 Kazuki Inoue
Kazuki Inoue Courtney Ng
Courtney Ng Yuhan Xia1
Yuhan Xia1 Baohong Zhao
Baohong Zhao- 1Arthritis and Tissue Degeneration Program, David Z. Rosensweig Genomics Research Center, Hospital for Special Surgery, New York, NY, United States
- 2Department of Medicine, Weill Cornell Medicine, New York, NY, United States
- 3Graduate Program in Cell and Developmental Biology, Weill Cornell Medicine Graduate School of Medical Sciences, New York, NY, United States
Osteoclasts are specialized bone-resorbing cells that contribute to physiological bone development and remodeling in bone metabolism throughout life. Abnormal production and activation of osteoclasts lead to excessive bone resorption in pathological conditions, such as in osteoporosis and in arthritic diseases with bone destruction. Recent epigenetic studies have shed novel insight into the dogma of the regulation of gene expression. microRNAs belong to a category of epigenetic regulators, which post-transcriptionally regulate and silence target gene expression, and thereby control a variety of biological events. In this review, we discuss miRNA biogenesis, the mechanisms utilized by miRNAs, several miRNAs that play important roles in osteoclast differentiation, function, survival and osteoblast-to-osteoclast communication, and their translational potential and challenges in bone biology and skeletal diseases.
Introduction
Bone loss related to multiple diseases, such as rheumatoid arthritis (RA), osteoporosis, psoriatic arthritis and periodontitis, is a prevalent cause of disability in patients (Schett and Gravallese, 2012; Goldring et al., 2013). Bone destruction significantly diminishes the quality of life and simultaneously can increase the risk of mortality of these patients. Osteoclasts are giant, multinucleated cells that are derived from monocyte/macrophage lineage and specialize in bone resorption through proteolytic degradation and acid decalcification of the bone matrix. Osteoclast-mediated bone resorption is essential for skeletal development and normal bone remodeling. However, abnormal osteoclast formation and activation in pathological conditions play a crucial role in osteolysis. A broader understanding of the regulatory mechanisms that govern osteoclastogenesis will facilitate development of novel therapeutic strategies for patients subjected to pathological bone resorption.
Myeloid osteoclast precursors first undergo differentiation into mononuclear osteoclasts in response to the master osteoclastogenic cytokine: receptor activator of nuclear factor-κB ligand (RANKL) accompanied by macrophage colony-stimulating factor (M-CSF) and immunoreceptor tyrosine-based activation motif (ITAM)-mediated co-stimulatory signaling pathways. Mononuclear osteoclast precursors then fuse into giant, mature polykaryons driven by the expression of cell-cell fusion genes, such as DC-Stamp, ATP6v0d2 and Gα13 (Yagi et al., 2005; Lee et al., 2006; Nakano et al., 2019). Osteoclastogenesis is delicately regulated and influenced in order to maintain the balance of osteoclastogenic and anti-osteoclastogenic mechanisms (Zhao and Ivashkiv, 2011). RANKL induces a broad range of signaling cascades including canonical and non-canonical NF-kB pathways, mitogen-activated kinase (MAPK) pathways and calcium signaling. These pathways activate downstream transcription factors to drive osteoclast differentiation, such as c-Fos, nuclear factor of activated T cells c1 (NFATc1), and B lymphocyte-induced maturation protein-1 (Blimp1). On the other hand, negative regulators mediating intrinsic anti-osteoclastogenic mechanisms, such as differentially expressed in FDCP 6 homolog (Def6), interferon regulatory factor (IRF8) and v-maf musculoaponeurotic fibrosarcoma oncogene family protein B (MafB), function as brakes to restrain excessive osteoclast formation and bone resorption (Humphrey et al., 2005; Asagiri and Takayanagi, 2007; Zhao B. et al., 2009; Nakashima and Takayanagi, 2011; Zhao and Ivashkiv, 2011; Xu and Teitelbaum, 2013; Li et al., 2014; Boyce et al., 2015; Binder et al., 2017). Some regulators were identified to preferentially limit pathological osteoclastic bone resorption, such as RBP-J and NF-κB p100 (Yao et al., 2009; Zhao et al., 2012). These regulators and associated mechanisms interact to form regulatory networks that coordinately modulate osteoclastic gene expression to govern osteoclast differentiation and function.
Over the past decade, epigenetic studies have gained rapidly growing research and clinical attention. Epigenetics investigates heritable changes in gene expression and cellular phenotype that do not involve alterations in DNA sequence, which provides a new level and dimension of understanding of gene expression. Major epigenetic mechanisms include DNA methylation, chromatin remodeling, histone posttranslational modifications and non-coding RNAs (ncRNAs) (Zhang and Cao, 2019). These mechanisms often act in concert to play crucial roles in the regulation of gene expression extensively involved in diverse biological and disease settings, such as cell proliferation and differentiation, development, cardiovascular disease, diabetes, autoimmune diseases, and cancer. In human genome, ncRNAs constitute more than 90% of the genome transcripts (Hangauer et al., 2013). ncRNAs consists of multiple classes of RNA that are not translated into proteins, which are classified into two categories by size; long non-coding RNAs (lncRNAs) (>200 nt) and small RNAs (<200 nt). Small RNAs include microRNAs (miRNAs), short interfering RNAs (siRNAs) and Piwi-interacting RNAs (piRNAs) (Kim et al., 2009; Esteller, 2011). Recently, a unique class of ncRNAs transcribed from sequences of enhancer regions, called enhancer RNAs (eRNAs), have been identified (Kim et al., 2010; Lam et al., 2014). Genome-wide cap analysis of gene expression (CAGE) identified eRNA regions at the Nrp2, Dcstamp, and Nfatc1 gene loci in osteoclasts. These eRNAs are necessary for the transcription of these genes that are important for osteoclast differentiation (Sakaguchi et al., 2018). To date, functions of lncRNAs in osteoclastogenesis are largely unknown. The most well-characterized ncRNAs in myeloid cells are miRNAs. In this review, we discuss miRNA biogenesis, several miRNAs that play important roles in osteoclast differentiation, function, survival, osteoblast-to-osteoclast communication, the mechanisms controlled by miRNAs, and their translational potential and challenges in bone biology and skeletal diseases (Table 1).
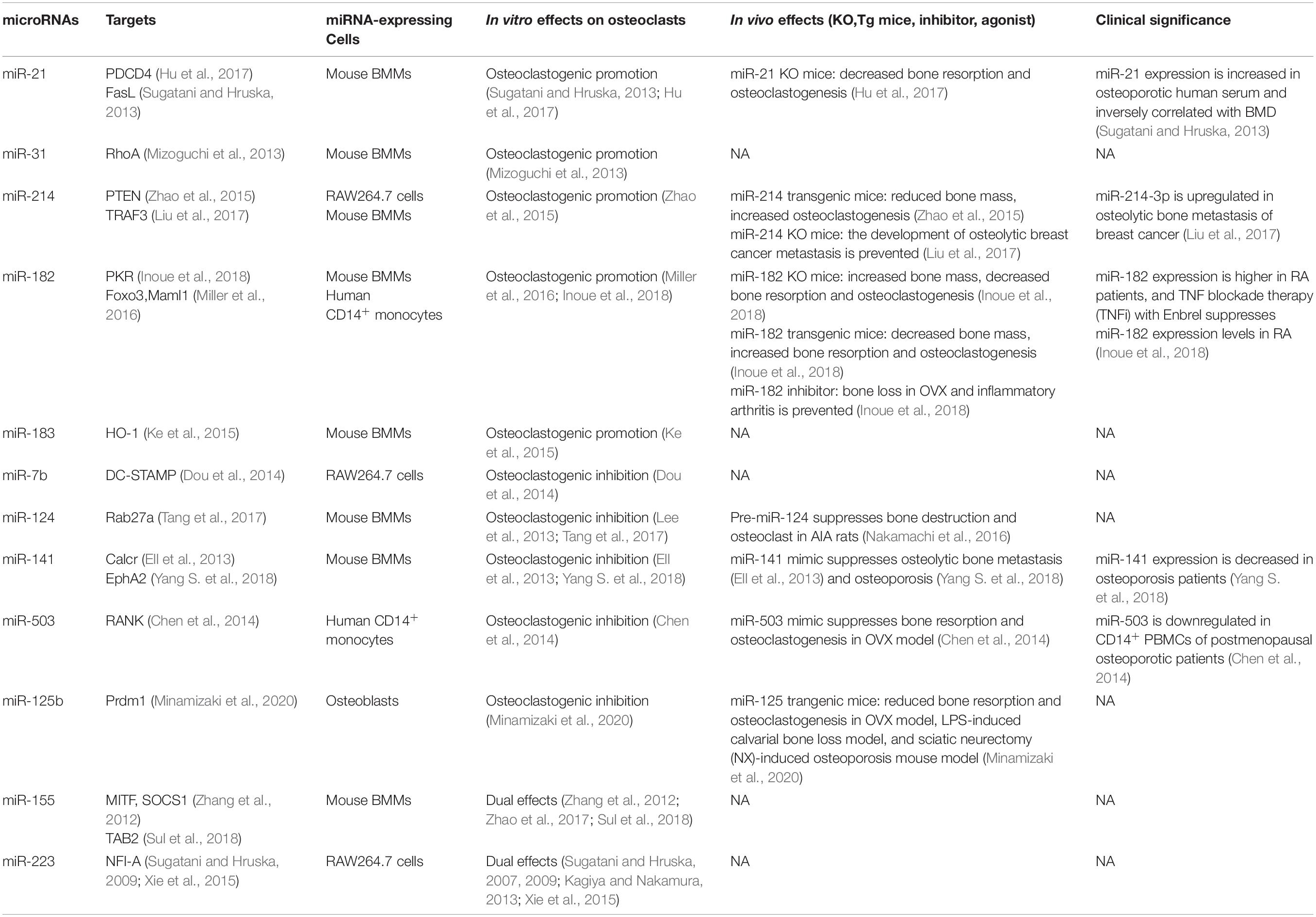
Table 1. Osteoclastogenesis and bone metabolism mediated by miRNAs.
Post-transcriptional Regulation of Target Gene Expression by miRNAs
MicroRNAs (miRNAs) constitute a class of small single-stranded non-coding RNAs (about 22 nucleotides), that function in diverse regulatory pathways through gene repression of their target mRNAs via post-transcriptional mechanisms (Lagos-Quintana et al., 2001; Bartel, 2004). miRNAs commonly bind to multiple target sequences at the 3′ untranslated region (3′-UTR) of mRNAs, through perfect complementary pairing of nucleotides 2–7 at the 5′ end of the miRNA (the miRNA seed region) (Bartel, 2009). These molecules repress gene expression via mRNA degradation or translational inhibition of their target mRNAs, or a combination of these mechanisms (Ling et al., 2013). A series of processes is required to generate mature miRNAs, beginning with the transcription of long precursor RNAs, called primary miRNAs (pri-miRNAs), by RNA polymerase II (Lee et al., 2004). In the nucleus, long pri-miRNAs are cropped into hairpin structures by the microprocessor complex, which consists of the RNase III enzyme, Drosha and RNA binding protein, DiGeorge syndrome critical region 8 (DGCR8). This process produces the precursor miRNAs (pre-miRNAs) that are approximately 60–70 nucleotides (Lee et al., 2003; Gregory et al., 2004). Pre-miRNAs are then exported from the nucleus by Exportin-5 (Exp5) into the cytoplasm where they are further cleaved by the RNase III enzyme, Dicer (Lund et al., 2004). Upon recognition, Dicer cleaves the pre-miRNA at the double-stranded RNA stem and the terminal loop to generate a mature ∼22 nucleotide miRNA duplex (Park et al., 2011). The miRNA duplex consists of two strands that are often referred to as a guide and a passenger strand. The passenger strand is typically degraded while the guide strand of the mature miRNA is incorporated into the RNA-induced silencing complex (RISC), which comprises Dicer, transactivating response RNA-binding protein (TRBP) and Argonaute proteins (AGO1-4) (Chendrimada et al., 2005; Peters and Meister, 2007). Active RISC is guided by miRNA to complementary target mRNAs and downregulates gene expressing by inducing mRNA degradation, translational repression, or a combination of the two mechanisms. AGO2 was identified to have catalytic slicer activity that can facilitate degradation of specific target mRNA. The target mRNAs sharing the same recognition sequences could compete for miRNA binding and influence the expression of each target gene. A single miRNA can recognize and regulate multiple mRNA targets, and consequently, may have a wide range and substantial effect on gene expression networks. Additionally, within the same cell types, the targets of an miRNA can be variable in response to differential environmental settings. Accordingly, miRNA-targeted gene regulation is both specific and sensitive to environmental changes. Multiple miRNAs can also be involved in the same biological processes, as exemplified by the miRNAs mentioned in this review, which play crucial and exclusive roles in osteoclast differentiation and function (Figure 1).
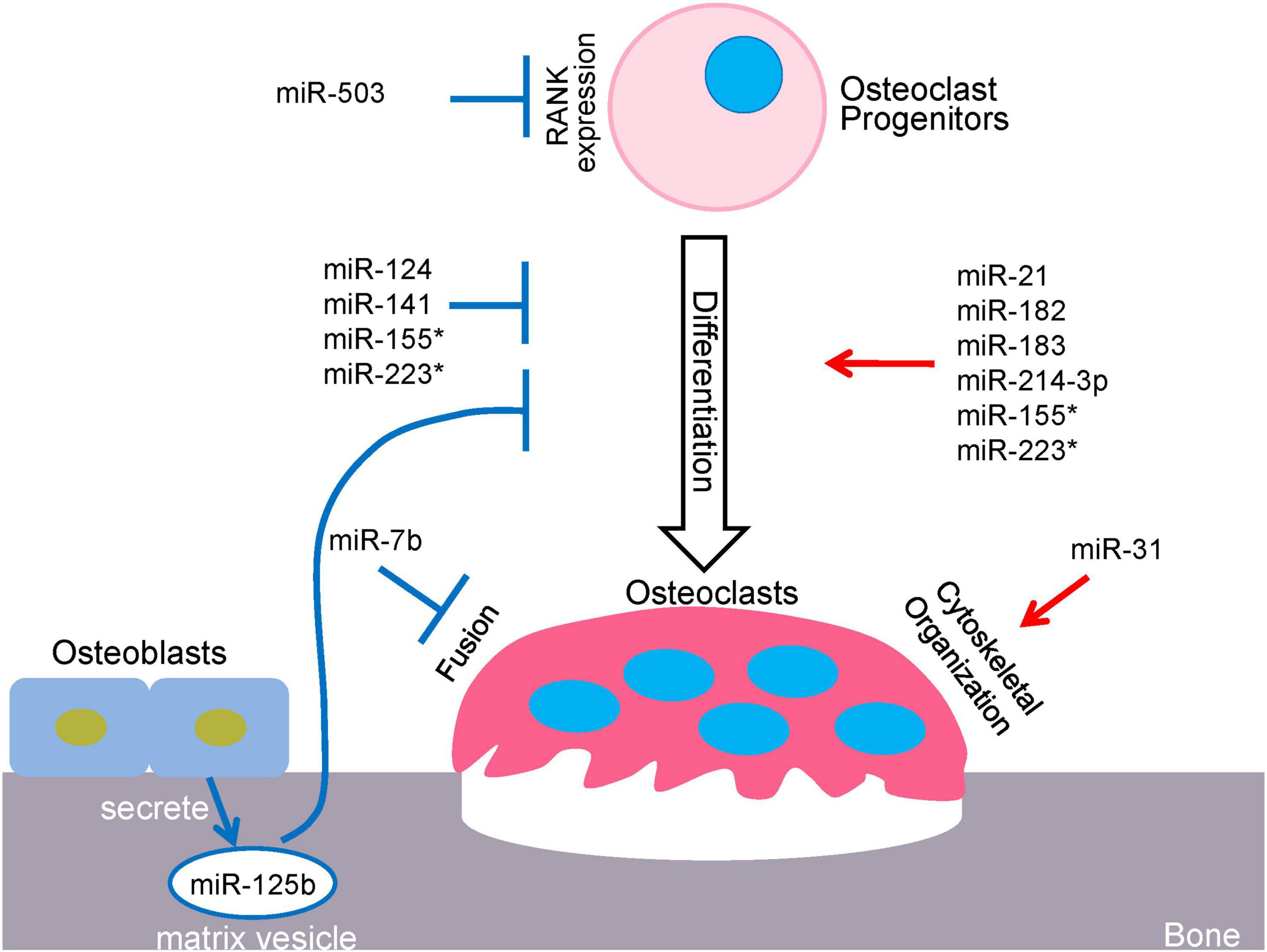
Figure 1. miRNA-mediated regulatory network in osteoclastogenesis. *miR-155 and miR223 play dual roles in osteoclastogenesis.
miRNAs That Positively Regulate Osteoclastogenesis
miR-21
In mammalian cells, miR-21 is most widely recognized as an oncogenic miRNA and its upregulation is associated to the progression of various cancers (Feng and Tsao, 2016). In addition, miR-21 is induced by inflammatory cytokines and plays regulatory roles in various cells. For example, miR-21 is induced by tumor necrosis factor-α (TNF-α) in endothelial cells (Yang D. et al., 2018), by interleukin-1β (IL-1β) in chondrocytes (Ma et al., 2020) and by interleukin-6 (IL-6) in myeloma cells (Loffler et al., 2007). Recently, miR-21 has gained recognition within the processes of bone metabolism and disease. In vitro studies have shown that RANKL induces miR-21 expression and promotes osteoclastogenesis via down-regulation of programmed cell death 4 (PDCD4), which subsequently regulates the c-Fos/NFATc1 axis (Sugatani et al., 2011). The pro-osteoclastic function of miR-21 is further demonstrated in vivo using miR-21 knockout mice, in which miR-21 deficiency inhibits bone resorption and osteoclastogenesis via targeting PDCD4 (Hu et al., 2017). miR-21 is found to be downregulated by bisphosphonate in lung cancer patients with osteolytic bone metastasis (Zhao et al., 2020). Another study revealed that estrogen suppresses miR-21 biogenesis, leading to upregulation of Fas Ligand (FasL), which is a target of miR-21 in osteoclasts and induces osteoclast cell death in an autocrine manner (Sugatani and Hruska, 2013). Thus, miR-21 also contributes to osteoclast survival and estrogen-mediated osteoclastogenesis.
miR-31
miR-31 is induced by RANKL, and positively regulates cytoskeletal organization during osteoclastogenesis and bone resorption activity. It does so by targeting expression of RhoA to optimize actin ring formation in osteoclasts, which is crucial for both cytoskeletal organization and bone resorption (Mizoguchi et al., 2013). In addition, inflammatory cytokines can regulate miR-31 expression. For instance, miR-31 is induced by TNF-α in cord blood mononuclear cells (Kim et al., 2020), by IL-1β in HUVEC endothelial cells (Zhong et al., 2017) and by IL-6 in HuCC-T1 cholangiocarcinoma cell line (Ishigami et al., 2018).
miR-214
The crosstalk and intercellular interaction between osteoclasts and osteoblasts are necessary for maintaining bone homeostasis and regulating bone remodeling. This communication involves secretory factors or cell-cell contact signaling, such as via RANKL, OPG, TGF-β1, IGF-1, Sema4D, Sema3A, ephrinB2/EphB4, Cthrc1, and Wnt16 (Zhao et al., 2006; Cao, 2011; Negishi-Koga et al., 2011; Hayashi et al., 2012; Takeshita et al., 2013; Moverare-Skrtic et al., 2014), and can be facilitated by bone-derived exosomes (Xie et al., 2017). Extracellular vesicles, or exosomes, are essential in coordinating intercellular communication through delivery of proteins, lipids, and nucleic acids including DNA, mRNA and miRNA. Moreover, tumor-derived exosomes are crucial in promoting tumor microenvironment and metastasis (Becker et al., 2016). Recent evidence has identified a role of osteoclast-derived exosomal miR-214-3p in mediating inhibition of osteoclast-directed osteoblast formation (Li et al., 2016). Osteoclastic miR-214-3p was found in association with reduced bone formation, and elevated levels of miR-214-3p were found in whole serum, serum exosome and bone tissue from elderly women with fractures and OVX mice. Osteoclast-specific miR-214-3p overexpression mice (OC-miR-214-3p) exhibited elevated exosomal miR-214-3p in serum and reduced bone formation, and this was rescued by injection of antagomiR-214-3p delivered by osteoclast-targeted (D-Asp8)-liposome (Liang et al., 2015). In vitro evidence revealed that osteoclastic exosomal miR-214-3p can transfer to osteoblasts and inhibit osteoblast activity. Consistently, in vivo osteoclast-targeted inhibition of miR-214-3p was able to enhance bone formation in aging OVX mice. These results highlight the impact of exosomal miR-214-3p as a mediator for bone cell communication on bone disease and related crosstalk. Wang et al. also provided evidence supporting the inhibitory role of miR-214 on bone formation and osteoblast activity via direct targeting of ATF4, an essential transcription factor for osteoblastogenesis and function (Yang et al., 2004; Wang et al., 2013; Li et al., 2016).
Moreover, miR-214-3p was found to be upregulated during osteoclastogenesis and positively regulates the differentiation process through activating PI3K/Akt/NFATc1 pathway by directly targeting phosphatase and tensin homolog (PTEN) (Zhao et al., 2015). Inhibition of miR-214-3p attenuated, while overexpression of miR-214-3p promoted, in vitro osteoclastogenesis. OC-miR-214-3p transgenic mice exhibited reduced PTEN expression levels, enhanced osteoclast activity and reduced bone mass, proposing potential as a therapeutic target for osteoporosis. Previous studies have also shown that PTEN is a target of miR-214-3p, which promotes T cell activation and proliferation (Jindra et al., 2010). Upregulation of miR-214-3p was also observed in osteolytic bone metastasis of breast cancer, which was consistently associated with elevated osteoclast activity in nude mice xenografted with human breast cancer cells (Liu et al., 2017). Breast cancer cells are able to induce miR-214-3p expression and interestingly, genetic ablation of osteoclast-specific miR-214-3p prevented the development of osteolytic breast cancer metastasis. This study also revealed that mechanistically, miR-214-3p promotes bone resorption through direct targeting of TNF-α Receptor Associated Factor 3 (TRAF3), and osteoclast-targeted treatment of Traf3 3′UTR plasmid attenuated excessive bone resorption in OC-miR-214-3p mice. In addition, miR-214 is decreased by TNF-α in HT29 colorectal adenocarcinoma cells (Li et al., 2017), induced by IL-1β in primary chondrocytes (Xu et al., 2021) and induced by IL-6 in human colonocytes (Polytarchou et al., 2015). This regulation of miR-214 expression by inflammatory cytokines suggests potential roles for miR-214 in inflammatory bone metabolism. Collectively, these studies strongly support the potential of miR-214-3p as a therapeutic target through regulating both osteoblasts and osteoclasts to treat bone disorders such as osteoporosis and cancer bone metastasis.
miR-182/miR-183
The miR-183 cluster, an miR family comprised of miR-182, miR-183 and miR-96, is well recognized to be functionally involved in development and also highly expressed and implicated in cancers, neurological disorders, autoimmune diseases (Dambal et al., 2015), and more recently, innate immunity (Singaravelu et al., 2019). miR96 was not detected during osteoclastogenesis (Miller et al., 2016), while miR-183 expression was found to be elevated by RANKL and promoted in vitro RANKL-induced osteoclastogenesis via targeting of heme oxygenase-1 (HO-1) (Ke et al., 2015). Recent studies revealed the functions of miR-182 in cell cycle, cell growth, cancer progression, T lymphocyte expansion and Th17 function (Stittrich et al., 2010; Ichiyama et al., 2016; Yao et al., 2019). miR-182 and miR-183 are induced by IL-6 but not IL-1β in Th17 cells and contribute to Th17 cell pathogenicity (Ichiyama et al., 2016).
Through genome-wide miRNA expression profiling via high-throughput miRNA-sequencing, our group identified miR-182 as a TNF-α-induced miRNA in mouse BMMs during inflammatory osteoclastogenesis (Miller et al., 2016). We further elucidated the role of miR-182 in regulating bone metabolism in both physiological and pathological conditions, such as those which occur in osteoporosis and rheumatoid arthritis (RA) (Inoue et al., 2018). Postmenopausal osteoporosis and RA are characterized by pathological bone destruction resulting from osteolysis conducted by osteoclasts. miRNA-based therapeutics for these diseases appear promising, though underdeveloped. Through a series of subsequent studies, we identified miR-182 as a key osteoclastogenic regulator, provided evidence revealing a novel regulatory network mediated by miR-182 in osteoclastogenesis, imparted therapeutic implications of targeting miR-182 to promote osteoprotection, and highlighted significant correlation between miR-182 and RA.
Our findings on miR-182 reveal great significance to the understanding of its role in bone metabolism, and potential impact on therapeutics and drug development for the treatment of bone diseases. Utilizing complementary genetic approaches (myeloid-specific miR-182 KO and Tg mice), miR-182 was identified as a key positive regulator of osteoclastogenesis. OVX-induced osteoporosis, which serves as a model for postmenopausal osteoporosis, and inflammatory arthritis, which mimics RA, were the disease models applied in this study for investigating translational significance. Genetic ablation or pharmacological inhibition of miR-182 prevented pathological bone loss in both models, suggesting a key role for miR-182 in bone protection. To test the efficacy of therapeutically targeting miR-182, chitosan (CH) nanoparticles were utilized to effectively and safely deliver miR-182 inhibitors to bone marrow and specifically target osteoclast lineage. The treatment with miR-182 inhibitors dramatically suppressed excessive osteoclast formation and bone resorption in these models without immune suppression, thus providing proof-of-principle that miR-182 inhibition presents significant clinical value for treating bone destruction. We also revealed and elucidated the miR-182-PKR-IFN-β axis through the identification of protein kinase double-stranded RNA-dependent (PKR), which is a crucial target of miR-182, and a novel inhibitor of osteoclastogenesis. PKR attenuates osteoclastogenesis by upregulating the endogenous IFN-β-mediated autocrine feedback loop. For the first time, a miR-182-PKR axis is revealed to be responsible for suppressing this IFN-β signaling induced by RANKL to activate osteoclastogenesis, and this network is involved in fine tuning the osteoclastogenic program. In the context of disease, the miR-182-PKR-IFN-β axis exhibits aberrant expression level changes including elevation of miR-182, and downregulation of PKR and IFN-β levels in the PBMCs from RA patients in comparison to healthy donors. Given the role of TNF-α in the pathogenesis of RA, TNF-α blockade therapy (TNFi) with Enbrel suppressed miR-182 expression levels and elevated PKR and IFN-β expression levels, and decreased osteoclastogenesis in RA patients-derived PBMCs. The osteoclastogenic capacity of PBMCs from RA patients is significantly correlated with the expression pattern of the miR-182-PKR-IFN-β axis. Both murine and human data provide strong evidence supporting the role of miR-182 in osteoclastogenesis, and translational promise of targeting miR-182 to treat pathological bone destruction.
miRNAs That Negatively Regulate Osteoclastogenesis
miR-7b
miR-7b has been reported to participate in the regulation of B cell receptor (BCR) signaling, which is enhanced in autoreactive B cells from systemic lupus erythematosus (SLE) (Wu et al., 2014). B cell hyper-responsiveness in SLE results from the downregulation of Phosphatase and tensin homolog (PTEN) by upregulated miR-7 (Wu et al., 2014). miR-7b is induced by a cocktail of IL-1β, IFN-γ, and TNF-α in the MIN6 insulinoma cell line (Bravo-Egana et al., 2012), suggesting its involvement in inflammation.
Within the process of osteoclastogenesis, maturation of multi-nucleated osteoclasts requires cell-cell fusion of mononuclear osteoclast precursors. Bone disease, such as osteoporosis can result from deregulated osteoclastic bone-resorbing activity when osteoclast multinucleation is defected. The role of miR-7b was revealed through its direct targeting of dendritic cell-specific transmembrane protein (DC-STAMP), which is a critical mediator of the osteoclast fusion process (Yagi et al., 2005; Dou et al., 2014). Positive regulation of DC-STAMP expression is mediated by NFATc1, c-Fos, and strawberry notch homolog 2 (Sbno2) (Maruyama et al., 2013). At the post-transcriptional level, miR-7b negatively regulates DC-STAMP, resulting in the inhibition of osteoclast fusion. Overexpression of miR-7b represses the expression of other fusogenic genes such as CD47, ATP6v0d2 and OC-STAMP, as well as osteoclast specific genes such as Nfatc1 and OSCAR via DC-STAMP inhibition. These studies characterize the crucial role of miR-7b in osteoclast multinucleation and function.
miR-124
miR-124 is, notably, abundantly expressed in neuronal cells, promoting cell differentiation (Lagos-Quintana et al., 2002; Makeyev et al., 2007), and also known as a critical regulator of immune function and implicated in inflammatory disorders (Qin et al., 2016). Expression of miR-124 is inversely correlated with activation of microglia during inflammation such as experimental autoimmune encephalomyelitis (EAE) (Ponomarev et al., 2011). Overexpression of miR-124 was found to promote microglia quiescence through directly targeting C/EBP-α and its downstream target, PU.1. This deactivation of macrophages suppresses CNS inflammation and inhibits EAE. In addition, inflammatory cytokines regulate the expression of miR-124. For example, TNF-α in keratinocytes (Yang et al., 2017) or IL-1β in lumbar (Willemen et al., 2012) was reported to decrease miR-124.
In addition to its effect on macrophage phenotypes in the CNS, miR-124 was shown to inhibit osteoclastogenesis in mouse BMMs by suppressing the expression of NFATc1 (Lee et al., 2013). miR-124 is downregulated in BMMs and osteoclasts from OVX mice, resulting in excessive bone resorption and destruction (Tang et al., 2017). This study also revealed a functional target of miR-124, Rab27a, which regulates multinucleation and transport of lysosome-related organelles (Shimada-Sugawara et al., 2015). Furthermore, miR-124 was reported to be downregulated in the ankles of adjuvant-induced arthritis (AIA) rats (Nakamachi et al., 2016). Administration of pre-miR-124 via injection into the ankles of AIA rats strongly suppressed bone destruction and reduced osteoclastogenesis via direct targeting of NFATc1. These findings support the novel role of miR-124 as a suppressor of osteoclastogenesis.
miR-141
miR-141 is identified as a tumor inhibitor, as well as a significant regulator participating in tumor-induced osteolytic bone metastasis. Conditioned medium (CM) from bone metastatic cell lines such as 4T1.2 and TSU-Pr-B2 cells were shown to be sufficient in inducing osteoclastogenesis, and miRNA microarray analysis revealed significant downregulation of miR-141 in osteoclasts under the treatment (Ell et al., 2013). Overexpression of miR-141 suppressed osteoclast differentiation via targeting of MITF and calcitonin receptor (CALCR), with observed downregulation of osteoclast marker expression such as NFATc1, CtsK, and PU.1. Systemic treatment of miR-141 was able to inhibit osteoclastogenesis and activity in vivo, as well as suppress osteolytic activity of breast cancer bone metastasis in vivo, accompanied with reduced osteoclast differentiation and activity. Further support of the inhibitory role of miR-141 on osteoclastogenesis and bone resorbing activity was established through the study of aged osteoporotic rhesus monkeys conducted by Yang S. et al. (2018). Both osteoporotic patients and aged rhesus monkeys exhibited downregulated expression of miR-141 in bone tissues. Targeted delivery of miR-141 to osteoclasts using Asp (Aspartic acid) 8-PU (polyurethane) nanoparticles inhibited bone loss, osteoclast differentiation and bone resorption activity in vivo, as well as in in vitro culture of rhesus monkey osteoclasts. They also demonstrated that miR-141 functionally targets Calcr and ephrin type-A receptor 2 precursor (EphA2) to inhibit osteoclastogenesis. Collectively, these data present the critical role of miR-141 in suppression of osteoclastogenesis and bone resorption, implicated in aberrant bone loss in bone disease and cancer.
miR-503
miR-503 has been identified in numerous cancer types via miRNA expression profiling analyses, including human retinoblastoma, glioblastoma, hepatocellular carcinomaand non-small cell lung cancer (NSLC), many of which support a tumor-suppressive role of miR-503 (Zhao J.J. et al., 2009; Zhou and Wang, 2011; Yang et al., 2014; Zhang et al., 2014). It has also been demonstrated that miR-503 can induce G1 cell cycle arrest to promote myogenic differentiation and similarly through promoting cell cycle quiescence, inhibit progression of osteosarcoma and colon cancer cell proliferation (Sarkar et al., 2010; Chong et al., 2014; Chang et al., 2015). In contrast, miR-503 expression has been reported to be upregulated in certain cancers and identified as a tumor-associated miRNA that directly targets Bcl2 apoptosis regulator to promote apoptosis of dendritic cells, indicating a mechanism for tumor immunotolerance (Min et al., 2013).
In the context of bone disorders, miR-503 was identified to be dramatically downregulated in CD14+ PBMCs of postmenopausal osteoporotic patients in comparison to postmenopausal healthy women. Following discontinuation of denosumab (an anti-RANKL antibody) treatment, miR-503 expression is decreased in patients with vertebral fractures (Anastasilakis et al., 2017). These findings suggest a crucial role for miR-503 in bone metabolism (Chen et al., 2014). Overexpression and silencing of miR-503 in CD14+ PBMCs confirmed its inhibitory role on osteoclastogenesis and its direct targeting of RANK, the receptor for RANKL. Treatment with agomiR-503 protected OVX mice from bone loss by inhibiting bone resorption and increased bone mass. Consistently, estrogen upregulated miR-503 expression in vitro and in vivo, confirming its contribution to the pathogenesis of postmenopausal osteoporosis and potential as a therapeutic target.
miR-125b
Bone matrix growth factors, such as TGF-β and IGF-1, play essential roles in coupling bone resorption and formation (Tang et al., 2009; Xian et al., 2012). Recently, miR-125b has been identified as a bone matrix-embedded coupling factor that is produced by osteoblasts (Minamizaki et al., 2020). miR-125b is enveloped by matrix vesicles (MVs) and accumulates in bone matrix. The osteoblast-derived MVs containing miR-125b are incorporated into osteoclast precursors and inhibit osteoclastogenesis. Mice overexpressing miR-125b in osteoblasts using human osteocalcin promoter show high bone mass phenotype and reduced number of bone-resorbing osteoclasts, without affecting osteoblasts and bone formation. miR-125b directly targets Prdm1, a key transcriptional factor of osteoclastogenesis, in osteoclast precursors. Overexpression of miR-125b in osteoblasts abrogates bone loss in OVX-induced osteoporosis mouse model, LPS-induced calvarial bone loss model, and sciatic neurectomy (NX)-induced osteoporosis mouse model. The MV containing miR-125b is a novel coupling factor of osteoblast-osteoclast communication. MVs containing miR-125b might be potential therapeutic targets for pathological bone loss.
miRNAs That Play Dual Roles in Osteoclastogenesis
miR-155
miR-155 plays crucial roles in cell development and function of various immune cells involved in both innate and adaptive immunity and is implicated in pathological conditions such as cancer (O’Connell et al., 2007, 2009; Thai et al., 2007; Vigorito et al., 2007; Teng et al., 2008). It has been shown that inflammatory cytokines and stimuli, such as TNF-α and lipopolysaccharide (LPS), can activate miR-155 expression, which in turn regulates proliferation, differentiation and function of macrophages and dendritic cells (O’Connell et al., 2007, 2009; Ceppi et al., 2009; Lind et al., 2015).
In addition to its role in immune cells, miR-155 has been revealed to inhibit RANKL-induced osteoclastogenesis through TGF-β1/Smad4 signaling and by mediating the suppressive effect of IFN-β on osteoclast differentiation through targeting microphthalmia-associated transcription factor (MITF), suppressor of cytokine signaling 1 (SOCS1), and PU.1 (Zhang et al., 2012; Zhao et al., 2017). However, in the context of LPS-mediated inflammatory bone loss, it has been recently reported that miR-155 is induced by LPS and enhances autophagy to increase osteoclast differentiation and activity through targeting transforming growth factor β-activated kinase 1-binding protein 2 (TAB2) (Sul et al., 2018). Taken together, miR-155 plays an inhibitory role in RANKL-induced osteoclastogenesis but enhances LPS-mediated inflammatory osteoclastogenesis.
miR-223
miR-223 was known to be preferentially expressed in hematopoietic cells and identified as a key regulator of myeloid cell differentiation and activation, particularly neutrophils and macrophages, and therefore, a regulator of innate immune response through myeloid cell homeostasis (Fukao et al., 2007; Yuan et al., 2018). Studies have demonstrated that miR-223 has the abilities to promote macrophage polarization to adopt anti-inflammatory phenotype and inhibit activation of macrophages and subsequent inflammatory response through targeting signal transducer and activator of transcription 3 (STAT3) (Chen et al., 2012) and inflammasome sensor-NLRP3 (Bauernfeind et al., 2012). In contrast, miR-223 expression is downregulated under inflammatory conditions activated by Toll-like receptor (TLR) ligands, such as LPS in macrophages (Chen et al., 2012; Zhang et al., 2017). However, elevated expression of miR-223 in CD68+ macrophages, CD14+ monocytes, and CD4+ T cells from synovium of rheumatoid arthritis (RA) patients was reported (Li et al., 2012; Shibuya et al., 2013). miR-223 expression is also induced by TNF-α in MH7A fibroblast-like synoviocyte cell line (Moriya et al., 2017). In osteoclast precursor cells, overexpression of miR-223 was shown to downregulate osteoclastogenesis (Sugatani and Hruska, 2007; Kagiya and Nakamura, 2013). However, conflicting studies have reported that in osteoclast precursor cells, a positive feedback loop exists whereby M-CSF induces transcription factor, PU.1, which stimulates miR-223 expression, resulting in downregulation of its target, nuclear factor I A (NFIA). The downregulation of NFIA expression allows for upregulation of M-CSF receptor levels and consequently, promotion of osteoclastogenesis (Sugatani and Hruska, 2009; Xie et al., 2015). PU.1, c-Fos, MITF, and NFATc1 levels are upregulated by M-CSF and RANKL during osteoclastogenesis. These results suggest that miR-223 is essential for osteoclast differentiation and function. However, the mechanisms underlying the dual effects of miR-223 on osteoclastogenesis are not well-understood.
Closing Remark
miRNAs are involved in numerous molecular pathways and cellular events through targeting specific genes. Recent studies have identified competing endogenous RNAs (ceRNAs), which comprise of mRNAs, pseudogenes, long non-coding RNAs (lncRNAs) and circular RNAs (circRNA) (Cesana et al., 2011; Salmena et al., 2011; Tay et al., 2014). These ceRNAs communicate with each other to compete for shared miRNAs, acting as natural miRNA “sponges,” to block miRNA binding and repression of its targets. ceRNAs impose a regulatory mechanism upon miRNAs, adding another level of complexity to miRNA-mediated gene regulation. The mechanisms of miRNA-ceRNA networks is another channel to unpack in further exploration and understanding of gene regulation by different RNA species in specific processes, pathways and tissues, which are especially important for understanding their translational implication.
Targeting miRNAs has shown promising therapeutic potential in a few clinical trials, such as in the treatment of diabetes, cancer and hepatitis C (Janssen et al., 2013; Chakraborty et al., 2017; Rupaimoole and Slack, 2017). These inspiring scientific advances highlight the clinical significance of the emerging field of miRNA and miRNA-based therapeutics. The miRNAs discussed in this review strongly support the potential of therapeutic targeting of miRNAs to prevent and treat pathological bone loss, such as that occurring in RA, osteoporosis and bone cancer metastasis. However, further work is needed in elucidating miRNA-mediated gene regulation networks specific to these diseases in order to successfully enhance the development of therapeutic approaches. In addition, generating an ideal miRNA-based drug delivery method presents challenges, such as ensuring stability, specificity and efficacy of targeting, and minimization of toxicities and off-target effects. Future studies and technological developments are expected to address these challenges in the clinical development of miRNA-based drugs.
Author Contributions
BZ designed, wrote, and edited the manuscript. KI wrote and edited the manuscript. CN and YX helped to writing and editing the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by grants from the National Institutes of Health (AR062047, AR068970, and AR071463 to BZ) and Tow Foundation (for the David Z. Rosensweig Genomics Center at the Hospital for Special Surgery).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We apologize to colleagues whose work could not be cited due to space limitation. The content of this manuscript is solely the responsibilities of the authors and does not necessarily represent the official views of the NIH.
References
Anastasilakis, A. D., Yavropoulou, M. P., Makras, P., Sakellariou, G. T., Papadopoulou, F., Gerou, S., et al. (2017). Increased osteoclastogenesis in patients with vertebral fractures following discontinuation of denosumab treatment. Eur. J. Endocrinol. 176, 677–683. doi: 10.1530/EJE-16-1027
Asagiri, M., and Takayanagi, H. (2007). The molecular understanding of osteoclast differentiation. Bone 40, 251–264. doi: 10.1016/j.bone.2006.09.023
Bartel, D. P. (2009). MicroRNAs: target recognition and regulatory functions. Cell 136, 215–233. doi: 10.1016/j.cell.2009.01.002
Bauernfeind, F., Rieger, A., Schildberg, F. A., Knolle, P. A., Schmid-Burgk, J. L., and Hornung, V. (2012). NLRP3 inflammasome activity is negatively controlled by miR-223. J. Immunol. 189, 4175–4181. doi: 10.4049/jimmunol.1201516
Becker, A., Thakur, B. K., Weiss, J. M., Kim, H. S., Peinado, H., and Lyden, D. (2016). Extracellular vesicles in Cancer: cell-to-cell mediators of metastasis. Cancer Cell 30, 836–848. doi: 10.1016/j.ccell.2016.10.009
Binder, N., Miller, C., Yoshida, M., Inoue, K., Nakano, S., Hu, X., et al. (2017). Def6 restrains osteoclastogenesis and inflammatory bone resorption. J. Immunol. 198, 3436–3447. doi: 10.4049/jimmunol.1601716
Boyce, B. F., Xiu, Y., Li, J., Xing, L., and Yao, Z. (2015). NF-kappaB-Mediated regulation of osteoclastogenesis. Endocrinol. Metab. 30, 35–44. doi: 10.3803/EnM.2015.30.1.35
Bravo-Egana, V., Rosero, S., Klein, D., Jiang, Z., Vargas, N., Tsinoremas, N., et al. (2012). Inflammation-Mediated regulation of MicroRNA expression in transplanted pancreatic islets. J. Transplant 2012:723614. doi: 10.1155/2012/723614
Cao, X. (2011). Targeting osteoclast-osteoblast communication. Nat Med 17, 1344–1346. doi: 10.1038/nm.2499
Ceppi, M., Pereira, P. M., Dunand-Sauthier, I., Barras, E., Reith, W., Santos, M. A., et al. (2009). MicroRNA-155 modulates the interleukin-1 signaling pathway in activated human monocyte-derived dendritic cells. Proc. Natl. Acad. Sci. U S A. 106, 2735–2740. doi: 10.1073/pnas.0811073106
Cesana, M., Cacchiarelli, D., Legnini, I., Santini, T., Sthandier, O., Chinappi, M., et al. (2011). A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell 147, 358–369. doi: 10.1016/j.cell.2011.09.028
Chakraborty, C., Sharma, A. R., Sharma, G., Doss, C. G. P., and Lee, S. S. (2017). Therapeutic miRNA and siRNA: moving from bench to clinic as next generation medicine. Mol. Ther. Nucleic Acids 8, 132–143. doi: 10.1016/j.omtn.2017.06.005
Chang, S. W., Yue, J., Wang, B. C., and Zhang, X. L. (2015). miR-503 inhibits cell proliferation and induces apoptosis in colorectal cancer cells by targeting E2F3. Int. J. Clin. Exp. Pathol. 8, 12853–12860.
Chen, C., Cheng, P., Xie, H., Zhou, H. D., Wu, X. P., Liao, E. Y., et al. (2014). MiR-503 regulates osteoclastogenesis via targeting RANK. J. Bone Miner. Res. 29, 338–347. doi: 10.1002/jbmr.2032
Chen, Q., Wang, H., Liu, Y., Song, Y., Lai, L., Han, Q., et al. (2012). Inducible microRNA-223 down-regulation promotes TLR-triggered IL-6 and IL-1beta production in macrophages by targeting STAT3. PLoS One 7:e42971. doi: 10.1371/journal.pone.0042971
Chendrimada, T. P., Gregory, R. I., Kumaraswamy, E., Norman, J., Cooch, N., Nishikura, K., et al. (2005). TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature 436, 740–744. doi: 10.1038/nature03868
Chong, Y., Zhang, J., Guo, X., Li, G., Zhang, S., Li, C., et al. (2014). MicroRNA-503 acts as a tumor suppressor in osteosarcoma by targeting L1CAM. PLoS One 9:e114585. doi: 10.1371/journal.pone.0114585
Dambal, S., Shah, M., Mihelich, B., and Nonn, L. (2015). The microRNA-183 cluster: the family that plays together stays together. Nucleic Acids Res. 43, 7173–7188. doi: 10.1093/nar/gkv703
Dou, C., Zhang, C., Kang, F., Yang, X., Jiang, H., Bai, Y., et al. (2014). MiR-7b directly targets DC-STAMP causing suppression of NFATc1 and c-Fos signaling during osteoclast fusion and differentiation. Biochimica et Biophysica Acta 1839, 1084–1096. doi: 10.1016/j.bbagrm.2014.08.002
Ell, B., Mercatali, L., Ibrahim, T., Campbell, N., Schwarzenbach, H., Pantel, K., et al. (2013). Tumor-Induced osteoclast miRNA changes as regulators and biomarkers of osteolytic bone metastasis. Cancer Cell 24, 542–556. doi: 10.1016/j.ccr.2013.09.008
Esteller, M. (2011). Non-coding RNAs in human disease. Nat. Rev. Genet. 12, 861–874. doi: 10.1038/nrg3074
Feng, Y. H., and Tsao, C. J. (2016). Emerging role of microRNA-21 in cancer. Biomed. Rep. 5, 395–402. doi: 10.3892/br.2016.747
Fukao, T., Fukuda, Y., Kiga, K., Sharif, J., Hino, K., Enomoto, Y., et al. (2007). An evolutionarily conserved mechanism for MicroRNA-223 expression revealed by MicroRNA gene profiling. Cell 129, 617–631. doi: 10.1016/j.cell.2007.02.048
Goldring, S. R., Purdue, P. E., Crotti, T. N., Shen, Z., Flannery, M. R., Binder, N. B., et al. (2013). Bone remodelling in inflammatory arthritis. Annals Rheumatic Dis. 72, (Suppl. 2), ii52–ii55. doi: 10.1136/annrheumdis-2012-202199
Gregory, R. I., Yan, K. P., Amuthan, G., Chendrimada, T., Doratotaj, B., Cooch, N., et al. (2004). The microprocessor complex mediates the genesis of microRNAs. Nature 432, 235–240. doi: 10.1038/nature03120
Hangauer, M. J., Vaughn, I. W., and McManus, M. T. (2013). Pervasive transcription of the human genome produces thousands of previously unidentified long intergenic noncoding RNAs. PLoS Genet. 9:e1003569. doi: 10.1371/journal.pgen.1003569
Hayashi, M., Nakashima, T., Taniguchi, M., Kodama, T., Kumanogoh, A., and Takayanagi, H. (2012). Osteoprotection by semaphorin 3A. Nature 485, 69–74. doi: 10.1038/nature11000
Hu, C. H., Sui, B. D., Du, F. Y., Shuai, Y., Zheng, C. X., Zhao, P., et al. (2017). miR-21 deficiency inhibits osteoclast function and prevents bone loss in mice. Sci. Rep. 7:43191. doi: 10.1038/srep43191
Humphrey, M. B., Lanier, L. L., and Nakamura, M. C. (2005). Role of ITAM-containing adapter proteins and their receptors in the immune system and bone. Immunol. Rev. 208, 50–65.
Ichiyama, K., Gonzalez-Martin, A., Kim, B. S., Jin, H. Y., Jin, W., Xu, W., et al. (2016). The MicroRNA-183-96-182 cluster promotes T Helper 17 cell pathogenicity by negatively regulating transcription factor foxo1 expression. Immunity 44, 1284–1298. doi: 10.1016/j.immuni.2016.05.015
Inoue, K., Deng, Z., Chen, Y., Giannopoulou, E., Xu, R., Gong, S., et al. (2018). Bone protection by inhibition of microRNA-182. Nat. Commun. 9:4108. doi: 10.1038/s41467-018-06446-0
Ishigami, K., Nosho, K., Koide, H., Kanno, S., Mitsuhashi, K., Igarashi, H., et al. (2018). MicroRNA-31 reflects IL-6 expression in cancer tissue and is related with poor prognosis in bile duct cancer. Carcinogenesis 39, 1127–1134. doi: 10.1093/carcin/bgy075
Janssen, H. L., Reesink, H. W., Lawitz, E. J., Zeuzem, S., Rodriguez-Torres, M., Patel, K., et al. (2013). Treatment of HCV infection by targeting microRNA. N. Engl. J. Med. 368, 1685–1694. doi: 10.1056/NEJMoa1209026
Jindra, P. T., Bagley, J., Godwin, J. G., and Iacomini, J. (2010). Costimulation-dependent expression of microRNA-214 increases the ability of T cells to proliferate by targeting Pten. J. Immunol. 185, 990–997. doi: 10.4049/jimmunol.1000793
Kagiya, T., and Nakamura, S. (2013). Expression profiling of microRNAs in RAW264.7 cells treated with a combination of tumor necrosis factor alpha and RANKL during osteoclast differentiation. J. Periodontal Res. 48, 373–385. doi: 10.1111/jre.12017
Ke, K., Sul, O. J., Rajasekaran, M., and Choi, H. S. (2015). MicroRNA-183 increases osteoclastogenesis by repressing heme oxygenase-1. Bone 81, 237–246. doi: 10.1016/j.bone.2015.07.006
Kim, J. H., Kim, J. Y., Park, M., Kim, S., Kim, T., Kim, J., et al. (2020). NF-kappaB-dependent miR-31/155 biogenesis is essential for TNF-alpha-induced impairment of endothelial progenitor cell function. Exp. Mol. Med. 52, 1298–1309. doi: 10.1038/s12276-020-0478-x
Kim, T. K., Hemberg, M., Gray, J. M., Costa, A. M., Bear, D. M., Wu, J., et al. (2010). Widespread transcription at neuronal activity-regulated enhancers. Nature 465, 182–187. doi: 10.1038/nature09033
Kim, V. N., Han, J., and Siomi, M. C. (2009). Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 10, 126–139. doi: 10.1038/nrm2632
Lagos-Quintana, M., Rauhut, R., Lendeckel, W., and Tuschl, T. (2001). Identification of novel genes coding for small expressed RNAs. Science 294, 853–858. doi: 10.1126/science.1064921
Lagos-Quintana, M., Rauhut, R., Yalcin, A., Meyer, J., Lendeckel, W., and Tuschl, T. (2002). Identification of tissue-specific microRNAs from mouse. Curr. Biol. 12, 735–739. doi: 10.1016/s0960-9822(02)00809-6
Lam, M. T., Li, W., Rosenfeld, M. G., and Glass, C. K. (2014). Enhancer RNAs and regulated transcriptional programs. Trends Biochem. Sci. 39, 170–182. doi: 10.1016/j.tibs.2014.02.007
Lee, S. H., Rho, J., Jeong, D., Sul, J. Y., Kim, T., Kim, N., et al. (2006). v-ATPase V0 subunit d2-deficient mice exhibit impaired osteoclast fusion and increased bone formation. Nat. Med. 12, 1403–1409. doi: 10.1038/nm1514
Lee, Y., Ahn, C., Han, J., Choi, H., Kim, J., Yim, J., et al. (2003). The nuclear RNase III Drosha initiates microRNA processing. Nature 425, 415–419. doi: 10.1038/nature01957
Lee, Y., Kim, H. J., Park, C. K., Kim, Y. G., Lee, H. J., Kim, J. Y., et al. (2013). MicroRNA-124 regulates osteoclast differentiation. Bone 56, 383–389. doi: 10.1016/j.bone.2013.07.007
Lee, Y., Kim, M., Han, J., Yeom, K. H., Lee, S., Baek, S. H., et al. (2004). MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 23, 4051–4060. doi: 10.1038/sj.emboj.7600385
Li, D., Liu, J., Guo, B., Liang, C., Dang, L., Lu, C., et al. (2016). Osteoclast-derived exosomal miR-214-3p inhibits osteoblastic bone formation. Nat. Commun. 7:10872. doi: 10.1038/ncomms10872
Li, J. A., Wang, Y. D., Wang, K., Wang, Z. L., Jia, D. Y., Yang, B. Y., et al. (2017). Downregulation of miR-214-3p may contribute to pathogenesis of ulcerative colitis via targeting STAT6. Biomed. Res. Int. 2017:8524972. doi: 10.1155/2017/8524972
Li, S., Miller, C. H., Giannopoulou, E., Hu, X., Ivashkiv, L. B., and Zhao, B. (2014). RBP-J imposes a requirement for ITAM-mediated costimulation of osteoclastogenesis. J. Clin. Invest. 124, 5057–5073. doi: 10.1172/JCI71882
Li, Y. T., Chen, S. Y., Wang, C. R., Liu, M. F., Lin, C. C., Jou, I. M., et al. (2012). Amelioration of collagen-induced arthritis in mice by lentivirus-mediated silencing of microRNA-223. Arthritis Rheumatism. 64, 3240–3245. doi: 10.1002/art.34550
Liang, C., Guo, B., Wu, H., Shao, N., Li, D., Liu, J., et al. (2015). Aptamer-functionalized lipid nanoparticles targeting osteoblasts as a novel RNA interference-based bone anabolic strategy. Nat. Med. 21, 288–294. doi: 10.1038/nm.3791
Lind, E. F., Millar, D. G., Dissanayake, D., Savage, J. C., Grimshaw, N. K., Kerr, W. G., et al. (2015). miR-155 upregulation in dendritic cells is sufficient to break tolerance in vivo by negatively regulating SHIP1. J. Immunol. 195, 4632–4640. doi: 10.4049/jimmunol.1302941
Ling, H., Fabbri, M., and Calin, G. A. (2013). MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat. Rev. Drug Discovery 12, 847–865. doi: 10.1038/nrd4140
Liu, J., Li, D., Dang, L., Liang, C., Guo, B., Lu, C., et al. (2017). Osteoclastic miR-214 targets TRAF3 to contribute to osteolytic bone metastasis of breast cancer. Sci. Rep. 7:40487. doi: 10.1038/srep40487
Loffler, D., Brocke-Heidrich, K., Pfeifer, G., Stocsits, C., Hackermuller, J., Kretzschmar, A. K., et al. (2007). Interleukin-6 dependent survival of multiple myeloma cells involves the Stat3-mediated induction of microRNA-21 through a highly conserved enhancer. Blood 110, 1330–1333. doi: 10.1182/blood-2007-03-081133
Lund, E., Guttinger, S., Calado, A., Dahlberg, J. E., and Kutay, U. (2004). Nuclear export of microRNA precursors. Science 303, 95–98. doi: 10.1126/science.1090599
Ma, S., Zhang, A., Li, X., Zhang, S., Liu, S., Zhao, H., et al. (2020). MiR-21-5p regulates extracellular matrix degradation and angiogenesis in TMJOA by targeting Spry1. Arthritis Res. Ther. 22:99. doi: 10.1186/s13075-020-2145-y
Makeyev, E. V., Zhang, J., Carrasco, M. A., and Maniatis, T. (2007). The MicroRNA miR-124 promotes neuronal differentiation by triggering brain-specific alternative pre-mRNA splicing. Mol. Cell 27, 435–448. doi: 10.1016/j.molcel.2007.07.015
Maruyama, K., Uematsu, S., Kondo, T., Takeuchi, O., Martino, M. M., Kawasaki, T., et al. (2013). Strawberry notch homologue 2 regulates osteoclast fusion by enhancing the expression of DC-STAMP. J. Exp. Med. 210, 1947–1960. doi: 10.1084/jem.20130512
Miller, C. H., Smith, S. M., Elguindy, M., Zhang, T., Xiang, J. Z., Hu, X., et al. (2016). RBP-J-Regulated miR-182 Promotes TNF-alpha-Induced Osteoclastogenesis. J. Immunol. 196, 4977–4986. doi: 10.4049/jimmunol.1502044
Min, S., Liang, X., Zhang, M., Zhang, Y., Mei, S., Liu, J., et al. (2013). Multiple tumor-associated microRNAs modulate the survival and longevity of dendritic cells by targeting YWHAZ and Bcl2 signaling pathways. J. Immunol. 190, 2437–2446. doi: 10.4049/jimmunol.1202282
Minamizaki, T., Nakao, Y., Irie, Y., Ahmed, F., Itoh, S., Sarmin, N., et al. (2020). The matrix vesicle cargo miR-125b accumulates in the bone matrix, inhibiting bone resorption in mice. Commun. Biol. 3:30. doi: 10.1038/s42003-020-0754-2
Mizoguchi, F., Murakami, Y., Saito, T., Miyasaka, N., and Kohsaka, H. (2013). miR-31 controls osteoclast formation and bone resorption by targeting RhoA. Arthritis Res. Ther. 15:R102. doi: 10.1186/ar4282
Moriya, N., Shibasaki, S., Karasaki, M., and Iwasaki, T. (2017). The impact of MicroRNA-223-3p on IL-17 receptor d expression in synovial cells. PLoS One 12:e0169702. doi: 10.1371/journal.pone.0169702
Moverare-Skrtic, S., Henning, P., Liu, X., Nagano, K., Saito, H., Borjesson, A. E., et al. (2014). Osteoblast-derived WNT16 represses osteoclastogenesis and prevents cortical bone fragility fractures. Nat. Med. 20, 1279–1288. doi: 10.1038/nm.3654
Nakamachi, Y., Ohnuma, K., Uto, K., Noguchi, Y., Saegusa, J., and Kawano, S. (2016). MicroRNA-124 inhibits the progression of adjuvant-induced arthritis in rats. Annals Rheumatic Dis. 75, 601–608. doi: 10.1136/annrheumdis-2014-206417
Nakano, S., Inoue, K., Xu, C., Deng, Z., Syrovatkina, V., Vitone, G., et al. (2019). G-protein Galpha13 functions as a cytoskeletal and mitochondrial regulator to restrain osteoclast function. Sci. Rep. 9:4236. doi: 10.1038/s41598-019-40974-z
Nakashima, T., and Takayanagi, H. (2011). New regulation mechanisms of osteoclast differentiation. Ann. N. Y. Acad. Sci. 1240, E13–E18.
Negishi-Koga, T., Shinohara, M., Komatsu, N., Bito, H., Kodama, T., Friedel, R. H., et al. (2011). Suppression of bone formation by osteoclastic expression of semaphorin 4D. Nat. Med. 17, 1473–1480. doi: 10.1038/nm.2489
O’Connell, R. M., Chaudhuri, A. A., Rao, D. S., and Baltimore, D. (2009). Inositol phosphatase SHIP1 is a primary target of miR-155. Proc. Natl. Acad. Sci. U S A. 106, 7113–7118. doi: 10.1073/pnas.0902636106
O’Connell, R. M., Taganov, K. D., Boldin, M. P., Cheng, G., and Baltimore, D. (2007). MicroRNA-155 is induced during the macrophage inflammatory response. Proc. Natl. Acad. Sci. U S A. 104, 1604–1609. doi: 10.1073/pnas.0610731104
Park, J. E., Heo, I., Tian, Y., Simanshu, D. K., Chang, H., Jee, D., et al. (2011). Dicer recognizes the 5′ end of RNA for efficient and accurate processing. Nature 475, 201–205. doi: 10.1038/nature10198
Peters, L., and Meister, G. (2007). Argonaute proteins: mediators of RNA silencing. Mol. Cell 26, 611–623. doi: 10.1016/j.molcel.2007.05.001
Polytarchou, C., Hommes, D. W., Palumbo, T., Hatziapostolou, M., Koutsioumpa, M., Koukos, G., et al. (2015). MicroRNA214 is associated with progression of ulcerative colitis, and inhibition reduces development of colitis and colitis-associated cancer in mice. Gastroenterology 149, 981–992.e11. doi: 10.1053/j.gastro.2015.05.057
Ponomarev, Veremeyko, T., Barteneva, N., Krichevsky, A. M., and Weiner, H. L. (2011). MicroRNA-124 promotes microglia quiescence and suppresses EAE by deactivating macrophages via the C/EBP-alpha-PU.1 pathway. Nat. Med. 17, 64–70. doi: 10.1038/nm.2266
Qin, Z., Wang, P. Y., Su, D. F., and Liu, X. (2016). miRNA-124 in immune system and immune disorders. Front. Immunol. 7:406. doi: 10.3389/fimmu.2016.00406
Rupaimoole, R., and Slack, F. J. (2017). MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discovery 16, 203–222. doi: 10.1038/nrd.2016.246
Sakaguchi, Y., Nishikawa, K., Seno, S., Matsuda, H., Takayanagi, H., and Ishii, M. (2018). Roles of Enhancer RNAs in RANKL-induced osteoclast differentiation identified by genome-wide cap-analysis of gene expression using CRISPR/Cas9. Sci. Rep. 8:7504. doi: 10.1038/s41598-018-25748-3
Salmena, L., Poliseno, L., Tay, Y., Kats, L., and Pandolfi, P. P. (2011). A ceRNA hypothesis: the rosetta stone of a hidden RNA language? Cell 146, 353–358. doi: 10.1016/j.cell.2011.07.014
Sarkar, S., Dey, B. K., and Dutta, A. (2010). MiR-322/424 and -503 are induced during muscle differentiation and promote cell cycle quiescence and differentiation by down-regulation of Cdc25A. Mol. Biol. Cell 21, 2138–2149. doi: 10.1091/mbc.E10-01-0062
Schett, G., and Gravallese, E. (2012). Bone erosion in rheumatoid arthritis: mechanisms, diagnosis and treatment. Nat. Rev. Rheumatol. 8, 656–664. doi: 10.1038/nrrheum.2012.153
Shibuya, H., Nakasa, T., Adachi, N., Nagata, Y., Ishikawa, M., Deie, M., et al. (2013). Overexpression of microRNA-223 in rheumatoid arthritis synovium controls osteoclast differentiation. Modern Rheumatol. 23, 674–685. doi: 10.1007/s10165-012-0710-1
Shimada-Sugawara, M., Sakai, E., Okamoto, K., Fukuda, M., Izumi, T., Yoshida, N., et al. (2015). Rab27A regulates transport of cell surface receptors modulating multinucleation and lysosome-related organelles in osteoclasts. Sci. Rep. 5:9620. doi: 10.1038/srep09620
Singaravelu, R., Ahmed, N., Quan, C., Srinivasan, P., Ablenas, C. J., Roy, D. G., et al. (2019). A conserved miRNA-183 cluster regulates the innate antiviral response. J. Biol. Chem. 294, 19785–19794. doi: 10.1074/jbc.RA119.010858
Stittrich, A. B., Haftmann, C., Sgouroudis, E., Kuhl, A. A., Hegazy, A. N., Panse, I., et al. (2010). The microRNA miR-182 is induced by IL-2 and promotes clonal expansion of activated helper T lymphocytes. Nat. Immunol. 11, 1057–1062. doi: 10.1038/ni.1945
Sugatani, T., and Hruska, K. A. (2007). MicroRNA-223 is a key factor in osteoclast differentiation. J. Cell. Biochem. 101, 996–999. doi: 10.1002/jcb.21335
Sugatani, T., and Hruska, K. A. (2009). Impaired micro-RNA pathways diminish osteoclast differentiation and function. J. Biol. Chem. 284, 4667–4678. doi: 10.1074/jbc.M805777200
Sugatani, T., and Hruska, K. A. (2013). Down-regulation of miR-21 biogenesis by estrogen action contributes to osteoclastic apoptosis. J. Cell. Biochem. 114, 1217–1222. doi: 10.1002/jcb.24471
Sugatani, T., Vacher, J., and Hruska, K. A. (2011). A microRNA expression signature of osteoclastogenesis. Blood 117, 3648–3657.
Sul, O. J., Sung, Y. B., Rajasekaran, M., Ke, K., Yu, R., Back, S. H., et al. (2018). MicroRNA-155 induces autophagy in osteoclasts by targeting transforming growth factor beta-activated kinase 1-binding protein 2 upon lipopolysaccharide stimulation. Bone 116, 279–289. doi: 10.1016/j.bone.2018.08.014
Takeshita, S., Fumoto, T., Matsuoka, K., Park, K. A., Aburatani, H., Kato, S., et al. (2013). Osteoclast-secreted CTHRC1 in the coupling of bone resorption to formation. J. Clin. Investigation 123, 3914–3924. doi: 10.1172/JCI69493
Tang, L., Yin, Y., Liu, J., Li, Z., and Lu, X. (2017). MiR-124 attenuates osteoclastogenic differentiation of bone marrow monocytes via targeting Rab27a. Cell Physiol. Biochem. 43, 1663–1672. doi: 10.1159/000484027
Tang, Y., Wu, X., Lei, W., Pang, L., Wan, C., Shi, Z., et al. (2009). TGF-beta1-induced migration of bone mesenchymal stem cells couples bone resorption with formation. Nat. Med. 15, 757–765. doi: 10.1038/nm.1979
Tay, Y., Rinn, J., and Pandolfi, P. P. (2014). The multilayered complexity of ceRNA crosstalk and competition. Nature 505, 344–352. doi: 10.1038/nature12986
Teng, G., Hakimpour, P., Landgraf, P., Rice, A., Tuschl, T., Casellas, R., et al. (2008). MicroRNA-155 is a negative regulator of activation-induced cytidine deaminase. Immunity 28, 621–629. doi: 10.1016/j.immuni.2008.03.015
Thai, T.-H., Calado, D. P., Casola, S., Ansel, K. M., Xiao, C., Xue, Y., et al. (2007). Regulation of the germinal center response by MicroRNA-155. Science 316, 604–608.
Vigorito, E., Perks, K. L., Abreu-Goodger, C., Bunting, S., Xiang, Z., Kohlhaas, S., et al. (2007). microRNA-155 regulates the generation of immunoglobulin class-switched plasma cells. Immunity 27, 847–859. doi: 10.1016/j.immuni.2007.10.009
Wang, X., Guo, B., Li, Q., Peng, J., Yang, Z., Wang, A., et al. (2013). miR-214 targets ATF4 to inhibit bone formation. Nat. Med. 19, 93–100. doi: 10.1038/nm.3026
Willemen, H. L., Huo, X. J., Mao-Ying, Q. L., Zijlstra, J., Heijnen, C. J., and Kavelaars, A. (2012). MicroRNA-124 as a novel treatment for persistent hyperalgesia. J. Neuroinflamm. 9:143. doi: 10.1186/1742-2094-9-143
Wu, X. N., Ye, Y. X., Niu, J. W., Li, Y., Li, X., You, X., et al. (2014). Defective PTEN regulation contributes to B cell hyperresponsiveness in systemic lupus erythematosus. Sci. Trans. Med. 6:246ra99. doi: 10.1126/scitranslmed.3009131
Xian, L., Wu, X., Pang, L., Lou, M., Rosen, C. J., Qiu, T., et al. (2012). Matrix IGF-1 maintains bone mass by activation of mTOR in mesenchymal stem cells. Nat. Med. 18, 1095–1101. doi: 10.1038/nm.2793
Xie, Y., Chen, Y., Zhang, L., Ge, W., and Tang, P. (2017). The roles of bone-derived exosomes and exosomal microRNAs in regulating bone remodelling. J. Cell Mol. Med. 21, 1033–1041. doi: 10.1111/jcmm.13039
Xie, Y., Zhang, L., Gao, Y., Ge, W., and Tang, P. (2015). The multiple roles of Microrna-223 in regulating bone metabolism. Molecules 20, 19433–19448. doi: 10.3390/molecules201019433
Xu, F., and Teitelbaum, S. L. (2013). Osteoclasts: new insights. Bone Res. 1, 11–26. doi: 10.4248/BR201301003
Xu, J., Pei, Y., Lu, J., Liang, X., Li, Y., Wang, J., et al. (2021). LncRNA SNHG7 alleviates IL-1beta-induced osteoarthritis by inhibiting miR-214-5p-mediated PPARGC1B signaling pathways. Int. Immunopharmacol. 90:107150. doi: 10.1016/j.intimp.2020.107150
Yagi, M., Miyamoto, T., Sawatani, Y., Iwamoto, K., Hosogane, N., Fujita, N., et al. (2005). DC-STAMP is essential for cell-cell fusion in osteoclasts and foreign body giant cells. J. Exp. Med. 202, 345–351. doi: 10.1084/jem.20050645
Yang, D., Tan, S., Yang, Z., Jiang, P., Qin, C., Yuan, Q., et al. (2018). Dihydromyricetin attenuates TNF-alpha-Induced endothelial dysfunction through miR-21-Mediated DDAH1/ADMA/NO signal pathway. Biomed. Res. Int. 2018:1047810. doi: 10.1155/2018/1047810
Yang, S., Zhang, W., Cai, M., Zhang, Y., Jin, F., Yan, S., et al. (2018). Suppression of bone resorption by miR-141 in aged rhesus monkeys. J. Bone Miner. Res. 33, 1799–1812. doi: 10.1002/jbmr.3479
Yang, X., Matsuda, K., Bialek, P., Jacquot, S., Masuoka, H. C., Schinke, T., et al. (2004). ATF4 is a substrate of RSK2 and an essential regulator of osteoblast biology; implication for coffin-lowry syndrome. Cell 117, 387–398.
Yang, Y., Liu, L., Zhang, Y., Guan, H., Wu, J., Zhu, X., et al. (2014). MiR-503 targets PI3K p85 and IKK-beta and suppresses progression of non-small cell lung cancer. Int. J. Cancer 135, 1531–1542. doi: 10.1002/ijc.28799
Yang, Z., Zeng, B., Wang, C., Wang, H., Huang, P., and Pan, Y. (2017). MicroRNA-124 alleviates chronic skin inflammation in atopic eczema via suppressing innate immune responses in keratinocytes. Cell Immunol. 319, 53–60. doi: 10.1016/j.cellimm.2017.08.003
Yao, H., Kong, F., and Zhou, Y. (2019). MiR-182 promotes cell proliferation, migration and invasion by targeting FoxF2 in endometrial carcinoma cells. Int. J. Clin. Exp. Pathol. 12, 1248–1259.
Yao, Z., Xing, L., and Boyce, B. F. (2009). NF-kappaB p100 limits TNF-induced bone resorption in mice by a TRAF3-dependent mechanism. J. Clin. Investigation 119, 3024–3034. doi: 10.1172/JCI38716
Yuan, X., Berg, N., Lee, J. W., Le, T. T., Neudecker, V., Jing, N., et al. (2018). MicroRNA miR-223 as regulator of innate immunity. J. Leukoc Biol. 104, 515–524. doi: 10.1002/JLB.3MR0218-079R
Zhang, J., Zhao, H., Chen, J., Xia, B., Jin, Y., Wei, W., et al. (2012). Interferon-beta-induced miR-155 inhibits osteoclast differentiation by targeting SOCS1 and MITF. FEBS Lett. 586, 3255–3262. doi: 10.1016/j.febslet.2012.06.047
Zhang, N., Fu, L., Bu, Y., Yao, Y., and Wang, Y. (2017). Downregulated expression of miR-223 promotes Toll-like receptor-activated inflammatory responses in macrophages by targeting RhoB. Mol. Immunol. 91, 42–48. doi: 10.1016/j.molimm.2017.08.026
Zhang, Q., and Cao, X. (2019). Epigenetic regulation of the innate immune response to infection. Nat. Rev. Immunol. 19, 417–432. doi: 10.1038/s41577-019-0151-6
Zhang, Y., Chen, X., Lian, H., Liu, J., Zhou, B., Han, S., et al. (2014). MicroRNA-503 acts as a tumor suppressor in glioblastoma for multiple antitumor effects by targeting IGF-1R. Oncol. Rep. 31, 1445–1452. doi: 10.3892/or.2013.2951
Zhao, B., Grimes, S. N., Li, S., Hu, X., and Ivashkiv, L. B. (2012). TNF-induced osteoclastogenesis and inflammatory bone resorption are inhibited by transcription factor RBP-J. J. Exp. Med. 209, 319–334. doi: 10.1084/jem.20111566
Zhao, B., and Ivashkiv, L. B. (2011). Negative regulation of osteoclastogenesis and bone resorption by cytokines and transcriptional repressors. Arthritis Res. Therapy 13:234. doi: 10.1186/ar3379
Zhao, B., Takami, M., Yamada, A., Wang, X., Koga, T., Hu, X., et al. (2009). Interferon regulatory factor-8 regulates bone metabolism by suppressing osteoclastogenesis. Nat. Med. 15, 1066–1071.
Zhao, C., Irie, N., Takada, Y., Shimoda, K., Miyamoto, T., Nishiwaki, T., et al. (2006). Bidirectional ephrinB2-EphB4 signaling controls bone homeostasis. Cell Metab. 4, 111–121. doi: 10.1016/j.cmet.2006.05.012
Zhao, C., Sun, W., Zhang, P., Ling, S., Li, Y., Zhao, D., et al. (2015). miR-214 promotes osteoclastogenesis by targeting Pten/PI3k/Akt pathway. RNA Biol. 12, 343–353. doi: 10.1080/15476286.2015.1017205
Zhao, H., Zhang, J., Shao, H., Liu, J., Jin, M., Chen, J., et al. (2017). Transforming growth factor beta1/Smad4 signaling affects osteoclast differentiation via regulation of miR-155 expression. Mol. Cells 40, 211–221. doi: 10.14348/molcells.2017.2303
Zhao, J. J., Yang, J., Lin, J., Yao, N., Zhu, Y., Zheng, J., et al. (2009). Identification of miRNAs associated with tumorigenesis of retinoblastoma by miRNA microarray analysis. Childs Nerv. Syst. 25, 13–20. doi: 10.1007/s00381-008-0701-x
Zhao, Q., Liu, C., Xie, Y., Tang, M., Luo, G., Chen, X., et al. (2020). Lung cancer cells derived circulating miR-21 promotes differentiation of monocytes into osteoclasts. Onco Targets Ther. 13, 2643–2656. doi: 10.2147/OTT.S232876
Zhong, L., Simoneau, B., Huot, J., and Simard, M. J. (2017). p38 and JNK pathways control E-selectin-dependent extravasation of colon cancer cells by modulating miR-31 transcription. Oncotarget 8, 1678–1687. doi: 10.18632/oncotarget.13779
Keywords: osteoclast, miRNA, bone resorption, rheumatoid arthritis, osteoporosis
Citation: Inoue K, Ng C, Xia Y and Zhao B (2021) Regulation of Osteoclastogenesis and Bone Resorption by miRNAs. Front. Cell Dev. Biol. 9:651161. doi: 10.3389/fcell.2021.651161
Received: 08 January 2021; Accepted: 12 May 2021;
Published: 18 June 2021.
Edited by:
Natalie A. Sims, University of Melbourne, AustraliaReviewed by:
Jiake Xu, University of Western Australia, AustraliaYankel Gabet, Tel Aviv University, Israel
Copyright © 2021 Inoue, Ng, Xia and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Baohong Zhao, emhhb2JAaHNzLmVkdQ==