 Lu Meng1,2†
Lu Meng1,2† Huimin Liu1,2†
Huimin Liu1,2† Tu Lan1,2
Tu Lan1,2 Lei Dong1,2
Lei Dong1,2 Haiyan Hu1,2,3
Haiyan Hu1,2,3 Shengguo Zhao1,2Yangdong Zhang1,2
Shengguo Zhao1,2Yangdong Zhang1,2 Nan Zheng1,2
Nan Zheng1,2 Jiaqi Wang1,2*
Jiaqi Wang1,2*- 1Laboratory of Quality and Safety Risk Assessment for Dairy Products of Ministry of Agriculture and Rural Affairs, Institute of Animal Sciences, Chinese Academy of Agricultural Sciences, Beijing, China
- 2Key Laboratory of Quality & Safety Control for Milk and Dairy Products of Ministry of Agriculture and Rural Affairs, Institute of Animal Sciences, Chinese Academy of Agricultural Sciences, Beijing, China
- 3College of Animal Science and Technology, Anhui Agricultural University, Hefei, China
Psychrotrophic bacteria in raw milk are most well known for their spoilage potential and the economic losses they cause to the dairy industry. Food-related psychrotrophic bacteria are increasingly reported to have antibiotic resistance features. The aim of this study was to evaluate the resistance patterns of Pseudomonas spp. isolated from bulk-tank milk. In total, we investigated the antibiotic susceptibility profiles of 86 Pseudomonas spp. isolates from raw milk. All strains were tested against 15 antimicrobial agents. Pseudomonas isolates were most highly resistant to imipenem (95.3%), followed by trimethoprim-sulfamethoxazole (69.8%), aztreonam (60.5%), chloramphenicol (45.3%), and meropenem (27.9%). Their multiple antibiotic resistance (MAR) index values ranged from 0.0 to 0.8. Whole-genome sequencing revealed the presence of intrinsic resistance determinants, such as BcI, ampC-09, blaCTX-M, oprD, sul1, dfrE, catA1, catB3, catI, floR, and cmlV. Moreover, resistance-nodulation-cell division (RND) and ATP-binding cassette (ABC) antibiotic efflux pumps were also found. This study provides further knowledge of the antibiotic resistance patterns of Pseudomonas spp. in milk, which may advance our understanding of resistance in Pseudomonas and suggests that antibiotic resistance of Pseudomonas spp. in raw milk should be a concern.
Introduction
Pseudomonas spp., which have been identified as predominantly psychrotrophic bacteria, are important spoilage bacteria in food (Marchand et al., 2009). The Pseudomonas genus is found extensively in environments such as water, soil, and sediment (Devarajan et al., 2016). They are ubiquitous Gram-negative bacteria that have a wide metabolic versatility, which allows them to acclimate to different habitats with temperatures ranging from 4 to 42°C (Quigley et al., 2013; Molina et al., 2014; Chakravarty and Gregory, 2015). Moreover, Pseudomonas can multiply at cold temperatures and accounts for more than half of all bacteria found in milk (Munsch-Alatossava and Alatossava, 2006; Fricker et al., 2011; Von Neubeck et al., 2015). Pseudomonas strains produce heat-stable extracellular peptidases and/or lipases that spoil raw milk, and some strains are opportunistic pathogens that are found in the environment, such as Pseudomonas aeruginosa (Jeukens et al., 2017).
The widespread administration of antimicrobial agents to animals used for food products has imposed a strong selective pressure that increases resistance among known pathogens and commensal bacteria (Beena et al., 2011; Decimo et al., 2016). Antimicrobial resistant Pseudomonas is also a concern as antimicrobial resistance is a key factor in the emergence of infectious diseases (Jones et al., 2008; Gomi et al., 2017). A study based on phenotypes and 16S rDNA gene sequences identified Pseudomonas isolates that were categorized as being a high risk for antibiotic resistance (Munsch-Alatossava et al., 2012). Moreover, Pseudomonas spp. have the ability to remain viable in the aquatic environment for long periods owing to innate resistance mechanisms, thereby increasing the risk of spreading antibiotic resistance genes and mobile genetic elements (Devarajan et al., 2016). The exchange of genetic material encoding resistance genes via mobile genetic elements, plasmids, or transposons can result in transfer of resistance between pathogenic and non-pathogenic bacteria (Mathur and Singh, 2005; Panelists, 2006; Decimo et al., 2016). Yomoda et al. (2003) reported that 22 of 32 Pseudomonas putida strains isolated from Japanese hospitals carried plasmids that could be transferred to P. aeruginosa by conjugation or transformation. Therefore, the transfer of antibiotic resistance determinants from non-pathogenic species to pathogens is a serious concern (Yomoda et al., 2003; Molina et al., 2014).
Antibiotic resistance has been a focus of research on food-related and mastitis pathogens in milk for many years. The potential for non-pathogenic commensal foodborne bacteria to become a biological hazard has also been investigated as resistance genes can be spread to humans via food (Decimo et al., 2016). The aim of this study was to evaluate the resistance of Pseudomonas spp. isolated from bulk-tank milk in China to different antibiotics. This study represents an extensive investigation of resistance of Pseudomonas spp. to a number of human and veterinary antimicrobial agents.
Materials and Methods
Bacterial Isolates
A total of 143 Pseudomonas strains were isolated from raw milk samples from 87 bulk tanks in Shaanxi province, China, in spring 2016 (average daily temperature >20°C) for a previous study of their proteolytic properties (Meng et al., 2017). All isolates were re-identified using the rpoD gene as rpoD provides higher resolution than 16S rDNA gene sequences for Pseudomonas species (Rajwar and Sahgal, 2016). In total, 86 isolates representing 11 different Pseudomonas spp. were subjected to antimicrobial susceptibility testing. A brief description of these 86 isolates is provided in Supplementary Table S1.
Antimicrobial Susceptibility Testing
Antimicrobial susceptibility tests were performed using broth microdilution with cation-adjusted Mueller–Hinton broth (BD, Franklin Lake, NJ, United States) following Clinical and Laboratory Standards Institute (Clinical and Laboratory Standards Institute [CLSI]) guidelines (2018). The antimicrobials used for susceptibility testing included 15 antimicrobial agents belonging to the following classes: penicillins (ampicillin, penicillin), monobactams (aztreonam), cephamycins (cefoxitin), phenicols (chloramphenicol), fluoroquinolones (ciprofloxacin, levofloxacin), lincosamides (clindamycin), lipopeptides (polymyxin B), aminoglycosides (gentamicin), carbapenems (imipenem, meropenem), tetracyclines (tetracycline), folate pathway inhibitors (trimethoprim-sulfamethoxazole), and β-lactams (ceftiofur). Of these antibiotics, ampicillin, penicillin, cefoxitin, ciprofloxacin, gentamicin, tetracycline, and ceftiofur are used as veterinary medicines (MOA, 2002). Our results were interpreted according to European Committee on Antimicrobial Susceptibility Testing (European Committee on Antimicrobial Susceptibility Testing [EUCAST], 2018) and Clinical and Laboratory Standards Institute [CLSI] criteria (2016, 2018). Multidrug-resistant (MDR) strains were defined as being resistant to three or more antimicrobial classes. However, because neither European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2018) nor Clinical and Laboratory Standards Institute [CLSI] (2018) criteria provide minimal inhibitory concentration (MIC) breakpoints for ampicillin, penicillin, cefoxitin, clindamycin, or ceftiofur for non-Enterobacteriaceae, results for these five antibiotics are provided as MIC values.
The multiple antibiotic resistance (MAR) index for each isolate was determined for 10 antimicrobial agents according to Blasco et al. (2008):
Whole Genome Sequencing
Strains exhibiting the same antimicrobial resistance patterns were excluded from sequencing analysis. After exclusion, 44 isolates were selected for sequencing. The genomic DNA of each isolate was extracted using a Wizard® Genomic DNA Purification Kit (Promega Corporation, Fitchburg, WI, United States) following the manufacturer’s protocol. DNA samples were sheared into 400- to 500-bp fragments using a Covaris M220 Focused-Ultrasonicator (ThermoFisher Scientific, Waltham, MA, United States) according to the manufacturer’s protocol. Illumina sequencing libraries were then prepared from the sheared fragments using a NEXTflexTM Rapid DNA-Seq Kit (Bioo Scientific, Austin, TX, United States). Briefly, 5’ prime ends were first end repaired and phosphorylated. Next, 3’ ends were A-tailed and ligated to sequencing adapters. Adapter-ligated products were then enriched using PCR. Finally, the prepared libraries were used for paired-end Illumina sequencing (2 × 150 bp) on an Illumina HiSeq X Ten machine (Illumina, San Diego, CA, United States).
Analysis of Sequence Data
Data generated from the Illumina platform were used for bioinformatics analysis. All analyses were performed using the I-Sanger Cloud Platform1 from Shanghai Majorbio BioTech Co., Ltd. (Shanghai, China). Briefly, the original image data were transferred into sequence data, defined as raw data or raw reads, by base calling and saved as a FASTQ file. A quality statistic was then applied for quality trimming to remove low-quality data from the clean data. Assembly of the clean reads was performed using SOAPdenovo2 (Luo et al., 2015). Finally, resistance genes were detected using the ResFinder web server and ARG-ANNOT database (Zankari et al., 2012; Gupta et al., 2014). All whole genome sequence data from this study have been deposited in the NCBI Sequence Read Archive database (accession numbers PRJNA523883, PRJNA523885, and PRJNA593738).
Results
Antimicrobial Susceptibility Profiles of Pseudomonas spp.
Isolates were tested for susceptibility to 15 antibiotics. Results were first interpreted according to European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2018), followed by Clinical and Laboratory Standards Institute [CLSI] (2016, 2018). Table 1 shows the Pseudomonas spp. that were found to belong to different interpretive categories, including susceptible (S), intermediate (I), and resistant (R), for which MIC breakpoints are available). Wide variability was observed in the susceptibility of isolates to gentamicin, ciprofloxacin, tetracycline, and levofloxacin. Approximately 95% of Pseudomonas strains were susceptible to gentamicin and ciprofloxacin, followed by 93.0% that were susceptible to tetracycline and 91.9% that were susceptible to levofloxacin. However, a high percentage of Pseudomonas strains were resistant to imipenem (95.3%). Resistance to trimethoprim-sulfamethoxazole (69.8%), aztreonam (60.5%), chloramphenicol (45.3%), and meropenem (27.9%) was also widespread in this study. The distribution of resistance patterns to the 10 antibiotics tested is shown in Figure 1. In total, 48 (55.8%) isolates were resistant to three or more drug classes and were defined as MDR strains. One isolate was not resistant to all eight classes of antibiotics, and one isolate could resist seven classes of antibiotics. Moreover, the 86 tested isolates had MAR index values ranging from 0.0 to 0.8 (Supplementary Table S2).
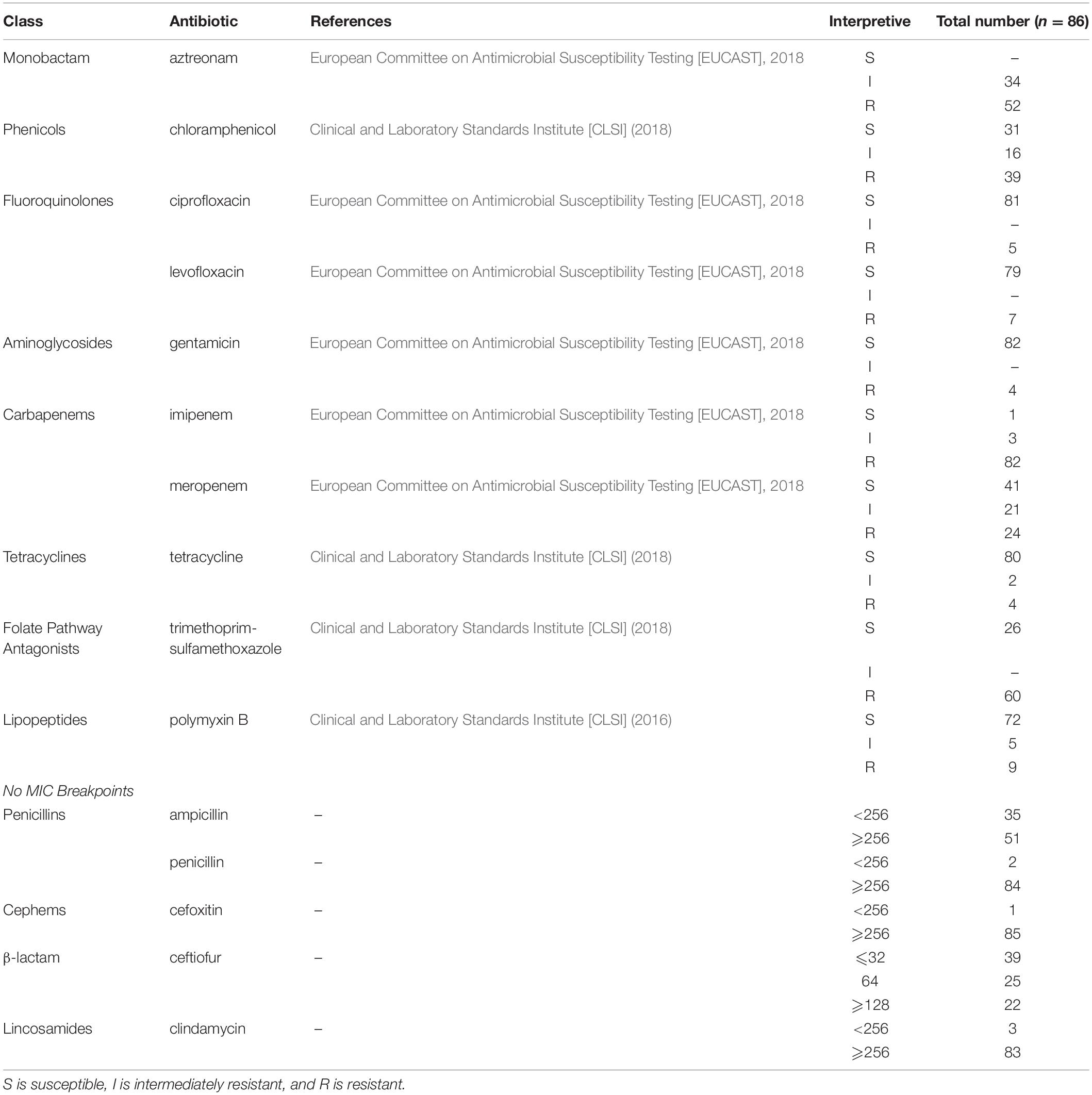
Table 1. The interpretive categories of Pseudomonas strains to the tested antibiotics.
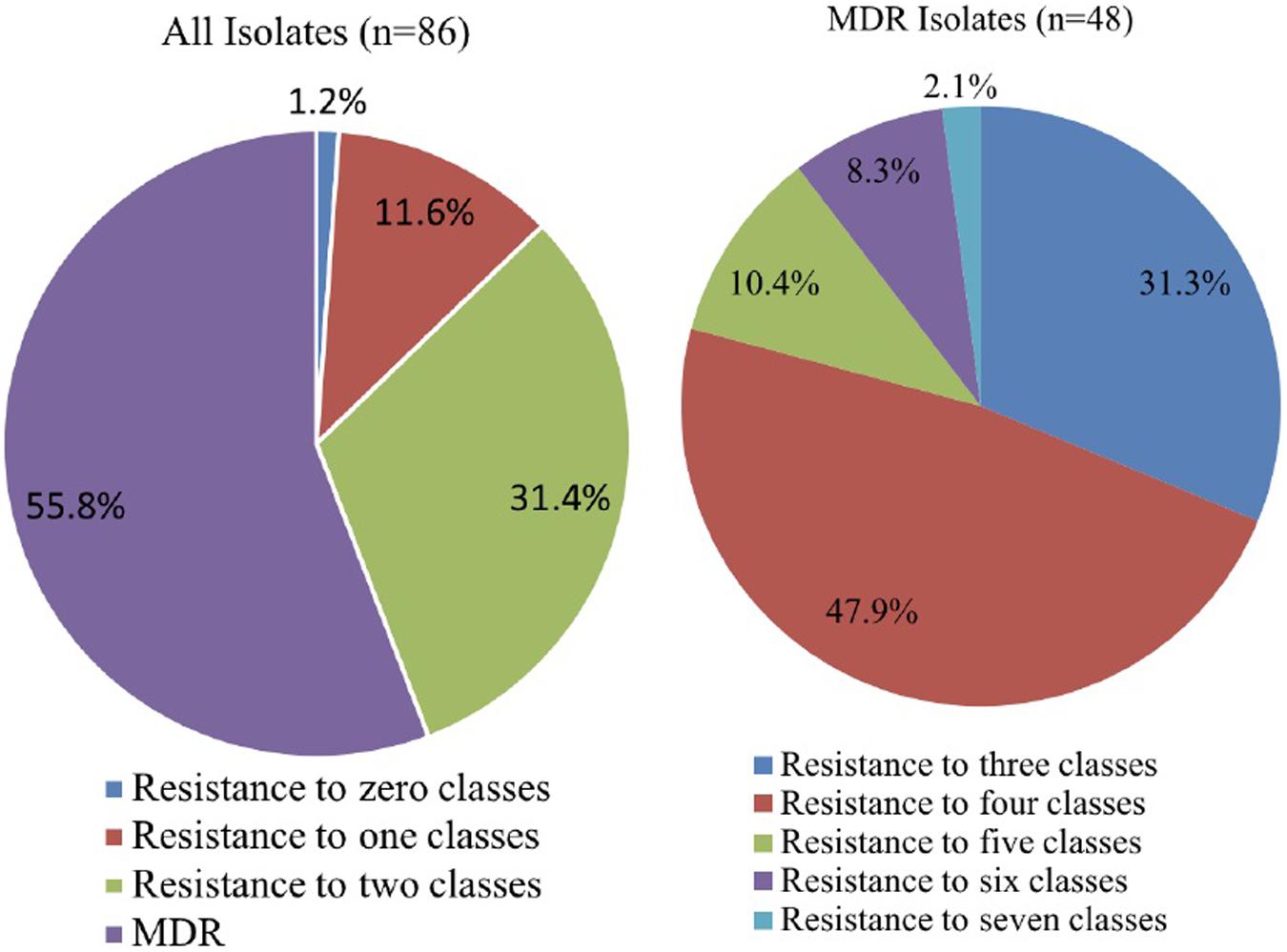
Figure 1. Distribution of resistance patterns.
For the antibiotics ampicillin, cefoxitin, ceftiofur, clindamycin, and penicillin, no European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2018) or Clinical and Laboratory Standards Institute [CLSI] (2016, 2018) breakpoints are available for Pseudomonas spp. We found MIC values ≥256 μg/mL for 85 isolates for cefoxitin, 84 isolates for penicillin, and 83 isolates for clindamycin. All MIC values are shown in Supplementary Table S1.
Resistance Determinants
Overall, more than 100 resistance determinants were found in 44 isolates (Table 2), and all of the isolates carried resistance determinants. Eleven different aminoglycoside resistance genes were detected. The most prevalent aminoglycoside resistance gene, aph(6)-Ic, is one of three resistance genes present in the composite transposon Tn5, which is found in Gram-negative bacteria (Steiniger-White et al., 2004). The ksgA gene, which confers resistance to kasugamycin (Zou et al., 2018), was also found in seven isolates. Genes ant(3”)-Ia, aph(6)-Id and aph(3’)-IIc as well as rpsL mutations, known to be responsible for streptomycin resistance (Ramirez and Tolmasky, 2010), were also found in these isolates (Supplementary Table S3).
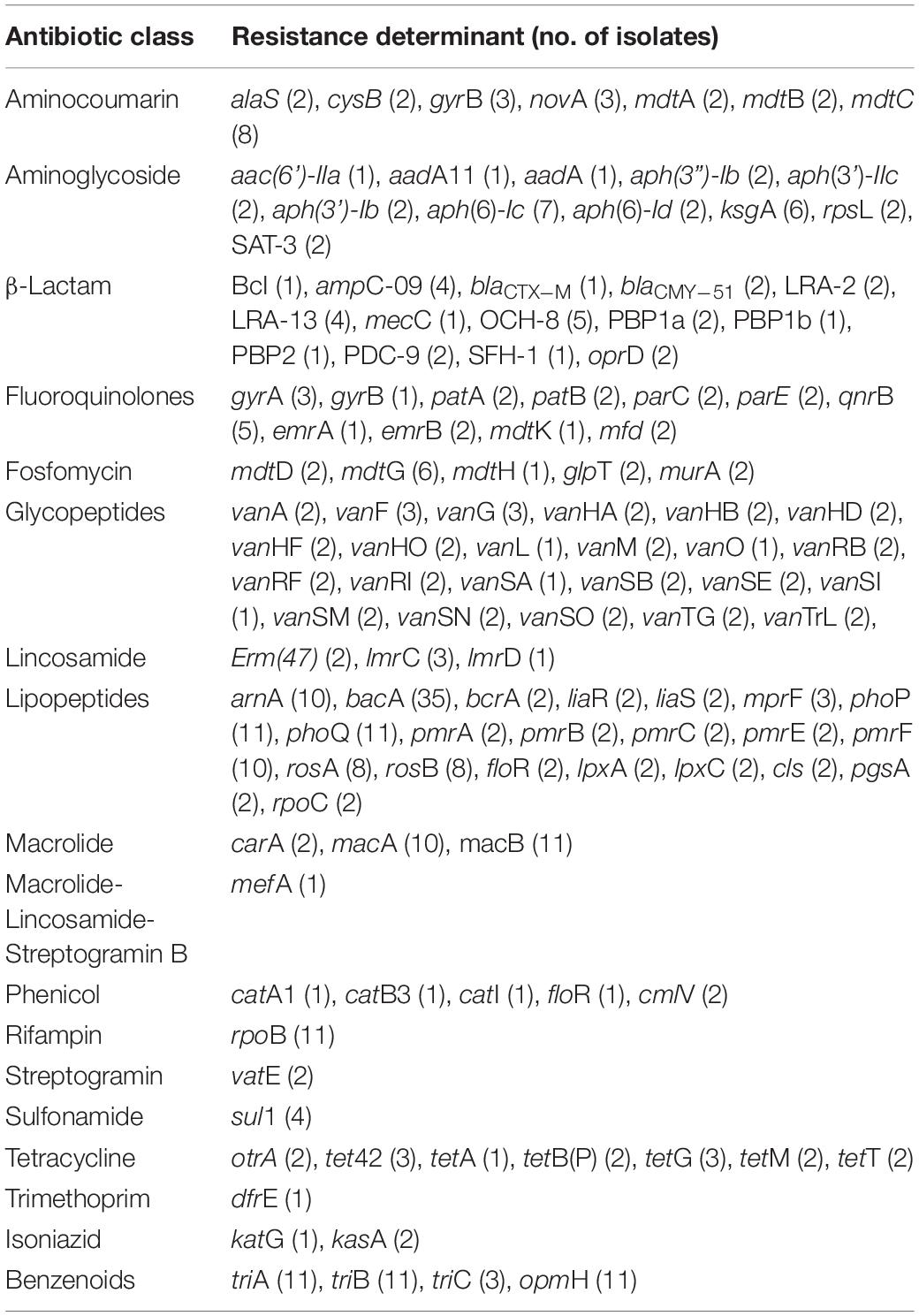
Table 2. Resistance determinants detected in 44 isolates.
Fourteen different β-lactam resistance genes were detected. Although the total number of β-lactam resistance genes was high, the prevalence of each gene was low. Of the β-lactam resistance determinants, OCH-8 confers resistance to third-generation cephalosporins (Alonso et al., 2017), whereas LRA-13 confers resistance to amoxicillin, ampicillin, cephalexin, and carbenicillin (Silveira et al., 2018). Penicillin-binding proteins (PBPs) are the physiological targets of β-lactam antibiotics (Spratt and Cromie, 1988), and mutations in the PBPs PBP1a, PBP1b, and PBP2, which were found to confer resistance to β-lactams (Di Guilmi et al., 2003; Stanhope et al., 2008; Unemo et al., 2012), have also been detected in four isolates (Table 2 and Supplementary Table S3). In addition, the remaining nine β-lactam resistance genes have been demonstrated to confer resistance to cephalosporin, carbapenem, monobactam, and cephamycin (Carfi et al., 1995; Saavedra et al., 2003; Rodriguez-Martinez et al., 2009; Milheirico et al., 2017).
Another important resistance gene group was found to confer resistance to lipopeptides. In total, 21 resistance genes have been detected with 12 genes conferring resistance to polymyxin, such as phoP and phoQ. The most common gene, bacA, is a bacitracin resistance gene (Shaaly et al., 2013), and bcrA is another one, but it is only found in two isolates. floR is a resistance gene against florfenicol (Arcangioli et al., 1999). Additionally, mutations in liaR, liaS, cls, pgsA, and rpoC, known to be responsible for daptomycin resistance (Yang et al., 2010; Arias et al., 2011; Peleg et al., 2012; Miller et al., 2013), were found in a few strains (Table 2 and Supplementary Table S3).
In total, 16 resistance genes were speculated from other bacteria, such as gyrA, gyrB, rpsL, PBP1a, PBP1b, PBP2, liaR, liaS, cls, pgsA, rpoC, rpoB, katG, and kasA. These genes confer resistance to aminocoumarin, aminoglycoside, β-lactam, fluoroquinolones, fosfomycin, lipopeptides, rifampin, and isoniazid. Moreover, it is speculated that all the rifampin and isoniazid resistance determinants were transferred from Escherichia coli, Mycobacterium tuberculosis, and Staphylococcus aureus.
In addition to these resistance genes, genes for resistance-nodulation-cell division (RND) and ATP-binding cassette (ABC) antibiotic efflux pumps, including AcrAB-TolC, MexAB-OprM, MexCD-OprJ, MexEF-OprN, MexJK-OpmH, MexMN-OprM, MexPQ-OpmE, MexVW-OprM, and EfrAB, were also detected (Depardieu et al., 2007; Chen and Duan, 2016). These RND efflux pumps can be mediated by the local repressor gene mutations, global regulatory gene mutations, or other mutations except MexAB-OprM and MexXY-OprM, which contribute to intrinsic multidrug resistance in P. aeruginosa PAO1 (Morita et al., 2001a; Piddock, 2006).
Discussion
Bacterial antibiotic resistance is considered a worldwide problem in the medical, environmental, and agricultural fields. Many researchers have focused on antibiotic resistance in pathogenic bacteria, which pose immediate risks to human health. However, more and more interests are focusing on commensal bacteria associated with food (Munsch-Alatossava et al., 2012; Decimo et al., 2016). Pseudomonas spp. isolated from raw milk shows extensive resistance to many antibiotics, and some even exhibit the highest known levels of antibiotic resistance (Straley et al., 2006; Munsch-Alatossava and Alatossava, 2007; Decimo et al., 2016).
EUCAST and CLSI criteria are the standards for bacterial resistance assessment. However, clinical breakpoints are based on parameters that are only relevant for therapeutic purposes. Therefore, assessing the antibiotic susceptibility of environmental bacteria using only CLSI criteria is inadequate. In contrast, EUCAST criteria are clinically and/or epidemiologically based and are more reliable for the interpretation of the antibiotic susceptibility of environmental bacteria (Berendonk et al., 2015; Gomi et al., 2017). Therefore, European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2018) criteria were used in the present study to assess the resistance phenotypes of Pseudomonas spp. to aztreonam, ciprofloxacin, levofloxacin, gentamicin, imipenem, and meropenem. However, Clinical and Laboratory Standards Institute [CLSI] (2016, 2018) criteria were used to interpret chloramphenicol, polymyxin B, tetracycline, and trimethoprim-sulfamethoxazole resistance as there is no MIC value for these antibiotics in the European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2018) criteria.
According to a previous survey, β-lactam antibiotics are commonly used for dairy mastitis therapy (Liu et al., 2017). Therefore, seven β-lactam antibiotics (ampicillin, aztreonam, cefoxitin, ceftiofur, imipenem, meropenem, and penicillin) were tested in the present study. Around 60% of Pseudomonas strains were resistant to aztreonam, which is higher than described by Decimo et al. (2016). A study conducted between 2010 and 2012 reported that 51–100% of Pseudomonas spp. in non-turbid drinking water were resistant to aztreonam (Flores Ribeiro et al., 2014). In contrast to our results, which showed that 95.3% of Pseudomonas strains were resistant to imipenem, Decimo et al. (2016) found that only 11% of Pseudomonas spp. from bulk-tank milk were resistant to imipenem, and Arslan et al. (2011) found that all Pseudomonas spp. isolates from cheese were susceptible to imipenem. In the present study, the meropenem resistance rate was only 27.9% (n = 24), which is similar to resistance rates reported in previous studies of Pseudomonas spp. from bulk-tank milk and the Danube River (Decimo et al., 2016; Kittinger et al., 2016). Although no breakpoints were available for the four other β-lactam antibiotics, we found that 59.3, 98.8, 2.3, and 97.7% of isolates had MIC value ≥256 μg/mL for ampicillin, cefoxitin, ceftiofur, and penicillin, respectively. In the 1980s, studies reported that P. aeruginosa that were the dominant cause of nosocomial infections were resistant to almost all aminoglycosides and β-lactams with the exception of cephalosporins and carbapenems (Neu, 1983; Acar, 1985). Penicillin G is still the most commonly used antibiotic for dry cow therapy (United States Department of Agriculture, Animal Plant Health Inspection Service National Animal Health Monitoring System [USDA APHIS], 2008), and one study found that 100% of Pseudomonas spp. isolates from cheese were resistant to penicillin G (Arslan et al., 2011).
In the present study, 14 different β-lactamase genes were detected in approximately 20 of the Pseudomonas isolates, which were resistant to at least one β-lactams antibiotic from among aztreonam, imipenem, and meropenem except 149-2 and 152-2. Gram-negative bacteria produce AmpC-type β-lactamases that can hydrolyze amino- and ureido-penicillins, cephamycin, and—at low levels—cephalosporin and monobactams (Rodriguez-Martinez et al., 2009). Four isolates carrying ampC-09 were all resistant to aztreonam, MIC value for cefoxitin ≥256 μg/mL, and MIC value for cefoxitin ≥64 μg/mL except 198-5. Two isolates 141-6 and 151-5 harbored blaCMY–51, an ampC variant first described in Citrobacter freundii (Porres-Osante et al., 2014), and were resistant to aztreonam, MIC value for cefoxitin ≥256 μg/mL, and MIC value for cefoxitin ≥128 μg/mL. The isolate 114-2 was intermediately resistant to aztreonam with the gene blaCTX–M, which has been demonstrated to hydrolyze aztreonam (Wang et al., 2011). All five isolates with OCH-8 and LRA-2 showed MIC value 128 μg/mL for ceftiofur. However, five isolates taking BcI, blaCTX–M, and LRA-13 showed a ceftiofur MIC value for ceftiofur ≤64 μg/mL. The resistance genes that have been found were much lower than the β-lactam resistance rates. Meanwhile, Pseudomonas spp. are reported to be naturally resistant to penicillin G as well as the majority of related β-lactam antibiotics (Decimo et al., 2016). The primary mechanisms of protection of Pseudomonas against β-lactams are the production of β-lactamases, the decrease or loss of the OprD porin in the outer membrane, and the overproduction of RND efflux pumps (Wolter and Lister, 2013). However, RND efflux pumps contribute to increased resistance to β-lactams, β-lactamase inhibitors, and certain carbapenems, including penicillin, aztreonam, meropenem, and ampicillin but not imipenem, when mediated by the local repressor gene mutations or global regulatory gene mutations. For example, a global regulatory gene, soxR, in P. aeruginosa has also been described to regulate efflux pumps (Palma et al., 2005; Piddock, 2006). And mutations in the local repressor genes acrR, nfxB, mexS, and mexL can induce overexpression of AcrAB-TolC, MexCD-OprJ, MexEF-OprN, and MexJK, respectively (Poole et al., 1996; Chuanchuen et al., 2002; Sobel et al., 2005).
Although no blaIMP, blaVIM, and blaNDM have been detected, carbapenem resistance genes PCD-9 and SFH-1 were found in three isolates. Of these, 68-1 and 103-1 showed resistance to imipenem and meropenem and 106-3 only to imipenem. Moreover, imipenem and meropenem resistance in P. aeruginosa have been found to be associated with the loss of the OprD porin combined with the activity of chromosomal β-lactamase (AmpC) (Livermore, 2001; El Amin et al., 2005). However, unlike imipenem, meropenem can be expelled by MexB-mediated efflux (Livermore, 2001). All imipenem-/meropenem-resistant isolates in the present study had oprD-loss mutations with the exception of two isolates, 141-6 and 151-5, that were resistant to both imipenem and meropenem but with oprD. Fang et al. (2014) demonstrated that reduced expression of oprD without oprD loss was the predominant mechanism of imipenem resistance.
In addition to β-lactam antibiotics, ciprofloxacin, sulfamethoxazole-trimethoprim, and gentamicin are also frequently used for dairy mastitis therapy in China (Liu et al., 2017). However, Pseudomonas spp. in the present study exhibited high susceptibility to fluoroquinolone (ciprofloxacin, 94.2%; levofloxacin, 91.9%) and aminoglycoside (gentamicin, 95.3%) antibiotics (Table 1). Our results confirmed the findings of Munsch-Alatossava and Alatossava (2007) and Decimo et al. (2016), who reported antibiotic resistance patterns of Gram-negative psychrotrophic bacteria from bulk-tank milk in Finland and Italy. However, psychrotrophic isolates from raw and pasteurized milk have been found to be resistant to gentamicin (Beena et al., 2011; Munsch-Alatossava et al., 2012). In addition to the 12 resistance genes found in 86 isolates, the enzymes ANT(3’), APH(3’), APH(6), and RpsL mutations confer resistance to streptomycin (Sreevatsan et al., 1996; Ramirez and Tolmasky, 2010). The gene aac(6’)-IIa, which can produce enzymes that actively mediate acetylation of gentamicin, was found in 114-2 that exhibited gentamicin resistance. Moreover, the RND efflux pump MexXY can be induced by sub-inhibitory concentrations of gentamicin and acquire resistance against aminoglycosides when overexpressed (Morita et al., 2001b).
Fluoroquinolone resistance has been associated with mutations in the genes gyrA and parC (Aoike et al., 2013). The mutations were found in two isolates, 141-6 and 151-5, that were resistant to ciprofloxacin and levofloxacin. Moreover, these two isolates also carried eight other quinolones resistance determinants, including mutations in the quinolone resistance-determining regions of DNA gyrase (gyrB) and DNA topoisomerase IV (parE). Isolate 103-1, which carried a gyrA mutation from E. coli, also exhibited resistance to ciprofloxacin and levofloxacin. Five isolates harbored the plasmid-mediated quinolone resistance gene qnrB; however, these isolates were all susceptible to ciprofloxacin and levofloxacin. Moreover, 60 isolates were resistant to trimethoprim-sulfamethoxazole. However, only five isolates carried the trimethoprim resistance gene dfrE or the sulfamethoxazole resistance gene sul1 (Toleman et al., 2007; Chen et al., 2019). Resistance to ciprofloxacin and levofloxacin in the other isolates may have been due to RND efflux pumps (Kohler et al., 1996; Li et al., 2015).
Polymyxin lipopeptide antibiotics are currently last-resort antibiotics for the treatment of MDR Gram-negative bacterial infections. Of 21 lipopeptide resistance genes, 12 confer resistance to polymyxin, including arnA, phoP, phoQ, pmrA, pmrB, pmrC, pmrE, pmrF, rosA, rosB, lpxA, and lpxC, with phoP and phoQ being the most common (McPhee et al., 2003; Gatzeva-Topalova et al., 2005; Lee and Ko, 2014; Zhang et al., 2017). Polymyxin B resistance has been found to be conferred by mutations in two-component regulatory systems of phoPQ, which are well known to contribute to polymyxin resistance in P. aeruginosa (Barrow and Kwon, 2009; Miller et al., 2011; Lee and Ko, 2014). The PmrF operon also plays an essential role in resistance to polymyxin and can be upregulated by PhoPQ (Marceau et al., 2004). In the present study, the number of mutations in phoPQ regions was higher than polymyxin B resistance rates, and a few polymyxin B–resistant isolates carried the fosmidomycin resistance genes rosA and rosB, which encode an efflux pump that confers resistance to polymyxin B in response to polymyxin B (Bengoechea and Skurnik, 2000).
In our study, resistance rates of Pseudomonas isolates to chloramphenicol and tetracycline were 45.3% (39/86) and 4.7% (4/86), respectively, which was much lower than the ratios of Pseudomonas aeruginosa isolated from milk samples and from major hospitals and laboratories in Jamaica (Brown and Izundu, 2004; Decimo et al., 2016). The chloramphenicol resistance is mainly caused by enzymatic inactivation by chloramphenicol acetyltransferases (CAT) and also efflux mechanisms mediated by cml genes (Schwarz et al., 2004). In total, cat and cml genes were found in five isolates, which is less than the number of chloramphenicol resistance isolates (n = 39). However, the effects of simultaneous expression of multidrug transporters, such as AcrAB–TolC or MexAB–OprM, can increase the chloramphenicol resistance of P. aeruginosa (Lee et al., 2000). Seven different tetracycline resistance genes have been found in this study, of which otrA, tetB(P), tetM, and tetT could code ribosomal protection proteins and tet42, tetA, and tetG are tetracycline efflux pump genes (Thaker et al., 2010). However, only two tetracycline-resistant isolates harbored the tetG gene. The other two resistant isolates harbored RND efflux pumps, such as MexCD-OprJ, which could help the isolates resistant to tetracycline (Li et al., 2015). The isolates that carried the resistance determinants were not resistant to tetracycline, especially 114-6 and 115-5, which had five and six different resistance genes, respectively. Moreover, MIC values of 83 isolates were ≥256 μg/mL for clindamycin. Gram-negative aerobic organisms are reported to be uniformly resistant to clindamycin, whereas only a few resistance genes have been detected (Ras et al., 1992).
One interesting finding was the presence of a triclosan efflux pump. The genes triA, triB, and triC, which were present in three isolates, are tightly coupled transcripts that form the TriABC protein and associate with OpmH to assemble a functional triclosan efflux pump (Mima et al., 2007). Triclosan is now used in many consumer products, including toothpastes, soaps, cosmetics, cutting boards, and mattress pads (Bhargava and Leonard, 1996; Chuanchuen et al., 2002). Triclosan contamination is raising concerns due to the high risk of it converting into toxic dioxin in aquatic environments, and attention should be paid to its usage (Yan et al., 2019).
In total, 48 isolates were MDR strains with 22 different combined groups (Supplementary Table S1). Although the isolates harbor ARGs, RND efflux pumps may play a role in resistance because they have wide antibiotic substrate spectrums (Depardieu et al., 2007). The MAR index is considered a good risk assessment tool. A MAR index value of 0.20 used to differentiate between low and high risk of contamination (Krumperman, 1983; Riaz et al., 2011). The MAR index values of the 86 isolates tested in this study were between 0.0 and 0.8 with 59.3% (51/86) of isolates having MAR indices >0.20, suggesting high antibiotic use and high selective pressure. Swetha et al. (2017) found that 19 P. aeruginosa isolates from raw milk samples had MAR index values ranging from 0.33 to 1.0 and were resistant to between four and 12 antibiotics. Pseudomonas isolates from wastewater treatment facilities in South Africa had MAR index values ranging from 0.26 to 0.58 and were resistant to between five and 11 antibiotics (Odjadjare et al., 2012). Moreover, some Pseudomonas isolates acquire resistance to antibiotics through horizontal gene transfer (HGT) of plasmids carrying genetic material encoding for antibiotic resistance. We found 16 resistance genes from E. coli, S. aureus, and M. tuberculosis, indicating that HGT played a key role in the development of these Pseudomonas strains. HGT can provide genes necessary for survival more quickly than spontaneous mutations (Charpentier et al., 2012). Although antibiotic resistance genes are an important public health concern in clinical and veterinary environments, it is unclear whether resistance genes of pathogenic microorganisms are transmitted from environmental bacteria or elsewhere (Forsberg et al., 2012). Therefore, HGT between Pseudomonas isolates and other bacteria should be further investigated.
Conclusion
Knowledge of the antibiotic resistance profiles of food-related bacteria, including Pseudomonas spp., is currently needed. In this study, we successfully characterized the antibiotic susceptibility profiles of Pseudomonas spp. isolates from raw milk and assessed their antibiotic resistance determinants using whole genome sequencing. Our results indicate that MDR Pseudomonas species were prevalent in raw milk in Shaanxi province, China. Moreover, whole genome sequencing revealed the presence of different resistance determinants in all of the isolates as well as acquired resistance genes from other bacteria, demonstrating that HGT occurred between these Pseudomonas isolates and other bacteria. Although this study was conducted in only one region, the emergence of MDR Pseudomonas species and HGT is an important public health issue. Therefore, our data support the need for further investigations into Pseudomonas species to prevent the spread of resistance to other pathogenic bacteria.
Data Availability Statement
The datasets generated for this study can be found in the NCBI Sequence Read Archive database (accession numbers PRJNA523883, PRJNA523885, and PRJNA593738).
Author Contributions
LM and HL contributed equally, designed and performed the research. TL, LD, and HH helped with antimicrobial susceptibility testing. SZ and YZ helped with the data analysis. NZ gave advices to the researchers. JW gave opinions on the research design.
Funding
This research was supported by the Special Fund for Project of Risk Assessment on Raw Milk (GJFP2019027), the Agricultural Science and Technology Innovation Program (ASTIP-IAS12), and the Modern Agro-Industry Technology Research System of China (CARS-36).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.01005/full#supplementary-material
Footnotes
References
Acar, J. F. (1985). Problems and changing patterns of resistance with gram-negative bacteria. Rev. Infect. Dis. 7(Suppl. 4), S545–S551.
Alonso, C. A., Kwabugge, Y. A., Anyanwu, M. U., Torres, C., and Chah, K. F. (2017). Diversity of Ochrobactrum species in food animals, antibiotic resistance phenotypes and polymorphisms in the blaOCH gene. FEMS Microbiol. Lett. 364:fnx178. doi: 10.1093/femsle/fnx178
Aoike, N., Saga, T., Sakata, R., Yoshizumi, A., Kimura, S., Iwata, M., et al. (2013). Molecular characterization of extraintestinal Escherichia coli isolates in Japan: relationship between sequence types and mutation patterns of quinolone resistance-determining regions analyzed by pyrosequencing. J. Clin. Microbiol. 51, 1692–1698. doi: 10.1128/JCM.03049-12
Arcangioli, M. A., Leroy-Setrin, S., Martel, J. L., and Chaslus-Dancla, E. (1999). A new chloramphenicol and florfenicol resistance gene flanked by two integron structures in Salmonella typhimurium DT104. FEMS Microbiol. Lett. 174, 327–332. doi: 10.1111/j.1574-6968.1999.tb13586.x
Arias, C. A., Panesso, D., Mcgrath, D. M., Qin, X., Mojica, M. F., Miller, C., et al. (2011). Genetic basis for in vivo daptomycin resistance in enterococci. N. Engl. J. Med. 365, 892–900. doi: 10.1056/NEJMoa1011138
Arslan, S., Eyi, A., and Ozdemir, F. (2011). Spoilage potentials and antimicrobial resistance of Pseudomonas spp. isolated from cheeses. J. Dairy Sci. 94, 5851–5856. doi: 10.3168/jds.2011-4676
Barrow, K., and Kwon, D. H. (2009). Alterations in two-component regulatory systems of phoPQ and pmrAB are associated with polymyxin B resistance in clinical isolates of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 53, 5150–5154. doi: 10.1128/AAC.00893-09
Beena, A. K., Ranjini, A. R., and Riya, T. G. (2011). Isolation of psychrotrophic multiple drug resistant Pseudomonas from pasteurised milk. Vet. World 4, 349–352.
Bengoechea, J. A., and Skurnik, M. (2000). Temperature-regulated efflux pump/potassium antiporter system mediates resistance to cationic antimicrobial peptides in Yersinia. Mol. Microbiol. 37, 67–80. doi: 10.1046/j.1365-2958.2000.01956.x
Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., et al. (2015). Tackling antibiotic resistance: the environmental framework. Nat. Rev. Microbiol. 13, 310–317. doi: 10.1038/nrmicro3439
Bhargava, H. N., and Leonard, P. A. (1996). Triclosan: applications and safety. Am. J. Infect. Control 24, 209–218. doi: 10.1016/s0196-6553(96)90017-6
Blasco, M. D., Esteve, C., and Alcaide, E. (2008). Multiresistant waterborne pathogens isolated from water reservoirs and cooling systems. J. Appl. Microbiol. 105, 469–475. doi: 10.1111/j.1365-2672.2008.03765.x
Brown, P. D., and Izundu, A. (2004). Antibiotic resistance in clinical isolates of Pseudomonas aeruginosa in Jamaica. Rev. Panam. Salud Publica 16, 125–130.
Carfi, A., Pares, S., Duee, E., Galleni, M., Duez, C., Frere, J. M., et al. (1995). The 3-D structure of a zinc metallo-beta-lactamase from Bacillus cereus reveals a new type of protein fold. EMBO J. 14, 4914–4921. doi: 10.1002/j.1460-2075.1995.tb00174.x
Chakravarty, S., and Gregory, G. (2015). “The genus Pseudomonas,” in Practical Handbook of Microbiology, eds E. Goldman and L. H. Green (New York, NY: CRC Press), 321–344.
Charpentier, X., Polard, P., and Claverys, J. P. (2012). Induction of competence for genetic transformation by antibiotics: convergent evolution of stress responses in distant bacterial species lacking SOS? Curr. Opin. Microbiol. 15, 570–576. doi: 10.1016/j.mib.2012.08.001
Chen, H., Chen, R., Jing, L., Bai, X., and Teng, Y. (2019). A metagenomic analysis framework for characterization of antibiotic resistomes in river environment: application to an urban river in Beijing. Environ. Pollut. 245, 398–407. doi: 10.1016/j.envpol.2018.11.024
Chen, L., and Duan, K. (2016). A PhoPQ-regulated ABC transporter system exports tetracycline in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 60, 3016–3024. doi: 10.1128/AAC.02986-15
Chuanchuen, R., Narasaki, C. T., and Schweizer, H. P. (2002). The MexJK efflux pump of Pseudomonas aeruginosa requires OprM for antibiotic efflux but not for efflux of triclosan. J. Bacteriol. 184, 5036–5044. doi: 10.1128/jb.184.18.5036-5044.2002
Clinical and Laboratory Standards Institute [CLSI] (2016). Performance Standards for Antimicrobial Susceptibility Testing; 26th Informational Supplement. CLSI Document M100-S26. Wayne, PA: Clinical and Laboratory Standards Institute.
Clinical and Laboratory Standards Institute [CLSI] (2018). Performance Standards for Antimicrobial Susceptibility Testing; 28th Informational Supplement. CLSI Document M100-S28. Wayne, PA: Clinical and Laboratory Standards Institute.
Decimo, M., Silvetti, T., and Brasca, M. (2016). Antibiotic resistance patterns of Gram-negative psychrotrophic bacteria from bulk tank milk. J. Food Sci. 81, M944–M951. doi: 10.1111/1750-3841.13250
Depardieu, F., Podglajen, I., Leclercq, R., Collatz, E., and Courvalin, P. (2007). Modes and modulations of antibiotic resistance gene expression. Clin. Microbiol. Rev. 20, 79–114. doi: 10.1128/cmr.00015-06
Devarajan, N., Laffite, A., Mulaji, C. K., Otamonga, J. P., Mpiana, P. T., Mubedi, J. I., et al. (2016). Occurrence of antibiotic resistance genes and bacterial markers in a tropical river receiving hospital and urban wastewaters. PLoS One 11:e0149211. doi: 10.1371/journal.pone.0149211
Di Guilmi, A. M., Dessen, A., Dideberg, O., and Vernet, T. (2003). Functional characterization of penicillin-binding protein 1b from Streptococcus pneumoniae. J. Bacteriol. 185, 1650–1658. doi: 10.1128/jb.185.5.1650-1658.2003
El Amin, N., Giske, C. G., Jalal, S., Keijser, B., Kronvall, G., and Wretlind, B. (2005). Carbapenem resistance mechanisms in Pseudomonas aeruginosa: alterations of porin OprD and efflux proteins do not fully explain resistance patterns observed in clinical isolates. APMIS 113, 187–196. doi: 10.1111/j.1600-0463.2005.apm1130306.x
European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2018). Breakpoints Tables for Interpretation of MICs and Zone Diameter, Version 8.0. 2018. Available online at: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_8.0_Breakpoint_Tables.pdf
Fang, Z. L., Zhang, L. Y., Huang, Y. M., Qing, Y., Cao, K. Y., Tian, G. B., et al. (2014). OprD mutations and inactivation in imipenem-resistant Pseudomonas aeruginosa isolates from China. Infect. Genet. Evol. 21, 124–128. doi: 10.1016/j.meegid.2013.10.027
Flores Ribeiro, A., Bodilis, J., Alonso, L., Buquet, S., Feuilloley, M., Dupont, J. P., et al. (2014). Occurrence of multi-antibiotic resistant Pseudomonas spp. in drinking water produced from karstic hydrosystems. Sci. Total Environ. 490, 370–378. doi: 10.1016/j.scitotenv.2014.05.012
Forsberg, K. J., Reyes, A., Wang, B., Selleck, E. M., Sommer, M. O., and Dantas, G. (2012). The shared antibiotic resistome of soil bacteria and human pathogens. Science 337, 1107–1111. doi: 10.1126/science.1220761
Fricker, M., Skanseng, B., Rudi, K., Stessl, B., and Ehling-Schulz, M. (2011). Shift from farm to dairy tank milk microbiota revealed by a polyphasic approach is independent from geographical origin. Int. J. Food Microbiol. 145(Suppl. 1), S24–S30. doi: 10.1016/j.ijfoodmicro.2010.08.025
Gatzeva-Topalova, P. Z., May, A. P., and Sousa, M. C. (2005). Structure and mechanism of ArnA: conformational change implies ordered dehydrogenase mechanism in key enzyme for polymyxin resistance. Structure 13, 929–942. doi: 10.1016/j.str.2005.03.018
Gomi, R., Matsuda, T., Matsumura, Y., Yamamoto, M., Tanaka, M., Ichiyama, S., et al. (2017). Whole-genome analysis of antimicrobial-resistant and extraintestinal pathogenic Escherichia coli in river water. Appl. Environ. Microbiol. 83:e02703-16. doi: 10.1128/AEM.02703-16
Gupta, S. K., Padmanabhan, B. R., Diene, S. M., Lopez-Rojas, R., Kempf, M., Landraud, L., et al. (2014). ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 58, 212–220. doi: 10.1128/AAC.01310-13
Jeukens, J., Freschi, L., Kukavica-Ibrulj, I., Emond-Rheault, J. G., Tucker, N. P., and Levesque, R. C. (2017). Genomics of antibiotic-resistance prediction in Pseudomonas aeruginosa. Ann. N. Y. Acad. Sci. 1435, 5–17. doi: 10.1111/nyas.13358
Jones, K. E., Patel, N. G., Levy, M. A., Storeygard, A., Balk, D., Gittleman, J. L., et al. (2008). Global trends in emerging infectious diseases. Nature 451, 990–993. doi: 10.1038/nature06536
Kittinger, C., Lipp, M., Baumert, R., Folli, B., Koraimann, G., Toplitsch, D., et al. (2016). Antibiotic resistance patterns of Pseudomonas spp. isolated from the river Danube. Front. Microbiol. 7:586. doi: 10.3389/fmicb.2016.00586
Kohler, T., Kok, M., Michea-Hamzehpour, M., Plesiat, P., Gotoh, N., Nishino, T., et al. (1996). Multidrug efflux in intrinsic resistance to trimethoprim and sulfamethoxazole in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 40, 2288–2290. doi: 10.1128/aac.40.10.2288
Krumperman, P. H. (1983). Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 46, 165–170. doi: 10.1128/aem.46.1.165-170.1983
Lee, A., Mao, W., Warren, M. S., Mistry, A., Hoshino, K., Okumura, R., et al. (2000). Interplay between efflux pumps may provide either additive or multiplicative effects on drug resistance. J. Bacteriol. 182, 3142–3150. doi: 10.1128/jb.182.11.3142-3150.2000
Lee, J. Y., and Ko, K. S. (2014). Mutations and expression of PmrAB and PhoPQ related with colistin resistance in Pseudomonas aeruginosa clinical isolates. Diagn. Microbiol. Infect. Dis. 78, 271–276. doi: 10.1016/j.diagmicrobio.2013.11.027
Li, X. Z., Plesiat, P., and Nikaido, H. (2015). The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin. Microbiol. Rev. 28, 337–418. doi: 10.1128/CMR.00117-14
Liu, H. M., Li, S. L., Meng, L., Dong, L., Zhao, S. G., Lan, X. Y., et al. (2017). Prevalence, antimicrobial susceptibility, and molecular characterization of Staphylococcus aureus isolated from dairy herds in northern China. J. Dairy Sci. 100, 8796–8803. doi: 10.3168/jds.2017-13370
Livermore, D. M. (2001). Of Pseudomonas, porins, pumps and carbapenems. J. Antimicrob. Chemother. 47, 247–250. doi: 10.1093/jac/47.3.247
Luo, R. B., Liu, B. H., Xie, Y. L., Li, Z. Y., Huang, W. H., Yuan, J. Y., et al. (2015). SOAPdenovo2: an empirically improved memory-efficient short-read de novo assembler. Gigascience 1:18.
Marceau, M., Sebbane, F., Ewann, F., Collyn, F., Lindner, B., Campos, M. A., et al. (2004). The pmrF polymyxin-resistance operon of Yersinia pseudotuberculosis is upregulated by the PhoP-PhoQ two-component system but not by PmrA-PmrB, and is not required for virulence. Microbiology 150, 3947–3957. doi: 10.1099/mic.0.27426-0
Marchand, S., Heylen, K., Messens, W., Coudijzer, K., De Vos, P., Dewettinck, K., et al. (2009). Seasonal influence on heat-resistant proteolytic capacity of Pseudomonas lundensis and Pseudomonas fragi, predominant milk spoilers isolated from Belgian raw milk samples. Environ. Microbiol. 11, 467–482. doi: 10.1111/j.1462-2920.2008.01785.x
Mathur, S., and Singh, R. (2005). Antibiotic resistance in food lactic acid bacteria–a review. Int. J. Food Microbiol. 105, 281–295. doi: 10.1016/j.ijfoodmicro.2005.03.008
McPhee, J. B., Lewenza, S., and Hancock, R. E. (2003). Cationic antimicrobial peptides activate a two-component regulatory system, PmrA-PmrB, that regulates resistance to polymyxin B and cationic antimicrobial peptides in Pseudomonas aeruginosa. Mol. Microbiol. 50, 205–217. doi: 10.1046/j.1365-2958.2003.03673.x
Meng, L., Zhang, Y., Liu, H., Zhao, S., Wang, J., and Zheng, N. (2017). Characterization of Pseudomonas spp. and associated proteolytic properties in raw milk stored at low temperatures. Front. Microbiol. 8:2158. doi: 10.3389/fmicb.2017.02158
Milheirico, C., De Lencastre, H., and Tomasz, A. (2017). Full-genome sequencing identifies in the genetic background several determinants that modulate the resistance phenotype in methicillin-resistant Staphylococcus aureus strains carrying the novel mecC gene. Antimicrob. Agents Chemother. 61:e02500-16. doi: 10.1128/AAC.02500-16
Miller, A. K., Brannon, M. K., Stevens, L., Johansen, H. K., Selgrade, S. E., Miller, S. I., et al. (2011). PhoQ mutations promote lipid A modification and polymyxin resistance of Pseudomonas aeruginosa found in colistin-treated cystic fibrosis patients. Antimicrob. Agents Chemother. 55, 5761–5769. doi: 10.1128/AAC.05391-11
Miller, C., Kong, J., Tran, T. T., Arias, C. A., Saxer, G., and Shamoo, Y. (2013). Adaptation of Enterococcus faecalis to daptomycin reveals an ordered progression to resistance. Antimicrob. Agents Chemother. 57, 5373–5383. doi: 10.1128/AAC.01473-13
Mima, T., Joshi, S., Gomez-Escalada, M., and Schweizer, H. P. (2007). Identification and characterization of TriABC-OpmH, a triclosan efflux pump of Pseudomonas aeruginosa requiring two membrane fusion proteins. J. Bacteriol. 189, 7600–7609. doi: 10.1128/jb.00850-07
MOA (2002). “Announcement No. 235”, in: Maximum Residue Limit of Veterinary Drugs in Animal Food. Beijing: Ministry of Agriculture of the People’s Republic of China.
Molina, L., Udaondo, Z., Duque, E., Fernandez, M., Molina-Santiago, C., Roca, A., et al. (2014). Antibiotic resistance determinants in a Pseudomonas putida strain isolated from a hospital. PLoS One 9:e81604. doi: 10.1371/journal.pone.0081604
Morita, Y., Kimura, N., Mima, T., Mizushima, T., and Tsuchiya, T. (2001a). Roles of MexXY- and MexAB-multidrug efflux pumps in intrinsic multidrug resistance of Pseudomonas aeruginosa PAO1. J. Gen. Appl. Microbiol. 47, 27–32. doi: 10.2323/jgam.47.27
Morita, Y., Komori, Y., Mima, T., Kuroda, T., Mizushima, T., and Tsuchiya, T. (2001b). Construction of a series of mutants lacking all of the four major mex operons for multidrug efflux pumps or possessing each one of the operons from Pseudomonas aeruginosa PAO1: MexCD-OprJ is an inducible pump. FEMS Microbiol. Lett. 202, 139–143. doi: 10.1111/j.1574-6968.2001.tb10794.x
Munsch-Alatossava, P., and Alatossava, T. (2006). Phenotypic characterization of raw milk-associated psychrotrophic bacteria. Microbiol. Res. 161, 334–346. doi: 10.1016/j.micres.2005.12.004
Munsch-Alatossava, P., and Alatossava, T. (2007). Antibiotic resistance of raw-milk-associated psychrotrophic bacteria. Microbiol. Res. 162, 115–123. doi: 10.1016/j.micres.2006.01.015
Munsch-Alatossava, P., Gauchi, J. P., Chamlagain, B., and Alatossava, T. (2012). Trends of antibiotic resistance in mesophilic and psychrotrophic bacterial populations during cold storage of raw milk. ISRN Microbiol. 2012:918208. doi: 10.5402/2012/918208
Neu, H. C. (1983). The emergence of bacterial resistance and its influence on empiric therapy. Rev. Infect. Dis. 5(Suppl. 1), S9–S20.
Odjadjare, E. E., Igbinosa, E. O., Mordi, R., Igere, B., Igeleke, C. L., and Okoh, A. I. (2012). Prevalence of multiple antibiotics resistant (MAR) Pseudomonas species in the final effluents of three municipal wastewater treatment facilities in South Africa. Int. J. Environ. Res. Public Health 9, 2092–2107. doi: 10.3390/ijerph9062092
Palma, M., Zurita, J., Ferreras, J. A., Worgall, S., Larone, D. H., Shi, L., et al. (2005). Pseudomonas aeruginosa SoxR does not conform to the archetypal paradigm for SoxR-dependent regulation of the bacterial oxidative stress adaptive response. Infect. Immun. 73, 2958. doi: 10.1128/iai.73.5.2958-2966.2005
Panelists, I. E. R. (2006). Antimicrobial resistance: implications for the food system. Compr. Rev. Food Sci. Food Saf. 5, 71–137. doi: 10.1111/j.1541-4337.2006.00004.x
Peleg, A. Y., Miyakis, S., Ward, D. V., Earl, A. M., Rubio, A., Cameron, D. R., et al. (2012). Whole genome characterization of the mechanisms of daptomycin resistance in clinical and laboratory derived isolates of Staphylococcus aureus. PLoS One 7:e28316. doi: 10.1371/journal.pone.0028316
Piddock, L. J. V. (2006). Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 19, 382–402. doi: 10.1128/cmr.19.2.382-402.2006
Poole, K., Gotoh, N., Tsujimoto, H., Zhao, Q., Wada, A., Yamasaki, T., et al. (1996). Overexpression of the mexC-mexD-oprJ efflux operon in nfxB-type multidrug-resistant strains of Pseudomonas aeruginosa. Mol. Microbiol. 21, 713–724.
Porres-Osante, N., Estepa, V., Seral, C., Rojo-Bezares, B., Salvo, S., Algarate, S., et al. (2014). First description of a blaVIM-2-Carrying Citrobacter freundii isolate in Spain. Antimicrob. Agents Chemother. 58, 6331–6332. doi: 10.1128/aac.03168-14
Quigley, L., O’sullivan, O., Stanton, C., Beresford, T. P., Ross, R. P., Fitzgerald, G. F., et al. (2013). The complex microbiota of raw milk. FEMS Microbiol. Rev. 37, 664–698. doi: 10.1111/1574-6976.12030
Rajwar, A., and Sahgal, M. (2016). Phylogenetic relationships of fluorescent pseudomonads deduced from the sequence analysis of 16S rRNA, Pseudomonas-specific and rpoD genes. 3 Biotech 6:80. doi: 10.1007/s13205-016-0386-x
Ramirez, M. S., and Tolmasky, M. E. (2010). Aminoglycoside modifying enzymes. Drug Resist Updat 13, 151–171. doi: 10.1016/j.drup.2010.08.003
Ras, G. J., Anderson, R., Taylor, G. W., Savage, J. E., Van Niekerk, E., Joone, G., et al. (1992). Clindamycin, erythromycin, and roxithromycin inhibit the proinflammatory interactions of Pseudomonas aeruginosa pigments with human neutrophils in vitro. Antimicrob. Agents Chemother. 36, 1236–1240. doi: 10.1128/aac.36.6.1236
Riaz, S., Faisal, M., and Hasnain, S. (2011). Antibiotic susceptibility pattern and multiple antibiotic resistances (MAR) calculation of extended spectrum β- lactamase (ESBL) producing Escherichia coli and Klebsiella species in Pakistan. Afr. J. Biotechnol. 10, 6325–6331.
Rodriguez-Martinez, J. M., Poirel, L., and Nordmann, P. (2009). Extended-spectrum cephalosporinases in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 53, 1766–1771. doi: 10.1128/AAC.01410-08
Saavedra, M. J., Peixe, L., Sousa, J. C., Henriques, I., Alves, A., and Correia, A. (2003). Sfh-I, a subclass B2 metallo-beta-lactamase from a Serratia fonticola environmental isolate. Antimicrob. Agents Chemother. 47, 2330–2333. doi: 10.1128/aac.47.7.2330-2333.2003
Schwarz, S., Kehrenberg, C., Doublet, B., and Cloeckaert, A. (2004). Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 28, 519–542. doi: 10.1016/j.femsre.2004.04.001
Shaaly, A., Kalamorz, F., Gebhard, S., and Cook, G. M. (2013). Undecaprenyl pyrophosphate phosphatase confers low-level resistance to bacitracin in Enterococcus faecalis. J. Antimicrob. Chemother. 68, 1583–1593. doi: 10.1093/jac/dkt048
Silveira, M. C., Catanho, M., and Miranda, A. B. (2018). Genomic analysis of bifunctional Class C-Class D beta-lactamases in environmental bacteria. Mem. Inst. Oswaldo Cruz 113:e180098. doi: 10.1590/0074-02760180098
Sobel, M. L., Neshat, S., and Poole, K. (2005). Mutations in PA2491 (mexS) promote MexT-dependent mexEF-oprN expression and multidrug resistance in a clinical strain of Pseudomonas aeruginosa. J. Bacteriol. 187, 1246–1253. doi: 10.1128/jb.187.4.1246-1253.2005
Spratt, B. G., and Cromie, K. D. (1988). Penicillin-binding proteins of gram-negative bacteria. Rev. Infect. Dis. 10, 699–711. doi: 10.1093/clinids/10.4.699
Sreevatsan, S., Pan, X., Stockbauer, K. E., Williams, D. L., Kreiswirth, B. N., and Musser, J. M. (1996). Characterization of rpsL and rrs mutations in streptomycin-resistant Mycobacterium tuberculosis isolates from diverse geographic localities. Antimicrob. Agents Chemother. 40, 1024–1026. doi: 10.1128/aac.40.4.1024
Stanhope, M. J., Lefebure, T., Walsh, S. L., Becker, J. A., Lang, P., Pavinski Bitar, P. D., et al. (2008). Positive selection in penicillin-binding proteins 1a, 2b, and 2x from Streptococcus pneumoniae and its correlation with amoxicillin resistance development. Infect. Genet. Evol. 8, 331–339. doi: 10.1016/j.meegid.2008.02.001
Steiniger-White, M., Rayment, I., and Reznikoff, W. S. (2004). Structure/function insights into Tn5 transposition. Curr. Opin. Struct. Biol. 14, 50–57. doi: 10.1016/j.sbi.2004.01.008
Straley, B. A., Donaldson, S. C., Hedge, N. V., Sawant, A. A., Srinivasan, V., Oliver, S. P., et al. (2006). Public health significance of antimicrobial-resistant gram-negative bacteria in raw bulk tank milk. Foodborne Pathog. Dis. 3, 222–233. doi: 10.1089/fpd.2006.3.222
Swetha, C. S., Babu, A. J., Rao, K. V., Bharathy, S., Supriya, R. A., and Rao, T. M. (2017). A study on the antimicrobial resistant patterns of Pseudomonas aeruginosa isolated from raw milk samples in and around Tirupati, Andhra Pradesh. Asian J. Dairy Food Res. 36, 100–105.
Thaker, M., Spanogiannopoulos, P., and Wright, G. D. (2010). The tetracycline resistome. Cell. Mol. Life Sci. 67, 419–431. doi: 10.1007/s00018-009-0172-6
Toleman, M. A., Bennett, P. M., Bennett, D. M., Jones, R. N., and Walsh, T. R. (2007). Global emergence of trimethoprim/sulfamethoxazole resistance in Stenotrophomonas maltophilia mediated by acquisition of sul genes. Emerg. Infect. Dis. 13, 559–565.
Unemo, M., Golparian, D., Nicholas, R., Ohnishi, M., Gallay, A., and Sednaoui, P. (2012). High-level cefixime- and ceftriaxone-resistant Neisseria gonorrhoeae in France: novel penA mosaic allele in a successful international clone causes treatment failure. Antimicrob. Agents Chemother. 56, 1273–1280. doi: 10.1128/AAC.05760-11
United States Department of Agriculture, Animal Plant Health Inspection Service National Animal Health Monitoring System [USDA APHIS] (2008). Antibiotic Use on U.S. Dairy Operations, 2002 and 2007 (Info Sheet, 5p, October, 2008). Available online at: www.aphis.usda.gov/animal_health/nahms/dairy/downloads/dairy07/Dairy07_is_AntibioticUse_1.pdf (accessed August 10, 2017).
Von Neubeck, M., Baur, C., Krewinkel, M., Stoeckel, M., Kranz, B., Stressler, T., et al. (2015). Biodiversity of refrigerated raw milk microbiota and their enzymatic spoilage potential. Int. J. Food Microbiol. 211, 57–65. doi: 10.1016/j.ijfoodmicro.2015.07.001
Wang, P., Hu, F. P., Xiong, Z. Z., Ye, X. Y., Zhu, D. M., Wang, Y. F., et al. (2011). Susceptibility of extended-spectrum-β-Lactamase-producing Enterobacteriaceae according to the new CLSI breakpoints. J. Clin. Microbiol. 49, 3127–3131. doi: 10.1128/JCM.01692-14
Wolter, D. J., and Lister, P. D. (2013). Mechanisms of beta-lactam resistance among Pseudomonas aeruginosa. Curr. Pharm. Des. 19, 209–222. doi: 10.2174/1381612811306020209
Yan, Z. R., Meng, H. S., Yang, X. Y., Zhu, Y. Y., Li, X. Y., Xu, J., et al. (2019). Insights into the interactions between triclosan (TCS) and extracellular polymeric substance (EPS) of activated sludge. J. Environ. Manage. 232, 219–225. doi: 10.1016/j.jenvman.2018.11.059
Yang, S. J., Nast, C. C., Mishra, N. N., Yeaman, M. R., Fey, P. D., and Bayer, A. S. (2010). Cell wall thickening is not a universal accompaniment of the daptomycin nonsusceptibility phenotype in Staphylococcus aureus: evidence for multiple resistance mechanisms. Antimicrob. Agents Chemother. 54, 3079–3085. doi: 10.1128/AAC.00122-10
Yomoda, S., Okubo, T., Takahashi, A., Murakami, M., and Iyobe, S. (2003). Presence of Pseudomonas putida strains harboring plasmids bearing the metallo-beta-lactamase gene bla(IMP) in a hospital in Japan. J. Clin. Microbiol. 41, 4246–4251. doi: 10.1128/jcm.41.9.4246-4251.2003
Zankari, E., Hasman, H., Cosentino, S., Vestergaard, M., Rasmussen, S., Lund, O., et al. (2012). Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 67, 2640–2644. doi: 10.1093/jac/dks261
Zhang, W. L., Aurosree, B., Gopalakrishnan, B., Balada-Llasat, J., Pancholi, V., and Pancholi, P. (2017). The role of LpxA/C/D and pmrA/B gene systems in colistin-resistant clinical strains of Acinetobacter baumannii. Front. Lab. Med. 1, 86–91. doi: 10.1016/j.flm.2017.07.001
Keywords: Pseudomonas spp., antibiotic resistance, whole genome sequencing, milk, multiple antibiotic resistance index
Citation: Meng L, Liu H, Lan T, Dong L, Hu H, Zhao S, Zhang Y, Zheng N and Wang J (2020) Antibiotic Resistance Patterns of Pseudomonas spp. Isolated From Raw Milk Revealed by Whole Genome Sequencing. Front. Microbiol. 11:1005. doi: 10.3389/fmicb.2020.01005
Received: 09 December 2019; Accepted: 24 April 2020;
Published: 03 June 2020.
Edited by:
Zhenbo Xu, University of Maryland, Baltimore, United StatesReviewed by:
Sanath H. Kumar, Central Institute of Fisheries Education (ICAR), IndiaCatherine Llanes, University of Franche-Comté, France
Cemal Sandalli, Recep Tayyip Erdoğan University, Turkey
Copyright © 2020 Meng, Liu, Lan, Dong, Hu, Zhao, Zhang, Zheng and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiaqi Wang, amlhcWl3YW5nQHZpcC4xNjMuY29t
†These authors have contributed equally to this work and share first authorship