 Vincent Le Moigne1
Vincent Le Moigne1 Audrey Bernut2†
Audrey Bernut2† Mélanie Cortès3
Mélanie Cortès3 Albertus Viljoen2
Albertus Viljoen2 Christian Dupont2Alexandre Pawlik4Jean-Louis Gaillard1,5
Christian Dupont2Alexandre Pawlik4Jean-Louis Gaillard1,5 Fabienne Misguich1
Fabienne Misguich1 Frédéric Crémazy1*
Frédéric Crémazy1* Laurent Kremer2,6
Laurent Kremer2,6 Jean-Louis Herrmann1,5*
Jean-Louis Herrmann1,5*- 12I, UVSQ, INSERM, Université Paris-Saclay, Versailles, France
- 2UMR 9004, Centre National de la Recherche Scientifique, Institut de Recherche en Infectiologie de Montpellier, Université de Montpellier, Montpellier, France
- 3VitamFero, Tours, France
- 4Unité de Pathogénomique Mycobactérienne, Institut Pasteur, Paris, France
- 5APHP, GHU PIFO, Hôpital Raymond-Poincaré – Hôpital Ambroise-Paré, Boulogne-Billancourt, France
- 6INSERM, Institut de Recherche en Infectiologie de Montpellier, Montpellier, France
Mycobacterium abscessus, a pathogen responsible for severe lung infections in cystic fibrosis patients, exhibits either smooth (S) or rough (R) morphotypes. The S-to-R transition correlates with inhibition of the synthesis and/or transport of glycopeptidolipids (GPLs) and is associated with an increase of pathogenicity in animal and human hosts. Lsr2 is a small nucleoid-associated protein highly conserved in mycobacteria, including M. abscessus, and is a functional homolog of the heat-stable nucleoid-structuring protein (H-NS). It is essential in Mycobacterium tuberculosis but not in the non-pathogenic model organism Mycobacterium smegmatis. It acts as a master transcriptional regulator of multiple genes involved in virulence and immunogenicity through binding to AT-rich genomic regions. Previous transcriptomic studies, confirmed here by quantitative PCR, showed increased expression of lsr2 (MAB_0545) in R morphotypes when compared to their S counterparts, suggesting a possible role of this protein in the virulence of the R form. This was addressed by generating lsr2 knock-out mutants in both S (Δlsr2-S) and R (Δlsr2-R) variants, demonstrating that this gene is dispensable for M. abscessus growth. We show that the wild-type S variant, Δlsr2-S and Δlsr2-R strains were more sensitive to H2O2 as compared to the wild-type R variant of M. abscessus. Importantly, virulence of the Lsr2 mutants was considerably diminished in cellular models (macrophage and amoeba) as well as in infected animals (mouse and zebrafish). Collectively, these results emphasize the importance of Lsr2 in M. abscessus virulence.
Introduction
Mycobacterium abscessus is a rapidly growing mycobacterium (RGM) increasingly acknowledged as a serious non-tuberculous mycobacterial (NTM) pathogen (Mougari et al., 2016; Diel et al., 2017). Although it causes extrapulmonary infections (Jeong et al., 2017) and disseminated pulmonary diseases among otherwise healthy individuals (Varghese et al., 2012), it has become notorious for the serious threat it poses to patients with cystic fibrosis (CF) and with other underlying lung disorders (Brown-Elliott and Wallace, 2002; Medjahed et al., 2010). M. abscessus infection correlates with a decline in pulmonary function and is an appreciable concern for CF patients requiring lung transplantation (Esther et al., 2010; Qvist et al., 2016; Smibert et al., 2016). M. abscessus has high intrinsic levels of resistance to many antibiotics, making infections with this mycobacterium hard to treat and eradicate (Brown-Elliott and Wallace, 2002; van Dorn, 2017).
Mycobacterium abscessus presents either smooth (S) or rough (R) colony morphotypes leading to different clinical outcomes (Howard et al., 2006; Catherinot et al., 2009; Medjahed and Reyrat, 2009; Ripoll et al., 2009; Roux et al., 2011). They also exhibit different morphological aspects (Howard et al., 2006; Sánchez-Chardi et al., 2011) and virulence phenotypes (Byrd and Lyons, 1999; Catherinot et al., 2007; Bernut et al., 2014a). These differences rely mainly on the presence (in S) or absence (in R) of surface-associated glycopeptidolipids (GPL) (Gutiérrez et al., 2018). The S variant is thought to be the colonizing form, capable of producing mature biofilms and exhibiting a significant motility on soft agar (Howard et al., 2006). The R variant, on the other hand, is impaired in these abilities and forms pronounced serpentine cords, a feature associated with hypervirulence (Byrd and Lyons, 1999; Catherinot et al., 2007; Bernut et al., 2014a; Halloum et al., 2016). Being the most pathogenic RGM, it is not surprising that both variants of M. abscessus resist phagocytosis by immune cells, a trait shared with pathogenic slow-growing mycobacterial (SGM) species, such as M. tuberculosis (Oberley-Deegan et al., 2010; Nessar et al., 2011; Roux et al., 2016). Therefore, the S-to-R transition, which occurs essentially in vivo and is responsible for a significant increase in pathogenicity (Jönsson et al., 2007; Rottman et al., 2007; Bernut et al., 2014b) likely represents an evolutionary adaptation mechanism to the host immune response (Pawlik et al., 2013; Roux et al., 2016).
Genome comparison of three distinct isogenic S/R couples of M. abscessus revealed the presence of genetic lesions in the R variant, such as single nucleotide polymorphisms and/or insertions/deletions, in genes belonging to the GPL biosynthesis and transport locus (Pawlik et al., 2013) and causing the S-to-R transition. Although these mutations can account for the change in colony morphology of M. abscessus, it is unlikely that they fully explain the virulence of the R variant observed in multiple cellular and animal models (Catherinot et al., 2007; Bernut et al., 2014b, 2017). This view is supported by transcriptomic studies performed in the same isogenic S/R pairs, which revealed differential expression for a large set of genes, including lsr2 (Pawlik et al., 2013).
Lsr2 is a nucleoid-associated protein (NAP) and a functional analog of the heat-stable nucleoid-structuring proteins or H-NS (Chen et al., 2008; Gordon et al., 2008; Qu et al., 2013) that is conserved in all actinomycetes and mycobacteria (Chen et al., 2008; Gordon et al., 2008; Qu et al., 2013). Lsr2 was originally shown to be an immunodominant antigen of Mycobacterium leprae (Laal et al., 1991) and has since been reported to regulate a broad range of processes (Chen et al., 2008; Colangeli et al., 2009; Gordon et al., 2010). In Mycobacterium smegmatis, lsr2 mutants show a different colony morphology and are defective in biofilm formation, a phenotype presumably resulting from an altered expression of key surface lipids, such as mycolyl-diacylglycerols (Chen et al., 2006; Arora et al., 2008; Kocíncová et al., 2008; Yang et al., 2017). Subsequent studies also demonstrated that Lsr2 is necessary for mycobacterial conjugal DNA transfer (Nguyen et al., 2010). In contrast to M. smegmatis, in which it is dispensable for planktonic growth, Lsr2 is considered essential in M. tuberculosis (Chen et al., 2006; Colangeli et al., 2007; Arora et al., 2008; Kocíncová et al., 2008; Yang et al., 2017). In addition, it has also been shown to contribute to antibiotic resistance (Colangeli et al., 2007) and to control the expression of a large panel of genes acquired by horizontal gene transfer in M. tuberculosis (Gordon et al., 2010). Like H-NS, Lsr2 preferentially binds to AT-rich sequences and can oligomerize through its protein-protein interaction domain to form filaments along the M. tuberculosis chromosome (Gordon et al., 2010, 2011). Moreover, it has the ability to bridge distant DNA segments, suggesting a role in the organization and compaction of the nucleoid (Chen et al., 2008).
Herein, we generated lsr2 knock-out mutants in both S and R variants of M. abscessus to define the contribution of Lsr2 in major physiological processes that are relevant to the context of increased pathogenicity of the R variant.
Materials and Methods
Strains and Culture Media
Mycobacterium abscessus subsp. abscessus S and R 19977-IP strains were used in this study and designated Mabs-S and Mabs-R, respectively. Mycobacterial strains were grown aerobically at 37°C in Middlebrook 7H9 broth or on Middlebrook 7H11 agar, supplemented with 0.2% glycerol and 1% glucose. When necessary; kanamycin, hygromycin, and zeocin were added to medium at 250 μg/ml, 500 μg/ml (or 1000 μg/ml for selection) and 25 μg/ml, respectively. After infection experiments, bacterial CFU were counted on agar plates (BioMérieux, France), either Columbia blood agar plates after infections in macrophages and amoeba, and H2O2 exposition, either VCAT (Vancomycin, Colistin sulfate, Amphotericin B, and Trimethoprim) chocolate agar plates after mice infections. The Escherichia coli TOP10 strain (Thermo Fisher) was grown in Lysogeny Broth (LB) medium with or without kanamycin (25 μg/ml).
Construction of lsr2 Mutants in M. abscessus
The lsr2 downstream and upstream regions were amplified by PCR using the primer pairs MC75/MC76 and MC80/MC81, respectively, then cloned with the zeocin resistance cassette yielding pMC34. Replacement of the endogenous lsr2 gene by the zeocin resistance cassette was performed as described previously (van Kessel and Hatfull, 2007; Medjahed and Reyrat, 2009; Bakala N’Goma et al., 2015). lsr2 knock-out mutants, designated Mabs-S-Δlsr2 and Mabs-R-Δlsr2 were confirmed by PCR (primer pairs lsr2-5/lsr2V and MC75/ZeoR) (Supplementary Figure S1) and Southern blotting (Figure 2B). Complementation was achieved by PCR amplification of lsr2 under the control of its endogenous promoter region (961 bp) using the primers Comp-MAB_0545+reg-AclI/Comp-MAB_0545-HpaI and insertion into the integrative vector pMV361. The resulting construct was then introduced into the Mabs-S-Δlsr2 strain. Mabs-R-Δlsr2 complementation was achieved by PCR amplification (primer pair Comp-MAB_0545-NdeI/ Comp-MAB_0545-HindIII) and cloning of lsr2 under the control of the hsp60 promoter in the replicative plasmid pVV16 (Grzegorzewicz et al., 2012) Expression of lsr2 mRNA transcripts was checked by quantitative RT-PCR analysis. To generate the Mabs-S-Δlsr2-Clsr2-FLAG strain used for the ChIP-qPCR experiments, lsr2 was fused to the FLAG tag by PCR amplification using the primer pair Comp-MAB_0545-reg-AclI/ Comp-MAB_0545-FLAG-HpaI and cloning with its endogenous promoter into the integrative plasmid pMVH361. All the primers used in this study are listed in Supplementary Table S1.
Quantitative RT-PCR Analysis
mRNA was reverse transcribed using the “iScript reverse transcription” kit (Bio-Rad). Quantitative PCR was performed with a MasterMix qPCR (Eurogentec) in a Chromo4 instrument (Bio-Rad), as described previously (Pawlik et al., 2013; Le Moigne et al., 2016).
ChIP-qPCR Analysis
For each ChIP library, 50 ml of Mabs-S-Δlsr2-Clsr2-FLAG was grown until an OD600 of 0.5 and fixed in 1% formaldehyde (Euromedex) and lysed using a Precellys grinder (3 cycles: 6,700 rpm −3 × 20 s ON/60 s OFF, Bertin Technologies) and VK05 beads. The bacterial chromatin was sheared in 100–300 bp fragments using a Bioruptor Pico (Diagenode). Chromatin Immunoprecipitation was performed as described previously (Grainger et al., 2006) using IgG M2 anti-FLAG (Sigma, F1804) and Anti-GFP (G6539, Sigma) mouse monoclonal antibodies attached to proteins A/G coupled to magnetic beads (Thermo fisher). The immunoprecipitated DNA and 1% of the total input were reverse-crosslinked and eluted using the iPure v2 kit (Diagenode). The quantitative PCR tests were performed using the SsoFast evergreen supermix (Bio-Rad). Enrichment of Lsr2 binding at the GPL operon operator was calculated using the “Percent Input” method using the primers MAB_4100c_qPCR1 and MAB_4100c_qPCR3 (Supplementary Table S1).
GPL Analysis
Apolar and polar lipid fractions were obtained from exponentially growing mycobacteria cultured as reported previously (Besra, 1998). The polar lipid fraction containing GPL was separated by TLC using silica gel 60 coated aluminum TLC plates (Merck) and chloroform/methanol (90:10; v/v) as solvent system, as previously described (Sondén et al., 2005; Pawlik et al., 2013). GPL were revealed by treating TLC plates with a mist of 0.2% (w/v) of Anthrone diluted in sulfuric acid, followed by charring.
Susceptibility Profile to H2O2
Exponential growth phase cultures (OD600 between 0.6 and 0.8) of M. abscessus were diluted to obtain an OD600 of 0.1 and transferred in two new tubes. One of the two cultures was exposed to 20 mM H2O2 and the other one to an equivalent volume of sterile water. CFUs were determined by plating aliquots at various time points after the addition of H2O2 (2, 4, 6, and 8 h) on blood agar plates. For each morphotype and mutant, three independent experiments were performed, and one experiment is shown.
Intracellular Survival in Macrophages and Amoeba After Infection
Murine J774.2 macrophages (MΦ) were grown at 37°C under 5% CO2 in DMEM medium supplemented with 5% heat-inactivated Fetal Calf Serum (FBS), penicillin (100 IU/ml) and streptomycin (100 μg/ml). MΦ infections with the M. abscessus strains were carried out, as described previously (Bakala N’Goma et al., 2015; Roux et al., 2016). CFU were counted after 3 to 5 days of incubation at 37°C. Amoeba infections were done using Acanthamoeba castellanii (ATCC30010), as described earlier (Bakala N’Goma et al., 2015; Dubois et al., 2018a). At 1, 2, and 3 days of co-culture, the A. castellanii monolayer was disrupted with 0.1% SDS for 30 min at 32°C and CFU were counted.
Systemic Infection in Mice
Six to eight-week-old BALB/c mice were infected with M. abscessus strains, as described earlier (Catherinot et al., 2007; Rottman et al., 2007; Bakala N’Goma et al., 2015; Bernut et al., 2015). A Student t-test was carried out to test significance of differences between groups. All procedures were performed according to the institutional and national ethical guidelines and approved by the Comité d’éthique en experimentation animale N°047 with agreement A783223 under the reference APAFIS#11465.
Zebrafish Infections
All zebrafish experiments were done according to European Union guidelines for handling of laboratory animals1 and approved by the Comité d’Ethique pour l’Expérimentation Animale de la région Languedoc Roussillon under the reference CEEALR36-1145. Experiments were performed using the golden mutant (Lamason et al., 2005). Zebrafish embryos were obtained, maintained and microinjected in the caudal vein of 30 hpf dechorionated embryos as described (Bernut et al., 2014a, 2015). Survival curves and statistics were carried out as described (Bernut et al., 2014a).
Results
Transcription Levels of lsr2 in M. abscessus R and S Strains
Our initial comparative genomic analyses performed on three isogenic M. abscessus S/R pairs identified several mutations in the GPL biosynthesis/transport locus that are responsible for the S-to-R transition (Pawlik et al., 2013). Microarray data obtained from RNAseq highlighted the differential expression pattern of several genes, including MAB_0545 (Pawlik et al., 2013; Supplementary Table S2). MAB_0545, encoding the Lsr2 protein, was found to be moderately more expressed in the R than in the S variants of the CIP104536 and ATCC19977 strains as well as in one clinical isolate, referred to as the CF R strain (Catherinot et al., 2009). To confirm these results, quantitative RT-PCR analysis was performed on lsr2 using mRNA isolated from both the S and R variants of all three strains.Figure 1 shows that transcription of lsr2 is increased in the R strains relative to their S counterparts, particularly in the two type strains, thus validating the microarray data. Because Lsr2 appears to be involved in the virulence and the antibiotic resistance of M. tuberculosis (Colangeli et al., 2009; Gordon et al., 2010), we reasoned that induction of Lsr2 expression in the R strain may explain, at least in part, the increase of virulence in the R over the S variant.
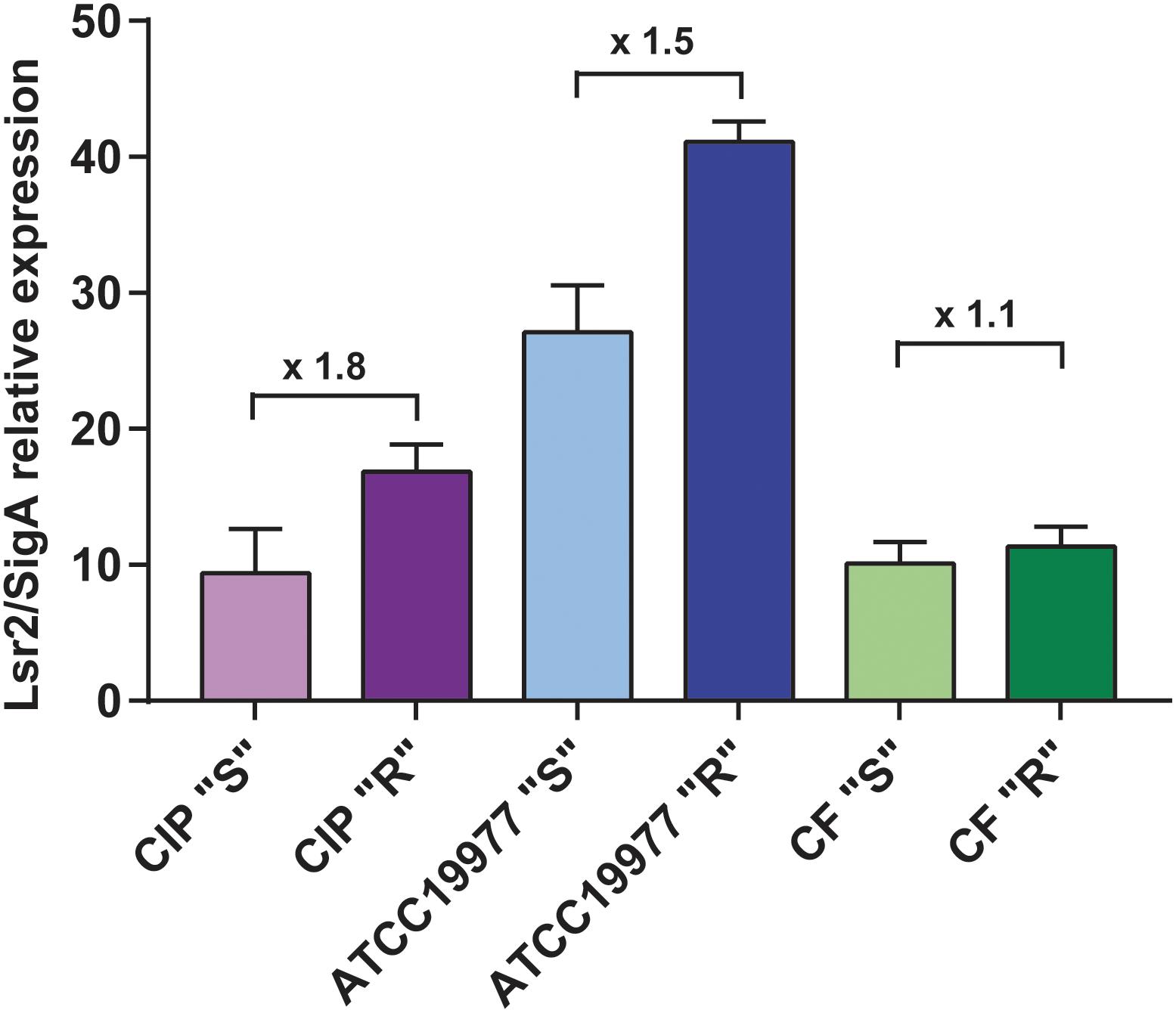
Figure 1. Increased transcriptional expression of lsr2 in M. abscessus R variants. qRT-PCR results of lsr2 mRNA expression in the R and S variants of M. abscessus CIP104536 (Collection Institut Pasteur), ATCC19977 type strains and in the CF clinical isolate. Results are expressed as lsr2/sigA ratio as described in Pawlik et al. (2013). For each assay, n = 3 and error bars are SEM.
Lsr2 Null Mutants Are Not Affected in Their Glycopeptidolipid Profile
To address the impact of Lsr2 on the M. abscessus morphotype, null mutants of lsr2 were generated in both variants, designated Mabs-S-Δlsr2 and Mabs-R-Δlsr2, using the recombineering method (van Kessel and Hatfull, 2007) that was adapted to M. abscessus (Bakala N’Goma et al., 2015; Halloum et al., 2016; Dubois et al., 2018b; Laencina et al., 2018). This was achieved by replacing lsr2 with a zeocin resistance cassette using homologous recombination (Figure 2A). Proper allelic exchange was subsequently confirmed by PCR (Supplementary Figure S1) and Southern blotting (Figure 2B). qRT-PCR revealed that complementation with a functional lsr2 gene was partial (Supplementary Figure S2).
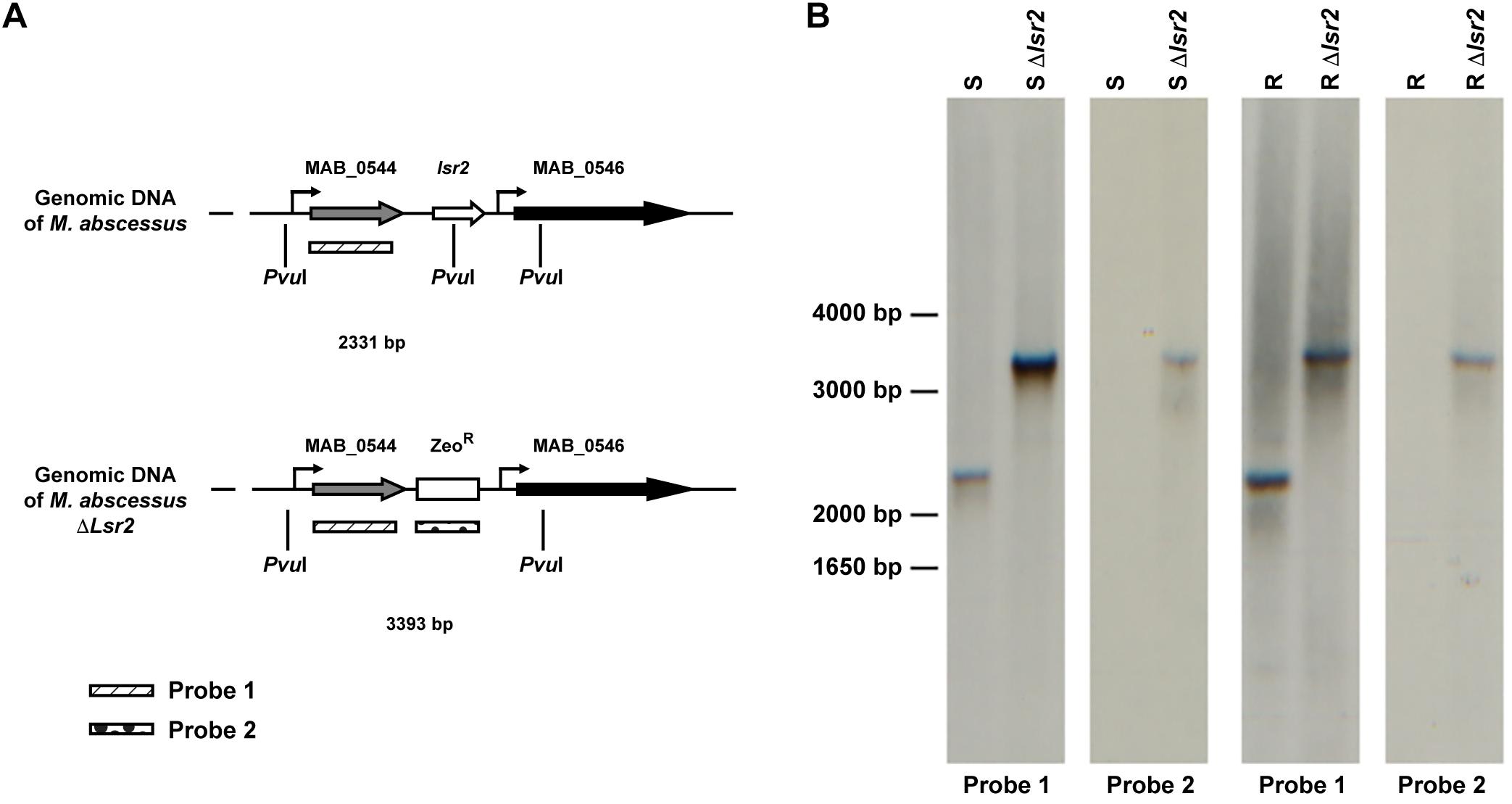
Figure 2. Generation of lsr2 null mutants. (A) Schematic representation of the genomic regions surrounding the lsr2 gene in the WT and lsr2 deletion mutant. (B) Southern blot of Mabs-S (S), Mabs-R (R), Mabs-S-Δlsr2 (S Δlsr2) and Mabs-R-Δlsr2 (R Δlsr2) using Probe 1 and Probe 2 depicted in panel (A). Genomic DNA was digested with the restriction enzyme PvuI, migrated on an agarose gel, transferred to a nitrocellulose membrane and hybridized with the two probes. Bands of the corresponding DNA fragments observed are of the expected size.
The viability of the mutant indicates that lsr2 is dispensable in M. abscessus, as previously reported in M. smegmatis (Chen et al., 2006; Arora et al., 2008; Cortes et al., 2011). However, Mabs-S-Δlsr2 and Mabs-R-Δlsr2 colonies were heterogenous in size with a tendency toward small colonies when compared to their wild-type progenitors (Supplementary Figure S3A). All strains exhibited a similar growth rate, suggesting that the reduced colony size did not affect in vitro growth in this liquid medium (Supplementary Figures S3B,C). Lsr2 has been reported to be a negative regulator of GPL expression through binding to an AT-rich element upstream of the coding region of mbtH and mps1, both known to be involved in GPL production in M. smegmatis ATCC607 (Kocíncová et al., 2008). However, two other independent studies reported an unaltered GPL profile in lsr2 mutants generated from the M. smegmatis mc2155 strain (Chen et al., 2006; Arora et al., 2008), which is the rough counterpart of the ATCC607 strain. We therefore addressed whether the inactivation of lsr2 may affect the GPL profile in both S and R M. abscessus variants. First, chromatin immunoprecipitation was carried out on lysates of Mabs-S-Δlsr2 in which a FLAG-tagged Lsr2 was introduced and Lsr2 enrichment in the AT-rich element within the promoter region of mbtH/mps1 was then measured (Figure 3A). Binding of Lsr2 was found to be significantly enriched in this region (Figure 3B). However, GPL analysis by thin layer chromatography failed to show differences in the GPL profiles in Mabs-S-Δlsr2 and in its parental S strain (Figure 3C), suggesting that Lsr2 inactivation alone is not sufficient to alter the morphotype of the mutant. As expected, Mabs-R-Δlsr2 also failed to produce GPL, which can be explained by the presence of severe genetic lesions that impair the transcription of genes such as mps1 and mmpL4b (Pawlik et al., 2013).
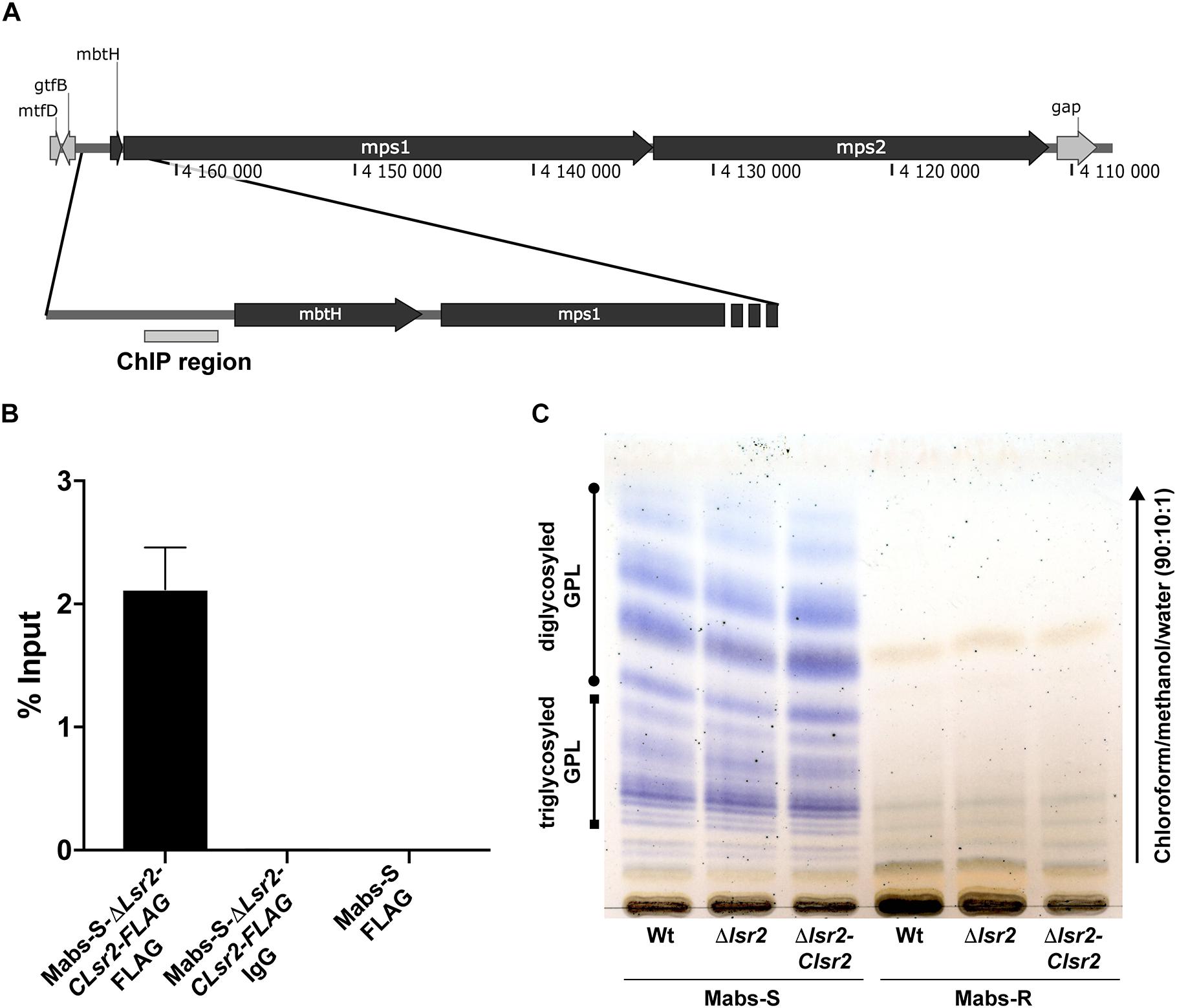
Figure 3. Measurement of Lsr2 enrichment in the promoter region upstream of the mbtH/mps1 genes. (A) Schematic representation of the GPL locus. The gray box upstream of the mbtH gene represents the region used to assay Lsr2 enrichment. (B) qPCR of ChIP performed using an anti-FLAG antibody. Enrichment of Lsr2 bound to the mbtH/mps1 promoter region in Mabs-S-Δlsr2-Clsr2-FLAG was calculated using the “percent input” method (% input = 1.894%). Negative controls included in ChIP assays using an unrelated IgG antibody on Mabs-S-Δlsr2-Clsr2-FLAG (% input = 0.00274%) and the anti-FLAG antibody on Mabs-S (% input = 0.00049%) confirm the specificity of the results. For each assay, n = 3 and error bars are SEM. (C) GPL profile characterizing the WT and Δlsr2 and lsr2 complemented strains in both S and R backgrounds. Following extraction, the GPL were separated by thin layer chromatography using chloroform/methanol (90:10, v/v) and revealed with a treatment of the TLC plate with a mist of 0.2% of Anthrone in concentrated sulfuric acid and charring.
Collectively, these data showed Lsr2 ability to bind the promoter region upstream of mbtH/msp1. However, loss of Lsr2 expression has no effect on production or composition of GPL in the S M. abscessus variant.
Lsr2 Promotes Resistance to Oxidative Species in the R Variant
An important function of Lsr2 resides in protecting the integrity of genomic DNA from the deleterious action of reactive oxygen species (ROS) during macrophage infection (Colangeli et al., 2009). We aimed at addressing the potential protective effect of the higher expression of lsr2 in M. abscessus R cultures under H2O2 exposure. Exponential phase cultures, diluted 10 times, were exposed to 20 mM H2O2 and CFU were counted at various time points during treatment. When comparing WT M. abscessus S and R variants, a high proportion of the R population survived in the presence of H2O2 (p < 0.001) (Figure 4A). Whether this advantage is conferred by a higher lsr2 expression was next investigated. The growth defects of Mabs-S and Mabs-S-Δlsr2 in the presence of H2O2 were similar (Supplementary Figure S4). In contrast, Mabs-R-Δlsr2 was more sensitive than its WT R progenitor, while genetic complementation in the R lsr2 mutant restored the WT R H2O2 resistance phenotype (Figure 4B). These results show a difference in the response of the M. abscessus S/R variants to H2O2, the R variant being less sensitive to oxidative stress than the S variant. However, the intrinsic resistance of the R variant was partially lost upon deletion of the lsr2 gene.
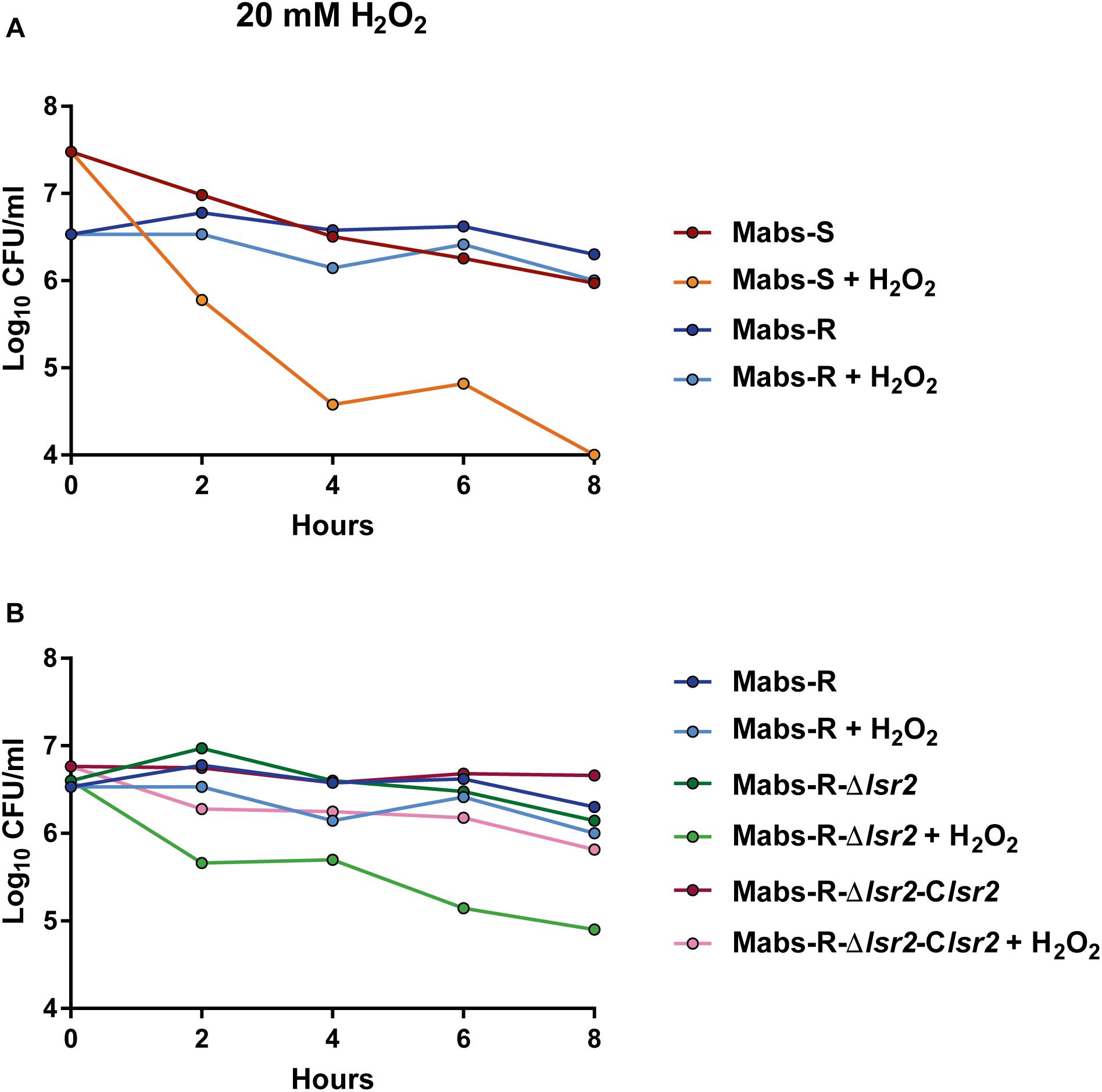
Figure 4. Susceptibility profile of M. abscessus toward oxidative derivatives. Strains were grown in broth medium until exponential phase and exposed to 20 mM H2O2. Growth was monitored by CFU counting at 2, 4, 6, and 8 h. (A) Comparative response of M. abscessus S and R variants to the treatment. (B) Comparative response of Mabs-R-Δlsr2, Mabs-R-Δlsr2 and Mabs-R Δlsr2-Clsr2.
The Intracellular Survival of lsr2 Mutants Is Reduced in Amoebae and Macrophages
The role of Lsr2 in the resistance of M. abscessus to oxidative species suggests that it may also protect the bacilli from the microbicidal activity of macrophages (Roux et al., 2016) and/or amoebae (Bakala N’Goma et al., 2015). This prompted us to investigate and compare the impact of lsr2 deletion on intracellular growth of the S and R mutants with their corresponding complemented strains (Mabs-S-Δlsr2-Clsr2 and Mabs-R-Δlsr2-Clsr2) and wild-type progenitors. Mabs-S-Δlsr2 (Figure 5A) and Mabs-R-Δlsr2 (Figure 5B) exhibited a pronounced defect inside murine MΦ as compared to the WT and complemented strains. This survival defect was already significant at 1 dpi for the S strain (p < 0.0001) and at 5 dpi for the R strain (p < 0.0001). Inside A. castellani, a similar decrease in bacterial viability was observed for both Mabs-S-Δlsr2 and Mabs-R-Δlsr2 mutants (Figures 5C,D). At 3 dpi, both mutants showed a strong difference (p < 0.0001) in intra-amoebal multiplication compared to their respective WT and complemented strains.
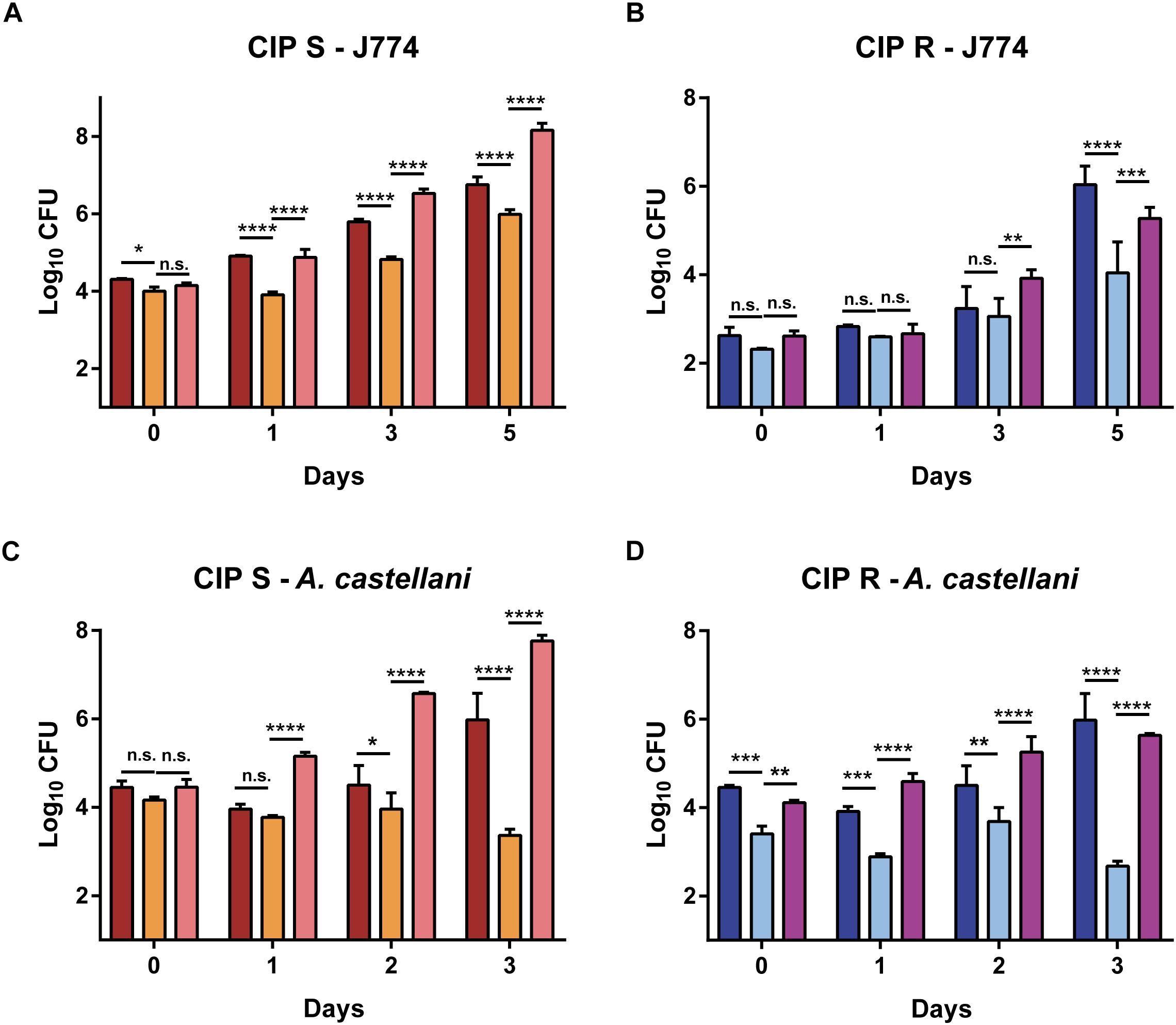
Figure 5. Intracellular growth of Δlsr2 mutants in macrophages (A,B) and in amoebae (C,D). Murine J774.2 macrophages were infected with CIP104536 (Collection Institut Pasteur) Mabs-S (in red) and Mabs-R (dark blue), Mabs-S-Δlsr2 (orange) and Mabs-R-Δlsr2 (light blue) and their respective complemented strains (C-Δlsr2; pink for S and mauve for R) at a MOI 1 whereas A. castellanii cells were infected with the same strains at a MOI 10. Intracellular growth was assessed by CFU counting for 5 days in macrophages and 3 days in amoebae. CFU histograms with error bars represent means ± SD calculated using data from two independent experiments. Differences between means were analyzed by two-way ANOVA and the Tukey post-test, allowing multiple comparisons. ns, non-significant, ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001, and ∗∗∗∗P < 0.0001.
Together, these results indicate that Lsr2 is required for intracellular survival of M. abscessus S/R forms in both macrophages and amoebae.
Lsr2 Plays a Key Role in M. abscessus Virulence in vivo
The impact of Lsr2 on mycobacterial virulence was originally suggested by demonstrating the preferential binding of Lsr2 to AT-rich regions in the M. tuberculosis genome, including several genomic islands acquired by horizontal gene transfer, encoding major virulence factors (Gordon et al., 2010). The behavior of the lsr2 mutants was first addressed by infecting zebrafish larvae (Bernut et al., 2014a) with the wild-type R (Mabs-R) and Mabs-R-Δlsr2 strains. A significant reduction in the virulence of Mabs-R-Δlsr2 was observed when compared with Mabs-R (Figure 6A). Whereas only 10% of zebrafish were killed when infected with Mabs-R-Δlsr2 at 12 dpi, around 40% of the larvae died at 6 dpi with Mabs-R while nearly 70% died at 10 dpi (Figure 6A). Complementing with lsr2 partially reversed the attenuated phenotype of Mabs-R-Δlsr2.
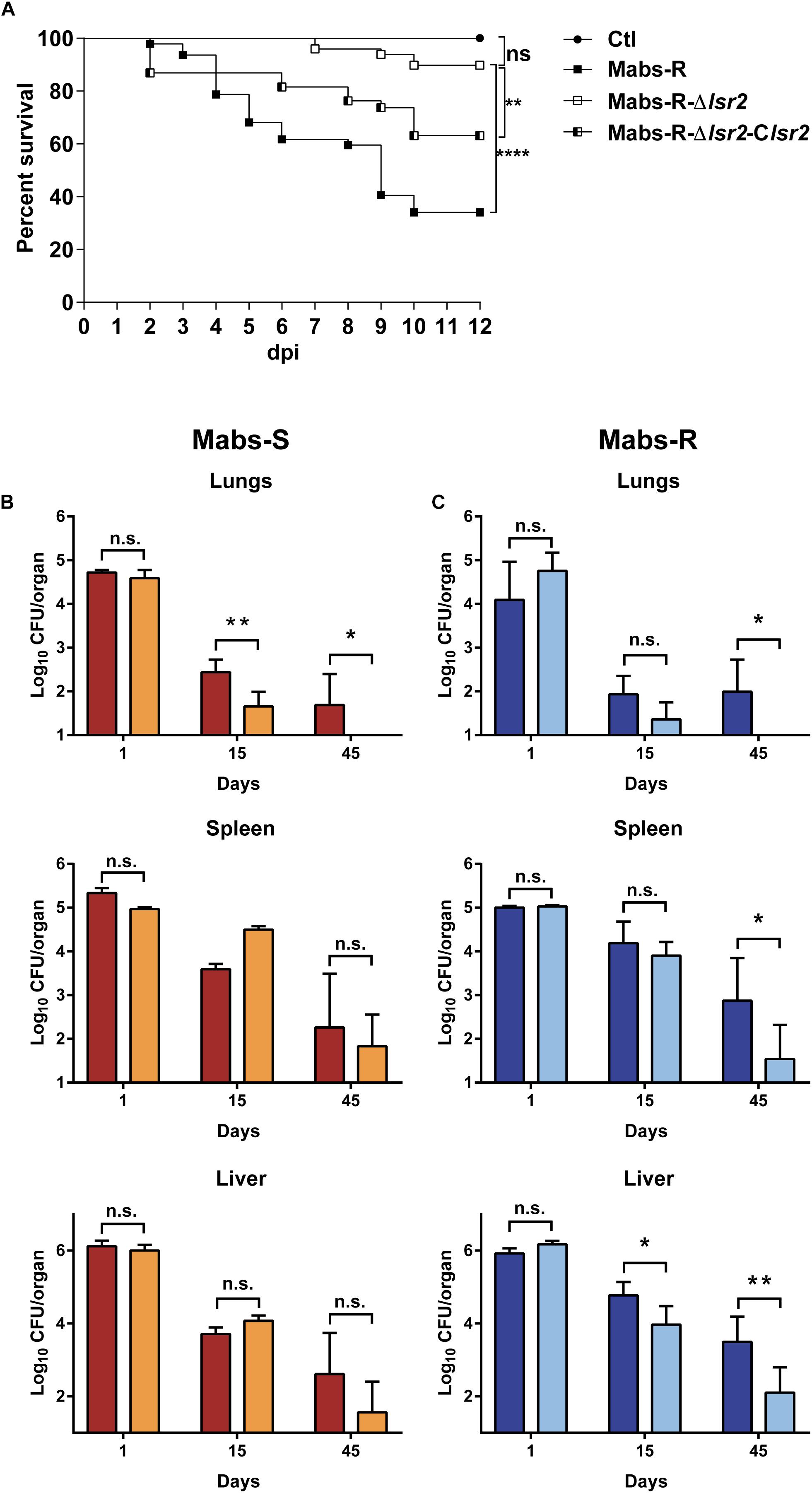
Figure 6. Lsr2 is required for survival of M. abscessus in zebrafish and mice. (A) Survival curve of infected embryos with 50–164 CFU of Mabs-R (n = 47 embryos), 160–282 CFU of Mabs-R-Δlsr2 (n = 49 embryos) and 149–290 CFU of the complemented strain (n = 37 embryos). Experiments were done twice. Significant differences were obtained using the Log-rank (Mantel-Cox) statistical test; ∗∗P < 0.01 and ∗∗∗∗P < 0.0001. (B,C) Comparative analysis of bacterial burden in different organs after autopsy of BALB/c mice infected intravenously with 106 CFU of Mabs-S (in red) and Mabs-S-Δlsr2 (orange) (B) or Mabs-R (dark blue) and Mabs-R-Δlsr2 (light blue) (C) in the lungs, spleen and liver at 1, 15, and 45 dpi. Student’s t-test and Fisher’s exact test were used. A P-value of 0.05 was considered significant (∗P < 0.05 and ∗∗P < 0.01).
To further evaluate the contribution of Lsr2 in pathogenesis of M. abscessus in a more complex animal host, BALB/c mice were intravenously infected with the wild-type S/R variants or Mabs-S/R-Δlsr2 strains (Catherinot et al., 2007; Rottman et al., 2007). Bacterial loads were monitored by measuring the CFU counts in the lung, spleen and liver homogenates from mice sacrificed at 1, 15, and 45 dpi. A significant reduction in the number of Mabs-S-Δlsr2 bacilli was observed in the lungs at 15 dpi when compared to the wild-type S progenitor (p < 0.01). This difference was further increased at 45 dpi, along with a complete clearance of Mabs-S-Δlsr2 (p < 0.05) (Figure 6B, upper panels). However, no significant differences in CFU counts in the spleen and the liver were observed between Mabs-S and Mabs-S-Δlsr2. Monitoring the bacterial burden of Mabs-R-Δlsr2 revealed a significant reduction in all three organs at 45 dpi as compared to the WT parental strain (p < 0.05 in lungs and spleen; p < 0.01 in liver) (Figure 6C, lower panels).
Overall, these results indicate that Lsr2 plays a critical role in M. abscessus virulence in different animal hosts and highlight the requirement of Lsr2 for persistence in mice, particularly in the lungs, which represents the major targeted organ during infection in patients, especially in those with already underlying pulmonary diseases.
Discussion
Lsr2, a unique nucleoid associated protein in the Actinomyces and Mycobacterium genera, has been extensively studied in M. tuberculosis (Colangeli et al., 2009; Gordon et al., 2010; Bartek et al., 2014). Like its ortholog H-NS, homodimers of Lsr2 can bind DNA cooperatively to form long oligomers on AT-rich sequences that can further interact to bridge distant DNA regions and contribute to loop formation (Chen et al., 2008; Liu and Gordon, 2012). At the genomic level, Lsr2 filaments are found at the promoters of 401 and 272 genes in M. tuberculosis and M. smegmatis, respectively, often extending into their coding regions (Gordon et al., 2010). This observation, as well as its known role as a negative regulator in several cellular functions, strongly suggest that like H-NS, Lsr2 represses transcription through promoter occlusion of RNA polymerase targets or by interfering with transcription elongation (Chen et al., 2008; Liu and Gordon, 2012; Landick et al., 2015). Lsr2 has been shown to be involved in M. tuberculosis virulence by binding to important genes, such as genes involved in the ESX secretion systems, the biosynthesis of the PDIM and PGL cell wall lipids or encoding antigenic proteins of the PE/PPE family (Gordon et al., 2010). Previous studies also demonstrated the implication of Lsr2 in multi-drug tolerance of M. tuberculosis (Colangeli et al., 2007) and in protection against reactive oxygen intermediates (Colangeli et al., 2009). All these phenotypes were investigated and confirmed in M. abscessus, except for the increased resistance to antibiotics (not shown), thus involving Lsr2 in increased resistance to oxidative species as well as in intracellular multiplication and survival of M. abscessus in zebrafish and mice.
The S-to-R transition of M. abscessus has only been found to occur during infection either in mice (Rottman et al., 2007) or in humans (Jönsson et al., 2007; Catherinot et al., 2009; Roux et al., 2009). In addition, the underlying genetic causes for this transition pointed to irreversible mechanisms, due to selective mutations such as indels that prevent the R form to reverse its morphotype to a S form (Pawlik et al., 2013). Thus, the S-to-R transition represents a clear advantage for M. abscessus during the course of the infection. When comparing pairs of isogenic S/R variants, it was found that lsr2 was expressed at a higher level in the R morphotype (Pawlik et al., 2013). This was confirmed in the present study using qRT-PCR analysis. By inducing a higher expression of lsr2, the R variant becomes more resistant to reactive oxygen species. Interestingly, these conditions look similar to those prevailing in the CF airways, characterized by polymicrobial and chronic infections, which maintain a very harsh and oxidative environment in the lungs. Similarly, CF patients receive frequent cures involving large-spectrum antibiotics, such as inhaled tobramycin therapy. Herein, we show that the R form is more resistant to H202 than the S form. But we were unable to confirm an increased resistance to antibiotics (not shown). This resistant phenotype was lost in the lsr2 mutant but restored to wild-type levels following complementation with a functional copy of lsr2.
A major outcome of this work is to point out the role of Lsr2 in the intracellular behavior of M. abscessus and on its requirement for persistence in infected mice and zebrafish. It was previously shown that a deletion of lsr2 in M. smegmatis has an impact on its survival in murine macrophages (Colangeli et al., 2009). In vivo studies of the role of lsr2 in virulence in mice were also performed on M. tuberculosis, using an attenuated strain that was complemented by a replicative plasmid expressing Lsr2 under the control of an inducible promoter (Ehrt et al., 2005) to circumvent the absence of viability of the lsr2 M. tuberculosis deletion mutant (Colangeli et al., 2007). Using this strategy, it was shown that lsr2 is important to protect M. tuberculosis against ROI during macrophage infection. The present study shows that despite being dispensable in M. abscessus, lsr2 remains essential for optimal intracellular growth and virulence in different vertebrates.
Previous work in M. smegmatis emphasized the contribution of Lsr2 in defining the lipid profile, colony morphology, bacterial motility and biofilm formation by specifically repressing transcription at the GPL locus in the rough morphotypes (Chen et al., 2006; Arora et al., 2008; Kocíncová et al., 2008; Yang et al., 2017). However, this study clearly shows that lsr2 mRNAs are detected in both M. abscessus S and R strains. Thanks to ChIP-qPCR, we also observed that Lsr2 is able to bind the promoter region of mbtH in M. abscessus S, despite the equivalent GPL profiles in the WT and the mutant strain. This suggests that binding of Lsr2 to this particular genomic region is not sufficient to regulate the biosynthetic GPL locus and that this process may involve other partners that remain to be identified. An alternative explanation is that the higher expression level of Lsr2 in M. abscessus R is needed to successfully reduce GPL production, eventually by increasing the formation of a DNA loop that can occlude RNA polymerase binding or inhibit transcription elongation, as proposed for H-NS in Gram-negative bacteria (Landick et al., 2015). One would have anticipated that increasing expression of Lsr2 in M. abscessus R would enhance the R phenotype by further repressing the genes of the GPL locus. However, deletion of lsr2 in M. abscessus R did not restore GPL production as it was previously observed in M. smegmatis (Kocíncová et al., 2008). As mentioned above, the GPL locus in the M. abscessus R variant suffers from irreversible genetic lesions, adding a layer of complexity to the molecular mechanisms involved in the regulation of this locus. The pleiotropic nature of Lsr2 implies that its deletion has a broader impact on the fitness of M. abscessus as well as on the general compaction of its nucleoid. Thus, additional analysis using genome-wide methods will help to unravel the details of the molecular role of Lsr2 during S-to-R transition and its impact on pathogenicity.
Taken together, this work showed the impact of Lsr2 as a key virulence factor on the intracellular and in vivo survival of M. abscessus. These results also strongly suggest that Lsr2, like other NAPs, might represent a target of choice for the development of new antimicrobials. Indeed, compounds that would either silence expression of lsr2 or inhibit its function could therefore be considered as a new anti-virulence approach against M. abscessus (Liu and Gordon, 2012).
Ethics Statement
All procedures involving mice were performed according to the institutional and national ethical guidelines and approved by the comité d’éthique en experimentation animale N°047 with agreement A783223 under the reference APAFIS#11465. All zebrafish experiments were done according to European Union guidelines for handling of laboratory animals (http://ec.europa.eu/environment/chemicals/lab_animals/home_en.htm) and approved by the Comité d’Ethique pour l’Expérimentation Animale de la région Languedoc Roussillon under the reference CEEALR36-1145.
Author Contributions
J-LH, LK, and FC designed the project and experiments. VLM, AB, MC, AV, CD, and AP performed the experiments. J-LG, FM, FC, LK, and J-LH wrote and corrected the manuscript.
Funding
This work was supported by the Vaincre la Mucoviscidose association (Post-doctoral fellowship to VLM #RF20110600446/1/3/130) and the ANR DIMYVIR Grant (ANR-13-BSV3-0007-01 to J-LH and LK) from the French National Research Agency Program.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.00905/full#supplementary-material
Footnotes
References
Arora, K., Whiteford, D. C., Lau-Bonilla, D., Davitt, C. M., and Dahl, J. L. (2008). Inactivation of lsr2 results in a hypermotile phenotype in Mycobacterium smegmatis. J. Bacteriol. 190, 4291–4300. doi: 10.1128/JB.00023-08
Bakala N’Goma, J. C., Le Moigne, V., Soismier, N., Laencina, L., Le Chevalier, F., et al. (2015). Mycobacterium abscessus phospholipase C expression is induced during coculture within amoebae and enhances M. abscessus virulence in mice. Infect. Immun. 83, 780–791. doi: 10.1128/IAI.02032-14
Bartek, I. L., Woolhiser, L. K., Baughn, A. D., Basaraba, R. J., Jacobs, W. R. Jr., Lenaerts, A. J., et al. (2014). Mycobacterium tuberculosis Lsr2 is a global transcriptional regulator required for adaptation to changing oxygen levels and virulence. mBio 5:e01106-14. doi: 10.1128/mBio.01106-14
Bernut, A., Dupont, C., Sahuquet, A., Herrmann, J.-L., Lutfalla, G., and Kremer, L. (2015). Deciphering and Imaging Pathogenesis and Cording of Mycobacterium abscessus in zebrafish embryos. J. Vis. Exp. 103:53130. doi: 10.3791/53130
Bernut, A., Herrmann, J.-L., Kissa, K., Dubremetz, J.-F., Gaillard, J.-L., Lutfalla, G., et al. (2014a). Mycobacterium abscessus cording prevents phagocytosis and promotes abscess formation. Proc. Natl. Acad. Sci. U.S.A. 111, E943–E952. doi: 10.1073/pnas.1321390111
Bernut, A., Le Moigne, V., Lesne, T., Lutfalla, G., Herrmann, J.-L., and Kremer, L. (2014b). In vivo assessment of drug efficacy against Mycobacterium abscessus using the embryonic zebrafish test system. Antimicrob. Agents Chemother. 58, 4054–4063. doi: 10.1128/AAC.00142-14
Bernut, A., Herrmann, J.-L., Ordway, D., and Kremer, L. (2017). The diverse cellular and animal models to decipher the physiopathological traits of Mycobacterium abscessus infection. Front. Cell. Infect. Microbiol. 7:100. doi: 10.3389/fcimb.2017.00100
Besra, G. S. (1998). Preparation of cell-wall fractions from mycobacteria. Methods Mol. Biol. 101, 91–107.
Brown-Elliott, B. A., and Wallace, R. J. (2002). Clinical and taxonomic status of pathogenic nonpigmented or late-pigmenting rapidly growing mycobacteria. Clin. Microbiol. Rev. 15, 716–746.
Byrd, T. F., and Lyons, C. R. (1999). Preliminary characterization of a Mycobacterium abscessus mutant in human and murine models of infection. Infect. Immun. 67, 4700–4707.
Catherinot, E., Clarissou, J., Etienne, G., Ripoll, F., Emile, J.-F., Daffé, M., et al. (2007). Hypervirulence of a rough variant of the Mycobacterium abscessus type strain. Infect. Immun. 75, 1055–1058.
Catherinot, E., Roux, A.-L., Macheras, E., Hubert, D., Matmar, M., Dannhoffer, L., et al. (2009). Acute respiratory failure involving an R variant of Mycobacterium abscessus. J. Clin. Microbiol. 47, 271–274. doi: 10.1128/JCM.01478-08
Chen, J. M., German, G. J., Alexander, D. C., Ren, H., Tan, T., and Liu, J. (2006). Roles of Lsr2 in colony morphology and biofilm formation of Mycobacterium smegmatis. J. Bacteriol. 188, 633–641.
Chen, J. M., Ren, H., Shaw, J. E., Wang, Y. J., Li, M., Leung, A. S., et al. (2008). Lsr2 of Mycobacterium tuberculosis is a DNA-bridging protein. Nucleic Acids Res. 36, 2123–2135. doi: 10.1093/nar/gkm1162
Colangeli, R., Haq, A., Arcus, V. L., Summers, E., Magliozzo, R. S., McBride, A., et al. (2009). The multifunctional histone-like protein Lsr2 protects mycobacteria against reactive oxygen intermediates. Proc. Natl. Acad. Sci. U.S.A. 106, 4414–4418. doi: 10.1073/pnas.0810126106
Colangeli, R., Helb, D., Vilchèze, C., Hazbon, M. H., Lee, C.-G., Safi, H., et al. (2007). Transcriptional regulation of multi-drug tolerance and antibiotic-induced responses by the histone-like protein Lsr2 in M. tuberculosis. PLoS Pathog. 3:e87. doi: 10.1371/journal.ppat.0030087
Cortes, M., Singh, A. K., Reyrat, J. M., Gaillard, J.-L., Nassif, X., and Herrmann, J.-L. (2011). Conditional gene expression in Mycobacterium abscessus. PLoS One 6:e29306. doi: 10.1371/journal.pone.0029306
Diel, R., Ringshausen, F., Richter, E., Welker, L., Schmitz, J., and Nienhaus, A. (2017). Microbiological and clinical outcomes of treating non-Mycobacterium avium complex nontuberculous mycobacterial pulmonary disease: a systematic review and meta-analysis. Chest 152, 120–142. doi: 10.1016/j.chest.2017.04.166
Dubois, V., Laencina, L., Bories, A., Le Moigne, V., Pawlik, A., Herrmann, J. L., et al. (2018a). Identification of virulence markers of Mycobacterium abscessus for intracellular replication in phagocytes. J. Vis. Exp. 139:e57766. doi: 10.3791/57766
Dubois, V., Viljoen, A., Laencina, L., Le Moigne, V., Bernut, A., Dubar, F., et al. (2018b). MmpL8MAB controls Mycobacterium abscessus virulence and production of a previously unknown glycolipid family. Proc. Natl. Acad. Sci. U.S.A. 115, E10147–E10156. doi: 10.1073/pnas.1812984115
Ehrt, S., Guo, X. V., Hickey, C. M., Ryou, M., Monteleone, M., Riley, L. W., et al. (2005). Controlling gene expression in mycobacteria with anhydrotetracycline and Tet repressor. Nucleic Acids Res. 33:e21. doi: 10.1093/nar/gni013
Esther, C. R., Esserman, D. A., Gilligan, P., Kerr, A., and Noone, P. G. (2010). Chronic Mycobacterium abscessus infection and lung function decline in cystic fibrosis. J. Cyst. Fibros. Soc. 9, 117–123. doi: 10.1016/j.jcf.2009.12.001
Gordon, B. R., Imperial, R., Wang, L., Navarre, W. W., and Liu, J. (2008). Lsr2 of Mycobacterium represents a novel class of H-NS like proteins. J. Bacteriol. 190, 7052–7059. doi: 10.1128/JB.00733-08
Gordon, B. R., Li, Y., Wang, L., Sintsova, A., van Bakel, H., Tian, S., et al. (2010). Lsr2 is a nucleoid-associated protein that targets AT-rich sequences and virulence genes in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. U.S.A. 107, 5154–5159. doi: 10.1073/pnas.0913551107
Gordon, B. R. G., Li, Y., Cote, A., Weirauch, M. T., Ding, P., Hughes, T. R., et al. (2011). Structural basis for recognition of AT-rich DNA by unrelated xenogeneic silencing proteins. Proc. Natl. Acad. Sci. U.S.A. 108, 10690–10695. doi: 10.1073/pnas.1102544108
Grainger, D. C., Hurd, D., Goldberg, M. D., and Busby, S. J. (2006). Association of nucleoid proteins with coding and non-coding segments of the Escherichia coli genome. Nucleic Acids Res. 34, 4642–4652. doi: 10.1093/nar/gkl542
Grzegorzewicz, A. E., Pham, H., Gundi, V. A. K. B., Scherman, M. S., North, E. J., Hess, T., et al. (2012). Inhibition of mycolic acid transport across the Mycobacterium tuberculosis plasma membrane. Nat. Chem. Biol. 8, 334–341. doi: 10.1038/nchembio.794
Gutiérrez, A. V., Viljoen, A., Ghigo, E., Herrmann, J.-L., and Kremer, L. (2018). Glycopeptidolipids, a double-edged sword of the Mycobacterium abscessus complex. Front. Microbiol. 9:1145. doi: 10.3389/fmicb.2018.01145
Halloum, I., Carrère-Kremer, S., Blaise, M., Viljoen, A., Bernut, A., Le Moigne, V., et al. (2016). Deletion of a dehydratase important for intracellular growth and cording renders rough Mycobacterium abscessus avirulent. Proc. Natl. Acad. Sci. U.S.A. 113, E4228–E4237. doi: 10.1073/pnas.1605477113
Howard, S. T., Rhoades, E., Recht, J., Pang, X., Alsup, A., Kolter, R., et al. (2006). Spontaneous reversion of Mycobacterium abscessus from a smooth to a rough morphotype is associated with reduced expression of glycopeptidolipid and reacquisition of an invasive phenotype. Microbiology 152, 1581–1590.
Jeong, S. H., Kim, S.-Y., Huh, H. J., Ki, C.-S., Lee, N. Y., Kang, C.-I., et al. (2017). Mycobacteriological characteristics and treatment outcomes in extrapulmonary Mycobacterium abscessus complex infections. Int. J. Infect. Dis. 60, 49–56. doi: 10.1016/j.ijid.2017.05.007
Jönsson, B. E., Gilljam, M., Lindblad, A., Ridell, M., Wold, A. E., and Welinder-Olsson, C. (2007). Molecular epidemiology of Mycobacterium abscessus, with focus on cystic fibrosis. J. Clin. Microbiol. 45, 497–504.
Kocíncová, D., Singh, A. K., Beretti, J. L., Ren, H., Euphrasie, D., Liu, J., et al. (2008). Spontaneous transposition of IS1096 or ISMsm3 leads to glycopeptidolipid overproduction and affects surface properties in Mycobacterium smegmatis. Tuberculosis 88, 390–398. doi: 10.1016/j.tube.2008.02.005
Laal, S., Sharma, Y. D., Prasad, H. K., Murtaza, A., Singh, S., Tangri, S., et al. (1991). Recombinant fusion protein identified by lepromatous sera mimics native Mycobacterium leprae in T-cell responses across the leprosy spectrum. Proc. Natl. Acad. Sci. U.S.A. 88, 1054–1058.
Laencina, L., Dubois, V., Le Moigne, V., Viljoen, A., Majlessi, L., Pritchard, J., et al. (2018). Identification of genes required for Mycobacterium abscessus growth in vivo with a prominent role of the ESX-4 locus. Proc. Natl. Acad. Sci. U.S.A. 115, E1002–E1011. doi: 10.1073/pnas.1713195115
Lamason, R. L., Mohideen, M. A., Mest, J. R., Wong, A. C., Norton, H. L., Aros, M. C., et al. (2005). SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science 310, 1782–1786.
Landick, R., Wade, J. T., and Grainger, D. C. (2015). H-NS and RNA polymerase: a love–hate relationship? Curr. Opin. Microbiol. 24, 53–59. doi: 10.1016/j.mib.2015.01.009
Le Moigne, V., Belon, C., Goulard, C., Accard, G., Bernut, A., Pitard, B., et al. (2016). MgtC as a host-induced factor and vaccine candidate against Mycobacterium abscessus infection. Infect. Immun. 84, 2895–2903. doi: 10.1128/IAI.00359-16
Liu, J., and Gordon, B. R. (2012). Targeting the global regulator Lsr2 as a novel approach for anti-tuberculosis drug development. Expert Rev. Anti. Infect. Ther. 10, 1049–1053. doi: 10.1586/eri.12.86
Medjahed, H., Gaillard, J.-L., and Reyrat, J.-M. (2010). Mycobacterium abscessus: a new player in the mycobacterial field. Trends Microbiol. 18, 17–23. doi: 10.1016/j.tim.2009.12.007
Medjahed, H., and Reyrat, J.-M. (2009). Construction of Mycobacterium abscessus defined glycopeptidolipid mutants: comparison of genetic tools. Appl. Environ. Microbiol. 75, 1331–1338. doi: 10.1128/AEM.01914-08
Mougari, F., Guglielmetti, L., Raskine, L., Sermet-Gaudelus, I., Veziris, N., and Cambau, E. (2016). Infections caused by Mycobacterium abscessus: epidemiology,diagnostic tools and treatment. Expert Rev. Anti. Infect. Ther. 14, 1139–1154.
Nessar, R., Reyrat, J.-M., Davidson, L. B., and Byrd, T. F. (2011). Deletion of the mmpL4b gene in the Mycobacterium abscessus glycopeptidolipid biosynthetic pathway results in loss of surface colonization capability, but enhanced ability to replicate in human macrophages and stimulate their innate immune response. Microbiology 157, 1187–1195. doi: 10.1099/mic.0.046557-0
Nguyen, K. T., Piastro, K., Gray, T. A., and Derbyshire, K. M. (2010). Mycobacterial biofilms facilitate horizontal DNA transfer between strains of Mycobacterium smegmatis. J. Bacteriol. 192, 5134–5142. doi: 10.1128/JB.00650-10
Oberley-Deegan, R. E., Rebits, B. W., Weaver, M. R., Tollefson, A. K., Bai, X., McGibney, M., et al. (2010). An oxidative environment promotes growth of Mycobacterium abscessus. Free Radic. Biol. Med. 49, 1666–1673. doi: 10.1016/j.freeradbiomed.2010.08.026
Pawlik, A., Garnier, G., Orgeur, M., Tong, P., Lohan, A., Le Chevalier, F., et al. (2013). Identification and characterization of the genetic changes responsible for the characteristic smooth-to-rough morphotype alterations of clinically persistent Mycobacterium abscessus. Mol. Microbiol. 90, 612–629. doi: 10.1111/mmi.12387
Qu, Y., Lim, C. J., Whang, Y. R., Liu, J., and Yan, J. (2013). Mechanism of DNA organization by Mycobacterium tuberculosis protein Lsr2. Nucleic Acids Res. 41, 5263–5272. doi: 10.1093/nar/gkt249
Qvist, T., Taylor-Robinson, D., Waldmann, E., Olesen, H. V., Hansen, C. R., Mathiesen, I. H., et al. (2016). Comparing the harmful effects of nontuberculous mycobacteria and Gram negative bacteria on lung function in patients with cystic fibrosis. J. Cyst. Fibros. 15, 380–385. doi: 10.1016/j.jcf.2015.09.007
Ripoll, F., Pasek, S., Schenowitz, C., Dossat, C., Barbe, V., Rottman, M., et al. (2009). Non-mycobacterial virulence genes in the genome of the emerging pathogen Mycobacterium abscessus. PLoS One 4:e5660. doi: 10.1371/journal.pone.0005660
Rottman, M., Soudais, C., Vogt, G., Renia, L., Emile, J.-F., Decaluwe, H., et al. (2007). Importance of T cells, gamma interferon, and tumor necrosis factor in immune control of the rapid grower Mycobacterium abscessus in C57BL/6 mice. Infect. Immun. 75, 5898–5907.
Roux, A.-L., Catherinot, E., Ripoll, F., Soismier, N., Macheras, E., Ravilly, S., et al. (2009). Multicenter study of prevalence of nontuberculous mycobacteria in patients with cystic fibrosis in France. J. Clin. Microbiol. 47, 4124–4128.
Roux, A.-L., Ray, A., Pawlik, A., Medjahed, H., Etienne, G., Rottman, M., et al. (2011). Overexpression of proinflammatory TLR-2-signalling lipoproteins in hypervirulent mycobacterial variants. Cell. Microbiol. 13, 692–704. doi: 10.1111/j.1462-5822.2010.01565.x
Roux, A.-L., Viljoen, A., Bah, A., Simeone, R., Bernut, A., Laencina, L., et al. (2016). The distinct fate of smooth and rough Mycobacterium abscessus variants inside macrophages. Open Biol. 6:160185.
Sánchez-Chardi, A., Olivares, F., Byrd, T. F., Julián, E., Brambilla, C., and Luquin, M. (2011). Demonstration of cord formation by rough Mycobacterium abscessus variants: implications for the clinical microbiology laboratory. J. Clin. Microbiol. 49, 2293–2295. doi: 10.1128/JCM.02322-10
Smibert, O., Snell, G. I., Bills, H., Westall, G. P., and Morrissey, C. O. (2016). Mycobacterium abscessus complex - a particular challenge in the setting of lung transplantation. Expert Rev. Anti. Infect. Ther. 14, 325–333.
Sondén, B., Kocíncová, D., Deshayes, C., Euphrasie, D., Rhayat, L., Laval, F., et al. (2005). Gap, a mycobacterial specific integral membrane protein, is required for glycolipid transport to the cell surface. Mol. Microbiol. 58, 426–440.
van Dorn, A. (2017). Multidrug-resistant Mycobacterium abscessus threatens patients with cystic fibrosis. Lancet Respir. Med. 5:15.
van Kessel, J. C., and Hatfull, G. F. (2007). Recombineering in Mycobacterium tuberculosis. Nat. Methods 4, 147–152.
Varghese, B., Shajan, S. E., Al Saedi, M. O., and Al-Hajoj, S. A. (2012). First case report of chronic pulmonary lung disease caused by Mycobacterium abscessus in two immunocompetent patients in Saudi Arabia. Ann. Saudi Med. 32, 312–314. doi: 10.5144/0256-4947.2012.312
Keywords: non-tuberculous mycobacteria, Mycobacterium abscessus, Lsr2, virulence, pathogenesis, zebrafish, mouse
Citation: Le Moigne V, Bernut A, Cortès M, Viljoen A, Dupont C, Pawlik A, Gaillard J-L, Misguich F, Crémazy F, Kremer L and Herrmann J-L (2019) Lsr2 Is an Important Determinant of Intracellular Growth and Virulence in Mycobacterium abscessus. Front. Microbiol. 10:905. doi: 10.3389/fmicb.2019.00905
Received: 05 January 2019; Accepted: 09 April 2019;
Published: 30 April 2019.
Edited by:
Thomas Dick, Center for Discovery and Innovation, Hackensack Meridian Health, United StatesReviewed by:
Thomas F. Byrd, The University of New Mexico, United StatesJoseph Oliver Falkinham, Virginia Tech, United States
Copyright © 2019 Le Moigne, Bernut, Cortès, Viljoen, Dupont, Pawlik, Gaillard, Misguich, Crémazy, Kremer and Herrmann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frédéric Crémazy, ZnJlZGVyaWMuY3JlbWF6eUB1dnNxLmZy Jean-Louis Herrmann, amVhbi1sb3Vpcy5oZXJybWFubkBhcGhwLmZy
†Present address: Audrey Bernut, Department of Infection, Immunity & Cardiovascular Disease, The Bateson Centre, The University of Sheffield, Sheffield, United Kingdom