 Solène Le Gal1,2*
Solène Le Gal1,2* Gautier Hoarau3
Gautier Hoarau3 Antoine Bertolotti4Steven Negri1
Antoine Bertolotti4Steven Negri1 Nathan Le Nan1
Nathan Le Nan1 Jean-Philippe Bouchara1
Jean-Philippe Bouchara1 Nicolas Papon1Denis Blanchet5,6Magalie Demar5,6
Nicolas Papon1Denis Blanchet5,6Magalie Demar5,6 Gilles Nevez1,2*
Gilles Nevez1,2*- 1Groupe d’Étude des Interactions Hôte-Pathogène (GEIHP) EA 3142, Université d’Angers-Université de Brest, Angers, France
- 2Laboratory of Mycology and Parasitology, CHRU de Brest, Brest, France
- 3Department of Microbiology, CHU La Réunion, Saint Pierre, France
- 4Department of Infectious Diseases, CHU La Réunion, Saint Pierre, France
- 5Laboratory of Mycology and Parasitology, Andrée Rosemon Hospital, Cayenne, French Guiana
- 6Equipe EA3593 - Ecosystèmes Amazoniens et Pathologie Tropicale, Université de Guyane, Cayenne, French Guiana
Data on Pneumocystis jirovecii characteristics from the overseas French territories are still scarce whereas numerous data on P. jirovecii genotypes are available for metropolitan France. The main objective of the present study was to identify P. jirovecii multilocus genotypes in patients living in Réunion and to compare them with those identified using the same method in metropolitan France and in French Guiana. Archival P. jirovecii specimens from immunosuppressed patients, 16 living in Réunion (a French island of the Indian ocean), six living in French Guiana (a South-American French territory), and 24 living in Brest (Brittany, metropolitan France) were examined at the large subunit rRNA (mtLSUrRNA) genes, cytochrome b (CYB), and superoxide dismutase (SOD) genes using PCR assays and direct sequencing. A total of 23 multi-locus genotypes (MLG) were identified combining mtLSUrRNA, CYB, and SOD alleles, i.e., six in Reunionese patients, three in Guianese patients, and 15 in Brest patients. Only one MLG (mtLSU1-CYB1-SOD2) was shared by Reunionese and Guianese patients (one patient from each region) whereas none of the 22 remaining MLG were shared by the 3 patient groups. A total of eight MLG were newly identified, three in Réunion and five in Brest. These results that were obtained through a retrospective investigation of a relatively low number of P. jirovecii specimens, provides original and first data on genetic diversity of P. jirovecii in Réunion island. The results suggest that P. jirovecii organisms from Réunion present specific characteristics compared to other P. jirovecii organisms from metropolitan France and French Guiana.
Introduction
Pneumocystis jirovecii is an opportunistic and transmissible fungus responsible for severe pneumonia Pneumocystis pneumonia (PCP) in immunocompromised patients. PCP remains the most frequent AIDS-defining illness in human immunodeficiency virus (HIV)-infected patients in metropolitan France (Cazein et al., 2015) and the West French Indies (Martinique and Guadeloupe) whereas in French Guiana, another French region of Americas, PCP occupies the fifth position of AIDS causes. PCP is also the most frequent AIDS-defining illness in Réunion, a French island of the Indian Ocean, located close to Capricorn tropic, 600 km from east coast of Madagascar (Cire Océan Indien, 2015). HIV-infection incidence is higher in the French regions of Americas than in metropolitan France whereas its incidence is lower in Réunion (Cazein et al., 2015). PCP is also a severe disease in other immunosuppressed patients who are not infected with HIV, such as patients treated with immunosuppressive and/or cytostatic therapies (Roux et al., 2014).
Data on P. jirovecii characteristics from the overseas French territories are still scarce. Indeed, there is only one report on this topic, which concerned P. jirovecii genotypes in French Guiana (Le Gal et al., 2015) located 7,000 km from metropolitan France whereas there are no data on genomic characteristics of P. jirovecii from Réunion, located 9,300 km from metropolitan France and 12,000 km from French Guiana (Figure 1). Conversely, numerous data on P. jirovecii genotypes are available for metropolitan France (Nevez et al., 2003; Totet et al., 2003; Le Gal et al., 2013; Maitte et al., 2013; Gits-Muselli et al., 2015; Alanio et al., 2016; Desoubeaux et al., 2016; Charpentier et al., 2017; Vindrios et al., 2017). In this context, the main objective of the present study was to identify P. jirovecii multilocus genotypes in patients living in Réunion and to compare these genotypes with those identified using the same method in patients living in metropolitan France or French Guiana.
Figure 1. Geographical locations of Reunion Island, French Guiana and Brest.
Materials and Methods
Pneumocystis jirovecii Specimens and Patients
Seventeen P. jirovecii specimens from 16 patients [sex ratio M/F 13/3, median age 52 years (limits, 30–72 years)] who developed PCP and who were monitored at South Réunion Island University Hospital, were retrospectively studied. The 16 patients were diagnosed with PCP from March 2015 through June 2017. Ten patients had hematological malignancies, five patients were HIV-infected, and one patient had non-X histiocytosis.
Six P. jirovecii specimens from six patients [sex ratio M/F 1/5; median age 33 years (range, 30–57)] monitored at Andrée Rosemon Hospital, Cayenne, French Guiana, were also studied retrospectively. The six patients were HIV-infected and developed PCP from November 2011 through October 2012.
Twenty-four P. jirovecii specimens from 24 patients [sex ratio M/F 16/8, median age 64 years (limits, 33–84 years)] monitored at Brest University Hospital, Brest, Brittany, metropolitan France, were also analyzed retrospectively. These 24 patients were diagnosed with PCP from January 2013 through June 2017. Clinical and biological data of the three patient groups are summarized in Table 1 and detailed in Supplementary Table S1. Data on Guianese patients were previously published elsewhere (Le Gal et al., 2015). Data on Brest patients were previously published in part elsewhere (Nevez et al., 2020).
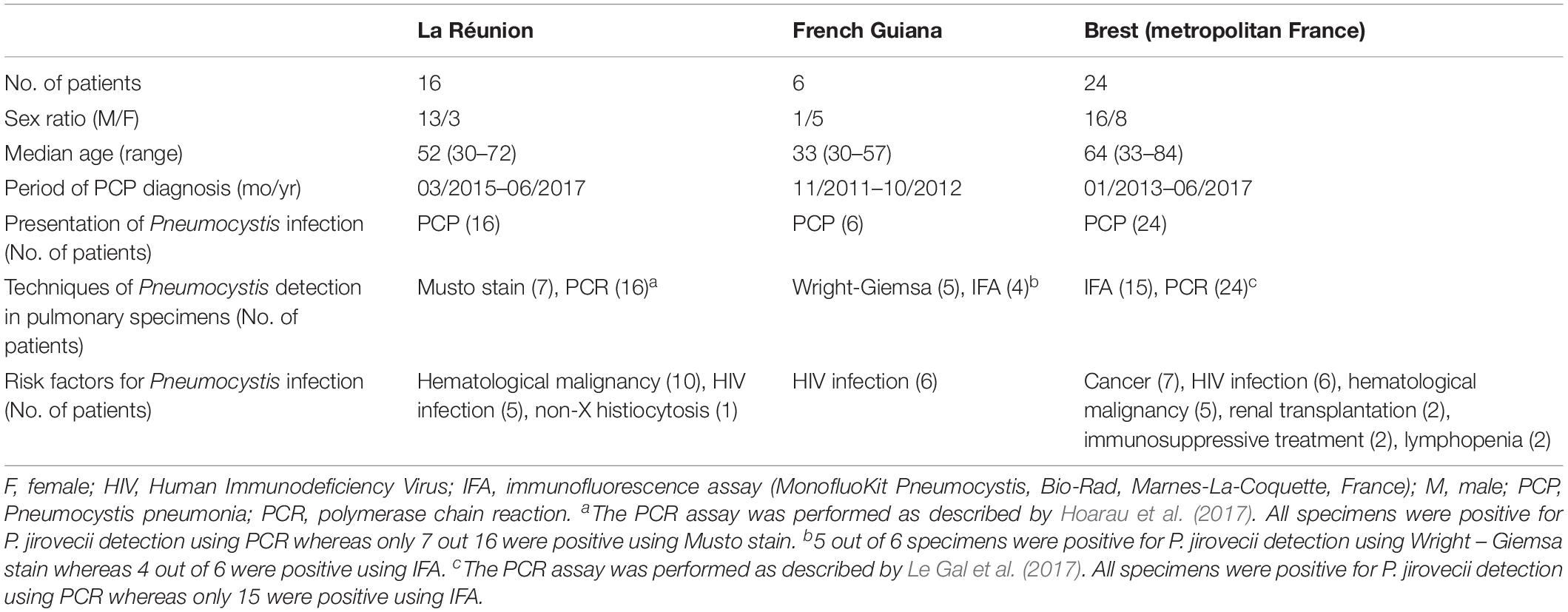
Table 1. Characteristics of the three groups of patients from whom Pneumocystis jirovecii specimens were genotyped.
Patients of the three groups had undergone a bronchoalveolar lavage (BAL) or induced sputum procedure to investigate pulmonary symptoms and/or fever. P. jirovecii had initially been detected in specimens by microscopy using Musto stain, Wright Giemsa stain, and/or an indirect immunofluorescence assay (MonofluoKit Pneumocystis, Bio-Rad, Marnes-La-Coquette, France), and/or PCR assays amplifying the mtLSUrRNA gene as described elsewhere (Hoarau et al., 2017; Le Gal et al., 2017). Extracted DNAs from the three patient groups were stored at −80°C until typing.
The study was non-interventional, and therefore did not require inform consents and ethical approval according to French laws and regulations (CSP Art L1121e1.1).
Pneumocystis jirovecii Typing
Extracted DNAs of BAL and induced sputum samples were examined for Pneumocystis genotyping based on unilocus and multilocus sequence typing (MLST) methods. Three loci, mtLSUrRNA, cytochrome b (CYB) and superoxide dismutase (SOD) genes were analyzed, as we previously described, using direct sequencing (Vindrios et al., 2017). Consensus sequences were aligned with reference sequences [GenBank accession numbers M58605 (mtLSUrRNA), AF074871 (CYB) and KT592355 (SOD)] (Sinclair et al., 1991; Walker et al., 1998; Singh et al., 2017) using the BioEdit software with the Clustal® W program. MtLSUrRNA alleles were named using the nomenclature described previously by Beard et al. (2000), CYB and SOD alleles were named using the nomenclature described previously by Esteves et al. (2010) and Maitte et al. (2013). According to Struelens (1996), the discriminatory power which was determined using Hunter index (H) (Hunter, 1990) was considered good if higher than 0.95. To avoid contamination, each step of the PCR assays was performed in different areas of the laboratory with different sets of micropipettes. Mix reagents were prepared in a laminar flow cabinet. To monitor for possible contamination, negative controls were included in each experiment and PCR round.
The Maximum Likelihood method implemented in MEGA (version 7.0.26) was used to reconstruct a phylogenetic tree based on the Hasegawa-Kishino-Yano model (Kumar et al., 2016). Substitution model was determined by Bayesian Information Criterion in jModelTest 0.1.1 (Posada, 2008). Bootstrap values for internal branches were generated from 1,000 replicates. MtLSUrRNA (209 bp), CYB (563 bp) and SOD (380 bp) sequences were concatenated and aligned to a reference sequence (SeqRef mtLSU CYB SOD). This reference sequence corresponds to a 209-bp portion of mtLSUrRNA reference sequence (M58605) concatenated with a 563-bp portion of CYB reference sequence (AF074871) and a 380-bp portion of SOD reference sequence (KT592355) (Sinclair et al., 1991; Walker et al., 1998; Singh et al., 2017).
Relatedness between Pneumocystis MLGs was evaluated with the minimum spanning tree (MST) method using GrapeTree, a free web browser application implementing Kruskal’s algorithm and Edmonds’ algorithm (Zhou et al., 2018). MLGs were treated as multistate categories based on an infinite allele model, i.e., all changes are equally likely.
Nucleotide Sequence Accession Numbers
The nucleotide sequences of the new CYB allele sequences with changes at scoring positions have been deposited in GenBank under accession numbers MN602710 and MN602711.
Results
Pneumocystis genotyping results are detailed in Table 2. MtLSUrRNA sequences were obtained for the 16 Reunionese patients (17 samples), the six Guianese patients, and the 24 Brest patients. Five alleles were identified, considering the mtLSU alleles previously described elsewhere (Beard et al., 2000; Esteves et al., 2010; Table 2). MtLSU1 was the most frequent allele in Reunionese patients (7 patients; 43.7%). MtLSU3 was the most frequent allele in Guianese patients (3 patients; 50%). MtLSU1, mtLSU2 and mtLSU3 were the most frequent alleles in Brest patients, these three genotypes being equally detected (seven patients each; 29.2%). MtLSU1, mtLSU2 and mtLSU3 were shared by the three patients’ groups while mtLSU4 was shared by Reunionese and Brest patients. MtLSU5 was identified only in one Reunionese patients (6.2%). The presence of more than one allele was observed in seven Reunionese patients (43.7%), two Guianese patients (33.3%) and six patients from Brest (25%). The Hunter index for mtLSU genotyping was evaluated to 0.76.
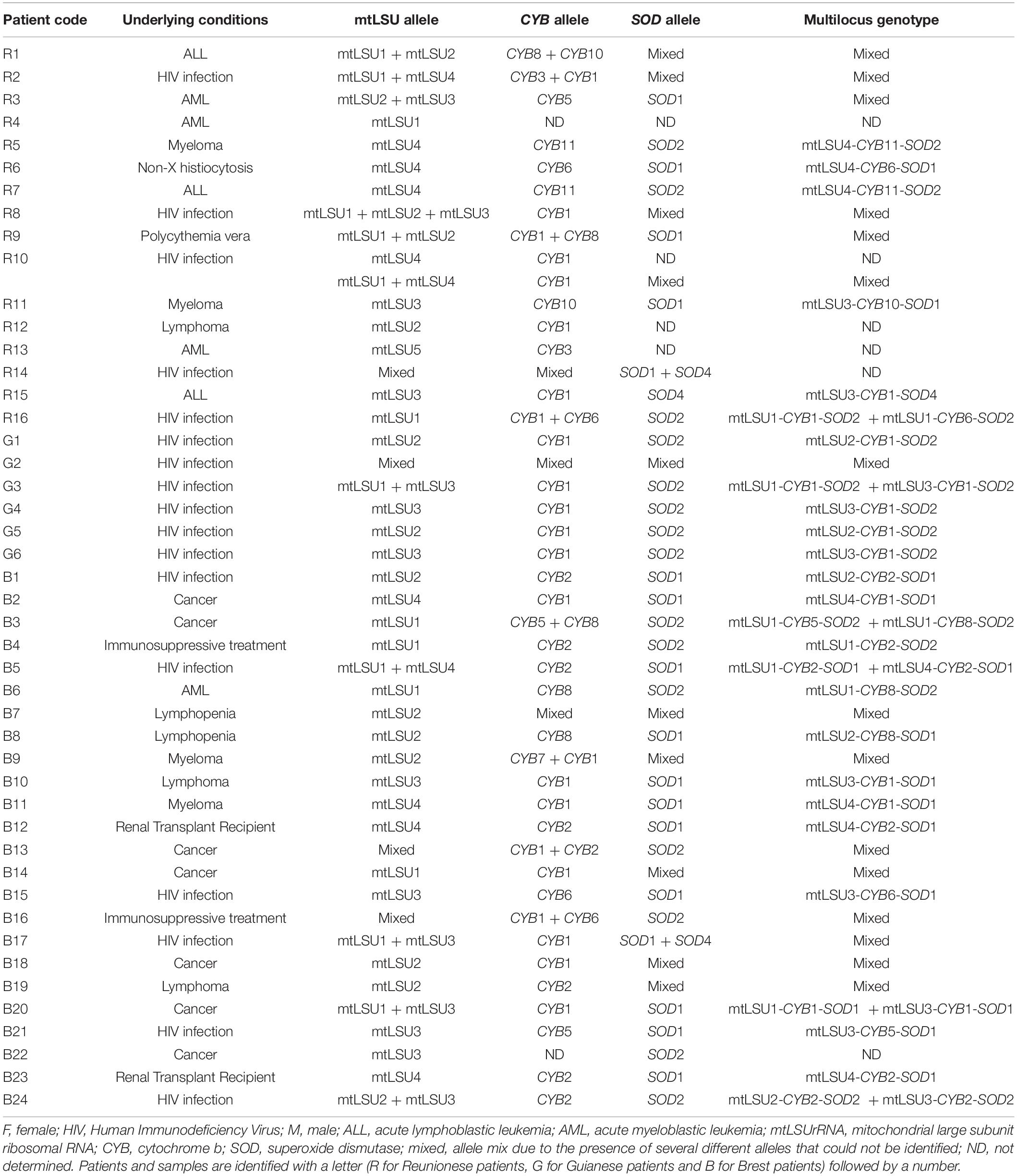
Table 2. Genotypes of Pneumocystis jirovecii identified in patients developing Pneumocystis pneumonia from Réunion, French Guiana and Brest.
CYB sequences were obtained from 15 out of 16 Reunionese patients (16 samples), as well as from the 6 Guianese patients and 23 out of 24 patients from Brest. Considering the CYB alleles previously described elsewhere (Esteves et al., 2010; Maitte et al., 2013), seven already known alleles (CYB1, CYB2, CYB3, and CYB5 to CYB8) and two new alleles (CYB10 and CYB11) were identified (Table 2), CYB1 being the most frequent in the three patient groups [seven Reunionese patients (46.7%), five Guianese patients (83.3%), 10 Brest patients (43.5%)]. The new CYB10 allele differs from CYB2 by a change from C to T residue at scoring position 279, and the new CYB11 allele differs from CYB1 by a change from C to T residue at scoring position 742. CYB1 was shared by the three populations. In contrast, CYB3, CYB10, and CYB11 were identified only in Reunionese patients [three (20%), three (20%) and two (13.3%) patients, respectively] while CYB2 and CYB7 were detected only in Brest patients [eight (37.8%) patients and one (4.3%) patient, respectively]. The presence of more than one allele was observed in five Reunionese patients (33.3%), one Guianese patient (16.7%) and five patients from Brest (21.7%). The Hunter index for CYB genotyping was evaluated to 0.76.
SOD sequences were obtained from 13 out of the 16 Reunionese patients, as well as from the six Guianese patients and the 24 patients from Brest. Considering the SOD alleles previously described elsewhere (Esteves et al., 2010; Maitte et al., 2013), three alleles were identified (Table 2), SOD1 being the most frequent in Reunionese and Brest patients (five (38.5%) and 12 (50%) patients, respectively) and SOD2 the most frequent in Guianese patients [five patients (83.3%)]. SOD2 was identified in the three patient groups. The presence of more than one allele was observed in five Reunionese patients (38.5%), one Guianese patient (16.7%) and six patients from Brest (25%). The Hunter index for SOD genotyping was evaluated to 0.59.
Combining mtLSUrRNA, CYB, and SOD alleles, 23 multi-locus genotypes (MLG) were identified (Table 2). MtLSU4-CYB11-SOD2, mtLSU3-CYB1-SOD2, and mtLSU4-CYB2-SOD1 were the most frequent MLG in Reunionese patients (two patients, 16.7%), Guianese patients (three patients, 50%), and Brest patients (three patients, 13%), respectively. Only one MLG (mtLSU1-CYB1-SOD2) was shared by the Reunionese and the Guianese patients (one patient from each region, 8.3 and 16.7% respectively) whereas the 22 remaining MLG were not shared by the three patient groups. The presence of more than one genotype was observed in seven Reunionese patients (58.3%), two Guianese patients (33.3%) and 12 patients from Brest (52.2%). The Hunter index for MLST was evaluated to 0.978.
The phylogenetic tree was generated based on sequence analysis of mtLSUrRNA, CYB, and SOD alleles through the MLST approach (Figure 2). The analysis showed that some genotypes detected in patients from metropolitan France and the Réunion island were close despite the rarity of MLST genotype sharing. Nonetheless, the bootstrap values were low (<50).
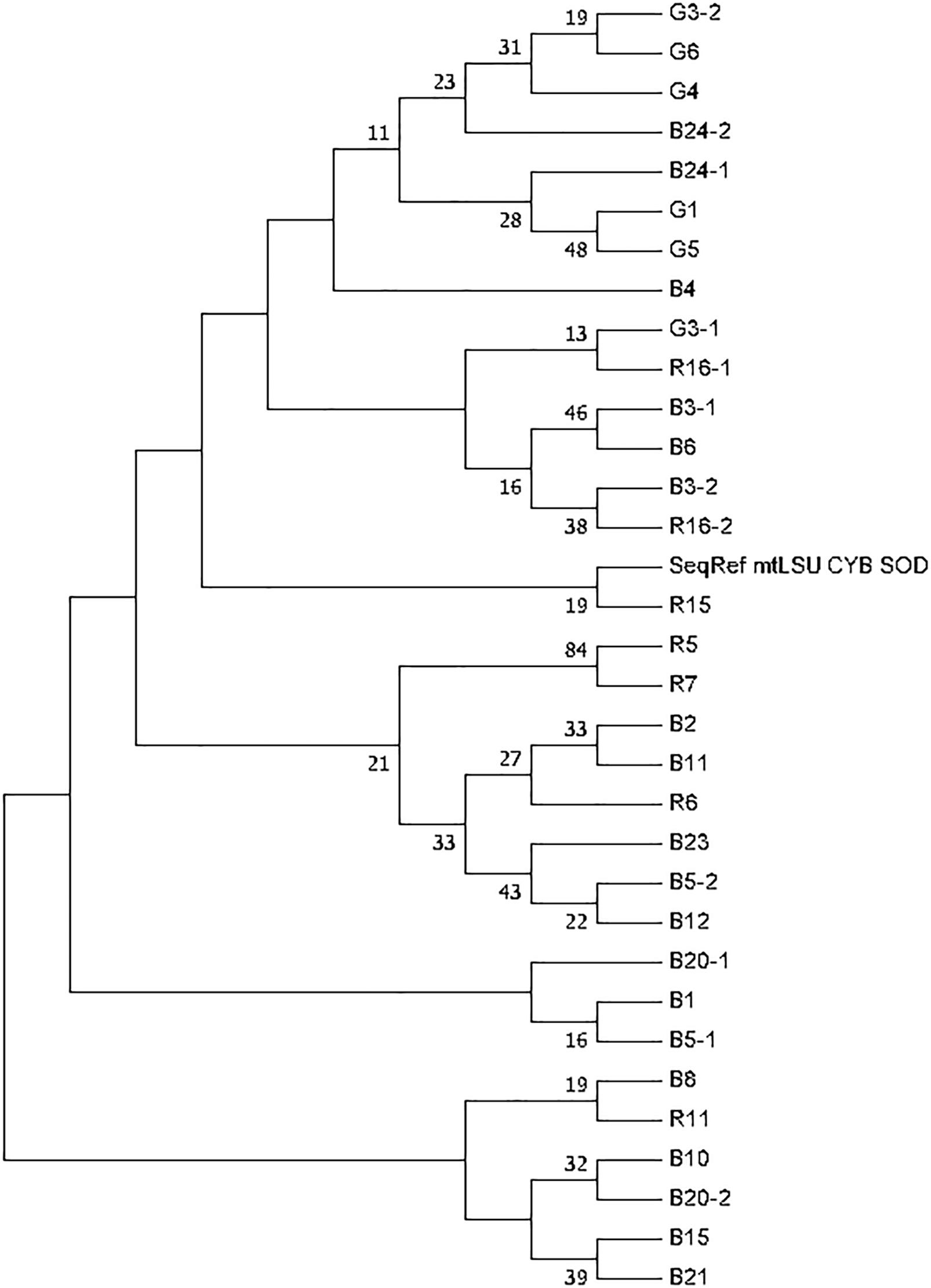
Figure 2. Phylogenetic tree generated based on Pneumocystis jirovecii MLST genotypes. The tree was constructed on the basis of concatenated loci (mtLSUrRNA, CYB, and SOD). Phylogenetic analysis used the Maximum Likelihood method based on the Hasegawa-Kishino-Yano model. The bootstrap consensus tree was inferred from 1,000 replicates. Branches corresponding to partitions reproduced in less than 50% bootstrap replicates are collapsed. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test are shown next to the branches. Only bootstrap values >10% are shown. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value. The analysis involved 33 nucleotide sequences. There were a total of 1152 positions in the final dataset. Evolutionary analyses were conducted in MEGA7. Patients and samples are identified with a letter (R for Reunionese patients, G for Guianese patients and B for Brest patients) followed by a number.
As well, MST analysis revealed that most of Pneumocystis isolates from the three geographic regions were close and belonged to the same genetic cluster since they had a single allelic mismatch with at least one other isolate (Figure 3). However, two isolates from Réunion who shared the same MLG (mtLSU4-CYB11-SOD2) were more distant from the other isolates since they had two allelic mismatches with the closest isolate.
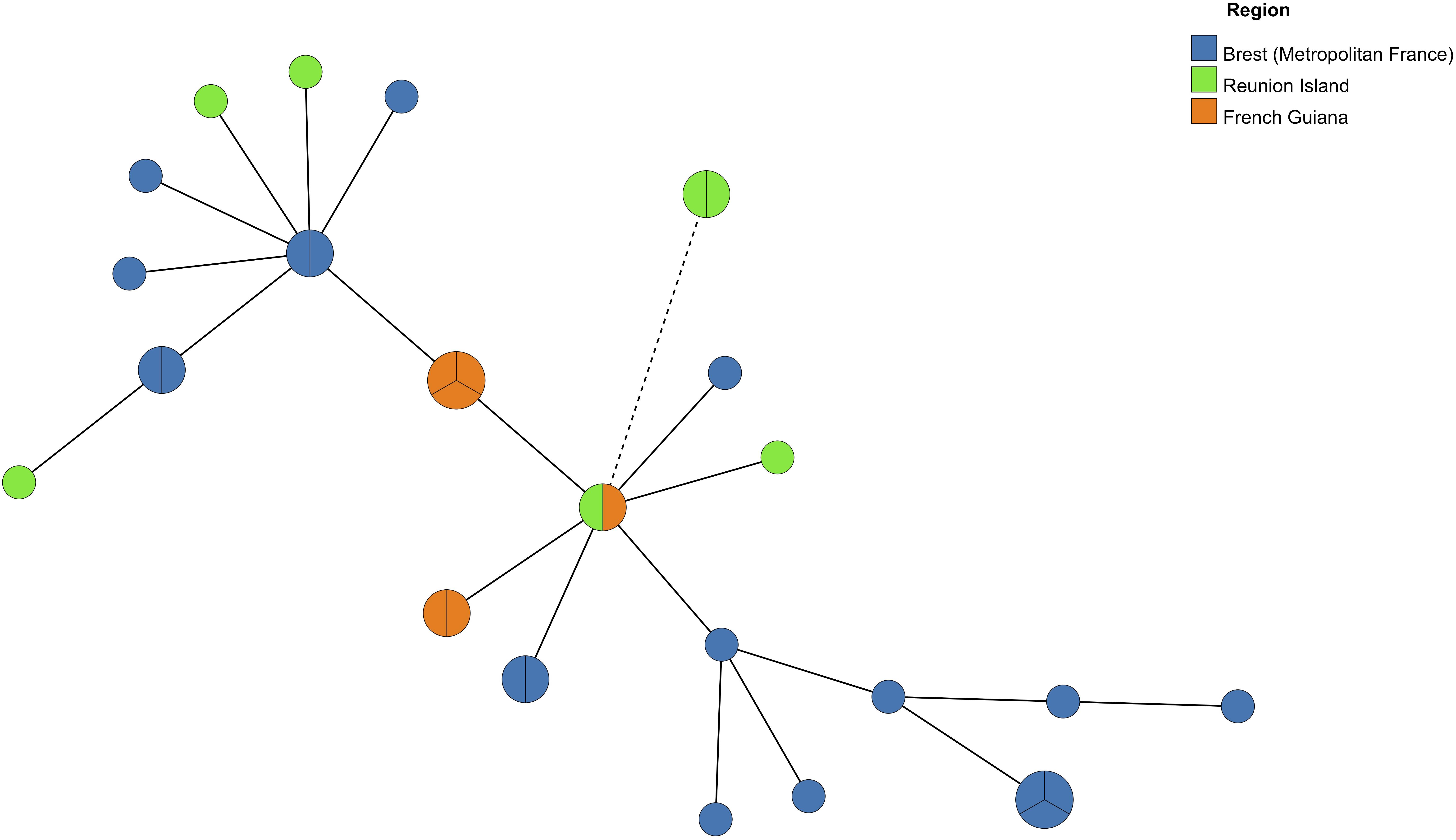
Figure 3. Minimum Spanning Tree analysis of Pneumocystis isolates from six Reunionese patients, five Guianese patients and 15 Brest (Metropolitan France) patients harboring one multilocus genotype (MLG) (one allele per locus) or two MLGs (two alleles for one locus). Each circle corresponds to one MLG. The size of the circles is correlated with the number of isolates possessing the corresponding MLG. The color of the circles indicates the origin region of the Pneumocystis isolates. A continuous connecting line corresponds to one different allele separating two MLGs. A dashed connecting line corresponds to two different alleles separating two MLGs.
Discussion
In this study, the first data on P. jirovecii genotypes in patients from Réunion, a French region in the Indian Ocean, were obtained. Using an unilocus approach, we identified mtLSUrRNA, CYB, and SOD common alleles in the three patient populations (Réunion, French Guiana, and Brest), suggesting that P. jirovecii organisms from Réunion, French Guiana and Metropolitan France share common characteristics. Nonetheless, some alleles were identified only from one patient population, e.g., mtLSU5, CYB10, and CYB11 were identified only from Reunionese patients.
We chose to analyze mtLSUrRNA, CYB, and SOD loci because this MLST scheme is highly discriminant as described by Maitte et al. (2013). Indeed, in the present study, the discriminatory power of our technique, based on the Hunter index, was evaluated to 0.978, which is considered to be good, i.e., >0.95, one criterion of those described by the ESCMID to validate a typing method (Struelens, 1996). The sequencing was performed using Sanger method, which is easy to be performed. However, it could be less sensitive to detect and identify alleles in a mixture than PCR-SSCP or high-throughput methods, such as ultra-deep sequencing or multiplex PCR-Single-Base-Extension (Hauser et al., 2001; Esteves et al., 2011, 2016; Alanio et al., 2016; Charpentier et al., 2017). In the present study mixed alleles were identified in 15 out of 46 patients (32.6%) for mtLSUrRNA, in 11 out of 44 patients (25%) for CYB, and in 12 out of 43 patients (27.9%) for SOD. Considering MLG analysis, mixed MLGs were identified in 21 out of 42 patients (50%). These rates are lower than those observed in studies using PCR-SSCP (Hauser et al., 2001) or molecular high-throughput methods, such as ultra-deep sequencing (Alanio et al., 2016; Charpentier et al., 2017), in which rates of mixed genotypes may reach 85% for nuclear loci and even 92% for mitochondrial loci. However, this potential weakness of our method should not question our analysis based on major genotypes. Moreover, the method we used is suitable for examining samples with low fungal loads (Vindrios et al., 2017; Nevez et al., 2018, 2020).
The alleles CYB10 and CYB11 identified in Reunionese patients were newly described in this study. Allele CYB10 differs from allele CYB2 described by Esteves et al. (2010) by the substitution from C to T residue at position 279, which is a silent mutation. However, this allele corresponds to haplotype 13 as described by Charpentier et al. (2017) using another nomenclature. It was identified in two patients from Grenoble, metropolitan France. Allele CYB11 differs from allele CYB1 by having a T residue at position 742. This substitution from C to T represents a non-synonymous mutation leading to the substitution from leucyl to phenylalanyl residue at position 275 (L275F). This mutation located at the quinol oxidation (Qo) site of the mitochondrial cytochrome bc1 may induce potential atovaquone resistance of P. jirovecii. Indeed, atovaquone which is used as second line treatment of PCP or PCP prophylaxis, is an analog of ubiquinone that binds the Qo site. This mutation, but not this allele, has previously been described by Kazanjian et al. (2001) among P. jirovecii isolates from patients with past history of atovaquone exposure. It is noteworthy that the two Reunionese patients, who were infected with cytochrome b mutant P. jirovecii organisms, were effectively subjected to atovaquone prophylaxis in a context of hematological malignancy (myeloma for one patient, and T-cell acute lymphoblastic leukemia for the other), at the time of PCP diagnosis. This observation was consistent with that we recently reported concerning the selection of cytochrome b mutants in heart transplant recipients in a context of PCP case clusters and the use of atovaquone prophylaxis (Argy et al., 2018).
In this study, allele mtLSU5 was identified only from one Reunionese patient. This infrequent allele has been previously identified in metropolitan patients from Brest and Lille (de Armas et al., 2012; Le Gal et al., 2015) and in a Guianese patient (patient G2 in the present study) (Le Gal et al., 2015) using cloning instead of direct sequencing of the mtLSUrRNA sequences. Thus, this allele does not represent a specific characteristic of P. jirovecii organisms from Réunion island.
There are available data on genotyping of P. jirovecii isolates from adult patients or children who lived in Zimbabwe or Mozambique (Africa), two countries relatively close to Réunion and from other adults or children who lived in Cuba (West Indies), a country relatively close to French Guiana (Miller et al., 2003, 2005; de Armas et al., 2012; Monroy-Vaca et al., 2014; Esteves et al., 2016). This geographical proximity deserves P. jirovecii genotype comparison. Considering the results of unilocus typing, mtLSU3 allele is the most frequent allele in adults from Cuba as well as in adults from French Guiana (de Armas et al., 2012), whereas mtLSU2 is the most frequent allele in children from Cuba (Monroy-Vaca et al., 2014). To the best of our knowledge, no information on SOD and CYB alleles in adults from Cuba is available. Conversely, in children, CYB1 and SOD1 are the most frequent alleles in Cuba whereas alleles CYB1 and SOD2 are the most frequent alleles in adults in French Guiana. No data on P. jirovecii genotyping in children from French Guiana are available.
MtLSU3 and mtLSU1 are the most frequent alleles in adults from Zimbabwe and Reunion respectively (Miller et al., 2003). In the same patient population, SOD2 and SOD1 are the most frequent alleles in Zimbabwe and Reunion respectively (Miller et al., 2003). Conversely, in children from Mozambique the most frequent allele is mtLSU2 and/or mtLSU5 (considering that information on scoring nucleotide position 248 is lacking) (Esteves et al., 2016). No data on CYB alleles from Zimbabwe and Mozambique, whichever patient population, are available.
Finally, due to differences in characteristics of the studied patient populations (adults vs. children, numbers of patients) and analyzed loci, it remains difficult to draw a conclusive analysis of this genotype comparison.
A total of 23 MLG were identified. It is noteworthy that 22 were not shared by the three patient group whereas only one MLG was shared by two of the three patient groups. Thus, the results of MLG analysis, due to the high discriminatory power of the method (Hunter index, 0.978), allow to discriminate P. jirovecii isolates into three different groups according to the geographic origin of the patients. Nonetheless, among the five MLG identified only in Reunionese patients, two (mtLSU1-CYB6-SOD2 and mtLSU3-CYB1-SOD4) has already been reported in patients from metropolitan France and Portugal (Esteves et al., 2010; Maitte et al., 2013). Conversely, the three other MLG (mtLSU3-CYB10-SOD1, mtLSU4-CYB11-SOD2, and mtLSU4-CYB6-SOD1) were reported for the first time in the present study. It is noteworthy that the MLG mtLSU4-CYB11-SOD2 consists in the combination of mtLSU4 allele, SOD2 allele and the newly described allele CYB11 which might have been selected in the course of atovaquone prophylaxis. Be that as it may, taken together, these results suggest that P. jirovecii organisms from Réunion island may present specific characteristics.
Three MLG were identified in Guianese patients; all of them have already been reported in patients from metropolitan France or Portugal (Esteves et al., 2010; Maitte et al., 2013; Desoubeaux et al., 2016; Charpentier et al., 2017). These results are not consistent with those previously obtained using another genotyping method (Le Gal et al., 2015), which suggested that specific genotypes and consequent specific characteristics of P. jirovecii organisms may exist in French Guiana. This could be explained by the fact that more data are available in literature on MLG combining mtLSUrRNA, CYB, and SOD alleles than on MLG combining ITS, DHPS, and mtLSUrRNA alleles, the method we used previously.
Fifteen MLG were identified in Brest patients, of which ten have already been reported in patients from metropolitan France and Portugal (Esteves et al., 2010; Maitte et al., 2013; Desoubeaux et al., 2016; Charpentier et al., 2017; Vindrios et al., 2017) whereas five were reported for the first time. These five MLG (mtLSU1-CYB2-SOD2, mtLSU1-CYB3-SOD1, mtLSU1-CYB5-SOD2, mtLSU2-CYB2-SOD2, and mtLSU3-CYB2-SOD2) may represent specific characteristics of P. jirovecii organisms in Brittany, western France. Likewise, the three MLG identified in Réunion island may represent specific characteristics of P. jirovecii organisms in this French overseas island. However, the results of the MST suggest that P. jirovecii organisms from the three French regions are closely related and belong to the same genetic cluster, excepting two isolates from Réunion. These two isolates share the same new MLG (mtLSU4-CYB11-SOD2) (see above).
Finally, a total of eight MLG were newly identified, three in Réunion and five in Brest, suggesting that specific characteristics in these two French regions, located 9,300 km apart may exist. However, genotyping results should not be too conclusive considering the low number of patients for whom MLG were identified in this study (six Reunionese patients, five Guianese patients, 15 patients from Brest). Moreover, the dates of P. jirovecii sampling in the three geographical regions were not identical, which may represent a bias of enrolment. Likewise, the underlying diseases of the patients were not strictly similar since Guianese patients were all HIV-infected contrary to Reunionese patients and patients from Brest, these disparities representing a bias as well. Furthermore, available data on MLG combining mtLSUrRNA, CYB, and SOD alleles are still limited since only four studies from France and one from Portugal were based on these sequences (Esteves et al., 2010; Maitte et al., 2013; Desoubeaux et al., 2016; Charpentier et al., 2017; Vindrios et al., 2017). Moreover, the results of phylogenetic tree and MST analysis are poorly informative. Be that as it may, despite these possible limitations, the present study brings original and first data on genetic diversity of P. jirovecii organisms from Réunion island and its comparison with other very distant French regions.
Data Availability Statement
The datasets generated for this study can be found in the GenBank database (accession numbers: MN602710 and MN602711).
Ethics Statement
Ethical review and approval was not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements.
Author Contributions
SL and GN analyzed the DNA sequences and wrote the manuscript. GH and AB performed P. jirovecii detection, provided P. jirovecii specimens from Réunion, and analyzed patients’ characteristics. SN and NL performed in part the genotyping. J-PB and NP contributed to the discussion and correction of the manuscript. DB and MD performed P. jirovecii detection and provided P. jirovecii specimens, and analyzed patients’ characteristics from French Guiana.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.00127/full#supplementary-material
References
Alanio, A., Gits-Muselli, M., Mercier-Delarue, S., Dromer, F., and Bretagne, S. (2016). Diversity of Pneumocystis jirovecii during infection revealed by ultra-deep pyrosequencing. Front. Microbiol. 7:733. doi: 10.3389/fmicb.2016.00733
Argy, N., Le Gal, S., Coppee, R., Song, Z., Vindrios, W., Massias, L., et al. (2018). Pneumocystis cytochrome B mutants associated with atovaquone prophylaxis failure as the cause of Pneumocystis infection outbreak among heart transplant recipients. Clin. Infect. Dis. 67, 913–919. doi: 10.1093/cid/ciy154
Beard, C. B., Carter, J. L., Keely, S. P., Huang, L., Pieniazek, N. J., Moura, I. N., et al. (2000). Genetic variation in Pneumocystis carinii isolates from different geographic regions: implications for transmission. Emerg. Infect. Dis. 6, 265–272. doi: 10.3201/eid0603.000306
Cazein, F., Pillonel, J., Le Strat, Y., Pinget, R., Le Vu, S., Brunet, S., et al. (2015). Découvertes de séropositivité VIH et de SIDA, France, 2003-2013. Bull. Epidemiol. Hebd. 9- 10 152–161.
Charpentier, E., Garnaud, C., Wintenberger, C., Bailly, S., Murat, J. B., Rendu, J., et al. (2017). Added value of next-generation sequencing for multilocus sequence typing analysis of a Pneumocystis jirovecii pneumonia outbreak1. Emerg. Infect. Dis. 23, 1237–1245. doi: 10.3201/eid2308.161295
Cire Océan Indien (2015). Surveillance des infections à VIH et Sida à la Réunion et Mayotte, région océan Indien de 2003 à 2014, données actualisées au 31/12/2014. Bull. Veille Sanitaire 29, 2–7.
de Armas, Y., Friaza, V., Capo, V., Durand-Joly, I., Govin, A., de la Horra, C., et al. (2012). Low genetic diversity of Pneumocystis jirovecii among Cuban population based on two-locus mitochondrial typing. Med. Mycol. 50, 417–420. doi: 10.3109/13693786.2011.607474
Desoubeaux, G., Dominique, M., Morio, F., Thepault, R. A., Franck-Martel, C., Tellier, A. C., et al. (2016). Epidemiological outbreaks of Pneumocystis jirovecii pneumonia are not limited to kidney transplant recipients: genotyping confirms common source of transmission in a liver transplantation unit. J. Clin. Microbiol. 54, 1314–1320. doi: 10.1128/JCM.00133-16
Esteves, F., Gaspar, J., De Sousa, B., Antunes, F., Mansinho, K., and Matos, O. (2011). Clinical relevance of multiple single-nucleotide polymorphisms in Pneumocystis jirovecii Pneumonia: development of a multiplex PCR-single-base-extension methodology. J Clin Microbiol. 49, 1810–1815. doi: 10.1128/JCM.02303-10
Esteves, F., Gaspar, J., Tavares, A., Moser, I., Antunes, F., Mansinho, K., et al. (2010). Population structure of Pneumocystis jirovecii isolated from immunodeficiency virus-positive patients. Infect., Genet. Evol. 10, 192–199. doi: 10.1016/j.meegid.2009.12.007
Esteves, F., de Sousa, B., Calderon, E. J., Huang, L., Badura, R., Maltez, F., et al. (2016). Multicentre study highlighting clinical relevance of new high-throughput methodologies in molecular epidemiology of Pneumocystis jirovecii pneumonia. Clin. Microbiol. Infect. 22:e569.e9–e566.e19. doi: 10.1016/j.cmi.2016.03.013
Gits-Muselli, M., Peraldi, M. N., de Castro, N., Delcey, V., Menotti, J., Guigue, N., et al. (2015). New short tandem repeat-based molecular typing method for Pneumocystis jirovecii reveals intrahospital transmission between patients from different wards. PloS One 10:e0125763. doi: 10.1371/journal.pone.0125763
Hauser, P., Blanc, D. S., Sudre, P., Senggen Manoloff, E., Nahimana, A., Bille, J., et al. (2001). Genetic diversity of Pneumocystis carinii in HIV-positive and -negative patients as revealed by PCR-SSCP typing. AIDS 15, 461–466. doi: 10.1097/00002030-200103090-00004
Hoarau, G., Le Gal, S., Zunic, P., Poubeau, P., Antok, E., Jaubert, J., et al. (2017). Evaluation of quantitative FTD-Pneumocystis jirovecii kit for Pneumocystis infection diagnosis. Diagn. Microbiol. Infect. Dis. 89, 212–217. doi: 10.1016/j.diagmicrobio.2017.08.001
Hunter, P. R. (1990). Reproducibility and indices of discriminatory power of microbial typing methods. J. Clin. Microbiol. 28, 1903–1905. doi: 10.1128/jcm.28.9.1903-1905.1990
Kazanjian, P., Armstrong, W., Hossler, P. A., Lee, C. H., Huang, L., Beard, C. B., et al. (2001). Pneumocystis carinii cytochrome b mutations are associated with atovaquone exposure in patients with AIDS. J. Infect. Dis. 183, 819–822. doi: 10.1086/318835
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093/molbev/msw054
Le Gal, S., Blanchet, D., Damiani, C., Gueguen, P., Virmaux, M., Abboud, P., et al. (2015). AIDS-related Pneumocystis jirovecii genotypes in French Guiana. Infect. Genet. Evol. 29, 60–67. doi: 10.1016/j.meegid.2014.10.021
Le Gal, S., Robert-Gangneux, F., Pepino, Y., Belaz, S., Damiani, C., Gueguen, P., et al. (2017). A misleading false-negative result of Pneumocystis real-time PCR assay due to a rare punctual mutation: a French multicenter study. Med. Mycol. 55, 180–184. doi: 10.1093/mmy/myw051
Le Gal, S., Rouille, A., Gueguen, P., Virmaux, M., Berthou, C., Guillerm, G., et al. (2013). Pneumocystis jirovecii haplotypes at the internal transcribed spacers of the rRNA operon in French HIV-negative patients with diverse clinical presentations of Pneumocystis infections. Med. Mycol. 51, 851–862. doi: 10.3109/13693786.2013.824123
Maitte, C., Leterrier, M., Le Pape, P., Miegeville, M., and Morio, F. (2013). Multilocus sequence typing of Pneumocystis jirovecii from clinical samples: how many and which loci should be used? J. Clin. Microbiol. 51, 2843–2849. doi: 10.1128/JCM.01073-13
Miller, R. F., Lindley, A. R., Ambrose, H. E., Malin, A. S., and Wakefield, A. E. (2003). Genotypes of Pneumocystis jiroveci isolates obtained in Harare, Zimbabwe, and London, United Kingdom. Antimicrob. Agents Chemother. 47, 3979–3981. doi: 10.1128/aac.47.12.3979-3981.2003
Miller, R. F., Lindley, A. R., Malin, A. S., Ambrose, H. E., and Wakefield, A. E. (2005). Isolates of Pneumocystis jirovecii from Harare show high genotypic similarity to isolates from London at the superoxide dismutase locus. Trans. R. Soc. Trop. Med. Hyg. 99, 202–206. doi: 10.1016/j.trstmh.2004.09.005
Monroy-Vaca, E. X., De Armas, Y., Illnait-Zaragozi, M. T., Diaz, R., Torano, G., Vega, D., et al. (2014). Genetic diversity of Pneumocystis jirovecii in colonized Cuban infants and toddlers. Infect. Genet. Evol. 22, 60–66. doi: 10.1016/j.meegid.2013.12.024
Nevez, G., Le Gal, S., Noel, N., Wynckel, A., Huguenin, A., Le Govic, Y., et al. (2018). Investigation of nosocomial pneumocystis infections: usefulness of longitudinal screening of epidemic and post-epidemic pneumocystis genotypes. J. Hosp. Infect. 99, 332–345. doi: 10.1016/j.jhin.2017.09.015
Nevez, G., Totet, A., Jounieaux, V., Schmit, J. L., Dei-Cas, E., and Raccurt, C. (2003). Pneumocystis jiroveci internal transcribed spacer types in patients colonized by the fungus and in patients with pneumocystosis from the same French geographic region. J. Clin. Microbiol. 41, 181–186. doi: 10.1128/jcm.41.1.181-186.2003
Nevez, G., Guillaud-Saumur, T., Cros, P., Papon, N., Vallet, S., Quinio, D., et al. (2020). Pneumocystis primary infection in infancy: additional french data and review of the literature. Med. Mycol. 58, 163–171. doi: 10.1093/mmy/myz040
Posada, D. (2008). JModelTest: phylogenetic model averaging. Mol. Biol. Evol. 25, 1253–1256. doi: 10.1093/molbev/msn083
Roux, A., Canet, E., Valade, S., Gangneux-Robert, F., Hamane, S., Lafabrie, A., et al. (2014). Pneumocystis jirovecii pneumonia in patients with or without AIDS, France. Emerg. Infect. Dis. 20, 1490–1497. doi: 10.3201/eid2009.131668
Sinclair, K., Wakefield, A. E., Banerji, S., and Hopkin, J. M. (1991). Pneumocystis carinii organisms derived from rat and human hosts are genetically distinct. Mol Biochem Parasitol. 45, 183–184. doi: 10.1016/0166-6851(91)90042-5
Singh, Y., Mirdha, B. R., Guleria, R., Khalil, S., Panda, A., Chaudhry, R., et al. (2017). Circulating genotypes of Pneumocystis jirovecii and its clinical correlation in patients from a single tertiary center in India. Eur. J. Clin. Microbiol. Infect. Dis. 36, 1635–1641. doi: 10.1007/s10096-017-2977-9
Struelens, M. J. (1996). Consensus guidelines for appropriate use and evaluation of microbial epidemiologic typing systems. Clin. Microbiol. Infect. 2, 2–11. doi: 10.1111/j.1469-0691.1996.tb00193.x
Totet, A., Pautard, J. C., Raccurt, C., Roux, P., and Nevez, G. (2003). Genotypes at the internal transcribed spacers of the nuclear rRNA operon of Pneumocystis jirovecii in nonimmunosuppressed infants without severe pneumonia. J. Clin. Microbiol. 41, 1173–1180. doi: 10.1128/jcm.41.3.1173-1180.2003
Vindrios, W., Argy, N., Le Gal, S., Lescure, F. X., Massias, L., Le, M. P., et al. (2017). Outbreak of Pneumocystis jirovecii infection among heart transplant recipients: molecular investigation and management of an interhuman transmission. Clin. Infect. Dis. 65, 1120–1126. doi: 10.1093/cid/cix495
Walker, D. J., Wakefield, A. E., Dohn, M. N., Miller, R. F., Baughman, R. P., Hossler, P. A., et al. (1998). Sequence polymorphisms in the Pneumocystis carinii cytochrome b gene and their association with atovaquone prophylaxis failure. J Infect Dis. 178, 1767–1775.
Keywords: Pneumocystis jirovecii, genotypes, Réunion, Pneumocystis pneumonia, multilocus sequence typing (MLST), French Guiana, France
Citation: Le Gal S, Hoarau G, Bertolotti A, Negri S, Le Nan N, Bouchara J-P, Papon N, Blanchet D, Demar M and Nevez G (2020) Pneumocystis jirovecii Diversity in Réunion, an Overseas French Island in Indian Ocean. Front. Microbiol. 11:127. doi: 10.3389/fmicb.2020.00127
Received: 18 October 2019; Accepted: 20 January 2020;
Published: 07 February 2020.
Edited by:
Olga Matos, New University of Lisbon, PortugalReviewed by:
Francisco Esteves, Universidade Nova de Lisboa, PortugalMagali Chabé, Université de Lille, France
Copyright © 2020 Le Gal, Hoarau, Bertolotti, Negri, Le Nan, Bouchara, Papon, Blanchet, Demar and Nevez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Solène Le Gal, c29sZW5lLmxlZ2FsQGNodS1icmVzdC5mcg==; Gilles Nevez, Z2lsbGVzLm5ldmV6QGNodS1icmVzdC5mcg==