 Qi Zhao
Qi Zhao Ruoyu Xiong
Ruoyu Xiong Jianhao Tang
Jianhao Tang- Key Laboratory of Germplasm Innovation and Genetic Improvement of Grain and Oil Crops, Co-construction by Ministry and Province, Ministry of Agriculture and Rural Affairs, Shanghai Academy of Agricultural Sciences, Shanghai, China
Introduction: Lodging is a critical limiting factor affecting stable rice production in the lower reaches of the Yangtze River in China.
Methods: This study aims to clarify the optimal spraying period and concentration of paclobutrazol for high-quality japonica rice by examining its effects on the mechanical, morphological, and metabolic characteristics of the stem. A clean water treatment (CK) was established alongside five spraying periods (S1: July 19; S2: July 26; S3: August 2; S4: August 9; S5: August 16) with three spraying concentrations (LP: 100 mg L-1; MP: 200 mg L-1; HP: 300 mg L-1).
Results and discussion: The results indicate that the MP treatment during the S4 period resulted in a yield increase of 2.3–11.8% while reducing the lodging index by 15.8-25.5%. In the early stages of spraying (S1, S2 and S3), the lodging resistance of rice was primarily enhanced by reducing plant height, but led to a significant decrease in the spikelets per panicle and grain filling, ultimately resulting in reduced yield. In the later stages of spraying (S4 and S5), lodging resistance was mainly improved by increasing internode diameter and culm wall thickness, which concurrently resulted in a significant increase in the grain filling. Although high concentrations of paclobutrazol in each period can reduce the lodging index, they are not conducive to yield formation. Correlation analysis revealed a significant negative correlation between the lodging index and both the internode diameter and the culm wall thickness. Non-target metabolism indicated that the significant increase in mandelonitrile and sarcosine content within cyanoamino acids metabolism, and glycine, serine, and threonine metabolism during S4, might be the primary metabolic factors promoting the development and thickness of stem walls. Additionally, the increase in pheophytin-a in porphyrin and chlorophyll metabolism, along with leucine in valine, leucine, and isoleucine degradation, may be the key contributors to the enhancement of rice lodging resistance and stable yield when treated with an appropriate concentration of paclobutrazol.
Highlights
● MP treatment (200 mg L-1 paclobutrazol) increased rice yield by 2.3%-11.8% and decreased lodging index by 15.8%-25.5% under S4 period.
● Paclobutrazol decreased lodging index by decreasing plant height in tillering rice stage.
● Paclobutrazol decreased lodging index by increasing culm wall thickness in jointing rice stage.
● The non-target metabolome indicated that paclobutrazol regulates the plant development by influencing its stem physiology.
1 Introduction
Approximately half of the global population relies predominantly on rice as a staple food. Consequently, enhancing rice yield remains a primary focus of research, driven by the increasing population, diminishing arable land, and various other challenges (Li et al., 2021). Previous studies have shown that rice production must increase by over 2.4% annually between 2010 and 2050 to satisfy the world’s food demands (Ray et al., 2013). In recent years, the frequent occurrence of extreme weather events worldwide has exacerbated the issue of rice lodging (Robinson et al., 2021). Rice lodging can reduce yields by 20%–50% and hinder the efficiency of mechanical harvesting (Lv et al., 2022). Therefore, improving the lodging resistance of rice is crucial to mitigate the risk of reduced grain yields. Lodging is defined as a permanent displacement of the vertical plant stem, resulting from a loss of balance within the plant (Zhang et al., 2014). The relationship between the lodging index (LI) and various characteristics including physical, morphological, anatomical, and physicochemical components has been extensively reported (Chen et al., 2021; Islam et al., 2007; Liao et al., 2023). Dwarf rice exhibits strong lodging resistance, and that appropriately shortening the basal internode can enhance the lodging resistance of rice populations (Liao et al., 2023). However, reducing the length of the upper internode may hinder ear development (Zhang et al., 2016). Significant positive correlations have been observed between stem thickness, stem wall thickness, and the ratio of stem diameter to pulp cavity diameter with the lodging resistance of rice populations (Zhang et al., 2013; Wang et al., 2006). The stem can be categorized into mechanical tissue, vascular bundles, epidermis, and basic tissue (Lv et al., 2022). The proportion of the cross-sectional area of mechanical tissue, the thickness of mechanical tissue, and the number of large and small vascular bundles are closely associated with the mechanical strength of the stems (Zhang et al., 2014). Lodging-resistant varieties tend to have a higher number of vascular bundles per unit area, whereas lodging-prone varieties typically exhibit thinner cell walls and larger pulp cavities (Perik et al., 2012). In the early growth stage, a substantial amount of photosynthetic products generated by the leaves is transported to the stem, facilitating lignification, fibrosis, and starch accumulation in the cell wall, which are positively correlated with the lodging resistance of rice (Zhang et al., 2020). However, during the vegetative growth period, carbohydrates in the stem sheath are redirected to the panicle kernel, leading to a decrease in internode weight, an increase in the height of the center of gravity, and, consequently, a heightened risk of lodging (Zhang et al., 2020; Pan et al., 2019).
Paclobutrazol, a triazole, is a commonly used plant growth regulator (Maheshwari et al., 2023). In rice, it has been reported that the application of paclobutrazol can resist salt stress and drought stress and enhance lodging resistance (Azarcon et al., 2022; Khunpon et al., 2018; Maheshwari et al., 2023). The application of paclobutrazol could improve seedling quality, increase tillering number, increase leaf chlorophyll content and net photosynthetic rate, and delay leaf senescence (Assuero et al., 2012). It also increases dry matter accumulation, promotes seed assimilation and distribution, and improves crop yield (Kamran et al., 2020). Previous studies have indicated that the application of paclobutrazol has three primary effects on various types and varieties of rice: it reduces plant height, thickens stems, and shortens internodes (Gai et al., 2023). These effects are primarily attributed to four molecular mechanisms. First, gibberellin is the principal hormone that promotes the elongation of plant cells. Paclobutrazol reduces plant height by inhibiting the biosynthesis pathway of gibberellin (Desta and Amare, 2021). Second, paclobutrazol enhances the activity of auxin oxidase, thereby decreasing the auxin content within the plant. Auxin plays a crucial role in cell elongation and division; thus, a reduction in its concentration contributes to reduced cell elongation and results in thicker stems (Opio et al., 2020). Third, paclobutrazol may increase the thickness and strength of the cell wall by regulating the expression of genes associated with cell wall synthesis (Lv et al., 2022). This enhancement of the cell wall improves the mechanical support capacity of the stem. Lastly, paclobutrazol can influence the expression of genes related to lodging, including those associated with stem strength and toughness, such as cellulose synthase genes and lignin synthesis-related genes (Lv et al., 2022). The upregulation of these genes enhances the mechanical properties of the stems.
The coastal japonica rice area in South China is the main production area to ensure food security in China. With the intensification of extreme climate change and frequent severe weather such as typhoon in recent years, the risk of rice lodging is increasing. As far as we know, in high-quality japonica rice, the optimal spray period and concentration of paclobutrazol and the mechanism of internode changes in metabolic level are still unclear. Therefore, the aims of this study are (1) to determine the best spray period and concentration of paclobutrazol in high-quality japonica rice and (2) to clarify the mechanism of changes in internode metabolism of paclobutrazol, in order to provide theoretical guidance for lodging resistance cultivation of high-quality japonica rice in coastal rice areas of South China.
2 Materials and methods
2.1 Site description
Field experiments were conducted at the Zhuanghang Comprehensive Experiment Station (121° 23′ E, 30° 53′ N) during 2023 and 2024. This area is characterized by the subtropical monsoon climate. The paddy soil is classified as clay soil (IUSS Working Group and WRB, 2006). The physicochemical properties of the topsoil measured at the 0- to 20-cm depth were pH 6.3, 18.5 g kg−1 soil organic matter, 4.7 mg kg−1 nitrate nitrogen, 21.5 mg kg−1 ammonium nitrogen, 40.3 mg kg−1 available phosphorus, and 129.3 mg kg−1 available potassium. The precipitation and temperature during this experiment are shown in the supplementary data in Supplementary Figure S1. The average daily temperatures from seedling transplanting to rice harvest in 2023 and 2024 were 27.4°C and 27.8°C, respectively, and the total rainfall was 61.6 and 828.1 mm, respectively.
2.2 Experimental design
To evaluate the effects of different paclobutrazol spraying periods and concentrations on the lodging characteristics and yield of high-quality japonica rice, a split-plot design was employed. The main plot corresponds to the paclobutrazol spraying periods: S1, sprayed on July 19, tillering initiation stage; S2, sprayed on July 26, tillering peak stage; S3, sprayed on August 2, tillering decline stage; S4, sprayed on August 9, jointing stage; and S5, sprayed on August 16, panicle stage. The sub-plot pertains to the spraying concentrations: low paclobutrazol concentration (LP) at 100 mg L−1, medium paclobutrazol concentration (MP) at 200 mg L−1, and high paclobutrazol concentration (HP) at 300 mg L−1. An equal amount of water was sprayed as a control (CK). In total, there are 16 treatments, each covering an area of 24 m² (length, 6 m; width, 4 m), with each treatment replicated three times. The planting density was 20 cm × 16 cm with two seedlings per hole. The total amount of N fertilizer was 270 kg ha−1 and was applied as base fertilizer:tillering fertilizer:panicle fertilizer = 5:3:2. Both phosphorus (P2O5) and potassium (K2O) were applied at a rate of 45 kg ha−1; phosphorus was applied as a base fertilizer once, and potassium was applied equally in base and panicle fertilizer. Other cultivation management practices were carried out in accordance with the local requirements for high yields and quality cultivation. Diseases and pests were intensively controlled. High-quality japonica rice cultivar Taian 1 was provided by Shanghai Academy of Agricultural Sciences in 2023 to 2024. Taian 1 was an inbred japonica rice variety cultivated as a single-cropping late rice in Shanghai. Its parent varieties include Ruanxiang rice (Maternal Parent) and Nanjing 46 (Paternal Parent) varieties, sown on 20 May, transplanted on 20 June, and harvested on 30 October in the 2 years.
2.3 Sampling and measurement
2.3.1 Grain yield and its components
Upon maturity, rice yield was measured from 2-m2 blocks by a moisture content of 13.5% to determine the actual yield, and 10 stump rice plants were selected for each plot to determine the spikelets per panicle and grain filling according to the number of effective panicles.
2.3.2 Morphological characteristics
Fifteen representative main stems were sampled in each plot for the measurements of the plant height, gravity center height, internode length, internode diameter, and culm wall thickness at 20 days after heading. Plant height and the length of internodes were measured by a ruler. Center of gravity height was measured as the distance from the base of the stem to the balance of the stem fulcrum (including the ear, leaf, and sheath). The first internode (I1) on the root was taken as the first internode (I1), and subsequent internodes were numbered as the second, third, fourth, and fifth internodes, denoted by I2, I3, I4, and I5, respectively. The internode diameter and culm wall thickness were measured with a digital caliper at the mid-region of the internode. The dry weight of each internode and the total dry weight were put into the oven after determining the morphological characteristics and dried to a constant weight.
2.3.3 Stem physical properties and lodging index
LI and physical properties were determined at 20 days after heading. Ten representative main stems were sampled in each plot. The breaking strength was measured at the I2, I3, and I4 with leaf sheath by using a prostrate tester (YYD-1, Zhejiang Top Instrument Co. Ltd., Hangzhou, China) according to the previous report by Lv et al. (2022). The parameters of stem physical properties were calculated as follows: (1) Bending moment of the whole plant (WP, g cm) = SL (cm) × FW (g), where SL is the length from the broken point to the panicle top and FW is the fresh weight from the broken point to the panicle top; (2) Breaking strength (M, g cm) = 1/4 × BL (kg) × L(cm), where BL is the force applied to break stem segment and L is the distance between two points; and (3) LI (%) = WP/M ×100.
2.3.4 Metabonomics
High-resolution non-targeted metabolomics analysis employs ultrahigh-performance liquid chromatography–tandem time-of-flight mass spectrometry or tandem Orbitrap mass spectrometry to detect metabolites within the sample. Baseline filtering, peak identification, integration, retention time correction, and peak alignment were conducted on the original data using the metabolomics processing software Progenesis QI (Waters Corporation, Milford, USA). This process resulted in the generation of a data matrix comprising retention time, mass–charge ratio, and peak intensity. The primary databases utilized included the Human Metabolome Database (http://www.hmdb.ca/) and the METLIN database (https://metlin.scripps.edu/), along with additional public databases and a self-built database. Subsequently, the data underwent the following preprocessing steps: (1) retention of only those variables with a non-zero value exceeding 80% in any sample group; (2) normalization of the total peak, followed by the exclusion of variables with a relative standard deviation (RSD) of QC samples ≥ 30%; and (3) transformation of the data using log10 to obtain the final data matrix for subsequent analyses. The data analysis encompassed univariate statistical analysis, multidimensional statistical analysis, differential metabolite screening, differential metabolite correlation analysis, and topological enrichment pathway analysis (Wang et al., 2024).
The analysis involves three spraying periods: 200 mg L−1 paclobutrazol in the S2 period corresponds to the number EP, 200 mg L−1 in the S4 period corresponds to the number MP, and 200 mg L−1 paclobutrazol in the S5 period corresponds to the number DP. Additionally, three paclobutrazol spray concentrations (100, 200, and 300 mg L−1) applied during the S4 period correspond to metabolites identified as LP, MP, and HP, respectively. Metabolome determinations were repeated six times for each treatment. For metabolite extraction, an 80% methanol (v/v) solution was used. Subsequently, proteins were removed by centrifugation at 12,000 × g for 15 min at 4°C. The supernatant was freeze-dried and then reconstituted with an acetonitrile–water (1:1) mixed solution for subsequent LC-MS analysis.
For non-targeted metabolomics analysis, a UHPLC-QTOF-MS system (Agilent 1290/6545, California, USA) equipped with a ZORBAX Eclipse Plus C18 chromatographic column (2.1 × 100 mm, 1.8 μm) was used. The mobile phase consisted of an aqueous solution containing 0.1% formic acid (Phase A) and acetonitrile (Phase B). A gradient elution program was applied (0–15 min, 5%–95% Phase B) with a flow rate of 0.3 mL/min. Mass spectrometry data were acquired in both positive and negative ion modes (m/z 50–1,000) at a scanning rate of 2 spectra per second. A quality control sample prepared by mixing all experimental samples was inserted every 10 injections to monitor the system stability.
The raw data were processed for peak extraction, alignment, and normalization using the eXtensible Computational Mass Spectrometry (XCMS). Metabolite annotation was accomplished by matching the accurate molecular weight (with a mass error <5 ppm) and the secondary mass spectrometry data against the HMDB and METLIN databases. Multivariate statistical analysis, including principal component analysis (PCA) and orthogonal partial least squares discriminant analysis (OPLS-DA), was performed using the SIMCA-P 14.1 (Sartorius Stedim Data Analytics AB, Uppsala, Sweden). The criteria for screening significantly different metabolites were as follows: a variable importance in projection (VIP) value >1.0 (based on the OPLS-DA model) and p < 0.05 (as determined by Student’s t-test). Pathway enrichment analysis was conducted using the MetaboAnalyst 5.0 platform (McGill University, Canada) with reference to the Kyoto Encyclopedia of Genes and Genomes (KEGG) database.
2.4 Statistical analysis
We used SPSS software (SPSS Inc, Chicago, USA) to calculate the mean values of yield, bending moment, breaking strength, LI, and morphological parameters, and determined the least significant difference (LSD) at the p < 0.05 level. Multifactor analysis of variance (ANOVA) was employed to determine the effects of spraying period and paclobutrazol concentration treatments (as independent variables) on yield, bending moment, breaking strength, LI, and morphological parameters, as well as their interaction levels, at significance levels of 0.05 and 0.01. Spearman correlation analysis was used to analyze the correlation between LI and morphological and mechanical parameters (n = 15). PCA of LI and mechanical parameters was performed using Origin software.
3 Results
3.1 Yield and its composition
The year (Y), spraying period (S), paclobutrazol concentration (P), and the interaction between S and P significantly influenced yield and its composition (Table 1). Additionally, the interaction between Y and S had significant effects on SPP, GF, and yield. However, the interactions between Y and P, as well as between Y, S, and P, were only found to significantly affect yield.
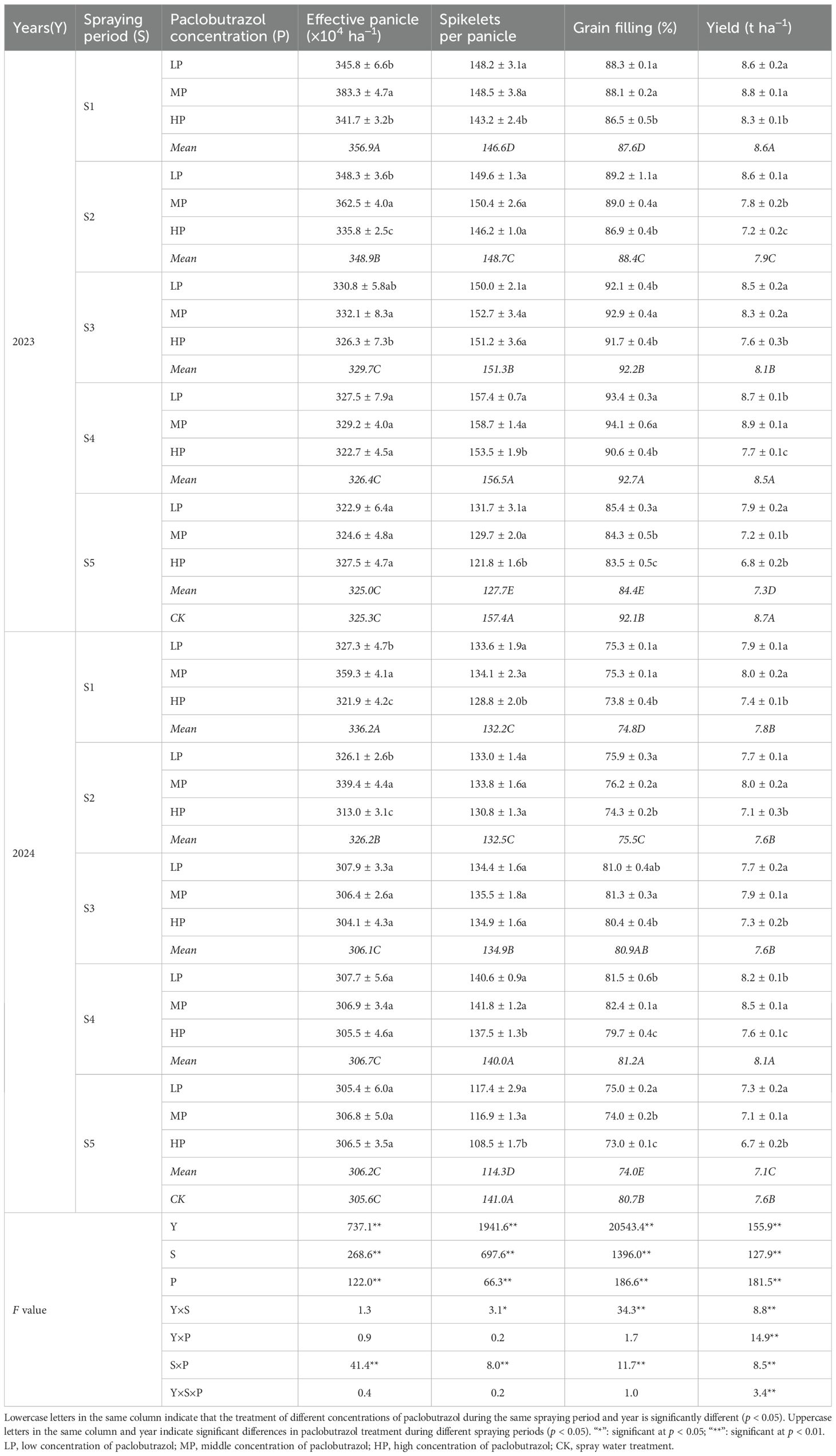
Table 1. Effects of different spraying periods and concentrations of paclobutrazol on rice yield from 2023 to 2024.
Compared with CK, there were no significant differences in yield under the S1 and S4 treatments in 2023. However, the S2, S3, and S5 periods significantly reduced yield by 9.2%, 6.9%, and 16.1%, respectively. In 2024, the S4 and S5 periods significantly altered yield, increasing or decreasing it by 6.6%, compared with CK. Compared with LP, the HP treatment significantly decreased yield by 3.5%–16.3% during the same spray period of paclobutrazol. The highest yield was observed under the MP treatment during the S4 period from 2023 to 2024. Analysis of yield components indicated that the highest yield was primarily attributed to increases in spikelets per panicle and grain filling, though the effective panicle showed no significant change compared with CK. Additionally, spray paclobutrazol significantly increased effective panicle in the S1 and S2 periods, with MP treatment demonstrating significantly higher than both LP and HP treatments. With the exception of the S4 period, spray paclobutrazol significantly reduced spikelets per panicle in the 2 years. Similarly, spray paclobutrazol significantly decreased grain filling over the 2 years, except during the S3 and S4 periods.
3.2 Lodging indices and physical parameters
S and P have significant effects on the LI, bending moment, and breaking strength at each internode (Table 2). The interaction between S and P significantly influences the breaking strength at the second and third internodes of the rice base, as well as the LI at the fourth internode of the rice base.
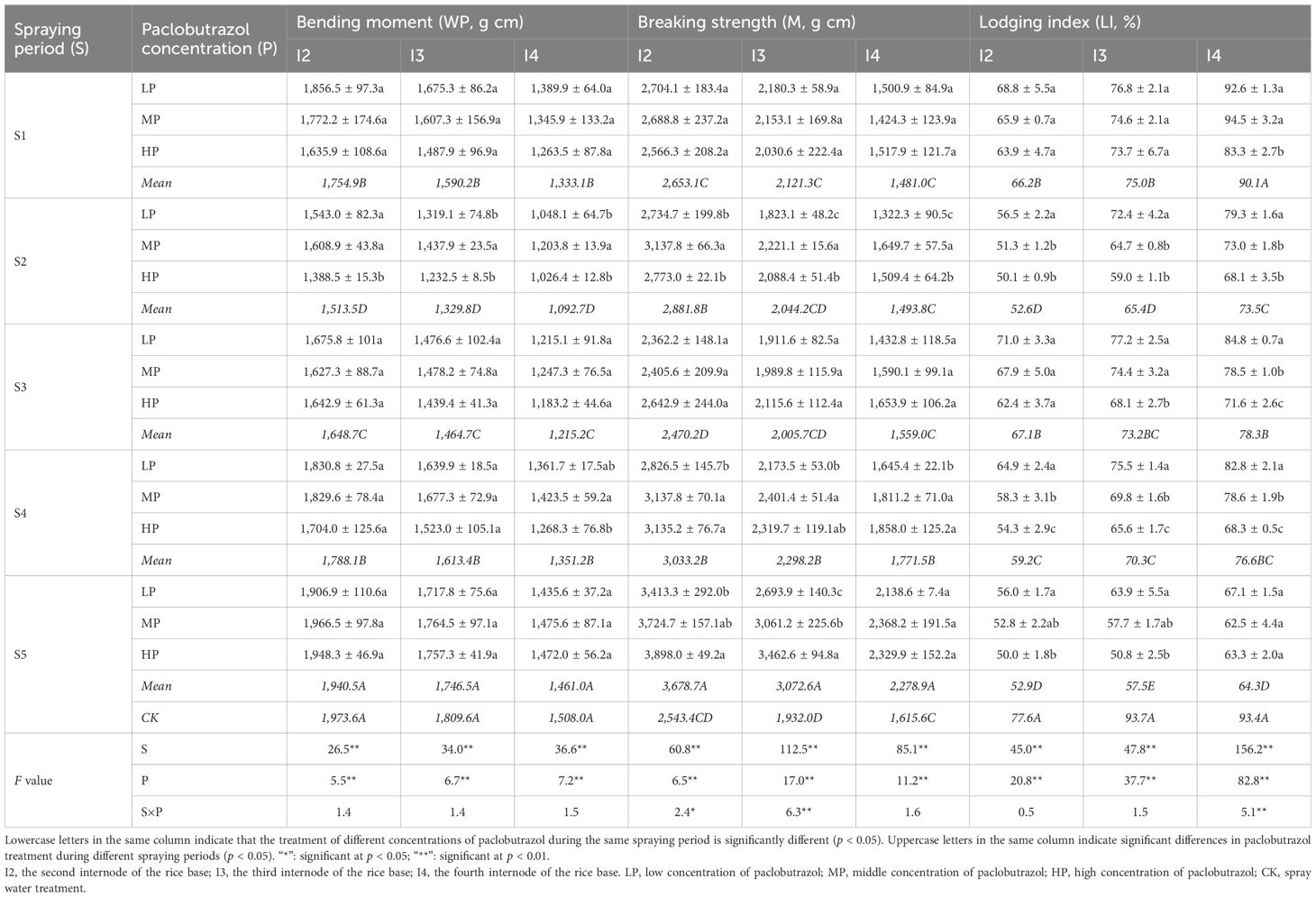
Table 2. Effects of different spraying periods and concentrations of paclobutrazol on the bending moment, breaking strength, and lodging index of rice in 2024.
Compared with CK, the LI of the second internode of the rice base decreased significantly by 14.7%, 32.2%, 13.5%, 23.7%, and 31.8% during the S1, S2, S3, S4, and S5 periods, respectively. Similarly, the LI of the third internode of the rice base showed significant reductions of 20.0%, 30.2%, 21.9%, 25.0%, and 38.6% in the S1, S2, S3, S4, and S5 periods, respectively, compared with CK. For the fourth internode of the rice base, the LI decreased significantly by 21.3%, 16.2%, 18.0%, and 31.2% in the S2, S3, S4, and S5 periods, respectively, while no significant change was observed in S1. The LI exhibited a decreasing trend as the concentration of paclobutrazol increased within the same treatment period. Notably, in the S5 period, the second and third internodes of the rice base lodging indices reached their lowest levels under HP treatment, decreasing significantly by 35.6% and 45.8%, respectively, compared with CK. Furthermore, the fourth internode of the rice base LI also reached its lowest level under MP treatment, with a significant decrease of 33.1% compared with CK.
The bending moment at each internode significantly decreased during the S1, S2, S3, and S4 periods, while showing no significant change during the S5 period, compared with CK. The bending moment of each internode initially decreased and then increased with the backward spray period of the paclobutrazol, reaching its lowest point during the S2 period. In the S2 period, the second, third, and fourth internodes of the rice base bending moment experienced significant reductions of 23.3%, 26.5%, and 27.5% compared to CK, respectively. Additionally, during the S2 period, the bending moment between internodes under MP treatment was the highest. Similarly, during the S4 period, the bending moment of the fourth internode of the rice base under MP treatment also reached its peak. Compared with CK, the breaking strength of the second internode of the rice base increased significantly during the S2, S4, and S5 periods by 13.3%, 19.3%, and 44.6%, respectively. The breaking strength of the third internode of the rice base also showed significant increases of 9.8%, 19.0%, and 59.0% in the S1, S4, and S5 periods, respectively, compared with CK. Conversely, the breaking strength of the fourth internode of the rice base significantly increased by 9.6% and 41.1% in the S4 and S5 periods, respectively, compared with CK. In the S2 period, the MP treatment significantly increased the bending strength between each node compared with both LP and HP treatments. During the S4 period, the bending strength of each node under MP treatment was significantly higher than that under LP treatment, while no significant difference was observed between HP and MP treatments. In the S5 period, the bending strength was highest under HP treatment in the second and third internodes of the rice base. Both PCA and correlation analysis showed a significant negative correlation between LI and breaking strength (Figure 1).
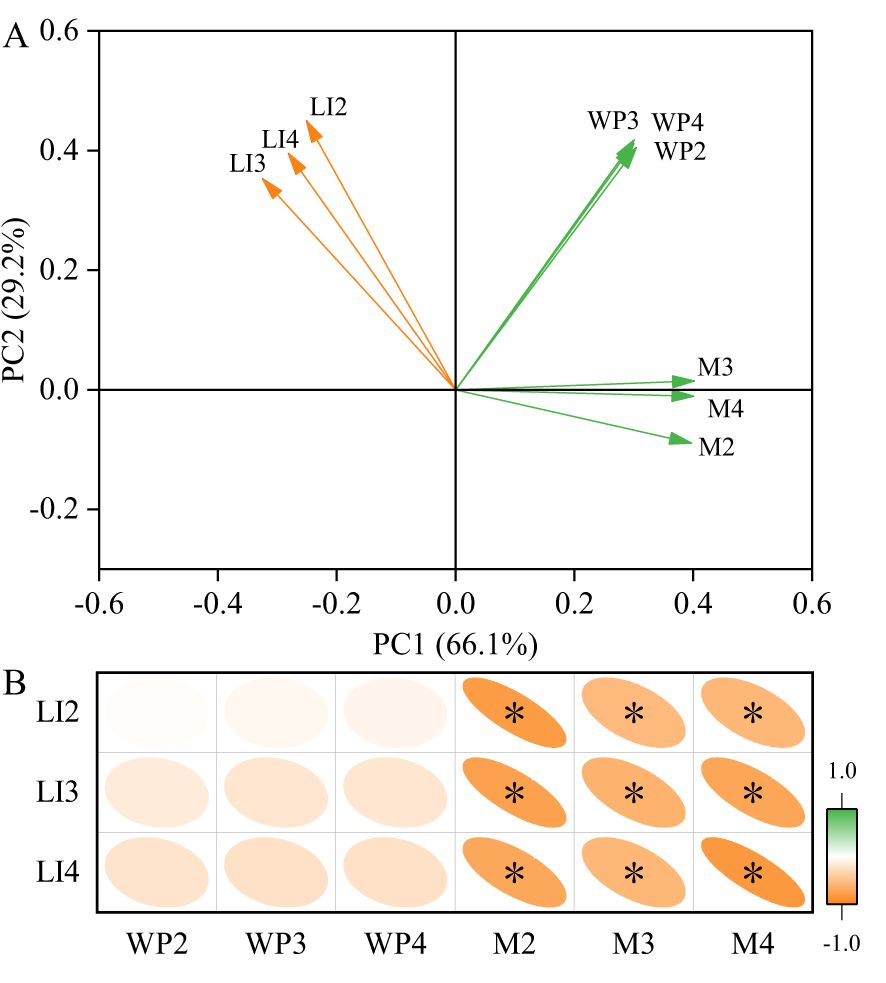
Figure 1. Principal component analysis (A) and Spearman correlation (B) between internode lodging index with bending moment and breaking strength (n = 15). “*”: significant at p < 0.05.
3.3 Plant height, gravity center height, and internode length
S had significant effects on plant height, gravity center height, and the fourth internode of rice base length (Figure 2; Table 3). P significantly affects plant height and the third and fourth internodes of rice base length. Additionally, the interaction between S and P significantly affected plant height and the fourth internode of length.
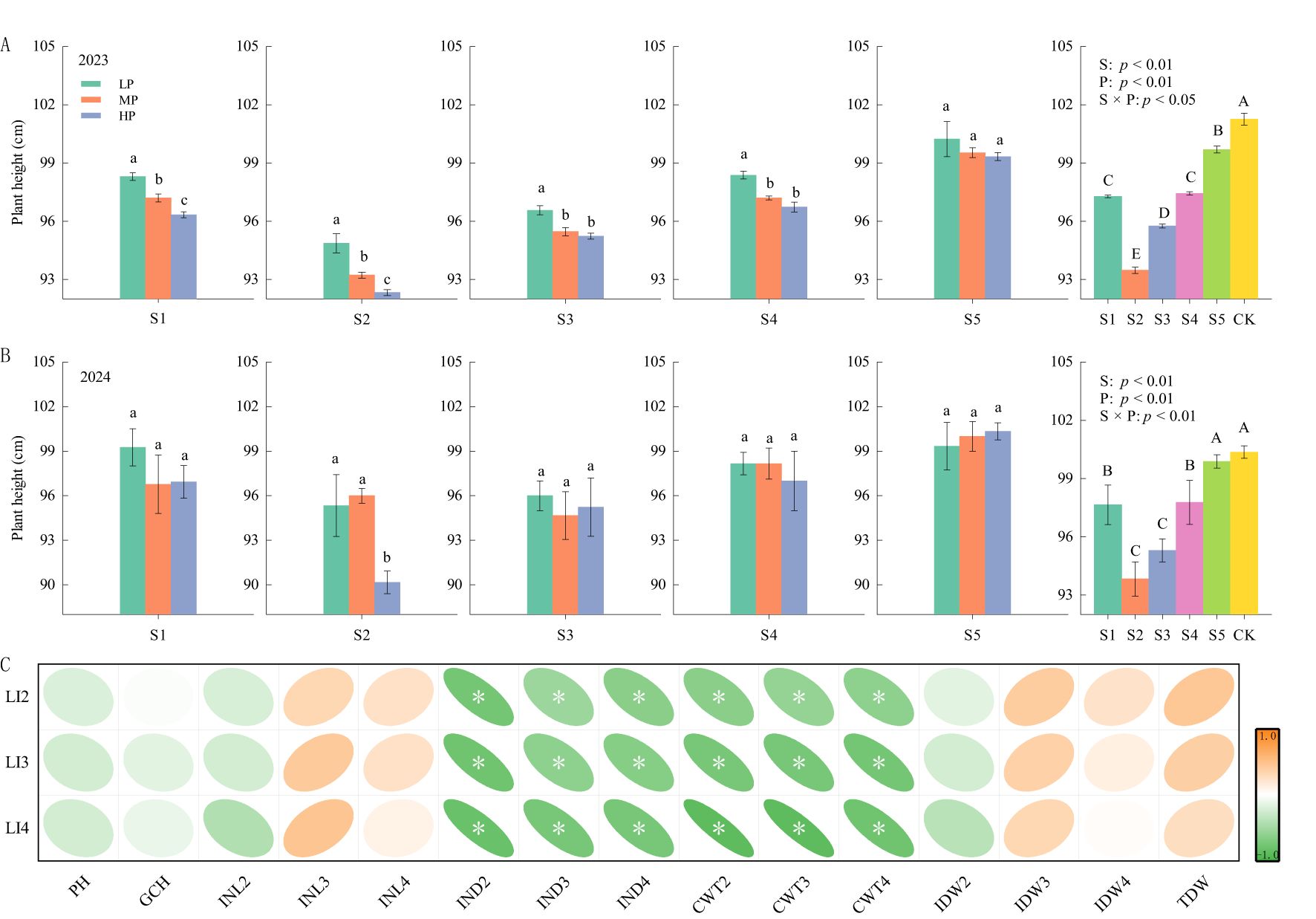
Figure 2. The effects of different spraying periods and concentrations of paclobutrazol on rice plant height in 2023 (A) and 2024 (B), and the Spearman correlation between internode lodging index and rice lodging parameters (C), n = 15. “*”: significant at p < 0.05. Lowercase letters indicate that the treatment of different concentrations of paclobutrazol during the same spraying period and year is significantly different (p<0.05). Capital letters indicate that the treatment of different spraying period of paclobutrazol during the same year is significantly different (p<0.05).
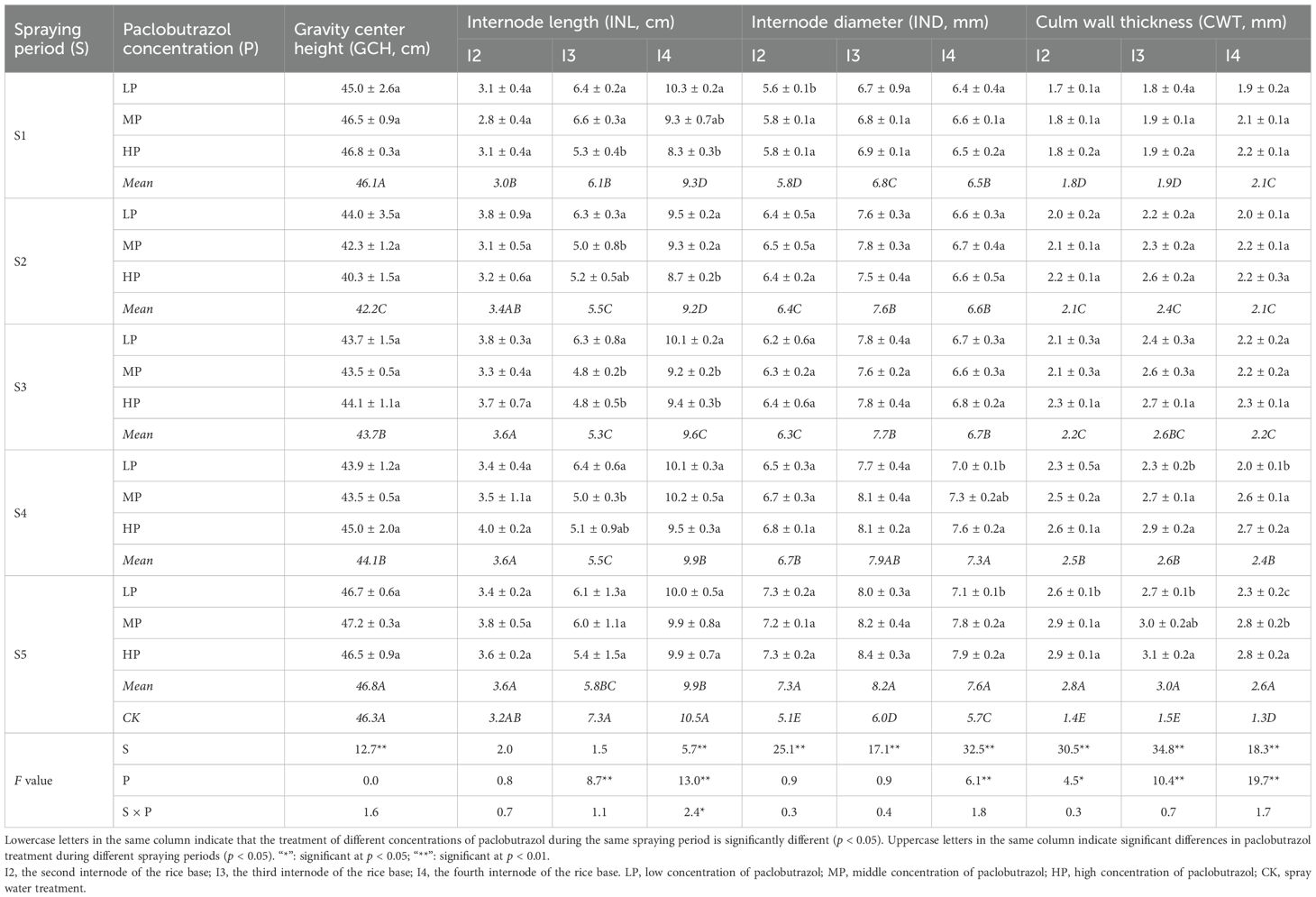
Table 3. Effects of different spraying periods and concentrations of paclobutrazol on the gravity center height, internode length, internode diameter, and culm wall thickness of rice in 2024.
Compared with CK, S1, S2, S3, and S4 resulted in significant reductions in plant height by 2.7%–3.9%, 6.5%–7.7%, 5.0%–5.4%, and 2.6%–3.8%, respectively, over 2 years. Additionally, in 2023, S5 exhibited a significant decrease in plant height by 1.5% (Figures 2A,B). In the same paclobutrazol spray period, a decrease in plant height was observed with increasing concentrations of paclobutrazol, with the lowest plant height recorded under HP treatment during the S2 period. The gravity center height initially decreased before subsequently increasing with the delaying paclobutrazol spray period (Table 3). Compared with CK, the S2, S3, and S4 periods exhibited significant decreases of 8.9%, 5.6%, and 4.8%, respectively. There was no significant effect on the gravity center height at different concentrations of paclobutrazol during the same spray period.
There was no significant difference in the second internode of rice base length, compared to the CK, across different paclobutrazol spraying periods, nor was there a significant difference between the various spraying concentrations within the same period (Table 3). The third internode of rice base length was significantly reduced by 16.4%, 24.7%, 27.4%, 24.7%, and 20.5% during the S1, S2, S3, S4, and S5 periods, respectively, compared to CK. Similarly, the fourth internode of rice base length was significantly reduced by 11.4%, 12.4%, 8.6%, 5.7%, and 5.7% during the S1, S2, S3, S4, and S5 periods, respectively. Additionally, during the S1 period, the lengths of both the third and fourth internodes of the rice base were significantly reduced under HP treatment. In the S2 period, the length of the third and fourth internodes of the rice base was at its lowest under MP and HP treatments, respectively. In the S3 period, both MP and HP treatments significantly reduced the length in the third and fourth internodes of the rice base compared to LP treatment. In the S4 period, only the fourth internode of rice base length decreased significantly under MP treatment.
3.4 Internode diameter and culm wall thickness
S had significant effects on the each internode of rice base diameter and culm wall thickness (Table 3). P significantly affected the fourth internode of rice base diameter and each internode of rice base culm wall thickness.
Paclobutrazol spraying resulted in a significant increase in internode diameter and culm wall thickness. Each internode diameter and culm wall thickness increased significantly with the delay of paclobutrazol spray periods. Compared with CK, each internode diameter increased significantly by 13.3%–14.0%, 15.8%–26.7%, 17.5%–28.3%, 28.1%–31.7%, and 33.3%–43.1% during the S1, S2, S3, S4, and S5 periods, respectively. Furthermore, the second internode of rice base diameter during the S2 period, as well as the fourth internode of the rice base during the S4 and S5 periods, increased significantly with higher concentrations of paclobutrazol. Compared with CK, each internode culm wall thickness increased significantly by 26.7%–61.5%, 38.1%–60.0%, 57.1%–73.3%, 73.3%–84.6%, and 93.3%–108.1% during the S1, S2, S3, S4, and S5 periods, respectively. The culm wall thickness of the third and fourth internodes of the rice base in the S4 period and the culm wall thickness of each internode in S5 period increased significantly with the increase of the concentration of polyphobulozole.
3.5 Dry weight of individual plants
S and P had significant effects on internode dry weight and total dry weight (Table 4). The interaction between S and P had a significant influence on the second and third internodes of the rice base.
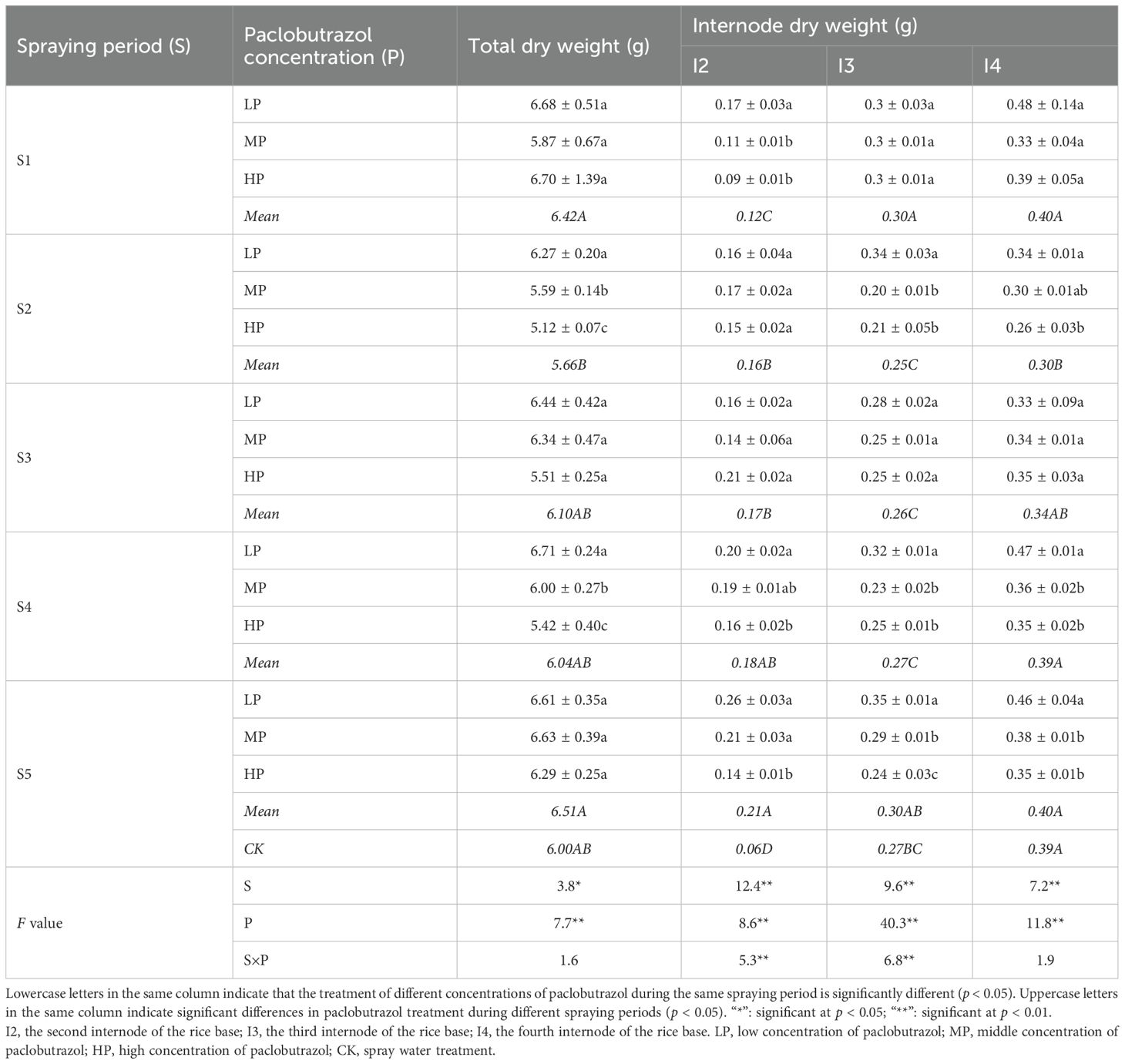
Table 4. Effects of different spraying periods and concentrations of paclobutrazol on the total and internode dry weight of rice in 2024.
Compared with CK, there was no significant effect on total dry weight during each spray period of polyphobulozole; however, a decreasing trend in total dry weight was observed during the S2 period. In both the S2 and S4 periods, total dry weight decreased significantly with the increase in spraying concentration. Notably, the dry weight of the rice base second internode exhibited a significant increase in each spraying period, compared with CK. Furthermore, with the delay of the spraying period, the dry weight of the rice base second internode continued to increase significantly. In contrast, the dry weight of the rice base third internode showed a significant increase only during the S1 period, while the dry weight of the rice base fourth internode showed a significant decrease in the S2 period. During the S1 period, the dry weight of the rice base second internode significantly decreased with increasing spraying concentration. In the S2 period, there was a notable decreasing trend in the dry weight of the rice base third and fourth internodes as the spraying concentration increased. Additionally, in the S4 and S5 periods, each internode dry weight significantly decreased with higher spraying concentrations.
3.6 The difference of non-target metabolome in different spray periods of paclobutrazol
Compared with CK, positive ions significantly upregulated 194, 97, and 88 metabolites, while downregulating 141, 195, and 184 metabolites under EP, MP, and DP treatments, respectively (Figure 3A). The negative ions significantly upregulated 115, 95, and 90 metabolites and downregulated 178, 176, and 157 metabolites under EP, MP, and DP treatments, respectively, compared with CK (Figure 3C). Additionally, there were 147 and 170 identical metabolites observed in the groups of positive and negative ions, respectively (Figures 3B, D). The MetPA analysis shows that the top five enrichment pathways of positive ions are glycerophospholipid metabolism, caffeine metabolism, riboflavin metabolism, arachidonic acid metabolism, and cyanoamino acid metabolism under different spray period treatments, compared with CK, respectively (Figures 4A–C). The top five enrichment pathways of negative ions are monobactam biosynthesis; lysine biosynthesis; glycine, serine, and threonine metabolism; caffeine metabolism; and C5-branched dibasic acid metabolism under different spray period treatments, compared with CK, respectively (Figures 4D–F).
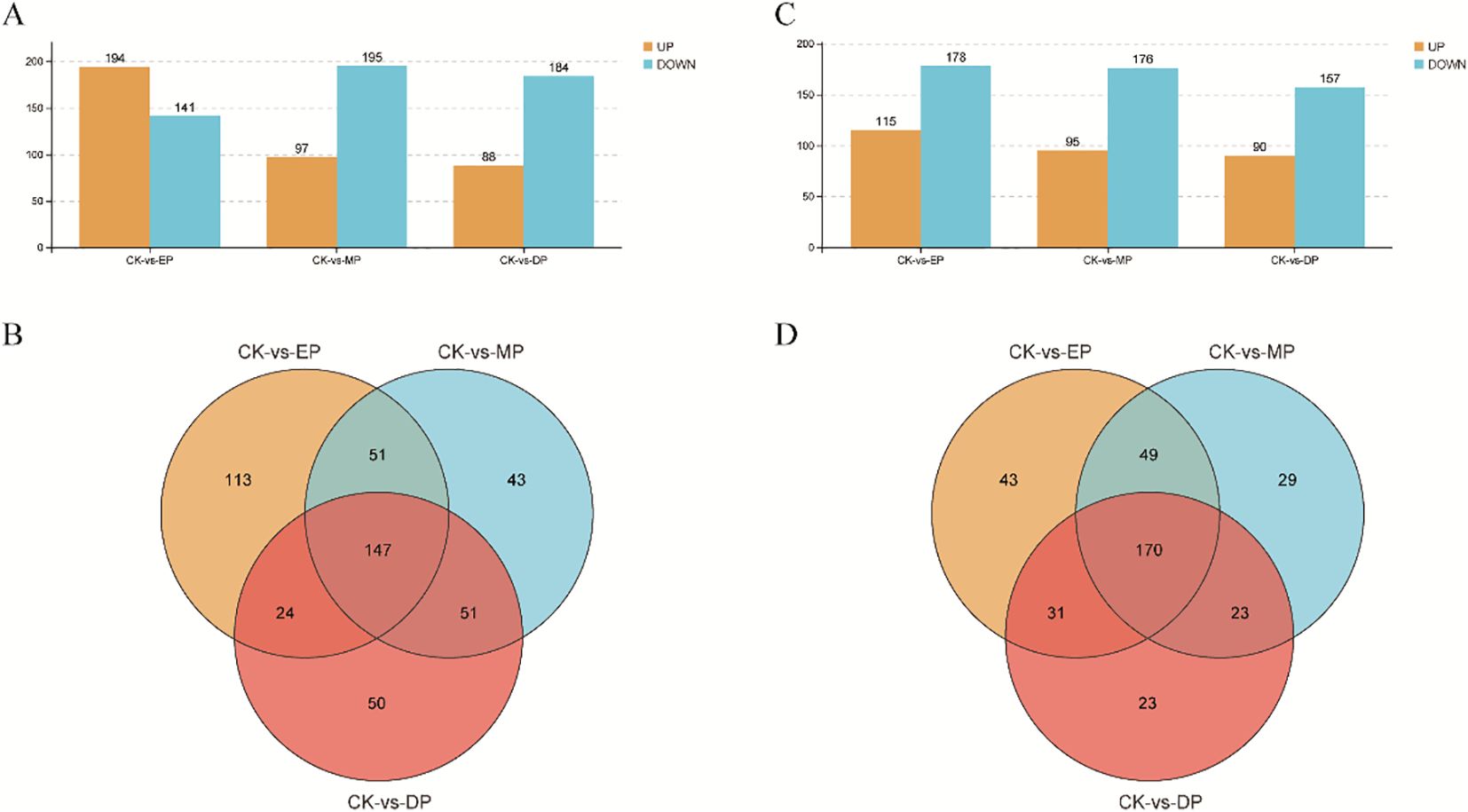
Figure 3. The metabolites of positive ions (A, B) and negative ions (C, D) were differentially expressed in the second internode of the rice base under different spraying periods of paclobutrazol. CK, spray water treatment; EP, 200 mg/L of paclobutrazol was sprayed on S1 stage (July 19); MP, 200 mg/L of paclobutrazol was sprayed on S3 stage (August 2); DP, 200 mg/L of paclobutrazol was sprayed on S5 stage (August 16).
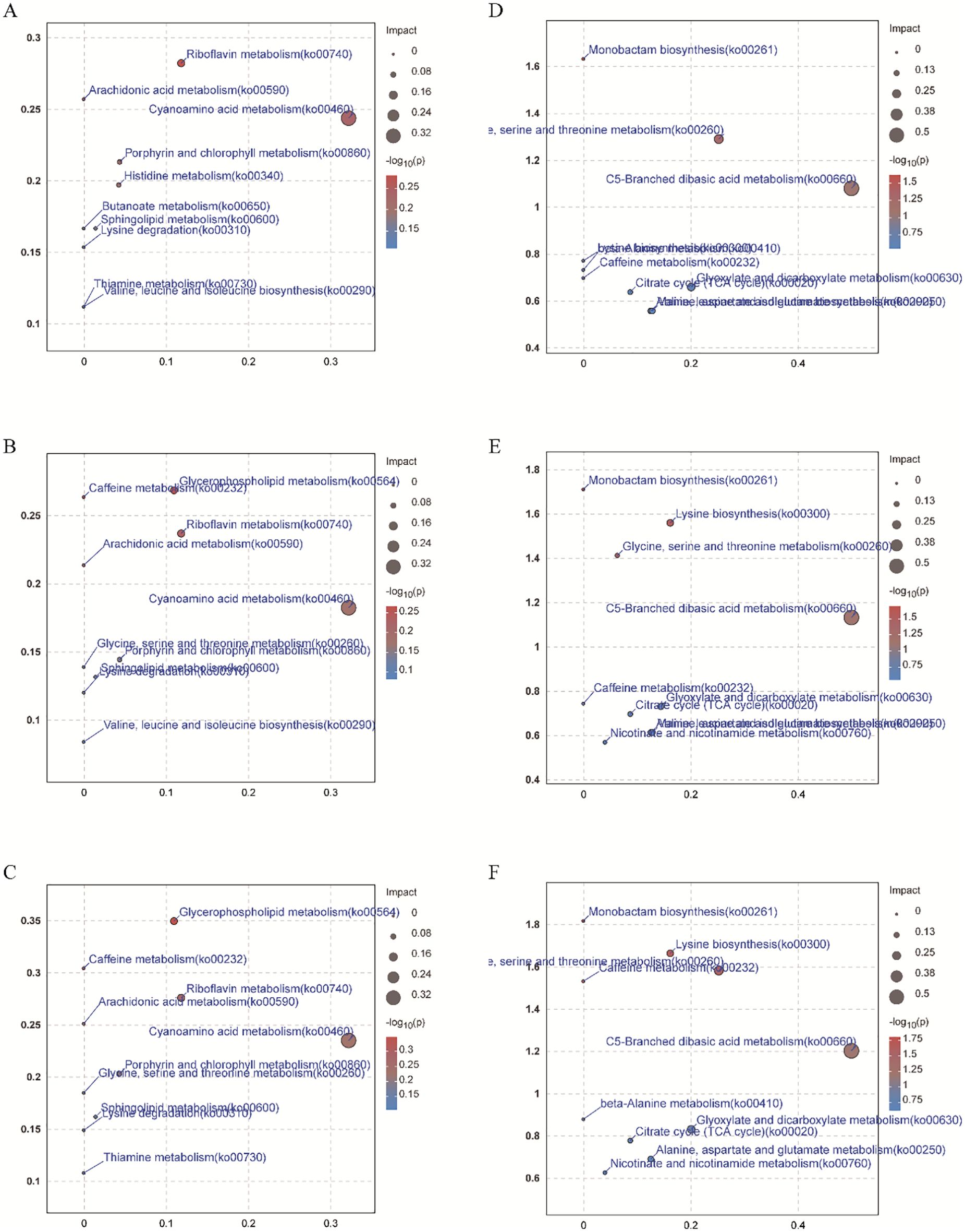
Figure 4. The MetPA analysis of positive ion (A, B, C) and negative ion (D, E, F) metabolites of paclobutrazol was different under EP (A, D), MP (B, E), and DP (C, F) treatments, respectively, compared with CK.
Further analysis of metabolites in the METPA-enriched metabolic pathway showed that glycerophosphocholine, diaminopimelic acid, and 1-methyluric acid decreased significantly with the delay of the paclobutrazol spray period (Figures 5, 6). However, (−)-riboflavin, S-methyl-5′-thioadenosine, prunasin, and phosphoric acid were significantly increased with the delay of the paclobutrazol spray period. Choline, phosphorylcholine, paraxanthine, and 1,7-dimethyluric acid reached their lowest levels under MP treatment, whereas mandelonitrile, lysine, and sarcosine reached their highest levels under MP treatment.
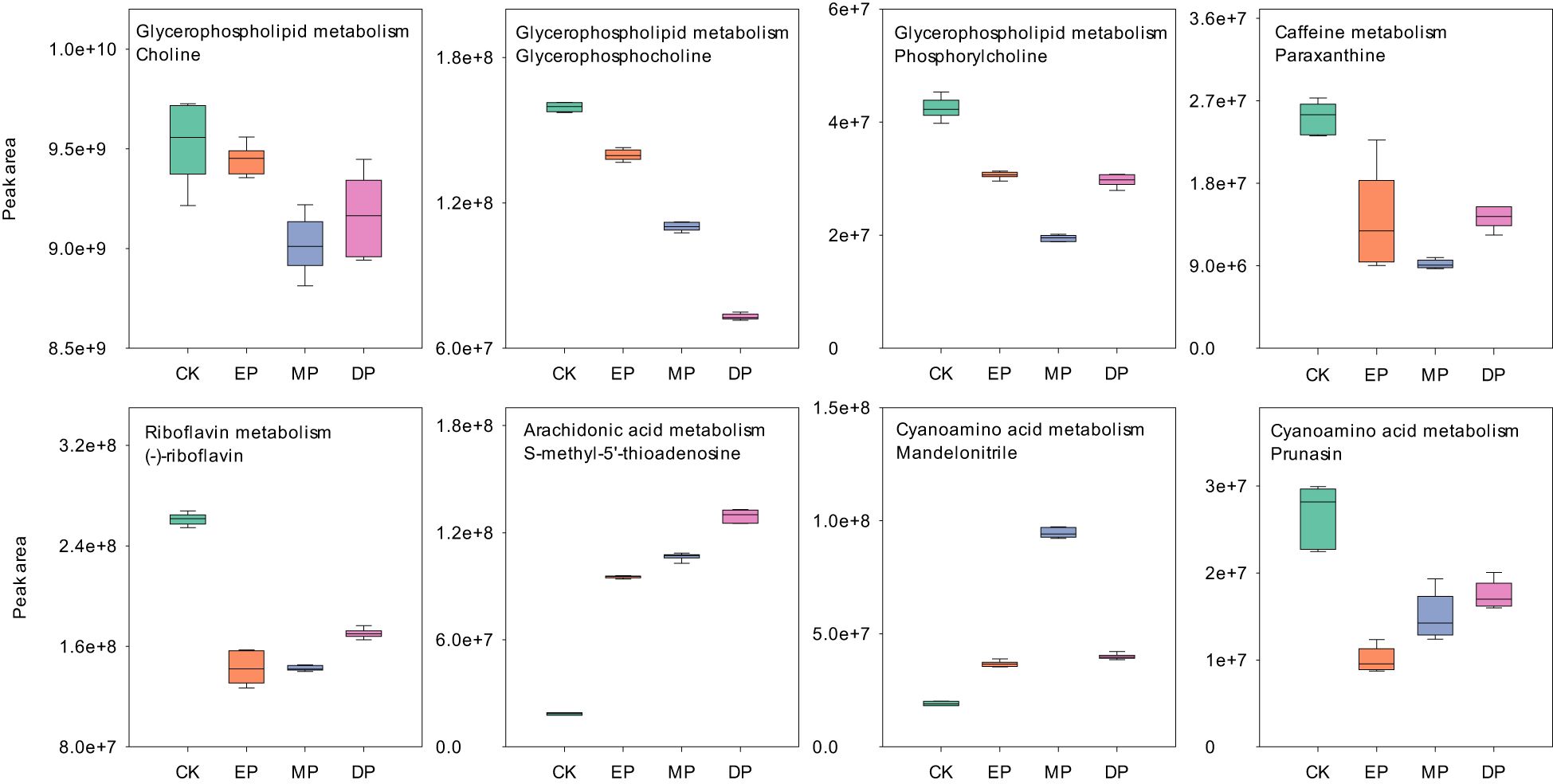
Figure 5. The MetPA analysis of positive ion differentially expressed metabolites under different spraying periods of paclobutrazol. CK, spray water treatment; EP, 200 mg/L of paclobutrazol was sprayed on S1 stage (July 19); MP, 200 mg/L of paclobutrazol was sprayed on S3 stage (August 2); DP, 200 mg/L of paclobutrazol was sprayed on S5 stage (August 16).
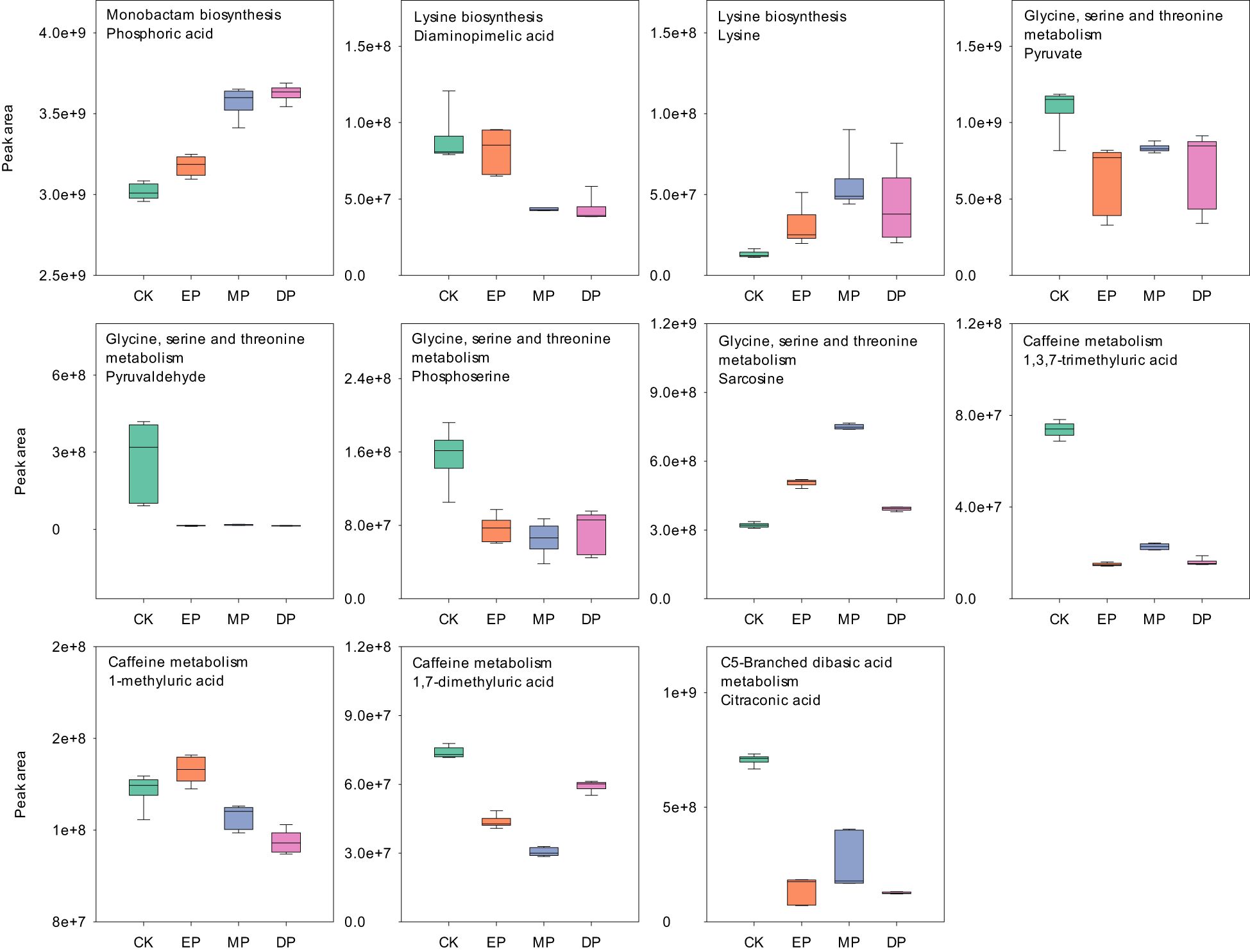
Figure 6. The MetPA analysis of negative ion differentially expressed metabolites under different spraying periods of paclobutrazol. CK, spray water treatment; EP, 200 mg/L of paclobutrazol was sprayed on S1 stage (July 19); MP, 200 mg/L of paclobutrazol was sprayed on S3 stage (August 2); DP, 200 mg/L of paclobutrazol was sprayed on S5 stage (August 16).
3.7 The difference of non-target metabolome in different concentrations of paclobutrazol
The positive ions significantly upregulated 139, 97, and 160 metabolites, while downregulating 151, 195, and 120 metabolites under LP, MP, and HP treatments, respectively, compared with CK (Figure 7A). The negative ions significantly upregulated 98, 95, and 124 metabolites and downregulated 167, 176, and 114 metabolites under LP, MP, and HP treatments, respectively, compared with CK (Figure 7C). There were 168 and 143 identical metabolites observed in the groups of positive and negative ions, respectively (Figures 7B,D). The MetPA analysis shows that the top five enrichment pathways of positive ions are biosynthesis of secondary metabolites–other antibiotics; riboflavin metabolism; porphyrin and chlorophyll metabolism; sphingolipid metabolism; and valine, leucine, and isoleucine degradation under different concentrations of paclobutrazol, compared with CK, respectively (Figures 8A–C). The top five enrichment pathways of negative ions are glyoxylate and dicarboxylate metabolism; glycine, serine, and threonine metabolism; caffeine metabolism; citrate cycle; and valine, leucine, and isoleucine biosynthesis under different concentration treatments, compared with CK, respectively (Figures 8D–F).
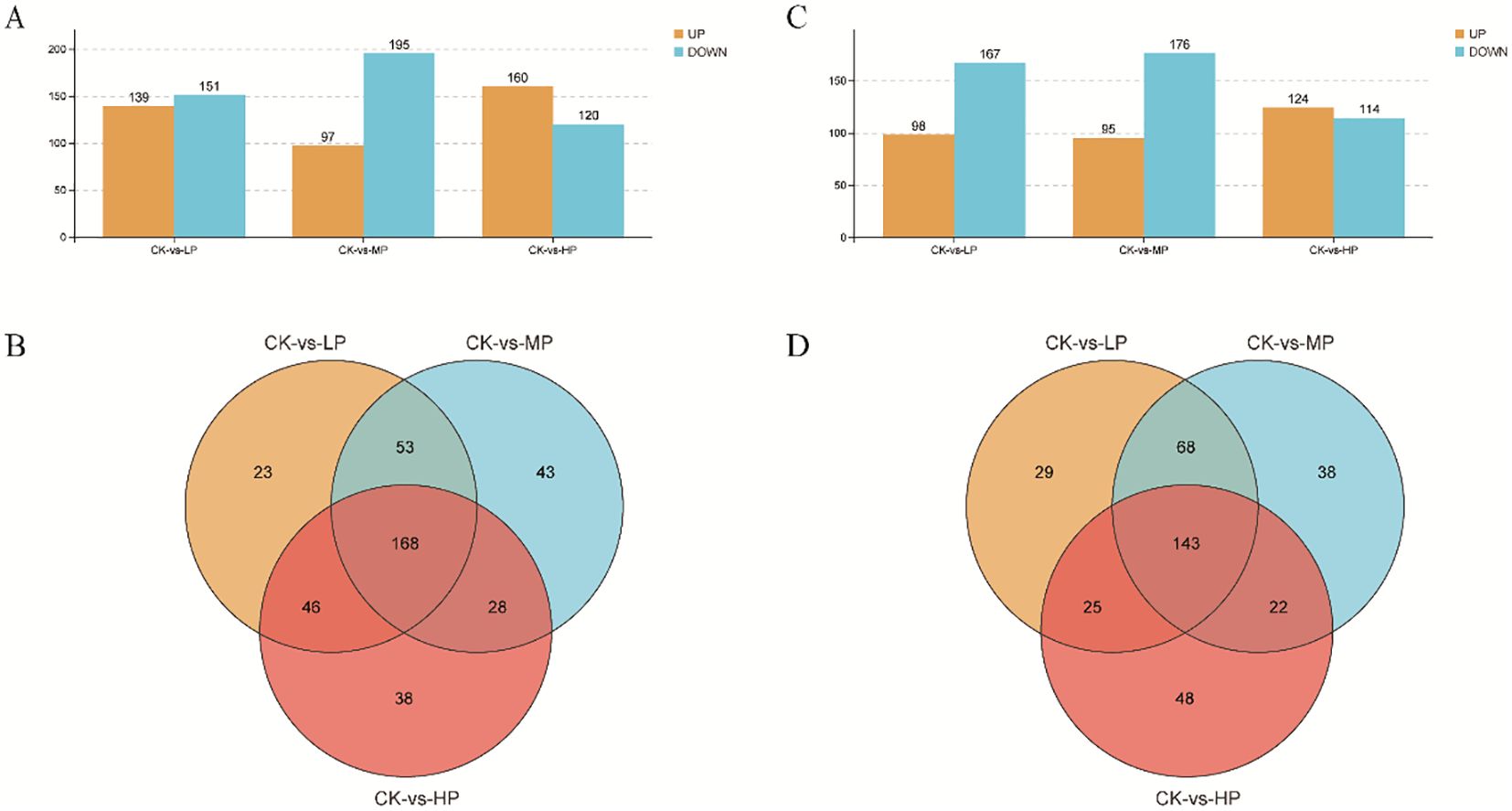
Figure 7. The metabolites of positive ions (A, B) and negative ions (C, D) were differentially expressed in the second internode of the rice base under different concentrations of paclobutrazol. CK, spray water treatment; LP, 100 mg/L of paclobutrazol was sprayed on S3 stage (August 2); MP, 200 mg/L of paclobutrazol was sprayed on S3 stage (August 2); HP, 300 mg/L of paclobutrazol was sprayed on S3 stage (August 2).
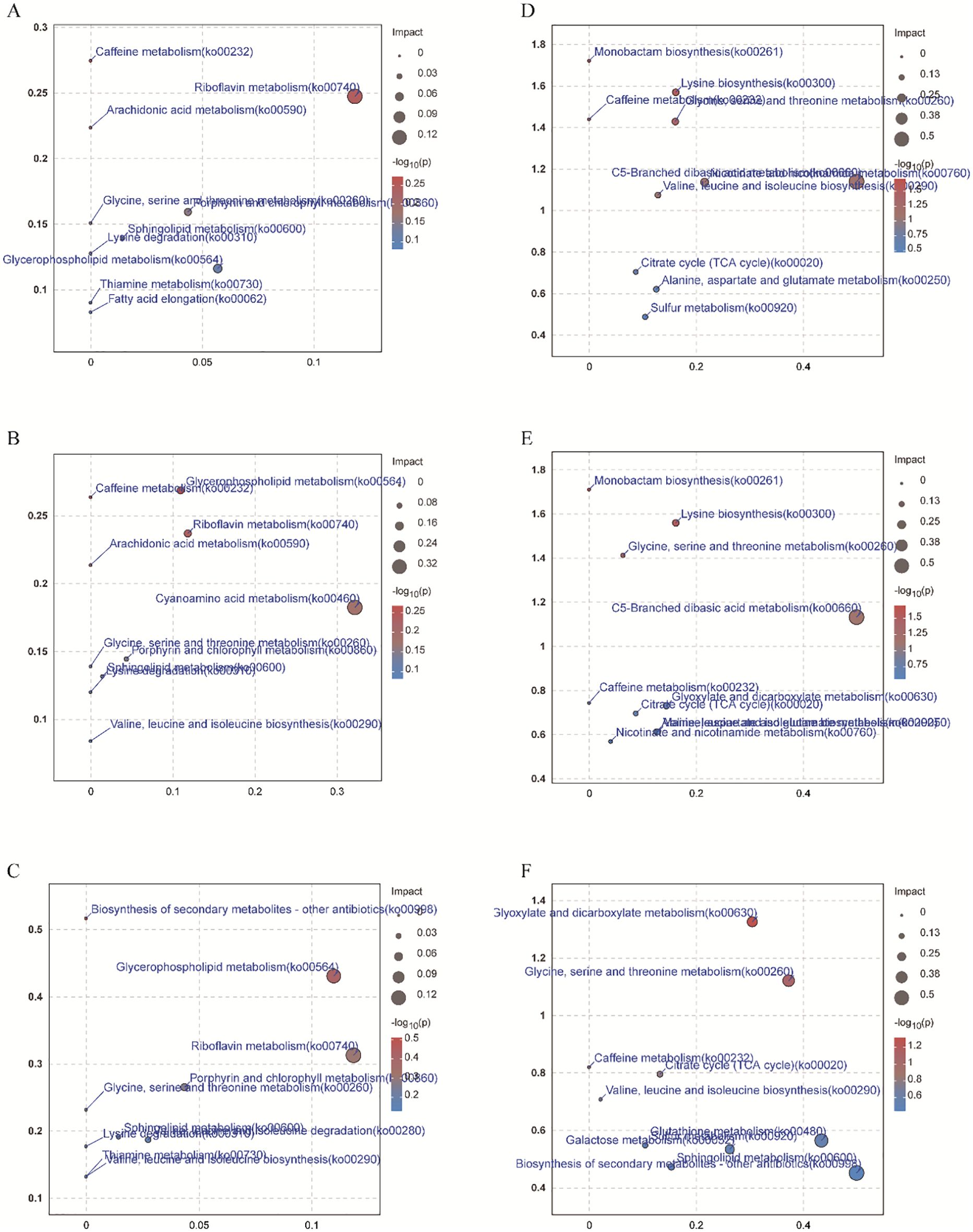
Figure 8. The MetPA analysis of positive ion (A, B, C) and negative ion (D, E, F) metabolites of paclobutrazol was different under LP (A, D), MP (B, E), and HP (C, F) treatments, respectively, compared with CK.
4 Discussion
Previous studies have demonstrated that a moderate concentration of paclobutrazol can increase rice yield, but a high concentration of paclobutrazol decreases rice yield (Sinniah et al., 2012; Syahputra et al., 2016). Our study showed that rice yield exhibited a significant decreasing trend with the increasing concentration of paclobutrazol during the same spraying period from 2023 to 2024 (Table 1). Syahputra et al. (2016) reported that high doses of paclobutrazol inhibited the development of rice spikelets. Analysis of yield components revealed that the decrease in yield was associated with the decrease in effective panicle, spikelets per panicle, and grain filling (Table 1). This might be attributed to the higher paclobutrazol application decreasing chlorophyll content and the net photosynthetic rate of the leaves, inhibiting rice dry matter accumulation, and reducing rice yield components (Assuero et al., 2012; Mehmood et al., 2021). The next step should be to explore the effects of paclobutrazol on rice yield from the perspectives of rice physiology such as enzymes, hormones, and grain filling characteristics. The spray period of paclobutrazol either decreased or had no significant effect on rice yield, with the exception of S4 in 2024 (Table 1). The significant reduction in spikelets per panicle and grain filling might be due to the inhibited rice growth and decreased dry matter accumulation during the tillering stage (S1 and S2 periods) (Liao et al., 2023). However, the spray period of paclobutrazol at the booting stage (S5 period) may affect anther activity or grain fertilization, thereby significantly reducing rice yield (Maheshwari et al., 2024). Notably, in 2024, the significant increase in yield and grain filling observed during the S4 period was attributed to varying degrees of rice lodging under other treatments, which were influenced by a typhoon.
Paclobutrazol is among the most widely utilized plant growth regulators, known for its ability to significantly enhance the mechanical strength of the internodes at the base of canola, sesame, and rice (Kuai et al., 2015; Mehmood et al., 2021; Liao et al., 2023). In this study, the LI was significantly reduced under different spraying periods of paclobutrazol in each internode (Table 2), corroborating the findings of previous research (Kuai et al., 2015; Liao et al., 2023). The lodging resistance of rice is influenced by various factors, including its physical, morphological, and anatomical characteristics, and the physicochemical components of the stem (Lv et al., 2022). The primary cause of crop lodging is the imbalance between the mechanical support capacity of the stem and the load imposed on the upper part of the plant (Lv et al., 2022). In this study, both PCA and correlation analysis indicated that the increase in breaking strength was the primary factor contributing to the significant decrease in each internode LI during the S5 period and under HP treatment (Table 2; Figure 1). Plant height, internode length, internode diameter, and culm wall thickness are critical indicators of stem morphology (Zhang et al., 2014). Previous studies have demonstrated that higher plant height increases the susceptibility of rice to lodging, while shorter internode lengths and larger internode diameters and culm wall thickness enhance lodging resistance (Lv et al., 2022). Our study indicates that internode diameter and culm wall thickness were negatively correlated with LI of rice (Figure 2). A strong source and stem is fundamental for achieving high and stable yields of rice (Wang et al., 2022). Numerous studies have demonstrated that the application of paclobutrazol in the jointing stage can increase the internode diameter and culm wall thickness, improve plant structure, and increase lodging resistance, thereby contributing to higher rice yields (Mehmood et al., 2021; Liao et al., 2023; Zhang et al., 2014). However, during the tillering stage, a significant decrease in plant height was observed following paclobutrazol injection, which may result in insufficient grain filling or even yield reduction (Wang et al., 2022). Similarly, we observed a significant reduction in plant height due to the application of paclobutrazol at the tillering stage (S2 period), whereas internode diameter and culm wall thickness were significantly increased by paclobutrazol at the jointing stage (S4 period) or booting stage (S5 period) (Table 3). In conclusion, the application of 200 mg/L of paclobutrazol at the jointing stage not only stabilizes rice yield but also significantly enhances lodging resistance by increasing internode diameter and culm wall thickness.
Previous studies have suggested that the application of paclobutrazol may inhibit the synthesis of gibberellin in rice, increase the activity of indoleacetic acid oxidase, and regulate the contents of indole-3-acetic acid (IAA) and abscisic acid (ABA) (Yan et al., 2016). These effects contribute to the inhibition of rice growth while promoting tillering, thereby enhancing lodging resistance and ultimately increasing yield. In our investigation of the effects of paclobutrazol spraying at various periods on rice stems, we identified the top five significant pathways associated with both positive and negative ion patterns through MetPA enrichment analysis (Supplementary Figures S2-S11). Furthermore, we performed a statistical analysis on the differential metabolites present within these pathways, as illustrated in Figures 5, 6. In this study, we found that the mandelonitrile content in the cyanoamino acid metabolism significantly increased during the jointing stage (Figure 5). This increase of mandelonitrile content may enhance the transport of essential nutrients in rice stems, facilitate the turnover process, and regulate oxidative stress (Arnaiz et al., 2023). Therefore, it promotes the elongation of the internode diameter of the stem (Table 3). The application of polybulobuzole at the jointing stage significantly increased the sarcosine content in glycine, serine, and threonine metabolism of rice stems (Figure 6). These results enhanced the production of glycine through oxidative demethylation, which positively influenced the growth and development of rice, promoting higher culm wall thickness and leaf area (Lahham et al., 2021). Our research also confirmed this view. Spraying paclobutrazol during the jointing stage may significantly promote the diameter of the stem internodes, especially the fourth internode, by significantly increasing the sarcosine content (Table 3). Glycerophospholipid metabolism is primarily associated with the functions of plant cell membranes (Nakamura, 2021). The application of paclobutrazol during the jointing stage significantly reduced the phosphatidylcholine content, compared to other paclobutrazol spray periods and CK (Figure 5). This reduction may inhibit certain aspects of the lipid metabolism process in rice stems (Nakamura, 2021). Similarly, the contents of paraxanthine and 1,7-dimethyluric acid are significantly decreased in caffeine metabolism (Figures 5, 6). The specific expression of these pathways and metabolites might be the main reason for the shortening of internode length during the jointing stage, especially the length of the third internode (Table 3). In future studies, the relationship between the alterations in metabolites induced by paclobutrazol spraying at varying time intervals and the morphological characteristics of rice stems will necessitate further verification across a broader range of rice varieties and over multiple years.
Plant growth regulators, such as paclobutrazol and prohexadione calcium, can enhance the lodging resistance of rice, and high concentrations of these regulators are detrimental to the growth and development of the plant (Lv et al., 2022; Liao et al., 2023). In the present study, the high concentrations of paclobutrazol significantly reduced the pinoresinol content in the biosynthesis of secondary metabolism of rice stems (Figure 9). Pinoresinol, a precursor in lignan synthesis, may impair the plant’s defense mechanisms and antioxidant capacity due to its reduced levels (Xiao et al., 2021). Previous studies have demonstrated that paclobutrazol can influence carbon and nitrogen metabolism, particularly in relation to organic acid and potassium metabolism (Sha et al., 2021). This might be the main reason for the significant reduction in internode dry weight and length at a high paclobutrazol concentration (Tables 3 and 4). Notably, isocitrate, a crucial intermediate in the tricarboxylic acid (TCA) cycle, was found to be significantly elevated following treatment with high concentrations of paclobutrazol, possibly disrupting the absorption and transport of potassium ions in stems (Sha et al., 2021), thereby reducing the dry weight and length of the internodes. Furthermore, our study revealed that isocitrate levels were also significantly increased in glyoxylate and dicarboxylate metabolism under high concentrations of paclobutrazol (Figure 10). This increase may have substantial implications for succinate, malate, metabolic energy regulation, and carbon skeleton supply (Kunze et al., 2006). The relationship between high-concentration paclobutrazol and the promotion of lateral internode growth necessitates further investigation. Pheophytin-a and leucine could increase rice stem nitrogen content, leaf photosynthetic utilization efficiency, stem growth, and stress resistance. Thus, the increase in pheophytin-a within porphyrin and chlorophyll metabolism, along with leucine in valine, leucine, and isoleucine degradation, might be the main reasons for the improvement of rice lodging resistance and stable yield through appropriate concentration of paclobutrazol treatment (Figure 9).
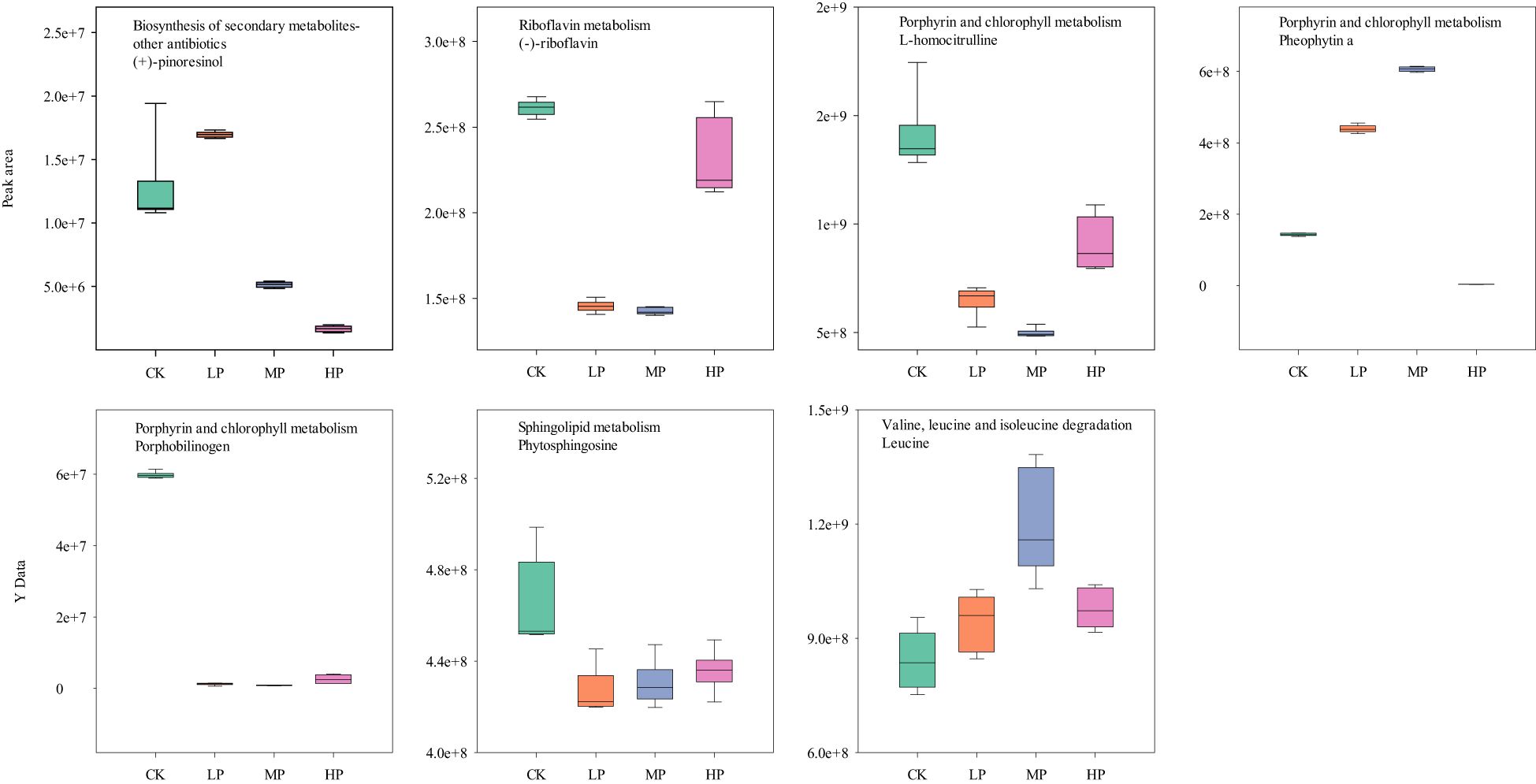
Figure 9. The MetPA analysis of positive ion differentially expressed metabolites under different concentrations of paclobutrazol. CK, spray water treatment; LP, 100 mg/L of paclobutrazol was sprayed on S3 stage (August 2); MP, 200 mg/L of paclobutrazol was sprayed on S3 stage (August 2); HP, 300 mg/L of paclobutrazol was sprayed on S3 stage (August 2).
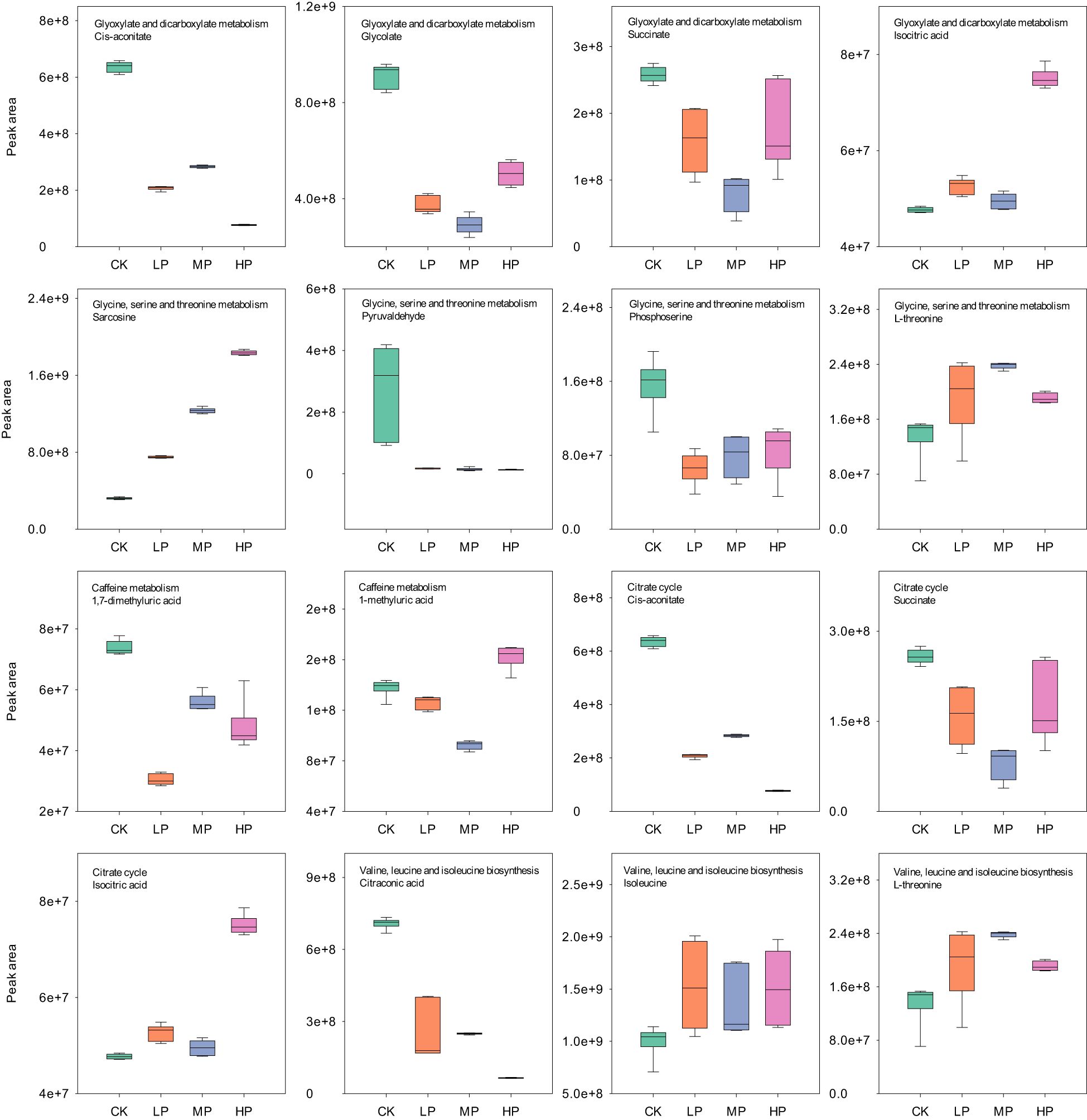
Figure 10. The MetPA analysis of negative ion differentially expressed metabolites under different concentrations of paclobutrazol. CK, spray water treatment; LP, 100 mg/L of paclobutrazol was sprayed on S3 stage (August 2); MP, 200 mg/L of paclobutrazol was sprayed on S3 stage (August 2); HP, 300 mg/L of paclobutrazol was sprayed on S3 stage (August 2).
5 Conclusions
Spraying paclobutrazol can significantly reduce the LI. However, spraying during the tillering stage (S1 and S2) and the young panicle stage (S5) results in decreased rice yield due to a reduction in the spikelets per panicle and grain filling. Additionally, high concentrations of paclobutrazol during these stages are not conducive to grain firming and yield formation. In contrast, applying paclobutrazol during the jointing stage (S4) under MP treatment can stabilize or even increase yield while simultaneously reducing the LI, making this the optimal period and concentration for spraying. Spraying paclobutrazol at the tillering stage primarily reduces plant height, internode length, gravity center height, and bending moment, thereby lowering the LI. Conversely, spraying during the jointing and young panicle stages primarily increases internode diameter and culm wall thickness, which also contributes to a reduction in the LI. The non-target metabolome results at various paclobutrazol spraying periods indicated a significant increase in mandelonitrile and sarcosine contents within cyanoamino acid metabolism and glycine, serine, and threonine metabolism during the S4 period, suggesting that it is beneficial to the development and thickening of stem walls. The non-target metabolome results from varying concentrations of paclobutrazol treatment suggest that the increase in pheophytin-a within porphyrin and chlorophyll metabolism, along with leucine in the degradation of valine, leucine, and isoleucine, might be key contributors to the enhancement of rice lodging resistance and stable yield under MP treatment.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
QZ: Writing – original draft, Writing – review & editing, Project administration. RX: Writing – original draft, Data curation, Methodology, Software, Visualization, Writing – review & editing. JT: Writing – original draft, Data curation, Methodology, Software, Visualization. RY: Data curation, Methodology, Software, Visualization, Writing – original draft. LC: Writing – review & editing, Conceptualization, Project administration, Validation, Writing – original draft. JB: Project administration, Writing – review & editing, Conceptualization, Validation, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The study was funded by the Shanghai Agricultural Science and Technology Innovation Program (Grant No. T2024316) and the Chongming Agriculture Innovation Project (Grant No. 2022CNKC-01-02).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2025.1603904/full#supplementary-material
References
Arnaiz A., Garcia J. L. V., Vallejos S., and Diaz I. (2023). Isolation and quantification of mandelonitrile from arabidopsis thaliana using gas chromatography/mass spectrometry. Bio-protocol. 13, e4700. doi: 10.21769/BioProtoc.4700
Assuero S., Lorenzo M., Ramírez N. P., V elázquez L., and Tognetti J. (2012). Tillering promotion by paclobutrazol in wheat and its relationship with plant carbohydrate status. New Z. J. Agric. Res. 55, 347–358. doi: 10.1080/00288233.2012.706223
Azarcon R. P., Vizmonte P. T., and Agustin A. (2022). Effects of paclobutrazol on growth, yield and water use efficiency of rice (Oryza sativa L.) under drought stress condition. Mindanao. J. Sci. Technology. 20, 38–60. doi: 10.61310/mndjstea.0959.22
Chen L. M., Yi Y. H., Wang W. X., Zeng Y. J., Tan X. M., Wu Z. M., et al. (2021). Innovative furrow ridging fertilization under a mechanical direct seeding system improves the grain yield and lodging resistance of early indica rice in South China. Field Crops Res. 270, 108184. doi: 10.1016/j.fcr.2021.108184
Desta B. and Amare G. (2021). Paclobutrazol as a plant growth regulator. Chem. Biol. Technol. Agric. 8, 1. doi: 10.1186/s40538-020-00199-z
Gai D. S., Liu W. Y., Liang J. N., Guo L. Y., Geng Y. Q., Zhang Q., et al. (2023). The Effects of paclobutrazol seed soaking on biomass production and yield formation in direct-seeded rice. Agronomy. 13, 1402. doi: 10.3390/agronomy13051402
Islam M. S., Peng S. B., Visperas R. M., Ereful N., Bhuiya M., and Julfiquar A. W. (2007). Lodging-related morphological traits of hybrid rice in a tropical irrigated ecosys-tem. Field Crops Res. 101, 240–248. doi: 10.1016/j.fcr.2006.12.002
IUSS Working Group and WRB (2006). World reference base for soil resources 2006. World Soil Resources Reports, No.103 Vol. 17) (Rome: FAO). Available at: http://refhun.elsevier.com/S0378-4290 (Access date December 27, 2024). 31810-5/sbref0095.
Kamran M., Ahmad S., Ahmad I., Hussain I., Meng X., Zhang X., et al. (2020). Paclobutrazol application favors yield improvement of maize under semiarid regions by delaying leaf senescence and regulating photosynthetic capacity and antioxidant system during grain-filling stage. Agronomy. 10, 187. doi: 10.3390/agronomy10020187
Khunpon B., Cha-um S., Faiyue B., Uthaibutra J., and Saengnil K. (2018). Paclobutrazol mitigates salt stress in indica rice seedlings by enhancing glutathione metabolism and glyoxalase system. Biologia 73, 1267–1276. doi: 10.2478/s11756-018-0132-4
Kuai J., Yang Y., Sun Y. Y., Zhou G. S., Zuo Q. S., Wu J. S., et al. (2015). Paclobutrazol increases canola seed yield by enhancing lodging and pod shatter resistance in Brassica napus L. Field Crops Res. 180, 10–20. doi: 10.1016/j.fcr.2015.05.004
Kunze M., Pracharoenwattana I., Smith S. M., and Hartig A. (2006). A central role for the peroxisomal membrane in glyoxylate cycle function. Biochim. Biophys. Acta (BBA). - Mol. Cell Res. 1763, 1441–1452. doi: 10.1016/j.bbamcr.2006.09.009
Lahham M., Jha S., Goj D., Macheroux P., and Wallner S. (2021). The family of sarcosine oxidases: Same reaction, different products. Arch. Biochem. Biophysics. 704, 108868. doi: 10.1016/j.abb.2021.108868
Li G. M., Tang J. Y., Zheng J. K., and Chu C. C. (2021). Exploration of rice yield potential: Decoding agronomic and physiological traits. Crop J. 9, 577–589. doi: 10.1016/j.cj.2021.03.014
Liao P., Bell S. M., Chen L., Huang S., Wang H. Y., Miao J. H., et al. (2023). Improving rice grain yield and reducing lodging risk simultaneously: A meta-analysis. Eur. J. Agronomy. 143, 126709. doi: 10.1016/j.eja.2022.126709
Lv R. J., Zhang W. J., Xie X. B., Wang Q. J., Gao K. G., Zeng Y. H., et al. (2022). Foliar application uniconazole enhanced lodging resistance of high-quality indica rice (Oryza sativa L.) by altering anatomical traits, cell structure and endogenous hormones. Field Crops Res. 277, 108425. doi: 10.1016/j.fcr.2021.108425
Maheshwari C., Garg N. K., Singh A., and Tyagi A. (2023). Optimization of paclobutrazol dose for mitigation of water-deficit stress in rice (Oryza Sativa L.). Biochem. Systematics Ecology. 107, 104596. doi: 10.1016/j.bse.2023.104596
Maheshwari C., Garg N. K., Singh A., and Tyagi A. (2024). Ameliorative effects of paclobutrazol via physio-biochemical and molecular manifestation in rice under water deficit stress. Rice Science. 31, 603–616. doi: 10.1016/j.rsci.2024.03.004
Mehmood M. Z., Qadir G., Afzal O., Din A. M. U., Raza M. A., Khan I., et al. (2021). Paclobutrazol improves sesame yield by increasing dry matter accumulation and reducing seed shattering under rainfed conditions. Int. J. Plant Production. 15, 337–349. doi: 10.1007/s42106-021-00132-w
Nakamura Y. (2021). Headgroup biosynthesis of phosphatidylcholine and phosphatidylethanolamine in seed plants. Prog. Lipid Res. 82, 101091. doi: 10.1016/j.plipres.2021.101091
Opio P., Tomiyama H., Saito T., Ohkawa K., Ohara H., and Kondo S. (2020). Paclobutrazol elevates auxin and abscisic acid, reduces gibberellins and zeatin and modulates their transporter genes in MArubakaido apple (Malus prunifolia Borkh. var. ringo Asami) rootstocks. Plant Physiol. Biochem. 155, 502–511. doi: 10.1016/j.plaphy.2020.08.003
Pan J. F., Zhao J. L., Liu Y. Z., Huang N. R., Tian K., Shah F., et al. (2019). Optimized nitrogen management enhances lodging resistance of rice and its morpho-anatomical, mechanical, and molecular mechanisms. Sci. Rep. 9, 848–856. doi: 10.1038/s41598-019-56620-7
Perik R. R., Raze D., Harkema H., Zhong Y., and Doorm W. G. V. (2012). Bending in cut Gerbera jamesonii flowers relates to adverse water relations and lack of stem sclerenchyma development, not to expansion of the stem central cavity or stem elongation. Postharvest. Biol. Technology. 74, 11–18. doi: 10.1016/j.postharvbio.2012.06.009
Ray D. K., Mueller N. D., West P. C., and Foley J. A. (2013). Yield trends are insufficient to double global crop production by 2050. PloS One 8, e66428. doi: 10.1371/journal.pone.0066428
Robinson A., Lehmann J., Barriopedro D., Rahmstorf S., and Coumou D. (2021). Increasing heat and rainfall extremes now far outside the historical climate. NPJ Climate Atmospheric. Science. 4, 45. doi: 10.1038/s41612-021-00202-w
Sha J. C., Ge S. F., Zhu Z. L., Du X., Zhang X., Xu X. X., et al. (2021). Paclobutrazol regulates hormone and carbon-nitrogen nutrition of autumn branches, improves fruit quality and enhances storage nutrition in ‘Fuji’ apple. Scientia. Horticulturae. 282, 110022. doi: 10.1016/j.scienta.2021.110022
Sinniah U. R., Wahyuni S., Syahputra B. S. A., and Gantait S. (2012). A potential retardant for lodging resistance in direct seeded rice (Oryza sativa L.). Can. J. Plant Science. 92, 13–18. doi: 10.4141/cjps2011-089
Syahputra B. S. A., Sinniah U. R., Ismail M. R., and Swamy M. K. (2016). Optimization of paclobutrazol concentration and application time for increased lodging resistance and yield in field-grown rice. Philippine. Agric. Scientist. 99, 221–228.
Wang M., Wang Y. P., Wang X. H., Wei G. W., Yang H. Y., Yang X., et al. (2024). Integrated physiological, biochemical, and transcriptomics analyses reveal the underlying mechanisms of high nitrogen use efficiency of black sesame. Plant Physiol. Biochem. 206, 108205. doi: 10.1016/j.plaphy.2023.108205
Wang H. X., Xiong R. Y., Zhou Y. Z., Tan X. M., Pan X. H., Zeng Y. J., et al. (2022). Grain yield improvement in high-quality rice varieties released in southern China from 2007 to 2017. Front. Sustain. Food Systems. 6. doi: 10.3389/fsufs.2022.986655
Wang J., Zhu J. M., Lin Q. Q., Li X. J., Teng N. J., Li Z. S., et al. (2006). Effects of stem structure and cell wall components on bending strength in wheat. Chin. Sci. Bulletin. 51, 815–823. doi: 10.1007/s11434-006-0815-z
Xiao Y., Shao K., Zhou J. W., Wang L., Ma X. Q., Wu D., et al. (2021). Structure-based engineering of substrate specificity for pinoresinol-lariciresinol reductases. Nat. Commun. 12, 2828. doi: 10.1038/s41467-021-23095-y
Yan K. L., Han Y., Tan T. G., Tang L. D., and Wu J. H. (2016). Effects of 15% paclobutrazol WP on growth, yield and quality in rice. Chin. J. Trop. Agriculture. 36, 73–76.
Zhang R., Jia Z., Ma X., Ma H., and Zhao Y. (2020). Characterising the morphological characters and carbohydrate metabolism of oat culms and their association with lodging resistance. Plant Biol. 22, 267–276. doi: 10.1111/plb.13058
Zhang J., Li G. H., Song Y. P., Liu Z. H., Yang C. D., Tang S., et al. (2014). Lodging resistance characteristics of high-yielding rice populations. Field Crops Res. 161, 64–74. doi: 10.1016/j.fcr.2014.01.012
Zhang J. E., Quan G. M., Huang Z. X., Luo S. M., and Ouyang Y. (2013). Evidence of duck activity induced anatomical structure change and lodging resistance of rice plant. Agroecology. Sustain. Food Systems. 37, 975–984. doi: 10.1080/21683565.2013.775688
Keywords: Japonica rice, lodging resistance, paclobutrazol, stem morphology, metabonomics
Citation: Zhao Q, Xiong R, Tang J, Yang R, Cao L and Bai J (2025) Effects of different spraying periods and concentrations of paclobutrazol on the lodging resistance and yield of high-quality japonica rice. Front. Agron. 7:1603904. doi: 10.3389/fagro.2025.1603904
Received: 01 April 2025; Accepted: 21 May 2025;
Published: 16 June 2025.
Edited by:
Ping Liao, Yangzhou University, ChinaCopyright © 2025 Zhao, Xiong, Tang, Yang, Cao and Bai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jianjiang Bai, YmFpamlhbmppYW5nMjAxMEAxNjMuY29t
†These authors have contributed equally to this work