 Paula Keiko Sato1,2*†
Paula Keiko Sato1,2*† Felipe Delatorre Busser1,2†
Felipe Delatorre Busser1,2† Flávia Mendes da Cunha Carvalho1,3‡
Flávia Mendes da Cunha Carvalho1,3‡ Alexandra Gomes dos Santos1,3
Alexandra Gomes dos Santos1,3 Aya Sadahiro4
Aya Sadahiro4 Constancia Lima Diogo1,2
Constancia Lima Diogo1,2 Adriana Satie Gonçalves Kono5
Adriana Satie Gonçalves Kono5 Maria Luiza Moretti6
Maria Luiza Moretti6 Olinda do Carmo Luiz7
Olinda do Carmo Luiz7 Maria Aparecida Shikanai-Yasuda1,2,3*
Maria Aparecida Shikanai-Yasuda1,2,3*- 1Laboratory of Medical Investigation in Immunology (LIM48), Faculdade de Medicina, Hospital das Clínicas, University of São Paulo, São Paulo, Brazil
- 2Institute of Tropical Medicine, Faculdade de Medicina, University of São Paulo, São Paulo, Brazil
- 3Department of Infectious and Parasitic Diseases, Faculdade de Medicina, University of São Paulo, São Paulo, Brazil
- 4Department of Parasitology, Instituto de Ciências Biológicas, Federal University of Amazonas, Manaus, Brazil
- 5Infectious Diseases Division, Hospital das Clínicas, University of São Paulo, São Paulo, Brazil
- 6Faculdade de Ciências Médicas, Hospital das Clínicas, State University of Campinas, Campinas, Brazil
- 7Department of Preventive Medicine, Faculdade de Medicina, University of São Paulo, São Paulo, Brazil
Paracoccidioidomycosis (PCM) is an important endemic, systemic disease in Latin America caused by Paracoccidioides spp. This mycosis has been associated with high morbidity and sequels, and its clinical manifestations depend on the virulence of the infecting strain, the degree and type of immune response, infected tissues, and intrinsic characteristics of the host. The T helper(Th)1 and Th17/Th22 cells are related to resistance and control of infection, and a Th2/Th9 response is associated with disease susceptibility. In this study, we focused on interleukin(IL)-12p35 (IL12A), IL-18 (IL18), and IFN-γ receptor 1 (IFNGR1) genetic polymorphisms because their respective roles have been described in human PCM. Real-time PCR was employed to analyze IL12A-504 G/T (rs2243115), IL18-607 C/A (rs1946518), and IFNGR1-611 A/G (rs1327474) single nucleotide polymorphisms (SNP). One hundred forty-nine patients with the acute form (AF), multifocal chronic (MC), or unifocal chronic (UC) forms of PCM and 110 non-PCM individuals as a control group were included. In the unconditional logistic regression analysis adjusted by ethnicity and sex, we observed a high risk of the IL18-607 A-allele for both AF [p = 0.015; OR = 3.10 (95% CI: 1.24–7.77)] and MC groups [p = 0.023; OR = 2.61 (95% CI: 1.14–5.96)] when compared with UC. The IL18-607 A-allele associated risk for the AF and MC groups as well as the protective role of the C-allele in UC are possibly linked to higher levels of IL-18 at different periods of the course of the disease. Therefore, a novel role of IL18-607 C/A SNP is shown in the present study, highlighting its importance in the outcome of PCM.
Introduction
Paracoccidioidomycosis (PCM) is one of the main endemic, systemic mycoses in Latin America, and it is caused by the thermally dimorphic fungi of the Paracoccidioides brasiliensis (P. brasiliensis) complex (Paracoccidioides spp.) and Paracoccidioides lutzii. PCM is associated with high morbidity and sequels; however, because it is not a compulsorily notified disease in Brazil, the actual data are based on reports of epidemiological surveys, case series, hospitalization, and mortality data (1). It is endemic in the southeast, central west, and south regions of Brazil and estimated at 0.71–2.70/100,000 inhabitants/year (2). Epidemic areas have been reported in the western Brazilian areas with a mean incidence of 9.4/100,000 and peaks of 37–39/100,000 inhabitants (3).
The inhalation of conidia of Paracoccidioides spp. can result in infection without symptoms, acute or chronic disease, or in sequelae. The clinical manifestations depend on the virulence of the infecting strain, the degree and type of immune response, infected tissues, and intrinsic characteristics of the host (4).
Some components of the innate immunity, such as neutrophils, dendritic cells, toll-like receptors, dectin-1, myeloid differentiation primary response 88 (MyD88), and NOD-like receptor P3 (NLRP3) inflammasome, have been evaluated in both experimental and human PCM (5–10). However, it is the adaptive response to P. brasiliensis that has been extensively studied with a well-established murine model and characterization of clinical forms of PCM by T helper (Th) responses and antibodies (11).
The acute form of PCM shows a mixed Th2/Th9 response: increased levels of IL-4, IL-5, IL-9, IL-10, TGF-β, and IL-27; low production of IFN-γ and TNF-α; and high levels of specific IgG4 and IgE antibodies. On the other hand, the chronic form presents a Th17/Th22 profile with high production of IL-17 and IL-22 and also secreting Th1-type cytokines, such as IFN-γ, TNF-α, and IL-2 as well as variable levels of IL-10 and IL-4 and increased levels of specific IgG1 antibodies (12, 13). Therefore, the Th2/Th9 responses can be associated with susceptibility to PCM, and the presence of Th1 and Th17/Th22 cells can contribute to more mild clinical manifestations with the axis IFN-γ/IL-12 directly associated with protection and control of the infection (11–16).
The high-affinity binding of IL-12 to its receptor results in the differentiation of naïve CD4 T cells into Th1 lymphocytes, which are the major producers of IFN-γ, alongside NK cells. In the presence of IL-12, IL-18 also stimulates the production of IFN-γ, inducing a Th1-mediated immune response; in the absence of IL-12, IL-18 can stimulate a Th2 response (17, 18).
Higher levels of IL-18 and sTNF-RII are described in the acute form of PCM when compared with the chronic form and controls, and IL-12 is also higher in patients than in controls (19). The same group reports higher levels of IL-18 during treatment and lower levels after antifungal treatment (20). As the disease is more severe in the acute form with high IL-18 levels, the authors suggest that this cytokine could be a useful marker of PCM severity.
Previous studies on murine PCM have shown that IL-12 and IL-18 secretions are associated with innate immunity factors. MyD88-deficient mice show a more severe disease after 8 weeks of infection with low levels of IL-12, and a protective role in murine pulmonary PCM was shown in the NLRP3 inflammasome, associated with IL-1β and IL-18 secretion and expansion of Th1 and Th17 cells and suppressive control of T-reg cells (7, 21).
The mechanisms controlling these effects are unclear, but reports on genetic background in both human and experimental disease have been shedding light on this matter. For instance, an autosomal dominant gene has been associated with resistance in murine PCM (22), and in human disease, reports show the influence of the human leukocyte antigen (HLA) in both susceptibility and outcomes. The HLA class I, HLA-B13, was found in a higher proportion in PCM patients compared to controls as well as a higher frequency of HLA-A9 in patients with the progressive pulmonary form (23). In parallel, increased risk of PCM development was associated with the presence of HLA-B40, which was found more in patients than in controls (24, 25). In 2011, the class II-HLA-DRB1*11 allele was reported in a higher frequency in patients with the more benign clinical presentation of this disease, the unifocal chronic (UC) form (26). Moreover, PCM patients were shown to have a higher proportion of the non-expressed C4B allele, C4B*Q0, of deficient C4 isotypes, suggesting to the authors a possible influence of different C4 isotype and allotype frequencies in the course of infection (27).
In addition, an enzyme phenotype (GLO-1 phenotype of glyoxalase I) was associated with PCM infection represented by a positive intradermic reaction, and a possible relationship with HLA antigens deserves further discussion because there is a close linkage between the GLO phenotype and HLA (28).
Our group and others have been studying single nucleotide polymorphisms (SNPs) and different mutations on cytokines and receptor genes. For instance, the inherited mutation Leu77Phe on the IL-12 receptor β1 subunit (IL-12Rβ1) gene (IL12RB1) associated with its loss of function and complete deficiency, resulting in a severe, acute form of PCM (29). Moreover, in Brazilian patients with PCM, reports of polymorphisms on IL-4 and IL-12Rβ1 genes have shown the relevance of IL4-590 C/T and IL12RB1 641 A/G SNPs in association with infection or clinical forms, contributing to a better understanding of the immunopathogenesis of this disease (30, 31).
Polymorphisms on the IFN-γ gene have also been described in infectious diseases, such as toxoplasmosis, tegumentar leishmaniasis, and PCM, and no association between alleles or genotypes and these diseases was observed (31–35). Furthermore, it is also necessary to consider that the expression of the IFN-γ receptor could be interfering in the axis IL-12/IFN-γ because its deficiency was already described in the more disseminated forms of histoplasmosis, coccidioidomycosis, mycobacteriosis, and disseminated BCG infection (36–39). This deficient expression could be associated with a mutation in the IFN-γ receptor 1 gene, such as the IFNGR1-611 A/G SNP, which has been associated with strong promoter activity and with decreased risk of pulmonary tuberculosis (40, 41).
IL-12p70, the bioactive form of IL-12, is a heterodimer of two subunits: p35 (encoded by the IL12A gene) and p40 (encoded by the IL12B gene). No association was found between IL12B +1188 A/C SNP and PCM (31). SNPs in the IL12A gene, on the other hand, have not yet been explored in this mycosis.
Studies show an association of IL18-607 C/A SNP (rs1946518) with protection or risk in several infections (42–45). However, as with the IL12A-504 G/T (rs2243115) and the IFNGR1-611 A/G (rs1327474) SNPs, there are still no studies associating this genetic polymorphism with systemic fungal infections.
Considering the interaction between IL-18, IL-12, and IFN-γ and the genetic aspects possibly involved in susceptibility to PCM and the lack of studies on this disease, the present study aimed to analyze the IL18-607 C/A (rs1946518), IL12A-504 G/T (rs2243115), and IFNGR1-611 A/G (rs1327474) SNPs, for the first time to our knowledge, in a cohort of Brazilian patients with PCM, also presenting a brief review of studies on these SNPs in infectious diseases.
Materials and Methods
Subjects
A total of 149 patients with PCM from the General Infirmary and Systemic Mycosis Outpatient Clinic from the Infectious Diseases Division (Hospital das Clínicas, Faculdade de Medicina, University of São Paulo—HCFMUSP, São Paulo, SP, Brazil) were included in the study (Table 2). Thirty-nine patients had the acute form of PCM (AF) and 110 had the chronic form (CF); 93 had the multifocal chronic form (MC) and 17 had the UC form. This classification was performed according to Franco et al. (46). UC includes only patients with mild, restricted disease to the skin, mucosae, or lymph nodes without lung or cerebral involvement. The control group (CO) included 110 non-PCM subjects. The study protocol was approved by the ethics committee of HCFMUSP (CAPPesq 10273; Plataforma Brasil 123334/2013), and all subjects gave written informed consent in accordance with the Declaration of Helsinki.
The inclusion criteria were (a) patients with PCM: identification of Paracoccidioides spp. by mycological and/or histopathological examination and/or presence of anti-P. brasiliensis serum antibodies (titers ≥32 on counterimmunoelectrophoresis test) at the moment of enrollment or proven in the past; and (b) CO: individuals considered healthy without a previous history of the disease, not sensitized in lymphoproliferation assays against the 43-kDa glycoprotein of P. brasiliensis, and absence of serum anti-P. brasiliensis antibodies (by immunodiffusion test). Subjects with comorbidities, such as neoplasia and other acute or chronic systemic infectious diseases, were excluded.
DNA Extraction
Genomic DNA was obtained from peripheral blood leukocytes by the salt precipitation method (DTAB/CTAB; dodecyl trimethyl ammonium bromide/cetyltrimethyl ammonium bromide, both from Sigma-Aldrich, Merck, St. Louis, MO, USA) as previously described (47). The concentration and purity of the extracted DNA were evaluated by a UV spectrophotometer (Nanodrop LITE, Thermo Fisher Scientific, Carlsbad, CA, USA).
Detection of SNPs
The SNPs in the IL-12p35 (IL12A-504 G/T, rs2243115), IL-18 (IL18-607 C/A, rs1946518), and IFN-γ receptor 1 (IFNGR1-611 A/G, rs1327474) genes were investigated by real-time PCR using specific oligonucleotides and probes labeled with VIC (wild allele) or FAM (mutated allele) fluorochromes and TaqMan™ Genotyping Master Mix (all from Molecular Probes, Thermo Fisher Scientific, Carlsbad CA, USA). The assays and results were performed and analyzed with the StepOne Plus Real Time PCR System and software (Applied Biosystems, Thermo Fisher Scientific, Foster City, CA, USA).
Statistical Analysis
The deviations from the Hardy–Weinberg equilibrium and the distribution of genotypic and allelic frequencies of the SNPs on IL12A, IL18, and IFNGR1 genes in the studied population were evaluated by Pearson's χ2 test. To estimate the risk of patients with AF, MC, and UC PCM associated with genotypes and alleles, odds ratios (ORs) and 95% confidence intervals (95% CIs) were calculated as approximations of relative risk using unconditional logistic regression analysis. For ordered variables, tests for linear trend were done by categorizing the exposure variables and entering the scores as continuous. To verify the strength of association between the final events, we performed univariate and multivariate logistic analyses adjusted by ethnicity and sex with the STATA 14.0 software (StataCorp, College Station, TX, USA), and p-values ≤ 0.05 were considered statistically significant.
Results
Considering the plausible role of genetic background in human PCM, we summarize all previously reported SNPs or mutations on immune-related genes and their main results on Brazilian patients in Table 1. Eight SNPs or mutations on DC-SIGN, HLA, IL-4, IL-10, IL-12Rβ1, and Vitamin D receptor genes were found to have an association with risk, outcome, or clinical forms of PCM. As the SNPs of this study had not yet been evaluated on PCM patients, we also compiled previous reports on the IL18 (-607 C/A, rs1946518 and -137 G/C, rs187238), IL12A (-504 G/T, rs2243115), and IFNGR1 (-611 A/G, rs13277474) SNPs and their association with diverse infectious diseases (Supplementary Tables 1–3, respectively).

Table 1. Polymorphisms on immune-related genes in Brazilian patients with PCM.
This study evaluated the IL18-607 C/A, IL12A-504 G/T, and IFNGR1-611 A/G SNPs in 149 patients with the AF and CF of PCM and 110 control individuals (CO). The CF group is subclassified as MC and UC forms. There were no statistically significant differences between cases and controls in the distribution of ethnic groups or sex (data not shown). Regarding the different groups of patients (AF, MC, and UC), no differences in ethnic distribution were found (p = 0.476). However, in the univariate analysis of sex distribution, there was a significant difference between these groups with a male majority on a ratio of 17.6 in the MC group (Table 2).
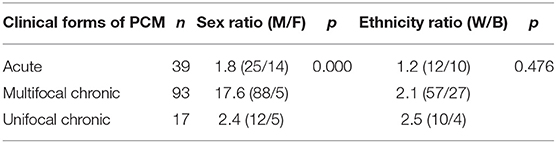
Table 2. Distribution (n), sex (M = Male; F = Female) and ethnicity (W = White; B = Black) ratio, and p-values among the groups of patients with acute, multifocal chronic and unifocal chronic forms of PCM.
Genotypic frequencies for the three evaluated SNPs were in Hardy–Weinberg equilibrium (data not shown). The distribution of genotypic and allelic frequencies of IL12A-504 G/T, IL18-607 C/A, and IFNGR1-611 A/G SNPs among patients and controls and among AF and CF groups is shown in Table 3. We found no statistical significances in the univariate analysis of genotypic frequencies in the codominant, dominant, and recessive models nor in the distribution and frequencies of alleles.
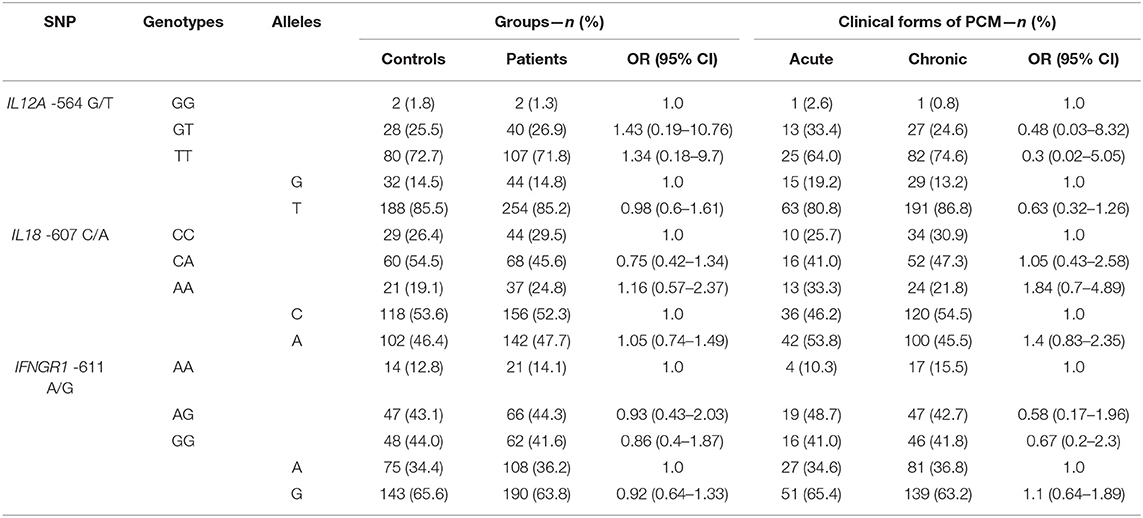
Table 3. Genotypic and allelic distributions and frequencies of IL12A-504 G/T, IL18-607 C/A, and IFNGR1-611 A/G SNPs, values of Odds Ratio (OR) and 95% Confidence Interval (95% CI) among the groups of Controls (n = 110) and Patients (n = 149), and among the groups of patients with Acute (n = 39) and Chronic (Multifocal + Unifocal) forms of PCM (n = 110).
In Table 4, we show the distribution of genotypic and allelic frequencies of IL12A-504 G/T, IL18-607 C/A, and IFNGR1-611 A/G SNPs among the groups of patients with the different clinical forms of PCM, analyzed by unconditional logistic regression with adjustments for ethnicity and sex. The comparison between AF, MC, and UC genotypic distribution did not result in a statistical difference.
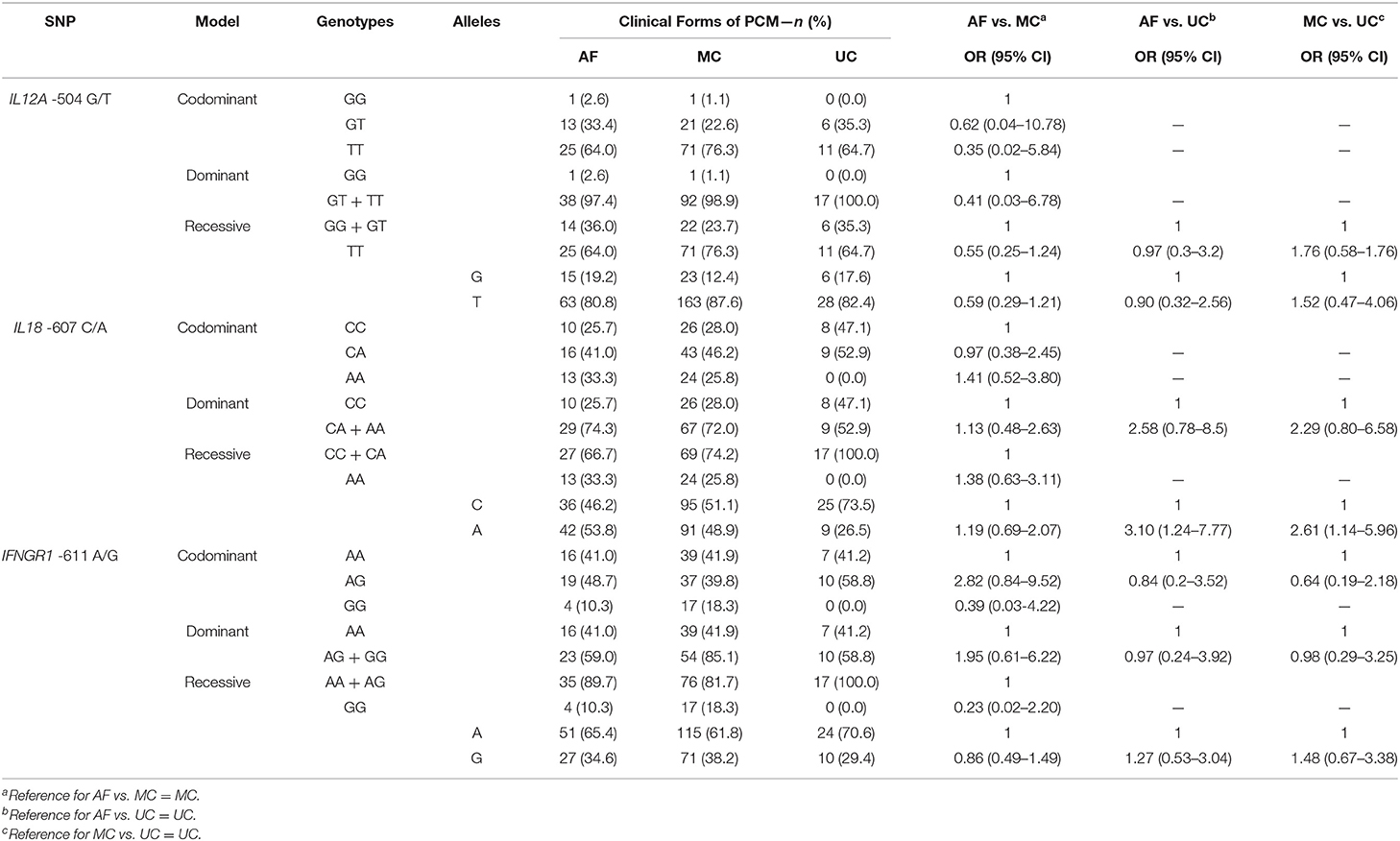
Table 4. Genotypic and allelic distributions and frequencies of IL12A-504 G/T, IL18-607 C/A, and IFNGR1-611 A/G SNPs, values of Odds Ratio (OR) and 95% Confidence Interval (95% CI) adjusted for sex and race by unconditional logistic regression analysis among the groups of patients with Acute (AF, n = 39), Multifocal Chronic (MC, n = 93) and Unifocal Chronic (UC, n = 17) forms of PCM.
Regarding the allelic distribution and frequencies of IL18-607 C/A SNP, unconditional logistic regression with adjustments for ethnicity and sex shows significant differences between AF and UC [p = 0.015; OR = 3.1 (95% CI: 1.24–7.77)] with a higher frequency of the A-allele in AF (53.8%) than in UC (26.5%) (Tables 4, 5). The same is observed in the comparison between MC and UC [p = 0.023; OR = 2.61 (95% CI: 1.14–5.96)]. Concerning alleles of IL12A-504 G/T and IFNGR1-611 A/G SNPs, no statistical differences were found. Furthermore, Table 5 shows the statistical data on sex and ethnicity as covariates of IL18-607 C/A SNP in the distribution of genotypes and alleles. There was a significant difference of sex distribution between AF and MC with a higher proportion of women in AF (p = 0.000); this is the opposite of the comparison between MC and UC, in which the proportion of female patients is significantly lower in the MC group (p < 0.015). This same effect occurred with the codominant, dominant, recessive, and allelic analyses, confirming and detailing the data of Table 2. There were no differences regarding sex distribution between AF and UC, and ethnicity had no statistically significant effect as a covariate in this analysis.
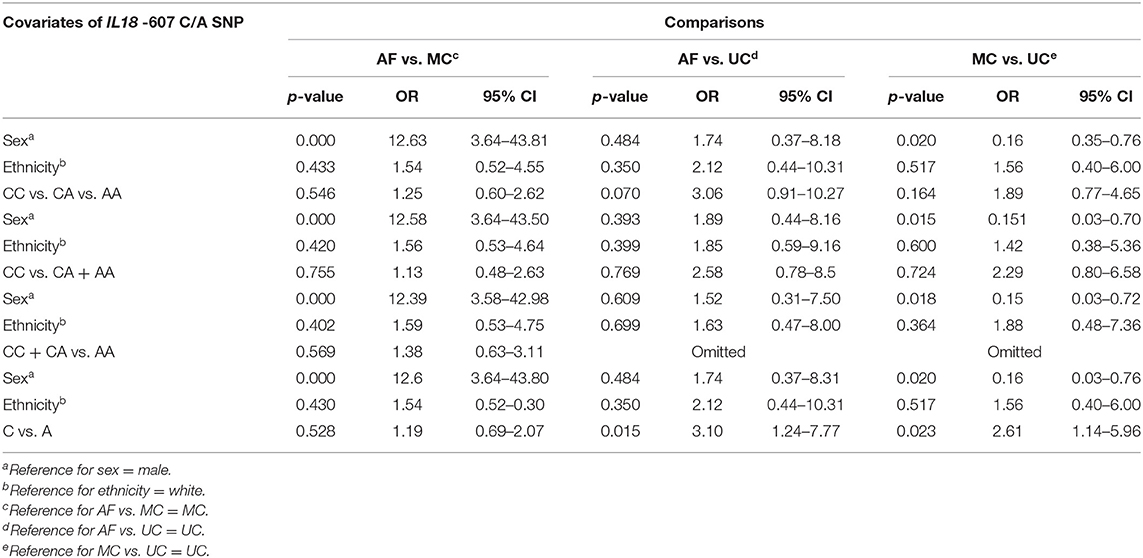
Table 5. Association studies between the groups of patients with Acute (AF), Multifocal Chronic (MC), and Unifocal Chronic (UC) forms of PCM including sex, ethnicity, and the IL18-607 C/A SNP as covariates and results of p-values, Odds Ratio (OR), and 95% Confidence Interval (95% CI).
Discussion
Genetic studies are relevant to understanding the mechanisms involved in the pathogenesis of diseases (50, 51). Some genetic polymorphisms are shown to directly interfere in cytokine expression, therefore directing the immune response of the host and possibly influencing the outcome of the disease.
In the present study, we investigated the possible association between the IL12A-504 G/T, IL18-607 C/A, and IFNGR1-611 A/G SNPs in Brazilian patients with PCM and disease susceptibility. The distribution of genotypes and alleles of the IL12A-504 G/T SNP was similar in all evaluated comparisons on our study. Although this SNP was related to immune responses against rubella vaccination and HBV and protection in tuberculosis, we did not observe a clear association with PCM (52–54).
As for IFNGR1-611 A/G SNP, genotypic and allelic distributions were similar in all evaluated comparisons, confirmed by unconditional logistic regression analysis (adjusted for sex and race). The absence of association with risk or protection reported here is similar to the data reported in tuberculosis and liver fibrosis progression due to recurrent hepatitis C (55–58). An association between tuberculosis and the IFNGR1-611 A/G and -56 T/C haplotype was observed by Bullak-Kardum et al. (41). In fact, the promoter activity is supposed to be stronger in -611 A/G than -56 T/C, and the variant A is estimated to decrease the binding of GATA-1 and TFIID factors to this site (40, 58). Because previous associations with risk or protection have been described in other SNPs, further studies should include the IFNGR1-611 A/G SNP.
In our analysis of the IL18-607 C/A SNP, we found no differences on allelic and genotypic distributions between patients and controls or between AF and CF groups of patients. On the other hand, the adjustment for sex and ethnicity in the unconditional regression logistic analysis confirmed the presence of the A-allele as a risk factor for the AF and MC groups when compared to the UC group.
The association of the A-allele/AA-genotype and higher risk is shown in various infectious diseases, such as chronic hepatitis B in Thailand and in India, gingivitis in the Czech Republic, pulmonary tuberculosis in China, Chagas disease in Colombia (mainly driven by rs360719), and infection by the hepatitis C virus in Egypt (37, 43, 52–62). Similarly to our results, the mutant allele/genotype of IL18-607 C/A is also associated with more severe outcomes in other infectious diseases: higher virus shedding of the severe acute respiratory syndrome-associated (SARS) coronavirus in Taiwan, lipodystrophy syndrome on HIV-positive Brazilian individuals, immune restoration disease on HIV–tuberculosis coinfected Indian patients, bacterial infections after liver transplantation in China, and hepatitis C–related hepatocellular carcinoma in Egypt (63–67).
The effect of the A-allele/AA-genotype on PCM and other infectious diseases could be explained by changes in IL-18 levels introduced by this mutant allele in the -607 position. In effect, the IL-18 human gene is composed of six exons and five introns with three very well-known SNPs in the promoter region: -656 G/T (rs1946519), -607 C/A (rs1946518), and -137 G/C (rs187238). Two of these positions, -607 and -137, are thought to be nuclear factor binding sites for cAMP responsive element binding protein and H4TF-1 nuclear factor, respectively, and mutation on both sites can affect the IL-18 levels (50).
The -607 C-allele/CC-genotype carriage has been associated with higher levels of IL-18 in the serum and/or of mRNA expression (50, 66, 68–70). In our study, we found higher C-allele carriage on patients with the UC form of PCM (73.5%), whereas the AF and MC groups (with 46.2 and 51.1% of C-allele carriage, respectively) had already been reported to have higher serum IL-18 levels than those from the former group (19, 20).
We hypothesize that the higher C-allele carriage on patients with the UC form of PCM may induce higher IL-18 levels at the early stages of infection, determining increased levels of IFN-γ and a more efficient cellular response that controls fungal dissemination and the consequential tissular inflammation. In parallel, previous work in our lab shows that phytohemagglutinin-stimulated cells from IL18-607 C-allele-unifocal patients apparently produce more IFN-γ than C-allele carriers from the acute or the chronic multifocal groups, although without statistically significant difference (unpublished data). However, because both infection and disease in humans are recognized in a later and undefined period after fungal entrance, those initial events can only be evaluated in experimental PCM and have been elegantly shown in deficient mice models. The intravenous infection of IL-18-deficient mice (IL-18−/−) with P. brasiliensis yeast cells resulted in a higher fungal burden in the lungs compared with wild-type (WT) animals and absence of granuloma formation (71). Furthermore, P. brasiliensis intratracheally infected mice deficient for NLRP3 inflammasome components (Nlrp3−/−, Casp1/11−/−, Asc−/−) as well as deficient for the ATP receptor (P2x7r−/−) also had a higher fungal burden in their lungs and liver; predominance of CD4+IL-4+, CD4+TGF-β+, and T-reg cells; a lower number of pulmonary PMN cells; and less IL-18 and IL-1β compared with their WT controls (7).
In this context, it is possible that patients with the less severe UC form of PCM present a more balanced IL-18/IL-12/IFN-γ axis, resulting in more localized and milder clinical manifestations compared with the other groups, in which the higher IL-18 levels are accompanied by other immunomodulating cytokines. The more severe form of PCM, the acute form, has been characterized by IL-4 with IL-18 inducing a Th2 immune response (high levels of IL-4, IL-5, and IL-13), by IL-4 and TGF-β inducing a Th9 pattern (high levels of IL-9), and both Th2 and Th9 inhibiting the Th1 response. Patients with chronic PCM were previously shown to have a mixed immune response of Th1 (IL-12 and IL-18 leading to IFN-γ production), Th17 (induced by IL-18, IL-1 and IL-23), and Th22 (induced by IL-18, IL-1, IL-23, and IL-21), which result in heterogeneous clinical symptoms (12, 13, 20).
In our study, the UC group included only patients with mild and restricted disease to the skin, mucosae, or lymph nodes. The exclusion of patients with lung involvement was based on the possible misclassification of MC patients who are more prone to develop pulmonary lesions. Although these criteria resulted in fewer patients in the UC group, it also revealed specific immunogenetic characteristics, such as the association of a more favorable outcome with the IL18-607 C-allele in the present study and the IL12RB1 641 AG-genotype and the HLA-DRB1*11 allele carriage, previously reported by our group (26, 31).
Additionally, the association of the IL18-607 A-allele with the more severe forms of PCM described herein collaborates with previous reports on different genes emphasizing the influence of genetic background on the outcome of this mycosis. The GG-genotype of IL10-1082 G/A SNP, the RP2/RP2-genotype of the intron-3 microsatellite polymorphism of the IL4 gene, the AA-genotype of IL12RB1 641 A/G SNP, the GG-genotype of the DCSIGN rs4804803 A/G SNP, and the CC-genotype of the VDR rs7975232 A/C SNP have all been previously associated with susceptibility or the more severe outcome in PCM (26, 29–31, 34, 35, 49). In parallel, HLA class I antigens and the GLO-1 phenotype of the glyoxalase enzyme have been associated with the progression of disease (pulmonary form) or infection (23–25, 27, 28).
We reinforce that our present data and all the genetic associations previously described may be considered in the same context of association of autosomal gene dominance with resistance in murine PCM, which is a model that reproduces several characteristics of the human disease (22).
The comparison between patients and controls in our study shows a greater number of male than female subjects, particularly in the MC group. As previously described, there has been a male predominance among cases of chronic PCM (72–74). This mycosis manifests more frequently in male farmers who are constantly working in direct contact with the soil where Paracoccidioides spp. probably occurs (1). In addition, infected women are less likely to manifest the chronic form because of the putative role of estrogen (17-β-estradiol) as a protective factor that impairs conidia transformation into yeast form during murine infection (75–77). Although this effect has been less commonly registered in human PCM, epidemiological data reported in Brazil show that the disease is rare in adult females (4.3%) and usually occurring in the menopausal period (91.3%) (78). Contrarily, it has been demonstrated that 17-β-estradiol can exert an anti-inflammatory role by decreasing TNF-α and IL-6 while increasing IL-4 levels, which further results in a Th2 response (79). In oral PCM lesions, a positive correlation between the amount of estradiol receptors and the fungal burden was observed only in female patients (80). These recent findings may explain the more even distribution of the acute form of PCM among sexes, and the higher frequency of females in UC and lower in MC could be evidence of protection for women in developing the more severe chronic form of the disease.
Possible limitations in our study are the inclusion of only one health center, mixed ethnic groups in the Brazilian population, and lack of detection of IL-18 levels and functional analyses. Although reflecting the distribution of PCM in endemic areas, our cohort with a low number of patients with the UC form is also a limitation for further inquiry.
In summary, our study did not show an association between PCM and the evaluated IL12A and IFNGR1 SNPs or between the acute and chronic forms or between multifocal and unifocal chronic forms. For the IL18-607 C/A SNP, no association was shown with infection or among acute and chronic forms. However, we show an association between the IL18-607 A-allele and the more severe clinical forms of PCM, acute and multifocal chronic forms, when compared to the UC form, the less severe form of this disease, associated with the IL18-607 C-allele. To the best of our knowledge, this is the first study that evaluates the association between IL12A, IFNGR1, and IL18 SNPs and PCM.
The present data suggest a novel role of IL18-607 C/A SNP as a contributor to a more favorable outcome of this disease, potentially leading to a more balanced and more efficient cellular response for the control of fungal dissemination at the early stages of infection in UC. Furthermore, our work highlights the need for new studies with other SNPs on the IL18 gene and on other components of the immune response for a better understanding of the pathophysiology and the clinical expression of PCM.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
The studies involving human participants were reviewed and approved by Research and Ethics Committee of Hospital das Clínicas, Faculdade de Medicina, University of São Paulo - HCFMUSP, São Paulo, SP, Brazil (CAPPesq 10273; Plataforma Brasil 123334/2013). The patients/participants provided their written informed consent to participate in this study.
Author Contributions
PS and MS-Y conceived and designed the experiments. MM and AK contributed with samples and patient selection. PS, FB, FC, CD, and AS performed the experiments. PS, AG, and OL analyzed the data. PS, FB, AG, FC, and MS-Y wrote the paper and/or performed bibliographic revision. PS, MS-Y, MM, AK, FB, FC, CD, AS, AG, and OL approved the paper. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP 2012/25192-7) and Fundação Faculdade de Medicina. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. FC was supported by grants from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), and FB was supported by grants from FAPESP (2014/07801-1).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Nídia P. O. Silva and Eron L. dos Santos for their contribution on DNA extraction of some of the samples and Marcia A. Ferreira, Carlos Jose Quinteiro, and Rodolfo E. Fujimori for providing older articles.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.542210/full#supplementary-material
References
1. Martinez R. New trends in paracoccidioidomycosis epidemiology. J Fungi. (2017) 3:1. doi: 10.3390/jof3010001
2. Martinez R. Epidemiology of paracoccidioidomycosis. Rev Inst Med Trop São Paulo. (2015) 57:11–20. doi: 10.1590/S0036-46652015000700004
3. Vieira G, de D, Alves T, da C, Lima SM, Camargo LM, Sousa CM. Paracoccidioidomycosis in a western Brazilian Amazon State: clinical-epidemiologic profile and spatial distribution of the disease. Rev Soc Bras Med Trop. (2014) 47:63–8. doi: 10.1590/0037-8682-0225-2013
4. Benard G. An overview of the immunopathology of human paracoccidioidomycosis. Mycopathologia. (2008) 165:209–21. doi: 10.1007/s11046-007-9065-0
5. Calich VL, Costa TA, Felonato M, Arruda C, Bernardino S, Loures FV, et al. Innate immunity to Paracoccidioides brasiliensis infection. Mycopathologia. (2008) 165:223–36. doi: 10.1007/s11046-007-9048-1
6. Calich VL, Pina A, Felonato M, Bernardino S, Costa TA, Loures FV. Toll-like receptors and fungal infections: the role of TLR2, TLR4 and MyD88 in paracoccidioidomycosis. FEMS Immunol Med Microbiol. (2008) 53:1–7. doi: 10.1111/j.1574-695X.2008.00378.x
7. Feriotti C, de Araújo EF, Loures FV, da Costa TA, Galdino NAL, Zamboni DS, et al. NOD-like receptor P3 inflammasome controls protective Th1/Th17 immunity against pulmonary paracoccidioidomycosis. Front Immunol. (2017) 10:786. doi: 10.3389/fimmu.2017.00786
8. Jannuzzi GP, de Almeida JRF, Amarante-Mendes GP, Romera LMD, Kaihami GH, Vasconcelos JR, et al. TLR3 is a negative regulator of immune responses against Paracoccidioides brasiliensis. Front Cell Infect Microbiol. (2019) 8:426. doi: 10.3389/fcimb.2018.00426
9. Rodrigues DR, Fernandes RK, Balderramas H, de A, Penitenti M, Bachiega TF, Calvi SA, et al. Interferon-gamma production by human neutrophils upon stimulation by IL-12, IL-15 and IL-18 and challenge with Paracoccidioides brasiliensis. Cytokine. (2014) 69:102–9. doi: 10.1016/j.cyto.2014.05.009
10. de Castro LF, Longhi LNA, Paião MR, Justo-Júnior ADS, de Jesus MB, Blotta MHSL, et al. NLRP3 inflammasome is involved in the recognition of Paracoccidioides brasiliensis by human dendritic cells and in the induction of Th17 cells. J Infect. (2018) 77:137–44. doi: 10.1016/j.jinf.2018.03.004
11. Calich VL, Vaz CA, Burger E. Immunity to Paracoccidioides brasiliensis infection. Res Immunol. (1998) 149:407–500. doi: 10.1016/S0923-2494(98)80764-5
12. Oliveira SJ, Mamoni RL, Musatti CC, Papaiordanou PM, Blotta MH. Cytokines and lymphocyte proliferation in juvenile and adult forms of paracoccidioidomycosis: comparison with infected and non-infected controls. Microbes Infect. (2002) 4:139–44. doi: 10.1016/S1286-4579(01)01521-0
13. de Castro LF, Ferreira MC, Silva RM, Blotta MHSL, Longhi LN, Mamoni RL. Characterization of immune response in human paracoccidioidomycosis. J Infect. (2013) 67:470–85. doi: 10.1016/j.jinf.2013.07.019
14. Calich VLG, Mamoni RL, Loures FV. Regulatory T cells in paracoccidioidomycosis. Virulence. (2018) 1:1–12. doi: 10.1080/21505594.2018.1483674
15. Benard G, Romano CC, Cacere CR, Juvenale M, Mendes-Giannini MJ, Duarte AJ. Imbalance of IL-2, IFN-gamma and IL-10 secretion in the immunosuppression associated with human paracoccidioidomycosis. Cytokine. (2001) 13:248–52. doi: 10.1006/cyto.2000.0824
16. Romano CC, Mendes-Giannini MJS, Duarte AJS, Benard G. The role of interleukin-10 in the differential expression of interleukin-12p70 and its β2 receptor on patients with active or treated paracoccidioidomycosis and healthy infected subjects. Clin Immunol. (2005) 114:86–94. doi: 10.1016/j.clim.2004.09.005
17. Nakanishi K, Yoshimoto T, Tsutsui H, Okamura H. Interleukin-18 is a unique cytokine that stimulates both Th1 and Th2 responses depending on its cytokine milieu. Cytokine Growth Factor Rev. (2001) 12:53–72. doi: 10.1016/S1359-6101(00)00015-0
18. Samarani S, Allam O, Sagala P, Aldabah Z, Jenabian MA, Mehraj V, et al. Imbalanced production of IL-18 and its antagonist in human diseases, and its implications for HIV-1 infection. Cytokine. (2016) 82:38–51. doi: 10.1016/j.cyto.2016.01.006
19. Corvino CL, Mamoni RL, Fagundes GZ, Blotta MH. Serum interleukin-18 and soluble tumour necrosis factor receptor 2 are associated with disease severity in patients with paracoccidioidomycosis. Clin Exp Immunol. (2007) 147:483–90. doi: 10.1111/j.1365-2249.2006.03308.x
20. Alves ABRM, David MA, de Castro LF, da Silva RM, Longhi LNA, Blotta MHSL, et al. Differential production of interleukin-1 family cytokines (IL-1β, IL-18, IL-33 and IL-37) in patients with paracoccidioidomycosis: correlation with clinical form and antifungal therapy. Med Mycol. (2018) 56:332–43. doi: 10.1093/mmy/myx050
21. Loures FV, Pina A, Felonato M, Feriotti C, de Araújo EF, Calich VL. MyD88 signaling is required for efficient innate and adaptive immune responses to Paracoccidioides brasiliensis infection. Infect Immun. (2011) 79:2470–80. doi: 10.1128/IAI.00375-10
22. Calich VL, Burger E, Kashino SS, Fazioli RA, Singer-Vermes LM. Resistance to Paracoccidioides brasiliensis in mice is controlled by a single dominant autosomal gene. Infect Immun. (1987) 55:1919–23. doi: 10.1128/IAI.55.8.1919-1923.1987
23. de Restrepo FM, Restrepo M, Restrepo A. Blood groups and HLA antigens in paracoccidioidomycosis. Sabouraudia. (1983) 21:35–9. doi: 10.1080/00362178385380061
24. Lacerda GB, Arce-Gomez B, Telles Filho FQ. Increased frequency of HLA-B40 in patients with paracoccidioidomycosis. J Med Vet Mycol. (1988) 26:253–6. doi: 10.1080/02681218880000351
25. Goldani LZ, Monteiro CM, Donadi EA, Martinez R, Voltarelli JC. HLA antigens in Brazilian patients with paracoccidioidomycosis. Mycopathologia. (1991) 114:89–91. doi: 10.1007/BF00436427
26. Sadahiro A, Roque AC, Shikanai-Yasuda MA. Generic human leukocyte antigen class II (DRB1 and DQB1) alleles in patients with paracoccidioidomycosis. Med Mycol. (2007) 45:35–40. doi: 10.1080/13693780600999132
27. de Messias IJ, Reis A, Brenden M, Queiroz-Telles F, Mauff G. Association of major histocompatibility complex class III complement components C2, BF, and C4 with Brazilian paracoccidioidomycosis. Complement Inflamm. (1991) 8:288–93. doi: 10.1159/000463198
28. Balarin MA, Freire-Maia DV. The association between glyoxalase I and paracoccidioidomycosis infection. Rev Soc Bras Med Trop. (1993) 26:141–3. doi: 10.1590/S0037-86821993000300002
29. Moraes-Vasconcelos D, Grumach AS, Yamaguti A, Andrade MEB, Fieschi C, Beaucoudrey L, et al. Paracoccidioides brasiliensis disseminated disease in a patient with inherited deficiency in the beta1 subunit of the interleukin (IL)-12/IL-23 receptor. Clin Infect Dis. (2005) 41:e31–7. doi: 10.1086/432119
30. Mendonça MS, Peraçolli TS, Silva-Vergara ML, Ribeiro SC, Oliveira RF, Mendes RP, et al. High interleukin-4 expression and interleukin-4 gene polymorphisms are associated with susceptibility to human paracoccidioidomycosis. Mem Inst Oswaldo Cruz. (2015) 110:781–5. doi: 10.1590/0074-02760150197
31. Carvalho FM, Busser FD, Freitas VL, Furucho CR, Sadahiro A, Kono AS, et al. Polymorphisms on IFNG, IL12B and IL12RB1 genes and paracoccidioidomycosis in the Brazilian population. Infect Genet Evol. (2016) 43:245–51. doi: 10.1016/j.meegid.2016.05.025
32. Neves E, de S, Curi AL, Albuquerque MC, Palhano-Silva CS, Silva LB, Bueno WF, et al. Genetic polymorphism for IFN-γ +874 T/A in patients with acute toxoplasmosis. Rev Soc Bras Med Trop. (2012) 45:751–60. doi: 10.1590/S0037-86822012000600020
33. Matos GI, Covas CJF, Bittar RC, Gomes-Silva A, Marques F, Maniero VC, et al. IFNG +874 T/A polymorphism is not associated with American tegumentary leishmaniasis susceptibility but can influence Leishmania induced IFN-γ production. BMC Infec Dis. (2007) 7:33. doi: 10.1186/1471-2334-7-33
34. Bozzi A, Pereira PP, Reis BS, Goulart MI, Pereira MC, Pedroso EP, et al. Interleukin-10 and tumor necrosis factor-alpha single nucleotide gene polymorphism frequency in paracoccidioidomycosis. Hum Immunol. (2006) 67:931–9. doi: 10.1016/j.humimm.2006.07.014
35. Bozzi A, Reis BS, Pereira PP, Pedroso EP, Goes AM. Interferon-gamma and interleukin-4 single nucleotide gene polymorphisms in Paracoccidioidomycosis. Cytokine. (2009) 48:212–7. doi: 10.1016/j.cyto.2009.07.011
36. Jouanguy E, Dupuis S, Pallier A, Döffinger R, Fondanèche MC, Fieschi C, et al. In a novel form of IFN-gamma receptor 1 deficiency, cell surface receptors fail to bind IFN-gamma. J Clin Invest. (2000) 105:1429–36. doi: 10.1172/JCI9166
37. Zerbe CS, Holland SM. Disseminated histoplasmosis in persons with interferon-γ receptor 1 deficiency. Clin Infect Dis. (2005) 41:e38–41. doi: 10.1086/432120
38. Vinh DC, Masannat F, Dzioba RB, Galgianai JN, Holland SM. Refractory disseminated coccidioidomycosis & mycobacteriosis in interferon-γ receptor deficiency. Clin Infect Dis. (2009) 49:e62–5. doi: 10.1086/605532
39. Marazzi MG, Chapgier A, Defilippi A, Pistioia V, Mangini S, Savioli C, et al. Disseminated Mycobacterium scrofulaceum infection in a child with interferon-γ receptor 1 deficiency. Int J Infect Dis. (2010) 14:167–70. doi: 10.1016/j.ijid.2009.03.025
40. Rosenzweig SD, Schäffer AA, Ding L, Sullivan R, Enyedi B, Yim JJ, et al. Interferon-gamma receptor 1 promoter polymorphisms: population distribution and functional implications. Clin Immunol. (2004) 112:113–9. doi: 10.1016/j.clim.2004.03.018
41. Bulat-Kardum L, Etokebe GE, Knezevic J, Balen S, Matakovic-Mileusnic N, Zaputovic L, et al. Interferon-gamma receptor-1 gene promoter polymorphisms (G-611A; T-56C) and susceptibility to tuberculosis. Scand J Immunol. (2006) 63:142–50. doi: 10.1111/j.1365-3083.2005.01694.x
42. Anyona SB, Kempaiah P, Raballah E, Ouma C, Were T, Davenport GC. Functional promoter haplotypes of interleukin-18 condition susceptibility to severe malarial anemia and childhood mortality. Infect Immun. (2011) 79:4923–32. doi: 10.1128/IAI.05601-11
43. Han M, Yue J, Lian YY, Zhao YL, Wang HX, Liu LR. Relationship between single nucleotide polymorphism of interleukin-18 and susceptibility to pulmonary tuberculosis in the Chinese Han population. Microbiol Immunol. (2011) 55:388–93. doi: 10.1111/j.1348-0421.2011.00332.x
44. Hirankarn N, Manonom C, Tangkijvanich P, Poovorawan Y. Association of interleukin-18 gene polymorphism (-607A/A genotype) with susceptibility to chronic hepatitis B virus infection. Tissue Antigens. (2007) 70:160–3. doi: 10.1111/j.1399-0039.2007.00865.x
45. Haas SL, Weiss C, Bugert P, Gundt J, Witt H, Singer MV, et al. Interleukin 18 promoter variants (−137G>C and −607C>A) in patients with chronic hepatitis C: association with treatment response. J Clin Immunol. (2009) 29:620–8. doi: 10.1007/s10875-009-9302-z
46. Franco M, Montenegro MR, Marques RS, Dillon L. Host-parasite relationship in Paracoccidioidomycosis. J Med Vet Mycol. (1987) 25:5–9. doi: 10.1080/02681218780000021
47. Gustincich S, Manfioletti G, Del Sal GS, Schneider G, Carninci PA. A fast method for high quality genomic DNA extraction from whole human blood. Biotechniques. (1991) 11:298–302.
48. Lozano VF, Lins TC, Teixeira MM, Vieira RG, Blotta MHS, Goes AM, et al. Polymorphism analysis of the CTLA-4 gene in paracoccidioidomycosis patients. Mem Inst Oswaldo Cruz. (2011) 106:220–6. doi: 10.1590/S0074-02762011000200017
49. Alves Pereira Neto T, Costa Pereira AA, Costa Hanemann JA, Coelho LFL, Malaquias LCC. DC-SIGN and VDR polymorphisms are associated with chronic form of paracoccidioidomycosis with oral manifestations. Mycoses. (2019) 62:186–92. doi: 10.1111/myc.12866
50. Giedraitis V, He B, Huang WX, Hillert J. Cloning and mutation analysis of the human IL-18 promoter: a possible role of polymorphisms in expression regulation. J Neuroimmunol. (2001) 112:146–52. doi: 10.1016/S0165-5728(00)00407-0
51. Cao W, Luo LL, Chen WW, Liang L, Zhang RR, Zhao YL, et al. Polymorphism in the EREG gene confers susceptibility to tuberculosis. BMC Med Genet. (2019) 20:7. doi: 10.1186/s12881-018-0729-z
52. Dhiman N, Haralambieva IH, Vierkant RA, Pankratz VS, Ryan JE, Jacobson RM, et al. Predominant inflammatory cytokine secretion pattern in response to two doses of live rubella vaccine in healthy vaccinees. Cytokine. (2010) 50:24–9. doi: 10.1016/j.cyto.2009.12.002
53. Pan L, Zhang W, Liang Z, Wu X, Zhu X, Li J, et al. Association between polymorphisms of the cytokine and cytokine receptor genes and immune response to hepatitis B vaccination in a Chinese Han population. J Med Virol. (2012) 84:26–33. doi: 10.1002/jmv.22251
54. Wang J, Tang S, Shen H. Association of genetic polymorphisms in the IL12-IFNG pathway with susceptibility to and prognosis of pulmonary tuberculosis in a Chinese population. Eur J Clin Microbiol Infect Dis. (2010) 29:1291–5. doi: 10.1007/s10096-010-0985-0
55. Awomoyi AA, Nejentsev S, Richardson A, Hull J, Koch O, Podinovskaia M, et al. No association between interferon-gamma receptor-1 gene polymorphism and pulmonary tuberculosis in a Gambian population sample. Thorax. (2004) 59:291–4. doi: 10.1136/thx.2003.013029
56. Velez DR, Hulme WF, Myers JL, Weinberg JB, Levesque MC, Stryjewskiet ME, et al. NOS2A, TLR4, and IFNGR1 interactions influence pulmonary tuberculosis susceptibility in African-Americans. Hum Genet. (2009) 126:643–53. doi: 10.1007/s00439-009-0713-y
57. Shin JG, Park BL, Kim LH, Namgoong S, Kim JO, Chang HS, et al. Association study of polymorphisms in interferon-γ receptor genes with the risk of pulmonary tuberculosis. Mol Med Rep. (2015) 12:1568–78. doi: 10.3892/mmr.2015.3544
58. Falleti E, Fabris C, Toniutto P, Fontanini E, Cussigh A, Caldato M, et al. Genetic polymorphisms of inflammatory cytokines and liver fibrosis progression due to recurrent hepatitis C. Interferon Cytokine Res. (2007) 27:239–46. doi: 10.1089/jir.2006.0062
59. Karra VK, Gumma PK, Chowdhury SJ, Ruttala R, Polipalli SK, Chakravarti A, et al. IL-18 polymorphisms in hepatitis B virus related liver disease. Cytokine. (2015) 73:277–82. doi: 10.1016/j.cyto.2015.02.015
60. Vokurka J, Klapusová L, Pantuckova P, Kukletova M, Kukla L, Holla LI. The association of MMP-9 and IL-18 gene promoter polymorphisms with gingivitis in adolescents. Arch Oral Biol. (2009) 54:172–8. doi: 10.1016/j.archoralbio.2008.09.002
61. Leon Rodriguez DA, Carmona FD, Echeverría LE, González CI, Martin J. IL18 gene variants influence the susceptibility to Chagas disease. PLoS Negl Trop Dis. (2016) 10:e0004583. doi: 10.1371/journal.pntd.0004583
62. Estfanous SZK, Ali SA, Seif SM, Soror SHA, Abdelaziz DHA. Inflammasome genes' polymorphisms in Egyptian chronic hepatitis c patients: influence on vulnerability to infection and response to treatment. Mediators Inflamm. (2019) 2019:3273645. doi: 10.1155/2019/3273645
63. Chen WJ, Yang JY, Lin JH, Fann CSJ, Osyetrov V, King CC, et al. Nasopharyngeal shedding of severe acute respiratory syndrome-associated coronavirus is associated with genetic polymorphisms. Clin Infect Dis. (2006) 42:1561–9. doi: 10.1086/503843
64. Castelar L, Silva MM, Castelli EC, Deghaide NHS, Mendes-Junior CT, Machado AA, et al. Interleukin-18 and interferon-gamma polymorphisms in Brazilian human immunodeficiency virus-1-infected patients presenting with lipodystrophy syndrome. Tissue Antigens. (2010) 76:126–30. doi: 10.1111/j.1399-0039.2010.01471.x
65. Affandi JS, Kumar M, Agarwal U, Singh S, Price P. The search for a genetic factor associating with immune restoration disease in HIV patients co-infected with Mycobacterium tuberculosis. Dis Markers. (2013) 34:445–9. doi: 10.1155/2013/246017
66. Shi BJ, Yu XY, Li H, Xing TH, Fan JW, Wang PW, et al. Association between donor and recipient interleukin-18 gene polymorphisms and the risk of infection after liver transplantation. Clin Invest Med. (2017) 40:E176–87. doi: 10.25011/cim.v40i5.28623
67. Bakr NM, Awad A, A Moustafa E. Association of genetic variants in the interleukin-18 gene promoter with risk of hepatocellular carcinoma and metastasis in patients with hepatitis C virus infection. IUBMB Life. (2018) 70:165–74. doi: 10.1002/iub.1714
68. Sakai K, Kita M, Sawai N, Shiomi S, Sumida Y, Kanemasa K, et al. Levels of interleukin-18 are markedly increased in Helicobacter pylori-infected gastric mucosa among patients with specific IL18 genotypes. J Infect Dis. (2008) 197:1752–61. doi: 10.1086/588196
69. Rezaeifar A, Eskandari-Nasab E, Moghadampour M, Kharazi-Nejad E, Hasani SSA, Asadi-Saghandi A, et al. The association of interleukin-18 promoter polymorphisms and serum levels with duodenal ulcer, and their correlations with bacterial CagA and VacA virulence factors. Scand J Infect Dis. (2013) 45:584–92. doi: 10.3109/00365548.2013.794301
70. Hasan FT, Naif HM. Association of gene polymorphisms and serum levels of IL-18 with the susceptibility to infection with hepatitis B virus. J Infect Dis Med. (2017) 2:3. doi: 10.4172/2576-1420.1000117
71. Ketelut-Carneiro N, Silva GK, Rocha FA, Milanezi CM, Cavalcanti-Neto FF, Zamboni DS, et al. IL-18 triggered by the Nlrp3 inflammasome induces host innate resistance in a pulmonary model of fungal infection. J Immunol. (2015) 194:4507–17. doi: 10.4049/jimmunol.1402321
72. Blotta MHSL, Mamoni RL, Oliveira SJ, Nouér SA, Papaiordanou PMO, Goveia A, et al. Endemic regions of paracoccidioidomycosis in Brazil: a clinical and epidemiologic study of 584 cases in the southeast region. Am J Trop Med Hyg. (1999) 61:390–4. doi: 10.4269/ajtmh.1999.61.390
73. Belissimo-Rodrigues F, Machado AA, Martinez R. Paracoccidioidomycosis epidemiological features of 1000-cases series from a hyperendemic area on the Southeast of Brazil. Am J Trop Med Hyg. (2011) 85:546–50. doi: 10.4269/ajtmh.2011.11-0084
74. Shikanai-Yasuda MA, Mendes RP, Colombo AL, Queiroz-Telles F, Kono ASG, Paniago AMM, et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Rev Soc Bras Med Trop. (2017) 50:715–40. doi: 10.1590/0037-8682-0230-2017
75. Aristizabal BH, Clemons KV, Stevens DA, Restrepo A. Morphological transition of Paracoccidioides brasiliensis conidia to yeast cells: in vivo inhibition in females. Infect Immun. (1998) 66:5587–91. doi: 10.1128/IAI.66.11.5587-5591.1998
76. Pinzan CF, Ruas LP, Casabona-Fortunato AS, Carvalho FC, Roque-Barreira MC. Immunological basis for the gender differences in murine Paracoccidioides brasiliensis infection. PLoS ONE. (2010) 5:e10757. doi: 10.1371/journal.pone.0010757
77. Shankar J, Wu TD, Clemons VK, Monteiro JP, Mirels LF, Stevens DA. Influence of 17-beta-estradiol on gene expression of Paracoccidioides during mycelia-to-yeast transition. PLoS ONE. (2011) 6:e28402. doi: 10.1371/journal.pone.0028402
78. Severo LC, Roesch EW, Oliveira EA, Rocha MM, Londero AT. Paracoccidioidomycosis in women. Rev Iberoam Micol. (1998) 15:88–9.
79. Santos RS, de Fatima LA, Frank AP, Carneiro EM, Clegg DJ. The effects of 17 alpha-estradiol to inhibit inflammation in vitro. Biol Sex Differ. (2017) 8:30. doi: 10.1186/s13293-017-0151-9
Keywords: paracoccidioidomycosis, Paracoccidioides spp., single nucleotide polymophism, IL18, IL12A, IFNGR1, clinical forms of paracoccidioidomycosis
Citation: Sato PK, Busser FD, Carvalho FMC, Gomes dos Santos A, Sadahiro A, Diogo CL, Kono ASG, Moretti ML, Luiz OC and Shikanai-Yasuda MA (2020) Polymorphism in the Promoter Region of the IL18 Gene and the Association With Severity on Paracoccidioidomycosis. Front. Immunol. 11:542210. doi: 10.3389/fimmu.2020.542210
Received: 11 March 2020; Accepted: 21 August 2020;
Published: 01 October 2020.
Edited by:
Adriana Malheiro, Federal University of Amazonas, BrazilReviewed by:
Yean Kong Yong, Xiamen University Malaysia, MalaysiaAnamelia Lorenzetti Bocca, University of Brasilia, Brazil
Copyright © 2020 Sato, Busser, Carvalho, Gomes dos Santos, Sadahiro, Diogo, Kono, Moretti, Luiz and Shikanai-Yasuda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paula Keiko Sato, cGF1bGEuc0BoYy5mbS51c3AuYnI=; Maria Aparecida Shikanai-Yasuda, bWFzeWFzdWRhQHlhaG9vLmNvbS5icg==
†These authors have contributed equally to this work
‡Present address: Flávia Mendes da Cunha Carvalho, Structural Biology Unit, Laboratory of Medical Investigation in Immunopathology of Schistosomiasis and Other Parasitoses (LIM06), Faculdade de Medicina, Hospital das Clínicas, University of São Paulo, São Paulo, Brazil