 Mohamed A. Nasr
Mohamed A. Nasr Radwa Ayman Salah†
Radwa Ayman Salah† M. Abd Elkodous
M. Abd Elkodous- Center of Excellence for Stem Cells and Regenerative Medicine (CESC), Zewail City of Science and Technology, 6th of October City, Egypt
Hepatocellular carcinoma (HCC) is one of the top causes of cancer mortality worldwide. Although HCC has been researched extensively, there is still a need for novel and effective therapeutic interventions. There is substantial evidence that initiation of carcinogenesis in liver cirrhosis, a leading cause of HCC, is mediated by cancer stem cells (CSCs). CSCs were also shown to be responsible for relapse and chemoresistance in several cancers, including HCC. MicroRNAs (miRNAs) constitute important epigenetic markers that regulate carcinogenesis by acting post-transcriptionally on mRNAs, contributing to the progression of HCC. We have previously shown that co-culture of cancer cells with mesenchymal stem cells (MSCs) could induce the reprogramming of MSCs into CSC-like cells. In this review, we evaluate the available data concerning the epigenetic regulation of miRNAs through methylation and the possible role of this regulation in stem cell and somatic reprogramming in HCC.
Hepatocellular Carcinoma
Hepatocellular carcinoma (HCC) is the most frequent primary malignancy of the liver. It is the third leading cause of mortality associated with cancer worldwide (Yang and Roberts, 2010; Dhanasekaran et al., 2012). HCC is a multifactorial disease that is influenced by several risk factors. It typically develops as a result of underlying liver disease and is commonly associated with cirrhosis (Huang et al., 2013). The major HCC-risk factors include viral infection with hepatitis B virus (HBV) and hepatitis C virus (HCV), which leads to liver cirrhosis and accounts for 75% of HCC cases (El-Serag, 2002). Other factors attributed to HCC include alcohol abuse, intake of food contaminated with aflatoxin and toxic chemical exposure, including dimethylformamide, dimethylacetamide, trichloroethylene, tetrachloroethylene, carbon tetrachloride and chloroform (Malaguarnera et al., 2012). In addition, Obesity is one of the highly recent factors that plays a significant role in developing non-alcoholic fatty liver disease (NAFLD) (Cholankeril et al., 2017). It can progress in many stages starting with lipid deposition in hepatocytes' cytoplasm and can lead to non-alcoholic steatohepatitis (NASH) (Marrero et al., 2002; Guzman et al., 2008; Reddy et al., 2012; White et al., 2012). NASH is the severe stage of NAFLD indicated by hepatocyte injury, uncontrolled inflammation, hepatocyte ballooning, cell death, infiltration of inflammatory cells, and collagen deposition (Guzman et al., 2008; Reddy et al., 2012). NASH has been determined to be one of the important events in promoting hepatic carcinogenesis (Ip and Wang, 2014).
Tissue damage and fibrosis result from chronic inflammation and oxidative stress, leading to cirrhosis and eventually tumor initiation, progression, and even metastasis (Lau and Lai, 2008; Shariff et al., 2009; Cabrera and Nelson, 2010). Although only ~10–20% of HCC patients are eligible for surgical interference at the time of diagnosis, liver transplantation remains the first choice for treatment (Ji et al., 2009a). Furthermore, patients suffer a high frequency of relapse, and in patients who experience curative resection, the 5-year survival rate is 30–40% (Budhu et al., 2008). The low detection and high recurrence rates for the curable stages of HCC have increased interest in investigations of the molecular mechanisms underlying this disease (He et al., 2015).
Cancer Stem Cells (CSCs)
The failure of conventional treatments to completely eliminate invasive tumor cells is thought to be due to the presence of a small subset of cancer cells, termed CSCs, which are accountable for cancer progression, metastasis, recurrence, and drug resistance. CSCs have been classified as immortal tumor-initiating cells which have pluripotent and self-renewal capacity (Chen et al., 2013). CSCs have been identified in numerous solid tumors, such as breast cancer, colon cancer, and HCC (Szotek et al., 2006; O'Brien et al., 2007; Kawai et al., 2015). CSCs were found to have a main contribution in tumor heterogeneity and to contribute to drug resistance (Beck and Blanpain, 2013; Bedard et al., 2013; Klein, 2013; Meacham and Morrison, 2013). While the origin of CSCs remains unclear, the proposed mechanisms for their generation include cell fusion, genetic mutations in stem cells, and regulatory factors within the tumor microenvironment (TME) (Bu and Cao, 2012). In addition, signaling pathways and genes that regulate stem cell differentiation, as Wnt/β-catenin, transforming growth factor β (TGF-β), and microRNAs (miRNAs), could contribute to the control and maintenance of CSC differentiation (Bedard et al., 2013; Meacham and Morrison, 2013). The Wnt/β-catenin signaling pathway seems to play major roles in the development of CSCs and in self-renewal, tumorigenesis, and cancer chemoresistance (Espada et al., 2009; Eaves and Humphries, 2010; Mohammed et al., 2016).
Characteristics of miRNAs
MiRNAs are small non-coding RNA molecules consisting of 21−23 nucleotides. They control gene expression by base pairing with the messenger RNAs (mRNAs) (Lu et al., 2005; Griffiths-Jones et al., 2006). Transcripts are regulated through either degradation or translational repression (Bartel, 2004). Full complementarity between a miRNA and an mRNA results in full degradation of the target mRNA. However, defects in perfect complementarity leads to less translation of the target gene without affecting the level of mRNA (Lewis et al., 2005; Cummins and Velculescu, 2006). MiRNAs target up to 90% of human genes (Miranda et al., 2006) and can be found in exons or introns of coding or non-coding genes, with their transcription dependent on genomic localization (Baskerville and Bartel, 2005; Lin et al., 2006). Although miRNAs have their own promoters and are self-sufficiently expressed some miRNAs that share the same transcriptional regulation are ordered in clusters. Hundreds of miRNAs have been known by molecular cloning and bioinformatics approaches in plants and animals (Bushati and Cohen, 2007; Liu et al., 2014). Interestingly, a subgroup of miRNAs, namely, epi-miRNAs, control the expression of epigenetic marks, such as DNA methyltransferases (DNMTs), histone deacetylases (HDACs), and polycomb genes, either directly or indirectly. DNA methylation has a key role in gene expression regulation via maintaining the stability of gene silencing. In mammals, DNA methylation takes place by covalent modification of cytosine residues through the addition of a methyl group to the fifth position of a cytosine ring, particularly in the CpG dinucleotides. This process is mediated by members of the DNMTs enzymes family (Chuang and Jones, 2007). Therefore, miRNAs function as both genetic and epigenetic regulators (Valeri et al., 2009).
miRNAs control many cellular processes in eukaryotes, such as rate of growth, development, differentiation potential, cell cycle progression, and apoptosis, and their abnormal expression affects many human diseases (Valeri et al., 2009; Krol et al., 2010; Wahid et al., 2010; Pritchard et al., 2012). In addition to serving as essential players in tumor development, miRNAs have a role as possible biomarkers for cancer (Calin and Croce, 2006). Indeed, miRNA profiles reflect the stages of tumors and their developmental lineages (George and Mittal, 2010). MiRNAs have been found to modulate CSCs and metastasis. They can also act as oncogenes and tumor suppressors. Due to their functions as oncogenes and tumor suppressor genes, these miRNAs have been referred to as oncomirs (George and Mittal, 2010).
miRNAs and HCC
Recent studies on liver miRNAs investigated the overexpression of specific miRNAs or the inhibition of other miRNAs both in vitro and in vivo. These studies showed the crucial biological roles of miRNAs for proper liver function (Takata et al., 2013). In HCC, Murakami et al. initially reported dysregulated miRNA expression, with four miRNAs, namely, miR-18, miR-92, miR-20, and precursor miR-18 being inversely associated with the extent of HCC development (Murakami et al., 2006). Later, several studies confirmed that miRNAs play an essential regulatory role in hepatic carcinogenesis progression and malignant transformation. Some miRNAs showed abnormal expression during the progression of liver cancer (Zhao et al., 2009). The expression of some miRNAs was shown to influence HCC development via dysregulation of a number of cancer-associated molecular pathways, including TGF-β, p53, WNT/β-catenin, P13K/AKT/mTOR, RAS/MAPS, MET, and MYC (Negrini et al., 2011). Many oncogenic miRNAs have shown aberrant expression in HCC, including miR-1275 (Shaalan et al., 2018), miR-17-5p (Habashy et al., 2016), miR-96-5p, miR-182-5p (Assal et al., 2015), miR-155 (El Tayebi et al., 2015), and miR-181a (Lashine et al., 2011). Other tumor suppressor miRNAs involved in HCC include miR-34a (Yacoub et al., 2016), miR-486-5p (Youness et al., 2016), miR-615-5p (El Tayebi et al., 2012), and miR-Let7i (Fawzy et al., 2016). Genome-wide approaches have identified hundreds of miRNAs in HCC tumor tissues that were to be dysregulated compared to non-tumor tissues (Borel et al., 2012). MiR-122 is among many unique and well-studied dysregulated miRNAs that are highly expressed specifically in human liver. In HCC patients, a shorter recurrence time were attributed to lower levels of miR-122. While elevated expression of cyclin G1, a target of miR-122, was associated with a lower survival rate. Moreover, miR-122 acts as a tumor suppressor in HCC, and was subsequently reported to be downregulated in around 70% of cases (Callegari et al., 2013). MiR-221 is another critical oncogenic miRNA that is upregulated in 70–80% of HCC cases. Its overexpression leads to enhanced proliferation potential, migration, invasion, rate of growth, and decreased the rate of apoptosis in HCC patients (Fornari et al., 2008). Additionally, miR-221 modulates several gene targets involved in cancer-related pathways, including PTEN (P13K/AKT/mTOR), CDKN1B/p27, and CDKN1C/p57 (Fornari et al., 2008; Garofalo et al., 2009).
Due to their non-invasive detection, good specificity, and sensitivity, miRNAs are considered effective biomarkers for HCC (Shen et al., 2016). MiR-155-5p, miR-206, miR-21-5p, and miR-212-3p. MiR-155-5p and miR-21-5p which are reported as biomarkers for the prognosis of HCC in tissues, were found to have upregulated expression levels. On the other hand others were down-regulated (Han et al., 2013; Wang et al., 2014; Yunqiao et al., 2014; Tu et al., 2015). Circulating miR-122-5p and miR-16-5p could be used as presumed biomarkers for HCC. MiR-122-5p and miR-16-5p belong to this group which were particularly detected to be up and down-regulated, respectively (Cho et al., 2015; El-Abd et al., 2015).
Most often, elevated expression of miR-18b-5p, miR-200a-3p, miR-200b-3p, miR-21-5p, miR-224-5p, and miR-29-5p in tissue were mostly reported to be HCC. In addition, miR-139-5p was down-regulated. Therefore, they were beneficial for diagnosis of HCC (Zhu et al., 2012; Murakami et al., 2013; Dhayat et al., 2014; Han et al., 2014; Li T. et al., 2014; Amr et al., 2016).
Circulating miRNAs were proposed as prognostic biomarkers and reported to be linked to tissue invasion and metastasis. Those biomarkers included miR-122-5p, miR-17-5p, miR-182-5p, miR-21-5p, miR-24-3p, and miR-331-3p, all were up-regulated in the group reported to have a low-survival rate (Zheng et al., 2013; Meng et al., 2014; Chen et al., 2015; Wang L.-J. et al., 2015; Xu Y. et al., 2015). Meanwhile, the serum miR-150-5p was highly expressed in HCC patients after surgical operation, however following tumor relapse its expression levels were reversed (Yu F. et al., 2015).
In tissues, high expression of miR-150-5p and miR-29a-5p in combination of low expression of miR-101-3p, miR-126-3p, miR-127-3p, miR-139-5p, and miR-214-3p have tumor-suppressor roles and consequently have potential use as diagnostic biomarkers for HCC (Zhu et al., 2012; Han et al., 2014; Li T. et al., 2014; Peveling-Oberhag et al., 2014; Xie et al., 2014; Zhou et al., 2014; Wang S. et al., 2015). The association between the circulating miR-101-3p, miR-122-5p, miR-125b-5p, miR-139-5p, miR-150-5p, miR-16-5p, miR-181a-5p, miR-199a-3p, miR-199a-5p, miR-203a-3p, miR-21-5p, miR-22-3p, miR-29b-3p, miR-375, let-7b-5p, and tumor suppressor render them potential biomarkers for differentiating HCC from healthy controls (Zhou J. et al., 2011; Luo et al., 2013; Li T. et al., 2014; Tan et al., 2014; Xie et al., 2014; Chen et al., 2015; Jiang et al., 2015; Wang S. et al., 2015; Yin et al., 2015; Yu F. et al., 2015; Hung et al., 2016). Contrarily, miR-101-3p, miR-122-5p, miR-125b-5p, miR-130a-3p, miR-146a-5p, miR-214-3p, and miR-99a-5p were known as tumor suppressors in HCC and played the role of prognostic indicators for HCC (Zhang et al., 2012; Wang et al., 2013; Li B. et al., 2014; Rong et al., 2014; Tsang et al., 2014; Xie et al., 2014; Xu Q. et al., 2015). Serum miR-1-3p, miR-101-3p, miR-122-5p, miR-150-5p, miR-203a-3p, and miR-30c-5p were linked to tumor suppression, and new independent parameters of overall survival in HCC (Köberle et al., 2013; Xie et al., 2014; Cho et al., 2015; Liu D. et al., 2015; Xu Y. et al., 2015; Yu F. et al., 2015).
As miRNAs expression levels can be used as biomarkers for HCC diagnosis and prognosis, miRNA specific methylation patterns are of importance for therapeutic applications as well. Acting as a tumor suppressor miRNA, decreased expression of miR-10a due to hypermethylation can be used as a biomarker for early HCC diagnosis and risk assessment (Shen et al., 2012). Furthermore, some miRNAs methylation patterns can be HCC cell-specific and therefore used as diagnostic biomarkers. Such miRNAs cell-specific diagnostic methylation patterns include the hypermethylation of miR-129-2, miR-34a, and miR-148a (Anwar et al., 2013; Lu et al., 2013). Also, hypermethylation of mir-9-1 has been shown to be a biomarker for poor diagnosis and aggressiveness (Anwar et al., 2013). In addition to their implications in diagnosis and prognosis, miRNA specific aberrant methylation patterns can be used for therapeutic applications. For example, administration of miR-124, which is hypermethylated in HCC, stopped HCC progression in animal models and was considered safe. Moreover, sorafenib (anti-cancer drug) increased the expression of miR-1274, which is hypermethylated in HCC, leading to an increased response to therapy (Zhou C. et al., 2011).
miRNAs and CSCs in HCC
miRNAs play essential roles in regulating CSCs (Garg, 2015), and in regulating apoptosis in CSCs by acting on mRNAs of apoptosis proteins or regulating mRNAs that are downstream targets in specific apoptotic pathways. These control mechanisms aid in the regulation of metastasis, drug resistance, tumor invasion, pluripotency, and self-renewal potential.
The tumorigenicity of liver CSCs was found to be significantly suppressed by inhibition of miR-181. This miRNA regulates the differentiation potential of liver CSCs through activating transcription factors, including caudal homeobox gene 2 (CDX2) and GATA6, and negatively regulating the Wnt/β-catenin pathway via nemo-like kinase (NLK) (Ji et al., 2009b; Leal and Lleonart, 2013; Bessède et al., 2014). MiR-Let-7 and miR-Lin28 have been reported to be related to the rate of growth and metastasis of HCC. Lin28 is highly expressed in normal embryonic stem cells (ESCs). It maintains the self-renewal of liver CSCs by inhibiting the interaction of Let-7 with the mature miRNA. Let-7 degradation, which is caused by excessively active Lin28 and c-MYC, dis-equilibrates liver CSCs, leading to accelerated growth and metastasis of HCC (Heo et al., 2008). MiRNAs positively regulate liver CSCs via high expression of EpCAM, which is a prominent marker of liver CSCs. This upregulation is mediated by inhibition of TGF-β by downstream transcription factors of miR-18, such as CDX2, GATA6, and NLK. The EpCAM intracellular domain (EpICD) enters the nucleus and induces overexpression of cyclin D1, c-MYC, and miR-181 after binding to LIM domain protein 2 (FHL2), β-catenin and lymphoid enhancer factor 1 (Lef-1) (Ji et al., 2009b). Another group showed that TGF-β1 downregulate TP53INP1 by targeting miR-155 and promote epithelial-mesenchymal transition (EMT) and liver CSC phenotypes (Liu F. et al., 2015). The Wnt/β-catenin signaling pathway that regulates tumor heterogeneity is mainly related to miRNA, but the mechanism by which this balance between liver CSCs and cancer cells is maintained has not been elucidated.
Based on previous studies, some miRNAs expression was reported to be dysregulated in both HCC and CSCs. In Table 1, we compiled the mutually dysregulated miRNAs to establish the links between these miRNAs and the initiation and progression of HCC.

Table 1. miRNAs whose expression was reported to be dysregulated in HCC and CSCs.
We classified the previously reported miRNAs as mutually upregulated, mutually downregulated, and mutually dysregulated but not mutually upregulated or mutually downregulated, as illustrated in Figure 1.
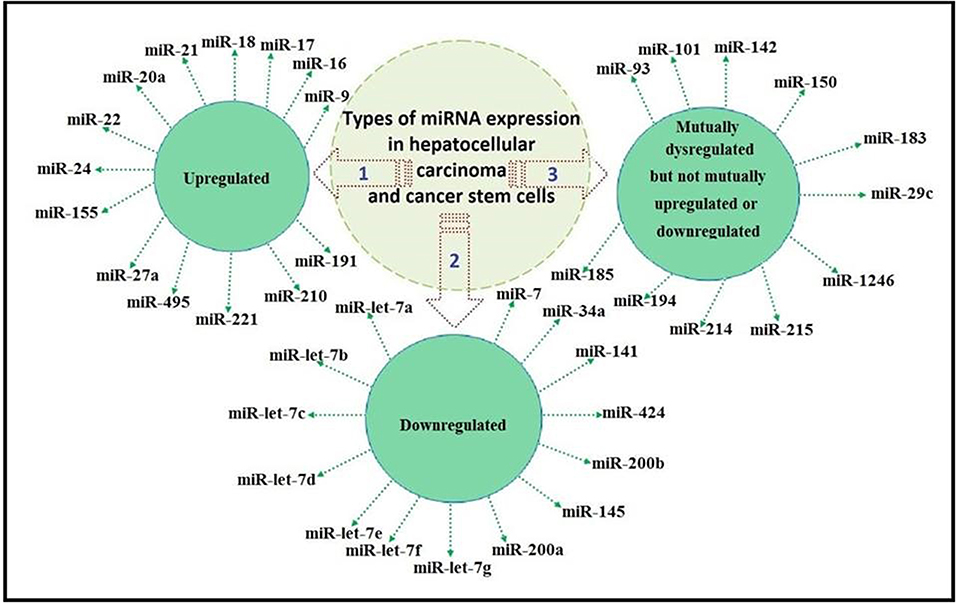
Figure 1. Classification of miRNA expression as mutually upregulated, mutually downregulated, and mutually dysregulated but not mutually upregulated or downregulated.
A schematic presentation of the role of dysregulated miRNAs in HCC initiation, progression, and aggressiveness is presented in Figure 2.
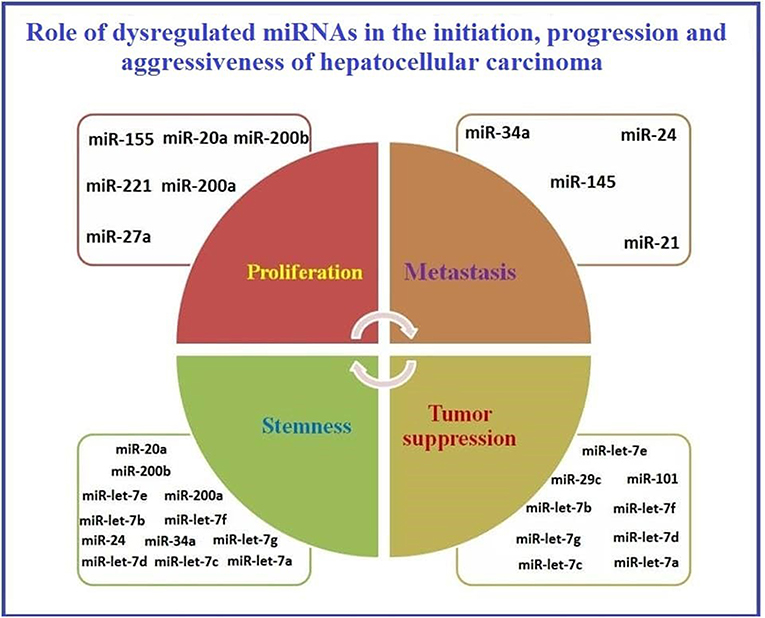
Figure 2. Representation of the role of dysregulated miRNAs in initiation, progression, and aggressiveness of Hepatocellular carcinoma.
As reported previously, MSCs aid in cancer development by enhancing the metastatic capability of tumor cells (Hill et al., 2017). This has been also reasoned by the fact that MSCs can home to tumor microenvironment mainly due to the action of stromal cell-derived factor 1 (SDF-1) (Gao et al., 2009). After homing, MSCs start to trans-differentiate into cancer associated fibroblasts mainly due to the action of TGF-beta1 (Ghaderi and Abtahi, 2018). Afterwards, cancer associated fibroblasts (CAFs) start to induce metastasis in neighboring tumor cells by inducing EMT (Wang et al., 2018). Some of the significant genes involved in such pro-metastatic signature of the tumor MSCs have been identified in lung cancer and they include GREM1, LOXL2, ADAMTS12, and ITGA11 (Fregni et al., 2018). Also, as investigated by our research group, soluble factors secreted from cancer cells when cocultured with MSCs have shown to induce cancer stem cell-like characteristics in the cocultured MSCs (El-Badawy et al., 2017). Relating the previous information, we are trying to highlight the mutual dysregulated miRNA in HCC, CSCs, and MSCs to investigate whether miRNAs play a vital role in the acquirement of MSCs to pro-metastatic characteristics or development into Cancer stem cells- like cells or even CAFs. So, in Table 2, we summarize the roles of miRNAs that are mutually dysregulated in HCC, CSC, and in MSCs. The functions of these miRNAs may provide insight into their regulatory roles in the development of cancer (Schraivogel et al., 2011). Based on these proposed functions (Table 2), we classified these miRNAs according to their roles in MSC differentiation.
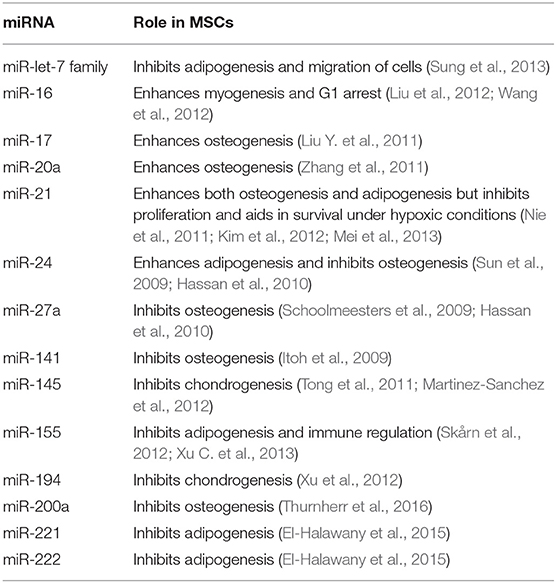
Table 2. The roles of the mutually dysregulated miRNAs in HCC and CSC, in MSC differentiation.
Figure 3 shows the potential pathways that may be involved in the reprogramming of MSCs and their acquisition of CSC-like characteristics after co-culture with cancer cells.
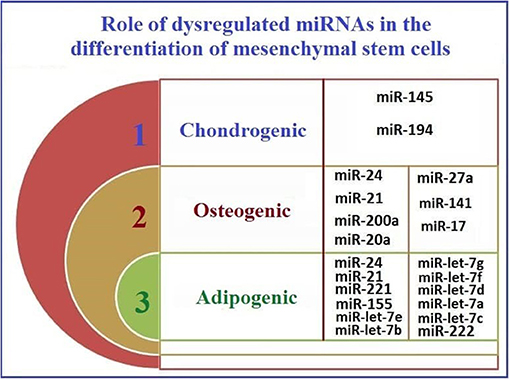
Figure 3. The roles of mutually dysregulated miRNAs in HCC and CSC, in MSC differentiation.
The relationship between the expression of miRNAs expression, their target genes' expression, and the fate of HCC is detailed in Table 3.
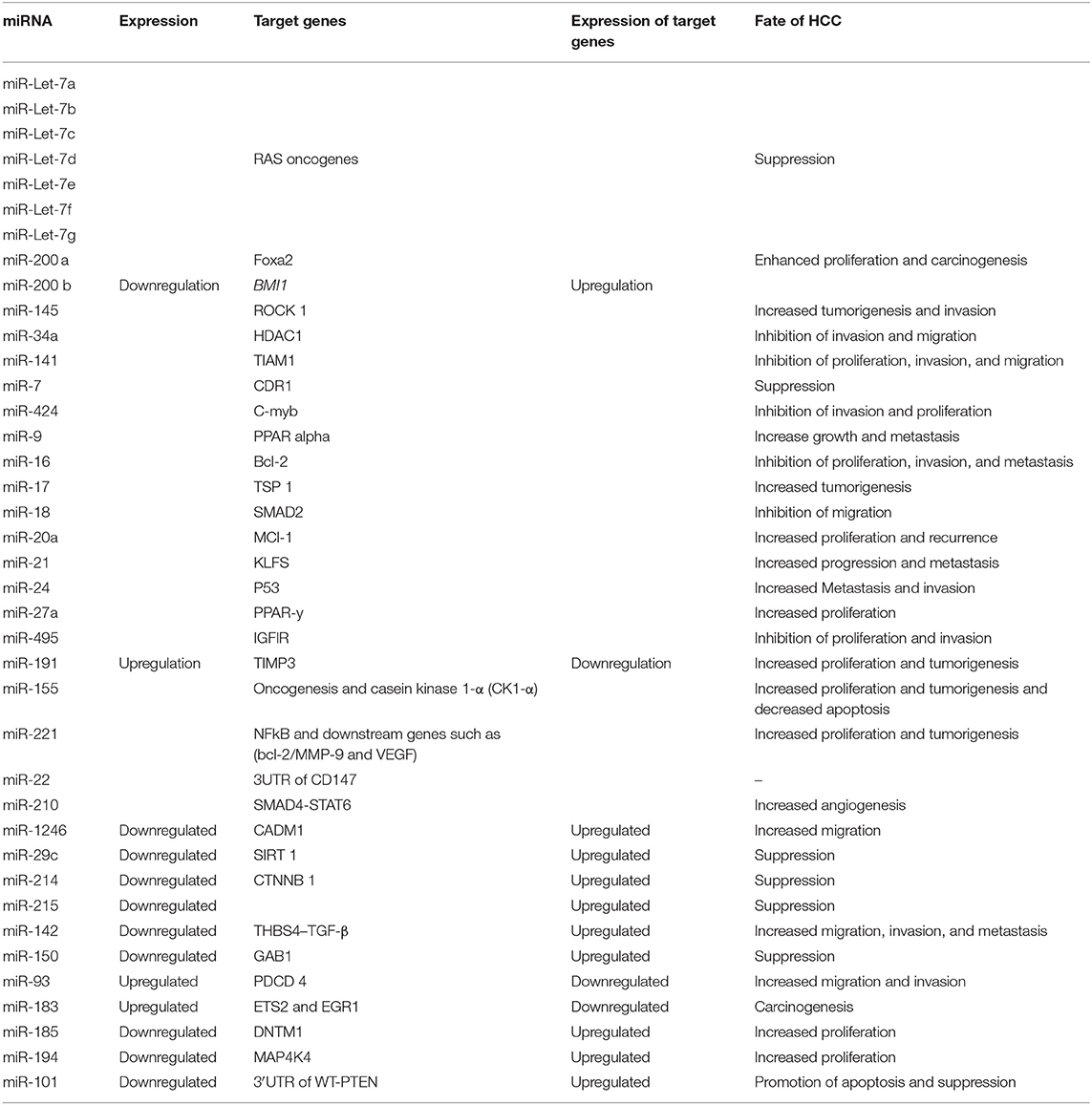
Table 3. The relationship among miRNAs' expression, target genes' expression, and the fate of HCC.
Dysregulated miRNA Methylation in HCC
Genome-wide abnormal DNA methylation of miRNA host genes in HCC was recently reported. One study analyzed tumor and neighboring normal non-tumor tissues in 62 HCC patients. This analysis was performed using Infinium Human Methylation Analysis Bead Chips. One hundred ten miRNAs from 64 different host genes were covered in this analysis through assessing the methylation of 254 CpG sites. Methylation levels were found to be significantly different at 54 CpG sites from 27 host genes between tumor and neighboring normal non-tumor tissues (Shen et al., 2012). In addition, the expression of three identified miRNAs were measured. MiR-10a was downregulated in tumor tissues and therefore its action on its oncogenic target genes as a tumor suppressor miRNA diminished. This decline appeared to be related to hypermethylation of the host genes. Accordingly, aberrant methylation and expression of miRNAs were considered valuable molecular biomarkers for HCC early diagnosis (Shen et al., 2012). In another study, miRNA genes, from HCC cells and normal liver hepatocytes, showed significantly different profiles of global DNA methylation. In the same study, in HCC cells, miRNAs CpG-poor regions were more commonly hypomethylated rather than being hypermethylation (He et al., 2015). Investigations using miRNA expression microarray data identified 10 dysregulated miRNAs in HCC that are regulated by DNA methylation. Of the 10 studied miRNAs, miR-23a/27a and miR-25/93/106b constituted two miRNA clusters in which five miRNAs were upregulated. On the other hand, the other five miRNAs including miR-375, miR-195, miR-497, miR-378, and miR-148a were downregulated (He et al., 2015). The cluster containing miR-25/93/106b, with upregulated expression, was required for cell proliferation including the anchorage-independent growth. It was also shown to target the E2F1 transcription factor in HCC, which inhibits apoptosis (Li Y. et al., 2009). Additionally, miR-331-3p was shown to target PHLPP, a protein that plays a central role in inducing apoptosis and reducing metastasis (Ma et al., 2010; Liu J. et al., 2011; Chang et al., 2014; Peuget et al., 2014). These data provide further evidence for the potential role of miR-331-3p in HCC metastasis. The miR-23a/27a cluster, with upregulated expression, enhanced anti-apoptotic pathways in addition to promoting cells proliferation in HCC (Huang et al., 2008). In another study, miR-429 functioned by manipulating liver tumor-initiating cells to target the RBBP4/E2F1/OCT4 axis and was upregulated in HCC due to four aberrant hypomethylated upstream sites (Li L. et al., 2015).
AEG-1 and ATG7 were found to be targets for miR-375, one of the previously mentioned epigenetically downregulated miRNAs, which makes miR-375 a tumor suppressor miRNA in HCC (Chang et al., 2012; He et al., 2012). When overexpressed, miR-375 inhibited both migration and invasion in HCC (He et al., 2012). Cell growth was also inhibited by the action of the miR-195/497, which targeted vital cell cycle regulators, leading to G1 arrest in HCC (Furuta et al., 2013). As reported using a bioinformatics approach, CDK4, which is involved in chemotherapy-mediated tumor cell apoptosis, was predicted to be a potential miR-195 target (Yang et al., 2011). MiR-378 suppressed HCC tumor growth, which was originally caused by HBV infection. MiR-378 was found to directly target the insulin-like growth factor 1 receptor (IGF1R) (Li et al., 2013). IGF2BP1, highly involved in liver cancer progression by promoting metastasis, was reported as an expected miR-378 target (Gutschner et al., 2014). Acting as a tumor suppressor miRNA by targeting c-Met which is an oncogene, miR-148 has also been shown to target DNMT1 in hepatocytes leading to the induction of liver-specific phenotype (Gailhouste et al., 2013). MiR-148a is among the five epigenetically downregulated miRNA by methylation and since DNMT1 and DNMT3B are considered to be two of its targets and responsible for its methylation, miR-148a together with DNMT1 and DNMT3B constitute a positive feedback mechanism which in the case of HCC leads to miR-148a downregulation (Duursma et al., 2008; Pan et al., 2010).
Due to several reports of miRNA hypomethylation in HCC, DNA hypomethylation was shown to play a significant role in miRNA regulation. Moreover, DNA methylation might be responsible for the abnormal expression of these miRNAs. Accordingly, further studies on these dysregulated miRNAs are needed (He et al., 2015).
In HCC, miRNAs have been shown to be regulated through epigenetic markers, specifically by DNA methylation. In Table 4, we review reported methylation-regulated miRNAs in HCC. One of the few studies in this area reported that methylation of miR-203 in CSCs plays a role in EMT and increases cancer progression (Taube et al., 2013). A summary of the available data concerning the epigenetic control of miRNA expression in HCC via methylation is provided in Table 4. Also, Table 4 shows the role the methylation of these miRNAs plays in the progression of HCC.

Table 4. miRNAs regulated by methylation in HCC.
Data in Table 4 shows some consistent pattern between the role of different miRNAs and their methylation patterns. To illustrate, miRNAs that act as tumor suppressors are hypermethylated while oncomiR are hypomethylated, which finally leads to HCC progression. On the other hand, in HCC, some miRNAs showed an opposite pattern. MiR-10a, miR-10b, miR-9, and miR-196b have been shown to be hypermethylated and their hypermethylation state would lead to reduced tumorigenesis. Therefore, further studies are needed to investigate other roles for these specific miRNAs, and especially the function of miR-596 in HCC progression.
Conclusion
Several studies showed how tumor microenvironment enhances cancer development and progression (Whiteside, 2008; Wang et al., 2017; Klymenko and Nephew, 2018). Studies from our laboratory have shown that soluble HCC factors play a vital role in the induction of chemoresistance properties in human bone marrow (hBM)-MSCs and trigger their transformation into CSC-like cells (El-Badawy et al., 2017). However, the mechanism of this transformation remains unclear. Although the initiation of HCC is known to be preceded by cirrhosis, the initiation mechanism itself is thought to involve CSCs. CSCs were proposed to be responsible for chemoresistance and relapse in most cancers. Previous data reported by our research group highlighted the role of CSCs in HCC initiation and progression (El-Badawy et al., 2017). Studies also demonstrated that liver CSCs are associated with liver cancer metastasis and relapse and that CSCs play a substantial role in the resistance of liver cancer to conventional treatment (Xu et al., 2009). These data indicate that targeting CSCs as a potential therapeutic approach for liver cancer holds huge promise for improving the treatment outcomes (Lou et al., 2018). Since reprogramming events responsible for the transformation and initiation processes are mainly controlled by epigenetic modifications, determining the role of such epigenetic fingerprints, including miRNAs and their methylation, in initiation, relapse, and chemoresistance in HCC and CSCs is central to understanding tumor biology and developing effective therapies.
Herein, we are presenting growing evidence supporting the central role of miRNAs in many biological processes (Brennecke et al., 2003; Ambros, 2004). In addition, miRNA dysfunction causes the development of diverse cancers (Iorio and Croce, 2009; Negrini et al., 2009). Recent studies show that HCC is associated with altered levels of miRNAs (Murakami et al., 2006; Jiang et al., 2008; Wong et al., 2008). Moreover, several miRNAs, such as miR-195, miR-122, miR-101, and miR-121, have been reported to regulate cell invasion, migration, apoptosis and growth (Fornari et al., 2008; Wang et al., 2008; Su et al., 2009). These findings suggest that dysregulation of miRNA may be attributed to hepatocarcinogenesis (Xiong et al., 2010).
Accumulating data on the expression profiles, roles, and regulation of miRNAs are essential for designing effective stem cell therapies for HCC. In this review, we highlighted the dysregulated expression of relevant miRNAs in HCC and CSCs. Based on the classification of miRNAs as mutually upregulated, mutually downregulated, or mutually dysregulated, we proposed their roles in cancer progression. Despite the lack of data on miRNA expression in MSCs, four miRNAs have dysregulated expression in HCC, CSCs, and MSCs. Based on their functions in MSCs, these miRNAs have been shown to mainly affect differentiation. It is clear, however, that more research is required on the expression profiles of miRNAs in MSCs under physiological conditions and from different tissue sources. Such studies are essential for determining how MSCs regulate and interact with cancer cells and CSCs in HCC.
In the context of clinical applications, miRNAs could represent an opportunity to develop safe strategies for achieving early diagnosis, monitoring disease status, and improving the effectiveness of non-invasive HCC treatment (Valeri et al., 2009). Several studies have shown the effect of miRNAs on enhancing the sensitivity of liver CSCs to treatment. Many dysregulated miRNAs in liver CSCs exert their roles by binding to specific target genes that are key molecules in many pathways. Targeting these miRNAs, their targeted genes, or respective pathways may thus effectively target CSCs, and disturb their role in metastasis, recurrence, and resistance to therapy (Lou et al., 2018). Together with conventional treatment, targeting specific miRNAs involved in tumor progression in combination therapy represent an attractive approach to multifactorial effective therapy to liver cancer (Tao et al., 2018). Although no miRNAs-based drugs are available in current clinical practice (Lou et al., 2018), the antitumor efficiency of modern anticancer drugs like sorafenib on HCC was significantly increased in vivo upon delivery of miR-122-exosome to the tumor (Blechacz and Gores, 2008; Lou et al., 2015). Such enhancement is promising to patients of unresectable HCC whose treatment with sorafenib was of limited efficacy, by prolonging survival for only 3 months (Blechacz and Gores, 2008; Kane et al., 2009). Investigating the expression signature of those candidate miRNAs in HCC diagnosis, prognosis, metastasis, and recurrence and determining how these miRNAs genetically and epigenetically regulate the transformation of somatic stem cells to a more chemoresistant phenotype is needed for translation to clinical practice (Valeri et al., 2009; El-Badawy et al., 2017).
Author Contributions
Each author has substantially contributed to conducting this study and drafting this manuscript. MN and RS wrote the manuscript. MN, ME, and SE analyzed data and moderate figures and tables. While, NE-B contributed in writing and editing of the manuscript.
Funding
This study was funded by the Egyptian Science and Technology Development Fund (STDF Grant ID: 5300).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Ali Hosseini Rad, S. M., Bavarsad, M. S., Arefian, E., Jaseb, K., Shahjahani, M., and Saki, N. (2013). The role of microRNAs in stemness of cancer stem cells. Oncol. Rev. 7, e8. doi: 10.4081/oncol.2013.e8
Alpini, G., Glaser, S. S., Zhang, J. P., Francis, H., Han, Y., Gong, J., et al. (2011). Regulation of placenta growth factor by microRNA-125b in hepatocellular cancer. J. Hepatol. 55, 1339–1345. doi: 10.1016/j.jhep.2011.04.015
Amr, K. S., Ezzat, W. M., Elhosary, Y. A., Hegazy, A. E., Fahim, H. H., and Kamel, R. R. (2016). The potential role of miRNAs 21 and 199-a in early diagnosis of hepatocellular carcinoma. Gene 575, 66–70. doi: 10.1016/j.gene.2015.08.038
Anwar, S. L., Albat, C., Krech, T., Hasemeier, B., Schipper, E., Schweitzer, N., et al. (2013). Concordant hypermethylation of intergenic microRNA genes in human hepatocellular carcinoma as new diagnostic and prognostic marker. Int. J. Cancer. 133, 660–670. doi: 10.1002/ijc.28068
Ashmawy, A. M., Elgeshy, K. M., Abdel Salam, E. T., Ghareeb, M., Kobaisi, M. H., Amin, H. A. A., et al. (2017). Crosstalk between liver-related microRNAs and Wnt/β-catenin pathway in hepatocellular carcinoma patients. Arab J. Gastroenterol. 18, 144–150. doi: 10.1016/j.ajg.2017.09.001
Assal, R. A., El Tayebi, H. M., Hosny, K. A., Esmat, G., and Abdelaziz, A. I. (2015). A pleiotropic effect of the single clustered hepatic metastamiRs miR-96-5p and miR-182-5p on insulin-like growth factor II, insulin-like growth factor-1 receptor and insulin-like growth factor-binding protein-3 in hepatocellular carcinoma. Mol. Med. Rep. 12, 645–650. doi: 10.3892/mmr.2015.3382
Bao, B., Ali, S., Ahmad, A., Azmi, A. S., Li, Y., Banerjee, S., et al. (2012). Hypoxia-induced aggressiveness of pancreatic cancer cells is due to increased expression of VEGF, IL-6, and miR-21, which can be attenuated by CDF treatment. PLoS ONE 7:e50165. doi: 10.1371/journal.pone.0050165
Bao, L., Zhao, J., Dai, X., Wang, Y., Ma, R., Su, Y., et al. (2014). Correlation between miR-23a and onset of hepatocellular carcinoma. Clin. Res. Hepatol. Gastroenterol. 38, 318–330. doi: 10.1016/j.clinre.2013.12.002
Bartel, D. P. (2004). MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116, 281–297. doi: 10.1016/S0092-8674(04)00045-5
Baskerville, S., and Bartel, D. P. (2005). Microarray profiling of microRNAs reveals frequent coexpression with neighboring miRNAs and host genes. RNA 11, 241–247. doi: 10.1261/rna.7240905
Beck, B., and Blanpain, C. (2013). Unravelling cancer stem cell potential. Nat. Rev. Cancer 13, 727–738. doi: 10.1038/nrc3597
Bedard, P. L., Hansen, A. R., Ratain, M. J., and Siu, L. L. (2013). Tumour heterogeneity in the clinic. Nature 501, 355–364. doi: 10.1038/nature12627
Bessède, E., Staedel, C., Acuña Amador, L. A., Nguyen, P. H., Chambonnier, L., Hatakeyama, M., et al. (2014). Helicobacter pylori generates cells with cancer stem cell properties via epithelial-mesenchymal transition-like changes. Oncogene 33, 4123–4131. doi: 10.1038/onc.2013.380
Blechacz, B., and Gores, G. J. (2008). Cholangiocarcinoma: advances in pathogenesis, diagnosis, and treatment. Hepatology 48, 308–321. doi: 10.1002/hep.22310
Borel, F., Konstantinova, P., and Jansen, P. L. (2012). Diagnostic and therapeutic potential of miRNA signatures in patients with hepatocellular carcinoma. J. Hepatol. 56, 1371–1383. doi: 10.1016/j.jhep.2011.11.026
Brennecke, J., Hipfner, D. R., Stark, A., Russell, R. B., and Cohen, S. M. (2003). Bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell 113, 25–36. doi: 10.1016/S0092-8674(03)00231-9
Budhu, A., Jia, H. L., Forgues, M., Liu, C. G., Goldstein, D., Lam, A., et al. (2008). Identification of metastasis-related microRNAs in hepatocellular carcinoma. Hepatology 47, 897–907. doi: 10.1002/hep.22160
Bushati, N., and Cohen, S. M. (2007). microRNA functions. Annu. Rev. Cell Dev. Biol. 23, 175–205. doi: 10.1146/annurev.cellbio.23.090506.123406
Cabrera, R., and Nelson, D. R. (2010). Review article: the management of hepatocellular carcinoma. Aliment. Pharmacol. Ther. 31, 461–476. doi: 10.1111/j.1365-2036.2009.04200.x
Calin, G. A., and Croce, C. M. (2006). MicroRNA signatures in human cancers. Nat. Rev. Cancer 6, 857–866. doi: 10.1038/nrc1997
Callegari, E., Elamin, B. K., Sabbioni, S., Gramantieri, L., and Negrini, M. (2013). Role of microRNAs in hepatocellular carcinoma: a clinical perspective. Onco. Targets. Ther. 6, 1167–1178. doi: 10.2147/OTT.S36161
Cao, J., Qiu, J., Wang, X., Lu, Z., Wang, D., Feng, H., et al. (2018). Identification of microRNA-124 in regulation of Hepatocellular carcinoma through BIRC3 and the NF-κB pathway. J. Cancer. 9, 3006–3015. doi: 10.7150/jca.25956
Caruso, S., Bazan, V., Rolfo, C., Insalaco, L., Fanale, D., Bronte, G., et al. (2012). MicroRNAs in colorectal cancer stem cells: new regulators of cancer stemness? Oncogenesis 1:e32. doi: 10.1038/oncsis.2012.33
Chang, R. M., Yang, H., Fang, F., Xu, J. F., and Yang, L. Y. (2014). MicroRNA-331-3p promotes proliferation and metastasis of hepatocellular carcinoma by targeting PH domain and leucine-rich repeat protein phosphatase. Hepatology 60, 1251–1263. doi: 10.1002/hep.27221
Chang, Y., Yan, W., He, X., Zhang, L., Li, C., Huang, H., et al. (2012). miR-375 inhibits autophagy and reduces viability of hepatocellular carcinoma cells under hypoxic conditions. Gastroenterology 143, 177–87 e8. doi: 10.1053/j.gastro.2012.04.009
Chapnik, E., Rivkin, N., Mildner, A., Beck, G., Pasvolsky, R., Metzl-Raz, E., et al. (2014). miR-142 orchestrates a network of actin cytoskeleton regulators during megakaryopoiesis. Elife 3:e01964. doi: 10.7554/eLife.01964
Chen, K., Huang, Y. H., and Chen, J. L. (2013). Understanding and targeting cancer stem cells: therapeutic implications and challenges. Acta Pharmacol. Sin. 34:732–740. doi: 10.1038/aps.2013.27
Chen, L., Chu, F., Cao, Y., Shao, J., and Wang, F. (2015). Serum miR-182 and miR-331-3p as diagnostic and prognostic markers in patients with hepatocellular carcinoma. Tumor Biol. 36, 7439–7447. doi: 10.1007/s13277-015-3430-2
Chen, S. Y., Ma, D. N., Chen, Q. D., Zhang, J. J., Tian, Y. R., Wang, Z. C., et al. (2017). MicroRNA-200a inhibits cell growth and metastasis by targeting Foxa2 in hepatocellular carcinoma. J. Cancer 8:617. doi: 10.7150/jca.17394
Cho, H. J., Kim, J. K., Nam, J. S., Wang, H. J., Lee, J. H., Kim, B. W., et al. (2015). High circulating microRNA-122 expression is a poor prognostic marker in patients with hepatitis B virus-related hepatocellular carcinoma who undergo radiofrequency ablation. Clin. Biochem. 48, 1073–1078. doi: 10.1016/j.clinbiochem.2015.06.019
Cholankeril, G., Patel, R., Khurana, S., and Satapathy, S. K. (2017). Hepatocellular carcinoma in non-alcoholic steatohepatitis: current knowledge and implications for management. World J. Hepatol. 9:533–543. doi: 10.4254/wjh.v9.i11.533
Chu, Y., Fan, W., Guo, W., Zhang, Y., Wang, L., Guo, L., et al. (2017). miR-1247-5p functions as a tumor suppressor in human hepatocellular carcinoma by targeting Wnt3. Oncol. Rep. 38, 343–351. doi: 10.3892/or.2017.5702
Chuang, J. C., and Jones, P. A. (2007). Epigenetics and microRNAs. Pediatr. Res. 61(5 Pt 2), 24R−29R. doi: 10.1203/pdr.0b013e3180457684
Connolly, E., Melegari, M., Landgraf, P., Tchaikovskaya, T., Tennant, B. C., Slagle, B. L., et al. (2008). Elevated expression of the miR-17-92 polycistron and miR-21 in hepadnavirus-associated hepatocellular carcinoma contributes to the malignant phenotype. Am. J. Pathol. 173, 856–864. doi: 10.2353/ajpath.2008.080096
Cummins, J. M., and Velculescu, V. E. (2006). Implications of micro-RNA profiling for cancer diagnosis. Oncogene 25, 6220–6227. doi: 10.1038/sj.onc.1209914
Dambal, S., Shah, M., Mihelich, B., and Nonn, L. (2015). The microRNA-183 cluster: the family that plays together stays together. Nucleic Acids Res. 43, 7173–7188. doi: 10.1093/nar/gkv703
Datta, J., Kutay, H., Nasser, M. W., Nuovo, G. J., Wang, B., Majumder, S., et al. (2008). Methylation mediated silencing of MicroRNA-1 gene and its role in hepatocellular carcinogenesis. Cancer Res. 68, 5049–5058. doi: 10.1158/0008-5472.CAN-07-6655
Dhanasekaran, R., Limaye, A., and Cabrera, R. (2012). Hepatocellular carcinoma: current trends in worldwide epidemiology, risk factors, diagnosis, and therapeutics. Hepat. Med. 4:19–37. doi: 10.2147/HMER.S16316
Dhayat, S. A., Mardin, W. A., Köhler, G., Bahde, R., Vowinkel, T., Wolters, H., et al. (2014). The microRNA-200 family—a potential diagnostic marker in hepatocellular carcinoma? J. Surg. Oncol. 110, 430–438. doi: 10.1002/jso.23668
Duursma, A. M., Kedde, M., Schrier, M., le Sage, C., and Agami, R. (2008). miR-148 targets human DNMT3b protein coding region. RNA 14, 872–877. doi: 10.1261/rna.972008
Eaves, C. J., and Humphries, R. K. (2010). Acute myeloid leukemia and the WNT pathway. N. Engl. J. Med. 362, 2326–2327. doi: 10.1056/NEJMcibr1003522
El Tayebi, H. M., Hosny, K. A., Esmat, G., Breuhahn, K., and Abdelaziz, A. I. (2012). miR-615-5p is restrictedly expressed in cirrhotic and cancerous liver tissues and its overexpression alleviates the tumorigenic effects in hepatocellular carcinoma. FEBS Lett. 586, 3309–3316. doi: 10.1016/j.febslet.2012.06.054
El Tayebi, H. M., Waly, A. A., Assal, R. A., Hosny, K. A., Esmat, G., and Abdelaziz, A. I. (2015). Transcriptional activation of the IGF-II/IGF-1R axis and inhibition of IGFBP-3 by miR-155 in hepatocellular carcinoma. Oncol. Lett. 10, 3206–3212. doi: 10.3892/ol.2015.3725
El-Abd, N. E., Fawzy, N. A., El-Sheikh, S. M., and Soliman, M. E. (2015). Circulating miRNA-122, miRNA-199a, and miRNA-16 as biomarkers for early detection of hepatocellular carcinoma in Egyptian patients with chronic hepatitis C virus infection. Mol. Diagn. Ther. 19, 213–220. doi: 10.1007/s40291-015-0148-1
El-Badawy, A., Ghoneim, M. A., Gabr, M. M., Salah, R. A., Mohamed, I. K., Amer, M., et al. (2017). Cancer cell-soluble factors reprogram mesenchymal stromal cells to slow cycling, chemoresistant cells with a more stem-like state. Stem Cell Res. Ther. 8, 254. doi: 10.1186/s13287-017-0709-9
El-Halawany, M. S., Ismail, H. M., Zeeneldin, A. A., Elfiky, A., Tantawy, M., Kobaisi, M. H., et al. (2015). Investigating the pretreatment miRNA expression patterns of advanced hepatocellular carcinoma patients in association with response to TACE treatment. Biomed. Res. Int. 2015:649750. doi: 10.1155/2015/649750
El-Serag, H. B. (2002). Hepatocellular carcinoma and hepatitis C in the United States. Hepatology. 36(5 Suppl. 1), S74–83. doi: 10.1053/jhep.2002.36807
Eshelman, M. A., and Yochum, G. S. (2016). The Wnt/β-catenin pathway is activated by miR-1246 in liver cancer stem cells. Transl. Cancer Res. S 1457–S1460. doi: 10.21037/tcr.2016.12.57
Espada, J., Calvo, M. B., Díaz-Prado, S., and Medina, V. (2009). Wnt signalling and cancer stem cells. Clin. Transl. Oncol. 11, 411–427. doi: 10.1007/s12094-009-0380-4
Fawzy, I. O., Hamza, M. T., Hosny, K. A., Esmat, G., and Abdelaziz, A. I. (2016). Abrogating the interplay between IGF2BP1, 2 and 3 and IGF1R by let-7i arrests hepatocellular carcinoma growth. Growth Factors. 34, 42–50. doi: 10.3109/08977194.2016.1169532
Fornari, F., Gramantieri, L., Ferracin, M., Veronese, A., Sabbioni, S., Calin, G. A., et al. (2008). MiR-221 controls CDKN1C/p57 and CDKN1B/p27 expression in human hepatocellular carcinoma. Oncogene 27, 5651–5661. doi: 10.1038/onc.2008.178
Fregni, G., Quinodoz, M., Möller, E., Vuille, J., Galland, S., Fusco, C., et al. (2018). Reciprocal modulation of mesenchymal stem cells and tumor cells promotes lung cancer metastasis. EBioMed. 29, 128–145. doi: 10.1016/j.ebiom.2018.02.017
Furuta, M., Kozaki, K., Tanimoto, K., Tanaka, S., Arii, S., Shimamura, T., et al. (2013). The tumor-suppressive miR-497-195 cluster targets multiple cell-cycle regulators in hepatocellular carcinoma. PLoS ONE 8:e60155. doi: 10.1371/journal.pone.0060155
Furuta, M., Kozaki, K. I., Tanaka, S., Arii, S., Imoto, I., Inazawa, J., et al. (2010). miR-124 and miR-203 are epigenetically silenced tumor-suppressive microRNAs in hepatocellular carcinoma. Carcinogenesis 31, 766–776. doi: 10.1093/carcin/bgp250
Gailhouste, L., Gomez-Santos, L., Hagiwara, K., Hatada, I., Kitagawa, N., Kawaharada, K., et al. (2013). miR-148a plays a pivotal role in the liver by promoting the hepatospecific phenotype and suppressing the invasiveness of transformed cells. Hepatology 58, 1153–1165. doi: 10.1002/hep.26422
Gao, H., Priebe, W., Glod, J., and Banerjee, D. (2009). Activation of signal transducers and activators of transcription 3 and focal adhesion kinase by stromal cell-derived factor 1 is required for migration of human mesenchymal stem cells in response to tumor cell-conditioned medium. Stem Cells 27, 857–865. doi: 10.1002/stem.23
Garg, M. (2015). Emerging role of microRNAs in cancer stem cells: implications in cancer therapy. World J. Stem Cells 7, 1078–1089. doi: 10.4252/wjsc.v7.i8.1078
Garofalo, M., Di Leva, G., Romano, G., Nuovo, G., Suh, S. S., Ngankeu, A., et al. (2009). miR-221&222 regulate TRAIL resistance and enhance tumorigenicity through PTEN and TIMP3 downregulation. Cancer Cell. 16, 498–509. doi: 10.1016/j.ccr.2009.10.014
George, G. P., and Mittal, R. D. (2010). MicroRNAs: potential biomarkers in cancer. Indian J. Clin. Biochem. 25, 4–14. doi: 10.1007/s12291-010-0008-z
Ghaderi, A., and Abtahi, S. (2018). Mesenchymal stem cells: miraculous healers or dormant killers? Stem Cell Rev. Rep. 14, 722–733. doi: 10.1007/s12015-018-9824-y
Gramantieri, L., Ferracin, M., Fornari, F., Veronese, A., Sabbioni, S., Liu, C. G., et al. (2007). Cyclin G1 is a target of miR-122a, a microRNA frequently down-regulated in human hepatocellular carcinoma. Cancer Res. 67, 6092–6099. doi: 10.1158/0008-5472.CAN-06-4607
Gregory, P. A., Bert, A. G., Paterson, E. L., Barry, S. C., Tsykin, A., Farshid, G., et al. (2008). The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell Biol. 10, 593–601. doi: 10.1038/ncb1722
Griffiths-Jones, S., Grocock, R. J., van Dongen, S., Bateman, A., and Enright, A. J. (2006). miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 34, D140–D144. doi: 10.1093/nar/gkj112
Gutschner, T., Hämmerle, M., Pazaitis, N., Bley, N., Fiskin, E., Uckelmann, H., et al. (2014). Insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) is an important protumorigenic factor in hepatocellular carcinoma. Hepatology 59, 1900–1911. doi: 10.1002/hep.26997
Guzman, G., Brunt, E. M., Petrovic, L. M., Chejfec, G., Layden, T. J., Cotler, S. J., et al. (2008). Does nonalcoholic fatty liver disease predispose patients to hepatocellular carcinoma in the absence of cirrhosis? Arch. Pathol. Lab. Med. 132, 1761–1766. doi: 10.1043/1543-2165-132.11.1761
Habashy, D. A., El Tayebi, H. M., Fawzy, I. O., Hosny, K. A., Esmat, G., and Abdelaziz, A. I. (2016). Interplay between microRNA-17-5p, insulin-like growth factor-II through binding protein-3 in hepatocellular carcinoma. World J. Hepatol. 8, 976–984. doi: 10.4254/wjh.v8.i23.976
Han, K., Li, J., Zhao, H., Liang, P., Huang, X., Zheng, L., et al. (2014). Identification of the typical miRNAs and target genes in hepatocellular carcinoma. Mol. Med. Rep. 10, 229–235. doi: 10.3892/mmr.2014.2194
Han, Z. B., Chen, H. Y., Fan, J. W., Wu, J. Y., Peng, Z. H., and Wang, Z. W. (2013). Expression and survival prediction of microRNA-155 in hepatocellular carcinoma after liver transplantation. Zhonghua Yi Xue Za Zhi. 93, 884–887.
Hassan, M. Q., Gordon, J. A., Beloti, M. M., Croce, C. M., van Wijnen, A. J., Stein, J. L., et al. (2010). A network connecting Runx2, SATB2, and the miR-23a~27a~24-2 cluster regulates the osteoblast differentiation program. Proc. Natl. Acad. Sci. U.S.A. 107, 19879–19884. doi: 10.1073/pnas.1007698107
He, X. X., Chang, Y., Meng, F. Y., Wang, M. Y., Xie, Q. H., Tang, F., et al. (2012). MicroRNA-375 targets AEG-1 in hepatocellular carcinoma and suppresses liver cancer cell growth in vitro and in vivo. Oncogene 31, 3357–3369. doi: 10.1038/onc.2011.500
He, X. X., Kuang, S. Z., Liao, J. Z., Xu, C. R., Chang, Y., Wu, Y. L., et al. (2015). The regulation of microRNA expression by DNA methylation in hepatocellular carcinoma. Mol. Biosyst. 11, 532–539. doi: 10.1039/C4MB00563E
He, Y., Cui, Y., Wang, W., Gu, J., Guo, S., Ma, K., et al. (2011). Hypomethylation of the hsa-miR-191 locus causes high expression of hsa-mir-191 and promotes the epithelial-to-mesenchymal transition in hepatocellular carcinoma. Neoplasia 13, 841–853. doi: 10.1593/neo.11698
Heo, I., Joo, C., Cho, J., Ha, M., Han, J., and Kim, V. N. (2008). Lin28 mediates the terminal uridylation of let-7 precursor MicroRN. Mol. Cell 32, 276–284. doi: 10.1016/j.molcel.2008.09.014
Higashi, T., Hayashi, H., Ishimoto, T., Takeyama, H., Kaida, T., Arima, K., et al. (2015). miR-9-3p plays a tumour-suppressor role by targeting TAZ (WWTR1) in hepatocellular carcinoma cells. Br. J. Cancer 113, 252–258. doi: 10.1038/bjc.2015.170
Hill, B. S., Pelagalli, A., Passaro, N., and Zannetti, A. (2017). Tumor-educated mesenchymal stem cells promote pro-metastatic phenotype. Oncotarget 8, 73296–73311. doi: 10.18632/oncotarget.20265
Hu, C., Shen, S. Q., Cui, Z. H., Chen, Z. B., and Li, W. (2015). Effect of microRNA-1 on hepatocellular carcinoma tumor endothelial cells. World J. Gastroenterol. 21, 5884–5892. doi: 10.3748/wjg.v21.i19.5884
Huang, S., He, X., Ding, J., Liang, L., Zhao, Y., Zhang, Z., et al. (2008). Upregulation of miR-23a approximately 27a approximately 24 decreases transforming growth factor-beta-induced tumor-suppressive activities in human hepatocellular carcinoma cells. Int. J. Cancer 123, 972–978. doi: 10.1002/ijc.23580
Huang, W., Li, J., Guo, X., Zhao, Y., and Yuan, X. (2016). miR-663a inhibits hepatocellular carcinoma cell proliferation and invasion by targeting HMGA2. Biomed. Pharmacother. 81, 431–438. doi: 10.1016/j.biopha.2016.04.034
Huang, X. B., Li, J., Zheng, L., Zuo, G. H., Han, K. Q., Li, H. Y., et al. (2013). Bioinformatics analysis reveals potential candidate drugs for HCC. Pathol. Oncol. Res. 19, 251–258. doi: 10.1007/s12253-012-9576-y
Huang, X. H., Wang, Q., Chen, J. S., Fu, X. H., Chen, X. L., Chen, L. Z., et al. (2009). Bead-based microarray analysis of microRNA expression in hepatocellular carcinoma: miR-338 is downregulated. Hepatol. Res. 39, 786–794. doi: 10.1111/j.1872-034X.2009.00502.x
Hung, C. H., Hu, T. H., Lu, S. N., Kuo, F. Y., Chen, C. H., Wang, J. H., et al. (2016). Circulating micro RNAs as biomarkers for diagnosis of early hepatocellular carcinoma associated with hepatitis B virus. Int. J. Cancer 138, 714–720. doi: 10.1002/ijc.29802
Hwang-Verslues, W. W., Chang, P. H., Wei, P. C., Yang, C. Y., Huang, C. K., Kuo, W. H., et al. (2011). miR-495 is upregulated by E12/E47 in breast cancer stem cells, and promotes oncogenesis and hypoxia resistance via downregulation of E-cadherin and REDD1. Oncogene 30, 2463–2474. doi: 10.1038/onc.2010.618
Iorio, M. V., and Croce, C. M. (2009). MicroRNAs in cancer: small molecules with a huge impact. J. Clin. Oncol. 27, 5848–5856. doi: 10.1200/JCO.2009.24.0317
Ip, B., and Wang, X.-D. (2014). Non-alcoholic steatohepatitis and hepatocellular carcinoma: implications for lycopene intervention. Nutrients 6, 124–162. doi: 10.3390/nu6010124
Itoh, T., Nozawa, Y., and Akao, Y. (2009). MicroRNA-141 and−200a are involved in bone morphogenetic protein-2-induced mouse pre-osteoblast differentiation by targeting distal-less homeobox 5. J. Biol. Chem. 284, 19272–19279. doi: 10.1074/jbc.M109.014001
Ji, C., Liu, H., Yin, Q., Li, H., and Gao, H. (2017). miR-93 enhances hepatocellular carcinoma invasion and metastasis by EMT via targeting PDCD4. Biotechnol. Lett. 39, 1621–1629. doi: 10.1007/s10529-017-2403-5
Ji, J., Shi, J., Budhu, A., Yu, Z., Forgues, M., Roessler, S., et al. (2009a). MicroRNA expression, survival, and response to interferon in liver cancer. N. Engl. J. Med. 361, 1437–1447. doi: 10.1056/NEJMoa0901282
Ji, J., Yamashita, T., Budhu, A., Forgues, M., Jia, H. L., Li, C., et al. (2009b). Identification of microRNA-181 by genome-wide screening as a critical player in EpCAM-positive hepatic cancer stem cells. Hepatology 50, 472–480. doi: 10.1002/hep.22989
Jiang, J., Gusev, Y., Aderca, I., Mettler, T. A., Nagorney, D. M., Brackett, D. J., et al. (2008). Association of MicroRNA expression in hepatocellular carcinomas with hepatitis infection, cirrhosis, and patient survival. Clin. Cancer Res. 14, 419–427. doi: 10.1158/1078-0432.CCR-07-0523
Jiang, L., Cheng, Q., Zhang, B. H., and Zhang, M. Z. (2015). Circulating microRNAs as biomarkers in hepatocellular carcinoma screening: a validation set from China. Medicine. 94:e13434. doi: 10.1097/MD.0000000000000603
Johnson, S. M., Grosshans, H., Shingara, J., Byrom, M., Jarvis, R., Cheng, A., et al. (2005). RAS is regulated by the let-7 microRNA family. Cell 120, 635–647. doi: 10.1016/j.cell.2005.01.014
Kane, R. C., Farrell, A. T., Madabushi, R., Booth, B., Chattopadhyay, S., Sridhara, R., et al. (2009). Sorafenib for the treatment of unresectable hepatocellular carcinoma. Oncologist 14, 95–100. doi: 10.1634/theoncologist.2008-0185
Kawai, T., Yasuchika, K., Ishii, T., Katayama, H., Yoshitoshi, E. Y., Ogiso, S., et al. (2015). Keratin 19, a cancer stem cell marker in human hepatocellular carcinoma. Clin. Cancer Res. 21, 3081–3091. doi: 10.1158/1078-0432.CCR-14-1936
Kim, Y. J., Hwang, S. H., Cho, H. H., Shin, K. K., Bae, Y. C., Jung, J. S., et al. (2012). MicroRNA 21 regulates the proliferation of human adipose tissue-derived mesenchymal stem cells and high-fat diet-induced obesity alters microRNA 21 expression in white adipose tissues. J. Cell. Physiol. 227, 183–193. doi: 10.1002/jcp.22716
Klein, C. A. (2013). Selection and adaptation during metastatic cancer progression. Nature 501, 365–372. doi: 10.1038/nature12628
Klymenko, Y., and Nephew, K. (2018). Epigenetic crosstalk between the tumor microenvironment and ovarian cancer cells: a therapeutic road less traveled. Cancers 10:E295. doi: 10.3390/cancers10090295
Köberle, V., Kronenberger, B., Pleli, T., Trojan, J., Imelmann, E., Peveling-Oberhag, J., et al. (2013). Serum microRNA-1 and microRNA-122 are prognostic markers in patients with hepatocellular carcinoma. Eur. J. Cancer. 49, 3442–3449. doi: 10.1016/j.ejca.2013.06.002
Krol, J., Loedige, I., and Filipowicz, W. (2010). The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 11, 597–610. doi: 10.1038/nrg2843
Lashine, Y. A., Seoudi, A. M., Salah, S., and Abdelaziz, A. I. (2011). Expression signature of microRNA-181-a reveals its crucial role in the pathogenesis of paediatric systemic lupus erythematosus. Clin. Exp. Rheumatol. 29, 351–357.
Lau, W. Y., and Lai, E. C. (2008). Hepatocellular carcinoma: current management and recent advances. HBPD INT 7, 237–257.
Law, P. T., Ching, A. K., Chan, A. W., Wong, Q. W., Wong, C. K., To, K. F., et al. (2012). MiR-145 modulates multiple components of the insulin-like growth factor pathway in hepatocellular carcinoma. Carcinogenesis 33, 1134–1141. doi: 10.1093/carcin/bgs130
Leal, J. A., and Lleonart, M. E. (2013). MicroRNAs and cancer stem cells: therapeutic approaches and future perspectives. Cancer Lett. 338, 174–183. doi: 10.1016/j.canlet.2012.04.020
Leung, W. K., He, M., Chan, A. W., Law, P. T., and Wong, N. (2015). Wnt/beta-catenin activates MiR-183/96/182 expression in hepatocellular carcinoma that promotes cell invasion. Cancer Lett. 362, 97–105. doi: 10.1016/j.canlet.2015.03.023
Lewis, B. P., Burge, C. B., and Bartel, D. P. (2005). Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 120, 15–20. doi: 10.1016/j.cell.2004.12.035
Li, B., Huang, P., Qiu, J., Liao, Y., Hong, J., and Yuan, Y. (2014). MicroRNA-130a is down-regulated in hepatocellular carcinoma and associates with poor prognosis. Med. Oncol. 31:230. doi: 10.1007/s12032-014-0230-2
Li, B., Lu, Y., Yu, L., Han, X., Wang, H., Mao, J., et al. (2017). miR-221/222 promote cancer stem-like cell properties and tumor growth of breast cancer via targeting PTEN and sustained Akt/NF-κB/COX-2 activation. Chem. Biol. Interact. 277, 33–42. doi: 10.1016/j.cbi.2017.08.014
Li, J., Fang, L., Yu, W., and Wang, Y. (2015). MicroRNA-125b suppresses the migration and invasion of hepatocellular carcinoma cells by targeting transcriptional coactivator with PDZ-binding motif. Oncol. Lett. 9, 1971–1975. doi: 10.3892/ol.2015.2973
Li, J., Fu, H., Xu, C., Tie, Y., Xing, R., Zhu, J., et al. (2010). miR-183 inhibits TGF-β1-induced apoptosis by downregulation of PDCD4 expression in human hepatocellular carcinoma cells. BMC Cancer. 10:354. doi: 10.1186/1471-2407-10-354
Li, L., Tang, J., Zhang, B., Yang, W., LiuGao, M., Wang, R., et al. (2015). Epigenetic modification of MiR-429 promotes liver tumour-initiating cell properties by targeting Rb binding protein 4. Gut 64, 156–167. doi: 10.1136/gutjnl-2013-305715
Li, L. H., Gao, Q., Wang, X. Y., and Guo, Z. J. (2013). [miR-378 suppresses HBV-related hepatocellular carcinoma tumor growth by directly targeting the insulin-like growth factor 1 receptor]. Zhonghua Gan Zang Bing Za Zhi. 21, 609–613. doi: 10.3760/cma.j.issn.1007-3418.2013.08.011
Li, N., Fu, H., Tie, Y., Hu, Z., Kong, W., Wu, Y., et al. (2009). miR-34a inhibits migration and invasion by down-regulation of c-Met expression in human hepatocellular carcinoma cells. Cancer Lett. 275, 44–53. doi: 10.1016/j.canlet.2008.09.035
Li, S., Li, J., Fei, B. Y., Shao, D., Pan, Y., Mo, Z. H., et al. (2015). MiR-27a promotes hepatocellular carcinoma cell proliferation through suppression of its target gene peroxisome proliferator-activated receptor γ. Chinese Med. J. 128, 941–947. doi: 10.4103/0366-6999.154302
Li, T., Yin, J., Yuan, L., Wang, S., Yang, L., Du, X., et al. (2014). Downregulation of microRNA-139 is associated with hepatocellular carcinoma risk and short-term survival. Oncol. Rep. 31, 1699–1706. doi: 10.3892/or.2014.3032
Li, Y., Tan, W., Neo, T. W., Aung, M. O., Wasser, S., Lim, S. G., et al. (2009). Role of the miR-106b-25 microRNA cluster in hepatocellular carcinoma. Cancer Sci. 100, 1234–1242. doi: 10.1111/j.1349-7006.2009.01164.x
Liao, C. G., Kong, L. M., Zhou, P., Yang, X. L., Huang, J. G., Zhang, H. L., et al. (2014). miR-10b is overexpressed in hepatocellular carcinoma and promotes cell proliferation, migration and invasion through RhoC, uPAR and MMPs. J. Transl. Med. 12, 234–234. doi: 10.1186/s12967-014-0234-x
Lin, S. L., Miller, J. D., and Ying, S. Y. (2006). Intronic microRNA (miRNA). J. Biomed. Biotechnol. 2006:26818. doi: 10.1155/JBB/2006/26818
Liu, B., Li, J., and Cairns, M. J. (2014). Identifying miRNAs, targets and functions. Brief. Bioinformatics 15, 1–19. doi: 10.1093/bib/bbs075
Liu, D., Wu, J., Liu, M., Yin, H., He, J., and Zhang, B. (2015). Downregulation of miRNA-30c and miR-203a is associated with hepatitis C virus core protein-induced epithelial–mesenchymal transition in normal hepatocytes and hepatocellular carcinoma cells. Biochem. Biophys. Res. Commun. 464, 1215–1221. doi: 10.1016/j.bbrc.2015.07.107
Liu, D. Z., Zhang, H. Y., Long, X. L., Zou, S. L., Zhang, X. Y., Han, G. Y., et al. (2015). MIR-150 promotes prostate cancer stem cell development via suppressing p27Kip1. Eur. Rev. Med. Pharmacol. Sci. 19, 4344–4352.
Liu, F., Kong, X., Lv, L., and Gao, J. (2015). TGF-beta1 acts through miR-155 to down-regulate TP53INP1 in promoting epithelial-mesenchymal transition and cancer stem cell phenotypes. Cancer Lett. 359, 288–298. doi: 10.1016/j.canlet.2015.01.030
Liu, J., Stevens, P. D., and Gao, T. (2011). mTOR-dependent regulation of PHLPP expression controls the rapamycin sensitivity in cancer cells. J. Biol. Chem. 286, 6510–6520. doi: 10.1074/jbc.M110.183087
Liu, J. L., Jiang, L., Lin, Q. X., Deng, C. Y., Mai, L. P., Zhu, J. N., et al. (2012). MicroRNA 16 enhances differentiation of human bone marrow mesenchymal stem cells in a cardiac niche toward myogenic phenotypes in vitro. Life Sci. 90, 1020–1026. doi: 10.1016/j.lfs.2012.05.011
Liu, L., Cai, X., Liu, E., Tian, X., and Tian, C. (2017). MicroRNA-18a promotes proliferation and metastasis in hepatocellular carcinoma via targeting KLF4. Oncotarget 8, 68263–68269. doi: 10.18632/oncotarget.19293
Liu, Y., Liu, W., Hu, C., Xue, Z., Wang, G., Ding, B., et al. (2011). MiR-17 modulates osteogenic differentiation through a coherent feed-forward loop in mesenchymal stem cells isolated from periodontal ligaments of patients with periodontitis. Stem Cells 29, 1804–1816. doi: 10.1002/stem.728
Lou, G., Song, X., Yang, F., Wu, S., Wang, J., Chen, Z., et al. (2015). Exosomes derived from miR-122-modified adipose tissue-derived MSCs increase chemosensitivity of hepatocellular carcinoma. J. Hematol. Oncol. 8:122. doi: 10.1186/s13045-015-0220-7
Lou, W., Liu, J., Gao, Y., Zhong, G., Ding, B., Xu, L., et al. (2018). MicroRNA regulation of liver cancer stem cells. Am. J. Cancer Res. 8, 1126–1141.
Lu, C. Y., Lin, K. Y., Tien, M. T., Wu, C. T., Uen, Y. H., and Tseng, T. L. (2013). Frequent DNA methylation of MiR-129-2 and its potential clinical implication in hepatocellular carcinoma. Genes Chromosomes Cancer 52, 636–643. doi: 10.1002/gcc.22059
Lu, J., Getz, G., Miska, E. A., Alvarez-Saavedra, E., Lamb, J., Peck, D., et al. (2005). MicroRNA expression profiles classify human cancers. Nature 435, 834–838. doi: 10.1038/nature03702
Luo, J., Chen, M., Huang, H., Yuan, T., Zhang, M., Zhang, K., et al. (2013). Circulating microRNA-122a as a diagnostic marker for hepatocellular carcinoma. Onco. Targets. Ther. 6, 577–583. doi: 10.2147/OTT.S44215
Ma, S., Tang, K. H., Chan, Y. P., Lee, T. K., Kwan, P. S., Castilho, A., et al. (2010). miR-130b Promotes CD133(+) liver tumor-initiating cell growth and self-renewal via tumor protein 53-induced nuclear protein 1. Cell Stem Cell 7, 694–707. doi: 10.1016/j.stem.2010.11.010
Malaguarnera, G., Cataudella, E., Giordano, M., Nunnari, G., Chisari, G., and Malaguarnera, M. (2012). Toxic hepatitis in occupational exposure to solvents. World J. Gastroenterol. 18, 2756–2766. doi: 10.3748/wjg.v18.i22.2756
Marrero, J. A., Fontana, R. J., Su, G. L., Conjeevaram, H. S., Emick, D. M., and Lok, A. S. (2002). NAFLD may be a common underlying liver disease in patients with hepatocellular carcinoma in the United States. Hepatology 36, 1349–1354. doi: 10.1002/hep.1840360609
Martinez-Sanchez, A., Dudek, K. A., and Murphy, C. L. (2012). Regulation of human chondrocyte function through direct inhibition of cartilage master regulator SOX9 by microRNA-145 (miRNA-145). J. Biol. Chem. 287, 916–924. doi: 10.1074/jbc.M111.302430
Meacham, C. E., and Morrison, S. J. (2013). Tumour heterogeneity and cancer cell plasticity. Nature 501, 328–337. doi: 10.1038/nature12624
Mei, Y., Bian, C., Li, J., Du, Z., Zhou, H., Yang, Z., et al. (2013). miR-21 modulates the ERK-MAPK signaling pathway by regulating SPRY2 expression during human mesenchymal stem cell differentiation. J. Cell. Biochem. 114, 1374–1384. doi: 10.1002/jcb.24479
Meng, F.-L., Wang, W., and Jia, D.- W. (2014). Diagnostic and prognostic significance of serum miR-24-3p in HBV-related hepatocellular carcinoma. Med. Oncol. 31:177. doi: 10.1007/s12032-014-0177-3
Mens, M. M. J., and Ghanbari, M. (2018). Cell cycle regulation of stem cells by MicroRNAs. Stem Cell Rev. Rep. 14, 309–322. doi: 10.1007/s12015-018-9808-y
Miranda, K. C., Huynh, T., Tay, Y., Ang, Y. S., Tam, W. L., Thomson, A. M., et al. (2006). A pattern-based method for the identification of MicroRNA binding sites and their corresponding heteroduplexes. Cell 126, 1203–1217. doi: 10.1016/j.cell.2006.07.031
Mohammed, M. K., Shao, C., Wang, J., Wei, Q., Wang, X., Collier, Z., et al. (2016). Wnt/beta-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance. Genes Dis. 3, 11–40. doi: 10.1016/j.gendis.2015.12.004
Murakami, Y., Tamori, A., Itami, S., Tanahashi, T., Toyoda, H., Tanaka, M., et al. (2013). The expression level of miR-18b in hepatocellular carcinoma is associated with the grade of malignancy and prognosis. BMC Cancer 13:566. doi: 10.1186/1471-2407-13-99
Murakami, Y., Yasuda, T., Saigo, K., Urashima, T., Toyoda, H., Okanoue, T., et al. (2006). Comprehensive analysis of microRNA expression patterns in hepatocellular carcinoma and non-tumorous tissues. Oncogene 25, 2537–2545. doi: 10.1038/sj.onc.1209283
Negrini, M., Gramantieri, L., Sabbioni, S., and Croce, C. M. (2011). microRNA involvement in hepatocellular carcinoma. Anticancer Agents Med. Chem. 11, 500–521. doi: 10.2174/187152011796011037
Negrini, M., Nicoloso, M. S., and Calin, G. A. (2009). MicroRNAs and cancer—new paradigms in molecular oncology. Curr. Opin. Cell Biol. 21, 470–479. doi: 10.1016/j.ceb.2009.03.002
Nie, Y., Han, B. M., Liu, X. B., Yang, J. J., Wang, F., Cong, X. F., et al. (2011). Identification of MicroRNAs involved in hypoxia- and serum deprivation-induced apoptosis in mesenchymal stem cells. Int. J. Biol. Sci. 7, 762–768. doi: 10.7150/ijbs.7.762
O'Brien, C. A., Pollett, A., Gallinger, S., and Dick, J. E. (2007). A human colon cancer cell capable of initiating tumour growth in immunodeficient mice. Nature 445, 106–110. doi: 10.1038/nature05372
Pan, L., Huang, S., He, R., Rong, M., Dang, Y., Chen, G., et al. (2014). Decreased expression and clinical significance of miR-148a in hepatocellular carcinoma tissues. Eur. J. Med. Res. 19, 68–68. doi: 10.1186/s40001-014-0068-2
Pan, W., Zhu, S., Yuan, M., Cui, H., Wang, L., Luo, X., et al. (2010). MicroRNA-21 and microRNA-148a contribute to DNA hypomethylation in lupus CD4+ T cells by directly and indirectly targeting DNA methyltransferase 1. J. Immunol. 184, 6773–6781. doi: 10.4049/jimmunol.0904060
Peter, M. E. (2009). Let-7 and miR-200 microRNAs: Guardians against pluripotency and cancer progression. Cell Cycle 8, 843–852. doi: 10.4161/cc.8.6.7907
Peuget, S., Bonacci, T., Soubeyran, P., Iovanna, J., and Dusetti, N. J. (2014). Oxidative stress-induced p53 activity is enhanced by a redox-sensitive TP53INP1 SUMOylation. Cell Death Differ. 21, 1107–1118. doi: 10.1038/cdd.2014.28
Peveling-Oberhag, J., Seiz, A., Döring, C., Hartmann, S., Köberle, V., Liese, J., et al. (2014). MicroRNA profiling of laser-microdissected hepatocellular carcinoma reveals an oncogenic phenotype of the tumor capsule. Transl. Oncol. 7, 672–680. doi: 10.1016/j.tranon.2014.09.003
Pode-Shakked, N., Shukrun, R., Mark-Danieli, M., Tsvetkov, P., Bahar, S., Pri-Chen, S., et al. (2013). The isolation and characterization of renal cancer initiating cells from human Wilms' tumour xenografts unveils new therapeutic targets. EMBO Mol. Med. 5, 18–37. doi: 10.1002/emmm.201201516
Potapova, A., Albat, C., Hasemeier, B., Haeussler, K., Lamprecht, S., Suerbaum, S., et al. (2011). Systematic cross-validation of 454 sequencing and pyrosequencing for the exact quantification of DNA methylation patterns with single CpG resolution. BMC Biotechnol. 11:6. doi: 10.1186/1472-6750-11-6
Pritchard, C. C., Cheng, H. H., and Tewari, M. (2012). MicroRNA profiling: approaches and considerations. Nat. Rev. Genet. 13, 358–369. doi: 10.1038/nrg3198
Reddy, S. K., Steel, J. L., Chen, H. W., DeMateo, D. J., Cardinal, J., Behari, J., et al. (2012). Outcomes of curative treatment for hepatocellular cancer in nonalcoholic steatohepatitis versus hepatitis C and alcoholic liver disease. Hepatology 55, 1809–1819. doi: 10.1002/hep.25536
Ren, F. H., Yang, H., He, R. Q., Lu, J. N., Lin, X. G., Liang, H. W., et al. (2018). Analysis of microarrays of miR-34a and its identification of prospective target gene signature in hepatocellular carcinoma. BMC Cancer 18:12. doi: 10.1186/s12885-017-3941-x
Rong, M., He, R., Dang, Y., and Chen, G. (2014). Expression and clinicopathological significance of miR-146a in hepatocellular carcinoma tissues. Ups. J. Med. Sci. 119, 19–24. doi: 10.3109/03009734.2013.856970
Roscigno, G., Puoti, I., Giordano, I., Donnarumma, E., Russo, V., Affinito, A., et al. (2017). MiR-24 induces chemotherapy resistance and hypoxic advantage in breast cancer. Oncotarget 8, 19507–19521. doi: 10.18632/oncotarget.14470
Schoolmeesters, A., Eklund, T., Leake, D., Vermeulen, A., Smith, Q., Force Aldred, S., et al. (2009). Functional profiling reveals critical role for miRNA in differentiation of human mesenchymal stem cells. PLoS ONE 4:e5605. doi: 10.1371/journal.pone.0005605
Schraivogel, D., Weinmann, L., Beier, D., Tabatabai, G., Eichner, A., Zhu, J. Y., et al. (2011). CAMTA1 is a novel tumour suppressor regulated by miR-9/9* in glioblastoma stem cells. EMBO J. 30, 4309–4322. doi: 10.1038/emboj.2011.301
Shaalan, Y. M., Handoussa, H., Youness, R. A., Assal, R. A., El-Khatib, A. H., Linscheid, M. W., et al. (2018). Destabilizing the interplay between miR-1275 and IGF2BPs by Tamarix articulata and quercetin in hepatocellular carcinoma. Nat. Prod. Res. 32, 2217–2220. doi: 10.1080/14786419.2017.1366478
Shariff, M. I., Cox, I. J., Gomaa, A. I., Khan, S. A., Gedroyc, W., and Taylor-Robinson, S. D. (2009). Hepatocellular carcinoma: current trends in worldwide epidemiology, risk factors, diagnosis and therapeutics. Expert Rev. Gastroenterol. Hepatol. 3, 353–367. doi: 10.1586/egh.09.35
Shen, J., Wang, S., Zhang, Y. J., Kappil, M. A., Chen Wu, H., Kibriya, M. G., et al. (2012). Genome-wide aberrant DNA methylation of microRNA host genes in hepatocellular carcinoma. Epigenetics 7, 1230–1237. doi: 10.4161/epi.22140
Shen, S., Lin, Y., Yuan, X., Shen, L., Chen, J., Chen, L., et al. (2016). Biomarker MicroRNAs for diagnosis, prognosis and treatment of hepatocellular carcinoma: a functional survey and comparison. Sci. Rep. 6:38311. doi: 10.1038/srep38311
Shi, W., Zhang, Z., Yang, B., Guo, H., Jing, L., Liu, T., et al. (2017). Overexpression of microRNA let-7 correlates with disease progression and poor prognosis in hepatocellular carcinoma. Medicine 96, e7764–e7764. doi: 10.1097/MD.0000000000007764
Shimono, Y., Mukohyama, J., Nakamura, S., and Minami, H. (2015). MicroRNA regulation of human breast cancer stem cells. J. Clin. Med. 5:E2. doi: 10.3390/jcm5010002
Shimono, Y., Zabala, M., Cho, R. W., Lobo, N., Dalerba, P., Qian, D., et al. (2009). Downregulation of miRNA-200c links breast cancer stem cells with normal stem cells. Cell 138, 592–603. doi: 10.1016/j.cell.2009.07.011
Skårn, M., Namløs, H. M., Noordhuis, P., Wang, M. Y., Meza-Zepeda, L. A., Myklebost, O., et al. (2012). Adipocyte differentiation of human bone marrow-derived stromal cells is modulated by microRNA-155, microRNA-221, and microRNA-222. Stem Cells Dev. 21, 873–883. doi: 10.1089/scd.2010.0503
Song, B., Wang, Y., Titmus, M. A., Botchkina, G., Formentini, A., Kornmann, M., et al. (2010). Molecular mechanism of chemoresistance by miR-215 in osteosarcoma and colon cancer cells. Mol. Cancer 9:96. doi: 10.1186/1476-4598-9-96
Song, J., Ouyang, Y., Che, J., Li, X., Zhao, Y., Yang, K., et al. (2017). Potential value of miR-221/222 as diagnostic, prognostic, and therapeutic biomarkers for diseases. Front. Immunol. 8:56. doi: 10.3389/fimmu.2017.00056
Stadler, B., Ivanovska, I., Mehta, K., Song, S., Nelson, A., Tan, Y., et al. (2010). Characterization of microRNAs involved in embryonic stem cell states. Stem Cells Dev. 19, 935–950. doi: 10.1089/scd.2009.0426
Su, H., Yang, J. R., Xu, T., Huang, J., Xu, L., Yuan, Y., et al. (2009). MicroRNA-101, down-regulated in hepatocellular carcinoma, promotes apoptosis and suppresses tumorigenicity. Cancer Res. 69, 1135–1142. doi: 10.1158/0008-5472.CAN-08-2886
Sun, C., Yao, X., Jiang, Q., and Sun, X. (2018). miR-106b targets DAB2 to promote hepatocellular carcinoma cell proliferation and metastasis. Oncol. Lett. 16, 3063–3069. doi: 10.3892/ol.2018.8970
Sun, F., Wang, J., Pan, Q., Yu, Y., Zhang, Y., Wan, Y., et al. (2009). Characterization of function and regulation of miR-24-1 and miR-31. Biochem. Biophys. Res. Commun. 380, 660–665. doi: 10.1016/j.bbrc.2009.01.161
Sun, T. Y., Xie, H. J., Li, Z., Kong, L. F., Gou, X. N., Li, D. J., et al. (2017). miR-34a regulates HDAC1 expression to affect the proliferation and apoptosis of hepatocellular carcinoma. Am. J. Transl. Res. 9, 103–114.
Sun, X., Liu, J., Xu, C., Tang, S. C., and Ren, H. (2016). The insights of Let-7 miRNAs in oncogenesis and stem cell potency. J. Cell. Mol. Med. 20, 1779–1788. doi: 10.1111/jcmm.12861
Sun, Z., Han, Q., Zhou, N., Wang, S., Lu, S., Bai, C., et al. (2013). MicroRNA-9 enhances migration and invasion through KLF17 in hepatocellular carcinoma. Mol. Oncol. 7, 884–894. doi: 10.1016/j.molonc.2013.04.007
Sung, S. Y., Liao, C. H., Wu, H. P., Hsiao, W. C., Wu, I. H., Jinpu, et al. (2013). Loss of let-7 microRNA upregulates IL-6 in bone marrow-derived mesenchymal stem cells triggering a reactive stromal response to prostate cancer. PLoS ONE 8:e71637. doi: 10.1371/journal.pone.0071637
Szotek, P. P., Pieretti-Vanmarcke, R., Masiakos, P. T., Dinulescu, D. M., Connolly, D., Foster, R., et al. (2006). Ovarian cancer side population defines cells with stem cell-like characteristics and Mullerian Inhibiting Substance responsiveness. Proc. Natl. Acad. Sci. U.S.A. 103, 11154–11159. doi: 10.1073/pnas.0603672103
Takahashi, R.-,u., Miyazaki, H., and Ochiya, T. (2014). The role of microRNAs in the regulation of cancer stem cells. Front. Genet. 4:295. doi: 10.3389/fgene.2013.00295
Takata, A., Otsuka, M., Yoshikawa, T., Kishikawa, T., Ohno, M., and Koike, K. (2013). MicroRNAs and liver function. Minerva Gastroenterol. Dietol. 59, 187–203.
Tan, Y., Ge, G., Pan, T., Wen, D., Chen, L., Yu, X., et al. (2014). A serum microRNA panel as potential biomarkers for hepatocellular carcinoma related with hepatitis B virus. PLoS ONE 9:e107986. doi: 10.1371/journal.pone.0107986
Tang, H. (2013). miR-10a regulates epithelial-mesenchymal transition and adhesion and angiogenesis in hepatoma. FASEB J. 27(1_supple), lb153–lb153. doi: 10.1371/journal.pone.0064273
Tang, T., Yang, Z., Zhu, Q., Wu, Y., Sun, K., Alahdal, M., et al. (2018). Up-regulation of miR-210 induced by a hypoxic microenvironment promotes breast cancer stem cell metastasis, proliferation, and self-renewal by targeting E-cadherin. FASEB J. 32, 6965–6981. doi: 10.1096/fj.201801013R
Tao, J., Jiang, L., and Chen, X. (2018). Roles of micro RNA in liver cancer. Liver Res. 2, 61–72. doi: 10.1016/j.livres.2018.06.002
Taube, J. H., Malouf, G. G., Lu, E., Sphyris, N., Vijay, V., Ramachandran, P. P., et al. (2013). Epigenetic silencing of microRNA-203 is required for EMT and cancer stem cell properties. Sci. Rep. 3:2687. doi: 10.1038/srep02687
Thurnherr, T., Mah, W. C., Lei, Z., Jin, Y., Rozen, S. G., and Lee, C. G. (2016). Differentially expressed miRNAs in hepatocellular carcinoma target genes in the genetic information processing and metabolism pathways. Sci. Rep. 6:20065. doi: 10.1038/srep20065
Tong, Y., Xu, W., Han, H., Chen, Y., Yang, J., Qiao, H., et al. (2011). Tanshinone IIA increases recruitment of bone marrow mesenchymal stem cells to infarct region via up-regulating stromal cell-derived factor-1/CXC chemokine receptor 4 axis in a myocardial ischemia model. Phytomedicine 18, 443–450. doi: 10.1016/j.phymed.2010.10.009
Tsai, S. C., Lin, C. C., Shih, T. C., Tseng, R. J., Yu, M. C., Lin, Y. J., et al. (2017). The miR-200b–ZEB1 circuit regulates diverse stemness of human hepatocellular carcinoma. Mol. Carcinog. 56, 2035–2047. doi: 10.1002/mc.22657
Tsang, F. H., Au, V., Lu, W. J., Shek, F. H., Liu, A. M., Luk, J. M., et al. (2014). Prognostic marker microRNA-125b inhibits tumorigenic properties of hepatocellular carcinoma cells via suppressing tumorigenic molecule eIF5A2. Dig. Dis. Sci. 59, 2477–2487. doi: 10.1007/s10620-014-3184-5
Tu, H., Wei, G., Cai, Q., Chen, X., Sun, Z., Cheng, C., et al. (2015). MicroRNA-212 inhibits hepatocellular carcinoma cell proliferation and induces apoptosis by targeting FOXA1. Onco. Targets. Ther. 8, 2227–2235. doi: 10.2147/OTT.S87976
Turchi, L., Debruyne, D. N., Almairac, F., Virolle, V., Fareh, M., Neirijnck, Y., et al. (2013). Tumorigenic potential of miR-18A* in Glioma initiating cells requires NOTCH-1 signaling. Stem Cells. 31, 1252–1265. doi: 10.1002/stem.1373
Ullmann, P., Nurmik, M., Schmitz, M., Rodriguez, F., Weiler, J., Qureshi-Baig, K., et al. (2019). Tumor suppressor miR-215 counteracts hypoxia-induced colon cancer stem cell activity. Cancer Lett. 450, 32–41. doi: 10.1016/j.canlet.2019.02.030
Valeri, N., Vannini, I., Fanini, F., Calore, F., Adair, B., and Fabbri, M. (2009). Epigenetics, miRNAs, and human cancer: a new chapter in human gene regulation. Mamm Genome. 20, 573–580. doi: 10.1007/s00335-009-9206-5
Wahid, F., Shehzad, A., Khan, T., and Kim, Y. Y. (2010). MicroRNAs: synthesis, mechanism, function, and recent clinical trials. Biochim. Biophys. Acta. 1803, 1231–1243. doi: 10.1016/j.bbamcr.2010.06.013
Wang, C., Wang, X., Su, Z., Fei, H., Liu, X., Pan, Q., et al. (2015). MiR-25 promotes hepatocellular carcinoma cell growth, migration and invasion by inhibiting RhoGDI1. Oncotarget 6, 36231–36244. doi: 10.18632/oncotarget.4740
Wang, J., Li, J., Wang, X., Zheng, C., and Ma, W. (2013). Downregulation of microRNA-214 and overexpression of FGFR-1 contribute to hepatocellular carcinoma metastasis. Biochem. Biophys. Res. Commun. 439, 47–53. doi: 10.1016/j.bbrc.2013.08.032
Wang, L.-J., He, C. C., Sui, X., Cai, M. J., Zhou, C. Y., Ma, J. L., et al. (2015). MiR-21 promotes intrahepatic cholangiocarcinoma proliferation and growth in vitro and in vivo by targeting PTPN14 and PTE. Oncotarget 6, 5932–5946. doi: 10.18632/oncotarget.3465
Wang, M., Zhang, J., Tong, L., Ma, X., and Qiu, X. (2015). MiR-195 is a key negative regulator of hepatocellular carcinoma metastasis by targeting FGF2 and VEGFA. Int. J. Clin. Exp. Pathol. 8, 14110–14120.
Wang, M., Zhao, J., Zhang, L., Wei, F., Lian, Y., Wu, Y., et al. (2017). Role of tumor microenvironment in tumorigenesis. J. Cancer. 8, 761–773. doi: 10.7150/jca.17648
Wang, S., Yin, J., Li, T., Yuan, L., Wang, D., He, J., et al. (2015). Upregulated circulating miR-150 is associated with the risk of intrahepatic cholangiocarcinoma. Oncol. Rep. 33, 819–825. doi: 10.3892/or.2014.3641
Wang, W. Y., Zhang, H. F., Wang, L., Ma, Y. P., Gao, F., Zhang, S. J., et al. (2014). miR-21 expression predicts prognosis in hepatocellular carcinoma. Clin. Res. Hepatol. Gastroenterol. 38, 715–719. doi: 10.1016/j.clinre.2014.07.001
Wang, X., Zhang, W., Sun, X., Lin, Y., and Chen, W. (2018). Cancer-associated fibroblasts induce epithelial-mesenchymal transition through secreted cytokines in endometrial cancer cells. Oncol. Lett. 15, 5694–5702. doi: 10.3892/ol.2018.8000
Wang, Y., Fan, H., Zhao, G., Liu, D., Du, L., Wang, Z., et al. (2012). miR-16 inhibits the proliferation and angiogenesis-regulating potential of mesenchymal stem cells in severe pre-eclampsia. FEBS J. 279, 4510–4524. doi: 10.1111/febs.12037
Wang, Y., Lee, A. T., Ma, J. Z., Wang, J., Ren, J., Yang, Y., et al. (2008). Profiling microRNA expression in hepatocellular carcinoma reveals microRNA-224 up-regulation and apoptosis inhibitor-5 as a microRNA-224-specific target. J. Biol. Chem. 283, 13205–13215. doi: 10.1074/jbc.M707629200
Wei, W., Wanjun, L., Hui, S., Dongyue, C., Xinjun, Y., Jisheng, Z., et al. (2013). miR-203 inhibits proliferation of HCC cells by targeting survivin. Cell Biochem. Funct. 31, 82–85. doi: 10.1002/cbf.2863
Wei, X., Tan, C., Tang, C., Ren, G., Xiang, T., Qiu, Z., et al. (2013). Epigenetic repression of miR-132 expression by the hepatitis B virus x protein in hepatitis B virus-related hepatocellular carcinoma. Cell. Signal. 25, 1037–1043. doi: 10.1016/j.cellsig.2013.01.019
Wheeler, B. M., Heimberg, A. M., Moy, V. N., Sperling, E. A., Holstein, T. W., Heber, S., et al. (2009). The deep evolution of metazoan microRNAs. Evol. Dev. 11, 50–68. doi: 10.1111/j.1525-142X.2008.00302.x
White, D. L., Kanwal, F., and El–Serag, H. B. (2012). Association between nonalcoholic fatty liver disease and risk for hepatocellular cancer, based on systematic review. Clin. Gastroenterol. Hepatol. 10, 1342–1359. e2. doi: 10.1016/j.cgh.2012.10.001
Whiteside, T. (2008). The tumor microenvironment and its role in promoting tumor growth. Oncogene 27:5904–5912. doi: 10.1038/onc.2008.271
Wong, Q. W., Lung, R. W., Law, P. T., Lai, P. B., Chan, K. Y., To, K. F., et al. (2008). MicroRNA-223 is commonly repressed in hepatocellular carcinoma and potentiates expression of Stathmin1. Gastroenterology. 135, 257–269. doi: 10.1053/j.gastro.2008.04.003
Xia, C., Shui, L., Lou, G., Ye, B., Zhu, W., Wang, J., et al. (2017). 0404 inhibits hepatocellular carcinoma through a p53/miR-34a/SIRT1 positive feedback loop. Sci. Rep. 7:4396. doi: 10.1038/s41598-017-04487-x
Xie, D., Yuan, P., Wang, D., Jin, H., and Chen, H. (2017). Expression and prognostic significance of miR-375 and miR-221 in liver cancer. Oncol. Lett. 14, 2305–2309. doi: 10.3892/ol.2017.6423
Xie, F., Yuan, Y., Xie, L., Ran, P., Xiang, X., Huang, Q., et al. (2017). miRNA-320a inhibits tumor proliferation and invasion by targeting c-Myc in human hepatocellular carcinoma. Onco. Targets. Ther. 10, 885–894. doi: 10.2147/OTT.S122992
Xie, Y., Yao, Q., Butt, A. M., Guo, J., Tian, Z., Bao, X., et al. (2014). Expression profiling of serum microRNA-101 in HBV-associated chronic hepatitis, liver cirrhosis, and hepatocellular carcinoma. Cancer Biol. Ther. 15, 1248–1255. doi: 10.4161/cbt.29688
Xiong, Y., Fang, J. H., Yun, J. P., Yang, J., Zhang, Y., Jia, W. H., et al. (2010). Effects of MicroRNA-29 on apoptosis, tumorigenicity, and prognosis of hepatocellular carcinoma. Hepatology 51, 836–845. doi: 10.1002/hep.23380
Xu, C., Ren, G., Cao, G., Chen, Q., Shou, P., Zheng, C., et al. (2013). miR-155 regulates immune modulatory properties of mesenchymal stem cells by targeting TAK1-binding protein 2. J. Biol. Chem. 288, 11074–11079. doi: 10.1074/jbc.M112.414862
Xu, J., Kang, Y., Liao, W. M., and Yu, L. (2012). MiR-194 regulates chondrogenic differentiation of human adipose-derived stem cells by targeting Sox5. PLoS ONE 7:e31861. doi: 10.1371/journal.pone.0031861
Xu, Q., Zhang, M., Tu, J., Pang, L., Cai, W., and Liu, X. (2015). MicroRNA-122 affects cell aggressiveness and apoptosis by targeting PKM2 in human hepatocellular carcinoma. Oncol. Rep. 34, 2054–2064. doi: 10.3892/or.2015.4175
Xu, W., Ji, J., Xu, Y., Liu, Y., Shi, L., Liu, Y., et al. (2015). MicroRNA-191, by promoting the EMT and increasing CSC-like properties, is involved in neoplastic and metastatic properties of transformed human bronchial epithelial cells. Mol. Carc. 54 (Suppl. 1), E148–E161. doi: 10.1002/mc.22221
Xu, X. L., Xing, B. C., Han, H. B., Zhao, W., Hu, M. H., Xu, Z. L., et al. (2009). The properties of tumor-initiating cells from a hepatocellular carcinoma patient's primary and recurrent tumor. Carcinogenesis 31, 167–174. doi: 10.1093/carcin/bgp232
Xu, Y., An, Y., Wang, Y., Zhang, C., Zhang, H., Huang, C., et al. (2013). miR-101 inhibits autophagy and enhances cisplatin-induced apoptosis in hepatocellular carcinoma cells. Oncol. Rep. 29, 2019–2024. doi: 10.3892/or.2013.2338
Xu, Y., Bu, X., Dai, C., and Shang, C. (2015). High serum microRNA-122 level is independently associated with higher overall survival rate in hepatocellular carcinoma patients. Tumor Biol. 36, 4773–4776. doi: 10.1007/s13277-015-3128-5
Yacoub, R. A., Fawzy, I. O., Assal, R. A., Hosny, K. A., Zekri, A. N., Esmat, G., et al. (2016). miR-34a: multiple opposing targets and one destiny in hepatocellular carcinoma. J. Clin. Transl. Hepatol. 4, 300–305. doi: 10.14218/JCTH.2016.00031
Yan, J. J., Zhang, Y. N., Liao, J. Z., Ke, K. P., Chang, Y., Li, P. Y., et al. (2015). MiR-497 suppresses angiogenesis and metastasis of hepatocellular carcinoma by inhibiting VEGFA and AEG-1. Oncotarget 6, 29527–29542. doi: 10.18632/oncotarget.5012
Yang, F., Zhang, L., Wang, F., Wang, Y., Huo, X. S., Yin, Y. X., et al. (2011). Modulation of the unfolded protein response is the core of microRNA-122-involved sensitivity to chemotherapy in hepatocellular carcinoma. Neoplasia 13, 590–600. doi: 10.1593/neo.11422
Yang, H., Cho, M. E., Li, T. W., Peng, H., Ko, K. S., Mato, J. M., et al. (2013). MicroRNAs regulate methionine adenosyltransferase 1A expression in hepatocellular carcinoma. J. Clin. Invest. 123, 285–298. doi: 10.1172/JCI63861
Yang, J. D., and Roberts, L. R. (2010). Epidemiology and management of hepatocellular carcinoma. Infect. Dis. Clin. North Am. 24, 899–919. doi: 10.1016/j.idc.2010.07.004
Yang, Y., Zhang, J., Xia, T., Li, G., Tian, T., Wang, M., et al. (2016). MicroRNA-210 promotes cancer angiogenesis by targeting fibroblast growth factor receptor-like 1 in hepatocellular carcinoma. Oncol. Rep. 36, 2553–2562. doi: 10.3892/or.2016.5129
Yata, K., Beder, L. B., Tamagawa, S., Hotomi, M., Hirohashi, Y., Grenman, R., et al. (2015). MicroRNA expression profiles of cancer stem cells in head and neck squamous cell carcinoma. Int. J. Oncol. 47, 1249–1256. doi: 10.3892/ijo.2015.3145
Yin, J., Hou, P., Wu, Z., Wang, T., and Nie, Y. (2015). Circulating miR-375 and miR-199a-3p as potential biomarkers for the diagnosis of hepatocellular carcinoma. Tumor Biol. 36, 4501–4507. doi: 10.1007/s13277-015-3092-0
Youness, R. A., El-Tayebi, H. M., Assal, R. A., Hosny, K., Esmat, G., and Abdelaziz, A. I. (2016). MicroRNA-486-5p enhances hepatocellular carcinoma tumor suppression through repression of IGF-1R and its downstream mTOR, STAT3 and c-Myc. Oncol. Lett. 12, 2567–2573. doi: 10.3892/ol.2016.4914
Yu, F., Lu, Z., Chen, B., Dong, P., and Zheng, J. (2015). microRNA-150: a promising novel biomarker for hepatitis B virus-related hepatocellular carcinoma. Diagn. Pathol. 10:129. doi: 10.1186/s13000-015-0369-y
Yu, L., Gong, X., Sun, L., Zhou, Q., Lu, B., Zhu, L., et al. (2016). The circular RNA Cdr1as act as an oncogene in hepatocellular carcinoma through targeting miR-7 expression. PLoS ONE 11:e0158347. doi: 10.1371/journal.pone.0158347
Yu, Y., Nangia-Makker, P., Farhana, L. G, Rajendra, S., Levi, E., Majumdar, A. P., et al. (2015). miR-21 and miR-145 cooperation in regulation of colon cancer stem cells. Mol. Cancer. 14:98. doi: 10.1186/s12943-015-0372-7
Yu, Z., Lin, X., Tian, M., and Chang, W. (2018). microRNA196b promotes cell migration and invasion by targeting FOXP2 in hepatocellular carcinoma. Oncol. Rep. 39, 731–738. doi: 10.3892/or.2017.6130
Yunqiao, L., Vanke, H., Jun, X., and Tangmeng, G. (2014). MicroRNA-206, down-regulated in hepatocellular carcinoma, suppresses cell proliferation and promotes apoptosis. Hepatogastroenterology 61, 1302–1307.
Zhang, H., Cai, K., Wang, J., Wang, X., Cheng, K., Shi, F., et al. (2014). MiR-7, inhibited indirectly by lincRNA HOTAIR, directly inhibits SETDB1 and reverses the EMT of breast cancer stem cells by downregulating the STAT3 pathway. Stem Cells. 32, 2858–2868. doi: 10.1002/stem.1795
Zhang, J. F., Fu, W. M., He, M. L., Xie, W. D., Lv, Q., Wan, G., et al. (2011). MiRNA-20a promotes osteogenic differentiation of human mesenchymal stem cells by co-regulating BMP signaling. RNA Biol. 8, 829–838. doi: 10.4161/rna.8.5.16043
Zhang, L., Huang, J., Yang, N., Greshock, J., Megraw, M. S., Giannakakis, A., et al. (2006). microRNAs exhibit high frequency genomic alterations in human cancer. Proc. Natl. Acad. Sci. U.S.A. 103:9136. doi: 10.1073/pnas.0508889103
Zhang, W. C., Chin, T. M., Yang, H., Nga, M. E., Lunny, D. P., Lim, E. K., et al. (2016). Tumour-initiating cell-specific miR-1246 and miR-1290 expression converge to promote non-small cell lung cancer progression. Nat. Commun. 7:11702. doi: 10.1038/ncomms11702
Zhang, Y., Guo, X., Xiong, L., Kong, X., Xu, Y., Liu, C., et al. (2012). MicroRNA-101 suppresses SOX9-dependent tumorigenicity and promotes favorable prognosis of human hepatocellular carcinoma. FEBS Lett. 586, 4362–4370. doi: 10.1016/j.febslet.2012.10.053
Zhao, Y., Jia, H. L., Zhou, H. J., Dong, Q. Z., Fu, L. Y., Yan, Z. W., et al. (2009). [Identification of metastasis-related microRNAs of hepatocellular carcinoma in hepatocellular carcinoma cell lines by quantitative real time PCR]. Zhonghua Gan Zang Bing Za Zhi. 17, 526–530.
Zheng, J., Dong, P., Gao, S., Wang, N., and Yu, F. (2013). High expression of serum miR-17-5p associated with poor prognosis in patients with hepatocellular carcinoma. Hepatogastroenterology 60, 549–552. doi: 10.5754/hge12754
Zhou, C., Liu, J., Li, Y., Liu, L., Zhang, X., Ma, C. Y., et al. (2011). microRNA-1274a, a modulator of sorafenib induced a disintegrin and metalloproteinase 9 (ADAM9) down-regulation in hepatocellular carcinoma. FEBS Lett. 585, 1828–1834. doi: 10.1016/j.febslet.2011.04.040
Zhou, J., Lu, S., Yang, S., Chen, H., Shi, H., Miao, M., et al. (2014). MicroRNA-127 post-transcriptionally downregulates Sept7 and suppresses cell growth in hepatocellular carcinoma cells. Cell. Physiol. Biochem. 33, 1537–1546. doi: 10.1159/000358717
Zhou, J., Yu, L., Gao, X., Hu, J., Wang, J., Dai, Z., et al. (2011). Plasma microRNA panel to diagnose hepatitis B virus-related hepatocellular carcinoma. J. Clin. Oncol. 29, 4781–4788. doi: 10.1200/JCO.2011.38.2697
Zhou, X., Yue, Y., Wang, R., Gong, B., and Duan, Z. (2017). MicroRNA-145 inhibits tumorigenesis and invasion of cervical cancer stem cells. Int. J. Oncol. 50, 853–862. doi: 10.3892/ijo.2017.3857
Zhu, H. T., Dong, Q. Z., Sheng, Y. Y., Wei, J. W., Wang, G., Zhou, H. J., et al. (2012). MicroRNA-29a-5p is a novel predictor for early recurrence of hepatitis B virus-related hepatocellular carcinoma after surgical resection. PLoS ONE 7:e52393. doi: 10.1371/journal.pone.0052393
Keywords: miRNA, cancer stem cell, methylation, mesencymal stem cells, hepatocellular carcinoma
Citation: Nasr MA, Salah RA, Abd Elkodous M, Elshenawy SE and El-Badri N (2019) Dysregulated MicroRNA Fingerprints and Methylation Patterns in Hepatocellular Carcinoma, Cancer Stem Cells, and Mesenchymal Stem Cells. Front. Cell Dev. Biol. 7:229. doi: 10.3389/fcell.2019.00229
Received: 25 July 2019; Accepted: 26 September 2019;
Published: 17 October 2019.
Edited by:
Mojgan Rastegar, University of Manitoba, CanadaReviewed by:
Kenneth K. W. To, The Chinese University of Hong Kong, ChinaZong Wei, Salk Institute for Biological Studies, United States
Copyright © 2019 Nasr, Salah, Abd Elkodous, Elshenawy and El-Badri. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nagwa El-Badri, bmVsYmFkcmlAemV3YWlsY2l0eS5lZHUuZWc=
†These authors have contributed equally to this work