 Rocío Velázquez
Rocío Velázquez Emiliano Zamora2
Emiliano Zamora2 Luis M. Hernández
Luis M. Hernández Manuel Ramírez
Manuel Ramírez- 1Departamento de Ciencias Biomédicas (Área de Microbiología), Facultad de Ciencias, Universidad de Extremadura, Badajoz, Spain
- 2Estación Enológica, Junta de Extremadura, Almendralejo, Spain
Torulaspora delbrueckii is becoming widely recommended for improving some specific characteristics of wines. However, its impact on wine quality is still far from satisfactory at the winery level, mostly because it is easily replaced by Saccharomyces cerevisiae-like yeasts during must fermentation. New T. delbrueckii killer strains were here isolated and selected for winemaking. They killed S. cerevisiae yeasts and were able to dominate and complete the fermentation of sterile grape must. Sequential yeast inoculation of non-sterile white must with T. delbrueckii followed by S. cerevisiae did not ensure T. delbrueckii dominance or wine quality improvement. Only a single initial must inoculation at high cell concentrations allowed the T. delbrueckii killer strains to dominate and complete the must fermentation to reach above 11% ethanol, but not the non-killer strains. None of the wines underwent malolactic fermentation as long as the must had low turbidity and pH. Although no statistically significant differences were found in the wine quality score, the S. cerevisiae-dominated wines were preferred over the T. delbrueckii-dominated ones because the former had high-intensity fresh fruit aromas while the latter had lower intensity, but nevertheless nice and unusual dried fruit/pastry aromas. Except for ethyl propanoate and 3-ethoxy-1-propanol, which were more abundant in the T. delbrueckii–dominated wines, most of the compounds with fresh fruit odor descriptors, including those with the greatest odor activity values (isoamyl acetate, ethyl hexanoate, and ethyl octanoate), were more abundant in the S. cerevisiae–dominated wines. The low relative concentrations of these fruity compounds made it possible to detect in the T. delbrueckii–dominated wines the low-relative-concentration compounds with dried fruit and pastry odors. An example was γ-ethoxy-butyrolactone which was significantly more abundant in these wines than in those dominated by S. cerevisiae.
Introduction
The non-Saccharomyces yeasts which are usually present in spontaneous must fermentations have been receiving ever more attention by the part of wine microbiologists because some of them can improve wine complexity. The yeasts which have lately been investigated for wine quality improvement belong to Candida, Kloeckera, Hanseniaspora, Zygosaccharomyces, Schizosaccharomyces, Torulaspora, Brettanomyces, Saccharomycodes, Pichia, and Williopsis genera (Jolly et al., 2006). Among them, Torulaspora delbrueckii is probably the most commonly used in winemaking. Controlled inoculation with this yeast is widely recommended for improving the complexity and for enhancing certain specific characteristics of wines (Jolly et al., 2006; Bely et al., 2008; Renault et al., 2009; Azzolini et al., 2012, 2015). This yeast can also be used to increase glycerol (Contreras et al., 2015) and mannoproteins (Comitini et al., 2011; Belda et al., 2015), or to reduce ethanol (Contreras et al., 2015) in the wine. However, its commercial impact on wine quality is still far from satisfactory, mostly because of the difficulty in reliably controlling the desired participating proportion of T. delbrueckii with respect to the other wine yeast species involved in the same must fermentation process, mainly Saccharomyces cerevisiae-like yeasts. It has been reported that the mixed inoculation of T. delbrueckii and S. cerevisiae reduces such off-flavor compounds as volatile acidity, acetaldehyde, and acetoin (Herraiz et al., 1990; Ciani et al., 2006; Bely et al., 2008), and leads to a systematic increase of 2-phenylethanol, terpenols, and lactones (Herraiz et al., 1990; Comitini et al., 2011; Azzolini et al., 2012; Sadoudi et al., 2012). However, results concerning ester production remain confusing. It has been reported that mixed inoculation can increase the total ester concentration (in particular that of isoamyl acetate and ethyl hexanoate, octanoate, and 3-hydroxybutanoate) relative to pure-culture inoculation (Herraiz et al., 1990). But the contrary has also been reported, i.e., that the total ester concentration of mixed inoculations was less than that of a pure S. cerevisiae culture, with a significant reduction in acetate esters, in particular of isoamyl acetate (Comitini et al., 2011; Sadoudi et al., 2012). Similarly, no difference in the overall ester concentrations was found between mixed T. delbrueckii/S. cerevisiae and single S. cerevisiae inoculation, although the level of some esters (ethyl 3-hydroxybutanoate, for instance) was higher in the mixed culture while that of others (such as isoamyl acetate) was lower (Azzolini et al., 2012). These apparently contradictory results concerning ester concentrations may depend on the proportion of each yeast species during must fermentation, or also on the eventual occurrence of malolactic fermentation, neither of which possibilities were discussed in any depth by those authors. Additionally, it has been shown that ester production by T. delbrueckii is strain dependent, and that the aromas resulting from this yeast can differ when it is associated with S. cerevisiae in mixed cultures (Renault et al., 2009).
As most non-Saccharomyces yeasts, T. delbrueckii has less fermentation vigor and a slower growth rate than S. cerevisiae under usual wine fermentation conditions, being quickly overcome by wild or inoculated S. cerevisiae strains (Mauricio et al., 1998; González-Royo et al., 2014). Thus, knowledge about the interactions between Saccharomyces and Torulaspora wine yeasts during wine fermentation needs to be improved to better predict the relative participation of each yeast species (Ciani et al., 2010). The availability of good-fermenting killer T. delbrueckii strains, able to kill the omnipresent wild Saccharomyces yeasts or to control the excessive growth of inoculated S. cerevisiae strains, could be an interesting tool with which to attain the desired domination of each inoculated yeast during must fermentation, and thus result in improved quality of the wine. The isolation of T. delbrueckii killer strains has been described previously (Sangorrin et al., 2007), but they have not been used and analyzed in depth for winemaking as it has been S. cerevisiae K2 strains (Pérez et al., 2001). The effect of S. cerevisiae killer strains on the growth of sensitive strains during must fermentation was seen to depend on the initial proportion of killer yeasts, the susceptibility of sensitive strains, and the treatment of the must. An initial proportion of 2–6% killer yeasts was enough to suppress isogenic sensitive strains in sterile filtered must, although a greater initial proportion of killer yeasts may be needed to get the same effect against non-isogenic strains. The suspended solids that remain in the must after cold-settling were seen to reduce the killer toxin effect due to inactivation by absorption onto the grape particles (Pérez et al., 2001).
The objective of the present work was to evaluate the use of new killer T. delbrueckii strains (Kbarr) for white wine making. We addressed the following issues: (i) capacity of Kbarr strains to dominate and complete must fermentation in the presence of S. cerevisiae yeasts; (ii) influence of must treatment on this Kbarr-1 strain domination; (iii) influence of Kbarr strains on malolactic fermentation; and (iv) analysis of the aroma profile of T. delbrueckii white wine as compared with S. cerevisiae white wine. The usefulness of killer T. delbrueckii strains for winemaking will be discussed.
Materials and Methods
Yeast Strains and Culture Media
EX85, EX85R, and E7AR1 are prototrophic and homothallic S. cerevisiae wine yeasts previously isolated from Spanish wineries, selected for winemaking (Regodón et al., 1997; Ramírez et al., 1998), and sold by Heral Enología SL (Almendralejo, Spain). EX85 is K2-killer, EX85R is virus-free killer-sensitive cycloheximide-resistant (cyhR), and E7AR1 is K2-killer cyhR. The S. cerevisiae K2-killer strains kill other killer-sensitive S. cerevisiae strains but do not kill T. delbrueckii yeasts. The new T. delbrueckii Kbarr wine yeasts are prototrophic strains isolated from spontaneous fermentations of grapes from vineyards of the Albarregas (Barraecas in Latin) river valley in Spain. They kill all S. cerevisiae killer and non-killer strains and the non-killer T. delbrueckii strains. The industrial use of these Kbarr yeasts is under patent application. The yeast strains used in this work are summarized in Table 1.
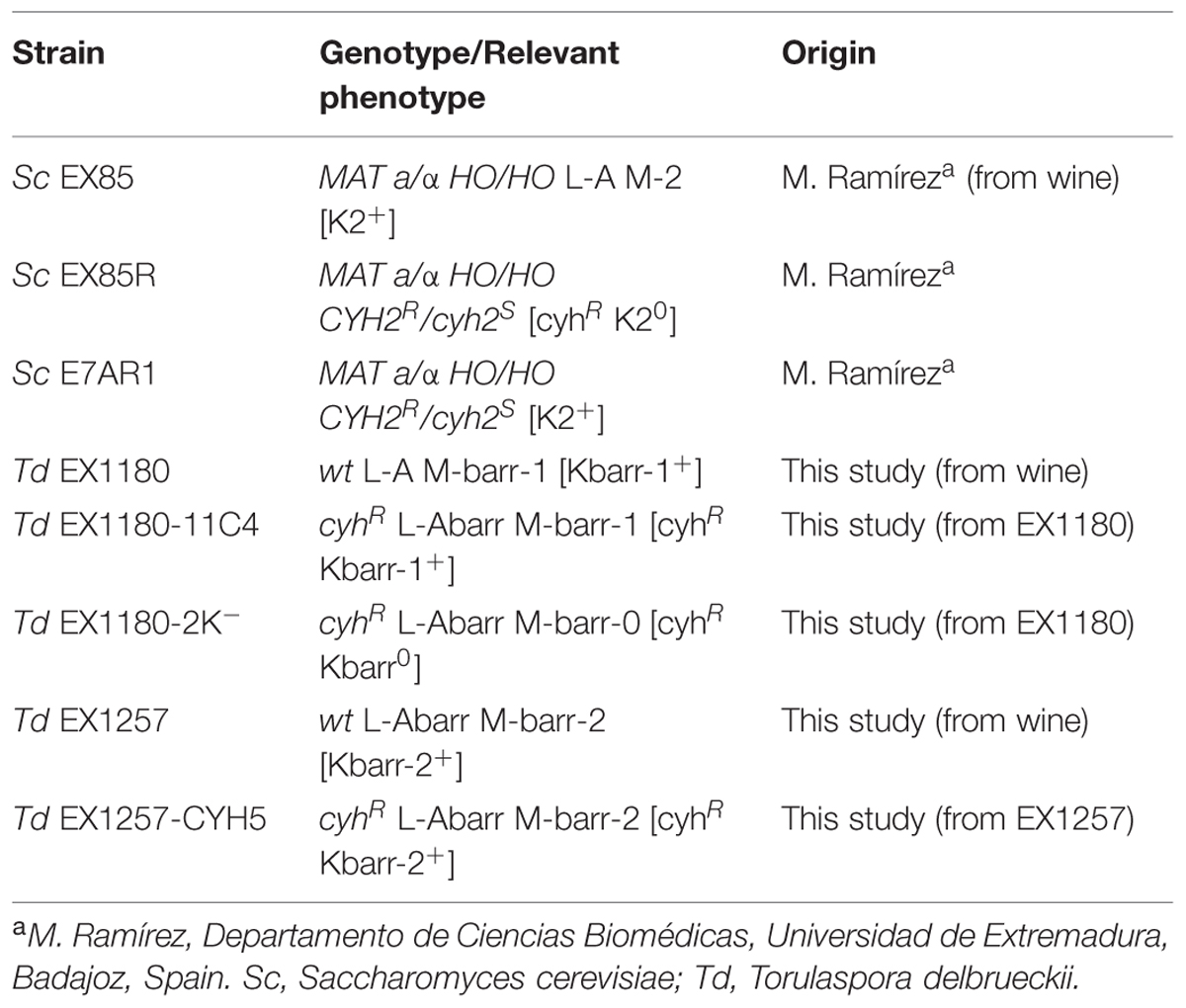
TABLE 1. Yeast strains used.
YEPD + cycloheximide (cyh) is YEPD-agar (1% Bacto-yeast extract, 2% Bacto-peptone, 2% glucose, 2% Bacto-agar) supplemented with cyh, prepared in a concentrated ethanol solution to a final concentration of 2 μg/mL (Pérez et al., 2000). Standard yeast genetics procedures were used for sporulation (Kaiser et al., 1994). Cells were grown on YEPD plates for 2 days at 30°C, transferred to sporulation plates (1% potassium acetate, 0.1% Bacto-yeast extract, 0.05% glucose, 2% Bacto-agar) and incubated for 7–30 days at 25°C.
Determination of Yeast Killer Activity
Killer activity was tested on low-pH (pH 3.3 or 4.0) methylene blue plates (3.3MB or 4.0MB; Kaiser et al., 1994) seeded with 100 μL of a 48-h grown culture of the sensitive strain (Ramírez et al., 2004). Depending on the experiments, the strains being tested for killer activity were either loaded as 4 μL aliquots of stationary phase cultures, patched from solid cultures, or replica plated onto the seeded MB plates. Then the plates were incubated for 4–8 days at 12 or 20°C.
Laboratory Must Fermentation
Must fermentation was carried out in 5-L Erlenmeyer flasks with 3.5 L of Cigüente grape must (18.0°Brix, pH 3.5, 50 mg/L SO2, and 0.3 g/L Actimax nutrients from Productos Agrovin S.A.) sterilized by membrane filtration through a Millipore system (0.45-μm membrane). Yeast cells were cultured in YEPD broth for 2 days at 30°C, washed twice (by centrifugation) with sterile water, and suspended in the must at the desired concentration. Fermentations were conducted at 18°C for 20 days. Yeast growth (determination of total yeast cells by counting with a Neubauer chamber, and viable cells by counting the yeast colonies that arose on YEPD-agar plates), and the °Brix were monitored. All experiments were done in triplicate.
Winery Vinification Trials
The yeast inocula were obtained in a pilot plant of the company Heral Enología SL following its industrial procedure. Cells were cultured in beet molasses broth [5% beet molasses, 0.2% Bacto-yeast extract, 0.075% (NH4)2HPO4, 0.1% MgSO4⋅7H2O, adjusted to pH 3.5 with HCl] for 18 h at 30°C with strong aeration, washed twice (by centrifugation) with sterile distilled water, and inoculated in 350-L stainless steel tanks with cold-settled white Cigüente (19.0–19.8°Brix, pH 3.42, 80–250 NTU, 50 mg/L SO2, and 0.3 g/L Actimax) or Macabeo (20.4–20.8°Brix, pH 3.29–3.55, 80–250 NTU, 50 mg/L SO2, and 0.3 g/L Actimax) grape must to a final concentration of 2–4 × 106 cells/mL for S. cerevisiae and 2–4 × 107 cells/mL for T. delbrueckii. The vinification process was conducted at 16–18°C. The density, °Brix, and yeast growth (total and viable yeast cells) were monitored throughout fermentation. The tanks were hermetically closed when reducing sugars reached around 1% to avoid oxidation problems. At the end of fermentation, the settled solids were discarded. An 800-mL centrifuged sample of each wine was taken for the analytical assays. The wines were stored at 12°C. After 30 days following the beginning of fermentation, settled solids were again discarded, a 2-L sample of each wine was taken for the first aroma compounds and organoleptic assays, and the wines were returned to store at 12°C. At 60 days, settled solids were discarded once again and the second aroma compounds and organoleptic assays were carried out. The organoleptic characteristics (flavor, color, and odor) of the wines were tested by a panel of 12 experts. Wines were presented in clear tulip-shaped wine glasses covered with glass Petri dishes. A sample of 50–70 mL of wine was poured into each glass immediately before being analyzed by each judge. The temperature of the samples was from 10 to 13°C. Sensory profiles of wines were evaluated for overall aromatic complexity, and fresh fruit and dried fruit/pastry aroma intensities. The judges scored the quality of the wines on a six-point scale (0 = very poor, 1 = deficient, 2 = acceptable, 3 = good, 4 = very good, and 5 = excellent). The maximum score possible (60 points) was considered 100% preference. All experiments were done in duplicate.
Determination of the Amount of Inoculated Yeasts during Must Fermentation
Determination of the percentage of genetically marked yeasts was done by the replica-plating method (Pérez et al., 2000). Samples from fermenting musts were diluted and plated onto YEPD-agar to obtain 100–300 colonies per plate. The detection of the cyhR mutants was accomplished by replica-plating these plates to either YEPD + cyh (2 μg/mL) plates using sterile velvets and then to other plates of YEPD-agar to detect wild yeasts sensitive to cyh. The time needed to easily observe growth of resistant yeasts on YEPD + cyh at 30°C varied between 1 and 3 days depending on the yeast strain.
The percentage of wild parent yeasts, or genetically marked yeasts for the replica-plating results validation, was mostly determined by analyzing the mtDNA restriction pattern as previously described (Maqueda et al., 2010).
The yeast spore (after yeast growth on sporulation medium for 7–30 days at 25°C) or vegetative cell morphology were also eventually analyzed for validation of the previous results obtained by the replica-plating or mtDNA restriction pattern analyses. This morphology analysis was done by microscopic observation in a Nikon Eclipse 600 microscope equipped with a Nomarski 60× objective.
Analytical Methods
Density, °Brix, pH, total acidity, volatile acid, reducing sugars, alcohol, and malic acid were determined according to the EC recommended methods (E.C, 1999). Lactic acid was determined using the EEC recommended method (E.E.C, 1990). Glycerol was determined with an enzymatic test (Roche, Germany). Mannoprotein content was measured as previously described (Quiros et al., 2012). T15 is the time needed to ferment 15% of the total sugars present in the must, and T100 is the time needed to ferment 100% of the total sugars (Ramírez et al., 1999).
The wine aroma compounds were analyzed by gas chromatography coupled to a mass detector. The minor aroma compounds were isolated and pre-concentrated following a solid-phase extraction (SPE) procedure (García-Carpintero et al., 2011). The analyses were carried out with an Agilent 6890 N gas chromatograph coupled to a Model 5973 mass detector and equipped with an autosampler. The column was a DB WAXETR (60 m × 0.25 mm, i.d; 0.25 μm film thickness). Quantitative data were obtained by calculating the peak area of each compound relative to that of the internal standard, interpolating with the corresponding calibration plot which had been constructed from the analysis of known amounts of the volatile aroma standards. For those compounds for which the authenticated standards were unavailable (ethyl 9-decenoate, diethyl 2-hydroxyglutarate, ethyl 2-hydroxy-3-phenylpropanoate, and γ-ethoxy-butyrolactone), the identification was based on spectral comparison with the Wiley A library data, and quantification was done using the calibration curves of standards with similar chemical structures obtained in the TIC mode. A total of 75 compounds were detected in the wines elaborated (Supplementary Table S1). The odor descriptor and the odor threshold concentration for each volatile compound were taken from the literature (Etievant and Maarse, 1991; Guth, 1997; Ferreira et al., 2000; Moyano et al., 2002; Zea et al., 2007; Muñoz et al., 2008; Pino and Queris, 2011). The odor activity value (OAV) is the ratio between the concentration of each individual aromatic compound and its odor threshold concentration (the minimal concentration that can be detected by the human nose). As no odor threshold concentration was available for some compounds, 1 mg/L was taken as the value for the ethyl 9-decenoate, ethyl 4-hydroxybutyrate, and 9-decenoic acid OAV calculations, and the value for γ-butyrolactone (0.035 mg/L) was taken for the γ-ethoxy-butyrolactone OAV calculation.
Statistical Analysis
Data were analyzed for statistical significance by a one-way analysis of variance (ANOVA, p < 0.05) with the software package SPSS version 20.0 for Windows (Chicago, IL, USA).
Results
Effect of T. delbrueckii Killer Yeasts on the S. cerevisiae Population during Sterile-must Fermentation
The influence of any given yeast on winemaking will mostly depend on its ability to dominate the must fermentation while reducing the influence of the other participating yeasts. Complementary and reliable methods to monitor the different wine yeasts in the fermenting must are required to accurately determine the degree of domination of each yeast strain. We isolated and characterized new spontaneous cyh-resistant (cyhR) mutants from new T. delbrueckii killer yeasts that had previously been isolated and selected for winemaking (Regodón et al., 1997; Ramírez et al., 2015). Some of these mutants, such as EX1180-2K- for instance, had lost the killer virus to become killer sensitive yeasts, but others, such as EX1180-11C4, retained the virus and the Kbarr-1 phenotype (Table 1). Both of these cyhR mutant types had good must fermentation capabilities, and were easily monitored by simple replica-plating on YEPD-CYH agar. Additionally, these T. delbrueckii killer yeasts can also be distinguished from the always present Saccharomyces by analyzing their cell morphology, spore forming process, or mtDNA RFLPs (Supplementary Figure S1). Moreover, the killer phenotype or viral dsRNA analysis can also be used for this same purpose (not shown). These alternative techniques were satisfactorily used to validate the results obtained from the simple replica-plating assay on YEPD-CYH agar plates.
To determine whether the T. delbrueckii killer yeasts can dominate the must fermentation in the presence of S. cerevisiae wine strains, sterile-must laboratory micro vinifications were inoculated with both yeasts. Each yeast species was monitored through the process by replica-plating on YEPD-CYH and by the aforementioned complementary methods (mostly mtDNA RFLP analysis) in at least two samples for each vinification. The results of the different methods showed full agreement, supporting the utility of cyhR as genetic marker to monitor T. delbrueckii in winemaking, as it was previously found for S. cerevisiae (Pérez et al., 2000; Ambrona et al., 2005, 2006). The must fermentation inoculated with S. cerevisiae alone or with two-yeast mixtures of S. cerevisiae + T. delbrueckii strains (one of them containing the cyhR genetic marker) showed faster kinetics than those single-inoculated with a T. delbrueckii strain, although all fermentations were completed after 10 days (Figure 1A). The total yeast cell concentration increased to above 2 × 108 cells/mL after 3–4 days from the start of fermentation, except for T. delbrueckii killer EX1180-11C4 which reached 2 × 108 cells/mL after 7 days (Figure 1B). The number of viable cells increased in parallel with the number of total cells, except for the vinification of filtered must inoculated with EX85 (10%) + EX1180-11C4 (90%), and for that of cold-settled must inoculated with EX85 (10%) + EX1180-11C4 (90%). In both cases, a decrease in viable cells was observed between days 2 and 4 of fermentation (Figure 1C), indicating that the S. cerevisiae yeasts were killed by the T. delbrueckii killer yeasts. The S. cerevisiae EX85 strain dominated the must fermentation when initially combined with 50% of the non-killer T. delbrueckii EX1180-2K- strain, that fell to 7% after 1 day of fermentation in filtered must (no grape particles present). This time required for S. cerevisiae EX85 domination was extended in filtered must fermentation when it was combined with the same initial proportion (50%) of the T. delbrueckii killer EX1180-11C4, which remained above 20% after 7 days. But this time was reduced again in cold-settled non-filtered must, where EX1180-11C4 disappeared after just 1 day (Figure 1D). A plausible explanation for this behavior is the presence of grape particles through the fermentation, which might adsorb and inactivate the toxin produced by T. delbrueckii, as it was previously shown for toxins produced by S. cerevisiae (Pérez et al., 2001). The S. cerevisiae EX85 strain also dominated the must fermentation when initially combined with 90% of the non-killer T. delbrueckii EX1180-2K-, although this latter strain remained at above 40% for 7 days in filtered must fermentation. On the contrary, the S. cerevisiae EX85 strain almost disappeared when initially combined with 90% of the killer T. delbrueckii EX1180-11C4, which was the dominating yeast throughout fermentation in filtered and in non-filtered grape must (Figure 1D).
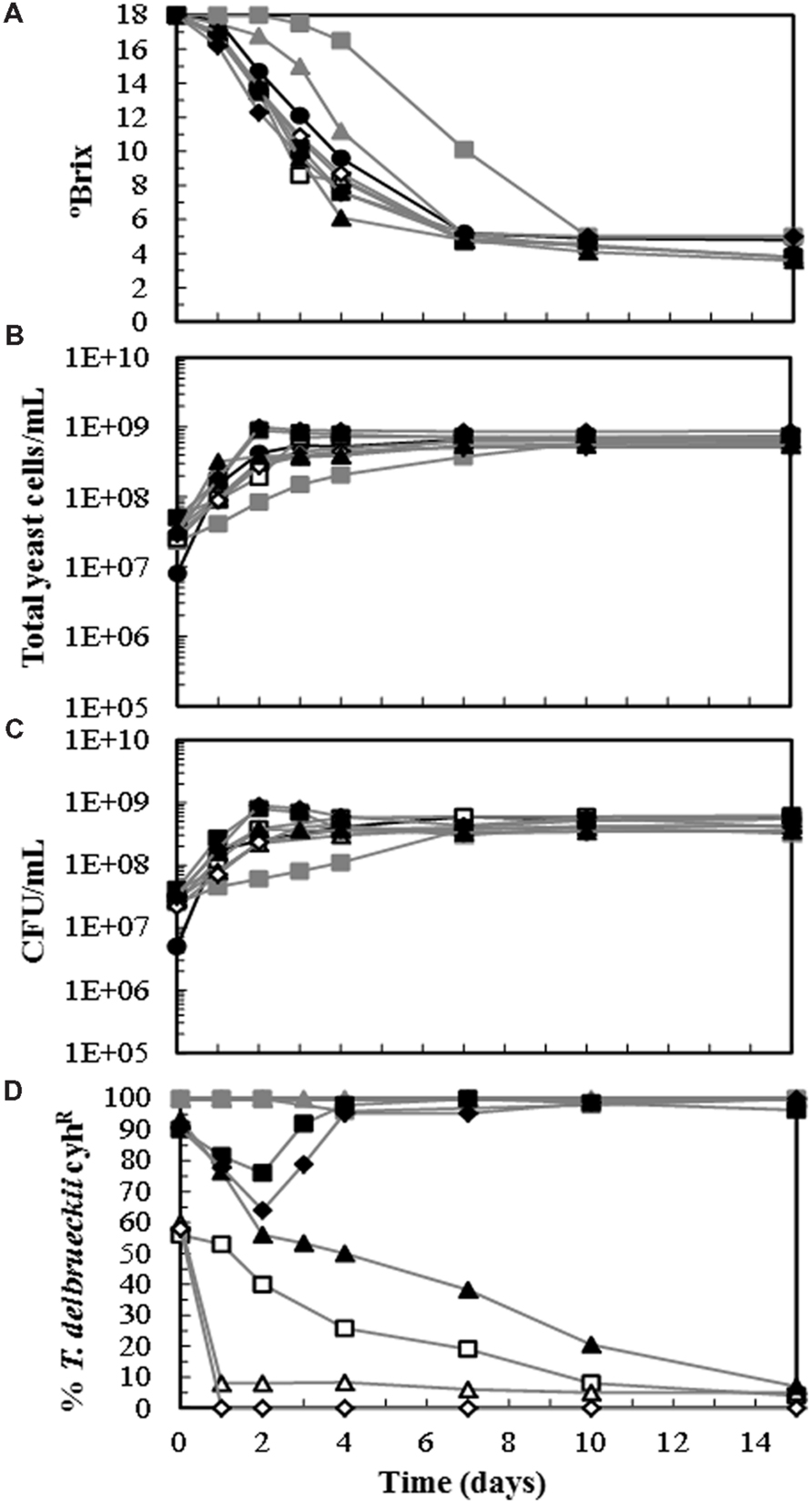
FIGURE 1. Fermentation kinetic and yeast population dynamics of sterile-filtered and cold-settled Cigüente grape must inoculated with different yeast strains of Saccharomyces cerevisiae: EX85 (K2, cyhS), or Torulaspora delbrueckii: EX1180-11C4 (Kbarr-1, cyhR) and EX1180-2K- (non-killer, cyhR). (A) °Brix. (B) Total yeast cells. (C) Viable yeasts. (D) Percentage of T. delbrueckii cyhR yeasts in each fermentation. Symbols: EX1180-11C4 in filtered musts (- -), EX1180-2K- in filtered musts (-
-), EX1180-2K- in filtered musts (- -), EX85 (50%) + EX1180-11C4 (50%) in filtered musts (-
-), EX85 (50%) + EX1180-11C4 (50%) in filtered musts (- -), EX85 (50%) + EX1180-11C4 (50%) cold-settled must (-
-), EX85 (50%) + EX1180-11C4 (50%) cold-settled must (- -), EX85 (50%) + EX1180-2K- (50%) in filtered musts (-
-), EX85 (50%) + EX1180-2K- (50%) in filtered musts (- -), EX85 (10%) + EX1180-11C4 (90%) in filtered musts (-
-), EX85 (10%) + EX1180-11C4 (90%) in filtered musts (- -), EX85 (10%) + EX1180-11C4 (90%) cold-settled must (-
-), EX85 (10%) + EX1180-11C4 (90%) cold-settled must (- -), and EX85 (10%) + EX1180-2K- (90%) in filtered musts (-
-), and EX85 (10%) + EX1180-2K- (90%) in filtered musts (- -).
-).
Winemaking with T. delbrueckii Killer and S. cerevisiae Yeasts
Once we had determined the T. delbrueckii cell concentration required to get its domination during must fermentation, new vinification trials were done using the commonest commercial recommendations: sequential yeast inoculation involving T. delbrueckii at the beginning (2–4 × 107 CFU/mL) followed by S. cerevisiae (2–4 × 106 CFU/mL) after 2 days of fermentation. In most vinifications, the T. delbrueckii viable population decreased to less than 10% of total viable yeast cells after around 1 day following S. cerevisiae inoculation. The wine obtained with these sequential mixed-yeast inoculations showed no relevant aromatic differences from those single-inoculated with a S. cerevisiae strain. This is probably because S. cerevisiae, which became the dominating yeast for most fermentation time, abolished the effect of T. delbrueckii on the wine aromatic compounds during the first two fermentation days. Sometimes, the S. cerevisiae domination was slower and less efficient, remaining more than 30% of T. delbrueckii killer yeasts at the end of a very slow fermentation. These fermentations were frequently not fully completed, mainly in those wines with ethanol concentrations greater than 11.5%. Therefore, in these cases, the wines obtained were not dry since they contained more than 6 g/L of reducing sugars.
In view of these disappointing results, new vinification trials were performed using single inoculation with T. delbrueckii (2–4 × 107 CFU/mL). The S. cerevisiae yeasts present in these fermentations were only those coming into the fresh cold-settled white must (less than 105 CFU/mL). This must was very well clarified (less than 100 NTU turbidity) and its pH was corrected to 3.3 by the addition of tartaric acid. As controls for comparison, vinifications were also performed using only a single initial inoculation with S. cerevisiae (2–4 × 106 CFU/mL). All the grape musts contained around 11°Be, but less than 11.5°Be to avoid the toxic effect of ethanol on T. delbrueckii yeasts and to facilitate the completion of fermentation. The fastest fermentations were always those inoculated with S. cerevisiae, while those inoculated with T. delbrueckii started quickly but slowed down as the ethanol concentration increased, and were very slow by the end of fermentation. Non-inoculated fermentations, performed mostly by wild Saccharomyces yeasts from the must, were the slowest in starting, but they finished the fermentation before the vinifications single-inoculated with T. delbrueckii yeasts (Figures 2A,B; Table 2). The inoculated S. cerevisiae dominated the fermentations (100%) from the beginning to the end. The T. delbrueckii killer strains also dominated the fermentation, although sometimes their proportion decreased to 75% at the end of fermentation (wine density less than 995 g/L, Figures 2C,D). These wines contained a certain amount of reducing sugars (5.98 ± 2.15), especially when this T. delbrueckii-domination was 100% throughout fermentation and no S. cerevisiae ethanol-resistant wild yeasts were present at the end of the process. T. delbrueckii non-killer strain did not dominate the fermentation. Sometimes its proportion decreased quickly to less than 1% or, after decreasing, it remained at a proportion of around 10%. All these T. delbrueckii-non-dominated fermentations were completed, and they were faster than those dominated by T. delbrueckii because S. cerevisiae ethanol-resistant wild yeasts were always present in increasing proportions (Figure 2). The main fermentation aroma of these T. delbrueckii-non-dominated vinifications was fresh fruit, while it was cooked/dried fruit and pastry for the vinifications dominated by T. delbrueckii killer yeasts. The main aroma of the non-inoculated control and the non-killer T. delbrueckii inoculated vinifications was very similar to those single-inoculated and dominated by S. cerevisiae, although the latter had greater fresh-fruit odor intensities.
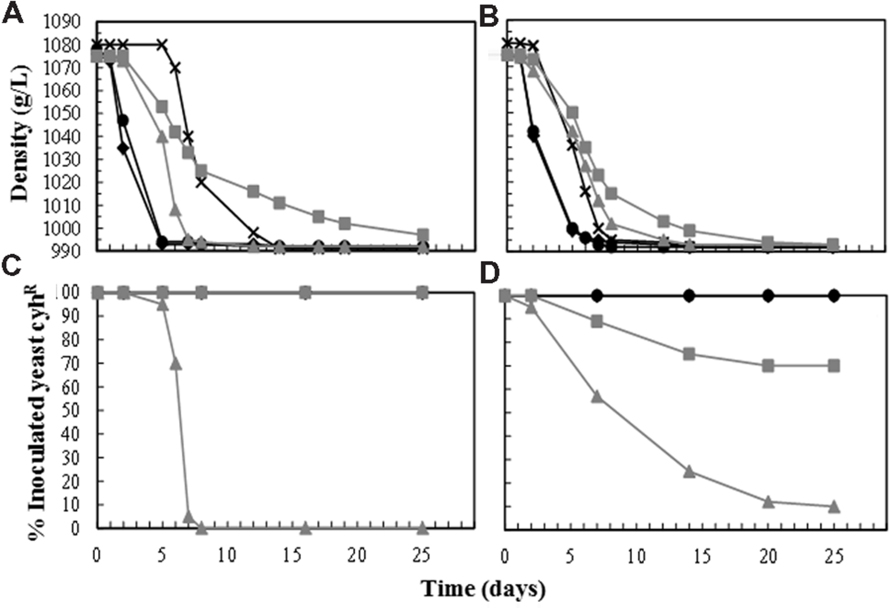
FIGURE 2. Must fermentation kinetics and yeast population dynamics of two independent sets of vinification trials done with two Macabeo grape musts (<100 NTU, pH < 3.5) in 2011 (A,C) and 2012 (B,D). Each yeast was single inoculated in the fresh must at a cell concentration of 2–4 × 106 CFU/mL for the S. cerevisiae strains E7AR1 (K2, cyhR) or EX85R (non-killer, cyhR), and 2–4 × 107 CFU/mL for T. delbrueckii strains EX1180-11C4 (Kbarr-1, cyhR) or EX1180-2K- (non-killer, cyhR). (A,B) Evolution of must-wine density. (C,D) Evolution of the percentage of each inoculated yeast (cyhR) during the must fermentation. Symbols: Non-inoculated control (-×-), E7AR1 (- -), EX85R (-
-), EX85R (- -), EX1180-11C4 (--), and EX1180-2K- (--).
-), EX1180-11C4 (--), and EX1180-2K- (--).
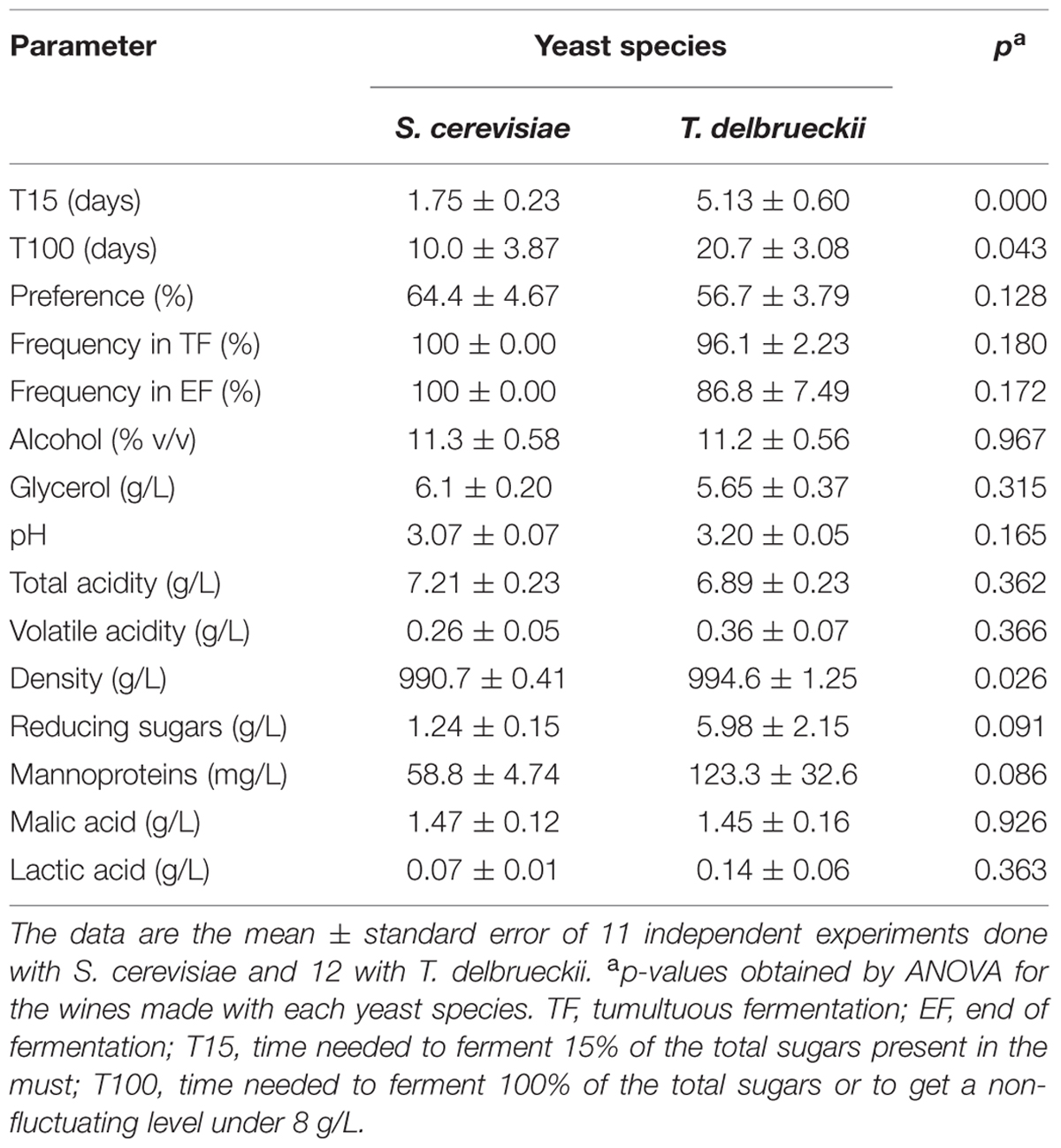
TABLE 2. Must fermentation parameters and white wine analysis results of independent winery vinifications made with Cigüente and Macabeo musts and of an ANOVA to study the effect of single initial inoculation with S. cerevisiae or T. delbrueckii yeasts.
None of these wines underwent malolactic fermentation, even those inoculated with T. delbrueckii killer strains that had slow fermentation kinetics and more than 5 g/L of reducing sugars (Table 2), conditions that usually favor the growth of lactic acid bacteria. However, the wines elaborated with the same grape must but of greater turbidity (around 250 NTU) and higher pH (3.55) did undergo malolactic fermentation (malic acid decreased, while lactic acid increased) when single-inoculated with T. delbrueckii killer yeasts, but not when single-inoculated with S. cerevisiae yeasts. Although both inoculated yeasts dominated the fermentation (100%) throughout the process, the fermentation inoculated with S. cerevisiae was faster than that inoculated with T. delbrueckii killer yeast (taking 7 and 14 days to complete fermentation, respectively; Figure 3).
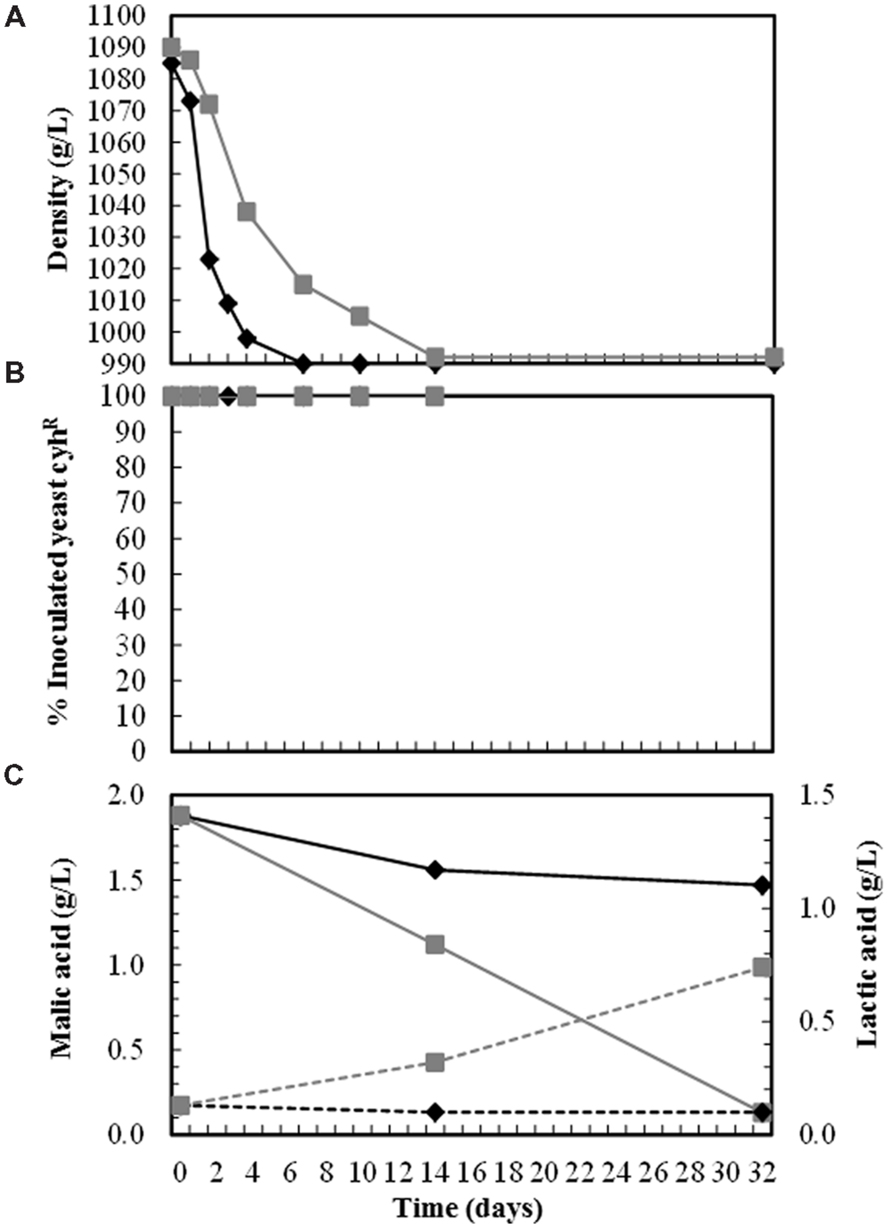
FIGURE 3. Must fermentation kinetics (A), yeast population dynamics (B), and malic/lactic acid degradation/production during the vinification trials done with turbid cold-settled Macabeo grape musts (around 250 NTU, pH 3.55). Each yeast was single inoculated in the fresh must at a cell concentration of 2–4 × 106 CFU/mL for the S. cerevisiae strain E7AR1 (K2, cyhR), and 2–4 × 107 CFU/mL for T. delbrueckii strain EX1180-11C4 (Kbarr-1, cyhR). Symbols: E7AR1 (--), EX1180-11C4 (--), malic acid (—), and lactic acid (- - - -).
Organoleptic and Physicochemical Analysis of the Wines
The wines made with S. cerevisiae or T. delbrueckii for which these yeasts dominated all or most of the fermentation process were compared. In particular, the wines inoculated with non-killer T. delbrueckii yeast that became replaced by wild S. cerevisiae yeast and those that underwent malolactic fermentation were not considered for this analysis. The wine parameter values were consistent with both wine types being non-defective, good-quality products. Significant differences were only found for the fermentation kinetics parameters (T15 and T100), wine density, and marginally significant differences for the amount of reducing sugars and mannoproteins (Table 2). However, although no statistically significant differences were found in the organoleptic quality score, the S. cerevisiae-dominated wines were preferred over the T. delbrueckii-dominated wines because the former had high-intensity fresh fruity aromas. The T. delbrueckii-dominated wines had low-intensity fresh fruit aroma, better flavor complexity, nice but unusual dried fruit (cooked fruit, pastry, and candy) aromas, a little sourness, and some aged/evolved taste. These unusual wine aromas were very similar to the aromas detected during the respective must fermentations of the same wines (see above), but less intense.
The total (summatory) amount of ethyl esters, acetate esters, organic acids, alcohols, monoterpenes, lactones, and carbonyl compounds was greater in the S. cerevisiae than in the T. delbrueckii wines, while the contrary was the case for the amount of furans + volatile phenols and norisoprenoid compounds. However, only the difference found for the amount of organic acids was statistically significant (Figure 4). Nevertheless, significant differences were found for 25 of the 75 volatile compounds analyzed independently (Figure 5). Only the amounts of ethyl propanoate (odor descriptor: banana, apple), 3-ethoxy-1-propanol (fruity), γ-ethoxy-butyrolactone (as with other lactones, probably cooked peach, coconut, caramel, or toasty odor notes), and isobutyric acid (cheese, sour, butter) were significantly greater in T. delbrueckii than in S. cerevisiae wines. In contrast, most compounds were more abundant in S. cerevisiae than in T. delbrueckii wines. These were principally ethyl esters (e.g., ethyl butyrate, ethyl hexanoate, ethyl octanoate, ethyl 3-hydroxybutyrate, ethyl decanoate, ethyl 9-decenoate, ethyl 4-hydroxybutyrate, ethyl laurate, ethyl palmitate) or acetate esters (e.g., isobutyl acetate, isoamyl acetate, hexyl acetate, and phenylethyl acetate), all with fresh fruit odors (Figure 5A). Taking the detection thresholds of these 25 aromatic compounds into account, the greatest OAVs corresponded to three compounds with fresh fruit odor descriptors that were more abundant in the S. cerevisiae than in the T. delbrueckii wines: isoamyl acetate (banana), ethyl hexanoate (banana, green apple), and ethyl octanoate (banana, pineapple, pear, floral; Figure 5B). No significant differences were found for the 75 compounds analyzed between the wines from T. delbrueckii-non-dominated and S. cerevisiae-dominated fermentations (data not shown).
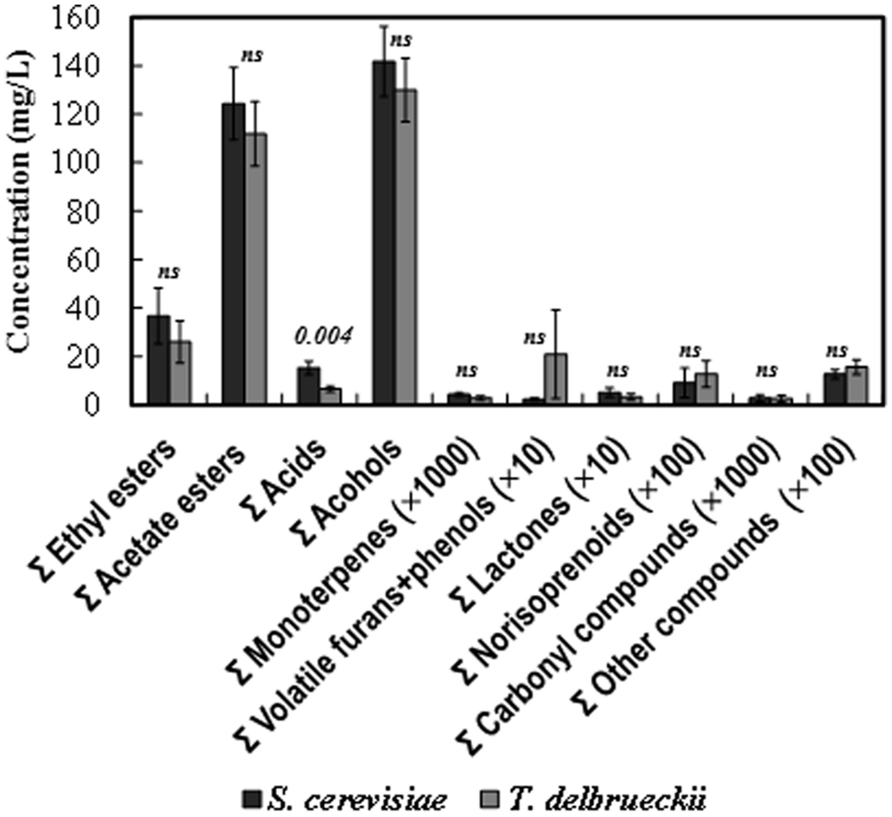
FIGURE 4. Aroma compound composition of the S. cerevisiae-dominated or T. delbrueckii-dominated wines. The amounts for the similar chemical compounds were pooled as summatory. The data are the mean ± standard error of 23 independent vinifications made in duplicate, 11 inoculated with S. cerevisiae and 12 with T. delbrueckii. Statistically significant difference (p) is stated in the top of the bars. ns, no significant difference.
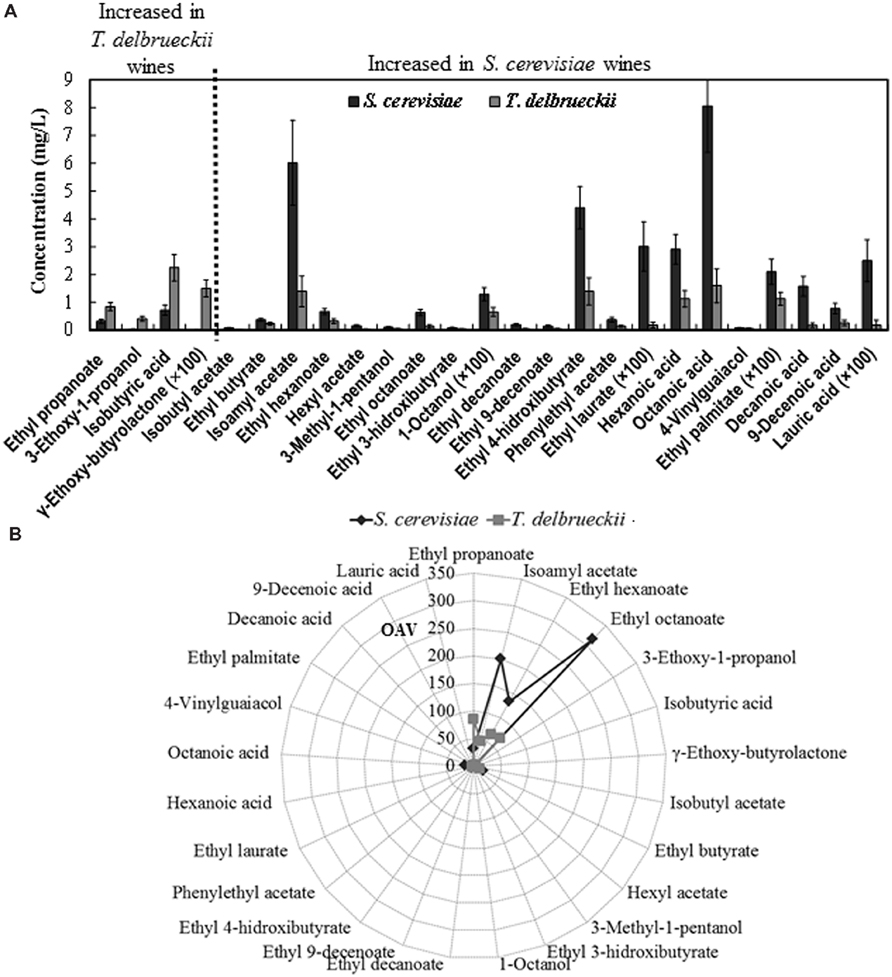
FIGURE 5. (A) Aromatic compounds from which statistically significant difference (p < 0.05) were found between the S. cerevisiae-dominated and T. delbrueckii-dominated wines. The data are the mean ± standard error of 23 independent vinifications made in duplicate, 11 inoculated with S. cerevisiae and 12 with T. delbrueckii. (B) Mean values of the odorant activity values (OAV) for the same compounds in S. cerevisiae-dominated and T. delbrueckii-dominated white wines.
Discussion
Influence of T. delbrueckii Killer Yeasts on the Must Fermentation Process
The new T. delbrueckii killer yeasts were reliably monitored during must fermentation by using spontaneous cyhR mutants, with the results being validated by complementary methods based on molecular polymorphisms or yeast cell morphology. In particular therefore, the results showing that the T. delbrueckii Kbarr-1 strain dominated the low-turbidity (<100 NTU) sterile must fermentation when co-inoculated in a 90% initial proportion with 10% of S. cerevisiae wine strains are reliable. This initial proportion was much greater than that required for the S. cerevisiae killer K2 strain to dominate must fermentation (Pérez et al., 2001), probably because of the faster growth and fermentation rates of S. cerevisiae relative to T. delbrueckii (Mauricio et al., 1998). Increased must turbidity to values that are frequent in industrial wineries (100–250 NTU) had no relevant inhibitory effect on this T. delbrueckii Kbarr-1 domination, and, in particular, much less than the inhibitory effect that had been found previously using S. cerevisiae killer-K2 strains (Pérez et al., 2001). This is probably because the T. delbrueckii Kbarr-1 strains had a more intense killer phenotype than the S. cerevisiae killer-K2 strains (data not shown), and the proportion of the Kbarr-1 toxin that remained unabsorbed onto the grape particles in the turbid must was active enough to kill the 10% of inoculated S. cerevisiae yeast. This T. delbrueckii Kbarr-1 domination decreased or disappeared when the initial proportion was reduced to 50%, or when the T. delbrueckii strain became non-killer. Thus, although the Kbarr-1 killer toxin kills S. cerevisiae and helps T. delbrueckii Kbarr-1 yeasts to dominate must fermentation, a high initial proportion of T. delbrueckii (90%) is required to overcome the greater growth rate of S. cerevisiae in the environmental conditions of the present study.
Sequential yeast inoculation with T. delbrueckii followed by S. cerevisiae did not ensure that the T. delbrueckii domination would continue beyond the first 2 days of fermentation. Most often the viable T. delbrueckii population quickly fell to less than 10% of total viable yeast cells, the aromatic wine profile was similar to those wines which were single-inoculated with S. cerevisiae, and the wines were often not fully dry. As has been shown for assimilable nitrogen limitation (Taillandier et al., 2014), the interference of the growths of the two yeasts could make any given yeast nutrient critically scarce, with the result that the S. cerevisiae population is unable to complete must fermentation under this limiting situation. Therefore, this sequential inoculation strategy does not seem appropriate for winemaking because it does not guarantee any relevant and reproducible effect of T. delbrueckii on wine quality.
Single T. delbrueckii inoculation allowed killer strains to dominate fresh-must fermentation (100–75%), but not the non-killer strains. The T. delbrueckii–dominated fermentations were rather slow at the end, and the resulting wines usually contained some reducing sugars. This was not a relevant issue, however, because part of this sugar was metabolized to reach wine dryness after 20–30 days of wine maturation (data not shown). The presence of low amounts of viable S. cerevisiae ethanol-resistant wild yeasts seems to ensure completion of the fermentation to give dry wines. This could be because there is none of the aforementioned two-yeast-growth interference at this maturation stage since most of the T. delbrueckii cells are dead and cannot secrete the required amount of active killer toxin to kill the ethanol-resistant S. cerevisiae cells. None of these wines presented malolactic fermentation as long as the musts were thoroughly clarified and their pH was 3.3 or lower. However, the T. delbrueckii–dominated wine from the same musts containing more grape particles and pH above 3.5 underwent malolactic fermentation, which is usually undesirable in white table wines. This was probably because of the larger wild bacteria population associated with the solid particles of the turbid must, and because that a pH above 3.5 did not greatly restrict the growth of lactic acid bacteria.
Influence of T. delbrueckii Killer Yeasts on the Organoleptic Quality and Aroma Compounds of the Wines
The main fermentation aroma of the T. delbrueckii–dominated fermentations and the resulting wines, dried/cooked fruit and pastry/candy, did not appear in the wines from T. delbrueckii–non-dominated fermentations, which were very similar to those from S. cerevisiae–dominated fermentations, fresh fruit aroma, as usual for young white wines. These results were coherent with the significant differences in the content of 25 aroma compounds found in the two wine types. Most of the compounds with fresh fruit odor descriptors were more abundant in the S. cerevisiae–dominated wines, including those with the greatest OAVs: isoamyl acetate, ethyl hexanoate, and ethyl octanoate (Figure 5B). However, no significant differences were found for the sum of compounds believed to be responsible for a dried/cooked fruit aroma, such as lactones (Hernandez-Orte et al., 2008; Azzolini et al., 2012; Figure 4), although a significantly greater amount of γ-ethoxy-butyrolactone was found in the T. delbrueckii–dominated than in the S. cerevisiae–dominated wines (Figure 5). However, a greater amount of ethyl 4-hydroxybutanoate (meringue) was detected in the S. cerevisiae–dominated wines. While this can potentially be responsible for some pastry odor, no such odor was detected in these wines by the trained judges. An explanation for these apparently contradictory results could be that, in the S. cerevisiae–dominated wines, the main compounds which had fresh-fruit-odor descriptors overcame the possibility of detecting the minor compounds which had dried fruit or pastry odor descriptors. On the contrary, the relative low concentrations of fresh-fruit-odor compounds in the T. delbrueckii–dominated wines made it possible to detect the dried fruit and pastry odors. Additionally, the slightly sour and evolved/aged flavor detected in the T. delbrueckii–dominated wines but not in the S. cerevisiae–dominated wines may have been due to the greater isobutyric acid concentration in the former (Figure 5), and which would be coherent with previous findings (Herraiz et al., 1990).
Overall, our results are partially in agreement with those previously reported for the influence of T. delbrueckii on the wine quality and aroma compound concentrations. The appearance of dried fruit/coconut aromas associated with the increase in some lactones and the decrease in some ethyl and acetate esters has also been observed in T. delbrueckii wine from synthetic white must (Hernandez-Orte et al., 2008). Similarly, the decrease in isoamyl acetate and ethyl esters of C4–C10 fatty acids has also been noted in T. delbrueckii dry white wine from Soave and Chardonnay grape musts, as well as in sweet “Vino Santo” wine from dried Nosiola grapes (Azzolini et al., 2015), although increases in lactones were found only in this last case. Also similarly to our results, that work’s T. delbrueckii dry wine had significantly lower freshness and acidity but higher flavor intensity, complexity, and persistence than the S. cerevisiae wines. The increased amount of lactones in the “Vino Santo” dessert wine was assumed to improve its organoleptic quality, although this point was not confirmed (Azzolini et al., 2015). Most esters were also found at much lower concentrations in T. delbrueckii than in S. cerevisiae Sauvignon Blanc dry wines (Renault et al., 2015), although some “minor” esters were considered as produced preferentially by T. delbrueckii, in particular ethyl propanoate (in agreement with our findings), ethyl isobutanoate, and ethyl dihydrocinnamate. Additionally, but contrary to our findings, isobutyl acetate and isoamyl acetate concentrations were systematically greater with mixed T. delbrueckii/S. cerevisiae inoculation although this increase did not correlate with the growth of either species, suggesting that this ester concentration enhancement was due to S. cerevisiae production in response to the presence of T. delbrueckii (Renault et al., 2015). This increase in isoamyl acetate (banana note) related to T. delbrueckii inoculation is rather unexpected given that the contrary has been reported several times (Comitini et al., 2011; Azzolini et al., 2012; Sadoudi et al., 2012), and there has also been a report of greater hydrolytic activity of isoamyl acetate (via esterase) with T. delbrueckii than with S. cerevisiae (Plata et al., 2003). In contrast, the increase in ethyl propanoate, ethyl isobutanoate, and ethyl dihydrocinnamate is in agreement with previous findings (Herraiz et al., 1990; Moreno et al., 1991; Plata et al., 2003; Hernandez-Orte et al., 2008; Renault et al., 2009; Sadoudi et al., 2012) and with this present work for the case of ethyl propanoate. One can find additional apparent disagreements in the literature for the relative amounts of other compounds produced by T. delbrueckii relative to S. cerevisiae, but those compounds are not thought to be as relevant for wine aroma as the aforementioned esters and lactones. Although these disagreements could be due to the different yeast strains inoculated in the winemaking (Renault et al., 2009), we did not find any significant differences among the T. delbrueckii strains used in this present study (data not shown). Therefore, we think that other vinification parameters are responsible for the disagreements, especially the degree of dominance of the inoculated yeasts because the S. cerevisiae-dominated wines had similar aroma profiles independently of whether or not they had previously been inoculated and partially fermented with T. delbrueckii. Only the wines from those vinifications inoculated and clearly dominated by T. delbrueckii had a differentiated aroma profile. We cannot evaluate the possible influence of the occurrence of malolactic fermentation on the T. delbrueckii wine because this aspect has as yet to be taken into account in any depth in previous studies.
In sum, it seems that T. delbrueckii has some common effects on wine quality and aroma composition independently of the winemaking condition as long as it is the most relevant yeast species during fermentation. These effects are reduction of the main ester concentrations, increase of some minor ethyl esters and lactone concentrations, and reduction of fresh fruit aromas. However, this yeast can lead to the production of some interesting wine aromas depending on the must type, the yeast inoculation procedure, the degree of the inoculated yeast’s dominance, yeast strain, etc. This variable behavior may determine the wine quality score given by the judges in the sensory evaluation. Therefore, further research on this topic is required to determine the best procedure for the use of T. delbrueckii at winery level in order to ensure the expected effect of this yeast on commercial wines’ complexity.
Notwithstanding this finding of variability in the T. delbrueckii wine aroma composition, a clear conclusion that can be drawn from this work is that the new T. delbrueckii killer strains had the additional advantage of dominating must fermentation in the presence of S. cerevisiae relative to the non-killer strains. They significantly decreased the amounts of the main ethyl and acetate ester compounds responsible for a fresh fruit wine aroma, while increasing some minor ethyl ester and lactone compounds that may be responsible for an improved wine complexity. These killer yeasts can be easily and reliably monitored during must fermentation by the incorporated cyhR genetic marker, cell/spore morphology, or molecular polymorphism analyses. Also, they were able to complete the must fermentation of white wines with less than 11.5% ethanol when single inoculated in low-turbidity low-pH must without favoring the growth of lactic acid bacteria.
Author Contributions
MR conceived the project. MR, RV, MÁ, and EZ designed and performed the experiments. MR, RV, EZ, and LH analyzed the data. MR wrote and edited the manuscript.
Conflict of InterestStatement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was funded by grants GR10088 from the Extremadura Regional Government and AGL2011-25711 from the Spanish Ministry of Education and Science. RV was the recipient of a studentship from the Extremadura Regional Government. We are grateful for the technical and human support provided by the SAIUEx Facility of Elemental and Molecular Analysis (financed by UEX, Junta de Extremadura, MICINN, FEDER, and FSE) for the analyses of the wine aroma compounds.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2015.01222
References
Ambrona, J., Maqueda, M., Zamora, E., and Ramírez, M. (2005). Sulfometuron resistance as genetic marker for yeast populations in wine fermentations. J. Agric. Food Chem. 53, 7438–7443. doi: 10.1021/jf0514468
Ambrona, J., Vinagre, A., Maqueda, M., Álvarez, M. L., and Ramírez, M. (2006). Rhodamine-pink as genetic marker for yeast populations in wine fermentations. J. Agric. Food Chem. 54, 2977–2984. doi: 10.1021/jf052844d
Azzolini, M., Fedrizzi, B., Tosi, E., Finato, F., Vagnoli, P., Scrinzi, C., et al. (2012). Effects of Torulaspora delbrueckii and Saccharomyces cerevisiae mixed cultures on fermentation and aroma of Amarone wine. Euro. Food Res. Technol. 235, 303–313. doi: 10.1007/s00217-012-1762-3
Azzolini, M., Tosi, E., Lorenzini, M., Finato, F., and Zapparoli, G. (2015). Contribution to the aroma of white wines by controlled Torulaspora delbrueckii cultures in association with Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 31, 277–293. doi: 10.1007/s11274-014-1774-1
Belda, I., Navascues, E., Marquina, D., Santos, A., Calderon, F., and Benito, S. (2015). Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 99, 1911–1922. doi: 10.1007/s00253-014-6197-2
Bely, M., Stoeckle, P., Masneuf-Pomarède, I., and Dubourdieu, D. (2008). Impact of mixed Torulaspora delbrueckii–Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 122, 312–320. doi: 10.1016/j.ijfoodmicro.2007.12.023
Ciani, M., Beco, L., and Comitini, F. (2006). Fermentation behaviour and metabolic interactions of multistarter wine yeast fermentations. Int. J. Food Microbiol. 108, 239–245. doi: 10.1016/j.ijfoodmicro.2005.11.012
Ciani, M., Comitini, F., Mannazzu, I., and Domizio, P. (2010). Controlled mixed culture fermentation: a new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 10, 123–133. doi: 10.1111/j.1567-1364.2009.00579.x
Comitini, F., Gobbi, M., Domizio, P., Romani, C., Lencioni, L., Mannazzu, I., et al. (2011). Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 28, 873–882. doi: 10.1016/j.fm.2010.12.001
Contreras, A., Hidalgo, C., Schmidt, S., Henschke, P. A., Curtin, C., and Varela, C. (2015). The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 205, 7–15. doi: 10.1016/j.ijfoodmicro.2015.03.027
E.C. (1999). N° 761. Amending regulation EEC N° 2676/90 determining community methods for the analysis of wines. Official J. Euro. Commun. 5–9.
E.E.C. (1990). N° 2676. Métodos de análisis comunitarios aplicables en el sector del vino. Official Rep. Euro. Commun. L 272, 191–191.
Etievant, P. X., and Maarse, H. (1991). “Wine,” in Volatile Compounds in Foods and Beverages, ed. H. Maarse (New York, NY: Marcel Dekker, Inc.), 483–532.
Ferreira, V., López, R., and Cacho, J. F. (2000). Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 80, 1659–1667.
García-Carpintero, E. G., Sánchez-Palomo, E., and González-Viñas, M. A. (2011). Aroma characterization of red wines from cv. Bobal grape variety grown in La Mancha region. Food Res. Int. 44, 61–70. doi: 10.1016/j.foodres.2010.11.013
González-Royo, E., Pascual, O., Kontoudakis, N., Esteruelas, M., Esteve-Zarzoso, B., Mas, A., et al. (2014). Oenological consequences of sequential inoculation with non-Saccharomyces yeasts (Torulaspora delbrueckii or Metschnikowia pulcherrima) and Saccharomyces cerevisiae in base wine for sparkling wine production. Euro. Food Res. Technol. 240, 999–1012. doi: 10.1007/s00217-014-2404-8
Guth, H. (1997). Quantitation and sensory studies of character impact odorants of different white wine varieties. J. Agric. Food Chem. 45, 3027–3032. doi: 10.1021/jf9608433
Hernandez-Orte, P., Cersosimo, M., Loscos, N., Cacho, J., Garcia-Moruno, E., and Ferreira, V. (2008). The development of varietal aroma from non-floral grapes by yeasts of different genera. Food Chem. 107, 1064–1077. doi: 10.1016/j.foodchem.2007.09.032
Herraiz, G., Reglero, M., Herraiz, P., Alvarez, M., and Cabezudo, M. (1990). The influence of the yeast and type of culture on the volatile composition of wine fermented without sulphur dioxide. Am. J. Enol. Viticult. 41, 313–318.
Jolly, N. P., Augustyn, O. P. H., and Pretorius, I. S. (2006). The effect of non-Saccharomyces yeasts in wine production. S. Afr. J. Enol. Vitic. 27, 15–39.
Kaiser, C., Michaelis, S., and Mitchell, A. (1994). Methods in Yeast Genetics. New York, NY: Cold Spring Harbor Laboratory Press.
Maqueda, M., Zamora, E., Rodríguez-Cousiño, N., and Ramírez, M. (2010). Wine yeast molecular typing using a simplified method for simultaneously extracting mtDNA, nuclear DNA and virus dsRNA. Food Microbiol. 27, 205–209. doi: 10.1016/j.fm.2009.10.004
Mauricio, J. C., Millán, C., and Ortega, J. M. (1998). Influence of oxygen on the biosynthesis of cellular fatty acids, sterols and phospholipids during alcoholic fermentation by Saccharomyces cerevisiae and Torulaspora delbrueckii. World J. Microbiol. Biotechnol. 14, 405–410. doi: 10.1023/A:1008873430077
Moreno, J., Millan, M. C., Ortega, J. M., and Medina, M. (1991). Analytical differentiation of wine fermentations using pure and mixed yeast cultures. J. Ind. Microbiol. 7, 181–190. doi: 10.1007/BF01575881
Moyano, L., Zea, L., Moreno, J., and Medina, M. (2002). Analytical study of aromatic series in sherry wines subjected to biological aging. J. Agric. Food Chem. 50, 7356–7361. doi: 10.1021/jf020645d
Muñoz, D., Peinado, R., Medina, M., and Moreno, J. (2008). Effect of Saccharomyces cerevisiae F12 on volatile compounds in wines at three different stages of industrial biological ageing. Aus. J. Grape Wine Res. 14, 71–77. doi: 10.1111/j.1755-0238.2008.00012.x
Pérez, F., Ramírez, M., and Regodón, J. A. (2001). Influence of killer strains of Saccharomyces cerevisiae on wine fermentation. Antonie van Leeuwenhoek J. Microbiol. 79, 393–399. doi: 10.1023/A:1012034608908
Pérez, F., Regodón, J. A., Valdés, M. E., De Miguel, C., and Ramírez, M. (2000). Cycloheximide resistance as marker for monitoring yeasts in wine fermentations. Food Microbiol. 17, 119–128. doi: 10.1006/fmic.1999.0271
Pino, J. A., and Queris, O. (2011). Analysis of volatile compounds of mango wine. Food Chem. 125, 1141–1146. doi: 10.1007/s12088-010-0028-7
Plata, C., Millan, C., Mauricio, J., and Ortega, J. (2003). Formation of ethyl acetate and isoamyl acetate by various species of wine yeasts. Food Microbiol. 20, 217–224. doi: 10.5803/jsfm.20.217
Quiros, M., Gonzalez, R., and Morales, P. (2012). A simple method for total quantification of mannoprotein content in real wine samples. Food Chem. 134, 1205–1210. doi: 10.1016/j.foodchem.2012.02.168
Ramírez, M., Pérez, F., and Regodón, J. A. (1998). A simple and reliable method for hybridization of homothallic wine strains of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 64, 5039–5041.
Ramírez, M., Regodon, J. A., Pérez, F., and Rebollo, J. E. (1999). Wine yeast fermentation vigor may be improved by elimination of recessive growth-retarding alleles. Biotechnol. Bioeng. 65, 212–218.
Ramírez, M., Velázquez, R., Maqueda, M., López-Piñeiro, A., and Ribas, J. C. (2015). A new wine Torulaspora delbrueckii killer strain with broad antifungal activity and its toxin-encoding double-stranded RNA virus. Front. Microbiol. 6:983. doi: 10.3389/fmicb.2015.00983
Ramírez, M., Vinagre, A., Ambrona, J., Molina, F., Maqueda, M., and Rebollo, J. E. (2004). Genetic instability of heterozygous hybrid populations of natural wine yeasts. Appl. Environ. Microbiol. 70, 4686–4691. doi: 10.1128/AEM.70.8.4686-4691.2004
Regodón, J. A., Pérez, F., Valdés, M. E., De Miguel, C., and Ramírez, M. (1997). A simple and effective procedure for selection of wine yeast strains. Food Microbiol. 14, 247–254. doi: 10.1006/fmic.1996.0091
Renault, P., Coulon, J., De Revel, G., Barbe, J.-C., and Bely, M. (2015). Increase of fruity aroma during mixed T. delbrueckii/S. cerevisiae wine fermentation is linked to specific esters enhancement. Int. J. Food Microbiol. 207, 40–48. doi: 10.1016/j.ijfoodmicro.2015.04.037
Renault, P., Miot-Sertier, C., Marullo, P., Hernández-Orte, P., Lagarrigue, L., Lonvaud-Funel, A., et al. (2009). Genetic characterization and phenotypic variability in Torulaspora delbrueckii species: potential applications in the wine industry. Int. J. Food Microbiol. 134, 201–210. doi: 10.1016/j.ijfoodmicro.2009.06.008
Sadoudi, M., Tourdot-Maréchal, R., Rousseaux, S., Steyer, D., Gallardo-Chacón, J.-J., Ballester, J., et al. (2012). Yeast–yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 32, 243–253. doi: 10.1016/j.fm.2012.06.006
Sangorrin, M. P., Lopes, C. A., Giraudo, M. R., and Caballero, A. C. (2007). Diversity and killer behaviour of indigenous yeasts isolated from the fermentation vat surfaces in four Patagonian wineries. Int. J. Food Microbiol. 119, 351–357. doi: 10.1016/j.ijfoodmicro.2007.04.010
Taillandier, P., Lai, Q. P., Julien-Ortiz, A., and Brandam, C. (2014). Interactions between Torulaspora delbrueckii and Saccharomyces cerevisiae in wine fermentation: influence of inoculation and nitrogen content. World J. Microbiol. Biotechnol. 30, 1959–1967. doi: 10.1007/s11274-014-1618-z
Keywords: Torulaspora delbrueckii, yeast, killer, must fermentation, winemaking, white table wine, aroma compounds
Citation: Velázquez R, Zamora E, Álvarez ML, Hernández LM and Ramírez M (2015) Effects of new Torulaspora delbrueckii killer yeasts on the must fermentation kinetics and aroma compounds of white table wine. Front. Microbiol. 6:1222. doi: 10.3389/fmicb.2015.01222
Received: 04 September 2015; Accepted: 19 October 2015;
Published: 03 November 2015.
Edited by:
Alberto Mas, Universitat Rovira i Virgili, SpainReviewed by:
Catarina Prista, Instituto Superior de Agronomia, PortugalAntonio Santos, Complutense University of Madrid, Spain
Copyright © 2015 Velázquez, Zamora, Álvarez, Hernández and Ramírez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuel Ramírez, bXJhbWlyZXpAdW5leC5lcw==.