 Eveline M. Ibeagha-Awemu
Eveline M. Ibeagha-Awemu Faith A. Omonijo
Faith A. Omonijo Laurie C. Piché
Laurie C. Piché Antony T. Vincent
Antony T. Vincent- 1Sherbrooke Research and Development Centre, Agriculture and Agri-Food Canada, Sherbrooke, QC, Canada
- 2Département des sciences animales, Université Laval, Québec, QC, Canada
The discovery of the growth promoting effects of antibiotics in the 1940s contributed to the economic efficacy of the livestock industry. In response to increased animal protein demand from the 1950s, antimicrobial use at sub-therapeutic levels for growth promotion and disease prevention (antimicrobial growth promoter, AGP) improved feed-to-weight ratio, meat quality and overall health of livestock. These benefits encouraged the heavy use of AGPs such that about 70% of global use of antimicrobials was for food animals. Despite the numerous benefits of AGPs, the emergence of antimicrobial resistance (AMR) associated with their use and impact on human and livestock health, establishes AMR as a global health plague, affecting man, animal and the environment. Although many countries have banned the use of AGPs in livestock production, efforts to identify effective alternatives have yielded inconsistent findings and only few effective alternative products are currently available. This highlights the need to intensify efforts toward identifying more effective AGP alternatives. While current strategies focus on evaluating the efficacy of single products/class of products that can enhance productivity and health, future strategies should focus on combining multiple approaches. It is also important to acknowledge that no single alternative can fully replicate the same mechanism of action attributed to antimicrobials. This comprehensive review presents recent research findings on AGP use trend before and after bans in many countries, the benefit/mode of action of reported AGP alternatives, the economic impact of AGP alternatives in the context of the One Health approach, the factors militating the search for effective AGP alternatives, research gaps and future action plans for AGP-free animal farm management.
1 Introduction
Since the discovery of antibiotics in the 1920s and the subsequent recognition of their growth-promoting effects in the 1940s (1), their use has played a pivotal role in enhancing livestock productivity and in improving the economic efficiency of animal production systems. In response to increased animal protein demand from the 1950s, antimicrobials have been used as feed supplements at sub-therapeutic doses to promote growth by enhancing feed efficiency and preventing disease occurrence in livestock production (2, 3), which ultimately contributed to the intensification of livestock production. Generally, antimicrobials are used in livestock production to (1) treat infected animals (disease treatment or therapy); (2) prevent disease occurrence even in animals that show no visible signs of illness (prophylactic use); (3) prevent the spread of disease to healthy animals in herds where infected animals are present (metaphylactic use or disease control use); and (4) promote faster growth or more efficient livestock growth (use for production purpose which may coincidentally prevent disease occurrence). Since the first use of antimicrobials as feed supplement for the purpose of promoting growth, coupled with advances in genetics, nutrition, management practices, and biosecurity measures, there has been enhanced growth of pigs and poultry, improved feed-to-weight ratio in livestock, improved feed efficiency, meat quality and overall animal health (4–6). These benefits encouraged the heavy non-therapeutic use of antimicrobials or antibiotics for growth promotion and disease prevention (AGPs) such that about 80% of antimicrobial use in the United States was for food animals and over 70% global sale of antimicrobials was for food animals, which is projected to increase by 67% by 2030 in low- and middle-income countries (7–9). Moreover, it was also projected that antimicrobial use in livestock production far exceeds its use for human health purpose (8). While antimicrobials as excellent assassins quickly eliminate their targets (microbes), they also quickly lose their ability to perform effectively as microbes develop defense mechanisms in the form of antimicrobial-resistance genes (ARGs). Antimicrobial-resistance genes circulating in the food chain causes the emergence of antimicrobial resistance (AMR) impacting human, livestock and environmental health (9). Antimicrobial-resistance genes or resistant bacteria of animal origin may be transmitted to humans through several routes including the environment, food products and by direct contact with agricultural workers (8), thus establishing AMR as a global health plague, affecting both man, animal and the environment. Therefore, AMR emanating from antimicrobial use in food animals is an issue within the One Health concept (Figure 1), requiring concerted efforts to address AGP use in livestock production and AMR. Moreover, antibiotic usage in animal feed is increasingly under scrutiny resulting from consumer concerns and demand for products from animals raised without AGPs.
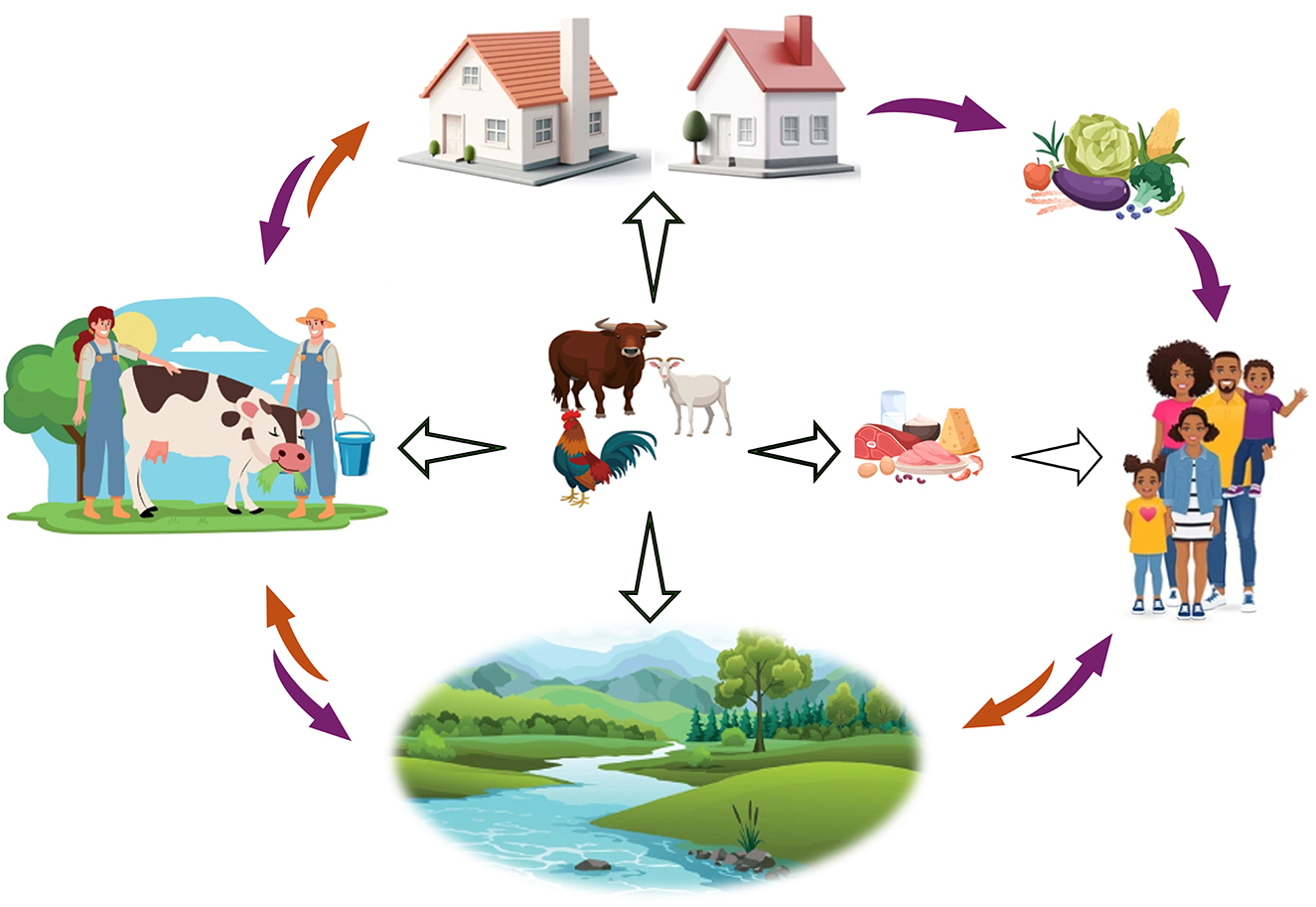
Figure 1. The role of animal farming in the One Health concept of antimicrobial resistance. Farm animals are at the center of the interactions and play a key role in the spread of antibiotic resistance genes and resistant bacteria. Antimicrobial-resistance genes or resistant bacteria of animal origin are transmitted to humans through several routes including the environment, food products and by direct contact with agricultural workers. Animals can directly contaminate fields and water through manure, farm workers, the environment, and food products derived from animals. Contaminated fields and water can also facilitate the transmission to farm workers, as well as to plant and animal products, which ultimately pose a risk to consumers. Finally, the environment can contribute to the contamination of both the farm workers and the general population.
Following the likening of AMR to a “ticking time bomb” in 2013 by Britain's Chief Medical Officer, Sally Davies, who also lobbied for the consideration of AMR as a threat of comparable magnitude to major coastal flooding or catastrophic terrorist attacks (7), there was a deluge of national action plans, expert reports, and pledges by many of the G20 nations and international organizations (World Health Organization (WHO), the Food and Agricultural Organization (FAO), the world Organization for Animal Health (OIE) and individual nations) to reduce antibiotic use (10–14). These pledges and planned actions included funding antibiotic research, tackling human overuse and commitment to reducing antibiotic use (therapeutic and AGPs) in food production. Moreover the guidelines for AMR surveillance established by these organizations allows for clear understanding of how AMR spreads across different settings and specific environments; which allows for the establishment of the relationships between antimicrobial use and AMR in different scenarios (animal, environment and human) and the outcome of interventions within and between the different sectors (11, 14–16).
As a result of the growing association of AGP use with AMR, many countries have banned/regulated the use of antimicrobials in growth promotion (Table 1) and concerted efforts have been put in place by various countries to develop alternatives to AGP in animal agriculture. An alternative to antibiotics is generally regarded as any substance that can prevent the need for antibiotic use in livestock production or be substituted for antimicrobial drugs (17, 18). Such efforts have seen reports on various AGP alternatives such as microbial related products (probiotics, engineered microbes, bacteriophages, bacteriophage-derived products, etc.), phytogenics (e.g., essential oils, phytonutrients, phytochemicals, acidifiers, trace minerals, etc.), immune-derived products (e.g., antimicrobial peptides, immunomodulators, etc.), prebiotics, vaccines, enzymes, minerals, metals and innovative animal drugs, etc. (Figure 2) and their mode of actions, which are varied and inconsistent in most cases, have been presented in several reviews (6, 19, 20). In spite of the ability of many existing alternatives to AGP to enhance animal production and health, only few effective alternatives to AGP with ability to effectively enhance productivity and prevent/control infections exist necessitating the search for more effective alternatives. This review therefore presents recent research findings on antimicrobial use trends before and after the ban of AGPs by many countries. It also discusses the benefit/mode of action of reported AGP alternatives, the economic impact of AGP alternatives in the context of the One Health approach, the factors militating the search for more effective AGP alternatives, research gaps and future action plan for AGP-free animal farm management.
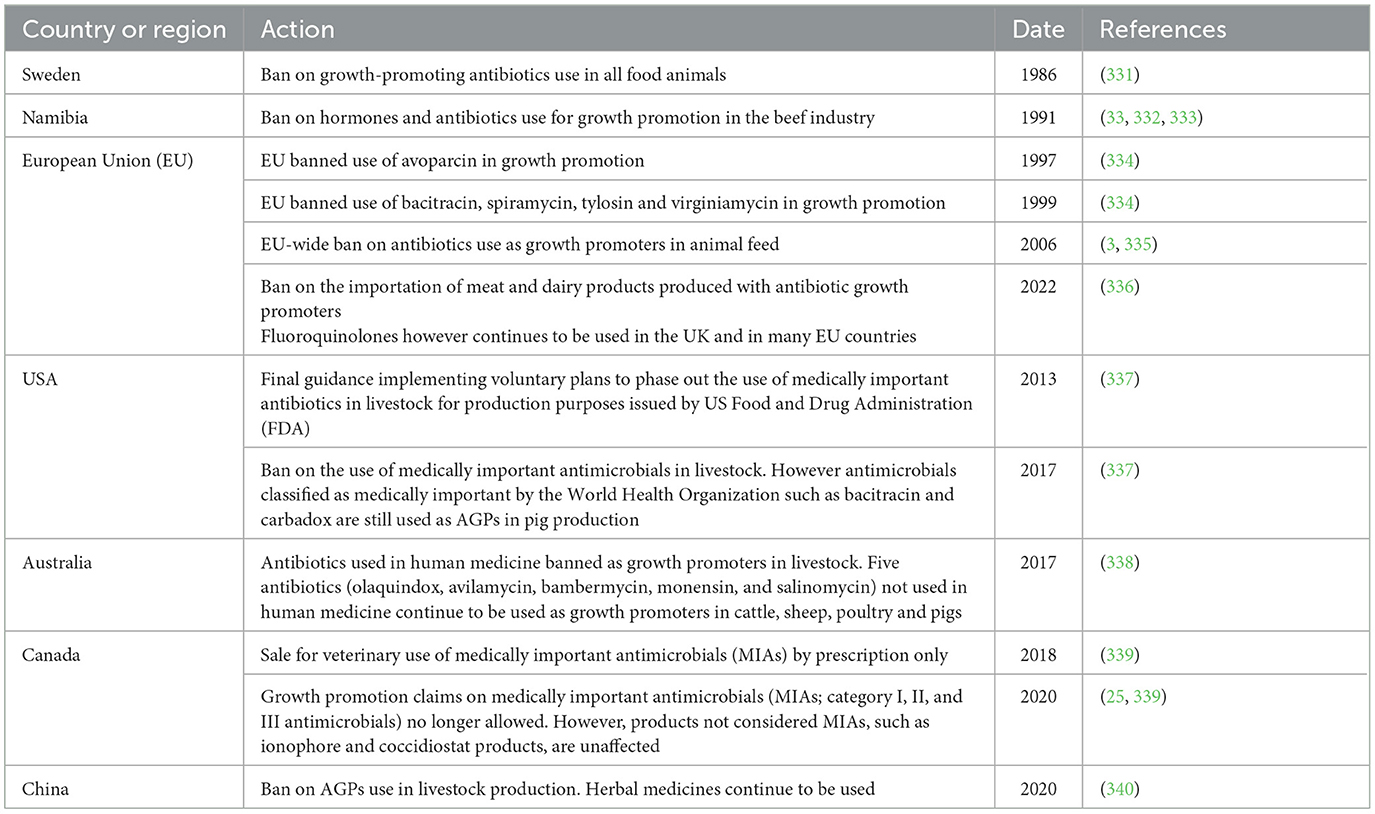
Table 1. Sample regulations and bans on the use of antibiotic growth promoters by various countries.
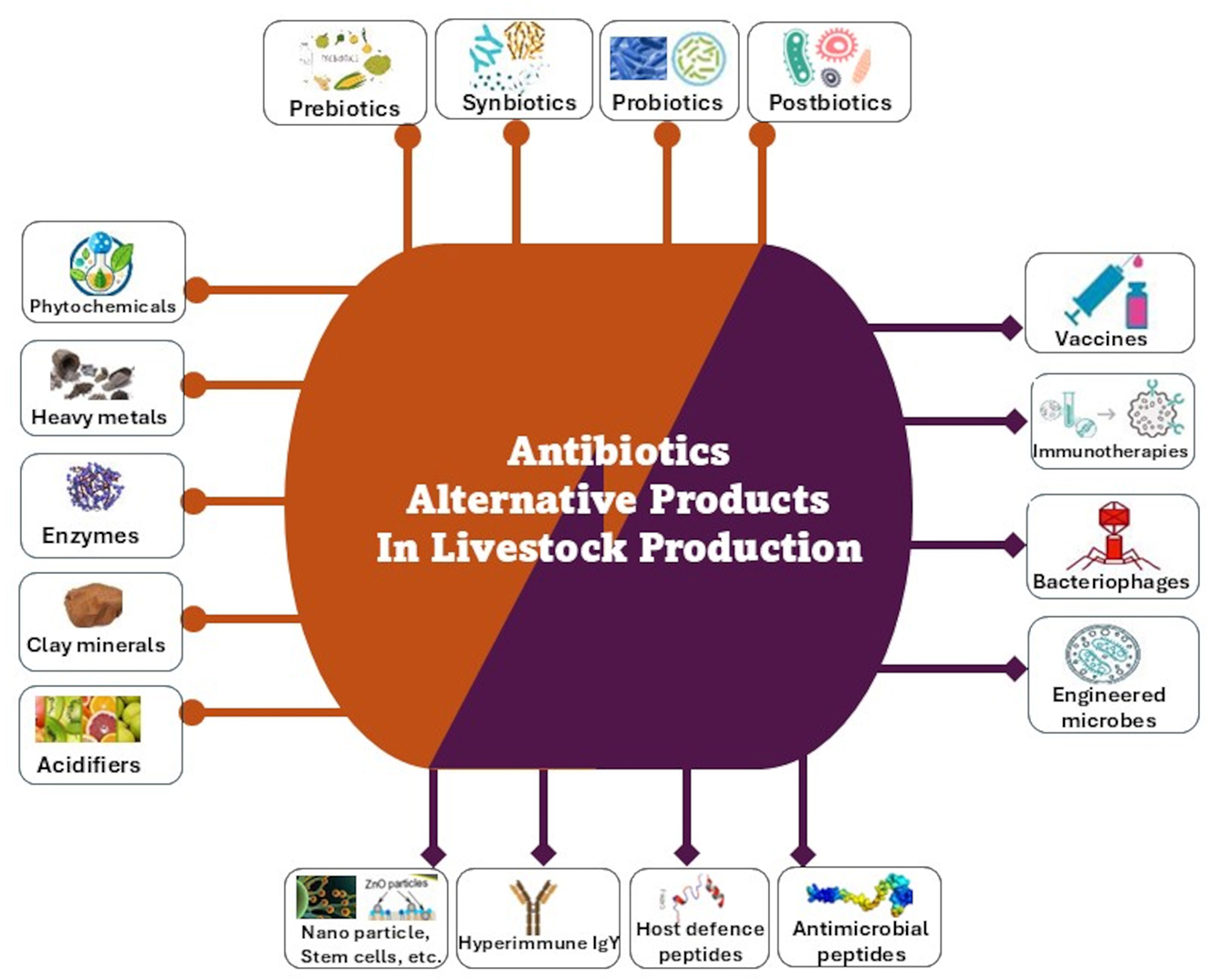
Figure 2. A summary of the health and growth-enhancer alternatives to antibiotics used in animal production or have been experimented. The brown lines point to those alternative products that are mainly administered through food/water. The purple lines points to immunotherapeutic and other microbial compounds or strategies.
2 Antibiotics use trend in livestock production before and after AGP bans by many countries
2.1 Trends in antimicrobial use in livestock production
Agricultural antibiotics use started with synthetic sulphonamides produced by Bayer (German pharmaceutical manufacturer) and marketed as Prontosil (sulfochrysoidine) in 1935, which was effective against Gram-positive bacterial infections (7). The success of Protonsil and other sulphonamides ushered in a new era of therapeutic drugs, which were marketed for livestock use as early as 1938, prompting the development of different antibiotics products by various United States and European companies for different applications in livestock. For example, gramicidin was used to treat an outbreak of mastitis infection (udder inflammation) in cows at a World Exhibition in New York in 1940 (21). The next decades will witness the growing and expanding use of antibiotics in livestock disease management, and as a production management tool, fueled by increasing demand for animal products. As early as 1948, sulfaquinoxaline (produced by Merk) was officially licensed for routine inclusion in poultry feeds, the so called “medicated feed,” to control coccidiosis. The use of medicated feeds/water helped to curb disease occurrence and spread in concentrated and mostly intensively management animal operations, and enhanced productivity by reducing labor time spent on animal care.
Interest in the non-therapeutic use of antibiotics in livestock production started with observations in 1949 that feeding antibiotic fermentation wastes as a viable alternative source of vitamin B12 feed supplement resulted in increased animal weight gain (22). Moreover, further investigations found that feeding low-dosed AGPs also prophylactically protected against bacterial infections (21, 23). These observations and perceived benefits fueled and expanded the adoption of AGPs in livestock enterprises, which quickly blurred the boundaries between growth promotion, therapy, and prophylaxis on-farm. An in-depth chronicle on the development and application of antimicrobials in livestock production to the emergence of AMR was presented recently (7).
A plethora of evidence abound on the benefits of AGP such as its positive effects on feed efficiency, meat quality, overall growth rate, reproduction performance, egg production and hatchability, etc. (4, 5). For example, it is known that pigs fed AGP require 10%−15% less feed to achieve desired growth (4) with the consequence of reduced cost of production since feed constitute a major expense in animal farming. AGPs ensure enhanced feed to animal product conversion efficiency and overall improvement. Meat from AGP supplemented livestock was of better quality and contained high amounts of proteins than fat compared to non-supplemented counterparts (24). The health of AGP supplemented animals improved while mortality decreased (4, 5). These benefits saw the explosion of the application of AGPs in livestock management for increased productivity and disease control worldwide. Moreover, a plethora of investigations have examined the antimicrobial use trends in many countries (8, 9, 25, 26), and concluded that the most important use of AGP in livestock production in the past 70 decades has been for growth promotion and disease control (prophylactic). Using information on livestock densities, antimicrobial consumption estimates in high-income countries and other variables to estimate antimicrobial use in livestock for years 2010 and 2030, it was found that the mean annual consumption of antimicrobials per kilogram of livestock produced was 148, 45, and 172 mg/kg for chicken, cattle, and pigs, respectively (8). Moreover, the same study estimated that the global consumption of antimicrobials will increase from 63,151 tons in 2010 to 105,596 tons by 2030, which represents a 67% increase (8). Using sales data in 41 countries, global sales of antimicrobials was estimated at 93 thousand tons in 2012 and projected to increase by 11.5% to 104 thousand tons by 2030 (27).
2.2 Regulation of antimicrobial use in livestock production
Although antibiotics use as feed additive provides numerous benefits to livestock health and welfare, its widespread use at sub-therapeutic levels poses significant risks to public health due to the development and dissemination of AMR. Moreover, antibiotic residues in animal products by entering the food supply chain is a major threat to human health and the environment (28, 29). These negative consequences have awakened global concerns which regards AMR as one of the greatest public health threats of today (30). To address the growing problem of AMR, many countries issued regulations and bans (Table 1) and action plans (30) aimed at regulating the use of antimicrobials in livestock production and the active search for alternative products (Figure 2). The regulations put in place by many countries have achieved some behavioral change in antimicrobial consumption in some countries. For example, following the ban on the use of medically important antibiotics in livestock production in the United States, there was a significant decrease in marketed volume of antibiotics in 2017, with decrease of 38% in sales volume in 2021 compared to peak sales in 2015, meanwhile annual sales have remained at reduced levels since 2017 (31). However, many developing countries continue unabated use of antimicrobials in livestock production (32, 33), which is not helping with global efforts to curb AMR.
2.3 Need to develop alternatives to antibiotics as growth promoters in livestock production
To reduce the over use of antimicrobials to stop the development of new resistant strains, health policy makers worldwide have prioritized tackling AMR because of its profound impact on human, animal, and environmental health (34). In 2015, the WHO published the Global Action Plan on AMR as well as the implementation of relevant strategies (10), emphasizing that elucidation of the mechanisms of AMR in pathogenic organisms from animal farms is essential for the development of new drugs and treatment strategies; and the need to develop alternative products. Finding alternatives to the use of antibiotics in farm animals would obviously have concrete positive consequences for producers, animal welfare, and the various players in the production chain. However, antibiotic resistance is a global phenomenon embedded in most ecosystems and spheres of human activity, and is an interesting example of the One Health concept. The term One Health was developed following the challenges posed by zoonoses and the need to establish collaborations between experts from diverse fields (35). Today, the One Health approach is regarded as a collaborative, multisectoral and multidisciplinary framework that acknowledges the interconnectedness of human, animal, and environmental health. Agriculture, including livestock farming, is a central component of human society and has a great impact on the development and spread of AMR (Figure 1). Overall, antibiotics such as tetracycline for example, are not completely metabolized by animals and therefore end up in the feces (36). Antibiotics in addition to promoting antibiotic resistance in the gut microbiome, may also induce an increase in resistance in the immediate environment. For example, Sun et al. (37) observed that veterinary medicine students experienced modulation of their microbiome, including an increase in ARGs, during a 3-month internship on pig farms. In fact, their microbiome evolved to resemble those of farm workers. Another study by Sun et al. (38) also demonstrated that the microbiome of agricultural workers was different from that of other villagers. Recently, Bai et al. (39) showed that bioaerosols from dairy farms and chickens containing ARGs and antibiotic-resistant bacteria, including Staphylococcus spp., can easily spread through the air over distances of up to 10 km. The use of antibiotics in animals is also known to increase the population of antibiotic-resistant bacteria in their feces. The presence of antibiotic residues in feces can be particularly problematic, among other things when feces are used as liquid manure. In fact, as already discussed in other reviews (40, 41), antibiotic residues from animal feces can contaminate plants, vegetables, the environment and people, through the consumption of contaminated products. The overuse of antibiotics in agriculture has sometimes unexpected repercussions. For example, Hammer et al. (42) demonstrated that giving antibiotics to cattle restructures the microbiota and increases the production of methane present in the dung and therefore increases greenhouse gases.
The aftermath of bans on AGP use in livestock production by some countries and the withdrawal of antibiotic use for growth promotion has resulted to increased enteric pathogenic challenges during the early growth phase of many livestock species including poultry and fish, causing immense economic losses in the livestock industry (43). Therefore, decreased AGP use must be matched with the use of products conferring similar benefits as antimicrobials to ensure maintenance of animal productivity, health and welfare, and food security worldwide. Therefore, alternative products that can replicate the effects of antimicrobials in livestock production must be developed. These products can be administered as nutritional additives or as microbial and immune enhancement compounds (Figure 2). The next sections present an overview of some of the progress made in developing alternatives to antimicrobials for use in livestock production.
3 Antimicrobial alternatives: microbial and immunotherapeutic strategies
3.1 Microbial strategies
A host's microbiome which is made up of microorganisms and their by-products (44) is increasingly recognized as a dynamic microbial organ that can evolve to respond to various factors (45). Livestock microbiomes are essential for health and growth and their dysbiosis has been associated with livestock pathologies such as mastitis (46) and calf diarrhea (47), among others. Moreover, as demonstrated by Verbeek et al. (48), microbial composition and metabolite production correlate with certain aggressive behaviors in pig, such as tail biting. In ruminants where the microbiome is an essential component of digestion, dysbiosis can lead to several pathologies, such as bovine respiratory disease (49), ruminal acidosis (50, 51) and pest des petits ruminants (52).
Antibiotics, being generally broad-spectrum molecules, have repercussions on the composition of the microbiome of animals, even after a single dose, especially if given at the early life (53). Jo et al. (54) demonstrated that finishing pigs treated with lincomycin had a greater incidence of diarrhea and that the animals' microbiota had a greater proportion of detrimental bacteria and fewer bacteria involved in fiber degradation. Currently, it is suggested that a restructuration of the microbiome (including the metabolites produced) and a modulation of the immune response or mitochondria function by antibiotics (55, 56) and other immunotherapeutic strategies (57) could be among the factors promoting animal growth. Although the exact mechanisms still remain hypothetical, the involvement of the microbiome was confirmed in 1963 by Coates et al. (58) who demonstrated that antibiotics had no effect on the growth of chicks without microbiota, compared to birds colonized by a microbial population. Thus, it is increasingly evident that it is necessary to have effective alternatives to antibiotics to treat animals with the hope to see a real reduction in antibiotic resistance. Research in livestock production has explored various methods for improving animal health and performance, including the use of microbiota transplantation (59), engineered microbes and phage therapy.
3.1.1 Microbiota transplantation
Integrating the microbiome as a new variable in animal husbandry for production and health management can reduce the environmental footprint while increasing food yield, and improving animal health. Microbiota transplantation, particularly through fecal microbiota transplantation (FMT), is a part of a broader shift toward understanding and leveraging the microbiome in livestock production. FMT consist of transferring feces from a healthy individual to the gastrointestinal tract (GIT) of another individual presenting symptoms of disease. The feces from the healthy donors contain both health-associated microbes and byproducts of fermentation, including anti-inflammatory compounds, and can resolve symptoms associated with dysbiosis or imbalance of the GIT microbiome. It is a promising method for improving the GIT microbiota in livestock with the potential to improve feed efficiency, alleviate diarrhea, reduce methane emissions, and enhance overall health and performance (60–64). For instance, a study on growing calves found that FMT ameliorated diarrhea and improved growth performance, with specific alterations in the gut microbiota and metabolomic profile correlating with these improvements (62). Similarly, recent reviews have highlighted the potential effect of FMT in pigs, specifically its role in improving GIT microbiota and potential to enhance health and production outcomes (60, 63). These data confirm that FMT can be used as an effective method of preventing or treating GIT disorders as well as improving livestock health and performance.
However, the use of FMT in livestock production also raises biosecurity and regulatory concerns, as the donor's microbiome may contain non-desirable microorganisms, which need to be addressed before widespread adoption (60). Lundberg et al. (65) highlighted the need for systematic experiments to evaluate the stability of microbial transplantations, considering recipient status and housing systems. Canibe et al. (60) discussed the potential application of FMT in pig production, emphasizing the need to identify and optimize factors that can influence its impact. McCormack et al. (66) cautions that FMT may not always lead to the desired outcomes, as seen in a study where FMT from highly feed-efficient pigs had detrimental effects on growth in offspring. These studies collectively underscore the need for further research to fully understand the potential and limitations of FMT in livestock production.
3.1.2 Engineered microbes
Advancement in gene-editing systems such as zinc finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindromic repeats (CRISPR)/Cas9 have dramatically increased the efficiency of producing gene edited (GE) livestock animals including pig, cattle, sheep and the expansion of the application of GE livestock animals beyond biomedicine (67–69). CRISPR/Cas9 nucleases have shown promise in producing sequence-specific antimicrobials in livestock production, with the potential to target specific bacterial strains and genes (70). Bikard et al. (71) and Citorik et al. (72) both demonstrated the potential of this technology in targeting specific genes in bacteria, such as virulence and antibiotic resistance genes, to selectively kill harmful bacteria. Bikard et al. (71) showed that the development of programmable, sequence-specific antimicrobials using the RNA-guided nuclease Cas9 delivered by a bacteriophage in a mouse skin colonization model had the ability to reprogram Cas9 to selectively kill virulent Staphylococcus aureus by targeting virulence genes and to destroy staphylococcal plasmids carrying ARGs (aph-3 kanamycin resistance gene), while immunizing avirulent staphylococci to prevent the spread of plasmid-borne resistance genes. Other studies further expanded on this by using CRISPR/Cas9 and other systems to induce gene insertion in cattle and other animals to improve health and other traits (73). For example, the insertion of NRAMP1 gene resulted in transgenic cattle with increased resistance to tuberculosis (74). These studies collectively highlight the promising applications of gene editing in developing antimicrobials and enhancing disease resistance in livestock.
However, there are limitations to the use of engineered microbes in livestock production. One primary concern is that CRISPR/Cas9, TALENS and ZFNs can produce off-target mutations in the genome. These unintended alterations can result from the guide RNA inadvertently pairing with unrelated sequences, leading to mutations in genes that are not the intended target (75–77). Various studies have noted that off-target effects could undermine the safety and reliability of CRISPR/Cas9, ZFNs and TALENS applications in livestock (75–77). Although anti-CRISPR proteins have been shown to mitigate off-target activity, yet their utility is limited due to challenges in delivery and biological compatibility (78, 79). Mosaicism is another significant challenge inherent to genome editing in livestock, which can occur post-modification. Mosaicism results from the introduction of edits in only a subset of cells during early embryonic development (80, 81). This genetic variation complicates efforts to produce uniform livestock populations with consistent phenotypes, as not all offspring of a genetically modified parent may express the intended traits (80, 81). Such heterogeneity can pose challenges in breeding programs aimed at enhancing specific desirable traits, limiting the effectiveness of these technologies in producing commercially viable livestock. The public perception of genetically modified animals remains a contentious issue. Concerns regarding animal welfare, environmental impact, and food safety influence consumer acceptance. Therefore, ethical considerations surrounding animal welfare and the potential for unintended consequences from genetic modifications must be addressed to gain public trust and facilitate the adoption of these technologies in production systems (82).
3.1.3 Bacteriophages
Bacteriophages (phages) are ubiquitous in virtually all environments and ecosystems (e.g., soil, ocean, feces, wastewater and the GIT of animals, etc.) (83, 84), and as natural predators of bacteria (85), they play an essential role in regulating bacterial populations (86). Indeed, they represent the most common and abundant biological entities on the planet with an estimated total number of 1031 (84, 87), and in some environments they are 10 times more numerous than bacteria (88). As bacteriophages can only infect specific bacteria (89, 90), they are increasingly being investigated as a novel solution to replace antibiotics.
3.1.3.1 Advantages and challenges of phage therapy
Among the existing alternative control methods, bacteriophages are promising as they offer several advantages over antibiotics for the control of bacterial infections and to combat multidrug-resistant bacteria. The existence of several phage-based products approved on the market as antibacterial agents for livestock production, mainly for poultry, but also growing pigs, calves and aquaculture, testifies to the effectiveness of bacteriophages (91). Further advantages associated with phages include their ability to target antibiotic-resistant bacteria and their safety for animals and humans, amongst others (Table 2).
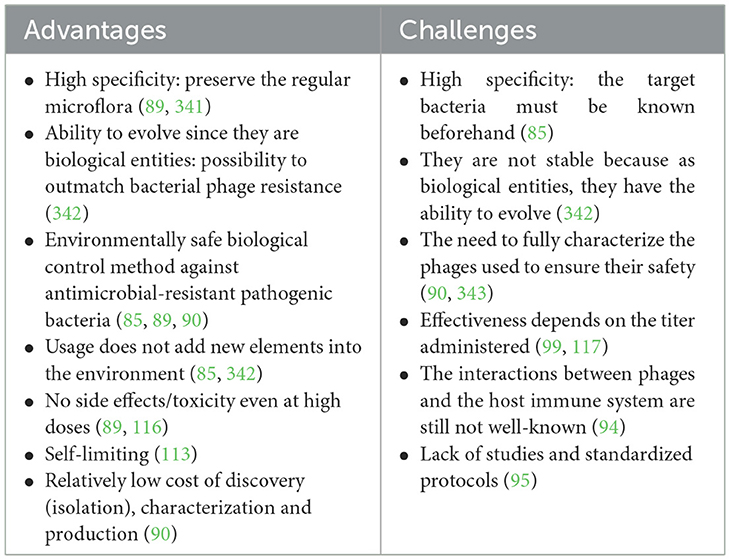
Table 2. Advantages and challenges of phage therapy in livestock production.
However, the use of phages in animal production also present challenges, especially for commercial production. Their high specificity requires prior identification of the target bacteria (85), and their efficacy can be influenced by various factors such as temperature, pH, and the physiological status of the host (92). Another challenge in using bacteriophages for livestock production is the feasibility of their administration to animals. Administering phages orally may necessitate antacids due to their sensitivity to low pH (93). Additionally, difficulties in cultivating host bacteria in vitro can complicate phage production. Fully characterizing phages through DNA sequencing and genome analysis is essential to ensure they are free of virulence or ARGs (90). The efficacy of phage therapy depends on delivering an appropriate phage titer to the infection site, as an insufficient dose can reduce effectiveness (96, 105, 117). Moreover, interactions between phages and the host immune system are not fully understood (94), and the lack of standardized protocols and regulatory guidelines further complicates their commercial use (95). Table 2 presents a summary of the advantages and the challenges of phage therapy in animal production.
3.1.3.2 Phage administration to livestock
Effective phage therapy requires bacteriophages to reach infection site directly, where the problematic bacteria are located (96–100). Numerous experiments have explored various phage delivery methods to prevent or treat diseases caused by pathogenic bacteria that lead to mortality or reduced performance in livestock, or to reduce GIT colonization by undesirable bacteria for which livestock are major reservoirs (Table 3). These delivery methods include oral gavage, injection, incorporation into feed, milk replacer or drinking water, aerosol spray, environmental dissemination, and in ovo injection at the hatchery (in the case of poultry production) (85, 99, 101–110). Keeping in mind that an effective phage therapy depends on whether an important titer of phages reaches the site of infection; the choice of the method used to administer treatment to livestock depends on the type of bacterial pathogen and the stage of infection, as these factors determine the infection site. Moreover, using a combination of administration methods often leads to more effective treatment outcomes. However, it is important to note that some administration methods such as oral gavage and injection are not suitable for some livestock species such as poultry and aquaculture because it would imply the manipulation of thousands of individuals, which is not feasible in a commercial farming context.
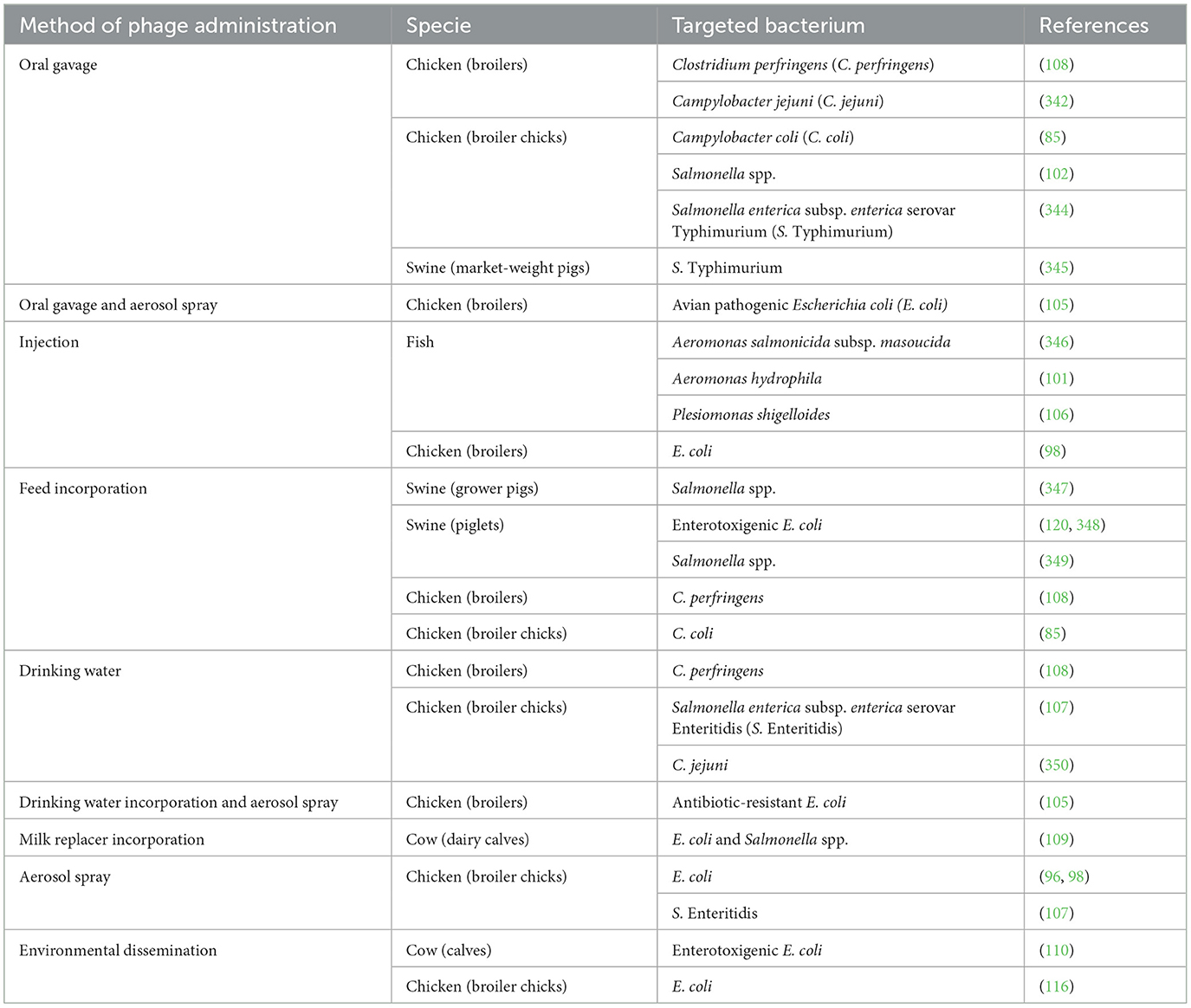
Table 3. In vivo phage trials using various methods of phage administration to livestock.
3.1.3.3. Phage selection process for cocktail preparation
Using a phage cocktail, which contains multiple phages targeting different receptors, is preferred over a single phage to prevent the emergence of resistant bacteria (111, 112). Korf et al. (113) found that a phage cocktail was effective in preventing resistance in Escherichia coli (E. coli), unlike using a single phage. Indeed, by using a phage cocktail, even if the target bacterium acquires resistance to one phage, it is highly likely to remain susceptible to at least one other phage in the cocktail (89, 113).
To control bacterial infections, only virulent phages, which undergo the lytic cycle and avoid DNA exchange with the host, should be used to prevent the spread of resistance or toxin genes (108). Also, the absence of non-desired genes, such as those responsible for AMR, bacterial virulence, and lysogeny, is crucial when selecting phages for a phage cocktail (114). To ensure that the selected phages are well-adapted and effective, it is advisable to isolate phages from the same environment as the target bacteria (115). Phages can be isolated from various environments depending on the target bacteria, including the GIT contents of infected carcasses (85), feces (113), contaminated water (113), wastewater treatment plants (96, 116, 117), slaughterhouse (118), or processing plant (96).
Overall, a good phage cocktail should include phages that specifically infect the target bacterium, are virulent and highly lytic, and can be easily produced with well-characterized host bacteria (113). For instance, Miller and colleagues administered (orally via drinking water, feed or oral gavage) a INT-401 phage cocktail comprising five bacteriophages at 105 PFU/ml to over 900 broiler chickens (0–42 days old) infected with Clostridium perfringens (C. perfringens) and found that administering INT-401 via their feed or drinking water effectively controlled necrotic enteritis and increased weight gain and feed conversion (108). Moreover, although no optimal lysis yields have been established, phages that replicate quickly and produce a high yield are preferred (119).
3.1.3.4 Bacteriophages as growth promoters
An interesting aspect of using bacteriophages in livestock production is their potential as growth promoters. Indeed, studies have demonstrated that adding phages to feed positively impacts the growth performance of antibiotic-free weaned piglets (120, 121) and grower pigs (122). Additionally, a study showed that supplementing chicken feed with a phage cocktail targeting non-pathogenic E. coli significantly improved the feed conversion ratio compared to the control group, and combining this phage cocktail with a commercial probiotic in the feed led to a significant improvement in growth performance (123).
3.1.3.5 Commercially available phage-based products for livestock production
Efforts to use bacteriophages for the fight against pathogenic bacteria has led to the development and emergence of various phage-based products for animal husbandry. Available commercial phage products for animal health have been listed in recent reviews (91, 124). Phage-based products for livestock production are incorporated as food additives in animal feed or drinking water, and some are approved for use in Canada (124). To date, the most targeted bacteria by phage products are Salmonella spp., E. coli and C. perfringens mainly for poultry, but also for swine and calves. A Canadian company (Cytophage Technologies Inc.) has developed AviPhage™, a water-soluble phage solution targeting Salmonella and E. coli in poultry, which is reported to promote animal growth, improve gut health, lower mortality risks, and reduce undesirable bacterial levels without side effects, while another bacteriophage-based product for bovine mastitis is under active development, but not yet available on the market (125).
The use of bacteriophages is still limited and not generally acceptable because of their potential risks of antibiotic resistance transmission. It was demonstrated recently that ARGs, such as sulfonamide-resistant dihydropteroate synthase (sul1), beta-lactamase (blaTEM), and erythromycin resistance gene (ermB) genes were found in bacteriophage DNA samples, while ermB and florfenicol-chloramphenicol resistance gene (fexA) were the most common ARGs in the bacterial population (126). Furthermore, limited experiments have been carried out to fully understand their mechanism of action and development of antibiotic resistance, and correct dosage of bacteriophage inclusion in animal diet (127, 128). Therefore, further research is warranted to elucidate the mechanism of action of bacteriophages and their impacts on livestock production.
3.2 Immunotherapies
3.2.1 Vaccines
Vaccines stimulate the host's immune system and enable the host to respond effectively to an infection. Vaccines function by presenting antigens to the immune system to generate an adaptive response, among other things through the production of B and T cells (129). They therefore have the considerable advantage, compared to antibiotics, of being passive and thus not require human intervention to make a diagnosis and administer medication. Before B and T cells are activated, the antigens must first be recognized by antigen-presenting cells, which triggers the immune process. Although this process occurs naturally, it is possible to increase its effectiveness by adding immunostimulating compounds, which are known as adjuvants, to the vaccine (130). The adjuvants can be compounds as varied as toll-like receptor agonists, cytokines or saponins. Mineral salts and microparticles also serve to protect the antigen and allow slow diffusion. Recently, polyphosphazenes are also being explored as an adjuvant in vaccines for various animals and could stimulate the innate immune response (131).
Vaccines have been instrumental in effectively controlling the incidence of some livestock pathogenic diseases. For example, Salmonella bacteria, a culprit in diarrhea in humans (132) and livestock, is a common pathogen of major food animals such as pig, poultry, and cattle (133). It is particularly worrisome since it can be found at all points of the food chain. In 2017, it caused 95.1 million illnesses worldwide and more than 50,000 deaths (134). In addition, it is considered by the WHO to be a priority two bacterium for antibiotic resistance (135). Fortunately, there are effective vaccines against Salmonella enterica. Smith et al. demonstrated in 2018 (136) that the use of a commercially available live attenuated vaccine (Salmoporc) in sows significantly reduced the prevalence of the bacterium in farrow-to-finishing herds. Recently, Schmidt et al. (137) demonstrated that the use of the Salmoporc vaccine induced an increase in cytokine-producing CD4+ T cells specific to Salmonella Typhimurium (S. Typhimurium) in all organs, with the most marked effects at the level of lamina propria lymphocytes of the jejunum and ileum. In cattle, although vaccines exist, their effectiveness remains limited. A recent study found a partial effect of two commercial vaccines against Salmonella Dublin (138). In poultry, vaccination against S. enterica is considered effective and reduces the presence of the bacterium at different points in the food chain (139). However, the diversity of Salmonella serotypes makes it difficult, if not impossible, to get rid of the bacteria completely on farms. However, new products are regularly developed to increase the effectiveness of vaccine strategies. For example, a new trivalent vaccine (Salmonella serovars Enteritidis [O:9, serogroup D], Typhimurium [O:4, serogroup B] and serovar Infantis [O:7, serogroup C1]) has been recently evaluated and shown to reduce the shedding of the bacteria (140). A recent study showed in chicks that taking antibiotic treatment before a live attenuated vaccine against S. Enteritidis reduced the effect of the vaccine (141); the vaccine strain being itself a bacterium sensitive to antibiotics. This therefore demonstrates the importance of knowing the parameters that can influence the vaccine effect.
Certain pathogenic strains of the bacterium E. coli are also particularly problematic for the food industry and are responsible for several poisonings. Globally, E. coli is a bacterium known to be prone to acquiring antibiotic resistance genes and therefore difficult to treat (142). This is the case, for example, of E. coli O157:H7 in cattle (143, 144). Weaned piglets are also particularly susceptible to E. coli infections, which can result in severe diarrhea. Vangroenweghe and Boone recently showed that the vaccination of piglets with an oral live non-pathogenic E. coli F4/F18 made it possible, in addition to offering protection against the bacterium, to increase the zootechnical performance of the animals and to reduce secondary infections by Streptococcus suis (145), another problematic pathogenic bacterium for the swine industry. This study illustrates well the interconnection between bacteria sharing the same ecosystem and that vaccines can influence more than one pathogen. Although vaccines exist, handling and associated costs can be prohibitive factors to their use (146, 147). Available vaccines for controlling mastitis in cattle and novel vaccine technologies in veterinary medicine were summarized recently (57, 148).
Noteworthy developments in vaccine technologies in the field of livestock production are undergoing significant transformation, driven by advances in molecular biology, immunology, and veterinary medicine. This evolving landscape highlights a shift from traditional vaccine modalities to more innovative approaches, including mRNA vaccines, viral vector vaccines, and DNA vaccines, which promise enhanced effectiveness against a range of infectious diseases (149–152). For instance, viral vector vaccines such as the ChAdOx1 RVF vaccine have demonstrated significant positive effects. Stedman et al. (152) have reported that this vaccine is not only safe and immunogenic but also offers robust protection across various livestock species against Rift Valley fever, with ongoing field trials supporting its future deployment in veterinary practice. Such advancements highlight the role of innovative vaccine platforms in addressing complex disease challenges in livestock, marking a notable departure from conventional inactivated or live vaccines.
Proper understanding of the roles of genes during infection have been used for the development of messenger RNA (mRNA) therapeutics/vaccines with high specificity, validity and safety for the management of many animal and human diseases, including cancer (153, 154). For instance, a study discovered that vascular endothelial growth factor A (VEGF-A) 165 mRNA transcript enhanced cardiac function when administered after 1 week of post mammary infection in swine (155). Currently, the development of mRNA vaccine has great potential due to their low risk of insertional mutagenesis, low cost of production and high potency. When the vaccine is introduced to the host's cell, the RNA is translated, and the antigen is expressed at the specific site (148). Several mRNA vaccines have entered clinical trials to prevent infectious diseases including influenza virus, Zika and rabies (156). RNA or mRNA vaccine uses part of mRNA that encodes antigens coated in vesicle carriers (148).
Furthermore, DNA vaccination emerges as another promising strategy. Choudhury et al. emphasize how recent developments in genetic engineering have led to the creation of subunit and recombinant vaccines specifically designed for ruminants. These vaccines enhance protective immune responses and have begun to replace traditional methods due to their safety and efficacy (150). Advances in delivery mechanisms and adjuvant formulations are also contributing to the effectiveness of these vaccines, further underscoring the move toward more sophisticated immunization approaches (148, 157).
Emerging research further indicates the necessity for cross-protection strategies, especially given the high variability of pathogens affecting livestock. As Davis et al. (158) stated, the identification of correlates of protection is vital for the rational design of vaccines against highly diverse viral pathogens like African swine fever and foot-and-mouth disease. This information guides the development of vaccines that can elicit cellular and humoral immunity, ideally leading to broader protection across different strains and species. The integration of biotechnological advances into the vaccine development pipeline presents additional opportunities to combat endemic diseases. For instance, irradiated vaccine technologies have been highlighted as solutions tailored to the specific needs of local livestock industries (159). This approach not only addresses disease prevention but also encourages stakeholder participation in vaccination programs, thereby improving overall vaccination coverage and efficacy in target populations (160, 161).
Although vaccines are an important weapon in our arsenal of defense against pathogenic microbes, especially in the context of antibiotic resistance, challenges remain. Vaccine development takes time and money. Also, antigenic variation and economic barriers can impede vaccine adoption (161). Therefore, future directions in research and the regulatory landscape will need to address these challenges to facilitate the successful implementation of new vaccine technologies in livestock health management. Recently, it has been proposed that a “One Health vaccinology” approach, consisting of using an inter-species vaccine strategy, could be a solution, especially for vaccines against zoonoses (162).
3.2.2 Immunoglobulins
Generally, the host first line of defense to infection is known as the innate immune response and is aimed at containing and eliminating the infectious organism from the host system (163). The innate host defenses activate adaptive immune responses by changing cell populations and soluble factors in the affected tissues. The innate host immune responses involve activating the local stroma, release of cytokine and chemokine messengers, attracting and activating neutrophils/heterophils to the affected sites, stimulating macrophages and natural killer cells, stimulating effector molecules like enzymes, collectins, acute phase proteins and host defense peptides, and triggering the complement system (163). Immunoglobulins, also known as antibodies, are effector molecules of humoral immunity and are among the innate immune responders that protect against infectious diseases.
Likewise, colostrum is the first milk produced by mammals after parturition and it consists of a very high level of antibodies (e.g., immunoglobulins) that provide passive immunity to the neonate, thereby protecting them from infectious disease pathogens in the first week of life. Passive immunity can be transferred to susceptible animals using hyperimmune plasma derived from animals with a very high immune response and it is a strategy that can be used to reduce antibiotic application by increasing the immune system's ability to fight infection (163, 164). However, there are several licensed immunoglobulin-based products available for passive immunization of animals, including E. coli-specific antibodies for calf; antibacterial bovine serum antibodies for cattle, calf and sheep; clostridial antitoxins for cattle, calf, goat, sheep, swine and horse; tetanus antitoxin for horse, cattle, sheep, swine and goat; anti-West Nile virus antibodies, anti-endotoxin antibodies, antibacterial plasma antibodies and equine plasma for horse; and spray dried plasma for weaned piglet (163). Slaughterhouse plasma, milk, and whey are natural sources of immunoglobulins that are readily available and inexpensive. In an experiment carried out by Hedegaard et al. (165), it was revealed that purified natural plasma IgG obtained from slaughterhouse contained antibody reactivity and possessed antibacterial effects against porcine bacteria, such as E. coli O138, E. coli F4, E. coli F18, and Salmonella enterica Diarizonae (165). However, some limitations to the use of immunoglobulins include: (1) purified immunoglobulin products need to be controllable, easy to take orally, and compatible with automatic feeding or drinking systems, (2) formulations should provide optimal shelf life at ambient temperature and resistance to denaturing and fragmentation caused by gut bacteria, (3) most importantly, immunoglobulins need to be certified that they are free of adventitious agents, including porcine circovirus type 2, porcine respiratory and reproductive syndrome virus, and porcine endemic diarrhea virus (163).
3.2.3 Host defense peptides
Host defense peptides (HDPs) have broad spectrum antimicrobial activities against several pathogens, including Gram-negative and Gram-positive bacteria, fungi, parasites and viruses, including multidrug-resistant strains and also regulates immunity, making them valuable in fighting microbial challenges. Several studies have proofed the ability of HDPs (e.g., human cathelicidin LL-37 and human β-defensin-3) in stimulating monocytes and other immune cells to secrete chemokines and cytokines, thus indirectly stimulating the recruitment of immune cells to infection sites (166–168). For instance, Cuperus et al. (166) reported that chicken cathelicidin-2 (CATH-2) displayed potent antibacterial properties while also modulating the immune response in chickens. This dual role enhances not only the immediate defense against pathogens but also the host's overall immune competence, which is particularly advantageous in intensive livestock production systems where stress and disease incidence are high. HDPs can enhance the resilience of the gut microbiota and promote the fitness of the host under pathogenic challenges. This is crucial for animals in high-density farming conditions, where disease outbreaks can significantly impact animal health and productivity (169, 170). A plethoral of investigations have indicated that the immunomodulatory effects of HDPs can stabilize the intestinal microbiota, which is essential for maintaining health and preventing disease outbreaks in livestock (review by Findlay-Greene et al. (171)). Not only that, the molecular characterization of HDPs has opened up avenues for recombinant technologies that can create tailored HDPs for specific pathogens affecting livestock (169). HDPs have low toxicity toward eukaryotic cells making them suitable for applications in livestock, where residue concerns associated with conventional antimicrobials are crucial (172). This characteristic aligns with consumer demands for safer meat products and animal welfare concerns surrounding the use of antibiotics in the livestock industry. Currently, research is ongoing on the possible inclusion of HDPs in animal feed to improve disease resistance and eradicate the use of antibiotics (163, 173). Other strategies include breeding animals with improved innate immunity against different strains of bacteria, including the expression of defense peptides specific to the host. This involves genetically selecting animals based on their ability to produce higher levels of immune peptides that will confer greater disease resistance to future generations.
Despite the advantages of HDPs, their use presents several limitations including stability of HDPs in the GIT of animals. The bioactivity of these peptides can be significantly affected by various conditions such as enzymatic degradation and changes in pH, which may lead to reduced efficacy when administered orally. Studies indicate that HDPs structural integrity can be compromised by the harsh enzymatic environment of the gut, limiting their therapeutic potential (174, 175). Moreover, the need for specific formulation strategies to protect these peptides from degradation represents a significant barrier for their practical application in livestock (175). The high cost associated with the purification and expression of HDPs poses a challenge for their widespread implementation in commercial settings (176). There is a need for scalable production methods that maintain cost-effectiveness in fostering the integration of HDPs into standard livestock management practices (176). Additionally, while HDPs are generally associated with low potential for the development of resistance compared to traditional antibiotics, there is emerging evidence that certain bacterial pathogens can develop mechanisms to evade the actions of these peptides (177). Moreover, the regulatory approval of HDPs for use in livestock can be a lengthy process. There is however a need for the establishment of guidelines on optimal dosing strategies and formulations that ensure the effectiveness and safety of HDPs use in livestock (178).
3.2.4 Antimicrobial peptides
Antimicrobial peptides (AMPs) have recently drawn attention due to their broad-spectrum antimicrobial activities, ability to kill rapidly, less toxicity, cell selectivity and immunomodulatory properties. AMPs are small-sized peptides that are crucial in host immune defense in living organisms, including animals, humans, and plants. Antimicrobial peptides are amphiphilic and positively charged and this feature allows them to bind and penetrate bacterial membrane bilayer to induce pores, causing intracellular leakage. Antimicrobial peptides are produced by most microorganisms and based on biological activity, they can be classified as antibacterial peptides, antifungal peptides, antiviral peptides, antiparasitic peptides, and anticancer peptides (179, 180). These features favors them as potential effective alternatives to antimicrobial use in livestock production. The most common AMPs used in feed supplements are divercin AS7 and microcin J25 (106).
Antimicrobial peptides have been found to improve growth performance, nutrient retention, intestinal histomorphology, and balance GIT microbiota (181, 182) (Table 4). Most AMPs modulate innate and adaptive immune responses by modulating pro- and anti-inflammatory cytokines and chemokines (183, 184). For example, supplementation of broiler feed with microcin J25, protected against several E. coli and Salmonella strains, promoted growth performance, improved intestinal morphology, reduced the secretion of pro-inflammatory cytokines (IL-1β, TNF-α, IL-6) (184), reduced intestinal inflammation (185), and increased the production of IL-10 and IFN-γ (186). Similar results were obtained when microcin J5 was added to piglet feed (184). The mechanism of action of AMPs is through their antimicrobial activity which is based primarily on the association of positively charged peptides with negatively charged bacterial membrane components (phospholipids and teichoic acids, lipopolysaccharide) that cause permeabilization of bacteria membrane, pore formation and cell lysis (20).
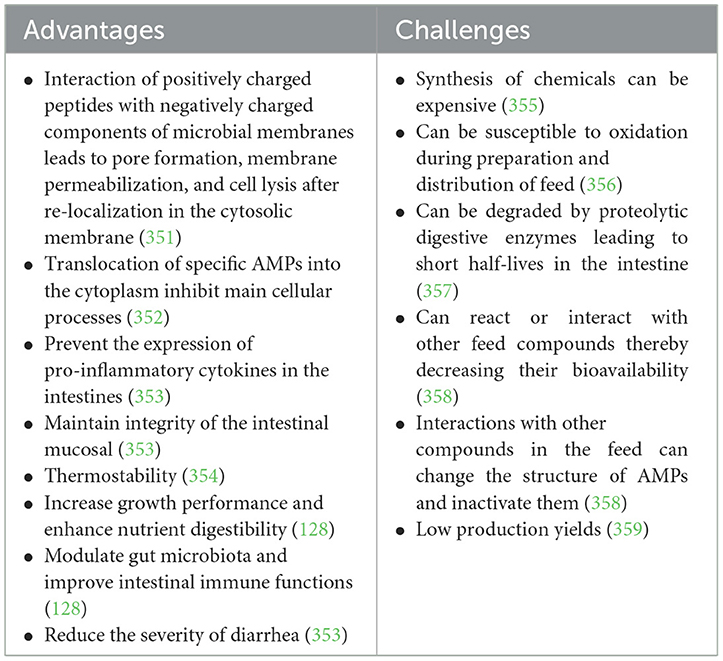
Table 4. Advantages and challenges of antimicrobial peptides as alternatives to antibiotic growth promoters.
It is important to consider the drawbacks to the use of AMPs. The translocation of AMPs into bacteria cytoplasm may inhibit cellular processes, such as synthesis of DNA and protein (187). There is a risk that bacteria can evolve and become resistant to the use of AMP over time. Increased resistance to some antibiotics was observed in variants resistant to an AMP (e.g., nisin-resistant mutants of Streptococcus bovis) (188) (Table 4). Other limitations to the use of AMPs include less stability during feed preparation and storage (183, 189) and lack of proper formulations to improve their bioavailability in the GIT (190).
3.2.5 Hyperimmune egg yolk antibodies
Hyperimmune egg yolk antibody (immunoglobulin Y, IgY) is emerging as a promising immune agent and alternative to antibiotics to combat infectious diseases because it is cost effective for mass production, particularly in livestock. Several egg yolk IgY products are commercially available for disease control in livestock. Egg yolk IgY is mostly administered orally, and it is the most common and convenient route that has been extensively investigated for controlling enteric pathogens (191). Purified egg yolk IgY can also be injected into the animal host via different routes (e.g., intramuscular, subcutaneous, and intraperitoneal) for the control of systemic infections caused by different pathogenic microbes (191).
Studies indicate that addition of IgY to livestock diet serve as preventive, treatment and control measures against pathogenic infections (e.g., E. coli, S. enterica, Campylobacter spp., rotavirus, Cryptosporidium parvum, and Eimeria spp.) as well as enhance host immunity (192, 193). They are less toxic and also environmentally friendly. Egg yolk IgY protect against various infections by preventing pathogen colonization in the GIT, alleviating morbidity or mortality rate and improving growth performance. For example, the induction of passive immunity by oral feeding of hyperimmune egg yolk to young broiler chicks infected with Eimeria tenella and Eimeria maxima improved body weight gains and enhanced immunity (194).
Despite the promising features of egg yolk IgY as a passive immune agent, the stability of egg yolk antibody IgY in the GIT is very crucial for their effectiveness, which is yet to be clearly elucidated. An in vitro study found that the potency of specific IgY was completely lost in a pepsin solution but was mostly retained in trypsin solution, which suggests the degradation of egg yolk IgY in the stomach (195). Likewise, a recent study showed that in vitro simulation of static digestion was easily degraded in intestinal phase but not in the gastric phase (196). The contrasting results in these two studies might be due to differences in the composition of the artificial gastric solutions and intestinal solutions. Egg yolk antibody was highly degraded in chicken gizzard (191), thus prompting suggestions for encapsulation for the controlled release and protection of orally administered egg yolk IgY in livestock. To date, the commercial use of IgY products are yet to be approved or endorsed by any regulatory authority (197). Furthermore, high purification is needed for passive immunization of host with IgY via parenteral routes (e.g., intraperitoneal, intramuscular, and subcutaneous routes). The limitations associated with the use of IgY in livestock include: lower stability of IgY due to its higher molecular weight, lower percentage of β-sheet structure and reduced flexibility, decreased activity at pH 3.5 or lower and loss of activity with irreversible change at pH 3 as well as sensitivity to pepsin digestion (198).
3.2.6 Cytokine immunotherapy
Recombinant DNA technology allows the production of large quantities of animal cytokines which can be used as cytokine immunotherapy for the control of livestock disease. Recombinant cytokines can ameliorate the outcome of mastitis infections in cows whose immune systems are impaired by recruiting effector cells into the mammary gland. In addition, they regulate acute inflammatory responses by stimulating phagocytic cell activity. A recent review reported on the therapeutic and prophylactic applications of recombinant interferons (IFNs) in livestock species (199). For instance, Sordillo and colleagues showed that IFN-gamma (γ) reduced the rate of mortality and morbidity caused by endotoxemia from bacterial toxins in cows (200). It was also added that the treatment of cow's mammary gland with IFN-γ before infecting with E. coli reduced infection of the udder quarters, reduced clinical scores, and shortened the time of infections when compared with the control (200). Recently, Fan et al. (201) assessed the antiviral effects of recombinant porcine interferons (PoIFN-α and PoIFN-γ) as an emergency treatment for pigs infected with African swine fever virus (ASFV), and showed that administering low doses (105 IU/kg) of recombinant IFNs significantly increased cytokine expression, reduced viral replication, and alleviated clinical symptoms during the early stages of infection. This suggests that recombinant porcine IFNs have high protective antiviral effects against ASFV, providing a new strategy for the prevention of the disease (201). It is however necessary to conduct further research in order to better understand the complex interactions between different pathogens and the host, and how cytokines affect their response. Molecular understanding of the role of cytokines in the host immune system will determine how well they can be used therapeutically in the future.
3.2.7 Others
Further developments already finding applications in human health management include epigenetic immunotherapy, regulatory non-coding RNA immunotherapy, nanoparticles therapy, extracellular vesicles for its role in intercellular communication and stem cell based therapies (202–208). For example, nanoparticles enable the examination and manipulation of genetic material and various types such as nano shells, carbon nanotubes, and gold nanoparticles, hold promise for applications such as diagnosis, treatment delivery, and molecular breeding. It offers tools for enhancing animal health, nutrition, and waste management (209). Antimicrobial nanoparticles have been used to replace antibiotics in extenders, as it prevents bacterial growth without inhibiting sperm viability (210). Nanotechnology holds promise for revolutionizing animal health and livestock production, with applications ranging from disease prevention, food safety and production efficiency to food preservation (211). Stem cell-based therapy presents an opportunity for using cells from livestock to enhance their ability to resist infections without the need for antibiotics application. Mesenchymal stem cells (MSCs) derived from various sources, including bone marrow, adipose tissue, and umbilical cord, have been investigated for their immunomodulatory and tissue repair properties (212). Limited trials in livestock indicate that stem cell-based therapies have potential in treating musculoskeletal disorders (213), reproductive disorders (214), and inflammatory conditions (213). More research in this area is needed to explore the therapeutic potential of MSCs in enhancing livestock health and productivity.
4 Antimicrobial alternatives: nutritional strategies
In recent years, several alternatives to AGPs have been explored and those administered to livestock as feed additives (Figure 2), their mode of action, their effects on animal health and productivity, and the advantages and limitations to their use are discussed in the following sections.
4.1 Phytochemicals
Phytochemicals, also referred to as phytobiotics, phytogenics, herbals, or botanicals are naturally occurring chemical compounds found in plants or part of plants including herbs and spices, and essential oils that are incorporated into animal feed to enhance animal productivity. Sample research findings on the impact of phytochemical supplements on animal performance are summarized in Table 5.
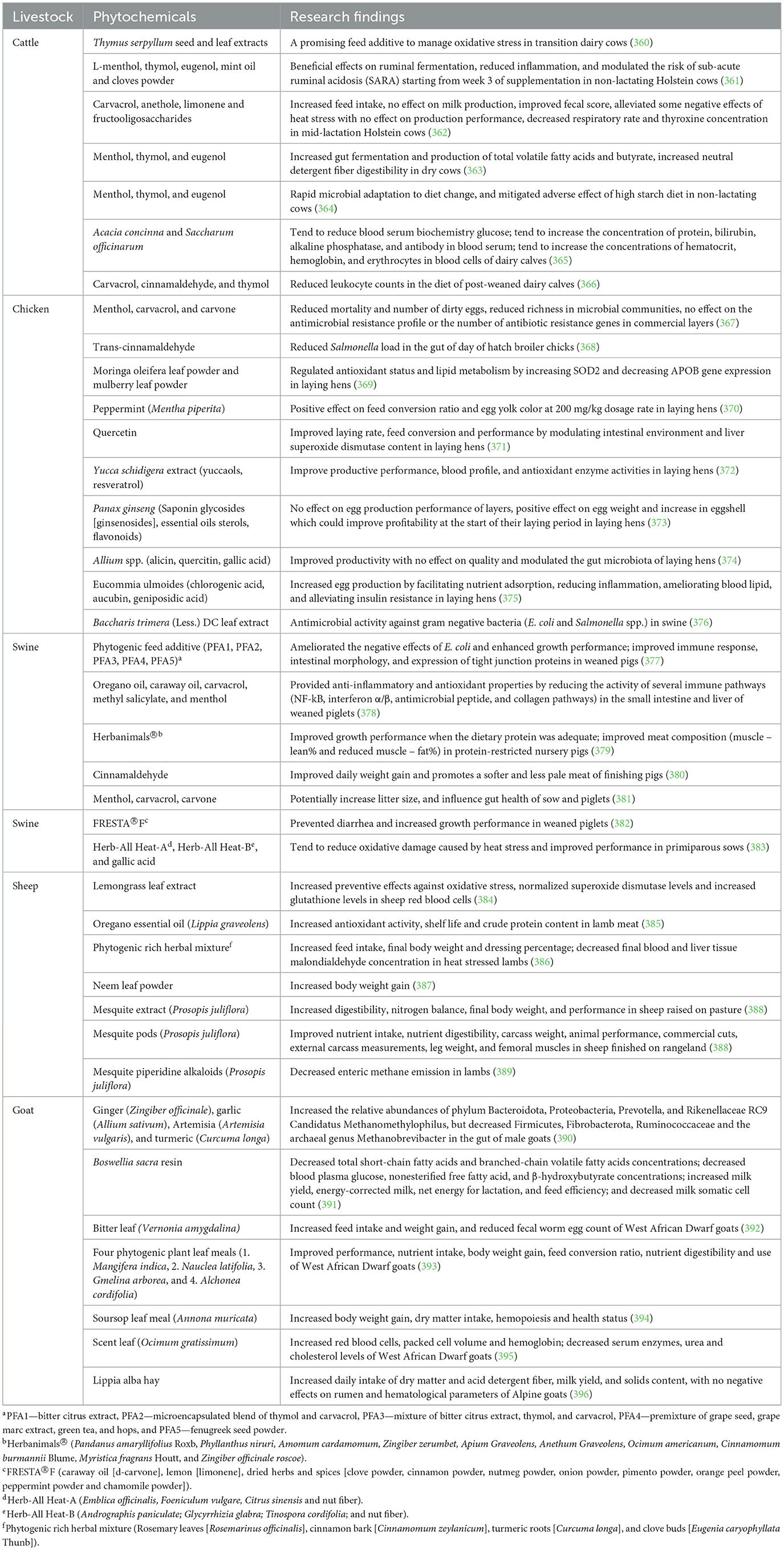
Table 5. Impact of phytochemical feed additives on livestock health and production.
4.1.1 Herbs and spices
Herbs and spices of different types (e.g., thyme, oregano, rosemary, marjoram, yarrow, garlic, ginger, green tea, black cumin, coriander, and cinnamon) have been used in poultry, swine, beef, and dairy production, for their potential as alternatives to AGPs (215, 216).
A range of studies have explored the potential of herbs and spices as alternatives to antibiotics in livestock production (217, 218) (Table 5). Thyme, turmeric, garlic, and cinnamon have all been shown to have growth-promoting effects and antimicrobial properties (218, 219). For instance, Seidavi et al. (218) and Díaz-Sánchez et al. highlighted the potential of various spices (thyme, coriander, and turmeric) in improving poultry health and performance. Turmeric has been found to improve growth performance, gut health, and short-chain fatty acid production in weaned piglets (220). In a recent study, administration of cinnamon and turmeric powder as antibiotic growth promoter substitutes in broiler chickens resulted to increased body weight gain, higher feed intake and feed conversion ratio, improved hematological parameters, and increased ileal and cecal Lactobacillus populations while decreasing coliforms and E. coli (219). In ovo supplementation of herbal compounds, such as grape pomace, resveratrol and chicoric acid, enhanced hatchability, chick quality, and post-hatch performance, while reducing infections and oxidative stress (221, 222). These natural alternatives can help improve poultry performance and reduce the risk of antibiotic-resistant pathogens.
Although some herbs and spices have shown potential as alternatives to AGPs, there are still some limitations to their use such as the requirement of optimal dosage to achieve desired health effects; highly lipophilic nature which limits delivery to enteric pathogens; offensive odor; inconsistent results due to variations in product compositions; can be toxic at high dosage; and high cost of production (19, 223).
4.1.2 Essential oils
Essential oils are becoming increasingly popular as potential antibiotic alternatives for animal production based on their antibacterial, antimicrobial, anti-inflammatory, and antioxidative properties, and ability to enhance feed palatability and gut health (19, 224). Essential oils from thymol, carvacrol, cinnamaldehyde, clove, coriander, star anise, ginger, garlic, rosemary, turmeric, basil, caraway, lemon, and sage are generally recognized as safe for their intended use, and they have been used either individually or synergistically to improve animal health and performance (215, 225).
The addition of essential oils to the diets of livestock has been shown to improve production performance (226) (Table 5). The inclusion of essential oils in the diet have been shown to alter and stabilize intestinal microflora and decrease toxic microbial metabolites in the GIT of animals (225). For instance, the synergistic use of carvacrol and thymol in broiler's feed increased their growth performance, enhanced the activities of digestive enzymes, antioxidant enzymes, composition of fatty acids, and immune responses (227). Essential oils could also exhibit their mode of action via immunomodulatory effects including increased production of immune cells and antibodies, and significant expression of cytokines. For instance, thymol attenuated lipopolysaccharide-induced inflammation in intestinal porcine enterocytes (IPEC-J2 cells) by blocking the production of reactive oxidative species, reduced the expression of IL-8 and TNF- α, reduced cell permeability and transepithelial electrical resistance (215). Supplementation of cinnamaldehyde to lactating Holstein dairy cattle diet improved nitrogen metabolism, reduced milk somatic cell count and increased efficiency of feed use (228).
While a plethora of investigations have documented positive impact of essential oils on animal growth and health, factors such as bad odors, inconsistent results due to variations in product compositions, high volatility, need of high doses to obtain desired results, toxicity and high cost of production limit their application (19, 20).
4.2 Probiotics
Probiotics, also known as direct fed microbials, are live microbial feed supplements which beneficially affect the host by improving intestinal microbial balance. Several species of bacteria (Bacillus, Bifidobacterium, Enterococcus, Lactobacillus, Streptococcus, and Lactococcus spp.; and yeast [Saccharomyces spp.]) have been used as probiotics in livestock feeding (poultry, dairy, swine, and pig) (229, 230).
Probiotics have been shown to improve growth performance and reduce pathogenic bacteria in animal GIT (229). A plethora of investigations have shown that probiotics, alone or combinations of different beneficial bacteria and/or yeast have growth-promoting effects on poultry (500), swine (20) and ruminants (231, 232) (Table 6). The mode of action by which probiotics exhibit their beneficial effect was recently reviewed (231, 232). Probiotics modulate gut microflora by competing with pathogenic bacteria for nutrients; favor the growth of beneficial bacteria against pathogenic bacteria; production of lactic acid and short chain fatty acids, and reduction of pH to create a hostile environment for pathogenic bacteria; secretion of antibacterial substances like bacteriocins; detoxification of inhibitory chemicals like amines or nitrates, and prevention of bacterial adherence and translocation (233). Furthermore, probiotics also function to maintain epithelial cell homeostasis and survival via the production of cytokines, prevent cell apoptosis and improve cell regeneration, improve barrier function by modulating cytoskeletal and epithelial tight junction, and increase synthesis of mucin (234, 235). Probiotics regulate host immune responses to pathogenic bacteria by reducing pro-inflammatory cytokines; increasing production of IgA; and promoting specific and non-specific immune responses against pathogens (236). Sample data on the impact of probiotics supplemental feeding on livestock health and production are summarized in Table 6.
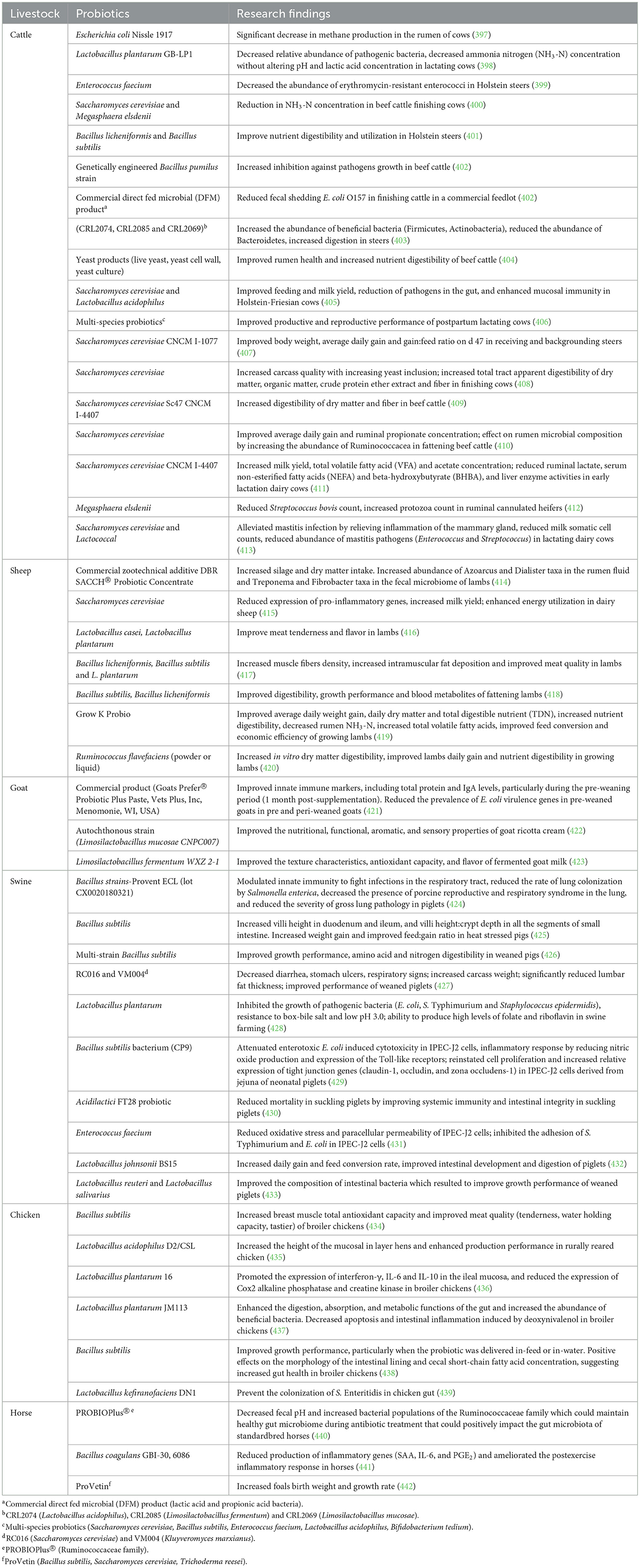
Table 6. Impact of probiotics feed additives on livestock production and health.
Despite the promising effects of probiotics as alternatives to AGPs; limitations to their use abound, such as differences in the quality and dose of some probiotic products, poor rate of survival in the GIT, inactivation of live beneficial bacteria during feed manufacturing, storage or transport, and allergy and transmission of antibiotic resistance genes (20, 237).
4.3 Prebiotics
Prebiotics are non-digestible feed components (non-starch polysaccharides or oligosaccharides such as mannan-oligosaccharide; fructans [fructooligosaccharide and inulin], galactans [galacto-oligosaccharide], malto-oligosaccharide, lactulose, lactitol, and gluco-oligosaccharides) and so on that stimulate the growth of beneficial bacteria in the GIT (238, 239). These non-digestible oligosaccharides are fermented in the GIT by beneficial bacteria and serve as a source of energy for the microbiota.
Prebiotics administration have shown promise in improving growth, immunity, and intestinal health in animals. Prebiotics perform several functions, including inhibition of pathogen attachment to cells, immunomodulation, increased the concentration of antibodies (IgG and IgM), alteration of gut morphology, reduction of intestinal pH, impact microbial populations (e.g., increase population of beneficial. bacterial strains such as Bifidobacterium spp. and Lactobacillus spp.) and decrease pathogenic bacteria (e.g., E. coli and Salmonella spp.), and promote fermentation-based synthesis of antimicrobial chemicals (20, 240). Prebiotics have also been shown to abate heat stress (241); and improve meat quality traits of chicken without affecting lower redness index, lightness and yellowness (242). A recent review on the prebiotic effects of seaweed polysaccharides demonstrated that they may be used to promote pig health throughout the production cycle, hence lowering antibiotic use (243). Sample studies showing the impact of prebiotics on livestock health and production are summarized in Table 7.
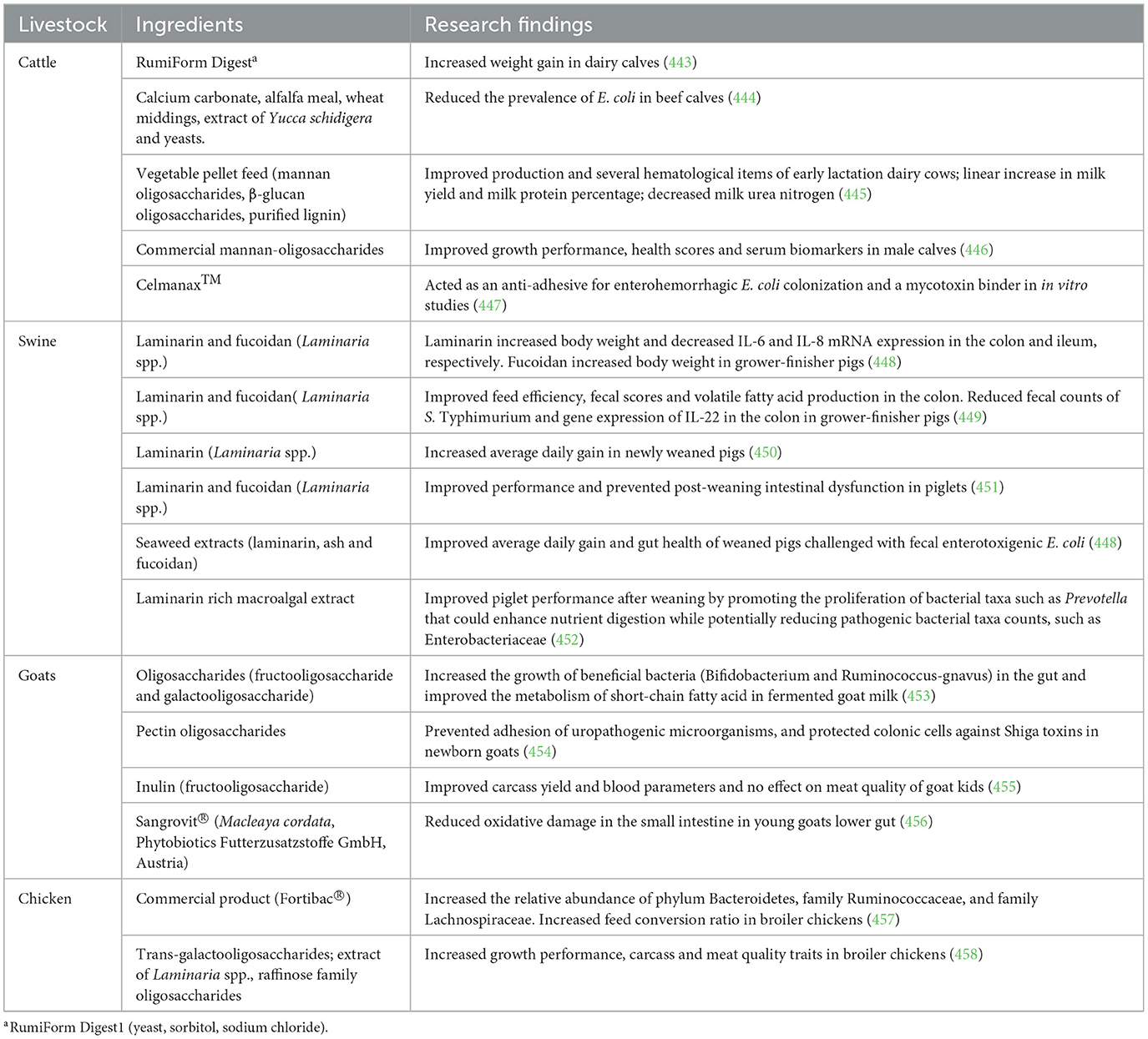
Table 7. Impact of prebiotics feed additives on livestock health and production.
Despite the benefits of prebiotics, its administration in large quantities might cause bloating or diarrhea (20, 244), and their effectiveness can be inconsistent across different animal species or environmental conditions (245, 246). Furthermore, competition from pathogenic organisms also limits the utility of prebiotics. Some pathogens can utilize prebiotics for their benefit, leading to increased resistance or growth rates of harmful bacteria. Fuhrmann et al. (246) reported that both single applications of the prebiotics (Inulin and fructooligosaccharides) failed to significantly reduce E. coli fitness. This suggests that adverse interactions can occur when prebiotics are included in certain dietary contexts, underscoring the complexity of gut microbiota interactions and the necessity for additional research to optimize prebiotic applications (247). Prebiotics also have the potential to develop resistance such as co- and cross-resistance with antibiotics (248).
4.4 Synbiotics
Synbiotics refers to the synergistic use of probiotics and prebiotics to counteract the challenges faced with the survival of probiotics in the intestines. The combined use of probiotics and prebiotics in livestock is reported to be more efficacious on gut microbiota (240), as it increase the production of short chain fatty acids and lactic acid, reduce the concentrations of branched chain fatty acids (20, 240), and increase digestibility and daily weight gain (501). For instance, lambs fed with Saccharomyces cerevisiae and mannan oligosaccharide had improved growth performance and daily weight gain with little effects on carcass traits and visceral mass (249). Sharma et al. (250) investigated the effect of synbiotics (fructo-oligosaccharide, and Lactobacillus plantarum CRD-7) on the growth performance, nutrient digestibility and fecal microflora in murrah bufallo calves and reported increased digestibility, average daily gain, immune response, antioxidant enzymes, fecal microbiota and metabolites, and reduced fecal score and incidence of diarrhea.
The mechanisms by which synbiotics affect the host include the use of prebiotic to promote the growth of probiotic bacteria in the intestines. Synbiotics impact the immune system of the host via the production and maturation of leukocytes. A study in broiler chickens showed that early in ovo treatment with synbiotics modulated the production and maturation of leukocytes. Likewise, the combined use of the Bifidobacterium breve probiotic and galacto-oligosaccharides prebiotic significantly enhanced defense against fatal intestinal infections due to multidrug-resistant Acinetobacter baumannii in a mouse model (251). Recent data on the impact of synbiotics on livestock production and health is summarized in Table 8.
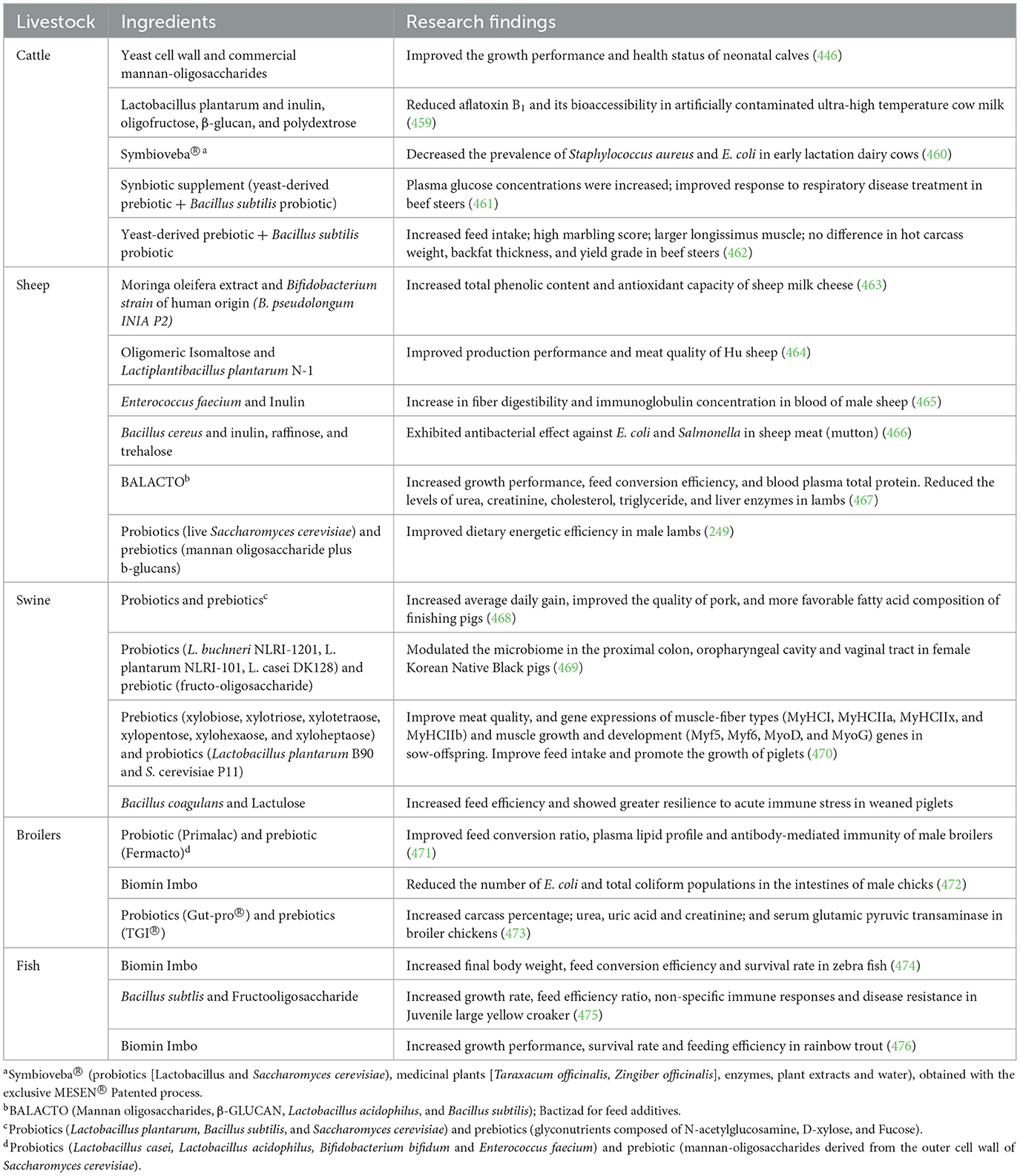
Table 8. Impact of synbiotic feed additives on livestock health and production.
4.5 Postbiotics
Postbiotics are also referred to as paraprobiotics, parapsychobiotics, ghost probiotics, metabiotics, tyndallized probiotics, and bacterial lysates. Postbiotics, as defined by the International Scientific Association of Probiotics and Prebiotics (ISAPP), are preparations of inanimate microorganisms and/or their components that provide health benefits to the host (252). A recent review updated this definition to include non-viable microbes or cell fragments, with or without metabolites, while emphasizing that purified metabolites alone do not qualify as postbiotics (253). Postbiotics have gained significant attention due to their enhanced safety, longer shelf life, and ease of production (254). They carry no risk of transferring antibiotic resistance genes and can be delivered to specific sites with encapsulation techniques, making them an attractive alternative in health and nutrition (255).
Several recent reviews have examined the effect of postbiotics use in livestock production (254, 256). Postbiotics improve growth performance (257, 258), increase nutrient digestibility and milk yield (259), improve gut health (260, 261), improve meat quality (262), anti-inflammatory and anti-oxidant properties (263, 264) in animals. For instance, Izuddin and colleagues reported that administering postbiotics from L. plantarum RG14, RG11 and TL1 affected antioxidant enzymes by increasing glutathione peroxidase in post-weaned lambs (264). Sample data on the impact of postbiotics feed additives on livestock health and performance is summarized in Table 9.
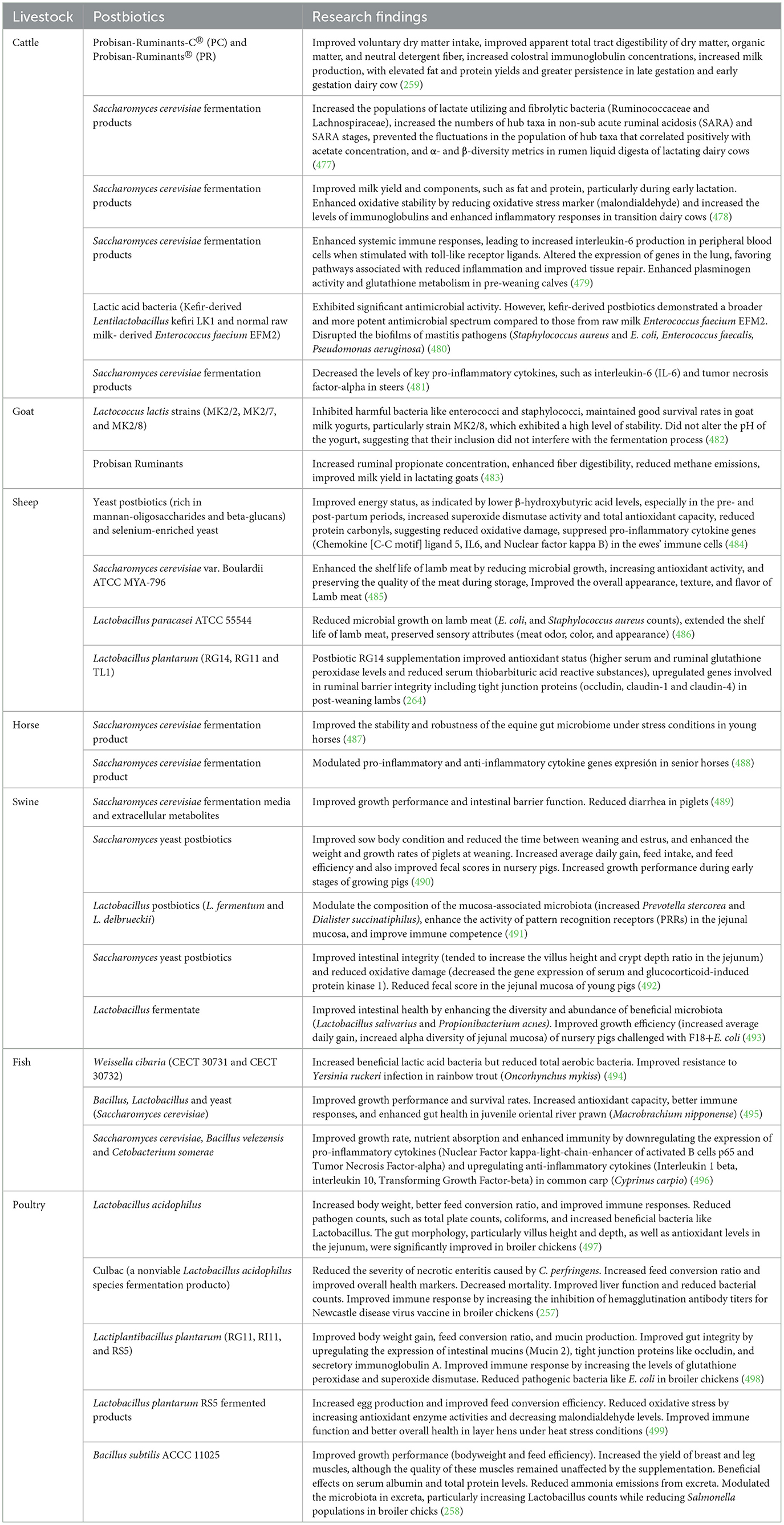
Table 9. Impact of postbiotic feed additives on livestock production and health.
The mode of action by which postbiotics exhibit their beneficial effects was recently reviewed (255). Postbiotics enhance gut health, nutrition, milk yield and composition, and immune response through various mechanisms. Postbiotics interfere with pathogen establishment and maintain the barrier function of the intestinal mucosa, thereby enhancing host–microbiome balance and contributing to a state of eubiosis. The effects of postbiotic treatments in reducing the severity of E. coli infections, improving gut health and growth performance in broilers and layers have been demonstrated (260, 261). The antimicrobial properties of postbiotics are mostly due to their metabolites (bacteriocins, short-chain fatty acids, etc.) which play significant roles in protecting against pathogen invasion and promoting gut health (265). Postbiotics modulate native probiotic strains, thereby maintaining intestinal microbiota and host homeostasis. Additionally, postbiotics can utilize bacterial extracellular vesicle secretion systems to mediate microbe–microbe communication (i.e. quorum sensing) and influence host signaling pathways (265). Postbiotics have longer shelf life, and are easy to produce (265).
Despite the numerous advantages associated with postbiotics, they also come with some limitations including lack of clear international regulatory standards, incomplete understanding of host-microorganism interactions which are responsible for producing specific postbiotics or a combination of postbiotics (266). Therefore, more research is needed to foster a more comprehensive approach and achieve harmonized regulatory frameworks.
4.6 Acidifiers
Acidifiers are compounds with acidic properties including organic and inorganic acids, and which demonstrate antimicrobial properties. Organic acids include benzoic, citric, formic, fumaric, lactic, and propionic acid or their salt counterparts (such as calcium, potassium, sodium formate, or sodium fumarate) (267, 268). Inorganic acids include hydrochloric, sulfuric, or phosphoric acids. Unlike inorganic acids, organic acids consist of one or several carboxyl (COOH) functional groups which play an important role in their activity. Some acidifiers can also form complexes with minerals including calcium (Ca2+) and zinc (Zn2+) cations that limit their absorption in the digestive tract. Acidifiers are generally recognized as safe agents and have been used in livestock diets and drinking water to improve their growth performance (269).
Organic acids have been found to improve gut health, nutrient digestibility and mineral utilization, inhibit inflammatory processes, and have antimicrobial effects (20, 270). The main mechanisms of the mode of action of acidifiers are: (i) reduction of the pH level in the gut which tend to reduce the numbers of pathogenic bacteria, (ii) direct killing of pathogenic bacteria by penetrating through their cell-wall, (iii) increase acid-tolerant beneficial species e.g., Lactobacilllus spp., (iv) increase nutrient digestibility by promoting retention of protein and dry matter, (v) improve absorption of mineral and use of phosphorous, and (vi) improve gut health via direct effects on epithelial cells (20). For decades, organic acids have been widely used as feed preservative due to their strong antibacterial and antifungal properties (271). The beneficial effects of organic acids are often enhanced when used as blends rather than as single products. Studies have demonstrated that organic acid blends improve feed conversion ratios in broiler chickens (272) and improve growth performance, serum antioxidant status, and intestinal health in weaned piglets (273), underscoring their potential as effective alternatives to antibiotics use in livestock production. Acidifiers positively influence intestinal microbiota, enhance immune response and growth performance, and reduce pathogen loads in livestock (20, 274). These beneficial effects of acidifiers in swine and poultry have been comprehensively reviewed (20, 275).
In spite of the demonstrated beneficial effects, the effectiveness of acidifiers can vary depending on factors such as the type of acid or form (acid, salt, coated or uncoated), chemical properties (pKa value, molecular weight, minimum inhibitory concentration of the acid), target microorganism, age of animal, diet composition and animals' health status and so on (20, 267, 270). For example, Ravindran and Kornegay reported that supplementing weaner pig diets with sulfuric acid posed a risk of reduced feed efficiency, likely due to alterations in electrolyte balance (276). Also, it was reported that although the inclusion of phosphoric acid effectively lowered pH, it did not improve nutrient utilization (277). Moreso, Walsh et al. (278) further reported that combining inorganic and organic acids could impair growth performance while the use of only the inorganic acid blend significantly improved growth performance compared to that of antibiotics or organic acid blends.
Acidifiers can be corrosive, potentially damaging equipment and posing handling issues for feed manufacturers (20). Acidifiers, when included at excessive levels, may reduce diet palatability and lead to decreased feed intake (267). For instance, studies by Shu et al. (279) and Mao et al. (280) reported that excessive consumption of benzoic acid can cause dysfunction and damage to the liver, spleen, and lungs, as well as alterations in gut morphology. In addition, acidifiers are very volatile, as a result, commercial acidifiers are often encapsulated with fatty acids or other materials to ensure controlled release in specific intestinal compartments (281). Although research evidence supports the use of organic acids for improving nursery pig growth, there is a lack of direct economic comparisons between commercial acidifier blends and conventional antimicrobials. This knowledge gap limits producers' ability to make well-informed, science-based decisions regarding their adoption in livestock production management (282).
4.7 Enzymes
Enzymes used as feed additives are biologically active proteins that promote the chemical breakdown of nutrients into smaller compounds for further digestion and absorption. Feed additive enzymes are derived from yeast, bacteria and fungi through fermentation (283). The most commonly used enzymes are phytase, carbohydrase, xylanase, α-galactosidase, β-mannanase, α-amylase, β-glucanase, proteases, lipases, and pectinase (284). Enzymes breakdown anti-nutritional factors found in plants such as phytic acid, non-starch polysaccharides, and cell-wall complex carbohydrates and improve nutrient utilization (284). Addition of enzymes to animal feed increase gut stability, enhance substrates for beneficial fermentation, and improve the ability of the intestines to fight against bacteria pathogens (285, 286). Yu et al. (286) found that the addition of carbohydrate enzymes and protease to the diet of finishing pigs improved the growth of beneficial bacteria (Actinobacteriota, Desulfobacterota, and Lactobacillus), reduced pathogenic Clostridium_sensu_stricto_1 and improved nutrient digestibility, average daily gain and growth performance of finishing pigs. Enzymes have also been used synergistically. A study discovered that the combined use of carbohydrases, phytase and acidifiers reduced E. coli counts and increased villus length in broiler chickens (287). Similarly, supplementation of multiparous early-lactation Holstein Friesian dairy cows' diet with fibrolytic (xylanase, cellulase and β-glucanase) and amylolytic (amylase) enzymes, enhanced α-amylase and xylanase activity levels in rumen fluid, improved milk production and feed efficiency (288).
The possible mechanisms of action of in-feed enzymes include the breakdown of plants anti-nutritional substances (e.g., phytic acid and non-starch polysaccharide); improve nutrient availability by degrading the cell-wall of complex polysaccharides; improve nutrient digestion of young animals having less developed digestive system; reduce feed costs and improve feed efficiency. Furthermore, in-feed enzymes exhibit antimicrobial effects by directly hydrolyzing bacterial cell walls or compromising the integrity of glycocalyx's (289).
The main caveats associated with the use of in-feed enzymes include the lack of proper understanding on inclusion rates, low enzyme activity, the dependence of enzyme function on diet composition, ambient temperature and pH, their susceptibility to inactivation by the high levels of gut acidity and low production and quality control standards (290). The beneficial effect of enzyme inclusion is sometimes inconsistent because of different diet composition, animal genetic variations and type of enzymes used (289). The effects of in-feed enzymes are variable on animal growth performance. Another limitation is the concern about the ability of hydrolytic enzymes to survive processing temperature and the animal digestive tract.
4.8 Clay minerals
Clay minerals (e.g., bentonite, sericite, Biotite V, zeolite, kaolin etc.) have stratified structures and as a result possess great adsorption capacity enabling them to bind aflatoxins, heavy metals, enterotoxins, pathogens, and plant metabolites (248). Clay minerals such as bentonite, zeolite and montmorillonite are characterized by their negatively charged and high surface areas, swelling ability, pore volume, and high cation exchange capacity. Moreover, mineral adsorbents such as hydrated sodium calcium aluminosilicate can bind or adsorb mycotoxins to their interlayer spaces, external surface and edges. These properties cause reductions in microbial metabolites, toxins, and enzymes in the intestine; thereby preventing irritation and damage and improving performance and morphological characteristics of the intestinal mucosa (291). Therefore, there is growing interest in the use of clay minerals due to their beneficial characteristics, absence of primary toxicity and ability to reduce animal diseases and improve animal production and safety of animal products.
The feeding of clay minerals to livestock has resulted to reduction in diarrhea, better feed conversion ratio, and improved health (292, 293). Clay minerals have been reported to fully or partly mitigate the toxic effects of mycotoxins in farm animals fed with contaminated diets (294, 295). The antibacterial mechanism of medicinal clays involves the release of soluble ferrous iron and aluminum ions (Fe2+ and Al3+) that attack cellular systems in pathogens (296). The antibacterial effectiveness of clays is influenced by their mineralogical variables, such as pH and Eh (redox potential) conditions (297). Clay minerals have also been used as drug carriers for antibiotics, demonstrating their potential in healthcare applications (298). In animals, the inclusion of 5 g/kg hydrated aluminosilicate into broiler diets significantly increased body weight gain of broilers at 1 and 3 weeks of age and increased the activity of serum amylase and lactate dehydrogenase (299).
The use of clay minerals has several limitations including incomplete understanding of the bacteriostatic mode of action (293). More so, when clay minerals are added to feed at high concentrations, they can bind to other nutrients like vitamins and trace elements, causing nutritional imbalance for animals (293). There can be negative effects induced in farm animals due to interaction with veterinary substances and micronutrients in feed as well as in vitro and in vivo toxicity of natural and modified mineral adsorbents. Davis et al. (300) reported that high adsorption capacity of thermally processed clays could lead to unintended adsorption of essential vitamins and trace minerals along with targeted biotoxins. Clay minerals utilization can lead to contamination of livestock feed (301).
4.9 Heavy metals and rare earth elements
Heavy metals (copper, zinc, iron, selenium, manganese, etc.) are known as trace minerals in animal nutrition and are mainly used to improve animal health and physiological functions (302). Rare earth elements are a group of 17 elements, including 14 chemical elements from group 3A of the periodic table called lanthanoids (atomic numbers 58–71) and three members in group 3B, namely scandium (Sc; atomic number 21), yttrium (Y; atomic number 39) and lanthanum (La; atomic number 57) (303).
Heavy metals are commonly used as in-feed supplements and they play significant roles in improving digestibility, physiological, and biosynthetic processes (285, 304). Zinc is known to play essential roles in cell proliferation, immune response, reproduction, gene regulation, and defense against oxidative stress and damage (305–307). For example, there was significant improvement in body weight gain of broilers supplemented with 80 mg/kg of zinc (308). Yusof et al. (309) observed that supplementation of broiler diet with 100 mg/kg of zinc oxide nanoparticles increased body weight gain and reduced pathogenic bacteria count (Enterococcus spp. and E. coli) without affecting the beneficial bacteria in the GIT. The growth-promoting effects of copper and the mechanisms of action have been reviewed recently (310). Divalent copper(II) sulfate (CuSO4) and monovalent copper oxide (Cu2O) are commonly used as growth promoter in pig (304, 311) and poultry (312).
A plethora of investigations have reported on the use of rare earth elements as alternative to antibiotics use in livestock production (303, 313, 314). For example, Xu et al. provide insights into the antibacterial activity of specific rare-earth ions (Yb3+, Gd3+, Sm3+, Tb3+, and La3+) against S. aureus and Pseudomonas aeruginosa, demonstrating their efficacy against both planktonic and biofilm-forming bacteria. The efficacy of the rare earth elements were compared to that of traditional agents like Cu2+, Ag+, and the antibiotic norfloxacin. However, while higher concentrations of rare-earth ions were required to achieve similar antibacterial effects, they exhibited significantly lower cytotoxicity toward mammalian cells. Notably, Gd3+ demonstrated potent bactericidal activity against both planktonic and biofilm bacteria with the lowest cytotoxicity, and all tested rare-earth ions showed excellent antifungal activity against Candida albicans, suggesting their potential as safer alternatives for developing novel antimicrobial agents (315).
Despite the benefits of using heavy metals and rare earth elements in livestock production, their application still faces certain limitations. For example, it has been observed that dietary supplementation of animal diets with heavy metals (e.g., Cu and Zn) can cause an increase in antibiotic resistance through co-selection and mobilization of ARGs (311). ARGs and metal resistance genes (MRGs) are genetically linked (i.e., co-resistance) if the resistance mechanism is the same with the resistance to both metals and antibiotics (i.e. cross-resistance), or if the expression of resistance systems to both metals and antibiotics is controlled by the same factor (i.e., co-regulation), these can lead to occurrence of co-selection (311, 316). Furthermore, inclusion of high dose of heavy metals can affect the absorption of essential trace elements and cause heavy metal residues, and it can be detrimental to the environment due to their accumulation in soil and surface water (308, 317). Likewise, excess dose of heavy metals could lead to immunosuppression, impaired health and negative effects on reproductive performance in livestock (318).
Moreover, the effectiveness of rare earth elements can vary depending on several factors, including bioavailability, the specific rare earth element compound used, the animal species, and the dosage (319). As cations, rare earth elements can interact with anions, potentially affecting their absorption and biological activity (319). The form in which the rare earth element is administered can influence its efficacy. For example, He et al. (319) reported that broilers supplemented with rare earth element citrate (an organic form) at a low dose of 70 mg/kg showed significantly improved growth performance, whereas the chloride form had no effect on their growth performance. Likewise, differences in animal species affect the effectiveness of rare earth elements (320). Therefore, further research is required to elucidate the underlying mechanisms of the effects of rare earth elements and to establish optimal use in livestock production.
5 Economics and the One Health implication of the use of alternatives to antibiotics in animal production
The use of effective and eco-responsible alternatives to antibiotics in agriculture will not only have a positive impact on industry and animals, but also on the environment and society in general. The economic value of livestock production without the use of antimicrobials is influenced by many factors including input and output prices, risk aversion, and profit variance. Other situations and factors include farmers' perceptions of costs and risks, and limited access to cheap sources of inputs (321, 322). Therefore, understanding these factors can help identify pathways to reduce antimicrobial use. However, while reducing antimicrobial use is crucial, there's no global consensus on acceptable levels, and a complete ban could have adverse effects on animal health and welfare (321). Many cost-effective antimicrobial alternatives are still experimental, requiring further research. Despite the lack of consensus, reducing antimicrobial use is widely agreed upon, and this serves as a basis for developing effective policies (322). Utilizing interdisciplinary systemic approaches facilitates the development of AMR policies and strategies that are feasible across technical, political, economic, and behavioral dimensions. This approach enables the identification of various key factors, including all factors influencing antimicrobial use in livestock production, the power dynamics between these factors, appropriate regulatory frameworks and interventions, optimal behavioral change strategies, and the responsible entities for implementation (323). Additionally, it allows for the cost-effective assessment of intervention combinations. However, AMR policies and strategies are often explored within different disciplines, lacking a holistic and systemic perspective. Therefore, advocating for more interdisciplinary collaboration is crucial to addressing the use of antimicrobials in livestock production and the emergence of AMR effectively, and further research opportunities in this area is warranted.
From an economic standpoint, antimicrobial use in livestock production generates negative externalities (e.g., development of AMR and the impact of AMR on the environment and society), and assessing the total economic value of antimicrobial use should consider economic losses due to the impact of AMR in the One Health continuum. This implies that the economic impact of AMR extends beyond increased healthcare costs and treatment failures in humans to include interruptions in agricultural productivity and international trade. For example, zoonotic disease outbreaks caused by multidrug-resistant pathogens, such as Salmonella and avian influenza have resulted to economic losses to the tune of billions of US dollars due to culling, decreased consumer confidence in livestock-derived products and trade restrictions (324). In dairy production, multidrug-resistant S. aureus strains causing mastitis significantly impacts milk production, leading to economic losses and increased dependance on antimicrobials for treatment (325). Also, many studies have reported that many commonly used intramammary antibiotic formulations contain critically important antimicrobial classes, such as aminoglycosides and cephalosporins, which are crucial for human medicine (326). The frequent use of these antibiotics not only accelerates resistance development but also increases the risk of economic loses and the presence of antimicrobial residues in dairy products, further complicating efforts to mitigate AMR.
Despite risks posed by AMR, many farmers, veterinarians and pharmaceutical companies continue to consider antimicrobials as essential components of livestock production, by prioritizing production efficiency and economic gains over the responsible use of antimicrobials. Recent surveys of dairy farmers in Scotland and Switzerland have identified key demographic and behavioral predictors of antimicrobial use trends, with younger farmers and those managing larger herds being more likely to engage in high antimicrobial usage as well as the tendency of farmers to rely on peer recommendations rather than veterinary guidance or self-prescribe antibiotics, which further complicates efforts to mitigate AMR (325, 326). Moreover, veterinarians as key decision-makers in antimicrobial use also face significant challenges in promoting antimicrobial stewardship as they are often pressured to prescribe antimicrobials based on farmers' preferences rather than scientific evidence (326). Despite the threat of AMR, a 2019 investigation by the New York Times found that Elanco, an animal pharma company, created a brochure that encouraged farmers to treat whole herds of pigs with antibiotics to prevent disease (327).
The challenges of limiting antimicrobial use in livestock production are particularly obvious in regions with limited veterinary oversight and weak regulatory frameworks (328). Therefore, addressing these issues requires the implementation of sustainable practices including the use of antimicrobial alternatives, enhanced antibiotic stewardship, strengthened interdisciplinary collaboration and emphasizing the critical need for a One Health approach by recognizing the interconnectedness of human, animal, and environmental health in safeguarding global public health and mitigating AMR (329). Such an approach is valuable for developing effective strategies for combating this common foe (AMR), including strengthening biosecurity measures, education of farmers on the implication of AMR and emphasizing responsible antimicrobial usage for curative purpose only, while eliminating use in growth promotion.
6 Research gaps, future perspectives and conclusion
Finding functional and more sustainable alternatives to antibiotics as growth promoters is expected to increase the wide range of different alternatives to be included in further research. Although only few available vaccines can effectively control disease occurrence and eliminate the therapeutic use of antimicrobials, and while few/no alternatives have emerged to replace antibiotics as growth promoters in livestock production, the options discussed in previous sections hold significant value that can be integrated into practical “no antimicrobial use as growth promoters” in livestock production programs. This involves strengthening collaborations with local farmers and animal husbandry practitioners to ensure real-world applicability of research findings; engaging stakeholders through international conferences and agricultural forums to raise awareness and foster interdisciplinary dialogue; promoting public-private partnerships and community-based projects to secure funding and logistical support for field trials; integrating knowledge-sharing platforms to disseminate best practices and align local efforts with global One Health goals. However, the use of most of these alternatives is still limited due to reliability, species-specificity, costs and complexity of production.
Currently, the mechanism of action of most antimicrobial alternatives are not known. For example, the intestinal physiology and microbiota of different breeds may affect the effectiveness of an alternative, be it probiotics or other alternatives presented above. It should be noted that the GIT consist of a wide array of nutritional and physicochemical environments which may impact the colonization and survival of supplemental probiotics. Moreover, there is a lack of understanding of the nature, function, and role of biofilm-forming microbial communities in pathophysiology and animal health. Considering that biofilm growth is common in GIT diseases, developing alternatives to antibiotics should take into consideration and explore how antibiotic alternatives in animal feed can affect biofilm growth and survival. The synergistic effect of probiotics and prebiotics on host physiology, the immune response and optimal ratios and rate of inclusion in livestock feed deserves scrutiny. The interactions between the microbiota, host, and feed components are not fully understood, which hampers their widespread use. Additionally, the potential for resistance development and co- and cross-resistance with antibiotics is a concern (20). To achieve maximum results, it is important to determine the exact dosage for each breed and species. In addition, microbial alternatives to AGPs should be broadly applicable, have low adverse effect levels, generally recognized as safe to livestock and the environment, and pose no threat to AMR development.
Understanding host-pathogen interactions and immune response to pathogens will improve the development of precise and personalized immunotherapies. Likewise, developing effective combination of immunotherapies will lead to synergistic effects and enhanced outcomes for disease prevention and treatment, following thorough evaluation and optimization of dosages, timing, and interactions between the components. For example, the use of AMPs shows promising positive impact on the host immune systems, however, more studies are needed to improve their use in animal production, and to properly understand their synergistic effect with other AGPs. The use of nanotechnology-based approaches, such as nanoparticle vaccines, can enhance the efficacy of vaccines and enable targeted delivery of immunotherapeutic agents to the site of infection, thereby enhancing their effectiveness and minimizing their side effects. Combination of different omics technologies, such as genomics, epigenomics, transcriptomics, etc. could provide important insights into the host immune response to livestock disease pathogens and the development of effective immunotherapies.
Antimicrobial alternatives delivered nutritionally (feed and water) pose numerous challenges needing concerted efforts to address them. Regarding phytochemicals, there is lack of understanding of the biochemical route or mechanism of action by herbs and spices, which calls for further research to understand their mechanisms of action and their rate of inclusion in livestock diets. Offensive characteristics (bad odors, variations in product compositions, high volatility, etc.) limits the application of essential oils in animal nutrition. Therefore, deeper understanding on their mechanism of action, optimal dosage and duration regime, and mode of administration is warranted to improve their efficiency and safety. Further research is also needed to optimize the delivery of essential oils into the lower GIT using microencapsulation and nanotechnology, understand the potential synergistic effects of combining different essential oils and other compounds, and dissect the mechanisms underlying the functions of essential oils using high-throughput systems technologies (19). Additionally, the potential for essential oils to interact with feed and the gut ecosystem of animals requires further investigation (330). Deep knowledge of organic acids and in-feed enzymes mode of action and potential interactions with other feed additives or medications is needed to fully understand their application as alternatives to AGPs in livestock production. Similarly, further studies are needed to certify the effectiveness and safety of utilizing clay minerals effectively in livestock nutrition while minimizing potential adverse effects, and to elucidate the safe use of heavy metals in animal nutrition.
While the antimicrobial alternatives discussed above present potential as replacements for antimicrobials in livestock production; perhaps, there should be a rethink of the goals for the search of effective AGP alternatives. For example, instead of searching for products with similar mode of action to antibiotics, the focus should be directed at products with ability to improve productivity and enhance animal health with the consequence of reduced need for therapeutic and sub-therapeutic antibiotics use. To focus in this direction, the mode of action of AGPs in health and production enhancement must be understood and management practices tailored to keep-away disease causing pathogens. However, AGP function is more complex than merely modulating microbial populations and functions, and their mode of action may be through direct or indirect modulation of the host immune system and consequently the immune response. Therefore, the key to finding new effective AGP alternatives lies in knowledge of the mechanisms associated with their action, the realization that no alternative can elicit the same mechanisms of action ascribed to AGPs and a focus on a combination of strategies (e.g., combine two or more AGP alternatives, AGP alternative and management practices, etc.) to achieve desired improvements without AGP. Moreover, a One Health approach should be applied to the search for AGP alternatives, with the aim to develop strategies that could simultaneously address animal health, human health, and the overall health of the ecosystem. A comprehensive understanding of these future perspectives will lead to significant advancements in the search for AGP alternatives for addressing the prevention, treatment, and control of livestock diseases, for the benefit of the livestock industry and animal and human welfare.
In conclusion, there are currently few or no alternative products that could completely replace antibiotics as a growth promoter in livestock production. However, the synergistic use of different alternatives could yield optimum breakthroughs in this field. While utilizing a combination of alternatives to antibiotics appears to be a promising strategy to combat drug resistance, it is not without its drawbacks. As we continue to strive toward reducing the use of antibiotics as growth promoters, it is crucial to give special attention to sustainable manufacturing, environmental impact, the likelihood of resistance development, the genetics of resistance evolution, and the risk of antibiotic cross-resistance. These factors should be carefully considered during the development and regulation of alternatives to antibiotics in livestock production.
Author contributions
EMI-A: Writing – review & editing, Methodology, Funding acquisition, Supervision, Data curation, Writing – original draft, Resources, Conceptualization, Project administration. FAO: Writing – review & editing, Writing – original draft. LCP: Writing – original draft, Writing – review & editing. ATV: Funding acquisition, Resources, Writing – review & editing, Supervision, Data curation, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The funding for this work was provided by Agriculture and Agri-Food Canada.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Aminov RI. A brief history of the antibiotic era: lessons learned and challenges for the future. Front Microbiol. (2010) 1:134. doi: 10.3389/fmicb.2010.00134
2. Brown K, Uwiera RRE, Kalmokoff ML, Brooks SPJ, Inglis GD. Antimicrobial growth promoter use in livestock: a requirement to understand their modes of action to develop effective alternatives. Int J Antimicrob Agents. (2017) 49:12–24. doi: 10.1016/j.ijantimicag.2016.08.006
3. Castanon JI. History of the use of antibiotic as growth promoters in European poultry feeds. Poult Sci. (2007) 86:2466–71. doi: 10.3382/ps.2007-00249
4. Chattopadhyay MK. Use of antibiotics as feed additives: a burning question. Front Microbiol. (2014) 5:334. doi: 10.3389/fmicb.2014.00334
5. Hao H, Cheng G, Iqbal Z, Ai X, Hussain HI, Huang L, et al. Benefits and risks of antimicrobial use in food-producing animals. Front Microbiol. (2014) 5:288. doi: 10.3389/fmicb.2014.00288
6. Stoica C, Cox G. Old problems and new solutions: antibiotic alternatives in food animal production. Can J Microbiol. (2021) 67:427–44. doi: 10.1139/cjm-2020-0601
7. Kirchhelle C. Pharming animals: a global history of antibiotics in food production (1935–2017). Palgrave Commun. (2018) 4:96. doi: 10.1057/s41599-018-0152-2
8. Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP, et al. Global trends in antimicrobial use in food animals. Proc Natl Acad Sci USA. (2015) 112:5649–54. doi: 10.1073/pnas.1503141112
9. Van Boeckel TP, Pires J, Silvester R, Zhao C, Song J, Criscuolo NG, et al. Global trends in antimicrobial resistance in animals in low- and middle-income countries. Science. (2019) 365:aaw1944. doi: 10.1126/science.aaw1944
10. WHO. Global Action Plan on Antimicrobial Resistance. Geneva: World Health Organization (WHO) (2015). Available online at: https://www.who.int/publications/i/item/9789241509763 (Accessed March 13, 2024).
11. WHO. Integrated Surveillance of Antimicrobial Resistance in Foodborne Bacteria. Geneva: World Health Organization (WHO) (2017). Available online at: http://www.who.int/foodsafety/publi-cations/agisar_guidance2017/en/ (Accessed March 13, 2024).
12. FAO. The Food and Agricultural Organization (FAO) Action Plan on Antimicrobial Resistance. Rome: Food and Agriculture Organization of the United Nations (2016). p. 3–25. Available online at: http://www.fao.org/3/a-i5996e.pdf (Accessed January 01, 2023).
13. G20. G20 Call to Action on Antimicrobial Resistance. (2022). Available online at: https://g7g20-documents.org/fileadmin/G7G20_documents/2022/G20/Indonesia/Sherpa-Track/Health%20Ministers/2%20Ministers%27%20Annex/G20%20Call%20to%20Action%20on%20Antimicrobial%20Resistance_28102022.pdf (Accessed June 20, 2024).
14. PHAC. Pan-Canadian Action Plan on Antimicrobial Resistance. Public Health Agency of Canada (PHAC) Report. (2023). Available online at: https://www.canada.ca/en/public-health/services/publications/drugs-health-products/pan-canadian-action-plan-antimicrobial-resistance.html (Accessed June 06, 2024).
15. Magouras I, Carmo LP, Stärk KDC, Schüpbach-Regula G. Antimicrobial usage and -resistance in livestock: where should we focus? Front Vet Sci. (2017) 4:148. doi: 10.3389/fvets.2017.00148
16. Queenan K, Häsler B, Rushton J. A One Health approach to antimicrobial resistance surveillance: is there a business case for it? Int J Antimicrob Agents. (2016) 48:422–7. doi: 10.1016/j.ijantimicag.2016.06.014
17. CAC. Codex Alimenatrius Commission. Antimicrobial Resistance and the Use of Antimicrobials in Animal Production (CX/RVDF 00/4). (2000). Available online at: https://ec.europa.eu/food/safety/international_affairs/standard_setting_bodies/codex/ccrdvf/archives_en (Accessed January 02, 2023).
18. Seal BS, Lillehoj HS, Donovan DM, Gay CG. Alternatives to antibiotics: a symposium on the challenges and solutions for animal production. Anim Health Res Rev. (2013) 14:78–87. doi: 10.1017/S1466252313000030
19. Omonijo FA, Ni L, Gong J, Wang Q, Lahaye L, Yang C. Essential oils as alternatives to antibiotics in swine production. Anim Nutr. (2018) 4:126–36. doi: 10.1016/j.aninu.2017.09.001
20. Rahman MRT, Fliss I, Biron E. Insights in the development and uses of alternatives to antibiotic growth promoters in poultry and swine production. Antibiotics. (2022) 11:766. doi: 10.3390/antibiotics11060766
22. Jukes TH. Some historical notes on chlortetracycline. Rev Infect Dis. (1985) 7:702–7. doi: 10.1093/clinids/7.5.702
23. Finlay MR, Marcus AI. Consumerist terrorists': battles over agricultural antibiotics in the United States and Western Europe. Agric Hist. (2016) 90:146–72. doi: 10.3098/ah.2016.090.2.146
24. Hughes P, Heritage J. Antibiotic Growth-promoters in Food Animals. Rome Food and Agriculture Organization of the United Nations (FAO) (2004). p. 152. Available online at: https://www.fao.org/4/y5159e/y5159e08.htm (Accessed February 12, 2023).
25. Bosman AL, Deckert AE, Carson CA, Poljak Z, Reid-Smith RJ, McEwen SA. Antimicrobial use in lactating sows, piglets, nursery, and grower-finisher pigs on swine farms in Ontario, Canada during 2017 and 2018. Porcine Health Manage. (2022) 8:17. doi: 10.1186/s40813-022-00259-w
26. Ruegg PL. Realities, challenges and benefits of antimicrobial stewardship in dairy practice in the United States. Microorganisms. (2022) 10:1626. doi: 10.3390/microorganisms10081626
27. Tiseo K, Huber L, Gilbert M, Robinson TP, Van Boeckel TP. Global trends in antimicrobial use in food animals from 2017 to 2030. Antibiotics. (2020) 9:918. doi: 10.3390/antibiotics9120918
28. Topi D, Spahiu J. Presence of veterinary antibiotics in livestock manure in two Southeastern Europe countries, Albania and Kosovo. Environ Sci Pollut Res. (2020) 27:44552–60. doi: 10.1007/s11356-020-10341-x
29. Van den Meersche T, Rasschaert G, Vanden Nest T, Haesebrouck F, Herman L, Van Coillie E, et al. Longitudinal screening of antibiotic residues, antibiotic resistance genes and zoonotic bacteria in soils fertilized with pig manure. Environ Sci Pollut Res Int. (2020) 27:28016–29. doi: 10.1007/s11356-020-09119-y
30. Kasimanickam V, Kasimanickam M, Kasimanickam R. Antibiotics use in food animal production: escalation of antimicrobial resistance: where are we now in combating AMR? Med Sci. (2021) 9:14. doi: 10.3390/medsci9010014
31. FDA. Federal Department of Agriculture (FDA) Releases Annual Summary Report on Antimicrobials Sold or Distributed in 2021 for use in food-producing animals. (2021). Available online at: https://www.fda.gov/animal-veterinary/cvm-updates/fda-releases-annual-summary-report-antimicrobials-sold-or-distributed-2021-use-food-producing (Accessed January 06, 2023).
32. Odey TOJ, Tanimowo WO, Afolabi KO, Jahid IK, Reuben RC. Antimicrobial use and resistance in food animal production: food safety and associated concerns in Sub-Saharan Africa. Int Microbiol. (2024) 27:1–23. doi: 10.1007/s10123-023-00462-x
33. Van TTH, Yidana Z, Smooker PM, Coloe PJ. Antibiotic use in food animals worldwide, with a focus on Africa: pluses and minuses. J Glob Antimicrob Resist. (2020) 20:170–7. doi: 10.1016/j.jgar.2019.07.031
34. Naranjo-Lucena A, Slowey R. Invited review: antimicrobial resistance in bovine mastitis pathogens: a review of genetic determinants and prevalence of resistance in European countries. J Dairy Sci. (2023) 106:1–23. doi: 10.3168/jds.2022-22267
35. Gibbs EP. The evolution of One Health: a decade of progress and challenges for the future. Vet Rec. (2014) 174:85–91. doi: 10.1136/vr.g143
36. Massé DI, Saady NM, Gilbert Y. Potential of biological processes to eliminate antibiotics in livestock manure: an overview. Animals. (2014) 4:146–63. doi: 10.3390/ani4020146
37. Sun J, Liao X-P, D'Souza AW, Boolchandani M, Li S-H, Cheng K, et al. Environmental remodeling of human gut microbiota and antibiotic resistome in livestock farms. Nat Commun. (20202) 11:1427. doi: 10.1038/s41467-020-15222-y
38. Sun J, Huang T, Chen C, Cao T-T, Cheng K, Liao X-P, et al. Comparison of fecal microbial composition and antibiotic resistance genes from swine, farm workers and the surrounding villagers. Sci Rep. (2017). 7:4965. doi: 10.1038/s41598-017-04672-y
39. Bai H, He L-Y, Wu D-L, Gao F-Z, Zhang M, Zou H-Y, et al. Spread of airborne antibiotic resistance from animal farms to the environment: dispersal pattern and exposure risk. Environ Int. (2022) 158:106927. doi: 10.1016/j.envint.2021.106927
40. Monger XC, Gilbert AA, Saucier L, Vincent AT. Antibiotic resistance: from pig to meat. Antibiotics. (2021) 10:1209. doi: 10.3390/antibiotics10101209
41. Zalewska M, Błażejewska A, Czapko A, Popowska M. Antibiotics and antibiotic resistance genes in animal manure - consequences of its application in agriculture. Front Microbiol. (2021) 12:610656. doi: 10.3389/fmicb.2021.610656
42. Hammer TJ, Fierer N, Hardwick B, Simojoki A, Slade E, Taponen J, et al. Treating cattle with antibiotics affects greenhouse gas emissions, and microbiota in dung and dung beetles. Proc R Soc B Biol Sci. (2016) 283:20160150. doi: 10.1098/rspb.2016.0150
43. Manyi-Loh C, Mamphweli S, Meyer E, Okoh A. Antibiotic use in agriculture and its consequential resistance in environmental sources: potential public health implications. Molecules. (2018) 23:795. doi: 10.3390/molecules23040795
44. Berg G, Rybakova D, Fischer D, Cernava T, Vergès M-CC, Charles T, et al. Microbiome definition re-visited: old concepts and new challenges. Microbiome. (2020) 8:103. doi: 10.1186/s40168-020-00875-0
45. Dhanaraju R, Rao DN. The human microbiome: an acquired organ? Resonance. (2022) 27:247–72. doi: 10.1007/s12045-022-1312-7
46. Couvillion SP, Mostoller KE, Williams JE, Pace RM, Stohel IL, Peterson HK, et al. Interrogating the role of the milk microbiome in mastitis in the multi-omics era. Front Microbiol. (2023) 14:1105675. doi: 10.3389/fmicb.2023.1105675
47. Jessop E, Li L, Renaud DL, Verbrugghe A, Macnicol J, Gamsjäger L, et al. Neonatal calf diarrhea and gastrointestinal microbiota: etiologic agents and microbiota manipulation for treatment and prevention of diarrhea. Vet Sci. (2024) 11:108. doi: 10.3390/vetsci11030108
48. Verbeek E, Keeling L, Landberg R, Lindberg JE, Dicksved J. The gut microbiota and microbial metabolites are associated with tail biting in pigs. Sci Rep. (2021) 11:20547. doi: 10.1038/s41598-021-99741-8
49. Gaeta NC, Lima SF, Teixeira AG, Ganda EK, Oikonomou G, Gregory L, et al. Deciphering upper respiratory tract microbiota complexity in healthy calves and calves that develop respiratory disease using shotgun metagenomics. J Dairy Sci. (2017) 100:1445–58. doi: 10.3168/jds.2016-11522
50. Khafipour E, Li S, Plaizier JC, Krause DO. Rumen microbiome composition determined using two nutritional models of subacute ruminal acidosis. Appl Environ Microbiol. (2009) 75:7115–24. doi: 10.1128/AEM.00739-09
51. Plaizier JC, Li S, Tun HM, Khafipour E. Nutritional models of experimentally-induced subacute ruminal acidosis (sara) differ in their impact on rumen and hindgut bacterial communities in dairy cows. Front Microbiol. (2016) 7:2128. doi: 10.3389/fmicb.2016.02128
52. Muritala I, Bemji MN, Busari MA, Sodimu BO, Ahmad SM, Negi A, et al. Nasal microbiome diversity in West African Dwarf goats with peste des petits ruminants viral infection. In: 39th International Society for Animal Genetics Conference, 2-7 July 2023, Cape Town, South Africa. ISAG 2023 Abstract Book. International Society for Animal Genetics (2023). p. 10–11. Available online at: https://www.isag.us/Docs/Proceedings/ISAG_2023_Abstracts.pdf
53. Schokker D, Zhang J, Zhang LL, Vastenhouw SA, Heilig HG, Smidt H, et al. Early-life environmental variation affects intestinal microbiota and immune development in new-born piglets. PLoS ONE. (2014) 9:e100040. doi: 10.1371/journal.pone.0100040
54. Jo HE, Kwon MS, Whon TW, Kim DW, Yun M, Lee J, et al. Alteration of gut microbiota after antibiotic exposure in finishing swine. Front Microbiol. (2021) 12:596002. doi: 10.3389/fmicb.2021.596002
55. Fernández Miyakawa ME, Casanova NA, Kogut MH. How did antibiotic growth promoters increase growth and feed efficiency in poultry? Poult Sci. (2024) 103:103278. doi: 10.1016/j.psj.2023.103278
56. Plata G, Baxter NT, Susanti D, Volland-Munson A, Gangaiah D, Nagireddy A, et al. Growth promotion and antibiotic induced metabolic shifts in the chicken gut microbiome. Commun Biol. (2022) 5:293. doi: 10.1038/s42003-022-03239-6
57. Saleem A, Saleem Bhat S, Omonijo FA, Ganai NA, Ibeagha-Awemu EM, Mudasir Ahmad S. Immunotherapy in mastitis: state of knowledge, research gaps and way forward. Vet Q. (2024) 44:1–23. doi: 10.1080/01652176.2024.2363626
58. Coates ME, Fuller R, Harrison GF, Lev M, Suffolk SF. A comparison of the growth of chicks in the Gustafsson germ-free apparatus and in a conventional environment, with and without dietary supplements of penicillin. Br J Nutr. (1963) 17:141–50. doi: 10.1079/BJN19630015
59. Wang JW, Kuo CH, Kuo FC, Wang YK, Hsu WH, Yu FJ, et al. Fecal microbiota transplantation: review and update. J Formos Med Assoc. (2019) 118:S23–31. doi: 10.1016/j.jfma.2018.08.011
60. Canibe N, O'Dea M, Abraham S. Potential relevance of pig gut content transplantation for production and research. J Anim Sci Biotechnol. (2019) 10:1–19. doi: 10.1186/s40104-019-0363-4
61. Guo H, Cui J, Li Q, Liang X, Li J, Yang B, et al. A Multi-omic assessment of the mechanisms of intestinal microbes used to treat diarrhea in early-weaned lambs. mSystems. (2024) 9:e0095323. doi: 10.1128/msystems.00953-23
62. Kim HS, Whon TW, Sung H, Jeong Y-S, Jung ES, Shin N-R, et al. Longitudinal evaluation of fecal microbiota transplantation for ameliorating calf diarrhea and improving growth performance. Nat Commun. (2021) 12:161. doi: 10.1038/s41467-020-20389-5
63. Nowland TL, Kirkwood RN. Faecal microbiota transplantation: is it the future for pig production? Microbiol Aust. (2020) 41:91–4. doi: 10.1071/MA20023
64. Wu D, Liang S, Du X, Xiao J, Feng H, Ren Z, et al. Effects of fecal microbiota transplantation and fecal virome transplantation on LPS-induced intestinal injury in broilers. Poult Sci. (2024) 103:103316. doi: 10.1016/j.psj.2023.103316
65. Lundberg R, Toft M, August B, Kornerup Hansen A, Hansen C. Antibiotic-treated versus germ-free rodents for microbiota transplantation studies. Gut Microbes. (2016) 7:68–74. doi: 10.1080/19490976.2015.1127463
66. McCormack UM, Curião T, Wilkinson T, Metzler-Zebeli BU, Reyer H, Ryan T, et al. Fecal microbiota transplantation in gestating sows and neonatal offspring alters lifetime intestinal microbiota and growth in offspring. mSystems. (2018) 3:e00134-17. doi: 10.1128/mSystems.00134-17
67. Menchaca A, Dos Santos-Neto P, Mulet A, Crispo M. CRISPR in livestock: from editing to printing. Theriogenology. (2020) 150:247–54. doi: 10.1016/j.theriogenology.2020.01.063
68. Sato M, Miyoshi K, Kawaguchi H, Inada E, Saitoh I, Tanimoto A. Recent advance in genome editing-based gene modification in pigs. In:Gardón Poggi JC, Ambrojo KS, , editors. Reproductive Biology and Technology in Animals. London: IntechOpen (2019). doi: 10.5772/intechopen.88022
69. Yi D, Zhou S-W, Qiang D, Bei C, Zhao X-E, Zhong S, et al. The CRISPR/Cas9 induces large genomic fragment deletions of MSTN and phenotypic changes in sheep. J Integr Agric. (2020) 19:1065–73. doi: 10.1016/S2095-3119(19)62853-4
70. de la Fuente-Núñez C, Lu TK. CRISPR-Cas9 technology: applications in genome engineering, development of sequence-specific antimicrobials, and future prospects. Integr Biol. (2017) 9:109–22. doi: 10.1039/c6ib00140h
71. Bikard D, Euler CW, Jiang W, Nussenzweig PM, Goldberg GW, Duportet X, et al. Exploiting CRISPR-Cas nucleases to produce sequence-specific antimicrobials. Nat Biotechnol. (2014) 32:1146–50. doi: 10.1038/nbt.3043
72. Citorik RJ, Mimee M, Lu TK. Sequence-specific antimicrobials using efficiently delivered RNA-guided nucleases. Nat Biotechnol. (2014) 32:1141–5. doi: 10.1038/nbt.3011
73. Van Eenennaam AL, De Figueiredo Silva F, Trott JF, Zilberman D. Genetic engineering of livestock: the opportunity cost of regulatory delay. Ann Rev Anim Biosci. (2021) 9:453–78. doi: 10.1146/annurev-animal-061220-023052
74. Gao Y, Wu H, Wang Y, Liu X, Chen L, Li Q, et al. Single Cas9 nickase induced generation of NRAMP1 knockin cattle with reduced off-target effects. Genome Biol. (2017) 18:1–15. doi: 10.1186/s13059-016-1144-4
75. Polkoff K, Piedrahita JA. The transformational impact of site-specific DNA modifiers on biomedicine and agriculture. Anim Reprod. (2018) 15:171–9. doi: 10.21451/1984-3143-AR2018-0065
76. Ruan J, Xu J, Chen-Tsai RY, Li K. Genome editing in livestock: are we ready for a revolution in animal breeding industry? Transgenic Res. (2017) 26:715–26. doi: 10.1007/s11248-017-0049-7
77. Zafar I, Singh S, Kumar J. Genome editing by programmable nucleases and their applications in livestock species. J Livest Sci. (2019) 10:32–47. doi: 10.33259/JLivestSci.2019.32-47
78. Lee SW, Tran KT, Vazquez-Uribe R, Gotfredsen CH, Clausen MH, Méndez BL, et al. Identification and optimization of novel small-molecule cas9 inhibitors by cell-based high-throughput screening. J Med Chem. (2022) 65:3266–305. doi: 10.1021/acs.jmedchem.1c01834
79. Pawluk A, Amrani N, Zhang Y, Garcia B, Hidalgo-Reyes Y, Lee J, et al. Naturally occurring off-switches for CRISPR-Cas9. Cell. (2016) 167:1829–38.e1829. doi: 10.1016/j.cell.2016.11.017
80. Mehravar M, Shirazi A, Nazari M, Banan M. Mosaicism in CRISPR/Cas9-mediated genome editing. Dev Biol. (2019) 445:156–62. doi: 10.1016/j.ydbio.2018.10.008
81. Hennig SL, Owen JR, Lin J, Young A, Ross PJ, Eenennaam ALV, et al. Evaluation of mutation rates, mosaicism and off target mutations when injecting Cas9 mRNA or protein for genome editing of bovine embryos. Sci Rep. (2020) 10:22309. doi: 10.1038/s41598-020-78264-8
82. Koralesky KE, Sirovica LV, Hendricks J, Mills KE, von Keyserlingk MAG, Weary DM. Social acceptance of genetic engineering technology. PLoS ONE. (2023) 18:e0290070. doi: 10.1371/journal.pone.0290070
83. Brüssow H, Hendrix RW. Phage genomics: small is beautiful. Cell. (2002) 108:13–6. doi: 10.1016/S0092-8674(01)00637-7
84. Sulakvelidze A, Morris JG Jr. Bacteriophages as therapeutic agents. Ann Med. (2001) 33:507–9. doi: 10.3109/07853890108995959
85. Carvalho CM, Gannon BW, Halfhide DE, Santos SB, Hayes CM, Roe JM, et al. The in vivo efficacy of two administration routes of a phage cocktail to reduce numbers of Campylobacter coli and Campylobacter jejuni in chickens. BMC Microbiol. (2010) 10:232. doi: 10.1186/1471-2180-10-232
86. Wittebole X, De Roock S, Opal SM. A historical overview of bacteriophage therapy as an alternative to antibiotics for the treatment of bacterial pathogens. Virulence. (2014) 5:226–35. doi: 10.4161/viru.25991
87. Huh H, Wong S, St Jean J, Slavcev R. Bacteriophage interactions with mammalian tissue: therapeutic applications. Adv Drug Deliv Rev. (2019) 145:4–17. doi: 10.1016/j.addr.2019.01.003
88. Dion MB, Oechslin F, Moineau S. Phage diversity, genomics and phylogeny. Nat Rev Microbiol. (2020) 18:125–38. doi: 10.1038/s41579-019-0311-5
89. Gabisoniaa T, Loladzea M, Chakhunashvilib N, Tamarashvilib N, Nadiradzeb M, Katamadzeb T, et al. Bacteriophage preparation for treatment and prevention of Salmonella infection in poultry. Ann Agrar Sci. (2019) 17:36–44. Available online at: https://journals.org.ge/index.php/aans/article/view/296
90. Loc-Carrillo C, Abedon ST. Pros and cons of phage therapy. Bacteriophage. (2011) 1:111–4. doi: 10.4161/bact.1.2.14590
91. Huang Y, Wang W, Zhang Z, Gu Y, Huang A, Wang J, et al. Phage products for fighting antimicrobial resistance. Microorganisms. (2022) 10:1324. doi: 10.3390/microorganisms10071324
92. Chhibber S, Kaur T, Kaur S. Essential role of calcium in the infection process of broad-spectrum methicillin-resistant Staphylococcus aureus bacteriophage. J Basic Microbiol. (2014) 54:775–80. doi: 10.1002/jobm.201300051
93. Jung L-s, Ding T, Ahn J. Evaluation of lytic bacteriophages for control of multidrug-resistant Salmonella Typhimurium. Ann Clin Microbiol Antimicrob. (2017) 16:66. doi: 10.1186/s12941-017-0237-6
94. Żbikowska K, Michalczuk M, Dolka B. The use of bacteriophages in the poultry industry. Animals. (2020) 10:872. doi: 10.3390/ani10050872
95. Colavecchio A, Goodridge LD. Phage therapy approaches to reducing pathogen persistence and transmission in animal production environments: opportunities and challenges. Microbiol Spectr. (2017) 5:10.1128/microbiolspec.pfs-0017-2017. doi: 10.1128/microbiolspec.PFS-0017-2017
96. Huff WE, Huff GR, Rath NC, Balog JM, Donoghue AM. Prevention of Escherichia coli infection in broiler chickens with a bacteriophage aerosol spray. Poult Sci. (2002) 81:1486–91. doi: 10.1093/ps/81.10.1486
97. Huff WE, Huff GR, Rath NC, Balog JM, Donoghue AM. Bacteriophage treatment of a severe Escherichia coli respiratory infection in broiler chickens. Avian Dis. (2003) 47:1399–405. doi: 10.1637/7041
98. Huff WE, Huff GR, Rath NC, Balog JM, Donoghue AM. Evaluation of aerosol spray and intramuscular injection of bacteriophage to treat an Escherichia coli respiratory infection. Poult Sci. (2003) 82:1108–12. doi: 10.1093/ps/82.7.1108
99. Huff WE, Huff GR, Rath NC, Balog JM, Xie H, Moore PA, et al. Prevention of Escherichia coli respiratory infection in broiler chickens with bacteriophage (SPR02). Poult Sci. (2002) 81:437–41. doi: 10.1093/ps/81.4.437
100. Huff WE, Huff GR, Rath NC, Donoghue AM. Evaluation of the influence of bacteriophage titer on the treatment of colibacillosis in broiler chickens. Poult Sci. (2006) 85:1373–7. doi: 10.1093/ps/85.8.1373
101. Feng C, Jia K, Chi T, Chen S, Yu H, Zhang L, et al. Lytic bacteriophage PZL-Ah152 as biocontrol measures against lethal aeromonas hydrophila without distorting gut microbiota. Front Microbiol. (2022) 13:898961. doi: 10.3389/fmicb.2022.898961
102. Gabisonia T, Loladze M, Chakhunashvili N, Tamarashvili N, Nadiradze M, Katamadze T, et al. Use of a bacteriophage cocktail to reduce Salmonella colonization in experimentally infected chickens. Ann Agrarian Sci. (2019) 17:397–402. Available online at: https://journals.org.ge/index.php/aans/article/view/322
103. Malik DJ. Bacteriophage encapsulation using spray drying for phage therapy. Curr Issues Mol Biol. (2021) 40:303–16. doi: 10.21775/cimb.040.303
104. Malik DJ, Sokolov IJ, Vinner GK, Mancuso F, Cinquerrui S, Vladisavljevic GT, et al. Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. Adv Colloid Interface Sci. (2017) 249:100–33. doi: 10.1016/j.cis.2017.05.014
105. Oliveira A, Sereno R, Azeredo J. In vivo efficiency evaluation of a phage cocktail in controlling severe colibacillosis in confined conditions and experimental poultry houses. Vet Microbiol. (2010) 146:303–8. doi: 10.1016/j.vetmic.2010.05.015
106. Zhang J, Xu H, Yang H, Li J, Xiao S, Hu S, et al. Screening of a Plesiomonas shigelloides phage and study of the activity of its lysis system. Virus Res. (2021) 306:198581. doi: 10.1016/j.virusres.2021.198581
107. Borie C, Albala I, Sànchez P, Sánchez ML, RamÍrez S, Navarro C, et al. Bacteriophage treatment reduces Salmonella colonization of infected chickens. Avian Dis. (2008) 52:64–7. doi: 10.1637/8091-082007-Reg
108. Miller RW, Skinner J, Sulakvelidze A, Mathis GF, Hofacre CL Bacteriophage therapy for control of Necrotic Enteritis of broiler chickens experimentally infected with Clostridium perfringens. Avian Dis. (2010) 54:33–40, 38. doi: 10.1637/8953-060509-Reg.1
109. Schmoeller E, Matos ADC, Rahal NM, Feijo JO, Brauner CC, Pino FABD, et al. Diarrhea duration and performance outcomes of pre-weaned dairy calves supplemented with bacteriophage. Can J Anim Sci. (2022) 102:165–74. doi: 10.1139/cjas-2021-0074
110. Smith HW, Huggins MB, Shaw KM. The control of experimental Escherichia coli diarrhea in calves by means of bacteriophages. Microbiology. (1987) 133:1111–26. doi: 10.1099/00221287-133-5-1111
111. Chan BK, Abedon ST, Loc-Carrillo C. Phage cocktails and the future of phage therapy. Future Microbiol. (2013) 8:769–83. doi: 10.2217/fmb.13.47
112. Gu J, Liu X, Li Y, Han W, Lei L, Yang Y, et al. A method for generation phage cocktail with great therapeutic potential. PLoS ONE. (2012) 7:e31698. doi: 10.1371/journal.pone.0031698
113. Korf IHE, Kittler S, Bierbrodt A, Mengden R, Rohde C, Rohde M, et al. In vitro evaluation of a phage cocktail controlling infections with Escherichia coli. Viruses. (2020) 12:1470. doi: 10.3390/v12121470
114. Shahin K, Barazandeh M, Zhang L, Hedayatkhah A, He T, Bao H, et al. Biodiversity of new lytic bacteriophages infecting Shigella spp. in freshwater environment. Front Microbiol. (2021) 12:619323. doi: 10.3389/fmicb.2021.619323
115. García P, Martínez B, Obeso JM, Rodríguez A. Bacteriophages and their application in food safety. Lett Appl Microbiol. (2008) 47:479–85. doi: 10.1111/j.1472-765X.2008.02458.x
116. El-Gohary FA, Huff WE, Huff GR, Rath NC, Zhou ZY, Donoghue AM. Environmental augmentation with bacteriophage prevents colibacillosis in broiler chickens. Poult Sci. (2014) 93:2788–92. doi: 10.3382/ps.2014-04282
117. Huff WE, Huff GR, Rath NC, Donoghue AM. Method of administration affects the ability of bacteriophage to prevent colibacillosis in 1-day-old broiler chickens. Poult Sci. (2013) 92:930–4. doi: 10.3382/ps.2012-02916
118. Sonalika J, Srujana AS, Akhila DS, Juliet MR, Santhosh KS. Application of bacteriophages to control Salmonella Enteritidis in raw eggs. Iran J Vet Res. (2020) 21:221–5. Available online at: https://ijvr.shirazu.ac.ir/article_5719.html
119. Gallet R, Kannoly S, Wang I-N. Effects of bacteriophage traits on plaque formation. BMC Microbiol. (2011) 11:181. doi: 10.1186/1471-2180-11-181
120. Lee S, Hosseindoust A, Goel A, Choi Y, Kwon IK, Chae B. Effects of dietary supplementation of bacteriophage with or without zinc oxide on the performance and gut development of weanling pigs. Ital J Anim Sci. (2016) 15:412–8. doi: 10.1080/1828051X.2016.1188676
121. Zeng Y, Wang Z, Zou T, Chen J, Li G, Zheng L, et al. Bacteriophage as an alternative to antibiotics promotes growth performance by regulating intestinal inflammation, intestinal barrier function and gut microbiota in weaned piglets. Front Vet Sci. (2021) 8:623899. doi: 10.3389/fvets.2021.623899
122. Kim KH, Ingale SL, Kim JS, Lee SH, Lee JH, Kwon IK, et al. Bacteriophage and probiotics both enhance the performance of growing pigs but bacteriophage are more effective. Anim Feed Sci Technol. (2014) 196:88–95. doi: 10.1016/j.anifeedsci.2014.06.012
123. Shaufi MA, Sieo CC, Chong CW, Geok Hun T, Omar AR, Han Ming G, et al. Effects of phage cocktail, probiotics, and their combination on growth performance and gut microbiota of broiler chickens. Animals. (2023) 13:1328. doi: 10.3390/ani13081328
124. Dec M, Wernicki A, Urban-Chmiel R. Efficacy of experimental phage therapies in livestock. Anim Health Res Rev. (2020) 21:69–83. doi: 10.1017/S1466252319000161
125. Cytophage Technologies Inc. The World's First Sustainable Solution to Bacterial Infection and Disease. (2019). Available online at: https://cytophage.com/ (Accessed November 4, 2024).
126. Principi N, Silvestri E, Esposito S. Advantages and limitations of bacteriophages for the treatment of bacterial infections. Front Pharmacol. (2019) 10:513. doi: 10.3389/fphar.2019.00513
127. Joshi D, Roy S, Banerjee S. Prebiotics: a functional food in health and disease. In:Mandal SC, Mandal V, Konishi T, , editors. Natural Products and Drug Discovery. Amsterdam: Elsevier (2018). p. 507–23. doi: 10.1016/B978-0-08-102081-4.00019-8
128. Wang G, Song Q, Huang S, Wang Y, Cai S, Yu H, et al. Effect of antimicrobial peptide microcin J25 on growth performance, immune regulation, and intestinal microbiota in broiler chickens challenged with Escherichia coli and Salmonella. Animals. (2020) 10:345. doi: 10.3390/ani10020345
129. Pollard AJ, Bijker EM. A guide to vaccinology: from basic principles to new developments. Nat Rev Immunol. (2021) 21:83–100. doi: 10.1038/s41577-020-00479-7
130. Burakova Y, Madera R, McVey S, Schlup JR, Shi J. Adjuvants for animal vaccines. Viral Immunol. (2018) 31:11–22. doi: 10.1089/vim.2017.0049
131. Chand DJ, Magiri RB, Wilson HL, Mutwiri GK. Polyphosphazenes as adjuvants for animal vaccines and other medical applications. Front Bioeng Biotechnol. (2021) 9:625482. doi: 10.3389/fbioe.2021.625482
132. Campos J, Mourão J, Peixe L, Antunes P. Non-typhoidal Salmonella in the pig production chain: a comprehensive analysis of its impact on human health. Pathogens. (2019) 8:19. doi: 10.3390/pathogens8010019
133. Davies RH, Kidd S, Mueller-Doblies D, Speed KCR. Salmonella Typhimurium in livestock in Great Britain – trends observed over a 32-year period. Epidemiol Infect. (2018) 146:409–22. doi: 10.1017/S095026881800002X
134. GBD. The global burden of non-typhoidal Salmonella invasive disease: a systematic analysis for the Global Burden of Disease Study 2017. Lancet Infect Dis. (2019) 19:1312–24. doi: 10.1016/S1473-3099(19)30418-9
135. WHO. World Health Organization (WHO) Publishes List of Bacteria for Which New Antibiotics are Urgently Needed. (2017). Available online at: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (Accessed February 27, 2024).
136. Smith RP, Andres V, Martelli F, Gosling B, Marco-Jimenez F, Vaughan K, et al. Maternal vaccination as a Salmonella Typhimurium reduction strategy on pig farms. J Appl Microbiol. (2018) 124:274–85. doi: 10.1111/jam.13609
137. Schmidt S, Sassu EL, Vatzia E, Pierron A, Lagler J, Mair KH, et al. Vaccination and infection of swine with Salmonella Typhimurium induces a systemic and local multifunctional CD4+ T-cell response. Front Immunol. (2021) 11:603089. doi: 10.3389/fimmu.2020.603089
138. Edrington TS, Arthur TM, Loneragan GH, Genovese KJ, Hanson DL, Anderson RC, et al. Evaluation of two commercially-available Salmonella vaccines on Salmonella in the peripheral lymph nodes of experimentally-infected cattle. Ther Adv Vaccines Immunother. (2020) 8:2515135520957760. doi: 10.1177/2515135520957760
139. Dórea FC, Cole DJ, Hofacre C, Zamperini K, Mathis D, Doyle MP, et al. Effect of Salmonella vaccination of breeder chickens on contamination of broiler chicken carcasses in integrated poultry operations. Appl Environ Microbiol. (2010) 76:7820–5. doi: 10.1128/AEM.01320-10
140. Crouch CF, Nell T, Reijnders M, Donkers T, Pugh C, Patel A, et al. Safety and efficacy of a novel inactivated trivalent Salmonella enterica vaccine in chickens. Vaccine. (2020) 38:6741–50. doi: 10.1016/j.vaccine.2020.08.033
141. Hu J, Che C, Zuo J, Niu X, Wang Z, Lian L, et al. Effect of antibiotics on the colonization of live attenuated Salmonella Enteritidis vaccine in chickens. Front Vet Sci. (2021) 8:784160. doi: 10.3389/fvets.2021.784160
142. Haulisah NA, Hassan L, Bejo SK, Jajere SM, Ahmad NI. High levels of antibiotic resistance in isolates from diseased livestock. Front Vet Sci. (2021) 8:652351. doi: 10.3389/fvets.2021.652351
143. Haile AF, Alonso S, Berhe N, Atoma TB, Boyaka PN, Grace D. Prevalence, antibiogram, and multidrug-resistant profile of E. coli O157: H7 in retail raw beef in Addis Ababa, Ethiopia. Front Vet Sci. (2022) 9:734896. doi: 10.3389/fvets.2022.734896
144. Tabaran A, Soulageon V, Chirila F, Reget OL, Mihaiu M, Borzan M, et al. Pathogenic E. coli from cattle as a reservoir of resistance genes to various groups of antibiotics. Antibiotics. (2022) 11:404. doi: 10.3390/antibiotics11030404
145. Vangroenweghe FACJ, Boone M. Vaccination with an Escherichia coli F4/F18 vaccine improves piglet performance combined with a reduction in antimicrobial use and secondary infections due to Streptococcus suis. Animals. (2022) 12:2231. doi: 10.3390/ani12172231
146. Matthews L, Reeve R, Gally DL, Low JC, Woolhouse MEJ, McAteer SP, et al. Predicting the public health benefit of vaccinating cattle against Escherichia coli O157. Proc Nat Acad Sci. (2013) 110:16265–70. doi: 10.1073/pnas.1304978110
147. Tonsor GT, Schroeder TC. Market impacts of E. coli vaccination in US feedlot cattle. Agric Food Econ. (2015) 3:7. doi: 10.1186/s40100-014-0021-2
148. Aida V, Pliasas VC, Neasham PJ, North J, McWhorter KL, Glover SR, et al. Novel vaccine technologies in veterinary medicine: a herald to human medicine vaccines. Front Vet Sci. (2021) 8:654289. doi: 10.3389/fvets.2021.654289
149. Chen T, Ding Z, Lan J, Wong G. Advances and perspectives in the development of vaccines against highly pathogenic Bunyaviruses. Front Cell Infect Microbiol. (2023) 13:1174030. doi: 10.3389/fcimb.2023.1174030
150. Choudhury SM, Ma X, Dang W, Li Y, Zheng H. Recent development of ruminant vaccine against viral diseases. Front Vet Sci. (2021) 8:697194. doi: 10.3389/fvets.2021.697194
151. Mahony TJ, Briody T, Ommeh SC. Can the revolution in mRNA-based vaccine technologies solve the intractable health issues of current ruminant production systems? Vaccines. (2024) 12:152. doi: 10.3390/vaccines12020152
152. Stedman A, Wright D, Schreur PJW, Clark M, Hill AVS, Gilbert SC, et al. Safety and efficacy of ChAdOx1 RVF vaccine against rift valley fever in pregnant sheep and goats. NPJ Vaccines. (2019) 4:44. doi: 10.1038/s41541-019-0138-0
153. Beck D, Ben Maamar M, Skinner MK. Integration of sperm ncRNA-directed DNA methylation and DNA methylation-directed histone retention in epigenetic transgenerational inheritance. Epigenetics Chromatin. (2021) 14:6. doi: 10.1186/s13072-020-00378-0
154. Deng LJ, Deng WQ, Fan SR, Chen MF, Qi M, Lyu WY, et al. m6A modification: recent advances, anticancer targeted drug discovery and beyond. Mol Cancer. (2022) 21:52. doi: 10.1186/s12943-022-01510-2
155. Carlsson L, Clarke JC, Yen C, Gregoire F, Albery T, Billger M, et al. Biocompatible, purified VEGF-A mRNA improves cardiac function after intracardiac injection 1 week post-myocardial infarction in swine. Mol Ther Methods Clin Dev. (2018) 9:330–46. doi: 10.1016/j.omtm.2018.04.003
156. Chavda VP, Soni S, Vora LK, Soni S, Khadela A, Ajabiya J. mRNA-based vaccines and therapeutics for COVID-19 and future pandemics. Vaccines. (2022) 10:2150. doi: 10.3390/vaccines10122150
157. Kozak M, Hu J. DNA vaccines: their formulations, engineering and delivery. Vaccines. (2024) 12:71. doi: 10.3390/vaccines12010071
158. Davis SK, Fan J, Wright QG, Islam MT, Bean AGD, Layton D, et al. Defining correlates of protection for mammalian livestock vaccines against high-priority viral diseases. Front Immunol. (2024) 15:1397780. doi: 10.3389/fimmu.2024.1397780
159. Unger H, Kangethe RT, Liaqat F, Viljoen G. Advances in irradiated livestock vaccine research and production addressing the unmet needs for farmers and veterinary services in FAO/IAEA member states. Front Immunol. (2022) 13:853874. doi: 10.3389/fimmu.2022.853874
160. Dione M, Traoré I, Kassambara H, Sow AN, Touré CO, Sidibé CAK, et al. Integrated approach to facilitate stakeholder participation in the control of endemic diseases of livestock: the case of peste des petits ruminants in Mali. Front Vet Sci. (2019) 6:392. doi: 10.3389/fvets.2019.00392
161. Huang Z. The evolution and impact of animal vaccines: advancements, applications, and future directions. Theor Nat Sci. (2024) 62:1–11. doi: 10.54254/2753-8818/62/20241451
162. Warimwe GM, Francis MJ, Bowden TA, Thumbi SM, Charleston B. Using cross-species vaccination approaches to counter emerging infectious diseases. Nat Rev Immunol. (2021) 21:815–22. doi: 10.1038/s41577-021-00567-2
163. van Dijk A, Hedegaard CJ, Haagsman HP, Heegaard PMH. The potential for immunoglobulins and host defense peptides (HDPs) to reduce the use of antibiotics in animal production. Vet Res. (2018) 49:68. doi: 10.1186/s13567-018-0558-2
164. Kim WH, Lillehoj HS. Immunity, immunomodulation, and antibiotic alternatives to maximize the genetic potential of poultry for growth and disease response. Anim Feed Sci Technol. (2019) 250:41–50. doi: 10.1016/j.anifeedsci.2018.09.016
165. Hedegaard CJ, Strube ML, Hansen MB, Lindved BK, Lihme A, Boye M, et al. Natural pig plasma immunoglobulins have anti-bacterial effects: potential for use as feed supplement for treatment of intestinal infections in pigs. PLoS ONE. (2016) 11:e0147373. doi: 10.1371/journal.pone.0147373
166. Cuperus T, Kraaij MD, Zomer A, Dijk A, Haagsman HP. Immunomodulation and effects on microbiota after in ovo administration of chicken Cathelicidin-2. PLoS ONE. (2018) 13:e0198188. doi: 10.1371/journal.pone.0198188
167. Mookherjee N, Hamill P, Gardy J, Blimkie D, Falsafi R, Chikatamarla A, et al. Systems biology evaluation of immune responses induced by human host defence peptide LL-37 in mononuclear cells. Mol Biosyst. (2009) 5:483–96. doi: 10.1039/b813787k
168. Petrov V, Funderburg N, Weinberg A, Sieg S. Human β defensin-3 induces chemokines from monocytes and macrophages: diminished activity in cells from HIV-infected persons. Immunology. (2013) 140:413–20. doi: 10.1111/imm.12148
169. Roca-Pinilla R, Lisowski L, Aris A, García-Fruitós E. The future of recombinant host defense peptides. Microb Cell Fact. (2022) 21:267. doi: 10.1186/s12934-022-01991-2
170. Sol A, Wang G, Blotnick E, Golla RM, Bachrach G, Mühlrád A. Interaction of the core fragments of the LL-37 host defense peptide with actin. RSC Adv. (2015) 5:9361–7. doi: 10.1039/C4RA13007C
171. Findlay-Greene F, Proudfoot L, Stevens C, Barlow PG. Cationic host defense peptides; novel antimicrobial therapeutics against category a pathogens and emerging infections. Pathog Glob Health. (2016) 110:137–47. doi: 10.1080/20477724.2016.1195036
172. Elhag O, Zhou D, Song Q, Soomro AA, Cai M, Zheng L, et al. Screening. expression, purification and functional characterization of novel antimicrobial peptide genes from Hermetia illucens (L). PLoS ONE. (2017) 12:e0169582. doi: 10.1371/journal.pone.0169582
173. Whitmore M, Tobin I, Burkardt A, Zhang G. Nutritional modulation of host defense peptide synthesis: a novel host-directed antimicrobial therapeutic strategy? Adv Nutr. (2024) 15:100277. doi: 10.1016/j.advnut.2024.100277
174. Robinson K, Ma X, Liu Y, Qiao S, Hou K, Zhang G. Dietary modulation of endogenous host defense peptide synthesis as an alternative approach to in-feed antibiotics. Anim Nutr. (2018) 4:160–9. doi: 10.1016/j.aninu.2018.01.003
175. Lyu Z, Yang P, Jian L, Zhao J. Biological function of antimicrobial peptides on suppressing pathogens and improving host immunity. Antibiotics. (2023) 12:1037. doi: 10.3390/antibiotics12061037
176. Sunkara LT. Biology, expression, and regulation of host defense peptides: a minireview. Adv Anim Vet Sci. (2015) 3:9–20. doi: 10.14737/journal.aavs/2015/3.3s.9.20
177. Balhuizen MD, Dijk A, Jansen JWA, Chris HAL, Veldhuizen EJ, Haagsman HP. Outer membrane vesicles protect gram-negative bacteria against host defense peptides. mSphere. (2021) 6:e00523-21. doi: 10.1128/mSphere.00523-21
178. DeLong MR, Tandon VJ, Lio ALD, Deming TJ, Cederna PS. Overview of host defense peptides and their applications for plastic and reconstructive surgeons. Plast Reconstr Surg. (2020) 146:91–103. doi: 10.1097/PRS.0000000000006910
179. Lei J, Sun L, Huang S, Zhu C, Li P, He J, et al. The antimicrobial peptides and their potential clinical applications. Am J Transl Res. (2019) 11:3919–31.
180. Saeed SI, Mergani A, Aklilu E, Kamaruzzaman NF. Antimicrobial peptides: bringing solution to the rising threats of antimicrobial resistance in livestock [mini review]. Front Vet Sci. (2022) 9:851052. doi: 10.3389/fvets.2022.851052
181. Liu Q, Yao S, Chen Y, Gao S, Yang Y, Deng J, et al. Use of antimicrobial peptides as a feed additive for juvenile goats. Sci Rep. (2017) 7:12254. doi: 10.1038/s41598-017-12394-4
182. Wang S, Zeng X, Yang Q, Qiao S. Antimicrobial peptides as potential alternatives to antibiotics in food animal industry. Int J Mol Sci. (2016) 17:603. doi: 10.3390/ijms17050603
183. Assoni L, Milani B, Carvalho MR, Nepomuceno LN, Waz NT, Guerra MES, et al. Resistance mechanisms to antimicrobial peptides in gram-positive bacteria. Front Microbiol. (2020) 11:593215. doi: 10.3389/fmicb.2020.593215
184. Zhang X, Zhao Q, Wen L, Wu C, Yao Z, Yan Z, et al. The effect of the antimicrobial peptide plectasin on the growth performance, intestinal health, and immune function of yellow-feathered chickens. Front Vet Sci. (2021) 8:688611. doi: 10.3389/fvets.2021.688611
185. Wang J, Dou X, Song J, Lyu Y, Zhu X, Xu L, et al. Antimicrobial peptides: promising alternatives in the post feeding antibiotic era. Med Res Rev. (2019) 39:831–59. doi: 10.1002/med.21542
186. Cutler SA, Lonergan SM, Cornick N, Johnson AK, Stahl CH. Dietary inclusion of colicin e1 is effective in preventing postweaning diarrhea caused by F18-positive Escherichia coli in pigs. Antimicrob Agents Chemother. (2007) 51:3830–5. doi: 10.1128/AAC.00360-07
187. Xiao H, Shao F, Wu M, Ren W, Xiong X, Tan B, et al. The application of antimicrobial peptides as growth and health promoters for swine. J Anim Sci Biotechnol. (2015) 6:1–6. doi: 10.1186/s40104-015-0018-z
188. Nazeer N, Uribe-Diaz S, Rodriguez-Lecompte JC, Ahmed M. Antimicrobial peptides as an alternative to relieve antimicrobial growth promoters in poultry. Br Poult Sci. (2021) 62:672–85. doi: 10.1080/00071668.2021.1919993
189. Ioannou F, Burnsteel C, Mackay DKJ, Gay CG. Regulatory pathways to enable the licencing of alternatives to antibiotics. Biologicals. (2018) 53:72–5. doi: 10.1016/j.biologicals.2018.03.003
190. Bedford MR. The evolution and application of enzymes in the animal feed industry: the role of data interpretation. Br Poult Sci. (2018) 59:486–93. doi: 10.1080/00071668.2018.1484074
191. Wang H, Zeng X, Cao L, He Q, Lin J. Passive immunization of chickens with anti-enterobactin egg yolk powder for campylobacter control. Vaccines. (2021) 9:569. doi: 10.3390/vaccines9060569
192. Hauck R, Macklin KS. Vaccination against poultry parasites. Avian Dis. (2024) 67:441–9. doi: 10.1637/aviandiseases-D-23-99989
193. Zeng M, Zhou R, He T, Hu F, Liu W, Gan N, et al. Bioluminescence assay for rapid detection of live Staphylococcus aureus based on the enrichment of egg yolk antibody modified magnetic metal organic framework immunobeads. Analyst. (2024) 149:876–84. doi: 10.1039/D3AN01564E
194. Lee SH, Lillehoj HS, Park DW, Jang SI, Morales A, García D, et al. Protective effect of hyperimmune egg yolk IgY antibodies against Eimeria tenella and Eimeria maxima infections. Vet Parasitol. (2009) 163:123–6. doi: 10.1016/j.vetpar.2009.04.020
195. Hatta H, Tsuda K, Akachi S, Kim M, Yamamoto T, Ebina T. Oral passive immunization effect of anti-human rotavirus IgY and its behavior against proteolytic enzymes. Biosci Biotechnol Biochem. (1993) 57:1077–81. doi: 10.1271/bbb.57.1077
196. Xia M, Ahn DU, Liu C, Cai Z. A basis for IgY-themed functional foods: digestion profile of oral yolk immunoglobulin (IgY) by INFOGEST static digestion model. Food Res Int. (2022) 162:112167. doi: 10.1016/j.foodres.2022.112167
197. Wang H, Zhong Q, Lin J. Egg yolk antibody for passive immunization: status, challenges, and prospects. J Agric Food Chem. (2023) 71:5053–61. doi: 10.1021/acs.jafc.2c09180
198. Gadde U, Rathinam T, Lillehoj HS. Passive immunization with hyperimmune egg-yolk IgY as prophylaxis and therapy for poultry diseases–a review. Anim Health Res Rev. (2015) 16:163–76. doi: 10.1017/S1466252315000195
199. Frazzini S, Riva F, Amadori M. Therapeutic and prophylactic use of oral, low-dose ifns in species of veterinary interest: back to the future. Vet Sci. (2021) 8:109. doi: 10.3390/vetsci8060109
200. Sordillo LM, Babiuk LA. Controlling acute Escherichia coli mastitis during the periparturient period with recombinant bovine interferon gamma. Vet Microbiol. (1991) 28:189–98. doi: 10.1016/0378-1135(91)90092-T
201. Fan W, Jiao P, Zhang H, Chen T, Zhou X, Qi Y, et al. Inhibition of African swine fever virus replication by porcine type I and type II interferons. Front Microbiol. (2020) 11:1203. doi: 10.3389/fmicb.2020.01203
202. Do DN, Dudemaine P-L, Mathur M, Suravajhala P, Zhao X, Ibeagha-Awemu EM. MiRNA regulatory functions in farm animal diseases, and biomarker potentials for effective therapies. Int J Mol Sci. (2021) 22:3080. doi: 10.3390/ijms22063080
203. Li Q, Ji C, Chen Y, Niu X, Sun F, Yin S, et al. Stem cell-derived extracellular vesicles (EVS) for disease therapy. In:Wang Q, Zheng F, , editors. Extracellular Vesicles: From Bench to Bedside. Cham: Springer (2024). p. 437–65. doi: 10.1007/978-981-99-8365-0_23
204. Moccia V, Sammarco A, Cavicchioli L, Castagnaro M, Bongiovanni L, Zappulli V. Extracellular vesicles in veterinary medicine. Animals. (2022) 12:2716. doi: 10.3390/ani12192716
205. Topper MJ, Vaz M, Marrone KA, Brahmer JR, Baylin SB. The emerging role of epigenetic therapeutics in immuno-oncology. Nat Rev Clin Oncol. (2020) 17:75–90. doi: 10.1038/s41571-019-0266-5
206. Wang M, Ibeagha-Awemu EM. Impacts of epigenetic processes on the health and productivity of livestock [review]. Front Genet. (2021) 11:613636. doi: 10.3389/fgene.2020.613636
207. Xiao X, Feng H, Liao Y, Tang H, Li L, Li K, et al. Identification of lncRNA-miRNA-mRNA regulatory network and therapeutic agents for skin aging by bioinformatics analysis. Biochem Genet. (2023) 61:1606–24. doi: 10.1007/s10528-023-10334-8
208. Xiong Y, Lou P, Xu C, Han B, Liu J, Gao J. Emerging role of extracellular vesicles in veterinary practice: novel opportunities and potential challenges. Front Vet Sci. (2024) 11:1335107. doi: 10.3389/fvets.2024.1335107
209. Ali A, Ijaz M, Khan YR, Sajid HA, Hussain K, Rabbani AH, et al. Role of nanotechnology in animal production and veterinary medicine. Trop Anim Health Prod. (2021) 53:508. doi: 10.1007/s11250-021-02951-5
210. Hargreaves CA, Rogers S, Hills F, Rahman F, Howell RJ, Homa ST. Effects of co-trimoxazole, erythromycin, amoxycillin, tetracycline and chloroquine on sperm function in vitro. Hum Reprod. (1998) 13:1878–86. doi: 10.1093/humrep/13.7.1878
211. Taouzinet L, Fatmi S, Lahiani-Skiba M, Skiba M, Iguer-Ouada M. Encapsulation nanotechnology in sperm cryopreservation: systems preparation methods and antioxidants enhanced delivery. CryoLetters. (2021) 42:1–12.
212. Marco V, Toscano F, Alex G, Torres M, Seqqat R. Stem cells and nanoparticles in therapy. Nanoscie Nanotechnol-Asia. (2021) 11:e010121189823. doi: 10.2174/2210681210999210101155808
213. Goel S, Gandhi S, Dubey S, Shah M, Saini S, Arora P, et al. Stem cell therapy: promises and challenges in treating animal diseases. In:Mukhopadhyay CS, Choudhary RK, Panwar H, Malik YS, , editors. Biotechnological Interventions Augmenting Livestock Health and Production. Cham: Springer (2023). p. 13–38. doi: 10.1007/978-981-99-2209-3_2
214. Malakar D, Malik HN, Kumar D, Saini S, Sharma V, Fatima S, et al. Stem cells: a potential regenerative medicine for treatment of diseases. in Advances in Animal Genomics. Amsterdam: Elsevier (2021). p. 33–48. doi: 10.1016/B978-0-12-820595-2.00003-5
215. Omonijo FA, Liu S, Hui Q, Zhang H, Lahaye L, Bodin JC, et al. Thymol improves barrier function and attenuates inflammatory responses in porcine intestinal epithelial cells during lipopolysaccharide (LPS)-induced inflammation. J Agric Food Chem. (2019) 67:615–24. doi: 10.1021/acs.jafc.8b05480
216. Rostami F, Ghasemi HA, Taherpour K. Effect of Scrophularia striata and Ferulago angulata, as alternatives to virginiamycin, on growth performance, intestinal microbial population, immune response, and blood constituents of broiler chickens. Poult Sci. (2015) 94:2202–9. doi: 10.3382/ps/pev198
217. Biswas S, Ahn JM, Kim IH. Assessing the potential of phytogenic feed additives: a comprehensive review on their effectiveness as a potent dietary enhancement for nonruminant in swine and poultry. J Anim Physiol Anim Nutr. (2024) 108:711–23. doi: 10.1111/jpn.13922
218. Seidavi A, Tavakoli M, Slozhenkina M, Gorlov I, Hashem NM, Asroosh F, et al. The use of some plant-derived products as effective alternatives to antibiotic growth promoters in organic poultry production: a review. Environ Sci Pollut Res. (2021) 28:47856–68. doi: 10.1007/s11356-021-15460-7
219. Khan K, Ahmad N. Using cinnamon (Cinnamomum zeylanicum) and turmeric (Curcuma longa L) powders as an antibiotic growth promoter substitutions in broiler chicken's diets. Anim Biotechnol. (2023) 34:4466–73. doi: 10.1080/10495398.2022.2157282
220. Recharla N, Balasubramanian B, Song M, Puligundla P, Kim S-k, Jeong JY, et al. Dietary turmeric (Curcuma longa L.) supplementation improves growth performance, short-chain fatty acid production, and modulates bacterial composition of weaned piglets. J Anim Sci Technol. (2021) 63:575. doi: 10.5187/jast.2021.e55
221. Akosile OA, Kehinde FO, Oni AI, Oke OE. Potential implication of in ovo feeding of phytogenics in poultry production. Transl Anim Sci. (2023) 7:txad094. doi: 10.1093/tas/txad094
222. Arain MA, Nabi F, Shah QA, Alagawany M, Fazlani SA, Khalid M, et al. The role of early feeding in improving performance and health of poultry: herbs and their derivatives. Worlds Poul Sci J. (2022) 78:499–513. doi: 10.1080/00439339.2022.2043133
223. Alves-Santos AM, Sugizaki CSA, Lima GC, Naves MMV. Prebiotic effect of dietary polyphenols: a systematic review. J Funct Foods. (2020) 74:104169. doi: 10.1016/j.jff.2020.104169
224. Zhang L, Gao F, Ge J, Li H, Xia F, Bai H, et al. Potential of aromatic plant-derived essential oils for the control of foodborne bacteria and antibiotic resistance in animal production: a review. Antibiotics. (2022) 11:1673. doi: 10.3390/antibiotics11111673
225. Valenzuela-Grijalva NV, Pinelli-Saavedra A, Muhlia-Almazan A, Domínguez-Díaz D, González-Ríos H. Dietary inclusion effects of phytochemicals as growth promoters in animal production. J Anim Sci Technol. (2017) 59:1–17. doi: 10.1186/s40781-017-0133-9
226. Campolina JP, Coelho SG, Belli AL, Neves LFM, Machado FS, Pereira LG, et al. Potential benefits of a blend of essential oils on metabolism, digestibility, organ development and gene expression of dairy calves. Sci Rep. (2023) 13:3378. doi: 10.1038/s41598-023-30088-y
227. Navarro M, Stanley R, Cusack A, Sultanbawa Y. Combinations of plant-derived compounds against Campylobacter in vitro. J Appl Poult Res. (2015) 24:352–63. doi: 10.3382/japr/pfv035
228. Cantet JM, Yu Z, Tucker HA, Ríus AG. A cinnamaldehyde feed additive improved feed use-efficiency in lactating dairy cows. Livest Sci. (2023) 272:105236. doi: 10.1016/j.livsci.2023.105236
229. Chaucheyras-Durand F, Durand H. Probiotics in animal nutrition and health. Benef Microbes. (2010) 1:3–9. doi: 10.3920/BM2008.1002
230. Kulkarni NA, Chethan HS, Srivastava R, Gabbur AB. Role of probiotics in ruminant nutrition as natural modulators of health and productivity of animals in tropical countries: an overview. Trop Anim Health Prod. (2022) 54:110. doi: 10.1007/s11250-022-03112-y
231. Chouraddi R, Kumar S, Gujjalkar PR, Vinay VV, Banakar P. A review on probiotics for animals and their mode of action. Pharma Innov J. (2023) 12:500–6. Available online at: https://www.thepharmajournal.com/archives/2023/vol12issue2/PartF/12-3-400-949.pdf
232. Das P, Meel M. Role of direct fed microbials in enhancing bovine productivity: a review. Int J Vet Sci Anim Husb. (2024) SP-9:586–90. Available online at: https://www.veterinarypaper.com/special-issue/2024/9/1S/S-9-1-34
233. Shaffi MS, Hameed MK. The role of probiotics in animal nutrition and health. World J Adv Res Rev. (2023) 17:276–80. doi: 10.30574/wjarr.2023.17.3.0396
234. Ciorba MA. A gastroenterologist's guide to probiotics. Clin Gastroenterol Hepatol. (2012) 10:960–8. doi: 10.1016/j.cgh.2012.03.024
235. Wang Y, Gu Q. Effect of probiotic on growth performance and digestive enzyme activity of Arbor Acres broilers. Res Vet Sci. (2010) 89:163–7. doi: 10.1016/j.rvsc.2010.03.009
236. Adjei-Fremah S, Ekwemalor K, Asiamah E, Ismail H, Worku M. Transcriptional profiling of the effect of lipopolysaccharide (LPS) pretreatment in blood from probiotics-treated dairy cows. Genomics Data. (2016) 10:15–8. doi: 10.1016/j.gdata.2016.08.016
237. Butt UD, Khan S, Liu X, Sharma A, Zhang X, Wu B. Present status, limitations, and prospects of using streptomyces bacteria as a potential probiotic agent in aquaculture. Probiotics Antimicrob Proteins. (2023) 16:426–42. doi: 10.1007/s12602-023-10053-x
238. Samal L, Behura N. Prebiotics: an emerging nutritional approach for improving gut health of livestock and poultry. Asian J Anim Vet Adv. (2015) 10:724–39. doi: 10.3923/ajava.2015.724.739
239. Yaqoob MU, El-Hack MEA, Hassan F, El-Saadony MT, Khafaga AF, Batiha GE, et al. The potential mechanistic insights and future implications for the effect of prebiotics on poultry performance, gut microbiome, and intestinal morphology. Poult Sci. (2021) 100:101143. doi: 10.1016/j.psj.2021.101143
240. Shehata AA, Yalçin S, Latorre JD, Basiouni S, Attia YA, Abd El-Wahab A, et al. Probiotics, prebiotics, and phytogenic substances for optimizing gut health in poultry. Microorganisms. (2022) 10:395. doi: 10.3390/microorganisms10020395
241. Adhikari PA, Kim WK. Overview of prebiotics and probiotics: focus on performance, gut health and immunity–a review. Ann Anim Sci. (2017) 17:949–66. doi: 10.1515/aoas-2016-0092
242. Wang Y, Sun J, Zhong H, Li N, Xu H, Zhu Q, et al. Effect of probiotics on the meat flavour and gut microbiota of chicken. Sci Rep. (2017) 7:1–13. doi: 10.1038/s41598-017-06677-z
243. Anadón A, Ares I, Martínez-Larrañaga MR, Martínez MA. Prebiotics and probiotics in feed and animal health. In:Gupta RC, Srivastava A, Lall R, , editors. Nutraceuticals in Veterinary Medicine. Cham: Springer (2019). p. 261–85. doi: 10.1007/978-3-030-04624-8_19
244. Roth N, Hofacre C, Zitz U, Mathis GF, Moder K, Doupovec B, et al. Prevalence of antibiotic-resistant E. coli in broilers challenged with a multi-resistant E coli strain and received ampicillin, an organic acid-based feed additive or a synbiotic preparation. Poult Sci. (2019) 98:2598–607. doi: 10.3382/ps/pez004
245. Cheon S, Kim G, Bae JH, Lee DH, Seong H, Kim DH, et al. Comparative analysis of prebiotic effects of four oligosaccharides using in vitro gut model: digestibility, microbiome, and metabolome changes. FEMS Microbiol Ecol. (2023) 99:fiad002. doi: 10.1093/femsec/fiad002
246. Fuhrmann L, Vahjen W, Zentek J, Günther R, Saliu E-M. The impact of pre- and probiotic product combinations on ex vivo growth of avian pathogenic Escherichia coli and Salmonella Enteritidis. Microorganisms. (2022) 10:121. doi: 10.3390/microorganisms10010121
247. Low CX, Tan LTH, Mutalib NSA, Pusparajah P, Goh BH, Chan KG, et al. Unveiling the impact of antibiotics and alternative methods for animal husbandry: a review. Antibiotics. (2021) 10:578. doi: 10.3390/antibiotics10050578
248. Thacker PA. Alternatives to antibiotics as growth promoters for use in swine production: a review. J Anim Sci Biotechnol. (2013) 4:35. doi: 10.1186/2049-1891-4-35
249. Estrada-Angulo A, Zapata-Ramírez O, Castro-Pérez BI, Urías-Estrada JD, Gaxiola-Camacho S, Angulo-Montoya C, et al. The effects of single or combined supplementation of probiotics and prebiotics on growth performance, dietary energetics, carcass traits, and visceral mass in lambs finished under subtropical climate conditions. Biology. (2021) 10:1137. doi: 10.3390/biology10111137
250. Sharma AN, Chaudhary P, Grover CR, Kumar S, Mondal G. Impact of synbiotics on growth performance and gut health in Murrah buffalo calves. Vet Res Commun. (2024) 48:179–90. doi: 10.1007/s11259-023-10194-y
251. Krumbeck JA, Walter J, Hutkins RW. Synbiotics for improved human health: recent developments, challenges, and opportunities. Annu Rev Food Sci Technol. (2018) 9:451–79. doi: 10.1146/annurev-food-030117-012757
252. Salminen S, Collado MC, Endo A, Hill C, Lebeer S, Quigley EM, et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat Rev Gastroenterol Hepatol. (2021) 18:649–67. doi: 10.1038/s41575-021-00440-6
253. Vinderola G, Sanders ME, Salminen S. The concept of postbiotics. Foods. (2022) 11:1077. doi: 10.3390/foods11081077
254. Waqas M, Nastoh N, Çinar A, Farooq M, Salman M. Advantages of the use of postbiotics in poultry production: a new concept. Braz J Poult Sci. (2024) 26:eRBCA-2024-1939. doi: 10.1590/1806-9061-2024-1939
255. Kumar A, Green KM, Rawat M. A comprehensive overview of postbiotics with a special focus on discovery techniques and clinical applications. Foods. (2024) 13:2937. doi: 10.3390/foods13182937
256. Sanli ER. Usability of postbiotics in ruminant nutrition and health. Turk J Agric-Food Sci Technol. (2024) 12:1104–9. doi: 10.24925/turjaf.v12i6.1104-1109.6710
257. Abd El-Ghany WA, Abdel-Latif MA, Hosny F, Alatfeehy NM, Noreldin AE, Quesnell RR, et al. Comparative efficacy of postbiotic, probiotic, and antibiotic against necrotic enteritis in broiler chickens. Poult Sci. (2022) 101:101988. doi: 10.1016/j.psj.2022.101988
258. Fang S, Fan X, Xu S, Gao S, Wang T, Chen Z, et al. Effects of dietary supplementation of postbiotic derived from Bacillus subtilis ACCC 11025 on growth performance, meat yield, meat quality, excreta bacteria, and excreta ammonia emission of broiler chicks. Poult Sci. (2024) 103:103444. doi: 10.1016/j.psj.2024.103444
259. Vicente F, Campo-Celada M, Menéndez-Miranda M, García-Rodríguez J, Martínez-Fernández A. Effect of postbiotic supplementation on nutrient digestibility and milk yield during the transition period in dairy cows. Animals. (2024) 14:2359. doi: 10.3390/ani14162359
260. Humam AM, Loh TC, Foo HL, Samsudin AA, Mustapha NM, Zulkifli I, et al. Effects of feeding different postbiotics produced by Lactobacillus plantarum on growth performance, carcass yield, intestinal morphology, gut microbiota composition, immune status, and growth gene expression in broilers under heat stress. Animals. (2019) 9:644. doi: 10.3390/ani9090644
261. Johnson CN, Kogut MH, Genovese K, He H, Kazemi S, Arsenault RJ. Administration of a postbiotic causes immunomodulatory responses in broiler gut and reduces disease pathogenesis following challenge. Microorganisms. (2019) 7:268. doi: 10.3390/microorganisms7080268
262. Humam AM, Loh TC, Foo HL, Izuddin WI, Awad EA, Idrus Z, et al. Dietary supplementation of postbiotics mitigates adverse impacts of heat stress on antioxidant enzyme activity, total antioxidant, lipid peroxidation, physiological stress indicators, lipid profile and meat quality in broilers. Animals. (2020) 10:982. doi: 10.3390/ani10060982
263. Humam AM, Loh TC, Foo HL, Izuddin WI, Zulkifli I, Samsudin AA, et al. Supplementation of postbiotic RI11 improves antioxidant enzyme activity, upregulated gut barrier genes, and reduced cytokine, acute phase protein, and heat shock protein 70 gene expression levels in heat-stressed broilers. Poult Sci. (2021) 100:100908. doi: 10.1016/j.psj.2020.12.011
264. Izuddin WI, Humam AM, Loh TC, Foo HL, Samsudin AA. Dietary postbiotic Lactobacillus plantarum improves serum and ruminal antioxidant activity and upregulates hepatic antioxidant enzymes and ruminal barrier function in post-weaning lambs. Antioxidants. (2020) 9:250. doi: 10.3390/antiox9030250
265. Zhao X, Liu S, Li S, Jiang W, Wang J, Xiao J, et al. Unlocking the power of postbiotics: a revolutionary approach to nutrition for humans and animals. Cell Metab. (2024) 36:725–44. doi: 10.1016/j.cmet.2024.03.004
266. Urban J, Kareem KY, Atanasov AG, Matuszewski A, Bień D, Ciborowska P, et al. Postbiotics, a natural feed additive for growth performance, gut microbiota and quality of poultry products. Curr Res Biotechnol. (2024) 8:100247. doi: 10.1016/j.crbiot.2024.100247
267. Ferronato G, Prandini A. Dietary supplementation of inorganic, organic, and fatty acids in pig: a review. Animals. (2020) 10:1740. doi: 10.3390/ani10101740
268. Tugnoli B, Giovagnoni G, Piva A, Grilli E. From acidifiers to intestinal health enhancers: how organic acids can improve growth efficiency of pigs. Animals. (2020) 10:134. doi: 10.3390/ani10010134
269. Xu QL, Liu C, Mo XJ, Chen M, Zhao XL, Liu MZ, et al. Drinking water supplemented with acidifiers improves the growth performance of weaned pigs and potentially regulates antioxidant capacity, immunity, and gastrointestinal microbiota diversity. Antioxidants. (2022) 11:809. doi: 10.3390/antiox11050809
270. Nguyen DH, Seok WJ, Kim IH. Organic acids mixture as a dietary additive for pigs—a review. Animals. (2020) 10:952. doi: 10.3390/ani10060952
271. Khan RU, Naz S, Raziq F, Qudratullah Q, Khan NA, Laudadio V, et al. Prospects of organic acids as safe alternative to antibiotics in broiler chickens diet. Environ Sci Pollut Res. (2022) 29:32594–604. doi: 10.1007/s11356-022-19241-8
272. Samanta S, Haldar S, Ghosh TK. Comparative efficacy of an organic acid blend and bacitracin methylene disalicylate as growth promoters in broiler chickens: effects on performance, gut histology, and small intestinal milieu. Vet Med Int. (2010) 2010:645150. doi: 10.4061/2010/645150
273. Ma J, Piao X, Shang Q, Long S, Liu S, Mahfuz S. Mixed organic acids as an alternative to antibiotics improve serum biochemical parameters and intestinal health of weaned piglets. Anim Nutr. (2021) 7:737–49. doi: 10.1016/j.aninu.2020.11.018
274. Scicutella F, Mannelli F, Daghio M, Viti C, Buccioni A. Polyphenols and organic acids as alternatives to antimicrobials in poultry rearing: a review. Antibiotics. (2021) 10:1010. doi: 10.3390/antibiotics10081010
275. USFDA. GRAS Substances (SCOGS) Database. U.S. Food and Drug Administration (USFDA) (2022). Available online at: https://www.fda.gov/food/generally-recognized-safe-gras/gras-substances-scogs-database (Accessed October 19, 2022).
276. Ravindran V, Kornegay E. Acidification of weaner pig diets: a review. J Sci Food Agric. (1993) 62:313–22. doi: 10.1002/jsfa.2740620402
278. Walsh M, Sholly D, Kelly D, Cobb M, Trapp S, Hinson R, et al. The Effects of Supplementing Weanling Pig Diets with Organic and Inorganic Acids on Growth Performance and Microbial Shedding. Swine Research Report. West Lafayette, IN: Purdue University (2003). p. 89–98.
279. Shu Y, Yu B, He J, Yu J, Zheng P, Yuan Z, et al. Excess of dietary benzoic acid supplementation leads to growth retardation, hematological abnormality and organ injury of piglets. Livest Sci. (2016) 190:94–103. doi: 10.1016/j.livsci.2016.06.010
280. Mao X, Yang Q, Chen D, Yu B, He J. Benzoic acid used as food and feed additives can regulate gut functions. Biomed Res Int. (2019) 2019:5721585. doi: 10.1155/2019/5721585
281. Wegl G, Rolinec M, Nagl V, Gierus M, Klose V. Effect of oxytetracycline as well as an acid-based feed supplement on the prevalence and abundance of selected antibiotic resistance genes in weaned piglets. Anim Husb Dairy Vet Sci. (2017) 1:1–6. doi: 10.15761/AHDVS.1000117
282. Dahmer PL, Jones CK. Evaluating dietary acidifiers as alternatives for conventional feed-based antibiotics in nursery pig diets. Transl Anim Sci. (2021) 5:txab040. doi: 10.1093/tas/txab040
283. Bennett SL, Arce-Cordero JA, Brandao VLN, Vinyard JR, Agustinho BC, Monteiro HF, et al. Effects of bacterial cultures, enzymes, and yeast-based feed additive combinations on ruminal fermentation in a dual-flow continuous culture system. Transl Anim Sci. (2021) 5:txab026. doi: 10.1093/tas/txab026
284. Ojha BK, Singh PK, Shrivastava N. Chapter 7 - Enzymes in the animal feed industry. In:Kuddus M, , editor. Enzymes in Food Biotechnology. New York, NY: Academic Press (2019). p. 93–109. doi: 10.1016/B978-0-12-813280-7.00007-4
285. Klein N, Sarpong N, Melzer T, Feuerstein D, Heyer CM, Camarinha-Silva A, et al. Effect of dietary calcium concentration and exogenous phytase on inositol phosphate degradation, mineral digestibility, and gut microbiota in growing pigs. J Anim Sci. (2023) 101:skad254. doi: 10.1093/jas/skad254
286. Yu SG, Lim CB, Jang SY, Zhang QQ, Kim IH. PSIII-13 supplementary effects of carbohydrate enzymes and protease to two protein levels in the diet of finishing pig on the growth performance, toxic gas emissions, meat quality, and colonic microbiota community. J Anim Sci. (2023) 101(Supplement_2):300–1. doi: 10.1093/jas/skad341.341
287. Anadón A, Ares I, Martínez-Larrañaga MR, Martínez MA. Enzymes in feed and animal health. In:Gupta R, Srivastava A, Lall R, , editors. Nutraceuticals in Veterinary Medicine. Cham: Springer (2019). p. 303–13. doi: 10.1007/978-3-030-04624-8_21
288. Liu ZK, Li Y, Zhao CC, Liu ZJ, Wang LM, Li XY, et al. Effects of a combination of fibrolytic and amylolytic enzymes on ruminal enzyme activities, bacterial diversity, blood profile and milk production in dairy cows. Animal. (2022) 16:100595. doi: 10.1016/j.animal.2022.100595
289. Jang JC, Kim KH, Kim DH, Jang SK, Hong JS, Heo PS, et al. Effects of increasing levels of palm kernel meal containing β-mannanase to growing-finishing pig diets on growth performance, nutrient digestibility, and pork quality. Livest Sci. (2020) 238:104041. doi: 10.1016/j.livsci.2020.104041
290. Bedford MR, Cowieson AJ. Matrix values for exogenous enzymes and their application in the real world. J Appl Poult Res. (2020) 29:15–22. doi: 10.1016/j.japr.2019.10.011
291. Lemos MJ, Calixto LFL, Alves OS, Souza DS, Moura BB, Reis TL. Kaolin in the diet and its effects on performance, litter moisture and intestinal morphology of broiler chickens. Ciência Rural. (2015) 45:1835–40. doi: 10.1590/0103-8478cr20141193
292. Damato A, Vianello F, Novelli E, Balzan S, Gianesella M, Giaretta E, et al. Comprehensive review on the interactions of clay minerals with animal physiology and production. Front Vet Sci. (2022) 9:889612. doi: 10.3389/fvets.2022.889612
293. Nadziakiewicza M, Kehoe S, Micek P. Physico-chemical properties of clay minerals and their use as a health promoting feed additive. Animals. (2019) 9:714. doi: 10.3390/ani9100714
294. Azzouz A, Arus VA, Platon N. Role of clay substrate molecular interactions in some dairy technology applications. Int J Mol Sci. (2024) 25:808. doi: 10.3390/ijms25020808
295. Muniyappan M, Shanmugam S, Kim IH. Effects of dietary supplementation of illite on growth performance, nutrient digestibility, and meat-carcass grade quality of growing-finishing pigs. J Anim Sci Technol. (2024) 66:115–24. doi: 10.5187/jast.2023.e24
296. Morrison KD, Misra R, Williams LB. Unearthing the antibacterial mechanism of medicinal clay: a geochemical approach to combating antibiotic resistance. Sci Rep. (2016) 6:19043. doi: 10.1038/srep19043
297. Morrison KD, Underwood JC, Metge DW, Eberl DD, Williams LB. Mineralogical variables that control the antibacterial effectiveness of a natural clay deposit. Environ Geochem Health. (2014) 36:613–31. doi: 10.1007/s10653-013-9585-0
298. Hamilton AR, Roberts M, Hutcheon GA, Gaskell EE. Formulation and antibacterial properties of clay mineral-tetracycline and -doxycycline composites. Appl Clay Sci. (2019) 179:105148. doi: 10.1016/j.clay.2019.105148
299. Prvulovic D, Kojic D, Grubor-Lajsic G, Kosarcic S. The effects of dietary inclusion of hydrated aluminosilicate on performance and biochemical parameters of broiler chickens. Turk J Vet Anim Sci. (2008) 32:159–66. Available online at: https://journals.tubitak.gov.tr/veterinary/vol32/iss3/6/
300. Davis EM, Liang Y, Wallace KP, Zimmerman AJ, Siebecker MG, Broadway PR, et al. A porous ceramic particle with or without a preservative blend did not impair apparent digestibility of macro- and micro-nutrients of postweaned pigs. Transl Anim Sci. (2022) 6:txac078. doi: 10.1093/tas/txac078
301. Boyd SA, Johnston CT, Pinnavaia TJ, Kaminski NE, Teppen BJ, Li H, et al. Suppression of humoral immune responses by 2,3,7,8-tetrachlorodibenzo-p-dioxin intercalated in smectite clay. Environ Toxicol Chem. (2011) 30:2748–55. doi: 10.1002/etc.701
302. Hejna M, Gottardo D, Baldi A, Dell'Orto V, Cheli F, Zaninelli M, et al. Review: nutritional ecology of heavy metals. Animal. (2018) 12:2156–70. doi: 10.1017/S175173111700355X
303. Tariq H, Sharma A, Sarkar S, Ojha L, Pal RP, Mani V. Perspectives for rare earth elements as feed additive in livestock — a review. Asian-Austral J Anim Sci. (2020) 33:373–81. doi: 10.5713/ajas.19.0242
304. van Baal J, Kruijt L, Binnendijk G, Durosoy S, Romeo A, Bikker P. Influence of copper source and dietary inclusion level on growth performance of weaned pigs and expression of trace element related genes in the small intestine. Animal. (2024) 18:101113. doi: 10.1016/j.animal.2024.101113
305. Lei XJ, Liu ZZ, Park JH, Kim IH. Novel zinc sources as antimicrobial growth promoters for monogastric animals: a review. J Anim Sci Technol. (2022) 64:187–96. doi: 10.5187/jast.2022.e1
306. Schokker D, Kar SK, Willems E, Bossers A, Dekker RA, Jansman AJM. Dietary supplementation of zinc oxide modulates intestinal functionality during the post-weaning period in clinically healthy piglets. J Anim Sci Biotechnol. (2023) 14:122. doi: 10.1186/s40104-023-00925-1
307. Younas Z, Mashwani ZUR, Ahmad I, Khan M, Zaman S, Sawati L, et al. Mechanistic approaches to the application of nano-zinc in the poultry and biomedical industries: a comprehensive review of future perspectives and challenges. Molecules. (2023) 28:1064. doi: 10.3390/molecules28031064
308. Burrell AL, Dozier WA, Davis AJ, Compton MM, Freeman ME, Vendrell PF, et al. Responses of broilers to dietary zinc concentrations and sources in relation to environmental implications. Br Poult Sci. (2004) 45:225–63. doi: 10.1080/00071660410001715867
309. Yusof MH, Abdul Rahman NA, Mohamad R, Zaidan UH, Arshad MA, Samsudin AA. Effects of dietary zinc oxide nanoparticles supplementation on broiler growth performance, zinc retention, liver health status, and gastrointestinal microbial load. J Trace Elem Miner. (2023) 4:100072. doi: 10.1016/j.jtemin.2023.100072
310. Espinosa CD, Stein HH. Digestibility and metabolism of copper in diets for pigs and influence of dietary copper on growth performance, intestinal health, and overall immune status: a review. J Anim Sci Biotechnol. (2021) 12:13. doi: 10.1186/s40104-020-00533-3
311. Brinck JE, Lassen SB, Forouzandeh A, Pan T, Wang Y-Z, Monteiro A, et al. Impacts of dietary copper on the swine gut microbiome and antibiotic resistome. Sci Total Environ. (2023) 857:159609. doi: 10.1016/j.scitotenv.2022.159609
312. Hamdi M, Solà D, Franco R, Durosoy S, Roméo A, Pérez JF. Including copper sulphate or dicopper oxide in the diet of broiler chickens affects performance and copper content in the liver. Anim Feed Sci Technol. (2018) 237:89–97. doi: 10.1016/j.anifeedsci.2018.01.014
313. Abdelnour SA, Abd El-Hack ME, Khafaga AF, Noreldin AE, Arif M, Chaudhry MT, et al. Impacts of rare earth elements on animal health and production: highlights of cerium and lanthanum. Sci Total Environ. (2019) 672:1021–32. doi: 10.1016/j.scitotenv.2019.02.270
314. Tommasi F, Thomas PJ, Lyons DM, Pagano G, Oral R, Siciliano A, et al. Evaluation of rare earth element-associated hormetic effects in candidate fertilizers and livestock feed additives. Biol Trace Elem Res. (2023) 201:2573–81. doi: 10.1007/s12011-022-03331-2
315. Xu Y, Luo W, Deng H, Hu X, Zhang J, Wang Y. Robust antibacterial activity of rare-earth ions on planktonic and biofilm bacteria. Biomed Mater. (2024) 19:045014. doi: 10.1088/1748-605X/ad4aa9
316. Poole K. At the nexus of antibiotics and metals: the impact of Cu and Zn on antibiotic activity and resistance. Trends Microbiol. (2017) 25:820–32. doi: 10.1016/j.tim.2017.04.010
317. Gadde U, Kim WH, Oh ST, Lillehoj HS. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: a review. Anim Health Res Rev. (2017) 18:26–45. doi: 10.1017/S1466252316000207
318. Afzal A, Mahreen N. Emerging insights into the impacts of heavy metals exposure on health, reproductive and productive performance of livestock. Front Pharmacol. (2024) 15:1375137. doi: 10.3389/fphar.2024.1375137
319. He M, Wehr U, Rambeck W. Effect of low doses of dietary rare earth elements on growth performance of broilers. J Anim Physiol Anim Nutr. (2010) 94:86–92. doi: 10.1111/j.1439-0396.2008.00884.x
320. Schuller S, Borger C, He ML, Henkelmann R, Jadamus A, Simon O, et al. [The effectiveness of rare earth elements as a possible alternative growth promoter for pigs and poultry]. Berl Munch Tierarztl Wochenschr. (2002) 115:16–23.
321. Nowakiewicz A, Zieba P, Gnat S, Matuszewski Ł. Last call for replacement of antimicrobials in animal production: modern challenges, opportunities, and potential solutions. Antibiotics. (2020) 9:883. doi: 10.3390/antibiotics9120883
322. Roskam JL, Oude Lansink AGJM, Saatkamp HW. The economic value of antimicrobial use in livestock production. Antibiotics. (2023) 12:1537. doi: 10.3390/antibiotics12101537
323. Baudoin F, Hogeveen H, Wauters E. Reducing antimicrobial use and dependence in livestock production systems: a social and economic sciences perspective on an interdisciplinary approach. Front Vet Sci. (2021) 8:584593. doi: 10.3389/fvets.2021.584593
324. Gilbert W. Thomas LF, Coyne L, Rushton J. Review: mitigating the risks posed by intensification in livestock production: the examples of antimicrobial resistance and zoonoses. Animal. (2021) 15:100123. doi: 10.1016/j.animal.2020.100123
325. Borelli E, Ellis K, Pamphilis NM, Tomlinson M, Hotchkiss E. Factors influencing Scottish dairy farmers' antimicrobial usage, knowledge and attitude towards antimicrobial resistance. Prev Vet Med. (2023) 221:106073. doi: 10.1016/j.prevetmed.2023.106073
326. Farrell S, McKernan C, Benson T, Elliott C, Dean M. Understanding farmers' and veterinarians' behavior in relation to antimicrobial use and resistance in dairy cattle: a systematic review. J Dairy Sci. (2021) 104:4584–603. doi: 10.3168/jds.2020-19614
327. Herrmann M, Carlilie C. ‘Narratives of Delay': How the Animal Pharma Industry Resists Moves to Curb the Overuse of Antibiotics on Farms. DeSmog International News (2023). Available online at https://www.desmog.com/2023/12/20/narratives-of-delay-how-the-animal-pharma-industry-resists-moves-to-curb-the-overuse-of-antibiotics-on-farms/ (Accessed June 14, 2025).
328. Magnusson U, Moodley A, Osbjer K. Antimicrobial resistance at the livestock–human interface: implications for veterinary services. Rev Sci Tech. (2021) 40:511–21. doi: 10.20506/rst.40.2.3241
329. Matheou A, Abousetta A, Pascoe AP, Papakostopoulos D, Charalambous L, Panagi S, et al. Antibiotic use in livestock farming: a driver of multidrug resistance? Microorganisms. (2025) 13:779. doi: 10.3390/microorganisms13040779
330. Akram M, Asghar MU, Jalal H. Essential oils as alternatives to chemical feed additives for maximizing livestock production. J Hell Vet Med Soc. (2021) 72:2595–610. doi: 10.12681/jhvms.26741
331. Bengtsson B, Wierup M. Antimicrobial resistance in Scandinavia after a ban of antimicrobial growth promoters. Anim Biotechnol. (2006) 17:147–56. doi: 10.1080/10495390600956920
332. Kaupitwa CJ, Nowaseb S, Godman B, Kibuule D. Analysis of policies for use of medically important antibiotics in animals in Namibia: implications for antimicrobial stewardship. Expert Rev Anti Infect Ther. (2022) 20:1365–79. doi: 10.1080/14787210.2022.2108404
333. WHO. Namibia's Ban on Antibiotics in Healthy Animals Drives Meat Exports. World Health Organization (WHO) (2017). Available online at: https://www.who.int/news-room/feature-stories/detail/namibia-s-ban-on-antibiotics-in-healthy-animals-drives-meat-exports (Accessed April 30, 2024).
334. Casewell M, Friis C, Marco E, McMullin P, Phillips I. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J Antimicrob Chemother. (2003) 52:159–61. doi: 10.1093/jac/dkg313
335. EU Regulation 1831. Ban on Antibiotics as Growth Promoters in Animal Feed Enters into Effect. (2003). Available online at: https://ec.europa.eu/commission/presscorner/detail/en/IP_05_1687 (Accessed January 02, 2023).
336. Van Boeckel TP, Glennon EE, Chen D, Gilbert M, Robinson TP, Grenfell BT, et al. Reducing antimicrobial use in food animals. Science. (2017) 357:1350–2. doi: 10.1126/science.aao1495
337. FDA. Food and Drug Administration (FDA) Reminds Retail Establishments of Upcoming Changes to the Use of Antibiotics in Food Animals, June 20, 2016. (2016). Available online at: http://www.fda.gov/AnimalVeterinary/NewsEvents/CVMUpdates/ucm507355.htm (Accessed January 01, 2023).
338. AGDHR. Australian Government Department of Health (AGDHR). Review of Published and Grey Literature on The Presence of Antimicrobial Resistance in Food in Australia and New Zealand. 2018. (2020). Available online at: https://www.amr.gov.au/file/1402/download (Accessed January 02, 2023).
339. GC. Responsible Use of Medically Important Antimicrobials in Animals. Ottawa, ON: Government of Canada (GC), Department of Health Canada (2024). Available online at:https://www.canada.ca/en/public-health/services/antibiotic-antimicrobial-resistance/animals/actions/responsible-use-antimicrobials.html (Accessed January 12, 2025).
340. MARAPRC. Announcement No. 194 of the Ministry of Agriculture and Rural Affairs of the People's Republic of China (MARAPRC). (2019). Available online at: http://www.xmsyj.moa.gov.cn/zcjd/201907/t20190710_6320678.htm (Accessed January 02, 2023).
341. Wernicki A, Nowaczek A, Urban-Chmiel R. Bacteriophage therapy to combat bacterial infections in poultry. Virol J. (2017) 14:179. doi: 10.1186/s12985-017-0849-7
342. Carrillo CL, Atterbury RJ, El-Shibiny A, Connerton PL, Dillon E, Scott A, et al. Bacteriophage therapy to reduce Campylobacter jejuni colonization of broiler chickens. Appl Environ Microbiol. (2005) 71:6554–63. doi: 10.1128/AEM.71.11.6554-6563.2005
343. Huff WE, Huff GR, Rath NC, Balog JM, Donoghue AM. Therapeutic efficacy of bacteriophage and Baytril (enrofloxacin) individually and in combination to treat colibacillosis in broilers. Poult Sci. (2004) 83:1944–7. doi: 10.1093/ps/83.12.1944
344. Ngu NT, Phuong LNN, Anh LH, Loc HT, Tam NT, Huan PKN, et al. The efficiency of bacteriophages against Salmonella Typhimurium infection in native noi broilers. Braz J Poult Sci. (2022) 24. doi: 10.1590/1806-9061-2021-1561
345. Wall Samantha K, Zhang J, Rostagno Marcos H, Ebner Paul D. Phage therapy to reduce preprocessing Salmonella infections in market-weight swine. Appl Environ Microbiol. (2010) 76:48–53. doi: 10.1128/AEM.00785-09
346. Xu Z, Jin P, Zhou X, Zhang Y, Wang Q, Liu X, et al. Isolation of a virulent Aeromonas salmonicida subsp. masoucida bacteriophage and its application in phage therapy in turbot (Scophthalmus maximus). Appl Environ Microbiol. (2021) 87:e01468-21. doi: 10.1128/AEM.01468-21
347. Yan L, Hong SM, Kim IH. Effect of bacteriophage supplementation on the growth performance, nutrient digestibility, blood characteristics, and fecal microbial shedding in growing pigs. Asian-Austral J Anim Sci. (2012) 25:1451–6. doi: 10.5713/ajas.2012.12253
348. Kim JS, Hosseindoust A, Lee SH, Choi YH, Kim MJ, Lee JH, et al. Bacteriophage cocktail and multi-strain probiotics in the feed for weanling pigs: effects on intestine morphology and targeted intestinal coliforms and Clostridium. Animal. (2017) 11:45–53. doi: 10.1017/S1751731116001166
349. Thanki AM, Mignard G, Atterbury RJ, Barrow P, Millard AD, Clokie MRJ. Prophylactic delivery of a bacteriophage cocktail in feed significantly reduces Salmonella colonization in pigs. Microbiol Spectr. (2022) 10:e00422-22. doi: 10.1128/spectrum.00422-22
350. Peh E, Szott V, Reichelt B, Friese A, Rösler U, Plötz M, et al. Bacteriophage cocktail application for Campylobacter mitigation- from in vitro to in vivo. BMC Microbiol. (2023) 23:209. doi: 10.1186/s12866-023-02963-1
351. Bechinger B, Gorr SU. Antimicrobial peptides: mechanisms of action and resistance. J Dent Res. (2017) 96:254–60. doi: 10.1177/0022034516679973
352. Yi T, Huang Y, Chen Y. Production of an antimicrobial peptide AN5-1 in Escherichia coli and its dual mechanisms against bacteria. Chem Biol Drug Des. (2015) 85:598–607. doi: 10.1111/cbdd.12449
353. Yi H, Zhang L, Gan Z, Xiong H, Yu C, Du H, et al. High therapeutic efficacy of Cathelicidin-WA against postweaning diarrhea via inhibiting inflammation and enhancing epithelial barrier in the intestine. Sci Rep. (2016) 6:25679. doi: 10.1038/srep25679
354. Pato U, Riftyan E, Ayu DF, Jonnaidi NN, Wahyuni MS, Feruni JA, et al. Antibacterial efficacy of lactic acid bacteria and bacteriocin isolated from Dadih's against Staphylococcus aureus. Food Sci Technol. (2021) 42:e27121. doi: 10.1590/fst.27121
355. Silveira RF, Roque-Borda CA, Vicente EF. Antimicrobial peptides as a feed additive alternative to animal production, food safety and public health implications: an overview. Anim Nutr. (2021) 7:896–904. doi: 10.1016/j.aninu.2021.01.004
356. Gao F, Ahmed A, Cong H, Yu B, Shen Y. Effective strategies for developing potent, broad-spectrum antibacterial and wound healing promotion from short-chain antimicrobial peptides. ACS Appl Mater Interfaces. (2023) 15:32136–47. doi: 10.1021/acsami.3c03069
357. dos Santos Clemente SM, Lúcio ÉC, Barros MR, de Oliveira Vilela SM, Junior JWP. Virulence genes and resistance profile of Escherichia coli isolated in meat meal samples. Acta Sci Vet. (2021) 49:114233. doi: 10.22456/1679-9216.114233
358. Udenigwe CC, Fogliano V. Food matrix interaction and bioavailability of bioactive peptides: two faces of the same coin? J Funct Foods. (2017) 35:9–12. doi: 10.1016/j.jff.2017.05.029
359. Wibowo D, Zhao C-X. Recent achievements and perspectives for large-scale recombinant production of antimicrobial peptides. Appl Microbiol Biotechnol. (2019) 103:659–71. doi: 10.1007/s00253-018-9524-1
360. Azhar J, Nadeem A, Javed M, Ahmad HI, Hassan F-u, Shah FS. Evaluation of phytochemicals from Thymus serpyllum as potential drug candidates to manage oxidative stress in transition dairy cows. J Biomol Struct Dyn. (2024) 42:2897–912. doi: 10.1080/07391102.2023.2209190
361. Rivera-Chacon R, Castillo-Lopez E, Ricci S, Petri RM, Reisinger N, Zebeli Q. Supplementing a phytogenic feed additive modulates the risk of subacute rumen acidosis, rumen fermentation and systemic inflammation in cattle fed acidogenic diets. Animals. (2022) 12:1201. doi: 10.3390/ani12091201
362. Bagheri Varzaneh M. Effects of a commercial blend of phytogenic compounds and prebiotic on the performance of mid-lactation dairy cows exposed to heat-stress. Iran J Appl Anim Sci. (2022) 12:489–95. Available online at: https://journals.iau.ir/article_694583.html
363. Castillo-Lopez E, Rivera-Chacon R, Ricci S, Reisinger N, Zebeli Q. Changes in fermentation profile of the reticulorumen and hindgut, and nutrient digestion in dry cows fed concentrate-rich diets supplemented with a phytogenic feed additive. J Dairy Sci. (2022) 105:5747–60. doi: 10.3168/jds.2022-21786
364. Ricci S, Pacífico C, Castillo-Lopez E, Rivera Chacon R, Schwartz H, Reisinger N, et al. Progressive microbial adaptation of the bovine rumen and hindgut in response to a step-wise increase in dietary starch and the influence of phytogenic supplementation. Front Microbiol. (2022) 13:920427. doi: 10.3389/fmicb.2022.920427
365. Lee-Rangel HA, Mendoza-Martinez GD, Martínez-García JA, Espinosa-Ayala E, Hernández-García PA, Cifuentes-López RO, et al. An Indian polyherbal phytogenic source improved blood serum biochemistry and immune response of dairy calves. Food Agric Immunol. (2022) 33:97–112. doi: 10.1080/09540105.2021.2024150
366. Molosse VL, Deolindo GL, Glombosky P, Pereira WAB, Carvalho RA, Zotti CA, et al. Curcumin or microencapsulated phytogenic blend to replace ionophore and non-ionophore antibiotics in weaned calves: effects on growth performance and health. Livest Sci. (2022) 263:105029. doi: 10.1016/j.livsci.2022.105029
367. Bajagai YS, Petranyi F, Yu SJ, Lobo E, Batacan R, Kayal A, et al. Phytogenic supplement containing menthol, carvacrol and carvone ameliorates gut microbiota and production performance of commercial layers. Sci Rep. (2022) 12:1–13. doi: 10.1038/s41598-022-14925-0
368. Lyte JM, Arsi K, Caputi V, Liyanage R, Assumpcao ALFV, Jesudhasan PR, et al. Inclusion of trans-cinnamaldehyde and caprylic acid in feed results in detectable concentrations in the chicken gut and reduces foodborne pathogen carriage. Poult Sci. (2024) 103:103368. doi: 10.1016/j.psj.2023.103368
369. Li T, Shen MM, Hou QR, Zhang SL, Huang HL, Guo P, et al. Effects of phytogenic feed on productive performance, egg quality, antioxidant activity and lipid metabolism of laying hens. J Anim Feed Sci. (2022) 32:50–8. doi: 10.22358/jafs/154977/2022
370. Rahman A, Bayram I, Gultepe EE. Effect of mentha on performance, haematological and biochemical parameters in laying hens. S Afr J Anim Sci. (2021) 51:221–30. doi: 10.4314/sajas.v51i2.10
371. Liu HN, Liu Y, Hu LL, Suo YL, Zhang L, Jin F, et al. Effects of dietary supplementation of quercetin on performance, egg quality, cecal microflora populations, and antioxidant status in laying hens. Poult Sci. (2014) 93:347–53. doi: 10.3382/ps.2013-03225
372. Alagawany M, El-Hack A, Mohamed E, El-Kholy MS. Productive performance, egg quality, blood constituents, immune functions, and antioxidant parameters in laying hens fed diets with different levels of Yucca schidigera extract. Environ Sci Pollut Res. (2016) 23:6774–82. doi: 10.1007/s11356-015-5919-z
373. Mutlu MIS, Yildirim A. Effect of dietary supplementation of Panax ginseng leaf extract on production performance and egg quality of hens at the beginning of their laying period. Large Anim Rev. (2020) 26:341–8.
374. Abad P, Arroyo-Manzanares N, Ariza JJ, Baños A, García-Campaña AM. Effect of Allium extract supplementation on egg quality, productivity, and intestinal microbiota of laying hens. Animals. (2020) 11:41. doi: 10.3390/ani11010041
375. Peng M-j, Huang T, Yang Q-l, Peng S, Jin Y-x, Wang X-s. Dietary supplementation Eucommia ulmoides extract at high content served as a feed additive in the hens industry. Poult Sci. (2022) 101:101650. doi: 10.1016/j.psj.2021.101650
376. Negri Brusamarello M, Brusamarello AP, Alves da Cunha MA, Busso C. Biological and phytochemical potential of Baccharis trimera (Less.) DC leaf extract on swine clinical isolates. Nat Prod Res. (2024) 38:1799–805. doi: 10.1080/14786419.2023.2222217
377. Chang SY, Song MH, Lee JH, Oh HJ, Kim YJ, An JW, et al. Phytogenic feed additives alleviate pathogenic Escherichia coli-induced intestinal damage through improving barrier integrity and inhibiting inflammation in weaned pigs. J Anim Sci Biotechnol. (2022) 13:1–12. doi: 10.1186/s40104-022-00750-y
378. Le Coz J, Ilic S, Fibi-Smetana S, Schatzmayr G, Zaunschirm M, Grenier B. Exploring with transcriptomic approaches the underlying mechanisms of an essential oil-based phytogenic in the small intestine and liver of pigs. Front Vet Sci. (2021) 8:650732. doi: 10.3389/fvets.2021.650732
379. Shili CN, Habibi M, Sutton J, Barnes J, Burch-Konda J, Pezeshki A. Effect of a phytogenic water additive on growth performance, blood metabolites and gene expression of amino acid transporters in nursery pigs fed with low-protein/high-carbohydrate diets. Animals. (2021) 11:555. doi: 10.3390/ani11020555
380. Moreira VS, Garbossa CAP, Guimarães EBB, Hirai WY, da Cruz TA, Alves LKS, et al. Evaluation of a phytogenic compound with minerals as a possible alternative to ractopamine for finishing pigs. Animals. (2022) 12:2311. doi: 10.3390/ani12182311
381. Nowland TL, Stanley D, Kirkwood RN, Torok VA, Bajagai YS, Gannon NJ, et al. Maternal supplementation with phytogenic additives influenced the faecal microbiota and reproductive potential in sows. AMB Express. (2021) 11:1–11. doi: 10.1186/s13568-021-01268-8
382. Caprarulo V, Turin L, Hejna M, Reggi S, Dell'Anno M, Riccaboni P, et al. Protective effect of phytogenic plus short and medium-chain fatty acids-based additives in enterotoxigenic Escherichia coli challenged piglets. Vet Res Commun. (2022) 47:217–31. doi: 10.1007/s11259-022-09945-0
383. Papatsiros VG, Katsogiannou EG, Papakonstantinou GI, Michel A, Petrotos K, Athanasiou LV. Effects of phenolic phytogenic feed additives on certain oxidative damage biomarkers and the performance of primiparous sows exposed to heat stress under field conditions. Antioxidants. (2022) 11:593. doi: 10.3390/antiox11030593
384. Hanuman DDV, Rao K, Nagarjuna G, Mounika K, Debbarma B, Sravathi V, et al. Antioxidant effect of lemongrass (Cymbopogon citratus) extract on paraquat-induced oxidative stress in sheep red blood cells using in-vitro phytochemical analysis. J Adv Biol Biotechnol. (2024) 27:730–8. doi: 10.9734/jabb/2024/v27i5835
385. Muñoz-Cuautle A, Ortega-Cerrilla ME, Herrera-Haro JG, Nava-Cuellar C, Gutiérrez-Olvera C, Ramírez-Bribiesca JE, et al. Effect of oregano (Lippia graveolens) essential oil as a phytogenic feed additive on productive performance, ruminal fermentation, and antioxidant activity in lamb meat. Agriculture. (2022) 12:973. doi: 10.3390/agriculture12070973
386. Hashemzadeh F, Rafeie F, Hadipour A, Rezadoust MH. Supplementing a phytogenic-rich herbal mixture to heat-stressed lambs: growth performance, carcass yield, and muscle and liver antioxidant status. Small Rumin Res. (2022) 206:106596. doi: 10.1016/j.smallrumres.2021.106596
387. El-Zaiat HM, Elshafie EI, Al-Marzooqi W, Dughaishi KA. Effects of Neem (Azadirachta indica) leaf powder supplementation on rumen fermentation, feed intake, apparent digestibility and performance in omani sheep. Animals. (2022) 12:3146. doi: 10.3390/ani12223146
388. Férrer JP, da Cunha MV, dos Santos MVF, Torres TR, da Silva JRC, Véras RML, et al. Mesquite (Prosopis juliflora) extract as a phytogenic additive for sheep finished on pasture in the semiarid region. Chil J Agric Res. (2021) 81:14–26doi: 10.4067/S0718-58392021000100014
389. Pereira MLA, Sousa LB, Tomich TR, Machado FS, Sousa LB, Pereira LGR. Methane mitigation potential of diets supplemented with mesquite piperidine alkaloids for sheep. In: EAAP Scientific Series. Wageningen: Wageningen Academic Publishers (2019). p. 4301–11. doi: 10.3920/978-90-8686-891-9_22
390. Rabee AE, Mohamed M, Ghandour M, Sallam A, Elwakeel EA, Mohammed RS, et al. Rumen fermentation and microbiota in Shami goats fed on condensed tannins or herbal mixture. BMC Vet Res. (2024) 20:35. doi: 10.1186/s12917-024-03887-2
391. Soltan YA, Morsy AS, Hashem NM, Sallam SM. Boswellia sacra resin as a phytogenic feed supplement to enhance ruminal fermentation, milk yield, and metabolic energy status of early lactating goats. Anim Feed Sci Technol. (2021) 277:114963. doi: 10.1016/j.anifeedsci.2021.114963
392. Adebayo K, Ojo V, Aderinboye R, Ajadi I, Hammed K. Vernonia amygdalina (bitter leaf) as a phytogenic additive in the diet of West African Dwarf goats. Trop Agric. (2022) 99:47–56. Available online at: https://journals.sta.uwi.edu/ojs/index.php/ta/article/view/7248
393. Jiwuba PC, Amaduruonye W, Okoye LE, Obasi KU. Haematology and serum biochemical profile of West African Dwarf Goats fed four phytogenic browse plant parts in their diets. Niger J Anim Sci Technol. (2021) 4:72–81.
394. Jiwuba PC, Azodo LN, Uzoma C. Nutritional and phytogenic properties of soursop (Annona muricata) leaf meal on the productive and physiological performances of West African Dwarf Goats. Zhivotnovadni Nauki. (2020) 57:20–8. Available online at: https://www.cabidigitallibrary.org/doi/pdf/10.5555/20203289821
395. Adebayo KO, Aderinboye RY, Sanwo KA, Oyewusi IK, Isah OA. Microbial population and blood parameters of West African dwarf goats fed scent leaf (Ocimum gratissimum) as additive. Niger J Anim Prod. (2019) 46:225–35. doi: 10.51791/njap.v46i1.2495
396. da Silva NIS, de Moura JFP, de Lucena Nascimento ME, Machado FCG, Costa TGP, de Araújo Filho JM, et al. Effect of Lippia alba hay as phytogenic feed additive on the lactation performance, milk composition, and rumen and blood parameters of Alpine goats. Small Rumin Res. (2022) 215:106767. doi: 10.1016/j.smallrumres.2022.106767
397. Park J, Kwak M-J, Kang M-G, Cho D-Y, Kim JN, Choi I-G, et al. Metabolic-methane mitigation by combination of probiotic Escherichia coli strain Nissle 1917 and biochar in rumen fluid in vitro fermentation of dairy cow. J Environ Chem Eng. (2024) 12:113977. doi: 10.1016/j.jece.2024.113977
398. Monteiro H, Lelis A, Fan P, Agustinho BC, Lobo R, Arce-Cordero J, et al. Effects of lactic acid-producing bacteria as direct-fed microbials on the ruminal microbiome. J Dairy Sci. (2022) 105:2242–55. doi: 10.3168/jds.2021-21025
399. Murray SA, Holbert AC, Norman KN, Lawhon SD, Sawyer JE, Scott HM. Effects of a direct-fed microbial and feedlot pen environmental change on phenotypic resistance among enterococci isolated from beef cattle feces. Preprints. (2021). doi: 10.20944/preprints202111.0474.v1
400. Monteiro HF, Agustinho BC, Vinyard JR, Harden T, Bennett SL, Arce-Cordero JA, et al. Megasphaera elsdenii and Saccharomyces Cerevisiae as direct fed microbials during an in vitro acute ruminal acidosis challenge. Sci Rep. (2022) 12:1–13. doi: 10.1038/s41598-022-11959-2
401. Pan L, Harper K, Queiroz O, Copani G, Cappellozza BI. Effects of a Bacillus-based direct-fed microbial on in vitro nutrient digestibility of forage and high-starch concentrate substrates. Transl Anim Sci. (2022) 6:txac067. doi: 10.1093/tas/txac067
402. Neujahr A, Doan T, Kubwimana F, Anderson C, Fernando S. PSIV-6 engineering microbes to develop novel direct fed microbials as an alternative to prophylactic antibiotic use in beef cattle production. J Anim Sci. (2022) 100(Supplement_2):188–9. doi: 10.1093/jas/skac064.317
403. Mansilla FI, Ficoseco CA, Miranda MH, Puglisi E, Nader-Macías MEF, Vignolo GM, et al. Administration of probiotic lactic acid bacteria to modulate fecal microbiome in feedlot cattle. Sci Rep. (2022) 12:1–17. doi: 10.1038/s41598-022-16786-z
404. Batista L, Cidrini I, Prados L, Cruz A, Torrecilhas J, Siqueira G, et al. A meta-analysis of yeast products for beef cattle under stress conditions: performance, health and physiological parameters. Anim Feed Sci Technol. (2022) 283:115182. doi: 10.1016/j.anifeedsci.2021.115182
405. Khalil F, Korany AM, Abdel Aziz S, Emeash H, Abdelghany AK. Comparing the efficacy of Saccharomyces and Lactobacillus probiotics on feeding behavior and milk production of dairy cattle. J Vet Med Res. (2022) 29:21–6. doi: 10.21608/jvmr.2022.123351.1051
406. Merati Z, Towhidi A. Effect of a multispecies probiotics on productive and reproductive performance of Holstein cows. Iran J Appl Anim Sci. (2022) 12:237–47.
407. Smith ZK, Karges K, Aguilar A. Evaluation of an active live yeast (Levucell Saccharomyces cerevisiae, CNCM l-1077) on receiving and backgrounding period growth performance and efficiency of dietary net energy utilization in low health risk beef steers. Transl Anim Sci. (2020) 4:txaa127. doi: 10.1093/tas/txaa127
408. Ovinge LA, Sarturi JO, Galyean ML, Ballou MA, Trojan SJ, Campanili PR, et al. Effects of a live yeast in natural-program finishing feedlot diets on growth performance, digestibility, carcass characteristics, and feeding behavior. J Anim Sci. (2018) 96:684–93. doi: 10.1093/jas/sky011
409. Cagle CM, Fonseca MA, Callaway TR, Runyan CA, Cravey MD, Tedeschi LO. Evaluation of the effects of live yeast on rumen parameters and in situ digestibility of dry matter and neutral detergent fiber in beef cattle fed growing and finishing diets. Appl Anim Sci. (2020) 36:36–47. doi: 10.15232/aas.2019-01888
410. Liu S, Shah AM, Yuan M, Kang K, Wang Z, Wang L, et al. Effects of dry yeast supplementation on growth performance, rumen fermentation characteristics, slaughter performance and microbial communities in beef cattle. Anim Biotechnol. (2021) 33:1150–60. doi: 10.1080/10495398.2021.1878204
411. Kumprechtová D, Illek J, Julien C, Homolka P, Jančík F, Auclair E. Effect of live yeast (Saccharomyces cerevisiae) supplementation on rumen fermentation and metabolic profile of dairy cows in early lactation. J Anim Physiol Anim Nutr. (2019) 103:447–55. doi: 10.1111/jpn.13048
412. Arik HD, Gulsen N, Hayirli A, Alatas MS. Efficacy of Megasphaera elsdenii inoculation in subacute ruminal acidosis in cattle. J Anim Physiol Anim Nutr. (2019) 103:416–26. doi: 10.1111/jpn.13034
413. Gao J, Liu Y-C, Wang Y, Li H, Wang X-M, Wu Y, et al. Impact of yeast and lactic acid bacteria on mastitis and milk microbiota composition of dairy cows. AMB Express. (2020) 10:1–12. doi: 10.1186/s13568-020-0953-8
414. Machado MO, Santana DAD, de Carvalho MB, Ollhoff RD, Weber SH, Sotomaior CS. Performance, ruminal and fecal microbiome of lambs fed diets supplemented with probiotics. Trop Anim Health Prod. (2024) 56:319. doi: 10.1007/s11250-024-04161-1
415. Mavrommatis A, Mitsiopoulou C, Christodoulou C, Karabinas D, Nenov V, Zervas G, et al. Dietary supplementation of a live yeast product on dairy sheep milk performance. Oxidative and immune status in peripartum period. J Fungi. (2020) 6:334. doi: 10.3390/jof6040334
416. Liu C, Hou Y, Su R, Luo Y, Dou L, Yang Z, et al. Effect of dietary probiotics supplementation on meat quality, volatile flavor compounds, muscle fiber characteristics, and antioxidant capacity in lambs. Food Sci Nutr. (2022) 10:2646–58. doi: 10.1002/fsn3.2869
417. Nie C, Hu Y, Chen R, Guo B, Li L, Chen H, et al. Effect of probiotics and Chinese medicine polysaccharides on meat quality, muscle fibre type and intramuscular fat deposition in lambs. Ital J Anim Sci. (2022) 21:811–20. doi: 10.1080/1828051X.2022.2067489
418. Mousa GA, Allak MA, Shehata MG, Hashem NM, Hassan OGJA. Dietary supplementation with a combination of fibrolytic enzymes and probiotics improves digestibility, growth performance, blood metabolites, and economics of fattening lambs. Animals. (2022) 12:476. doi: 10.3390/ani12040476
419. Mohamed M, Abdou S, Hassan E, Suliman A. Effect of probiotics supplementation on productive performance of growing lambs. Arch Agric Sci J. (2022) 5:21–33. doi: 10.21608/aasj.2022.123927.1108
420. Hassan A, Gado H, Anele UY, Berasain MA, Salem AZJAB. Influence of dietary probiotic inclusion on growth performance, nutrient utilization, ruminal fermentation activities and methane production in growing lambs. Anim Biotchnol. (2020) 31:365–72. doi: 10.1080/10495398.2019.1604380
421. Ndegwa E, Elhadedy DE, Richey C, Kim C, Yousuf AB Differential age-based response induced by a commercial probiotic supplementation in pastured goats. Probiotics Antimicrob Proteins. (2024) 1–15. doi: 10.1007/s12602-024-10337-w
422. Viana MGS, Vasconcelos DKM, Campos MIF, de Carvalho LM, Olegário LS, de Sousa Galvão M, et al. Improving the physicochemical and sensory properties and volatile profile of goat ricotta cream with Limosilactobacillus mucosae CNPC007 supplementation. Int Dairy J. (2024) 157:106011. doi: 10.1016/j.idairyj.2024.106011
423. Liu D, Zhao F, Li L, Zhang J, Wu S, Lü X, et al. Enhancing the antioxidant capacity and quality attributes of fermented goat milk through the synergistic action of Limosilactobacillus fermentum WXZ 2-1 with a starter culture. J Dairy Sci. (2024) 107:1928–49. doi: 10.3168/jds.2023-24135
424. Zuckermann FA, Husmann R, Chen W, Roady P, Pfeiff J, Leistikow KR, et al. Bacillus-based direct-fed microbial reduces the pathogenic synergy of a coinfection with Salmonella enterica, Serovar choleraesuis and porcine reproductive and respiratory syndrome virus. Infect Immun. (2022) 90:e00574-21. doi: 10.1128/iai.00574-21
425. González F, Cervantes M, Morales A, Valle JA, Camacho Reyna L, ngélica Morales-Becerra A, et al. Effect of supplementing a Bacillus subtilis-based probiotic on performance, intestinal integrity, and serum antioxidant capacity and metabolites concentrations of heat-stressed growing pigs. J Anim Sci. (2024) 102:skae012. doi: 10.1093/jas/skae012
426. Lewton JR, Woodward AD, Moser RL, Thelen KM, Moeser AJ, Trottier NL, et al. Effects of a multi-strain Bacillus subtilis-based direct-fed microbial on weanling pig growth performance and nutrient digestibility. Transl Anim Sci. (2021) 5:txab058. doi: 10.1093/tas/txab058
427. Magnoli AP, Parada J, de la Torre FC, Watson S, Poloni V, Fochesato A, et al. Respiratory tract clinometry, fat thickness, haematology and productive parameters associated with direct-fed microbials used as growth promoter antibiotic alternative in weaned piglets. Vet Anim Sci. (2022) 16:100246. doi: 10.1016/j.vas.2022.100246
428. Nguyen VTH, Tung QN, Lien BT, Trang NH. Efficacy of biosynthesizing folate, riboflavin and typical probiotic traits reveal the potential use of Lactobacillus plantarum LCN13 as a feed additive for swine farming. Acad J Biol. (2022) 44:73–82. doi: 10.15625/2615-9023/16628
429. Sudan S, Zhan X, Li JJM. A novel probiotic Bacillus subtilis strain confers cytoprotection to host pig intestinal epithelial cells during Enterotoxic Escherichia coli infection. Microbiol Spectr. (2022) 10:e01257-21. doi: 10.1128/spectrum.01257-21
430. Sarkar VK, De UK, Kala A, Verma AK, Chauhan A, Paul BR, et al. Early-life intervention of lactoferrin and probiotic in suckling piglets: effects on immunoglobulins, intestinal integrity, and neonatal mortality. Probiotics Antimicrob Proteins. (2022) 15:149–59. doi: 10.1007/s12602-022-09964-y
431. Palkovicsné Pézsa N, Kovács D, Gálfi P, Rácz B, Farkas OJN. Effect of Enterococcus faecium NCIMB 10415 on gut barrier function, internal redox state, proinflammatory response and pathogen inhibition properties in porcine intestinal epithelial cells. Nutrients. (2022) 14:1486. doi: 10.3390/nu14071486
432. Xin J, Zeng D, Wang H, Sun N, Zhao Y, Dan Y, et al. Probiotic Lactobacillus johnsonii BS15 promotes growth performance, intestinal immunity, and gut microbiota in piglets. Probiotics Antimicrob Proteins. (2020) 12:184–93. doi: 10.1007/s12602-018-9511-y
433. Yang J, Wang C, Huang K, Zhang M, Wang J, Pan XJAM, et al. Compound Lactobacillus sp. administration ameliorates stress and body growth through gut microbiota optimization on weaning piglets. Appl Microbiol Biotechnol. (2020) 104:6749–65. doi: 10.1007/s00253-020-10727-4
434. Mohammed AA, Mahmoud MA, Zaki RS, Cheng HW. Effect of a probiotic supplement (Bacillus subtilis) on struggling behavior, immune response, and meat quality of shackled broiler chickens exposed to preslaughter stress. Poult Sci. (2024) 103:104051. doi: 10.1016/j.psj.2024.104051
435. Forte C, Manuali E, Abbate Y, Papa P, Vieceli L, Tentellini M, et al. Dietary Lactobacillus acidophilus positively influences growth performance, gut morphology, and gut microbiology in rurally reared chickens. Poult Sci. (2018) 97:930–6. doi: 10.3382/ps/pex396
436. Wu Y, Wang B, Zeng Z, Liu R, Tang L, Gong L, et al. Effects of probiotics Lactobacillus plantarum 16 and Paenibacillus polymyxa 10 on intestinal barrier function, antioxidative capacity, apoptosis, immune response, and biochemical parameters in broilers. Microbiol Food Safety. (2019) 98:5028–39. doi: 10.3382/ps/pez226
437. Wu S, Liu Y, Duan Y, Wang F, Guo F, Yan F, et al. Intestinal toxicity of deoxynivalenol is limited by supplementation with Lactobacillus plantarum JM113 and consequentially altered gut microbiota in broiler chickens. J Anim Sci Biotechnol. (2018) 9:1–13. doi: 10.1186/s40104-018-0286-5
438. Oladokun S, Koehler A, MacIsaac J, Ibeagha-Awemu EM, Adewole DI. Bacillus subtilis delivery route: effect on growth performance, intestinal morphology, cecal short-chain fatty acid concentration, and cecal microbiota in broiler chickens. Poult Sci. (2021) 100:100809. doi: 10.1016/j.psj.2020.10.063
439. Bae D, Kim D-H, Chon J-W, Song K-Y, Seo K-HJPS. Synergistic effects of the early administration of Lactobacillus kefiranofaciens DN1 and Kluyveromyces marxianus KU140723-05 on the inhibition of Salmonella Enteritidis colonization in young chickens. Micobiol Food Safety. (2020) 99:5999–6006. doi: 10.1016/j.psj.2020.07.032
440. Lagounova M. The Use of Dietary Probiotics in Horses: Microbial Activity During Antibiotic Treatment and Bacterial Characterization [Master of Science Dissertation]. University of Guelph, Guelph (2022).
441. Jacobs RD, Grum D, Trible B, Ayala DI, Karnezos TP, Gordon ME. Oral probiotic administration attenuates postexercise inflammation in horses. Transl Anim Sci. (2024) 8:txae124. doi: 10.1093/tas/txae124
442. Ivannikova R, Smirnova E, Navruzshoeva G, Pavlova A, Kalinin I. Effect of the feed additive ProVetin on the development of foals. E3S Web Conf. (2024) 494:04020. doi: 10.1051/e3sconf/202449404020
443. Šebková A, Zábranský L, Vachoušková J, Brabenec V, Polívková D, Sezimová K, et al. Importance of prebiotic, probiotic and phytobiotic feed supplements in calf nutrition. Nutrinet. (2024) 2024:221. doi: 10.11118/978-80-7509-994-5-0221
444. Grispoldi L, Bertero F, Franceschini S, Mastrosimone F, Sechi P, Iulietto MF, et al. Prevalence and characterisation of shigatoxigenic Escherichia coli isolated from beef cattle fed with prebiotics. Ital J Food Safety. (2017) 6:6958. doi: 10.4081/ijfs.2017.6958
445. Boushehri M, Sadeghi AA, Chamani M, Aminafshar M. Effects of antioxidants and prebiotics as vegetable pellet feed on production performance, hematological parameters and colostrum immunoglobulin content in transition dairy cows. Ital J Anim Sci. (2021) 20:1863–9. doi: 10.1080/1828051X.2021.1987158
446. Raza M, Yousaf MS, Ahmad J, Rashid MA, Majeed KA, Tahir SK, et al. Prebiotics supplementation modulates pre-weaning stress in male cattle calves by improving growth performance, health scores and serum biomarkers. Czech J Anim Sci. (2022) 67:102–13. doi: 10.17221/70/2021-CJAS
447. Baines D, Erb S, Lowe R, Turkington K, Sabau E, Kuldau G, et al. A prebiotic, Celmanax™, decreases Escherichia coli O157: H7 colonization of bovine cells and feed-associated cytotoxicity in vitro. BMC Res Notes. (2011) 4:1–13. doi: 10.1186/1756-0500-4-110
448. Heim G, Sweeney T, O'Shea CJ, Doyle DN, O'doherty JV. Effect of maternal dietary supplementation of laminarin and fucoidan, independently or in combination, on pig growth performance and aspects of intestinal health. Anim Feed Sci Technol. (2015) 204:28–41. doi: 10.1016/j.anifeedsci.2015.02.007
449. Bouwhuis MA, Sweeney T, Mukhopadhya A, McDonnell MJ, O'Doherty JV. Maternal laminarin supplementation decreases Salmonella Typhimurium shedding and improves intestinal health in piglets following an experimental challenge with S. Typhimurium post-weaning. Anim Feed Sci Technol. (2017) 223:156–68. doi: 10.1016/j.anifeedsci.2016.11.007
450. Bouwhuis MA, Sweeney T, Mukhopadhya A, Thornton K, McAlpine PO, O'Doherty JV. Zinc methionine and laminarin have growth-enhancing properties in newly weaned pigs influencing both intestinal health and diarrhoea occurrence. J Anim Physiol Anim Nutr. (2017) 101:1273–85. doi: 10.1111/jpn.12647
451. Rattigan R, Sweeney T, Maher S, Thornton K, Rajauria G, O'Doherty JV. Laminarin-rich extract improves growth performance, small intestinal morphology, gene expression of nutrient transporters and the large intestinal microbial composition of piglets during the critical post-weaning period. Br J Nutr. (2020) 123:255–63. doi: 10.1017/S0007114519002678
452. Vigors S, O'Doherty JV, Rattigan R, McDonnell MJ, Rajauria G, Sweeney T. Effect of a laminarin rich macroalgal extract on the caecal and colonic microbiota in the post-weaned pig. Mar Drugs. (2020) 18:157. doi: 10.3390/md18030157
453. Ma H, Abubaker MA, Al-Wraikat M, Li L, Liu Y. Effect of co-fermentation with non-digestible oligosaccharides on the quality of fermented goat milk: physicochemical properties, microbial composition, and small intestine modulations in mice. Food Biosci. (2024) 61:104878. doi: 10.1016/j.fbio.2024.104878
454. Angulo M, Reyes-Becerril M, Cepeda-Palacios R, Tovar-Ramírez D, Esteban MÁ, Angulo C. Probiotic effects of marine Debaryomyces hansenii CBS 8339 on innate immune and antioxidant parameters in newborn goats. Appl Microbiol Biotechnol. (2019) 103:2339–52. doi: 10.1007/s00253-019-09621-5
455. da Silva CI, Schneider CR, Hygino B, Duarte V, Teixeira UHG, Alcalde CR, et al. Performance, carcass characteristics, and meat quality of goat kids supplemented with inulin. Livest Sci. (2022) 265:105094. doi: 10.1016/j.livsci.2022.105094
456. Chen K, Liu Y, Cheng Y, Yan Q, Zhou C, He Z, et al. Supplementation of Lactobacillus plantarum or Macleaya cordata extract alleviates oxidative damage induced by weaning in the lower gut of young goats. Animals. (2020) 10:548. doi: 10.3390/ani10040548
457. Mosa M, Salem H, Hassan M, El-Saied M, Bastamy M, Amer M. Application of 16S rRNA gene sequencing in evaluation of prebiotics or probiotics administration to restore gut dysbiosis induced by infectious bursal disease virus in broiler chickens. Ger J Vet Res. (2024) 4:86–99. doi: 10.51585/gjvr.2024.3.0101
458. Tavaniello S, Maiorano G, Stadnicka K, Mucci R, Bogucka J, Bednarczyk M. Prebiotics offered to broiler chicken exert positive effect on meat quality traits irrespective of delivery route. Poult Sci. (2018) 97:2979–87. doi: 10.3382/ps/pey149
459. Vasconcelos RAM, Kalschne DL, Wochner KF, Moreira MCC, Becker-Algeri TA, Centenaro AI, et al. Feasibility of L. plantarum and prebiotics on Aflatoxin B 1 detoxification in cow milk. Food Sci Technol. (2020) 41:627–32. doi: 10.1590/fst.34120
460. Abdennebi I, Messai CR, Ouchene N, Ouchene-Khelifi NA, Ghallache L, Ait-Oudhia K, et al. Symbiotic effect on some microbiological species and physicochemical properties in milk in subclinical mastitis of dairy cows. Agric Sci Technol. (2020) 12:8. doi: 10.15547/ast.2020.04.058
461. Colombo EA, Cooke RF, Brandão AP, Wiegand JB, Schubach KM, Sowers CA, et al. Performance, health, and physiological responses of newly received feedlot cattle supplemented with pre-and probiotic ingredients. Animal. (2021) 15:100214. doi: 10.1016/j.animal.2021.100214
462. Pickett A, Cooke RF, Colombo E, Batista LF, Brandão AP. Effects of supplementing pre-and pro-biotic feed ingredients on performance, health, and physiological responses in feedlot cattle. J Anim Sci. (2023) 101(Supplement_1):31–2. doi: 10.1093/jas/skad068.037
463. Rodríguez-Mínguez E, Calzada J, Sánchez C, Vázquez M, Ávila M, Garde S, et al. Symbiotic sheep milk cheese containing Moringa oleifera extract and Bifidobacterium pseudolongum INIA P2. Int J Food Microbiol. (2025) 427:110942. doi: 10.1016/j.ijfoodmicro.2024.110942
464. Zhou Z, Xu X, Luo D, Zhou Z, Zhang S, He R, et al. Effect of dietary supplementation of Lactiplantibacillus plantarum N-1 and its synergies with Oligomeric isomaltose on the growth performance and meat quality in Hu sheep. Foods. (2023) 12:1858. doi: 10.3390/foods12091858
465. Kazemi-Bonchenari M, Ghasemi HA, Khodaei-Motlagh M, Khaltabadi-Farahani AH, Ilani M. Influence of feeding synbiotic containing Enterococcus faecium and inulin on blood metabolites, nutrient digestibility and growth performance in sheep fed alfalfa-based diet. Sci Res Essays. (2013) 8:853−7. Available online at: http://www.academicjournals.org/article/article1381490720_Kazemi-Bonchenari%20et%20al.pdf
466. Hassanzadeh P, Nouri Gharajalar S, Mohammadzadeh S. Antimicrobial effects of different synbiotic compounds against pathogenic bacteria isolated from beef, mutton, and chicken. Arch Razi Inst. (2022) 77:2105–13. doi: 10.22092/ARI.2022.357834.2107
467. Elliethy MA, Fattah A, Marwan AA. Influence of prebiotic, probiotic and synbiotic supplementation on digestibility, haemobiochemical profile and productive performance in barki lambs. Egypt J Nutr Feeds. (2022) 25:199–210. doi: 10.21608/ejnf.2022.256903
468. Munezero O, Cho S, Kim IH. The effects of synbiotics-glyconutrients on growth performance, nutrient digestibility, gas emission, meat quality, and fatty acid profile of finishing pigs. J Anim Sci Technol. (2024) 66:310–25. doi: 10.5187/jast.2023.e52
469. Bugenyi AW, Song K-D, Lee H-K, Heo J. Bacillus-and Lactobacillus-based dietary synbiotics are associated with shifts in the oropharyngeal, proximal colonic, and vaginal microbiomes of korean native black pigs. Fermentation. (2023) 9:359. doi: 10.3390/fermentation9040359
470. Zhu Q, Azad MAK, Dong H, Li C, Li R, Cheng Y, et al. Sow-offspring diets supplemented with probiotics and synbiotics are associated with offspring's growth performance and meat quality. Int J Mol Sci. (2023) 24:7668. doi: 10.3390/ijms24087668
471. Ghasemi HA, Kasani N, Taherpour K. Effects of black cumin seed (Nigella sativa L.). A probiotic, a prebiotic and a synbiotic on growth performance, immune response and blood characteristics of male broilers. Livest Sci. (2014) 164:128–34. doi: 10.1016/j.livsci.2014.03.014
472. Dibaji SM, Seidavi A, Asadpour L, da Silva FM. Effect of a synbiotic on the intestinal microflora of chickens. J Appl Poult Res. (2014) 23:1–6. doi: 10.3382/japr.2012-00709
473. Ahmed KS, Hasan M, Asaduzzaman M, Khatun A, Islam K. Effects of probiotics and synbiotics on growth performance and haemato-biochemical parameters in broiler chickens. J Sci. (2015) 10:926–9.
474. Nekoubin H, Rakhshanipour G, Hatefi S, Sudagar M, Montajami S. Effects of feeding frequency on growth performance and survival rate of Zebra fish (Danio rerio). Adv J Agric Res. (2013) 1:7–10.
475. Ai Q, Xu H, Mai K, Xu W, Wang J, Zhang W. Effects of dietary supplementation of Bacillus subtilis and fructooligosaccharide on growth performance, survival, non-specific immune response and disease resistance of juvenile large yellow croaker, Larimichthys crocea. Aquaculture. (2011) 317:155–61. doi: 10.1016/j.aquaculture.2011.04.036
476. Mehrabi Z, Firouzbakhsh F, Jafarpour A. Effects of dietary supplementation of synbiotic on growth performance, serum biochemical parameters and carcass composition in rainbow trout (Oncorhynchus mykiss) fingerlings. J Anim Physiol Anim Nutr. (2012) 96:474–81. doi: 10.1111/j.1439-0396.2011.01167.x
477. Guo J, Zhang Z, Guan LL, Yoon I, Plaizier JC, Khafipour E. Postbiotics from Saccharomyces cerevisiae fermentation stabilize microbiota in rumen liquid digesta during grain-based subacute ruminal acidosis (SARA) in lactating dairy cows. J Anim Sci Biotechnol. (2024) 15:101. doi: 10.3389/fmicb.2024.1409659
478. Dai D, Kong F, Han H, Shi W, Song H, Yoon I, et al. Effects of postbiotic products from Saccharomyces cerevisiae fermentation on lactation performance, antioxidant capacity, and blood immunity in transition dairy cows. J Dairy Sci. (2024) 107:10584–98. doi: 10.3168/jds.2023-24435
479. Maina TW, McDonald PO, Rani Samuel BE, Sardi MI, Yoon I, Rogers A, et al. Feeding Saccharomyces cerevisiae fermentation postbiotic products alters immune function and the lung transcriptome of preweaning calves with an experimental viral-bacterial coinfection. J Dairy Sci. (2024) 107:2253–67. doi: 10.3168/jds.2023-23866
480. Kim H-J, Youn H-Y, Moon J-S, Kim H, Seo K-H. Comparative anti-microbial and anti-biofilm activities of postbiotics derived from kefir and normal raw milk lactic acid bacteria against bovine mastitis pathogens. LWT-Food Sci Technol. (2024) 191:115699. doi: 10.1016/j.lwt.2023.115699
481. Henige M, Anklam K, Aviles M, Buettner J, Henschel S, Yoon I, et al. (2024). The effect of Saccharomyces cerevisiae fermentation product supplementation on pro-inflammatory cytokines in holstein friesian cattle experimentally inoculated with digital dermatitis Animals, 14. (3260). doi: 10.3390/ani14223260
482. Lauková A, Madar M, Zábolyová N, Troscianczyk A, Pogány Simonová M. Fortification of goat milk yogurts with encapsulated postbiotic active Lactococci. Life. (2024) 14:1147. doi: 10.3390/life14091147
483. Fernández C, Romero T, Badiola I, Díaz-Cano J, Sanzol G, Loor JJ. Postbiotic yeast fermentation product supplementation to lactating goats increases the efficiency of milk production by enhancing fiber digestibility and ruminal propionate, and reduces energy losses in methane. J Anim Sci. (2022) 101:skac370. doi: 10.1093/jas/skac370
484. Christodoulou C, Skourtis A, Kyriakaki P, Satolias FF, Karabinas D, Briche M, et al. The effect of dietary supplementation with probiotic and postbiotic yeast products on ewes milk performance and immune oxidative status. J Fungi. (2023) 9:1139. doi: 10.3390/jof9121139
485. Abbasi A, Sabahi S, Bazzaz S, Tajani AG, Lahouty M, Aslani R, et al. An edible coating utilizing Malva sylvestris seed polysaccharide mucilage and postbiotic from Saccharomyces cerevisiae var. boulardii for the preservation of lamb meat. Int J Biol Macromol. (2023) 246:125660. doi: 10.1016/j.ijbiomac.2023.125660
486. Ozma MA, Abbasi A, Sabahi S. Characterization of postbiotics derived from Lactobacillus paracasei ATCC 55544 and its application in Malva sylvestris seed mucilage edible coating to the improvement of the microbiological, and sensory properties of lamb meat during storage. Biointerface Res Appl Chem. (2022) 13:267–1. doi: 10.33263/BRIAC133.267
487. Ganda E, Chakrabarti A, Sardi MI, Tench M, Kozlowicz BK, Norton SA, et al. Saccharomyces cerevisiae fermentation product improves robustness of equine gut microbiome upon stress. Front Vet Sci. (2023) 10:1134092. doi: 10.3389/fvets.2023.1134092
488. Norton SA, Adams AA. PSXV-17 Dietary postbiotic mediates cytokine gene expression in senior horses. J Anim Sci. (2021) 99(Supplement_3):360–61. doi: 10.1093/jas/skab235.660
489. Yan H, Xing Q, Xiao X, Yu B, He J, Mao X, et al. Effect of Saccharomyces cerevisiae postbiotics and essential oil on growth performance and intestinal health of weanling pigs during K88 ETEC infection. J Anim Sci. (2024) 102:skae007. doi: 10.1093/jas/skae007
490. Kim SW, Duarte ME. Saccharomyces yeast postbiotics supplemented in feeds for sows and growing pigs for its impact on growth performance of offspring and growing pigs in commercial farm environments. Anim Biosci. (2024) 37:1463–73. doi: 10.5713/ab.23.0467
491. Duarte ME, Deng Z, Kim SW. Effects of dietary Lactobacillus postbiotics and bacitracin on the modulation of mucosa-associated microbiota and pattern recognition receptors affecting immunocompetence of jejunal mucosa in pigs challenged with enterotoxigenic F18+ Escherichia coli. J Anim Sci Biotechnol. (2024) 15:139. doi: 10.1186/s40104-024-01098-1
492. Duarte ME, Kim SW. Efficacy of Saccharomyces yeast postbiotics on cell turnover, immune responses, and oxidative stress in the jejunal mucosa of young pigs. Sci Rep. (2024) 14:19235. doi: 10.1038/s41598-024-70399-2
493. Xu X, Duarte ME, Kim SW. Postbiotic effects of Lactobacillus fermentate on intestinal health, mucosa-associated microbiota, and growth efficiency of nursery pigs challenged with F18+Escherichia coli. J Anim Sci. (2022) 100:skac210. doi: 10.1093/jas/skac210
494. Quintanilla-Pineda M, Ibañez FC, Garrote-Achou C, Marzo F. A novel postbiotic product based on weissella cibaria for enhancing disease resistance in rainbow trout: aquaculture application. Animals. (2024) 14:744. doi: 10.3390/ani14050744
495. Wang J, Li S, Jian Y, Song J, Zheng J, Zhou D, et al. Dietary postbiotics supplementation improves growth, survival rate, antioxidant capacity, non-specific immunity and gut health of juvenile oriental river prawn (Macrobrachium nipponense). Aquacult Rep. (2023) 33:101771. doi: 10.1016/j.aqrep.2023.101771
496. Meng D, Hao Q, Zhang Q, Yu Z, Liu S, Yang Y, et al. A compound of paraprobiotic and postbiotic derived from autochthonous microorganisms improved growth performance, epidermal mucus, liver and gut health and gut microbiota of common carp (Cyprinus carpio). Aquaculture. (2023) 570:739378. doi: 10.1016/j.aquaculture.2023.739378
497. Monika M, Tyagi JS, Sonale N, Biswas A, Murali D, Sky, et al. Evaluating the efficacy of Lactobacillus acidophilus derived postbiotics on growth metrics, health, and gut integrity in broiler chickens. Sci Rep. (2024) 14:24768. doi: 10.1038/s41598-024-74078-0
498. Chang HM, Loh TC, Foo HL, Lim ETC. Lactiplantibacillus plantarum postbiotics: alternative of antibiotic growth promoter to ameliorate gut health in broiler chickens. Front Vet Sci. (2022) 9:883324. doi: 10.3389/fvets.2022.883324
499. Farran M, El Masry B, Kaouk Z, Shaib H. Impact of dietary Lactobacillus plantarum postbiotics on the performance of layer hens under heat stress conditions. Open J Vet Med. (2024) 14:39–55. doi: 10.4236/ojvm.2024.143004
500. Zhang ZF, Kim IH. Effects of multistrain probiotics on growth performance, apparent ileal nutrient digestibility, blood characteristics, cecal microbial shedding, and excreta odor contents in broilers. Poult Sci. (2014) 93:364–70. doi: 10.3382/ps.2013-03314
Keywords: antibiotic growth promoter (AGP), antimicrobial resistance, antimicrobial alternative, AGP-free livestock production, One Health approach
Citation: Ibeagha-Awemu EM, Omonijo FA, Piché LC and Vincent AT (2025) Alternatives to antibiotics for sustainable livestock production in the context of the One Health approach: tackling a common foe. Front. Vet. Sci. 12:1605215. doi: 10.3389/fvets.2025.1605215
Received: 03 April 2025; Accepted: 07 July 2025;
Published: 13 August 2025.
Edited by:
Francis Sena Nuvey, Friedrich Loeffler Institute, GermanyReviewed by:
Massimo Canali, University of Bologna, ItalyGisele R. Rodrigues, Catholic University of Brasilia (UCB), Brazil
Helena Dela, University of Ghana, Ghana
Jose Maria Carrera Chavez, Universidad Autónoma de Ciudad Juárez, Mexico
Copyright © 2025 Ibeagha-Awemu, Omonijo, Piché and Vincent. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eveline M. Ibeagha-Awemu, ZXZlbGluZS5pYmVhZ2hhLWF3ZW11QGFnci5nYy5jYQ==; Antony T. Vincent, YW50b255LnZpbmNlbnRAZnNhYS51bGF2YWwuY2E=