachter, I., Bohl, J., Schmitt, U., Kojro, E., et al. (2004). A disintegrin-metalloproteinase prevents amyloid plaque formation and hippocampal defects in an Alzheimer disease mouse model. J. Clin. Invest. 113, 1456–1464. doi: 10.1172/jci20864
Powers, M. E., Kim, H. K., Wang, Y., and Bubeck Wardenburg, J. (2012). ADAM10 mediates vascular injury induced by Staphylococcus aureus α-Hemolysin. J. Infect. Dis. 206, 352–356. doi: 10.1093/infdis/jis192
Prox, J., Bernreuther, C., Altmeppen, H., Grendel, J., Glatzel, M., D’Hooge, R., et al. (2013). Postnatal disruption of the disintegrin/metalloproteinase ADAM10 in brain causes epileptic seizures, learning deficits, altered spine morphology, and defective synaptic functions. J. Neurosci. 33, 12915–12928, 12928a.
Pruessmeyer, J., Hess, F. M., Alert, H., Groth, E., Pasqualon, T., Schwarz, N., et al. (2014). Leukocytes require ADAM10 but not ADAM17 for their migration and inflammatory recruitment into the alveolar space. Blood 123, 4077–4088. doi: 10.1182/blood-2013-09-511543
Rodney, T., Osier, N., and Gill, J. (2018). Pro- and anti-inflammatory biomarkers and traumatic brain injury outcomes: a review. Cytokine 110, 248–256. doi: 10.1016/j.cyto.2018.01.012
Schäfer, M. K., Pfeiffer, A., Jaeckel, M., Pouya, A., Dolga, A. M., and Methner, A. (2014). Regulators of mitochondrial Ca(2+) homeostasis in cerebral ischemia. Cell Tissue Res. 357, 395–405. doi: 10.1007/s00441-014-1807-y
Schäfer, M. K. E., and Tegeder, I. (2018). NG2/CSPG4 and progranulin in the posttraumatic glial scar. Matrix Biol. 6, 571–588. doi: 10.1016/j.matbio.2017.10.002
Schulte, M., Reiss, K., Lettau, M., Maretzky, T., Ludwig, A., Hartmann, D., et al. (2007). ADAM10 regulates FasL cell surface expression and modulates FasL-induced cytotoxicity and activation-induced cell death. Cell Death Differ. 14, 1040–1049. doi: 10.1038/sj.cdd.4402101
Schulz, B., Pruessmeyer, J., Maretzky, T., Ludwig, A., Blobel, C. P., Saftig, P., et al. (2008). ADAM10 regulates endothelial permeability and T-Cell transmigration by proteolysis of vascular endothelial cadherin. Circ. Res. 102, 1192–1201. doi: 10.1161/circresaha.107.169805
Sépult, C., Bellefroid, M., Rocks, N., Donati, K., Gérard, C., Gilles, C., et al. (2019). ADAM10 mediates malignant pleural mesothelioma invasiveness. Oncogene 38, 3521–3534. doi: 10.1038/s41388-018-0669-2
Shackleton, B., Crawford, F., and Bachmeier, C. (2016). Inhibition of ADAM10 promotes the clearance of Aβ across the BBB by reducing LRP1 ectodomain shedding. Fluids Barriers CNS 13:14.
Sifringer, M., Stefovska, V., Zentner, I., Hansen, B., Stepulak, A., Knaute, C., et al. (2007). The role of matrix metalloproteinases in infant traumatic brain injury. Neurobiol. Dis. 25, 526–535. doi: 10.1016/j.nbd.2006.10.019
Smith, D. H., Soares, H. D., Pierce, J. S., Perlman, K. G., Saatman, K. E., Meaney, D. F., et al. (1995). A model of parasagittal controlled cortical impact in the mouse: cognitive and histopathologic effects. J. Neurotrauma 12, 169–178. doi: 10.1089/neu.1995.12.169
Sun, L., Zhao, M., Zhang, J., Liu, A., Ji, W., Li, Y., et al. (2017). MiR-144 promotes beta-amyloid accumulation-induced cognitive impairments by targeting ADAM10 following traumatic brain injury. Oncotarget 8, 59181–59203. doi: 10.18632/oncotarget.19469
Thornton, E., Vink, R., Blumbergs, P. C., and Van Den Heuvel, C. (2006). Soluble amyloid precursor protein alpha reduces neuronal injury and improves functional outcome following diffuse traumatic brain injury in rats. Brain Res. 1094, 38–46. doi: 10.1016/j.brainres.2006.03.107
Tippmann, F., Hundt, J., Schneider, A., Endres, K., and Fahrenholz, F. (2009). Up-regulation of the alpha-secretase ADAM10 by retinoic acid receptors and acitretin. FASEB J. 23, 1643–1654. doi: 10.1096/fj.08-121392
Toshkezi, G., Kyle, M., Longo, S. L., Chin, L. S., and Zhao, L. R. (2018). Brain repair by hematopoietic growth factors in the subacute phase of traumatic brain injury. J. Neurosurg. 129, 1286–1294. doi: 10.3171/2017.7.jns17878
Tsang, J. Y. S., Lee, M. A., Chan, T. H., Li, J., Ni, Y. B., Shao, Y., et al. (2018). Proteolytic cleavage of amyloid precursor protein by ADAM10 mediates proliferation and migration in breast cancer. EBioMedicine 38, 89–99. doi: 10.1016/j.ebiom.2018.11.012
Tsenter, J., Beni-Adani, L., Assaf, Y., Alexandrovich, A. G., Trembovler, V., and Shohami, E. (2008). Dynamic changes in the recovery after traumatic brain injury in mice: effect of injury severity on T2-weighted MRI abnormalities, and motor and cognitive functions. J. Neurotrauma 25, 324–333. doi: 10.1089/neu.2007.0452
Tuttolomondo, A., Pecoraro, R., and Pinto, A. (2014). Studies of selective TNF inhibitors in the treatment of brain injury from stroke and trauma: a review of the evidence to date. Drug Design Dev. Ther. 8, 2221–2238. doi: 10.2147/dddt.s67655
Vajtr, D., Benada, O., Kukacka, J., Prùsa, R., Houstava, L., Toupalík, P., et al. (2009). Correlation of ultrastructural changes of endothelial cells and astrocytes occurring during blood brain barrier damage after traumatic brain injury with biochemical markers of BBB leakage and inflammatory response. Physiol. Res. 58, 263–268. doi: 10.33549/physiolres.931253
Vezzoli, E., Caron, I., Talpo, F., Besusso, D., Conforti, P., Battaglia, E., et al. (2019). Inhibiting pathologically active ADAM10 rescues synaptic and cognitive decline in Huntington’s disease. J. Clin. Invest. 129, 2390–2403. doi: 10.1172/jci120616
Warren, K. M., Reeves, T. M., and Phillips, L. L. (2012). MT5-MMP, ADAM-10, and N-cadherin act in concert to facilitate synapse reorganization after traumatic brain injury. J. Neurotrauma 29, 1922–1940. doi: 10.1089/neu.2012.2383
Werner, C., and Engelhard, K. (2007). Pathophysiology of traumatic brain injury. Br. J. Anaesth. 99, 4–9.
Wetzel, S., Seipold, L., and Saftig, P. (2017). The metalloproteinase ADAM10: a useful therapeutic target? Biochim. Biophys. Acta (BBA) Mol. Cell Res. 1864, 2071–2081. doi: 10.1016/j.bbamcr.2017.06.005
Yang, H., Qiu, B., Chen, S., Xun, Y., Pan, Y., Chen, M., et al. (2019). Soluble CXCL16 promotes TNF-α-induced apoptosis in DLBCL via the AMAD10-NF-κB regulatory feedback loop. Cell Biol. Int. 43, 863–874. doi: 10.1002/cbin.11154
Zhang, S. Q., Sawmiller, D., Li, S., Rezai-Zadeh, K., Hou, H., Zhou, S., et al. (2013). Octyl gallate markedly promotes anti-amyloidogenic processing of APP through estrogen receptor-mediated ADAM10 activation. PLoS One 8:e71913. doi: 10.1371/journal.pone.0071913
Zhang, Z., Larner, S. F., Liu, M. C., Zheng, W., Hayes, R. L., and Wang, K. K. (2009). Multiple alphaII-spectrin breakdown products distinguish calpain and caspase dominated necrotic and apoptotic cell death pathways. Apoptosis 14, 1289–1298. doi: 10.1007/s10495-009-0405-z
Keywords: traumatic brain injury, ADAM10 (a disintegrin and metalloprotease 10), GI254023X, neuroprotection, neuroinflammation, axonal injury
Citation: Appel D, Hummel R, Weidemeier M, Endres K, Gölz C and Schäfer MKE (2021) Pharmacologic Inhibition of ADAM10 Attenuates Brain Tissue Loss, Axonal Injury and Pro-inflammatory Gene Expression Following Traumatic Brain Injury in Mice. Front. Cell Dev. Biol. 9:661462. doi: 10.3389/fcell.2021.661462
Received: 30 January 2021; Accepted: 23 February 2021;
Published: 15 March 2021.
Edited by:
David Lutz, Ruhr-University Bochum, GermanyReviewed by:
Igor Jakovcevski, Helmholtz Association of German Research Centers (HZ), GermanyAdam Bachstetter, University of Kentucky, United States
Copyright © 2021 Appel, Hummel, Weidemeier, Endres, Gölz and Schäfer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael K. E. Schäfer, TWljaGFlbC5TY2hhZWZlckB1bmltZWRpemluLW1haW56LmRl; orcid.org/0000-0001-6055-6244
†These authors have contributed equally to this work and share first authorship