 Adrien Bosseboeuf1
Adrien Bosseboeuf1 Célia Seillier1
Célia Seillier1 Nicolas Mennesson1
Nicolas Mennesson1 Sophie Allain-Maillet1
Sophie Allain-Maillet1 Maeva Fourny1Anne Tallet2
Maeva Fourny1Anne Tallet2 Eric Piver2,3
Eric Piver2,3 Philippe Lehours4,5
Philippe Lehours4,5 Francis Mégraud4,5Laureline Berthelot6
Francis Mégraud4,5Laureline Berthelot6 Jean Harb1,6,7
Jean Harb1,6,7 Edith Bigot-Corbel1,7
Edith Bigot-Corbel1,7 Sylvie Hermouet1,8*
Sylvie Hermouet1,8*- 1CRCINA, Inserm, Université de Nantes, Université d'Angers, Nantes, France
- 2Laboratoire de Biochimie, CHU de Tours, Tours, France
- 3Inserm UMR966, Tours, France
- 4Inserm U1053, Université de Bordeaux, Bordeaux, France
- 5Laboratoire de Bactériologie, Centre National de Reference des Campylobacters et des Hélicobacters, CHU de Bordeaux, Bordeaux, France
- 6Centre de Recherche en Transplantation et Immunologie UMR1064, Inserm, Université de Nantes, Nantes, France
- 7Laboratoire de Biochimie, CHU de Nantes, Nantes, France
- 8Laboratoire d'Hématologie, CHU de Nantes, Nantes, France
Previous studies showed that monoclonal immunoglobulins G (IgGs) of “monoclonal gammopathy of undetermined significance” (MGUS) and myeloma were hyposialylated, thus presumably pro-inflammatory, and for about half of patients, the target of the monoclonal IgG was either a virus—Epstein–Barr virus (EBV), other herpes viruses, hepatitis C virus (HCV)—or a glucolipid, lysoglucosylceramide (LGL1), suggesting antigen-driven disease in these patients. In the present study, we show that monoclonal IgAs share these characteristics. We collected 35 sera of patients with a monoclonal IgA (6 MGUS, 29 myeloma), and we were able to purify 25 of the 35 monoclonal IgAs (6 MGUS, 19 myeloma). Monoclonal IgAs from MGUS and myeloma patients were significantly less sialylated than IgAs from healthy volunteers. When purified monoclonal IgAs were tested against infectious pathogens and LGL1, five myeloma patients had a monoclonal IgA that specifically recognized viral proteins: the core protein of HCV in one case, EBV nuclear antigen 1 (EBNA-1) in four cases (21.1% of IgA myeloma). Monoclonal IgAs from three myeloma patients reacted against LGL1. In summary, monoclonal IgAs are hyposialylated and as described for IgG myeloma, significant subsets (8/19, or 42%) of patients with IgA myeloma may have viral or self (LGL1) antigen-driven disease.
Introduction
Myeloma is preceded by an asymptomatic stage termed monoclonal gammopathy of undetermined significance (MGUS) (1–4). In MGUS and myeloma, clonal plasma cells produce large quantities of a so-called “monoclonal” immunoglobulin (Ig). In MGUS, clonal plasma cells represent <10% of bone marrow cells, and the amount of monoclonal Ig in blood is <30 g/L. Following the acquisition of genetic alterations in clonal plasma cells, a fraction of MGUS progress over time toward smoldering, then overt myeloma (5, 6). The rate of transformation of MGUS toward myeloma is 1% per year per individual. In myeloma, patients suffer from diverse renal, bone, and hematological complications; clonal plasma cells represent >10% of bone marrow cells, and the amount of monoclonal Ig in blood is >30 g/L (6). For 55–60% of MGUS and myeloma patients, the monoclonal Ig is type G, for 20–25%, it is type A, and for <5%, it is type D, M, or E; light chain myeloma represent ~15% cases (7).
The causes of MGUS have long remained unknown, although it is established that chronic infection may lead to the emergence of oligoclonal and eventually, monoclonal plasma cells and the subsequent production of a so-called “monoclonal Ig.” In addition, it is thought that certain genetic backgrounds, ethnicity, lipid disorders, and obesity may increase the risk of developing myeloma. Consistent with these observations, the study of Ig genes in malignant plasma cells had revealed restricted heavy-chain variable-region use and highly hypermutated Ig heavy- and light-chain genes, which supports antigen-mediated selection of the malignant clone (8, 9). Recent studies identified several types of antigens that are specifically recognized by monoclonal IgGs, notably lysoglucosylceramide (LGL1) (10, 11), and infectious antigens (12–14). Thus, it has been proposed that chronic simulation by glucolipidic auto-antigens or by infectious antigens may underlie the pathogenesis of subsets of IgG MGUS and myeloma.
Identification of LGL1 as a frequent target of the plasma cell clone in MGUS and in myeloma resulted from the study of patients with Gaucher disease (10, 11). Gaucher disease is a genetic disorder in which a glucocerebroside (or glucosylceramide) accumulates, and MGUS and myeloma are more frequent in Gaucher patients than in the general population. Nair et al. demonstrated that the monoclonal Ig of patients with Gaucher disease frequently target LGL1, a glucosylceramide present in excess in these individuals (10, 11). Moreover, up to a third of monoclonal Igs from patients without Gaucher disease—with sporadic MGUS or myeloma—may also target LGL1, which suggested a link between chronic stimulation by a self-antigen and the development of MGUS and myeloma (10, 11). In parallel, our group investigated whether an abnormal immune response to latent infection may lead to MGUS and eventually, myeloma. We designed a new assay, called the multiplexed infectious antigen micro-array (MIAA), which carries proteins and lysates from nine infectious pathogens, to analyze the specificity of infectious antigen recognition of purified monoclonal IgGs from MGUS or myeloma patients (15). Using the MIAA assay, we found that purified monoclonal IgGs reacted with several infectious pathogens known to cause latent infection. These pathogens include herpesviruses, especially Epstein–Barr virus (EBV), and hepatitis C virus (HCV) (12–15). EBV and HCV are oncogenic viruses associated with solid cancers and B-cell malignancies such as Hodgkin and non-Hodgkin lymphoma, mostly directly via cell infection and transformation (16–19). In contrast, EBV DNA is rarely detected in malignant plasma cells (20). Thus, in MGUS and myeloma with EBV-specific IgG, malignant transformation presumably occurs indirectly, without infection of tumor cells, via chronic antigen-driven stimulation of the B-cell receptor (BCR) that has identical heavy and light chain specificity to the secreted IgG. Interestingly, monoclonal IgGs may contribute to the inflammation associated with MGUS and myeloma, since they have a very low level of sialylation of the Fc fragment, a characteristic typically associated with a pro-inflammatory action (21).
In contrast to the monoclonal IgGs of MGUS and myeloma patients, the characteristics and antigenic targets of monoclonal IgAs have not been studied. IgA myeloma is relatively rare and differs from IgG myeloma by a worse prognosis and shorter survival: patients with IgA myeloma are considered more at risk of bone destruction, extra-medullary disease, infection, and hyper-viscosity facilitated by greater polymerization of IgAs compared to IgGs (22–28). In the present study, we were able to collect serum from 35 patients with a monoclonal IgA; 25/35 monoclonal IgAs were successfully separated from other Igs. The specificity of antigenic recognition of the purified monoclonal IgAs was analyzed using the MIAA and an adapted LGL1 assay; the isotype of monoclonal IgAs was also determined, and their degree of sialylation was quantified.
Materials and Methods
Patients
We examined 35 patients with a monoclonal IgA (6 MGUS, 29 myeloma). Among those, 6 MGUS and 22 myeloma were diagnosed at the University Hospitals (CHUs) in Tours and Bordeaux (France) over the 2010–2016 period. Samples of blood serum from seven additional patients with IgA myeloma from international cohorts of relapsed myeloma were provided by Novartis (Basel, Switzerland).
Purification of Monoclonal IgAs and Determination of Isotype
After clotting, blood samples were centrifuged at 2,200 × g (4°C), and serum aliquots were frozen. Measurement of Ig concentration, separation of monoclonal IgAs from other Igs, and verification of purity were performed as described previously (12–15, 21). Briefly, the presence of a monoclonal IgA in serum is first established in clinical laboratories, then purification of the monoclonal IgA is performed. The protocol of purification starts with a high resolution agarose gel electrophoresis (SAS-MX high resolution; Helena Biosciences, Gateshead, UK), which allows us to elute the monoclonal Ig from the gel, for elution in PBS. The purity of the monoclonal IgA preparation is then verified by isoelectrofocusing on an agarose gel (pH 3–10) followed by blotting and immunorevelation by an anti-human IgA alpha chain antibody labeled with peroxidase. In some cases, the monoclonal IgA preparation still contains very small amounts of other IgAs (oligo- or poly-clonal), always in very low concentration and not detectable by our techniques. Moreover, eventual contamination by IgG is not relevant here because all further assays are revealed using anti-human IgA alpha chain antibodies.
To determine the A1/A2 isotype, 96-well plates (Nunc MaxiSorp™) were coated overnight at 4°C with 50 μl of goat anti-human IgA antibody (Southern Biotech, Birmingham, AL, USA) diluted 1:500 in 25 mM borate buffer pH 9. After washing with PBS-Tween 0.05%, wells were saturated for 2 h at 37°C with 0.25% B-grade bovine gelatin (Sigma, St. Louis, MO, USA) diluted in 0.1% PBS-Tween; 50 μl of monoclonal IgA (400 ng/ml) was then added (2-h incubation, 37°C). After washing, 50 μl of biotinylated mouse anti-human IgA1 or IgA2 antibody (0.5 μg/ml; Southern Biotech, Birmingham, AL, USA) was added (2-h incubation, 37°C). After washing, 50 μl of streptavidin-HRP (1 μg/ml; Vector Laboratories, Burlingame, CA, USA) was added (1-h incubation, 37°C). After washing, 50 μl of 3,3′,5,5′-tetramethylbenzidine (TMB) was added. The reaction was stopped with 50 μl of sulfuric acid (0.5 M). Optical density was read at 450 nm using a Spark 10 M multimode microplate reader (Tecan, Männedorf, Switzerland).
The MIAA Assay
The MIAA assay allows testing for panels of commercially available proteins, antigens, or/and lysates from EBV, herpes simplex virus 1 (HSV-1), HSV-2, cytomegalovirus (CMV), varicella zoster virus (VZV), HCV, Helicobacter pylori (H. pylori), Toxoplasma gondii (T. gondii), and Borrelia burgdorferi (B. burgdorferi) (12–15, 21). For incubation on MIAA arrays, Ig concentrations were adjusted to 400 μg/ml (serum) or 50–200 μg/ml (purified monoclonal IgAs) in 80 μl. After washing, MIAA slides were incubated with DylightTM 680-labeled goat anti-human IgA Fc antibody (1:2,500; 0.4 μg/ml; Immuno Reagents, Raleigh, NC, USA). Fluorescence signals were detected with the Odyssey infrared imaging system scanner at 21-μm resolution (LI-COR Biosciences, Lincoln, NE, USA) and quantified using the GenePix® Pro 4 Microarray Acquisition and Analysis Software (Molecular Devices, Sunnyvale, CA, USA) (12–15, 21). Five fluorescence thresholds of specific positivity were determined using positive and negative controls: 500, for HCV, H. pylori, T. gondii; 1,000, for HSV-1 and HSV-2; 1,200, for CMV; 1,400, for EBV and VZV; and 1,800 for B. burgdorferi. Fluorescent signals below the thresholds were considered negative (12, 21).
Dot Blotting Assays
Nitrocellulose membranes (Amersham, Buckinghamshire, UK) were spotted with recombinant EBNA-1 protein, relevant and irrelevant EBV peptides, or HCV core protein, then dried (12). Saturation of membranes, and incubation with serum or purified monoclonal IgA were performed as published (12). Antigen–IgA complexes were revealed using an HRP-labeled goat anti-human IgA (α chain) from Bethyl Laboratories (Montgomery, TX, USA).
LGL1 Immunoblotting Assay
For LGL1-specific immunoblotting, polyvinylidene fluoride (PVDF) membranes were incubated for 90 min in 100 μg/ml of LGL1 in 0.1 M sodium bicarbonate, rinsed 3 times in PBS and 0.1% Tween 20 detergent, then blocked for 2 h with 5% bovine serum albumin (BSA) (10, 29). Samples of serum or purified monoclonal IgAs were submitted to agarose gel electrophoresis; then, the gels were blotted onto the LGL1-saturated membranes by diffusion blotting during 12 min (10, 30). After blocking for 1 h with 2.5% BSA in PBS and 0.1% Tween 20, membranes were incubated with anti-human IgA horseradish peroxidase (HRP)-conjugated secondary antibody for 1 h, then washed and revealed with Super Signal West Pico chemiluminescent substrate (Thermo Scientific).
Analysis of the Sialylation of Serum IgAs
An enzyme linked lectin assay (ELLA) was developed to analyze IgA sialylation, and an enzyme linked immuno-sorbent assay (ELISA) was used for the detection of total IgAs, as previously described (21). Ninety-six well plates (Nunc MaxiSorp™) were coated overnight at 4°C with 50 μl of goat anti-human IgA (Bethyl Laboratories, Montgomery, TX, USA) diluted 1:250 (4.0 μg/ml; ELLA) and 1:1,000 (1.0 μg/ml; ELISA) in 25 mM borate buffer pH 9. After three washes with 200 μl of PBS-Tween 0.05% (Sigma, Saint Louis, USA), 100 μl of periodic acid (5 mM) per well was added for 10 min at room temperature, protected from light. The plates were then saturated with 100 μl of 0.25% B-grade bovine gelatin (Sigma, St. Louis, MO, USA) in PBS-Tween 0.01%, at 37°C, for 2 h. After three washes, samples were diluted in PBS-Tween 0.1% and deposited in triplicate wells containing 1.25 ng of Ig for the detection of total IgA, or 2.5 ng of IgA for sialylation studies. The total IgA quantity was assessed by incubating the plates with 50 μl of HRP-coupled goat anti-human IgA diluted 1:1,000 for 1 h (Bethyl Laboratories, Montgomery, TX, USA). Sialic acid was revealed using 50 μl of biotinylated Sambucus nigra agglutinin (SNA) diluted 1:750 (2 μg/ml; Glycodiag, Orléans, France) for 90 min and then 50 μl of streptavidin HRP diluted 1:1,000 (1 μg/ml; Vector laboratories, Burlingame, CA, USA) for 1 h, at 37°C. Then, 50 μl of TMB, the chromogenic substrate for HRP (Sigma-Aldrich, St. Louis, MO, USA) was added, and the reaction was stopped by 50 μl of 0.5 M sulfuric acid, after 3 min for IgA detection, and after 5 min for sialic acid detection. Optical densities (OD) were measured using a Spark 10 M multimode microplate reader (Tecan, Männedorf, Switzerland) at 450 nm. The percentage of sialylation was calculated as follows: [SNA OD signal/IgA OD signal]/[ng IgA in SNA well/ng IgA in IgA well] × 100. In all experiments, internal controls were used to assess reproducibility.
Statistics
Data analysis was performed by GraphPad Prism 6.01 software. Patient parameters were expressed as medians and ranges, or/and means ± standard error of the mean (SEM). The Chi-2 test was used. For continuous variables, a Mann–Whitney U-test or a Kruskal–Wallis test followed by Dunn's post-hoc test was performed. The tests used are indicated in the legends of figures and tables. A value of p < 0.05 was considered statistically significant.
Study Approval
The study was promoted by the CHU of Nantes, France (# RC12 0085) with the approval of the local and national ethical committee [Comité Consultatif de Protection des Personnes dans la Recherche Biomédicale, Commission Nationale de l'Informatique et des Libertés (CNIL #912335)]. Written informed consents were obtained from patients and healthy donors, by the blood bank (Etablissement Français du Sang (EFS), Nantes, France). A convention was signed between CRCINA and EFS Pays de La Loire.
Results
Characteristics of Patients With a Monoclonal IgA
In this retrospective study, 35 patients with a monoclonal IgA were recruited (6 MGUS, 29 myeloma). Annotated clinical data were available for 26 patients (6 MGUS, 20 myeloma); the biological and clinical characteristics of patients are shown in Table 1. The male/female ratios were 33.3% for MGUS and 45.0% for myeloma, and the median age of MGUS and myeloma patients with monoclonal IgA at the time of diagnosis was 76.6 and 75.1, respectively. Thus, in this cohort, patients with IgA myeloma were older than in the series of 135 IgG myeloma patients we studied previously (median age at the time of diagnosis: 75.1 years for IgA myeloma vs. 63.8 years for IgG myeloma) (12, 21). Compared to IgG myeloma, the quantity of monoclonal Ig produced at the time of diagnosis of IgA myeloma was low (median quantity of monoclonal Ig: 17.0 g/L for IgA vs. 26.7 g/L for IgG), and the median percentage of plasma cells in the bone marrow was high: 52 vs. 33% for IgG myeloma (12). All but one patients with IgA myeloma presented with bone lesions, and the International Staging System (ISS) and Durie–Salmon Staging (DSS) scores indicated that 50.0% of patients presented with ISS stage III at the time of diagnosis (median DSS stage III: 59.1%).
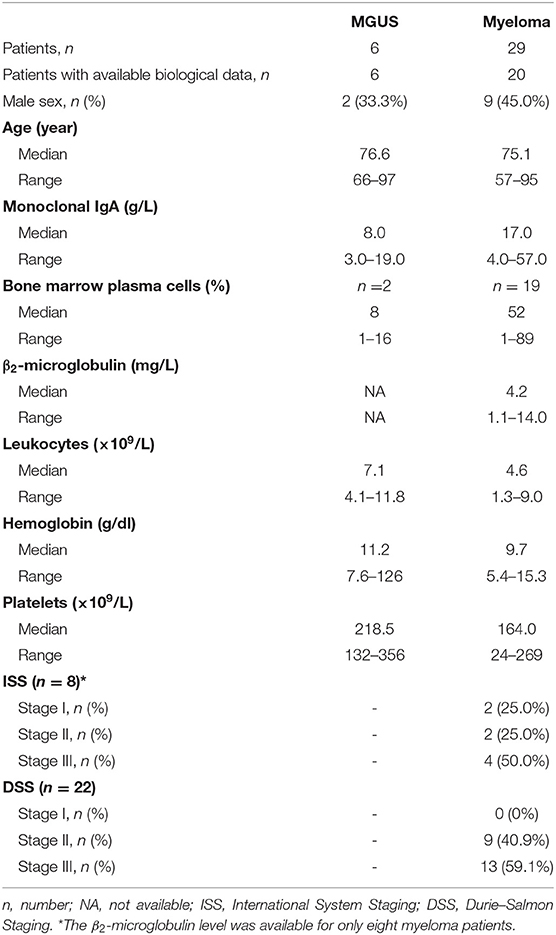
Table 1. Characteristics of patients with IgA monoclonal gammopathy of undetermined significance (MGUS) or IgA myeloma.
Serological Status of MGUS and Myeloma Patients With a Monoclonal IgA
The unseparated IgG and IgA serological status was determined for 32 patients (6 MGUS, 26 myeloma) using the MIAA, which tests for reactivity to lysates and/or antigens representing nine infectious pathogens (12, 15, 21). The MIAA assay can be revealed either with a fluorescent goat anti-human IgG Fc antibody (for IgG serology) or with a fluorescent goat anti-human IgA Fc antibody (for IgA serology), which allowed us to analyze in parallel IgG and IgA reactivity in the serum of patients (Table 2). Overall, the rates of positive IgG serology for EBV, CMV, HSV-1, T. gondii of MGUS, and myeloma patients were comparable to those of our previous studies and to those observed in the general population. The rates of positive IgG serology differed for HSV-2 (high frequency of positivity in the IgA cohort) and VZV (likely underestimated by the MIAA) (12, 15, 21). Patients also had a positive IgA serology for EBV, CMV, HSV-1, as reported for the general population (31–35). The rates of positive IgA serology were significantly lower than the rates observed for IgG serology for EBV, CMV, HSV-1 (p < 0.00001, Fisher exact test), and HSV-2 (p = 0.0272, Fisher exact test) (Table 2). They were similar to those of IgG for H. pylori and T. gondii, and increased for VZV.
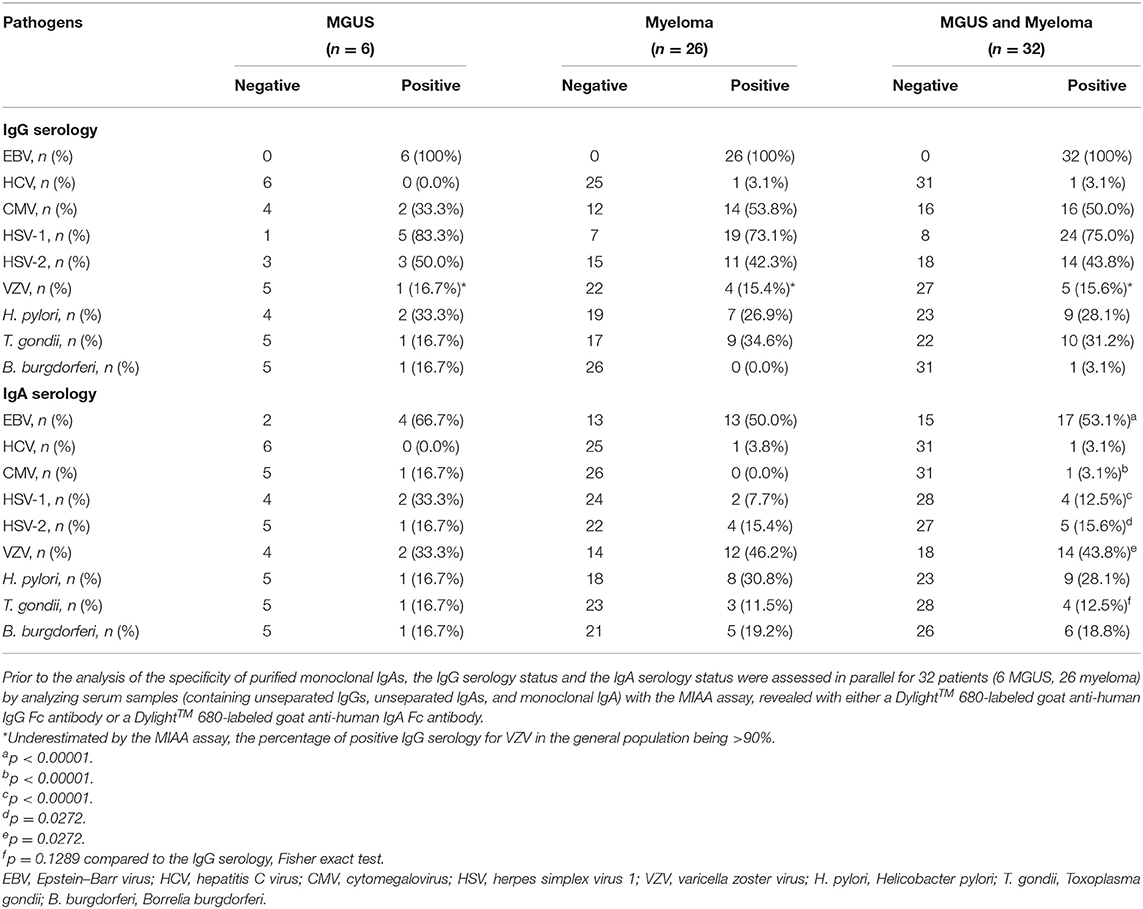
Table 2. IgG and IgA serological status of MGUS and myeloma patients with monoclonal IgA, as determined with the multiplexed infectious antigen micro-array (MIAA) assay.
Identification of the Infectious Targets of Purified Monoclonal IgAs
Monoclonal IgAs were separated individually from other Igs from blood serum, then incubated on the MIAA. In general, monoclonal IgAs were more difficult to purify than monoclonal IgGs because of their migration in agarose in the beta–gamma zone (Figure 1). Altogether, purity was achieved for 25 (71.4%) monoclonal IgAs (6 MGUS, 19 myeloma). The A1 or A2 isotype was determined for 21 monoclonal IgAs: 20 were IgA1s (5 MGUS, 15 myeloma) and 1 was an IgA2 (1 MGUS, X01). None of the monoclonal IgAs from MGUS patients recognized any infectious pathogen in the MIAA (Figure S1), whereas the purified monoclonal IgAs from five (26.3%) myeloma patients in this series specifically recognized a single recombinant protein from a single pathogen. Four recognized EBV nuclear antigen-1 (EBNA-1) (Figures 2A–E), and one recognized HCV core protein (Figure 2F). MIAA results of other myeloma patients are shown in Figure S1. Dot blots with recombinant EBNA-1 or HCV core proteins confirmed a positive reaction for the monoclonal IgAs specific for EBNA-1 (Figure 3A) or HCV core (Figure 3B). EBNA-1-specific monoclonal IgAs were then tested against an immuno-dominant B-cell public epitope sequence, PGRRPFF (EBNA-1 residues 400–406), reported to be a target for polyclonal Igs from 86.3% of the general population (36). However, we previously found that the PGRRPFF sequence was recognized by only 2/32 (6.25%) of EBNA-1-specific monoclonal IgGs (12). In this cohort of IgA myelomas, none of the four EBNA-1-specific monoclonal IgAs recognized PGRRPFF (data not shown).
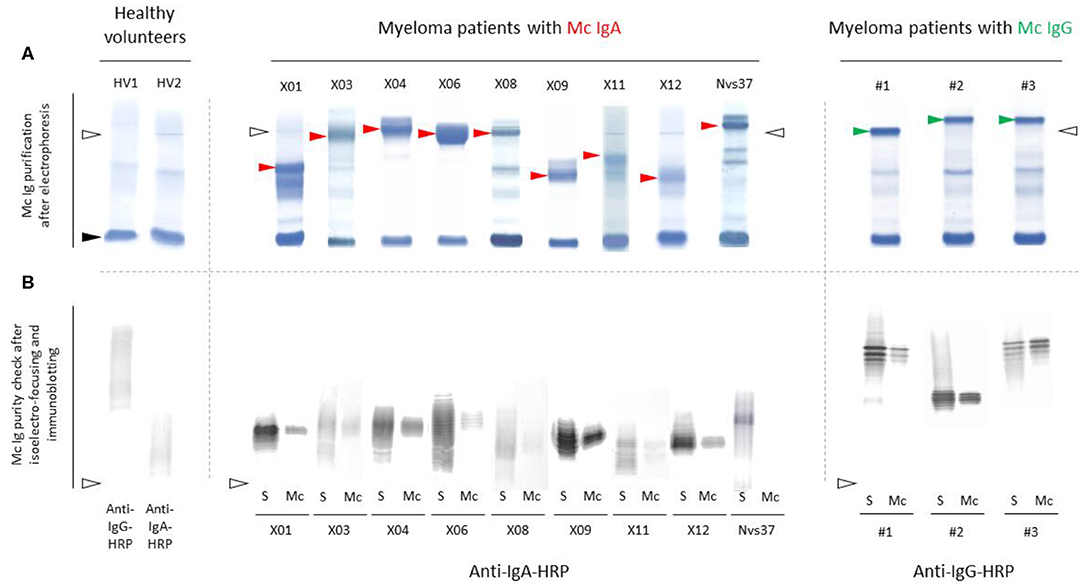
Figure 1. Separation of monoclonal IgAs from other Igs in serum. (A) Monoclonal IgAs (red arrowheads) and monoclonal IgGs used as controls (green arrowheads) were separated from other Igs in serum as published (9–11, 19). In the serum of healthy volunteers (HV), only polyclonal Igs are found; thus, no band is seen, only a smear. Note that monoclonal IgAs migrate in the β zone, are less well-separated than monoclonal IgGs, and thus are more difficult to purify than monoclonal IgGs. (B) The purity of monoclonal IgAs and IgGs was verified using isoelectric focusing (IEF) and immunoblotting. (A,B) Nine examples are shown for monoclonal IgAs and three for monoclonal IgGs (S, serum; Mc, monoclonal Ig separated from polyclonal Igs). Polyclonal Igs are represented by smears, whereas monoclonal Igs are represented by a single band in serum protein electrophoresis (A) and due to different migration according to different degrees of sialylation during IEF, as a stack of bands (B). The lines of sample deposit are indicated by white arrowheads. The albumin band is shown with a black arrowhead (A).
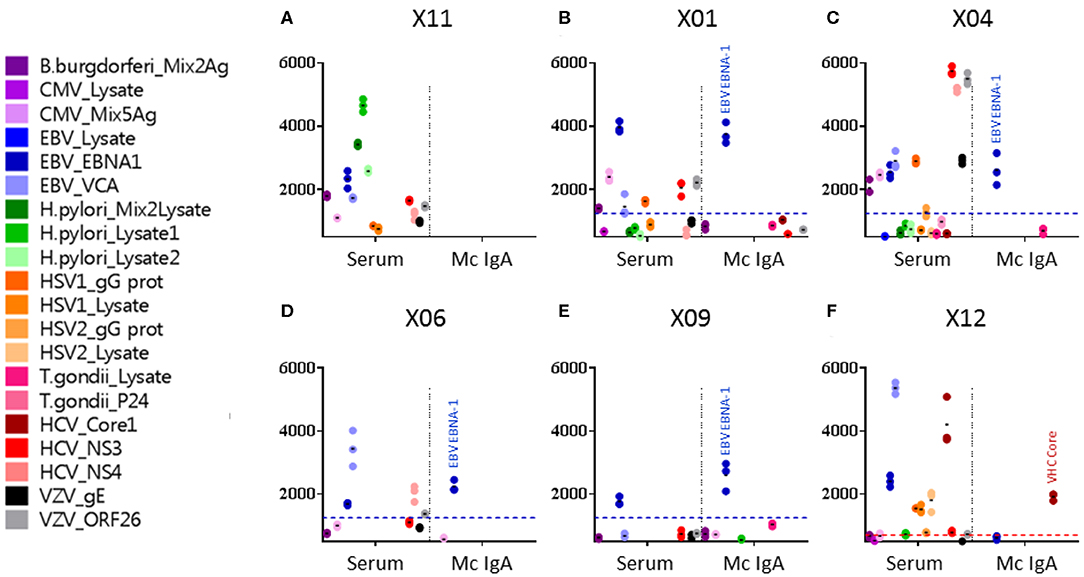
Figure 2. Viral targets of purified monoclonal IgAs from myeloma patients, as determined by the multiplexed infectious antigen micro-array (MIAA) revealed using a DylightTM 680-labeled goat anti-human IgA Fc antibody. For each patient, serum and purified monoclonal (Mc) IgA were incubated in parallel in the MIAA assay; results shown as fluorescent intensity represent either unseparated IgAs (left) or the patient's monoclonal IgA (right). (A) A patient with a Mc IgA that does not react with any pathogen of the MIAA. (B–E) Four patients with Epstein–Barr virus (EBV)-specific Mc IgAs. (F) One patient with a hepatitis C virus (HCV)-specific Mc IgA. EBV nuclear antigen (EBNA-1) signals are shown in dark blue dots, HCV core signals in red dots, and positive thresholds are shown in dotted lines. (A) For patient X11, the serum contained IgAs that recognized Borrelia burgdorferi, EBV EBNA-1, EBV VCA, Helicobacter pylori lysates 1 and 2, HCV NS3, and varicella zoster virus (VZV) ORF26 protein, whereas the purified Mc IgA did not recognize anything on the MIAA array. (B) For patient X01, the serum contained IgAs that recognized a mix of cytomegalovirus (CMV) antigens, EBV EBNA-1, EBV VCA, herpes simplex virus (HSV-1) gG, HCV NS3, and VZV ORF26; the purified Mc IgA recognized EBV EBNA-1 only. (C) For patient X04, the serum contained IgAs that recognized B. burgdorferi, CMV antigens, EBV EBNA-1, EBV VCA, HSV-1 gG, HCV NS3, HCV NS4, VZV gE, and ORF26, whereas the purified Mc IgA recognized EBV EBNA-1 only. (D) For patient X06, the serum contained IgAs that recognized EBV EBNA-1, EBV VCA, and HCV NS3; the purified Mc IgA recognized EBV EBNA-1 only. (E) For patient X09, both IgAs in serum and the purified Mc IgA recognized the EBV EBNA-1 protein only. (F) For patient X12, the serum contained IgAs that recognized EBV EBNA-1, EBV VCA, HSV-1 gG, HSV-1 lysate, HSV-2 lysate, and HCV core, whereas the purified Mc IgA recognized HCV core only. (B–F) The fluorescence values shown for EBV EBNA-1 or HCV core were obtained after subtraction of the non-specific fluorescent background. Thresholds of specific positivity were defined for each viral pathogen or protein (1,400 for EBV EBNA-1, blue threshold; 500 for HCV core, red threshold) (9, 14, 19). Note that dots may be superimposed; horizontal bars represent the means of results obtained for a pathogen, Ag, or lysate. Experiments were performed in triplicates, repeated at least once.

Figure 3. Confirmation of the specificity of recognition of EBV EBNA-1 or HCV core proteins by purified monoclonal IgAs. (A) Dot blotting assays with purified recombinant EBNA-1 were performed in parallel with PBS, as control (CTRL), and with the serum and the purified monoclonal IgA from six patients. As assessed by the MIAA array, both serum and purified monoclonal IgAs of the patient X05 did not recognize EBV EBNA-1, and only unseparated IgAs from the serum of the patient X11 recognized EBNA-1 (negative controls). For patients X01, X04, X06, and X09, both serum and purified monoclonal IgAs recognized EBV EBNA-1, thus confirming the results obtained with the MIAA array. (B) A dot blotting assay with purified recombinant HCV core protein was performed in parallel with PBS, as control (CTRL), and with the serum and purified monoclonal IgA of patients. As assessed by the MIAA array, both the serum and the purified monoclonal IgA of patient X08 did not recognize the HCV core (negative control). For patient X12, both the serum and the purified monoclonal IgA recognized the HCV core, confirming the results obtained with the MIAA array. Experiments were performed at least twice.
Identification of Monoclonal IgAs That Target LGL1
Purified monoclonal IgAs from MGUS and myeloma patients were analyzed with the LGL1 assay adapted from Nair et al. (10). As shown in Figure 4, only three monoclonal IgAs reacted with LGL1, and all were from myeloma patients (X03, X08, Nvs37). The monoclonal IgA of each of the three patients did not recognize any pathogen of the MIAA assay (their MIAA assay was negative, an indirect proof of purity of the monoclonal IgA preparation). Thus, 3/19 (15.8%) of myeloma monoclonal IgA targeted LGL1 in this series.
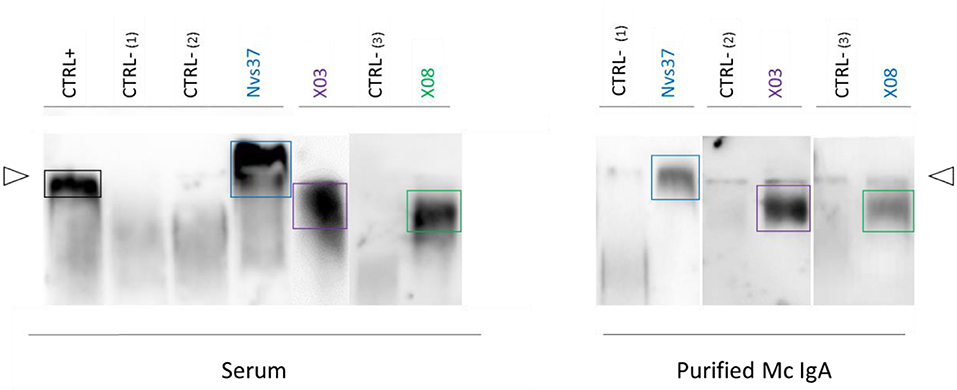
Figure 4. Lysoglucosylceramide (LGL1) is specifically recognized by subsets of purified monoclonal IgAs. LGL1-specific immunoblotting assays were performed as described in the Materials and Methods section (7, 27, 28). Samples of serum (left) or purified monoclonal IgAs (right) were first submitted to agarose gel electrophoresis; then, the gels were blotted onto LGL1-saturated membranes. After blocking for 1 h, membranes were incubated with anti-human IgA horseradish peroxidase (HRP)-conjugated secondary antibody, then washed and revealed by chemiluminescence. The positive control (CTRL+, left) was a sample of serum from a patient known to have LGL1-specific IgAs. Negative controls (CTRL–) were samples of serum without LGL1-reactive IgAs (one from a healthy volunteer, two from patients). The lines of sample deposit are indicated by white arrowheads. The positive signals characteristic of LGL1-reactive Igs are encircled. Patterns of migration may differ for serum and purified monoclonal IgAs because serum may contain both monoclonal and polyclonal LGL1-reactive IgAs.
Characteristics of Myeloma Patients With a Monoclonal IgA Specific for EBNA1 or LGL1
We compared the clinical characteristics of the four myeloma patients with EBNA-1-specific monoclonal IgA [“EBNA-1(+)” patients] with those of the 17 myeloma patients with a monoclonal IgA that was non-reactive for pathogens of the MIAA assay [“MIAA(–)” patients] (Table 3). Compared to other IgA myeloma patients, myeloma patients with EBNA-1-specific IgA were relatively young at the time of diagnosis (≤63 years old), as reported for myeloma patients with EBNA-1-specific monoclonal IgG (12). Regarding patients with an LGL1-specific monoclonal IgA, Nair et al. reported a tendency toward a mild form of disease for LGL1-associated myeloma (10). Here, clinical and biological characteristics were available for only two patients. The data did not suggest a mild disease since both had bone lesions: one had >50% plasma cells in the bone marrow, and one had a DSS stage III (Table 4).
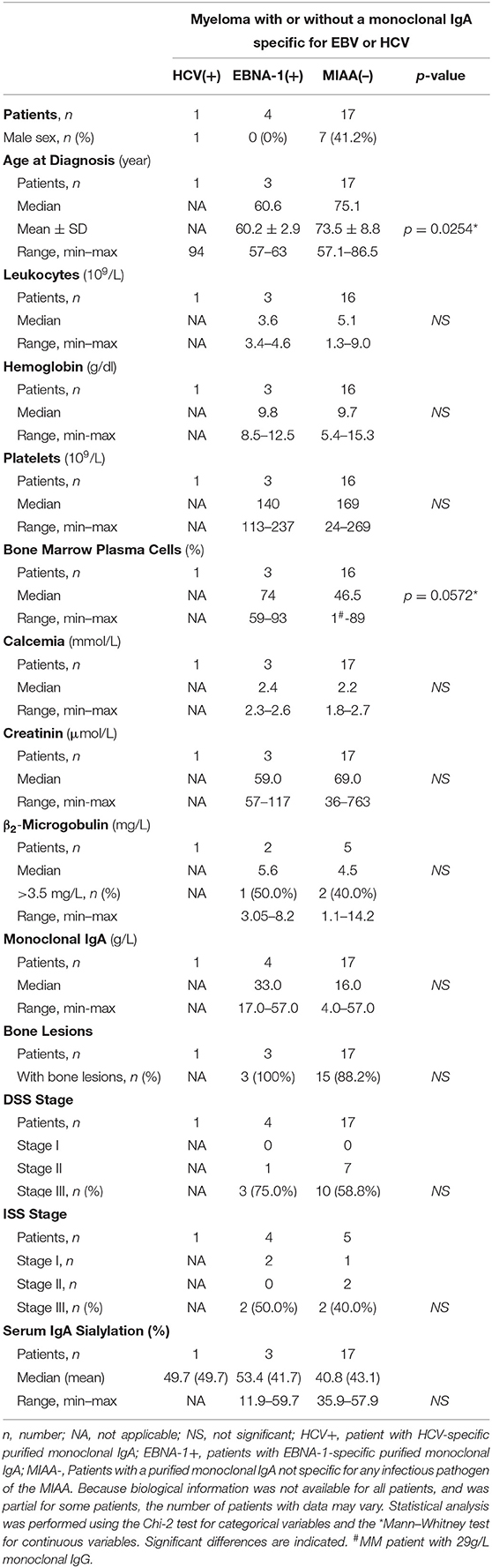
Table 3. Characteristics of myeloma patients with a monoclonal IgA specific for EBV nuclear antigen 1 (EBNA-1).
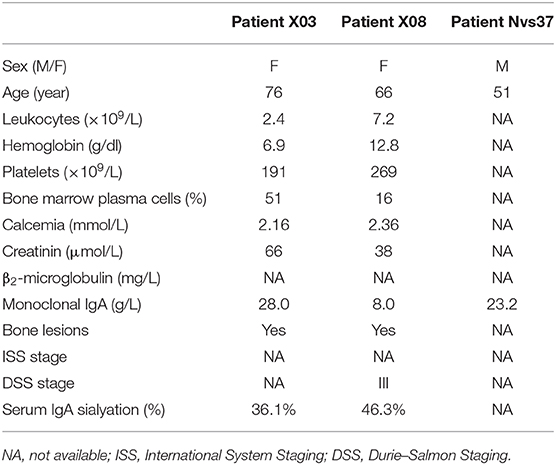
Table 4. Characteristics of myeloma patients with a lysoglucosylceramide (LGL1)-specific monoclonal IgA.
Glycosylation of IgAs
As published for monoclonal IgGs, the sialylation level of unseparated, total IgAs, assessed in serum, expressed in % sialylation, was lower for MGUS and myeloma patients with a monoclonal IgA than for healthy volunteers (41.2 vs. 63.4%, respectively; ***p < 0.0001, Mann–Whitney U-test) (Figure 5A) (12, 21). No difference was observed according to age (healthy donors under or over 60 years), nor between MGUS and myeloma (Figure 5B). In addition, there was no difference in IgA sialylation depending on the antigenic specificity (MIAA+ vs. MIAA–) of the monoclonal IgA (Figure 5C, Table 4). However, due to the small size of the cohorts, these results will need to be confirmed in larger studies.
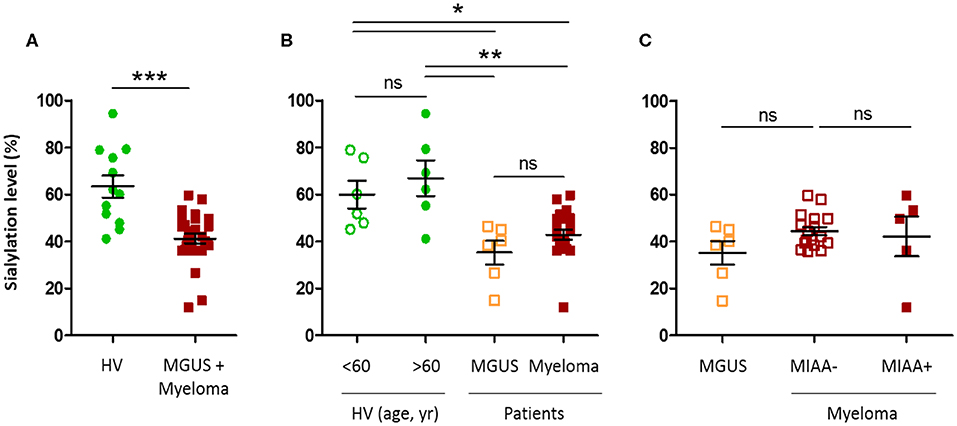
Figure 5. Hyposialylation of IgAs from monoclonal gammopathy of undetermined significance (MGUS) and myeloma patients. The sialylation level of IgAs in the serum of healthy volunteers (HV, n = 12), MGUS (n = 6), and myeloma (n = 22) patients was assessed using an enzyme-linked lectin assay (ELLA) technique, as described in the Materials and Methods section. Unfortunately, we were not able to constitute a control cohort of patients with excessive amounts of non-clonal IgAs. Results are expressed as percentages of sialylated forms of IgAs in serum. (A) Sialylation level of IgAs from HV (green dots) and MGUS and myeloma (MM) patients (red dots); ***p < 0.001, Mann–Whitney U-test. (B) Sialylation level according to age, in HV under or over 60 (green dots), and according to the diagnosis of MGUS (orange squares) or MM (red squares). (C) Sialylation level of IgAs from MM patients with a pathogen-specific monoclonal IgA, as determined by the MIAA (filled red squares, MIAA+) and MM patients with a monoclonal IgA of undetermined specificity (open red squares, MIAA–), compared to MGUS patients (orange squares). Bars indicate means±SEM. (B,C) Significant differences are indicated by stars; *p < 0.05 and **p < 0.01, Kruskal–Wallis test followed by Dunn's post-hoc test (ns, not significant).
Discussion
This is the first analysis of the antigenic origins of IgA myeloma. In this small cohort, no antigenic target was identified for the monoclonal IgAs from MGUS patients, but for patients with IgA myeloma, two viruses (EBV, HCV) were candidate targets for the monoclonal IgA of 5/19 patients (26.3%). In addition, the monoclonal IgA of 3/19 myeloma patients (15.8%) reacted with LGL1, a glucolipidic auto-antigen. These results imply that chronic stimulation by viral antigens or auto-antigen LGL1 may underlie the initiation of ~40% of IgA myeloma, as reported for MGUS and myeloma with a monoclonal IgG (10, 12).
In this study, myeloma patients with a monoclonal IgA were typically older and had stage III disease, consistent with IgA myeloma being more severe than IgG myeloma (22–28). Analysis of the IgG and IgA serology status of MGUS and myeloma patients revealed that despite stage III disease, patients maintained detectable levels of polyclonal IgGs and IgAs directed against common pathogens. The rates of positive IgG serology against EBV, CMV, HSV-1, and HSV-2 observed for IgA myeloma patients were similar to those of the general population. The rates of positive IgA serology of these patients were lower than those of IgG for EBV, CMV, HSV-1, and HSV-2, as expected (31–35). They were similar to the rates of positive IgG serology for H. pylori, T. gondii, B. burgdorferi, and higher for VZV. Thus, this cohort of patients with IgA myeloma maintained IgG- and/or IgA-mediated protection against EBV, CMV, HSV-1, HSV-2, and VZV.
Two viral proteins, EBV EBNA-1 and HCV core, were specifically recognized by monoclonal IgAs from certain myeloma patients; these proteins are also the targets of monoclonal IgGs in both MGUS and myeloma (12–15). Because IgAs are linked with the digestive tract and other mucosal tissues, it is not surprising that monoclonal IgAs reacted with pathogens found in mucosal or digestive and hepatic tissues (HCV) or in saliva (EBV) (31, 35, 37). The PGRRPFF sequence, identified as a frequent EBNA-1 epitope for the general population, was not recognized by EBNA-1-reactive monoclonal IgAs, an observation also made for EBNA-1-reactive monoclonal IgGs (12, 36). It will be important to determine whether the amino acid sequences targeted by monoclonal Igs from myeloma patients differ from those of Igs from healthy individuals. Future characterization of the EBNA-1 sequences recognized by monoclonal IgGs and IgAs should help determine whether certain epitope “hotspots” are overrepresented in the BCR specificity of malignant B cells in myeloma. For instance, knowing the viral sequences linked to MGUS and myeloma would permit their elimination from the future EBV vaccines in development (38, 39). In addition, for 15.8% of patients with IgA myeloma in this cohort, the purified monoclonal IgA reacted with LGL1, a self-antigen initially described as a target of monoclonal Igs in the context of Gaucher disease (10, 11). In future studies, it would be of interest to determine whether MGUS and myeloma patients with an anti-LGL1 monoclonal Ig present a mild, unsuspected metabolic deficiency resulting in sphingolipid accumulation.
Altogether, our findings suggest that for a significant fraction (>30%) of patients with IgA or IgG myeloma, the initial cause of disease may be chronic antigen stimulation due to a viral infection (particularly by EBV or HCV) or autoimmunity against LGL1 (10–14). These observations may be compared to the ~50% chronic lymphocytic leukemia (CLL) where the malignant clone displays somatically mutated Ig heavy (H) chain variable (IGHV) genes, indicative of antigen-driven disease (40, 41). Importantly, antigen-driven disease may be associated with a distinct prognosis: patients with antigen-driven CLL seem to have a more favorable clinical course than other CLL patients, whereas myeloma patients with EBNA-1-associated myeloma tend to present with more severe disease (12, 42, 43). In this small cohort, myeloma patients with EBNA-1-specific monoclonal IgA were relatively young at diagnosis (≤63 years), with severe (59–93%) plasma cell infiltration of the bone marrow, characteristics similar to those reported for myeloma patients with EBNA-1-specific monoclonal IgG (12). Regarding LGL1-associated myeloma, Nair et al. suggested that it may represent a mild form of myeloma (10). In the present study, the two myeloma patients with LGL1-specific IgA did not have mild disease. Studies performed on large cohorts of well-annotated patients (with cytometry, cytogenetics, genetic data) are necessary for the full characterization of myeloma linked to LGL-1 or EBV EBNA-1.
The antigenic targets of malignant clones of B-cell lineage have been studied in the context of CLL, using different technical approaches (phage-display technology, mass spectrometry). Several auto-antigens have been associated with CLL, notably cytoskeleton components (non-muscle myosin heavy chain IIA, vimentin, cofilin-1, filamin B), cardiolipin, proline-rich acidic protein-1 (PRAP-1), dUTPase, and auto-antigens at the surface of apoptotic cells and bacteria (Streptococcus pneumoniae for instance) (44–51). Evidence of virus (HCV, HIV)-driven CLL has been less reported (52–54). Regarding EBV, EBV DNA is typically not detected in malignant CLL or myeloma cells (20). The variability of EBV DNA loads in blood and the patterns of anti-EBV Ig responses of patients have been well-analyzed in CLL, but the findings of these studies appear to mostly reflect the deficient immune system of aged CLL patients (55–58). Thus, formal evidence of EBV antigen-driven CLL disease is still lacking, and identified CLL-associated antigens are predominantly auto-antigens linked to bacterial infection and/or apoptotic cell removal, and to a lesser degree, viral antigens (44, 45, 52–54). In contrast, the most frequent antigenic targets associated so far with MGUS and myeloma are viral proteins (especially from EBV, HSV, HCV) and a ganglioside, LGL1 (10–14). Of note, several groups reported that gangliosides facilitate cell entry of viruses (59). Knowing whether anti-LGL1 monoclonal Igs can counter virus cell entry would be of interest.
IgA glycosylation was also analyzed. Monoclonal IgAs differed from IgAs from healthy donors by their low level of sialylation, a characteristic observed for monoclonal IgGs and associated with a pro-inflammatory action of the Ig Fc fragment upon binding to FcγR, notably in monocytes and macrophages (21). However, sialylation did not differ depending on the antigenic specificity of the monoclonal IgA. Further studies are needed to determine whether monoclonal IgAs from MGUS and myeloma patients contribute to the production of pro-inflammatory cytokines, as reported for patients with hyposialylated IgGs (polyclonal or/and monoclonal) (21).
In aging populations, the incidence of MGUS and the subsequent risk of myeloma and other MGUS-associated diseases increase (60, 61). The detection of antigen-initiated MGUS and myeloma cases, and the determination of the antigenic target of the monoclonal Ig, should be useful additions to the diagnostic work-up of MGUS and myeloma because they allow new possibilities of prevention and treatment. First, disease-initiating antigenic targets could serve as new risk markers. Second, MGUS patients, who are not treated presently, and myeloma patients could benefit from antigen-reduction treatments. Supporting this approach, several groups reported that the addition of anti-viral treatment to myeloma protocols resulted in disease regression and/or improved response to chemotherapy, notably for HCV-associated myeloma (62, 63). Drugs that target BCR signaling may also be considered (64, 65). Moreover, new drugs are currently being developed that specifically target EBV (66, 67). Clearly, if one could clear the MGUS-associated underlying chronic infection early on, it may be possible to prevent the development of myeloma (68). Regarding myeloma patients with an LGL1-specific monoclonal Ig, reduction of LGL1 levels may be envisioned as a complementary treatment. Indeed, LGL1 reduction has been successfully achieved in Gaucher patients for many years (69–73). Importantly, glucolipid reduction prevents associated B-cell malignancies in murine models (74, 75). Recently, Nair et al. reported that glucolipid reduction treatment resulted in decreased amount of monoclonal Ig in Gaucher patients with monoclonal gammopathy (76).
In conclusion, EBV EBNA-1, the HCV core protein, and LGL1, a glucolipid, were identified as candidate antigenic targets of the purified monoclonal Igs of patients with IgA myeloma. An abnormal immune response to these viruses or to LGL1 may therefore be part of the pathogenesis of IgA myeloma, as reported for IgG myeloma. Detecting patients who present with LGL1- or virus-associated MGUS or myeloma is important since it is possible to add antigen target reduction to classic treatments.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
The studies involving human participants were reviewed and approved by Local ethics committee, CHU Nantes. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
SH, AB, EB-C, and JH designed the research, analyzed the data, and wrote the manuscript. AB, CS, NM, SA-M, MF, EB-C, and JH performed the experiments and edited the manuscript. AT, EP, PL, and FM contributed patient samples and data. All approved the version to be submitted for publication and agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the article are appropriately investigated and resolved.
Funding
The study has been supported by grants to SH from the Ligue Nationale contre le Cancer, the Cancéropôle Grand Ouest and Région Pays de la Loire (2015–2016), and Janssen USA. EP benefited from a grant from the Cancéropôle Grand Ouest and Région Centre (2015–2016). The salary of AB was supported by the Cancéropôle Grand Ouest/Région Pays de la Loire (HII-GO project, June 2015 to December 2016), then Janssen USA (January 2017 to December 2017). The salary of NM was supported by Janssen USA (September 2016 to April 2019).
Conflict of Interest
The authors declare that this study received funding from Janssen (USA). Janssen had no role in study design, data collection and analysis, or preparation of the manuscript.
Acknowledgments
We thank all the colleagues from the Departments of Hematology or Internal Medicine of the University Hospitals in Tours and Bordeaux, who contributed to the diagnosis and care of patients in this study, and Novartis (Basel, Switzerland) for providing seven IgA myeloma serum samples. We also thank Dr. Pierre Weigel and Dr. Cathy Charlier (CNRS UMR6286) and the IMPacT platform (impact-plateforme.com) at the University of Nantes, in Nantes, France.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.00854/full#supplementary-material
References
1. Kyle RA. Monoclonal gammopathy of undetermined significance. Natural history in 241 cases. Am J Med. (1978) 64:814–26. doi: 10.1016/0002-9343(78)90522-3
2. Kyle RA, Therneau TM, Rajkumar SV, Offord JR, Larson DR, Plevak MF, et al. A long-term study of prognosis of monoclonal gammopathy of undetermined significance. N Engl J Med. (2002) 346:564–9. doi: 10.1056/NEJMoa01133202
3. Dhodapkar MV. MGUS to myeloma: a mysterious gammopathy of underexplored significance. Blood. (2016) 128:2599–606. doi: 10.1182/blood-2016-09-692954
4. Kyle RA, Larson DR, Therneau TM, Dispenzieri A, Kumar S, Cerhan JR, et al. Long-term follow-up of monoclonal gammopathy of undetermined significance. New Engl J Med. (2018) 378:241–9. doi: 10.1056/NEJMoa1709974
5. Kyle RA, Greipp PR. Smoldering multiple myeloma. N Engl J Med. (1980) 302:1347–9. doi: 10.1056/NEJM198006123022405
6. Rajkumar SV, Dimopoulos MA, Palumbo A, Blade J, Merlini G, Mateos MV, et al. International myeloma working group updated criteria for the diagnosis of multiple myeloma. Lancet Oncol. (2014) 15:e538–48. doi: 10.1016/S1470-2045(14)70442-5
7. Bergón E, Miravalles E. Retrospective study of monoclonal gammopathies detected in the clinical laboratory of a Spanish healthcare district: 14-year series. Clin Chem Lab Med. (2007) 45:190–6. doi: 10.1515/CCLM.2007.029
8. Vescio RA, Cao J, Hong CH, Lee JC, Wu CH, Der Danielian M, et al. Myeloma Ig heavy chain V region sequences reveal prior antigenic selection and marked somatic mutation but no intraclonal diversity. J Immunol. (1995) 155:2487–97.
9. Sahota SS, Leo R, Hamblin TJ, Stevenson FK. Myeloma VL and VH gene sequences reveal a complementary imprint of antigen selection in tumor cells. Blood. (1997) 89:219–26. doi: 10.1182/blood.V89.1.219.219_219_226
10. Nair S, Branagan AR, Liu J, Boddupalli CS, Mistry PK, Dhodapkar M V. Clonal immunoglobulin against lysolipids in the origin of myeloma. N Engl J Med. (2016) 374:555–61. doi: 10.1056/NEJMoa1508808
11. Nair S, Sng J, Sekhar Boddupalli C, Seckinger A, Chesi M, Fulciniti M, et al. Antigen-mediated regulation in monoclonal gammopathies and myeloma. J Clin Invest Insight. (2018) 3:e98259. doi: 10.1172/jci.insight.98259
12. Bosseboeuf A, Feron D, Tallet A, Rossi C, Charlier C, Garderet L, et al. Monoclonal IgG in MGUS and multiple myeloma target infectious pathogens. J Clin Invest Insight. (2017) 2:e95367. doi: 10.1172/jci.insight.95367
13. Hermouet S, Corre I, Gassin M, Bigot-Corbel E, Sutton CA, Casey JW. Hepatitis C virus, human herpesvirus 8, and the development of plasma-cell leukemia. N Engl J Med. (2003) 348:178–9. doi: 10.1056/NEJM200301093480219
14. Bigot-Corbel E, Gassin M, Corre I, Le Carrer D, Delaroche O, Hermouet S. Hepatitis C virus (HCV) infection, monoclonal immunoglobulin specific for HCV core protein, and plasma-cell malignancy. Blood. (2008) 112:4357–8. doi: 10.1182/blood-2008-07-167569
15. Feron D, Charlier C, Gourain V, Garderet L, Coste-Burel M, Le Pape P, et al. Multiplexed infectious protein microarray immunoassay suitable for the study of the specificity of monoclonal immunoglobulins. Anal Biochem. (2013) 433:202–9. doi: 10.1016/j.ab.2012.10.012
16. Plummer M, de Martel C, Vignat J, Ferlay J, Bray F, Franceschi S. Global burden of cancers attributable to infections in 2012: a synthetic analysis. Lancet Glob Health. (2016) 4:e609–16. doi: 10.1016/S2214-109X(16)30143-7
17. Seifert M, Scholtysik R, Küppers R. Origin and pathogenesis of B cell lymphomas. Methods Mol Biol. (2013) 971:1–25. doi: 10.1007/978-1-62703-269-8_1
18. Saha A, Robertson ES. Epstein-Barr virus-associated B-cell lymphomas: pathogenesis and clinical outcomes. Clin Cancer Res. (2011) 17:3056–63. doi: 10.1158/1078-0432.CCR-10-2578
19. Franceschi S, Lise M, Trépo C, Berthillon P, Chuang SC, Nieters A, et al. Infection with hepatitis B and C viruses and risk of lymphoid malignancies in the European prospective investigation into cancer and nutrition (EPIC). Cancer Epidemiol Biomarkers Prev. (2011) 20:208–14. doi: 10.1158/1055-9965.EPI-10-0889
20. Hermouet S, Sutton CA, Rose TM, Greenblatt RS, Corre I, Garand R, et al. Qualitative and quantitative analysis of human herpesvirus expression in chronic and acute B-cell lymphocytic leukemia and in multiple myeloma. Leukemia. (2003) 17:185–95. doi: 10.1038/sj.leu.2402748
21. Bosseboeuf A, Allain S, Mennesson N, Tallet A, Rossi C, Garderet L, et al. Pro-inflammatory state in MGUS and myeloma is characterized by low sialylation of pathogen-specific and other monoclonal and polyclonal immunoglobulin G. Front Immunol. (2017) 8:1347. doi: 10.3389/fimmu.2017.01347
22. Nair B, Waheed S, Szymonifka J, Shaughnessy JD, Crowley J Jr, Barlogie B. Immunoglobulin isotypes in multiple myeloma: laboratory correlates and prognostic implications in total therapy protocols. Br J Haematol. (2009) 145:134–7. doi: 10.1111/j.1365-2141.2008.07547.x
23. Krejci M, Hajek R, Buchler T, Krivanova A, Svobodnik A, Pour L, et al. Simple variables predict survival after autologous transplantation: a single centre experience in 181 multiple myeloma patients. Neoplasma. (2007) 54:143–8.
24. Pasqualetti P, Colantonio D, Collacciani A, Casale R, Natali G. Classification and prognostic evaluation in multiple myeloma. A retrospective study of relationship of survivals and responses to chemotherapy to immunological types, 20 single prognostic factors, 15 clinical staging systems, and 6 morphological classifications. Panminerva Med. (1991) 33:93–110.
25. Wang L, Jin F-Y, Li Y, Sun J-N, Zhang J-J, Tang R, et al. IgA type multiple myeloma, clinical features, and prognosis. Chinese Med J. (2018) 131:1249–50. doi: 10.4103/0366-6999.231513
26. Gemenetzi K, Agathangelidis A, Papalexandri A, Medina A, Genuardi E, Moysiadis T, et al. Distinct immunogenetic signatures in IgA versus IgG multiple myeloma. 58th American Society of Hematology (ASH) Annual Meeting & Exposition. San Diego, CA: Poster (2016) doi: 10.1182/blood.V128.22.2062.2062
27. Roberts-Thomson PJ, Mason DY, MacLennan ICM. Relationship between paraprotein clinical features in IgA polymerization and myeloma. Br J Haematol. (1976) 33:117–30. doi: 10.1111/j.1365-2141.1976.tb00978.x
28. Kyle RA, Remstein ED, Therneau TM, Dispenzieri A, Kurtin PJ, Hodnefield JM, et al. Clinical course and prognosis of smoldering (asymptomatic) multiple myeloma. New Engl J Med. (2007) 356:2582–90. doi: 10.1056/NEJMoa070389
29. Nooij FJ, Van der Sluijs-Gelling AJ, Jol-Van der Zijde CM, Van Tol MJ, Haas H, Radl J. Immunoblotting techniques for the detection of low level homogeneous immunoglobulin components in serum. J Immunol Methods. (1990) 134:273–81. doi: 10.1016/0022-1759(90)90389-D
30. Braun W, Abraham R. Modified diffusion blotting for rapid and efficient protein transfer with PhastSystem. Electrophoresis. (1989) 10:249–53. doi: 10.1002/elps.1150100406
31. Hadar T, Rahima M, Kahan E, Sidi J, Rakowsky E, Sarov B, et al. Significance of specific epstein-barr virus IgA and elevated IgG antibodies to viral capsid antigens in nasopharyngeal carcinoma patients. J Med Virol. (1986) 20:329–39. doi: 10.1002/jmv.1890200405
32. Levy E, Margalith M, Sarov B, Sarov I, Rinaldo CR, Detels R, et al. Cytomegalovirus IgG and IgA serum antibodies in a study of HIV infection and HIV-related diseases in homosexual men. J Med Virol. (1991) 35:174–9. doi: 10.1002/jmv.1890350306
33. Friedman MG, Kimmel N. Herpes Simplex virus-specific serum immunoglobulin a: detection in patients with primary or recurrent herpes infections and in healthy adults. Infect Immun. (1982) 1:374–7. doi: 10.1128/IAI.37.1.374-377.1982
34. Wittek AE, Arvin AM, Koropchak CM. Serum immunoglobulin A antibody to varicella-zoster virus in subjects with primary varicella and herpes zoster infections and in immune subjects. J Clin Microbiology. (1983) 18:1146–9. doi: 10.1128/JCM.18.5.1146-1149.1983
35. Urita Y, Hike K, Torii N, Kikuchi Y, Kurakata H, Kanda E, et al. Comparison of serum IgA and IgG antibodies for detecting helicobacter pylori infection. Int Med. (2004) 43:548–52. doi: 10.2169/internalmedicine.43.548
36. Xu GJ, Kula T, Xu Q, Li MZ, Vernon SD, Ndung'u T, et al. Viral immunology. Comprehensive serological profiling of human populations using a synthetic human virome. Science. (2015) 348:aaa0698. doi: 10.1126/science.aaa0698
37. Gonzàlez-Quintela A, Alende MR, Gamallo R, Gonzàlez-Gil P, López-Ben S, Tomé S, et al. Serum immunoglobulins (IgG, IgA, IgM) in chronic hepatitis C. A comparison with non-cirrhotic alcoholic liver disease. Hepatogastroenterol. (2003) 54:2121–6.
38. Cohen JI. Epstein-Barr virus vaccines. Clin Translat Immunol. (2015) 4:e32. doi: 10.1038/cti.2014.27
39. Cohen JI. Vaccine development for epstein-barr virus. Adv Exp Med Biol. (2018) 1045:477–93. doi: 10.1007/978-981-10-7230-7_22
40. Fais F, Ghiotto F, Hashimoto S, Sellars B, Valetto A, Allen SL, et al. Chronic lymphocytic leukemia B cells express restricted sets of mutated and unmutated antigen receptors. J Clin Invest. (1998) 102:1515–25. doi: 10.1172/JCI3009
41. Henriques A, Rodríguez-Caballero A, Criado I, Langerak AW, Nieto WG, Lécrevisse Q, et al. Molecular and cytogenetic characterization of expanded B-cell clones from multiclonal versus monoclonal B-cell chronic lymphoproliferative disorders. Haematologica. (2014) 99:897–907. doi: 10.3324/haematol.2013.098913
42. Damle RN, Wasil T, Fais F, Ghiotto F, Valetto A, Allen SL, et al. Ig V gene mutation status and CD38 expression as novel prognostic indicators in chronic lymphocytic leukemia. Blood. (1999) 94:1840–7. doi: 10.1182/blood.V94.6.1840.418k06_1840_1847
43. Hamblin TJ, Davis Z, Gardiner A, Oscier DG, Stevenson FK. Unmutated Ig VH genes are associated with a more aggressive form of chronic lymphocytic leukemia. Blood. (1999) 94:1848–54. doi: 10.1182/blood.V94.6.1848.418k05_1848_1854
44. Chiorazzi N, Hatzi K, Albesiano E. B-cell chronic lymphocytic leukemia, a clonal disease of B lymphocytes with receptors that vary in specificity for (auto)antigens. Ann N Y Acad Sci. (2005) 1062:1–12. doi: 10.1196/annals.1358.002
45. Karp M, Giannopoulos K. Antigen stimulation in the development of chronic lymphocytic leukemia. Postepy Hig Med Dosw. (2013) 67:1204–13. doi: 10.5604/17322693.1078852
46. Catera R, Silverman GJ, Hatzi K, Seiler T, Didier S, Zhang L, et al. Chronic lymphocytic leukemia cells recognize conserved epitopes associated with apoptosis and oxidation. Mol Med. (2008) 14:665–74. doi: 10.2119/2008-00102.Catera
47. Chu CC, Catera R, Hatzi K, Yan XJ, Zhang L, Wang XB, et al. Chronic lymphocytic leukemia antibodies with a common stereotypic rearrangement recognize nonmuscle myosin heavy chain IIA. Blood. (2008) 112:5122–9. doi: 10.1182/blood-2008-06-162024
48. Lanemo Myhrinder A, Hellqvist E, Sidorova E, Söderberg A, Baxendale H, Dahle C, et al. A new perspective: molecular motifs on oxidized LDL, apoptotic cells, and bacteria are targets for chronic lymphocytic leukemia antibodies. Blood. (2008) 111:3838–48. doi: 10.1182/blood-2007-11-125450
49. Seiler T, Woelfle M, Yancopoulos S, Catera R, Li W, Hatzi K, et al. Characterization of structurally defined epitopes recognized by monoclonal antibodies produced by chronic lymphocytic leukemia B cells. Blood. (2009) 114:3615–24. doi: 10.1182/blood-2009-01-197822
50. Rosén A, Murray F, Evaldsson C, Rosenquist R. Antigens in chronic lymphocytic leukemia–implications for cell origin and leukemogenesis. Semin Cancer Biol. (2010) 20:400–9. doi: 10.1016/j.semcancer.2010.09.004
51. Williams M, Ariza ME. EBV positive diffuse large B cell lymphoma and chronic lymphocytic leukemia patients exhibit increased Anti-dUTPase antibodies. Cancers (Basel). (2018) 10:129. doi: 10.3390/cancers10050129
52. Casabonne D, Almeida J, Nieto WG, Romero A, Fernández-Navarro P, Rodriguez-Caballero A, et al. Common infectious agents and monoclonal B-cell lymphocytosis: a cross-sectional epidemiological study among healthy adults. PLoS ONE. (2012) 7:e52808. doi: 10.1371/journal.pone.0052808
53. Arcaini L, Merli M, Volpetti S, Rattotti S, Gotti M, Zaja F. Indolent B-cell lymphomas associated with HCV infection: clinical and virological features and role of antiviral therapy. Clin Dev Immunol. (2012) 2012:638185. doi: 10.1155/2012/638185
54. Hwang K-K, Trama AM, Kozink DM, Chen X, Wiehe K, Cooper AJ, et al. IGHV1-69 B cell chronic lymphocytic leukemia antibodies cross-react with HIV-1 and hepatitis C virus antigens as well as intestinal commensal bacteria. PLoS ONE. (2014) 9 :e90725. doi: 10.1371/journal.pone.0090725
55. Visco C, Falisi E, Young KH, Pascarella M, Perbellini O, Carli G, et al. Epstein-Barr virus DNA load in chronic lymphocytic leukemia is an independent predictor of clinical course and survival. Oncotarget. (2015) 6:18653–63. doi: 10.18632/oncotarget.4418
56. Teras LR, Rollison DE, Pawlita M, Michel A, Brozy J, de Sanjose S, et al. Epstein-Barr virus and risk of non-Hodgkin lymphoma in the cancer prevention study-II and a meta-analysis of serologic studies. Int J Cancer. (2015) 136:108–16. doi: 10.1002/ijc.28971
57. Casabonne D, Benavente Y, Robles C, Costas L, Alonso E, Gonzalez-Barca E, et al. Aberrant epstein-barr virus antibody patterns and chronic lymphocytic leukemia in a Spanish multicentric case-control study. Infect Agent Cancer. (2015) 10:5. doi: 10.1186/1750-9378-10-5
58. Ferrajoli A, Ivan C, Ciccone M, Shimizu M, Kita Y, Ohtsuka M, et al. Epstein-barr virus micrornas are expressed in patients with chronic lymphocytic leukemia and correlate with overall survival. EBioMedicine. (2015) 2:572–82. doi: 10.1016/j.ebiom.2015.04.018
59. Martínez MA, López S, Arias CF, Isa P. Gangliosides have a functional role during rotavirus cell entry. J Virol. (2013) 87:1115–22. doi: 10.1128/JVI.01964-12
60. Therneau TM, Kyle RA, Melton LJ 3rd, Larson DR, Benson JT, Colby CL, et al. Incidence of monoclonal gammopathy of undetermined significance and estimation of duration before first clinical recognition. Mayo Clin Proc. (2012) 87:1071–9. doi: 10.1016/j.mayocp.2012.06.014
61. Bida JP, Kyle RA, Therneau TM, Melton LJ 3rd, Plevak MF, Larson DR, et al. Disease associations with monoclonal gammopathy of undetermined significance: a population-based study of 17,398 patients. Mayo Clin Proc. (2009) 84:685–93. doi: 10.4065/84.8.685
62. Panfilio S, D'Urso P, Annechini G, D'Elia GM, De Angelis F, Stefanizzi C, et al. Regression of a case of multiple myeloma with antiviral treatment in a patient with chronic HCV infection. Leuk Res Rep. (2013) 2:39–40. doi: 10.1016/j.lrr.2013.01.002
63. Rodríguez García A, Linares M, Mennesson N, Sanchez-Vega B, Sanchez R, Alonso Fernandez R, et al. The role of Hepatitis C virus in the development of multiple myeloma: A case study. 60th Annual Meeting of the American Society of Hematology (ASH), San Diego, CA, USA, December 1-4, (2018). Abstract Nbr: 5592, Submission ID: 112842. Blood. (2018) 132:5592. doi: 10.1182/blood-2018-99-112842
64. Pal Singh S, Dammeijer F, Hendriks RW. Role of Bruton's tyrosine kinase in B cells and malignancies. Mol Cancer. (2018) 17:57. doi: 10.1186/s12943-018-0779-z
65. Burger JA, Chiorazzi N. B cell receptor signaling in chronic lymphocytic leukemia. Trends Immunol. (2013) 34:592–601. doi: 10.1016/j.it.2013.07.002
66. Pagano JS, Whitehurst CB, Andrei G. Antiviral drugs for EBV. Cancers (Basel). (2018) 10:E197. doi: 10.3390/cancers10060197
67. Verma D, Thompson J, Swaminathan S. Spironolactone blocks epstein-barr virus production by inhibiting EBV SM protein function. Proc Natl Acad Sci USA. (2016) 113:3609–14. doi: 10.1073/pnas.1523686113
68. Tomasson MH, Ali M, De Oliveira V, Xiao Q, Jethava Y, Zhan F, et al. Prevention is the best treatment: the case for understanding the transition from monoclonal gammopathy of undetermined significance to myeloma. Int J Mol Sci. (2018) 19:E3621. doi: 10.3390/ijms19113621
69. Weinreb NJ, Charrow J, Andersson HC, Kaplan P, Kolodny EH, Mistry P, et al. Effectiveness of enzyme replacement therapy in 1028 patients with type 1 gaucher disease after 2 to 5 years of treatment: a report from the gaucher registry. Am J Med. (2002) 113:112–9. doi: 10.1016/S0002-9343(02)01150-6
70. Weinreb NJ, Goldblatt J, Villalobos J, Charrow J, Cole JA, Kerstenetzky M, et al. Long-term clinical outcomes in type 1 Gaucher disease following 10 years of imiglucerase treatment. J Inherit Metab Dis. (2013) 36:543–53. doi: 10.1007/s10545-012-9528-4
71. Mistry PK, Lukina E, Ben Turkia H, Amato D, Baris H, Dasouki M, et al. Effect of oral eliglustat on splenomagealy in patients with Gaucher disease type 1 : the ENGAGE randomized clinical trial. JAMA. (2015) 313:695–706. doi: 10.1001/jama.2015.459
72. Wasserstein MP, Diaz GA, Lachmann RH, Jouvin MH, Nandy I, Ji AJ, et al. Olipudase alfa for treatment of acid sphingomyelinase deficiency (ASMD): safety and efficacy in adults treated for 30 months. J Inherit Metab Dis. (2018) 41:829–38. doi: 10.1007/s10545-017-0123-6
73. Lachmann RH. Treating lysosomal storage disorders: what have we learnt? J Inherit Metab Dis. (2019) 43:125–32. doi: 10.1002/jimd.12131
74. Lwin ST, Olechnowicz SW, Fowler JA, Edwards CM. Diet-induced obesity promotes a myeloma-like condition in vivo. Leukemia. (2015) 29:507–10. doi: 10.1038/leu.2014.295
75. Pavlova EV, Archer J, Wang S, Dekker N, Aerts JM, Karlsson S, et al. Inhibition of UDP-glucosylceramide synthase in mice prevents Gaucher disease-associated B-cell malignancy. J Pathol. (2015) 235:113–24. doi: 10.1002/path.4452
Keywords: monoclonal immunoglobulin A (IgA), multiple myeloma, monoclonal gammopathy of undetermined significance (MGUS), infectious antigens, Epstein–Barr virus, hepatitis C virus, lysoglucosylceramide (LGL-1), sialylation
Citation: Bosseboeuf A, Seillier C, Mennesson N, Allain-Maillet S, Fourny M, Tallet A, Piver E, Lehours P, Mégraud F, Berthelot L, Harb J, Bigot-Corbel E and Hermouet S (2020) Analysis of the Targets and Glycosylation of Monoclonal IgAs From MGUS and Myeloma Patients. Front. Immunol. 11:854. doi: 10.3389/fimmu.2020.00854
Received: 20 September 2019; Accepted: 14 April 2020;
Published: 27 May 2020.
Edited by:
Harry W. Schroeder, University of Alabama at Birmingham, United StatesReviewed by:
Masaki Hikida, Akita University, JapanDagmar Scheel-Toellner, University of Birmingham, United Kingdom
Copyright © 2020 Bosseboeuf, Seillier, Mennesson, Allain-Maillet, Fourny, Tallet, Piver, Lehours, Mégraud, Berthelot, Harb, Bigot-Corbel and Hermouet. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sylvie Hermouet, c3lsdmllLmhlcm1vdWV0QHVuaXYtbmFudGVzLmZy