 Christine L. Xu1,2†
Christine L. Xu1,2† Galaxy Y. Cho1,2,3†
Galaxy Y. Cho1,2,3† Jesse D. Sengillo1,2,4Karen S. Park1,2Vinit B. Mahajan5,6*
Jesse D. Sengillo1,2,4Karen S. Park1,2Vinit B. Mahajan5,6* Stephen H. Tsang1,2,7,8*
Stephen H. Tsang1,2,7,8*- 1Jonas Children's Vision Care, Bernard & Shirlee Brown Glaucoma Laboratory, Columbia University, New York, NY, United States
- 2Department of Ophthalmology, Columbia University, New York, NY, United States
- 3Frank. H. Netter MD School of Medicine, Quinnipiac University, North Haven, CT, United States
- 4State University of New York Downstate Medical Center, Brooklyn, NY, United States
- 5Omics Lab, Department of Ophthalmology, Byers Eye Institute, Stanford University, Palo Alto, CA, United States
- 6Palo Alto Veterans Administration, Palo Alto, CA, United States
- 7Department of Pathology & Cell Biology, Columbia University, New York, NY, United States
- 8Institute of Human Nutrition, College of Physicians and Surgeons, Columbia University, New York, NY, United States
In recent years, there has been accelerated growth of clustered regularly interspaced short palindromic repeats (CRISPR) genome surgery techniques. Genome surgery holds promise for diseases for which a cure currently does not exist. In the field of ophthalmology, CRISPR offers possibilities for treating inherited retinal dystrophies. The retina has little regenerative potential, which makes treatment particularly difficult. For such conditions, CRISPR genome surgery methods have shown great potential for therapeutic applications in animal models of retinal dystrophies. Much anticipation surrounds the potential for CRISPR as a therapeutic, as clinical trials of ophthalmic genome surgery are expected to begin as early as 2018. This mini-review summarizes preclinical CRISPR applications in the retina and current CRISPR clinical trials.
Background
After the first application of clustered regularly interspaced short palindromic repeats (CRISPR)-mediated gene-editing in human cells in 2013 (Cong et al., 2013; Jinek et al., 2013; Mali et al., 2013), clinical applications of the CRISPR system have become highly anticipated. The CRISPR genome surgery tool is able to edit, delete, insert, activate, repress, epigenetically modify, and induce directed evolution (Doudna and Charpentier, 2014; Hess et al., 2016; Cabral et al., 2017). The growth in the diversity of techniques has potentiated translation from bench to bedside. Conventional gene therapy, which helps restore expression of functional gene products by wild-type gene supplementation, is limited to recessively-inherited diseases and conditions of haploinsufficiency (Sengillo et al., 2016). When a wild-type (WT) gene is supplemented to a cell with a dominantly expressed pathogenic gene, there is no effect on disease progression (Lin et al., 2015). Thus, correction of the pathogenic gene is imperative. As such, the CRISPR system's potential to approach dominantly-inherited conditions is of particular interest (Sengillo et al., 2016; Cabral et al., 2017; Tsai et al., 2018).
While the CRISPR system has great potential for expanding the range of possible treatments for inherited diseases, it cannot be considered a perfect system. Possibly of greatest concern is off-targeting, or unexpected mutations that arise in the process of CRISPR activity (Jamal et al., 2016; Cho et al., 2017; Schaefer et al., 2017). Control of or better understanding of off-targeting should be addressed before CRISPR can be implemented in a broader range of clinical applications.
Current Clinical Trials
In October 2016, the first CRISPR clinical trial officially began when Sichuan University recruited the first subject (NCT02793856). So far, 10 CRISPR clinical trials that utilize the CRISPR genome editing tool are registered on ClinicalTrials.gov (Table 1). All of these trials focus on treating various malignancies such as neoplasms and HIV-infection (Table 1). Of the trials that seek to target cancer, six utilize CRISPR-Cas9 to engineer programmed cell death protein (PD-1) knockout T cells. PD-1 is a T cell immune checkpoint receptor that interacts with the programmed cell death ligand (PD-L1) on dendritic cells (Su et al., 2016). Normally, when PD-L1 binds to PD-1, the immune response is dampened via peripheral tolerance, which suppresses T-cell proliferation and reduces cytokine production (Fife and Pauken, 2011; Kuol et al., 2018). Cancer cells, however, also express PD-L1. This hijacks the dampening effects of PD-1/PD-L1 interactions. The downside to the currently employed method to block PD-1 receptors—monoclonal antibody delivery—is that it may have unwanted effects on peripheral tolerance by disrupting endogenous dendritic cell/T cell interactions and immune homeostasis (Su et al., 2016). Thus, an ex-vivo approach may be favored to inhibit the PD-1 gene in T cells, which may help avoid problems such as adverse autoimmune effects (Lloyd et al., 2013), yielding a more targeted and efficacious T-cell based therapy.
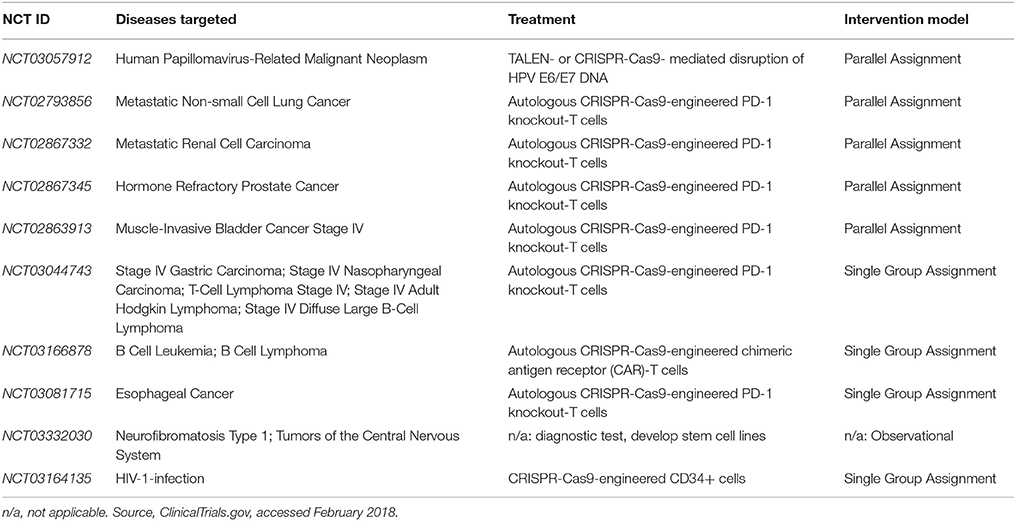
Table 1. CRISPR-Cas9 Clinical Trials as of February 2018.
Preclinical Ophthalmic Gene-Editing
As progress in clinical application of CRISPR genome surgery is made in the field of oncology, it is expected that ophthalmology will soon see clinical advances as well. One of the reasons for this expectation is that ophthalmic genome surgery offers many advantages such as relative immune privileged status due to the blood-retina barrier (Yang et al., 2015). Furthermore, the effects of treatment can be precisely monitored at the resolution of a single cell with non-invasive adaptive optics imaging (Park et al., 2013; Bae et al., 2014; Song et al., 2015, 2018; Yang et al., 2015; Zhang et al., 2015). Another reason for expectation of clinical advances is the demonstrated success in pre-clinical in vivo models of retinal dystrophies (Table 2). Table 2 outlines CRISPR-Cas9-mediated successful gene rescue and disease model rescue of various ophthalmic pathogenic genes. While there exists many other successful experiments showcasing the correction of pathogenic genes using CRISPR-Cas9, the pre-clinical in vivo models are outlined in this review because in vivo models point most directly toward potential clinical applications. Briefly, other applications of CRISPR-Cas9 technology in the retina include disease model generation (Zhong et al., 2015; Arno et al., 2016; Collery et al., 2016) and expanded understanding of disease mechanism (Bassuk et al., 2016; Latella et al., 2016; Yiu et al., 2016; Dong et al., 2017; Kim et al., 2017; Ruan et al., 2017; Yu et al., 2017).
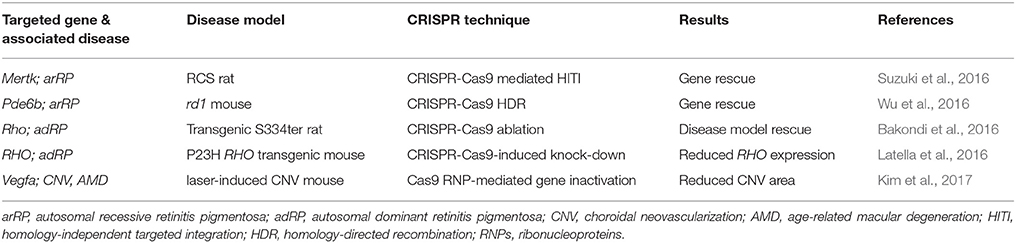
Table 2. In vivo CRISPR-Cas9 corrections of preclinical retinal disease models.
One of the new frontiers for CRISPR research in the retina is the modeling and treatment of Leber congenital amaurosis (LCA). LCA, a retinal dystrophy, is the largest cause of childhood blindness worldwide and it can involve up to 20 different genes (Chacon-Camacho and Zenteno, 2015; Maeder et al., 2015). Thus, LCA has been categorized into many subtypes. The gene CEP290 is implicated in LCA10, which causes 30% of all LCA (Maeder et al., 2015). CEP290 is very large (containing 54 exons and 7,440 bp in the open reading frame) and thus canonical gene augmentation techniques are difficult due to the limited carrying capacity of the adeno-associated virus (AAV) (Ruan et al., 2017). In May 2015, Maeder et al. reported that CRISPR-Cas9 could fix the cryptic splice site causing the most common mutation in CEP290 (IVS26 C.2991+1655 A>G mutation) in fibroblasts from LCA10 patients (Maeder et al., 2015). CRISPR-Cas9 fixed the single point mutation—what scientists now call the IVS26 mutation—using two sgRNAs that target sites flanking the aberrant splice site. This allowed S. aureus Cas9 to make two double stranded cuts and “repair” the mutant splice site by non-homologous end joining (NHEJ). Expression of the correct CEP290 transcript due to the removal of the premature stop codon was checked by qRT-PCR.
In February 2017, Ruan et al. successfully removed the IVS26 mutation in CEP290 from HEK293FT cells carrying the IVS26 mutation (Ruan et al., 2017). They achieved genomic deletion with a pair of sgRNAs, which respectively flanked the splice site in both upstream and downstream directions. After a proof-of-concept in vitro experiment, WT mice were used to study in vivo applications of CRISPR mediated Cep290 editing. Wild-type mice were chosen because an animal disease model for LCA10 does not currently exist. Two sgRNAs and SpCas9 were packaged into separate AAV plasmids in a dual AAV approach, which mediated the deletion of an intronic fragment of the Cep290 gene in WT mice. The purpose of the dual AAV approach was to circumvent the limitations of AAV carrying capacity.
Translation of CEP290 CRISPR-Cas9 treatment from bench to bedside may be imminent, because Editas Medicine (Cambridge, MA, USA) recently announced in a press release that clinical trials for CEP290-associated LCA treated with CRISPR-Cas9 technology would begin in 2018 (Taylor, 2017). Further developments in clinical application for other causative mutation retinal dystrophies are highly anticipated as patients suffering from blinding inherited eye disease may gain options for previously untreatable conditions (Doudna and Charpentier, 2014; Sengillo et al., 2016, 2017; Cabral et al., 2017). While expectations are rising, an important aspect of the CRISPR system to consider is undesired off-target mutagenesis, a by-product of CRISPR-Cas9 editing because CRISPR-Cas9 does not have 100% specificity (Tsai and Joung, 2016; Schaefer et al., 2017). However, it should also be acknowledged that there is progress toward understanding off-targeting and reducing off-targeting effects (Tsai and Joung, 2016). One example is Shin et al.'s recent study which demonstrated that the delivery of anti-CRISPR protein AcrIIIA4 in human cells could reduce CRISPR-Cas9 active time. Of note, the team reported reduced off-target editing while on-target CRISPR-Cas9 genome editing remained intact (Shin et al., 2017). Other efforts to address off-targeting include improvements in off-targeting prediction and detection (Tsai et al., 2015; Tsai and Joung, 2016). While the consequences of off-targeting are potentially serious, there is continued development in improving CRISPR-Cas9 specificity (Tsai and Joung, 2016).
Conclusion
CRISPR genome surgery techniques hold great potential to treat previously unapproachable conditions. While the CRISPR genome surgery system cannot be considered infallible, accelerated progress in recent years has allowed development toward a more specific CRISPR-Cas9. As further developments in CRISPR technology are made to increase on-target accuracy and decrease off-target cleavage, it is expected that the list of CRISPR clinical trials will continue to grow. Especially in ophthalmology, preclinical in vivo successes point toward future CRISPR applications in the clinical setting.
Author Contributions
CX and GC contributed equally to this work. CX, GC, JS, and KP wrote and edited manuscript. VM and ST oversaw the writing process.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer LG and handling Editor declared their shared affiliation.
Acknowledgments
Supported, in part, by grants from National Eye Institute, NIH; P30EY019007, R01EY018213, R01EY024698, National Cancer Institute Core [5P30CA013696]. ST is a member of the RD-CURE Consortium and is supported by the Tistou and Charlotte Kerstan Foundation, the Schneeweiss Stem Cell Fund, New York State [C029572], the Foundation Fighting Blindness New York Regional Research Center Grant [C-NY05-0705-0312], the Crowley Family Fund, and the Gebroe Family Foundation. VM is supported by NIH grants [R01EY024665, R01EY025225, R01EY024698], and Research to Prevent Blindness (RPB), New York, NY.
References
Arno, G., Agrawal, S. A., Eblimit, A., Bellingham, J., Xu, M., Wang, F., et al. (2016). Mutations in REEP6 cause autosomal-recessive retinitis pigmentosa. Am. J. Hum. Genet. 99, 1305–1315. doi: 10.1016/j.ajhg.2016.10.008
Bae, E. J., Kim, K. R., Tsang, S. H., Park, S. P., and Chang, S. (2014). Retinal damage in chloroquine maculopathy, revealed by high resolution imaging: a case report utilizing adaptive optics scanning laser ophthalmoscopy. Kor. J. Ophthalmol. 28, 100–107. doi: 10.3341/kjo.2014.28.1.100
Bakondi, B., Lv, W., Lu, B., Jones, M. K., Tsai, Y., Kim, K. J., et al. (2016). In vivo CRISPR/Cas9 gene editing corrects retinal dystrophy in the S334ter-3 rat model of autosomal dominant retinitis pigmentosa. Mol. Ther. 24, 556–563. doi: 10.1038/mt.2015.220
Bassuk, A. G., Zheng, A., Li, Y., Tsang, S. H., and Mahajan, V. B. (2016). Precision medicine: genetic repair of retinitis pigmentosa in patient-derived stem cells. Sci. Rep. 6:19969. doi: 10.1038/srep19969
Cabral, T., DiCarlo, J. E., Justus, S., Sengillo, J. D., Xu, Y., and Tsang, S. H. (2017). CRISPR applications in ophthalmologic genome surgery. Curr. Opin. Ophthalmol. 28, 252–259. doi: 10.1097/ICU.0000000000000359
Chacon-Camacho, O. F., and Zenteno, J. C. (2015). Review and update on the molecular basis of Leber congenital amaurosis. World J. Clin. Cases 3, 112–124. doi: 10.12998/wjcc.v3.i2.112
Cho, G. Y., Abdulla, Y., Sengillo, J. D., Justus, S., Schaefer, K. A., Bassuk, A. G., et al. (2017). CRISPR-mediated ophthalmic genome surgery. Curr. Ophthalmol. Rep. 5, 199–206. doi: 10.1007/s40135-017-0144-1
Collery, R. F., Volberding, P. J., Bostrom, J. R., Link, B. A., and Besharse, J. C. (2016). Loss of Zebrafish Mfrp causes nanophthalmia, hyperopia, and accumulation of subretinal macrophages. Invest. Ophthalmol. Vis. Sci. 57, 6805–6814. doi: 10.1167/iovs.16-19593
Cong, L., Ran, F. A., Cox, D., Lin, S., Barretto, R., Habib, N., et al. (2013). Multiplex genome engineering using CRISPR/Cas systems. Science 339, 819–823. doi: 10.1126/science.1231143
Dong, Y., Cai, X., Wu, Y., Liu, Y., Deng, L., and Chen, H. (2017). Insights from genetic model systems of retinal degeneration: role of epsins in retinal angiogenesis and VEGFR2 signaling. J. Nat. Sci. 3:e281.
Doudna, J. A., and Charpentier, E. (2014). Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 346:1258096. doi: 10.1126/science.1258096
Fife, B. T., and Pauken, K. E. (2011). The role of the PD-1 pathway in autoimmunity and peripheral tolerance. Ann. N.Y. Acad. Sci. 1217, 45–59. doi: 10.1111/j.1749-6632.2010.05919.x
Hess, G. T., Fresard, L., Han, K., Lee, C. H., Li, A., Cimprich, K. A., et al. (2016). Directed evolution using dCas9-targeted somatic hypermutation in mammalian cells. Nat. Methods 13, 1036–1042. doi: 10.1038/nmeth.4038
Jamal, M., Khan, F. A., Da, L., Habib, Z., Dai, J., and Cao, G. (2016). Keeping CRISPR/Cas on-Target. Curr. Issues Mol. Biol. 20, 1–12. doi: 10.21775/cimb.020.001
Jinek, M., East, A., Cheng, A., Lin, S., Ma, E., and Doudna, J. (2013). RNA-programmed genome editing in human cells. Elife 2:e00471. doi: 10.7554/eLife.00471
Kim, K., Park, S. W., Kim, J. H., Lee, S. H., Kim, D., Koo, T., et al. (2017). Genome surgery using Cas9 ribonucleoproteins for the treatment of age-related macular degeneration. Genome Res. 27, 419-426 doi: 10.1101/gr.219089.116
Kuol, N., Stojanovska, L., Nurgali, K., and Apostolopoulos, V. (2018). PD-1/PD-L1 in disease. Immunotherapy 10, 149–160. doi: 10.2217/imt-2017-0120
Latella, M. C., Di Salvo, M. T., Cocchiarella, F., Benati, D., Grisendi, G., Comitato, A., et al. (2016). In vivo editing of the human mutant rhodopsin gene by electroporation of plasmid-based CRISPR/Cas9 in the mouse retina. Mol. Ther. Nucleic Acids 5:e389. doi: 10.1038/mtna.2016.92
Lin, M. K., Tsai, Y.-T., and Tsang, S. H. (2015). Emerging Treatments for Retinitis Pigmentosa: Genes and stem cells, as well as new electronic and medical therapies, are gaining ground. Retinal physician 12, 52–55.
Lloyd, A., Vickery, O. N., and Laugel, B. (2013). Beyond the antigen receptor: editing the genome of T-cells for cancer adoptive cellular therapies. Front. Immunol. 4:221. doi: 10.3389/fimmu.2013.00221
Maeder, M. L., Shen, S., Burnight, E. R., Gloskowski, S., Mepani, R., Friedland, A. E., et al. (2015). Therapeutic correction of an LCA-causing splice defect in the CEP290 Gene by CRISPR/Cas-mediated genome editing. Mol. Ther. 23, S273–S274. doi: 10.1016/S1525-0016(16)34296-4
Mali, P., Yang, L., Esvelt, K. M., Aach, J., Guell, M., DiCarlo, J. E., et al. (2013). RNA-guided human genome engineering via Cas9. Science 339, 823–826. doi: 10.1126/science.1232033
Park, S. P., Chung, J. K., Greenstein, V., Tsang, S. H., and Chang, S. (2013). A study of factors affecting the human cone photoreceptor density measured by adaptive optics scanning laser ophthalmoscope. Exp. Eye Res. 108, 1–9. doi: 10.1016/j.exer.2012.12.011
Ruan, G. X., Barry, E., Yu, D., Lukason, M., Cheng, S. H., and Scaria, A. (2017). CRISPR/Cas9-Mediated genome editing as a therapeutic approach for leber congenital amaurosis 10. Mol. Ther. 25, 331–341. doi: 10.1016/j.ymthe.2016.12.006
Schaefer, K. A., Wu, W. H., Colgan, D. F., Tsang, S. H., Bassuk, A. G., and Mahajan, V. B. (2017). Unexpected mutations after CRISPR-Cas9 editing in vivo. Nat. Methods 14, 547–548. doi: 10.1038/nmeth.4293
Sengillo, J. D., Justus, S., Cabral, T., and Tsang, S. H. (2017). Correction of monogenic and common retinal disorders with gene therapy. Genes (Basel) 8:E53. doi: 10.3390/genes8020053.
Sengillo, J. D., Justus, S., Tsai, Y. T., Cabral, T., and Tsang, S. H. (2016). Gene and cell-based therapies for inherited retinal disorders: an update. Am. J. Med. Genet. C Semin. Med. Genet. 172, 349–366. doi: 10.1002/ajmg.c.31534
Shin, J., Jiang, F., Liu, J. J., Bray, N. L., Rauch, B. J., Baik, S. H., et al. (2017). Disabling Cas9 by an anti-CRISPR DNA mimic. Sci. Adv. 3:e1701620. doi: 10.1126/sciadv.1701620
Song, H., Rossi, E. A., Latchney, L., Bessette, A., Stone, E., Hunter, J. J., et al. (2015). Cone and rod loss in Stargardt disease revealed by adaptive optics scanning light ophthalmoscopy. JAMA Ophthalmol. 133, 1198–1203. doi: 10.1001/jamaophthalmol.2015.2443
Song, H., Rossi, E. A., Stone, E., Latchney, L., Williams, D., Dubra, A., et al. (2018). Phenotypic diversity in autosomal-dominant cone-rod dystrophy elucidated by adaptive optics retinal imaging. Br. J. Ophthalmol. 102, 136–141. doi: 10.1136/bjophthalmol-2017-310498
Su, S., Hu, B., Shao, J., Shen, B., Du, J., Du, Y., et al. (2016). CRISPR-Cas9 mediated efficient PD-1 disruption on human primary T cells from cancer patients. Sci. Rep. 6:20070. doi: 10.1038/srep20070
Suzuki, K., Tsunekawa, Y., Hernandez-Benitez, R., Wu, J., Zhu, J., Kim, E. J., et al. (2016). In vivo genome editing via CRISPR/Cas9 mediated homology-independent targeted integration. Nature 540, 144–149. doi: 10.1038/nature20565
Taylor, N. P. (2017). Editas delays IND for Allergan-Partnered CRISPR Program. Newton, MA: FierceBiotech.
Tsai, S. Q., and Joung, J. K. (2016). Defining and improving the genome-wide specificities of CRISPR-Cas9 nucleases. Nat. Rev. Genet. 17, 300–312. doi: 10.1038/nrg.2016.28
Tsai, S. Q., Zheng, Z., Nguyen, N. T., Liebers, M., Topkar, V. V., Thapar, V., et al. (2015). GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases. Nat. Biotechnol. 33, 187–197. doi: 10.1038/nbt.3117
Tsai, Y.-T., Wu, W.-H., Lee, T.-T., Wu, W.-P., Xu, C. L., Park, K. S., et al. (2018). CRISPR-based genome surgery for the treatment of autosomal dominant retinitis pigmentosa. Ophthalmology doi: 10.1016/j.ophtha.2018.04.001
Wu, W. H., Tsai, Y. T., Justus, S., Lee, T. T., Zhang, L., Lin, C. S., et al. (2016). CRISPR repair reveals causative mutation in a preclinical model of retinitis pigmentosa. Mol. Ther. 24, 1388–1394. doi: 10.1038/mt.2016.107
Yang, T., Justus, S., Li, Y., and Tsang, S. H. (2015). BEST1: the best target for gene and cell therapies. Mol. Ther. 23, 1805–1809. doi: 10.1038/mt.2015.177
Yiu, G., Tieu, E., Nguyen, A. T., Wong, B., and Smit-McBride, Z. (2016). Genomic disruption of VEGF-A expression in human retinal pigment epithelial cells using CRISPR-Cas9 endonuclease. Invest. Ophthalmol. Vis. Sci. 57, 5490–5497. doi: 10.1167/iovs.16-20296
Yu, W., Mookherjee, S., Chaitankar, V., Hiriyanna, S., Kim, J. W., Brooks, M., et al. (2017). Nrl knockdown by AAV-delivered CRISPR/Cas9 prevents retinal degeneration in mice. Nat. Commun. 8:14716. doi: 10.1038/ncomms14716
Zhang, J., Yang, Q., Saito, K., Nozato, K., Williams, D. R., and Rossi, E. A. (2015). An adaptive optics imaging system designed for clinical use. Biomed. Opt. Express 6, 2120–2137. doi: 10.1364/BOE.6.002120
Keywords: genome surgery, CRISPR-Cas9, retinal dystrophies, clinical trials, off-target effect
Citation: Xu CL, Cho GY, Sengillo JD, Park KS, Mahajan VB and Tsang SH (2018) Translation of CRISPR Genome Surgery to the Bedside for Retinal Diseases. Front. Cell Dev. Biol. 6:46. doi: 10.3389/fcell.2018.00046
Received: 14 November 2017; Accepted: 12 April 2018;
Published: 23 May 2018.
Edited by:
Yuk Fai Leung, Purdue University, United StatesReviewed by:
Zhijian Wu, National Institutes of Health (NIH), United StatesLogan Ganzen, Purdue University, United States
Copyright © 2018 Xu, Cho, Sengillo, Park, Mahajan and Tsang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vinit B. Mahajan, bWFoYWphbmxhYkBnbWFpbC5jb20=
Stephen H. Tsang, c2h0MkBjdW1jLmNvbHVtYmlhLmVkdQ==
†These authors have contributed equally to this work.