Abstract
Lymphedema is a chronic inflammatory disease without an effective treatment method, and it results in a high disease burden and psychological distress in patients. Although there have been significant advances in targeted therapies, there are still no effective options to treat this refractory disease. In recent years, new advances and breakthroughs have been made in signaling pathways, including RAS/MAPK, PI3K/AKT, VEGF-C/VEGFR-3, HGF/MET, and TGF-β1, which are important for understanding the pathogenesis and disease progression of lymphedema. Mutations in genes encoding cell junctions affect the formation of junctions in lymphatic endothelial cells (LECs), causing abnormal lymphatic valve development and the impairment of lymphatic vessels. A vicious cycle of oxidative stress and chronic inflammation of lymphatic vessels leads to lymphedema. Moreover, the interactions and information communication of T-cell subsets, neutrophils, macrophages, dendritic cells (DCs), and fibroblasts with LECs play equally important roles in the progression of lymphedema. Therefore, this paper summarizes the reported signaling pathways, cell junctions, oxidative stress, and cell communication involved in lymphedema, with the goal of providing ideas and a basis for understanding the pathogenesis, disease progression and targeted therapy of lymphedema. By integrating current findings on signaling dysregulation, cell junctions, and cellular crosstalk, this review provides a conceptual framework for developing multitarget therapeutic strategies to restore lymphatic homeostasis and develop potential therapies for treating lymphedema.
1 Background
Lymphedema is a chronic disease of the lymphatic system that is characterized by localized fluid retention, swelling, inflammation, fibrosis, deposition of fatty tissue, and susceptibility to infection (Gousopoulos et al., 2016b; Tashiro et al., 2017; Grada and Phillips, 2017; Bowman and Rockson, 2024). The long-term limitation of limb movement can cause inconveniences and great psychological pressure on patients’ lives, seriously affecting their quality of life (Engel et al., 2003; Khan et al., 2012; Stuiver et al., 2015).
Lymphedema can be classified as either primary lymphedema or secondary lymphedema (Grada and Phillips, 2017; Yamamoto et al., 2015; Pinto et al., 2013) (Table 1). Primary lymphedema results from dysplasia or dysfunction of LECs and their associated vasculature development caused by mutations in genes involved in lymphatic development or function, leading to impaired lymphatic system function (Ferrell, 2002; Karkkainen et al., 2001; Karkkainen et al., 2000). Primary lymphedema is hereditary, as it has been reported that up to 29 genetic abnormalities are related to lymphedema (Michelini et al., 2020). Secondary lymphedema occurs after damage to the lymphatic vessels, such as bacterial or viral infection, cancer treatment, or filarial infection (Cheon et al., 2023; Rockson et al., 2019; Coriddi et al., 2017; Cormier et al., 2010). In secondary lymphedema, filariasis-associated lymphedema is the most common globally, whereas cancer-related lymphedema is more prevalent in Western countries. Global epidemiological patterns reveal significant geographic disparities: filarial lymphedema predominates in certain regions of India and Tanzania, with 250 districts in India alone accounting for more than 40% of the global burden of lymphatic filariasis. Moreover, cancer-related lymphedema affects 21%–28% of breast cancer survivors in Western countries, with the prevalence increasing to 75% among those with gynecological malignancies. (Grada and Phillips, 2017; Walsh et al., 2016; Fimbo et al., 2020; DiSipio et al., 2013; Dayan et al., 2018). Most current lymphedema treatments include massage, manual lymphatic drainage, compression bandages, therapeutic exercise, and dietary intervention (Shimizu et al., 2023). However, there is still a lack of targeted therapies for lymphedema, and there are no treatments based on molecular targets. A few lymphatic venous anastomoses are performed, supplemented by drug therapy, and an effective treatment plan is urgently needed (Lu et al., 2023). Therefore, this review summarizes some of the key signaling pathways involved in lymphedema, the cell junctions between LECs, and cell-to-cell information exchange in lymphedema, with the hope that insights from these studies will ultimately contribute to the development of effective management strategies for lymphedema.
TABLE 1
| Category | Primary lymphedema | Secondary lymphedema | |||
|---|---|---|---|---|---|
| Pathogenesis | LEC dysplasia/vasculature dysfunction | Injury lymphatic dysfunction | |||
| Key mechanisms | Genic mutation | Heredity | Infection Filariasis (global), virus, bacterial | Cancer treatment: Surgery, radiotherapy (Western prevalence) | Regional hotspots: India (40% global filariasis burden) |
| Epidemiology | Rare | Filarial: 250 endemic districts in India Cancer-related: 21%–28% breast cancer survivors (West), 75% gynecological malignancies | |||
| Core Therapies | Massage, manual lymphatic drainage, compression bandages, therapeutic exercise, and dietary intervention | ||||
The background information of lymphedema.
2 Physiology of the lymphatic system
The lymphatic system is a tubular network that, together with blood vessels, constitutes the body’s circulatory system and ensures normal physiological activities (Adams and Alitalo, 2007). It is known to be a unidirectional blind-ended system consisting of lymph nodes and lymphatic vessels, including lymphatic capillaries, pre-collecting and collecting vessels (Swartz, 2001; Leak, 1976; Brix et al., 2021). In peripheral tissues, a dense network of nonconventional lymphatic capillaries composed of a single layer of LECs is responsible for pressure-dependent drainage of interstitial fluid, cells, and macromolecules from the interstitial space into the lymphatic system (Bollinger and Partsch, 1985; Castenholz, 1984; Ohtani, 1987; Lynch et al., 2007; Scallan et al., 2022). Special button-like junctions, discontinuous basement membranes, a lack of pericyte coverage, and connections with the surrounding extracellular matrix constitute the high drainage capacity of lymphatic capillaries. Lymph fluid is transported through the anterior collecting ducts containing valves and sporadic smooth muscle cells (SMCs) to the collecting lymphatic vessels (Mandelbrot and Mandelbrot, 1982; Ducoli and Detmar, 2021). The collecting lymphatic vessels have intact basement membranes and “zipper-like” tight junctions and are surrounded by SMCs (Baluk et al., 2007). Both the anterior collecting and collecting lymphatic vessels contain mitral structures or valves that prevent the backflow of lymph nodes and ensure unidirectional lymphatic flow. SMCs, which surround lymphatic vessels, mediate the inward constriction of collecting lymphatic vessels, allowing the upward transport of lymph through multiple lymph nodes. However, there are exceptions (von der Weid, 2013). The contraction of lymphatic vessels lacking SMC coverage, such as pulmonary lymphatic vessels, mainly depends on the force generated by the surrounding tissues (Zawieja, 2009). After ascending through several lymph nodes, the lymph eventually returns to the vasculature through major lymphatic trunks, such as the thoracic duct. Because of this highly structured network, the lymphatic system is well suited to maintain fluid homeostasis. More importantly, the lymphatic system serves as a transport route for antigens and immune cells to initiate immune responses (Liao and von der Weid, 2015). In this context, LECs must follow specific and robust regulatory programs expressing specific lymphatic markers, which ensures not only their correct cell identity and function but also proper maturation and organization of the lymphatic network (Oliver et al., 2005).
3 Abnormal signaling pathway of lymphedema
A growing body of evidence has revealed important signaling pathways involved in lymphedema-related mechanisms. Signaling pathways such as the RAS/MAPK pathway, the PI3K/AKT pathway, the TGF-β1 pathway, and the HGF/MET signaling pathway have been confirmed to play important roles in the development and progression of lymphedema (Visser et al., 2012; Tidyman and Rauen, 2008; Fritz-Six et al., 2008; Agollah et al., 2014; Korhonen et al., 2022; Suzuki et al., 2022; Finegold et al., 2008; Koksharova et al., 2024; Alpaslan et al., 2024) (Table 2). The effects of these pathways are closely correlated with the upstream VEGF-C/VEGFR-3 signaling axis. Moreover, some of the less reported signaling pathways, such as the S1P, Notch1, NF-κB, EphrinB2/EphB4, Dchs1 and Fat4 signaling pathways, are similarly discussed below. As information on these pathways in relation to lymphedema remains limited, they provide new perspectives for future research and potential new targets for lymphedema treatment. The exchange of cell-to-cell information between different gene mutations, regulatory molecules, and key components of pathways is involved in the progression of lymphedema. According to the classification of signaling pathways, while introducing regulatory molecules and gene mutations related to lymphedema, we generated tables and pictures for reference (Figure 1).
TABLE 2
| Signaling pathway | Genes | Experimental model | Mechanisms of action | Types of lymphedema | Data sources |
|---|---|---|---|---|---|
| RAS/MAPK | AM (Fritz-Six et al., 2008) CALCRL (Fritz-Six et al., 2008) RAMP2 (Fritz-Six et al., 2008) | Knockout mice LECs HUVECs | Necessary for murine lymphatic vascular development Enhanced activation of ERK signal transduction | Primary | Human Mice |
| KRAS (Sheppard et al., 2023) | HDLECs Zebrafish | variants activate the RAS/MAPK pathway leading to lymphatic dysplasia and edema | Primary | Human Zebrafish | |
| NRAS (Carta et al., 2006; Ekvall et al., 2015) CBL (Hanson et al., 2014; Rodriguez-Viciana et al., 2006; Young et al., 2018; Brand et al., 2014; Castel et al., 2019) LZTR1 (Castel et al., 2019; Motta et al., 2019) PTPN11 (Bonetti et al., 2022; Martinelli et al., 2010; Tartaglia et al., 2002) SOS1 (Tidyman and Rauen, 2009; Roberts et al., 2007) SPRED1 (Cirstea et al., 2010) NF1 (Cirstea et al., 2010) RAF1 (Cirstea et al., 2010) BRAF (Cirstea et al., 2010) MEK1 (Cirstea et al., 2010) MEK2 (Cirstea et al., 2010) | NS patients | Mutations cause disruption of the Ras/MAPK signaling pathway | Primary | Human | |
| SHOC2 (Hanson et al., 2014; Rodriguez-Viciana et al., 2006; Young et al., 2018; Brand et al., 2014; Castel et al., 2019) MRAS (Hanson et al., 2014; Rodriguez-Viciana et al., 2006; Young et al., 2018; Brand et al., 2014; Castel et al., 2019) PP1 (Hanson et al., 2014; Rodriguez-Viciana et al., 2006; Young et al., 2018; Brand et al., 2014; Castel et al., 2019) | HEK293 DLD-1 | Mutations in MRAS, SHOC2, and PP1 contribute directly to NS by enhancing formation of a ternary complex | Primary | Human | |
| RIT1 (Castel et al., 2019; Motta et al., 2019) | Mutant mouse model HEK293 | Mutations cause disruption of the Ras/MAPK signaling pathway | Primary | Human Mice | |
| NSD1 (Visser et al., 2012) | SoS patients | Mutations cause disruption of the Ras/MAPK signaling pathway | Primary | Human | |
| PI3K/AKT | INPPL1 (Agollah et al., 2014) | Lymphatic disease family HDLECs | Negative regulation of the PI3K signaling pathway, whose mutation is associated with lymphatic dysfunction | Primary | Human |
| ANG2 (Korhonen et al., 2022) Tie2 (Korhonen et al., 2022) | Knockout mice | Gene deletion leads to abnormal development and function of embryonic lymphatic vessels, resulting in lymphatic dysfunction | Primary | Human Mice | |
| ARAP3 (Kartopawiro et al., 2014) | Mouse model Zebrafish | Lymphatic vessel development is essential | Primary | Mice Zebrafish | |
| PTEN (Goberdhan and Wilson, 2003; Kataru et al., 2021) | Knockout mice | The loss of PTEN leads to increased phosphorylation of AKT, thereby increasing LEC proliferation and differentiation | Primary | Mice | |
| KIF11 (Ostergaard et al., 2012) PIEZO1 (Fotiou et al., 2015) | MLCRD Patients Genetic and data analysis | Both KIF11 and PIEZO1 can activate the PI3K/AKT signaling pathway and increase cytoplasmic Ca2+ concentration Mutations affect the development of lymphatic structures | Primary | Human | |
| VEGF-C/VEGFR-3 | FLT 4 (Gordon et al., 2013; Irrthum et al., 2000) | Several animal models | Plays an important role in lymphatic vessel formation; mutations prevent phosphorylation | Primary | Several animal models |
| CCBE 1 (Brouillard et al., 2017; Bos et al., 2011; Au et al., 2010) ADAMTS3 (Brouillard et al., 2017; Bos et al., 2011; Au et al., 2010) | Hennekam syndrome patients Mouse model | Mutations can cause lymphatic abnormalities and even death | Primary | Human Mice | |
| PTPN14 (Brouillard et al., 2017; Bos et al., 2011; Au et al., 2010) | Mouse model | Lymphatic hyperplasia with lymphoedema following genetic defect | Primary | Human Mice | |
| ANG2 (Korhonen et al., 2022) Tie2 (Korhonen et al., 2022) | Knockout mice | Gene deletion leads to abnormal development and function of embryonic lymphatic vessels, resulting in lymphatic dysfunction | Primary | Human Mice | |
| GATA2 (Kazenwadel et al., 2015; Mansour et al., 2010) | Emberger syndrome patients Mouse model | It is required for lymphatic vessel valve development and maintenance | Primary | Huamn Mice | |
| SOX18 (Irrthum et al., 2003; Brouillard et al., 2014) | Clinical patients | Mutations can cause lymphatic dysfunction and lymphoedema | Primary | Human | |
| Prox1 (Kazenwadel et al., 2015; Mansour et al., 2010; Wigle and Oliver, 1999) | Knockout mice LECs | Essential for lymphatic system development Critical for the differentiation of LECs | Primary | Human Mice | |
| HGF/MET | HGF (Finegold et al., 2008) MET (Finegold et al., 2008) | Lymphedema patient Mouse model LECs | Promotes LEC proliferation, migration and tube formation Promotes lymphatic vessel formation in mice | Primary Secondary | Human Mice |
| TGF-β1 | TGF-β1 (Sano et al., 2020) Smad (Sano et al., 2020) | Mouse model Lymphedemapatients | TGFB1 is a key regulator of accelerated fibrosis while inhibiting lymphangiogenesis | Secondary | Human Mice |
| NF-κB | IKBKG (Roberts et al., 2010) | Lymphedema patient | Affects activation of the NF-κB signaling pathway, and defects in this pathway may lead to lymphatic vessel dysfunction | Primary | Human |
| STAT3 (Bowman and Rockson, 2024; Satapathy et al., 2006; Levy and Lee, 2002) | Lymphocyte Mouse model | Role in the inflammatory and fibrotic processes of lymphedema | Secondary | Human Mice | |
| NLRP3 (Zhou et al., 2023) | Mouse model | Promotes macrophage M2 polarization | Secondary | Mice | |
| TLR4 (Li et al., 2024) VEGF-C (Li et al., 2024) | Mouse model | Promote lymphatic angiogenesis to reduce lymphedema | Secondary | Mice | |
| EphrinB2/EphB4 | EphrinB2 (Frye et al., 2020) EphB4 (Frye et al., 2020) CLDN5 (Frye et al., 2020) | Mouse model HDLECSs | LECs connections are required and selectively regulate lymphatic vessel permeability and function | Primary Secondary | Human Mice |
| Dchs1 and Fat4 | Dchs1 (Pujol et al., 2017) Fat4 (Pujol et al., 2017) | Knockout mice Hennekam patients | Controls endothelial cell polarization during lymphatic valve morphogenesis | Primary | Human Mice |
| Notch1 | Notch1 (Jannaway et al., 2023) DLL4 (Jannaway et al., 2023) | Lymphedema patients Mouse model | Functional blockade can lead to lymphedema disorders | Primary | Human Mice |
| S1P | S1P (Kim et al., 2023) | Mouse model LECs | Signaling abnormalities exacerbate lymphatic dysfunction and tissue inflammation | Secondary | Human Mice |
| TRML1 | TRPML1 (Yang et al., 2024) AQPs (Yang et al., 2024) | Mouse model HUVECs LECs | Alteration of cell permeability through regulation of AQPs localization triggers lymphedema with chronic inflammation | Secondary | Human Mice |
| Wnt/β-catenin | Wnt (Cha et al., 2018) β-catenin (Cha et al., 2018) | Knockout mice HLECs LECs | Plays a key role in the development of both lymphatic vessels and valves | Primary | Human Mice |
Genes involved in the lymphedema signaling pathway.
FIGURE 1
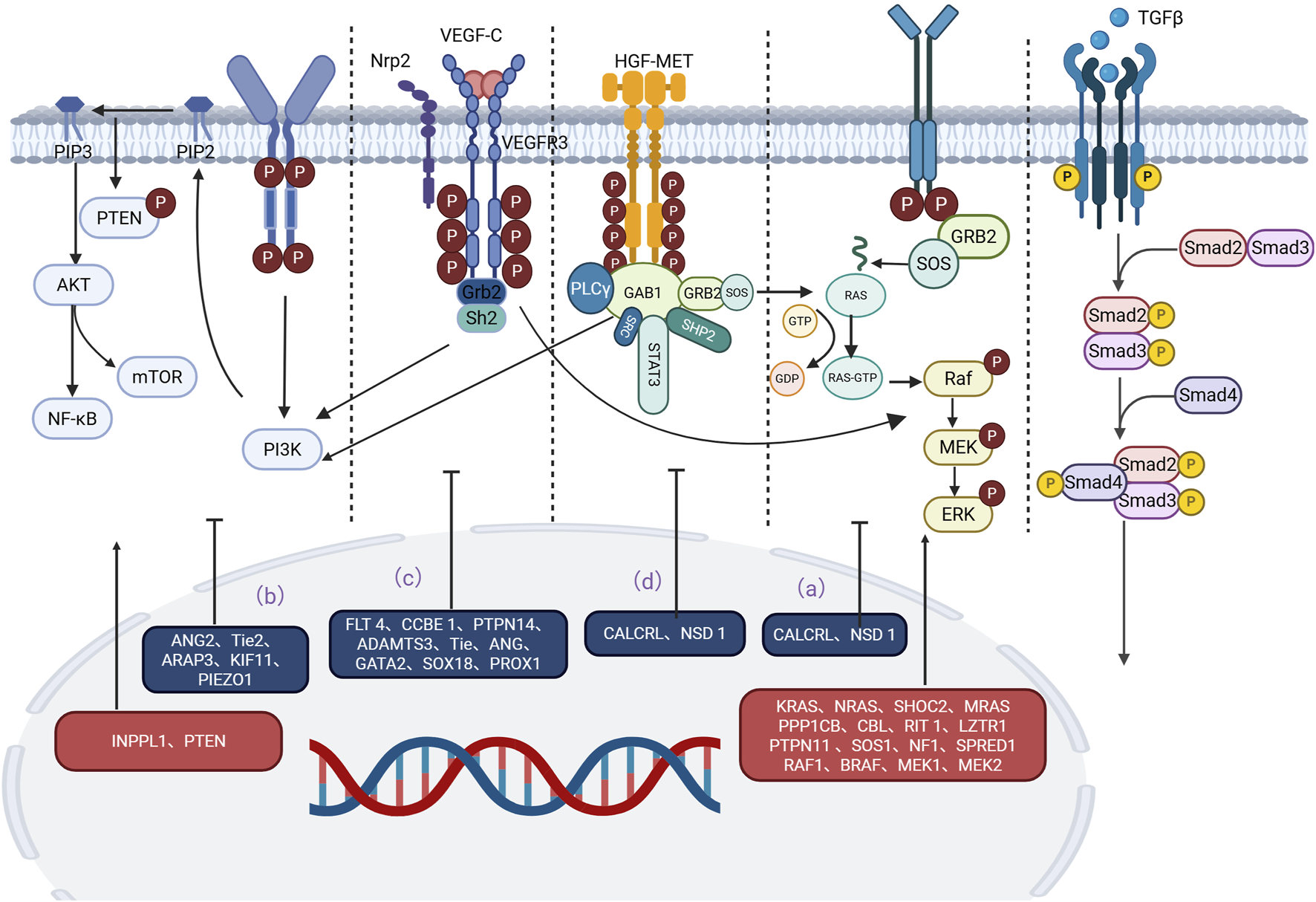
Main signaling pathways involved in the occurrence and development of lymphedema. Mutations in key proteins or regulators in these pathways can lead to overactivation or inhibition of signaling pathways, which can trigger lymphedema. (a) RAS‒MAPK pathway: Mutations in CALCRL and NSD1 weaken the RAS‒MAPK signaling pathway. Mutations in KRAS, MEK, SHOC2, MRAS, PP1, SOS1, CBL, RIT1, LZTR1, PTPN11, NF1, SPRED1, RAF1, BRAF, and MEK1/2 often lead to an abnormal MAPK cascade that increases signaling and ultimately leads to lymphedema. (b) P13K/AKT pathway: Mutations in INPPL1, ANG2, TIE2, ARAP3, PTEN, KIF11, and PIEZO1 are also strongly associated with lymphedema. (c) VEGF-C/VEGFR-3 pathway: Mutations in the LT4, CCBE1, PTPN14, ADAMTS3, TIE, ANG, GATA2, Prox1, Prox1, and SOX18 genes interfere with the normal conduction of the VEGF-C/VEGFR-3 pathway, ultimately leading to lymphedema. (d) HGF/MET pathway: Mutations in HGF and MET disrupt signal transduction in the HGF/MET system and lead to lymphedema. An in-depth study of these signaling pathways is expected to provide new targets for the prevention and treatment of lymphedema.
3.1 RAS/MAPK signaling pathways
The RAS/MAPK pathway is closely associated with lymphedema development. For example, Noonan syndrome (NS), cardio-facio-cutaneous (CFC) syndrome, and Sotos syndrome are all lymphedema syndromes associated with RAS/MAPK pathway abnormalities (Visser et al., 2012; Tidyman and Rauen, 2008). Many regulatory molecules and key components play important roles in the progression of lymphedema. For example, mutation or deletion of AM, CALCRL and RAMP2 leads to a lack of normal activation of the MAPK/ERK signaling pathway, resulting in reduced LEC proliferation, hypoplastic lymphatic vessels, and ultimately interstitial lymphedema (Fritz-Six et al., 2008). Sheppard SE et al. reported that the expression of activated KRAS variants increased the phosphorylation of ERK and increased MAPK signaling in human dermal lymphatic endothelial cells (HDLECs), leading to lymphatic dysplasia and edema (Sheppard et al., 2023). In the following text, we address the existing reports of NS, CFC, and Sotos syndrome with primary lymphedema separately.
NS, caused by germline pathogenic variants of genes within the RAS/MAPK signaling pathway, is characterized mainly by symptoms of lymphatic dysplasia and abnormalities of the lymphatic system (Sleutjes et al., 2022). NS patient species were also detected for germline mutations in both KRAS and NRAS, although in a small proportion (Carta et al., 2006; Ekvall et al., 2015). Most genes involved in NS encode proteins indispensable to the RAS/MAPK pathway, and pathogenic mutations often increase signaling through this pathway (Schubbert et al., 2007). For example, the gene encoding the SHOC2–MRAS–PP1 complex is a positive regulator of the RAS-ERK signaling cascade, and the gene encoding CBL can negatively regulate intracellular signaling downstream of the receptor tyrosine kinase. Mutations in the genes encoding SHOC2, MRAS, PP1 and CBL can cause symptoms similar to those of NS (Hanson et al., 2014; Rodriguez-Viciana et al., 2006; Young et al., 2018; Brand et al., 2014; Castel et al., 2019). Moreover, NS is closely associated with aberrant mutations in RIT1, LZTR1, PTPN11 and SOS1, which regulate the RAS/MAPK pathway. Castel P and Motta M et al. reported that the RIT1 and LZTR1 genes regulate the progression of severe bilateral lower limb lymphedema in adolescent patients through abnormal activation of the RAS/MAPK pathway (Castel et al., 2019; Motta et al., 2019). Tartaglia M et al. reported that 50% of NS cases are caused by missense gain-of-function mutations in PTPN11, which affect the domain of SHP2 and thus the signaling in the RAS/MAPK pathway (Maheshwari et al., 2002; Tartaglia et al., 2001). Missense mutations in SOS1, which occur in approximately 13% of cases of NS, increase RAS activity and subsequently enhance RAS/MAPK signaling pathway transduction by disrupting the autoinhibition of RAS-GEF activity (Tidyman and Rauen, 2009; Roberts et al., 2007). In addition, a report by Cirstea IC reported that germline mutations in the RAS regulatory factors NF1 and SPRED1, as well as downstream signaling factors such as RAF1, BRAF, MEK1, and MEK2, are involved. These mutations typically lead to abnormalities in the MAPK cascade, resulting in increased signal flow (Cirstea et al., 2010). Studies and case reports of lymphatic abnormalities in NS patients caused by RRAS2 gene mutations are still rare and are valuable for further investigations. In conclusion, NS mutations in patients, such as those in KRAS, NRAS, SHOC2, MRAS, PP1, CBL, RIT1, LZTR1, PTPN11, SOS1, NF1, SPRED1, RAF1, BRAF, MEK1, and MEK2, deserve further investigation of the specific mechanism of action in primary lymphedema.
Studies of the RAS/MAPK pathway in CFC and Sotos syndrome remain scarce. The NSD1 gene product is postulated to be a corepressor protein of growth-promoting genes. Sotos syndrome is a childhood overgrowth syndrome caused by haploinsufficiency of the NSD1 gene (Kurotaki et al., 2002). McClelland J reported the presence of primary lymphedema in female patients with Sotos syndrome caused by NSD1 internal mutations (McClelland et al., 2016). Genetic mutations in NSD1 are accompanied by lower phosphorylation levels of the MAPK kinase (Visser et al., 2012). Similarly, CFC, caused by genetic mutations in BRAF, MEK1/2, HRAS, and KRAS that lead to altered signaling by the RAS/MAPK pathway, is a very rare sporadic disease accompanied by symptoms of primary lymphedema (Morcaldi et al., 2015).
Currently, abnormalities in the RAS/MAPK pathway are reflected in the different syndromes associated with primary lymphedema, and more genes remain to be discovered. Moreover, studying the abnormal manifestations of the RAS/MAPK signaling pathway in primary lymphedema is highly valuable for the study of pathogenesis and for the further development of therapeutic methods.
3.2 PI3K/AKT signaling pathway
The PI3K/AKT signaling pathway plays a critical role in many aspects of cell growth, proliferation, survival, metabolism, and migration and is one of the core pathways regulating cell function (Sheng et al., 2003; Zhao et al., 2004). Both VEGF-C/VEGFR-3 signaling and HGF/MET signaling can activate the PI3K/AKT pathway. Similar to the MAPK/ERK signaling pathway, the activation of the PI3K/AKT signaling pathway plays a crucial role in the development of lymphedema. For example, Coso S et al. reported that the activation of PI3K/AKT by VEGF-C/VEGFR-3 leads to the phosphorylation of P70S6K, eNOS, PLCγ1, and ERK1/2, thus participating in the formation of lymphatic vessels (Coso et al., 2012). Yang Y et al. reported that VE-cadherin regulates AKT signaling, which is involved in lymphatic valve formation and maintenance (Yang et al., 2019). The role of VEGF-C/VEGFR-3 signaling and HGF/MET signaling in the activation of components of the PI3K/AKT signaling pathway in the pathogenesis of lymphedema should be further explored.
Many regulatory molecules, such as SHIP-2, VEGF-C, and H2S, are able to modulate the PI3K/AKT signaling pathway to play important roles in lymphedema. Agollah GD et al. reported that mutations in the INPPL1 gene encoding SHIP-2 lead to hyperactivation of PI3K/AKT signaling, which is negatively regulated by SHIP-2 to mediate various forms of lymphedema, due to reduced LEC proliferation, adhesion, migration, and lumen formation (Agollah et al., 2014). Korhonen EA et al. reported that the activation of Ang2/Tie/PI3K signaling in LECs induced by VEGF-C promotes lymphatic vessel formation, which provides further investigation and potential therapeutic means for the targeted treatment of secondary lymphedema (Korhonen et al., 2022). Suzuki J et al. reported that H2S activates the PI3K/AKT signaling pathway in LECs, thus improving secondary lymphedema (Suzuki et al., 2022). Some of these regulatory molecules are gene expression products of LECs, and some are endogenous molecules that regulate PI3K/AKT signaling and are involved in the progression of lymphedema.
Genetic mutations in PIK3CA, ARAP3, PIEZO1, and PTEN also play important roles in the PI3K/AKT signaling pathway in the development of lymphedema. Mutations in genes encoding proteins that regulate the PI3K/AKT signaling pathway can also contribute to the progression of lymphedema. The PI3K/AKT pathway can be activated by two auxiliary proteins: KIF11 and PIEZO1. KIF11 activates PI3KA and increases the cytoplasmic Ca2+ concentration (Bonetti et al., 2022). Ostergaard P noted that KIF11 mutations are associated with congenitallymphedema (Ostergaard et al., 2012). Fotiou E noted that novel mutations in PIEZO1 can also lead to primary lymphedema (Fotiou et al., 2015). Furthermore, Krugmann S et al. reported that ARAP3 is also a genuine effector of the PI3K signaling system (Krugmann et al., 2002). Moreover, Kartopawiro J et al. reported that the deletion of ARAP3 caused subcutaneous lymphatic vascular dysplasia and hereditary lymphedema (Kartopawiro et al., 2014). In comparison, Kataru RP and colleagues reported that in a PTEN knockout mouse model, the inhibition of PTEN promoted PI3K/AKT signaling in LECs, thereby increasing LECs proliferation. This provides a potential target for improving lymphatic growth and function (Goberdhan and Wilson, 2003; Kataru et al., 2021). Gene mutations affecting the PI3K/AKT signaling pathway still deserve further investigation and discussion.
Interestingly, there is crosstalk between the PI3K/AKT pathway and the MAPK/ERK pathway in RAS and PI3KA. PI3KA can also be activated by RAS, connecting the RAS/MAPK and PI3K/AKT pathways (Bui and Hong, 2020). Rodriguez-Viciana P et al. reported that PI3K activation by RAS is correlated with direct binding between RAS and p110 (Rodriguez-Viciana et al., 1994; Chan et al., 2002). Zimmermann S et al. reported that the phosphorylation of RAF by AKT inhibited the RAS–RAF–MEK–ERK cascade (Zimmermann et al., 1999). The interaction of the PI3K/AKT pathway and the MAPK/ERK pathway in lymphedema is important for mechanistic studies. The links between these signaling pathways should be further discussed.
3.3 VEGF-C/VEGFR-3 signaling pathway
VEGF-C and VEGF-D bind VEGFR3 on the surface of LECs and induce lymphatic proliferation, migration, and survival by activating the intracellular PI3K-AKT and MAPK-ERK signaling pathways (Kuonqui et al., 2023). Many studies have confirmed that mutations in the FLT4 gene encoding VEGFR3 cause Milroy’s disease, an inherited primary lymphedema (Gordon et al., 2013; Irrthum et al., 2000). Many regulatory proteins are involved in lymphedema and are regulated by the VEGF-C/VEGFR-3 axis. Mutations in CCBE1, PTPN14, and ADAMTS3 can lead to abnormal VEGF-C/VEGFR-3 signaling, causing lymphedema (Brouillard et al., 2017; Bos et al., 2011; Au et al., 2010). Korhonen et al. reported that deletion of the Tie receptor or blockade of Ang2 also reduced lymphangiogenesis induced by VEGF-C (Korhonen et al., 2022). Several transcription factors and gene mutations are similarly involved in regulating the VEGF-C/VEGFR-3 axis to influence the development of lymphedema. Mutations in the GATA2 gene, the transcription factor controlling Prox1 and FOXC2 expression, cause primary lymphedema with myelodysplasia (Kazenwadel et al., 2015; Mansour et al., 2010). Prox1 is a key transcription factor in the differentiation of LECs. Prox1 can repress BEC-specific markers and upregulate LEC-specific genes. Sunju Lee et al. reported that blood vascular endothelial cells (BECs) in the main vein are able to migrate and form primitive lymphatic vessels and that both VEGF-C and Prox1 are indispensable regulators of this process, accompanied by the suppression of BEC-specific markers and an increase in the levels of LEC-specific genes, such as VEGFR-3 and FGFR-3 (Lee et al., 2009). In mice, Prox1-deficient mice are unable to form a lymphatic system resulting in severe edema and death (Wigle and Oliver, 1999). In addition, abnormal lymphatic leakage from lymphatic vessels promoted by Prox1 haploinsufficiency contributes to adult-onset obesity (Harvey et al., 2005). Lymphatic leakage also creates an immunosuppressive environment in Prox1+/− mice defective in the Prox1 gene (Herrada et al., 2022). Mutations in SOX18 may indirectly regulate VEGFR3 expression by affecting Prox1, resulting in lymphedema (Irrthum et al., 2003; Brouillard et al., 2014). Moreover, the transcription factors FOXC2 and NFATc1 are both involved in lymphatic valve development and the maturation of the lymphatic system, which may provide potential therapeutic means for the treatment of lymphedema (Norrmén et al., 2009). VEGFR3 was once thought to maintain button junctions, but recent studies have shown that it is not required throughout the life cycle after the formation of button junctions (Jannaway et al., 2023). The VEGF-C/VEGFR-3 signaling pathway is important in the mechanism of lymphedema and in the formation of normal lymphatic vessels.
3.4 HGF/MET signaling pathway
Currently, an increasing number of case studies support the potential role of HGF mutations in primary and secondary lymphedema. By sequencing the HGF and MET genes in patients with lymphedema and patients with secondary lymphedema after cancer treatment, Finegold DN et al. reported that four HGF mutations and two MET gene mutations were closely associated with multiple types of lymphedema, which may provide therapeutic targets for lymphedema (Finegold et al., 2008). Koksharova G et al. reported a close association between HGF gene mutation and clinical symptoms of lymphedema. Genetic mutations in HGF are also involved in the pathogenesis of lymphedema (Koksharova et al., 2024). In a study of 770 patients, Alpaslan M et al. reported that mutations in the HGF gene result in mRNA degradation or impaired protein function, which impairs the activation of the AKT and ERK1/2 pathways, leading to primary lymphedema, mainly in the lower limbs, with the onset of the disease extending from childhood to adulthood (Alpaslan et al., 2024). Regulatory molecules of the HGF/MET signaling pathway are equally able to participate in the progression of lymphedema. Bonetti G reported that CBL and PTPN11 are also involved in MET signal transduction. Among them, PTPN11 can activate RAS and AKT, thus acting on both the AKT/PI3K pathway and the RAS/MAPK pathway. Functional mutations in CBL and PTPN11 can cause NS with lymphedema (Bonetti et al., 2022; Martinelli et al., 2010; Tartaglia et al., 2002). The HGF/MET signaling pathway plays an important role in the pathogenesis of lymphedema, which is closely related to the interaction of RAS/M APK signaling and PI3K/AKT signaling.
3.5 TGF-β1 signaling pathway
The activation of the TGF-β1 signaling pathway has been identified as a pivotal factor in the progression of fibrosis in lymphedema. Sano M et al. utilized a rat model to elucidate the onset of dermal fibrosis with activation of the TGF-β1/Smad signaling pathway, thereby providing the first evidence of the role of this pathway in the development of secondary lymphedema in humans (Sano et al., 2020). Increased expression and abnormal activation of TGF-β1 promote skin fibrosis in patients with secondary lower extremity lymphedema (Di et al., 2016). Recent studies have shown that TGF-β1-activated fibroblast subpopulations are closely associated with the disease stage and fibrosis progression of lymphedema in human and mouse lymphedema tissues (Will et al., 2024a). As some research findings suggest, dysregulated TGF-β1 promotes extracellular matrix deposition in patients with breast cancer-related lymphedema (BCRL) and in mouse models (Baik et al., 2022). The inhibition of TGF-β1, a multifaceted mediator of fibrosis and extracellular matrix deposition, has been shown to reduce T-cell inflammatory infiltration (Avraham et al., 2010). This finding is further supported by the observations of Sun D et al., who reported that TGF-β1 is overexpressed in the skin of patients with symptomatic secondary lymphedema. In addition, studies have demonstrated that its inhibition in vivo can increase neolymphangiogenesis and alleviate lymphedema in a rodent tail model (Di et al., 2016; Avraham et al., 2010; Yan et al., 2011). Furthermore, Baik JE et al. reported that TGF-β1 is an essential regulator of ECM deposition in secondary lymphedema and that inhibition of this response is a promising means of treating lymphedema (Baik et al., 2022). Notably, TGF-β1 also possesses immunosuppressive properties, which could contribute to chronic inflammation secondary to lymphedema. TGF-β1 influences immunoinflammation by modulating the Th17-Treg balance (Yoshimura and Muto, 2011). It has also been shown to inhibit inflammatory signaling pathways such as the NF-κB pathway and reduce the expression of inflammatory factors and chemokines, including IL-6, IFN-γ, and RANTES (Shiou et al., 2013; Dai et al., 2011). LECs has been observed to secrete factors such as TGF-β1, which functions to inhibit DC maturation, thereby reducing inflammation in lymphoid tissues (Christiansen et al., 2016). However, Avraham T’s study revealed that the inhibition of TGF-β function reduces chronic inflammation, thereby rendering the role of TGF-β in secondary lymphedema more complex and elusive. Consequently, the inhibition of TGF-β1 has emerged as a promising treatment for secondary lymphedema. The role of TGF-β1 in the development and progression of lymphedema warrants further exploration, and we anticipate providing novel concepts and methodologies for targeting TGF-β1 in the treatment of lymphedema.
3.6 Other signaling pathways
3.6.1 NF-κB signaling pathway
Roberts CML et al. reported that the occurrence of lymphedema in a 6-year-old boy was closely related to the mutation of the IKBKG gene, which encodes the NEMO protein (IKK γ) to regulate the activation of the NF-κB signaling pathway (Roberts et al., 2010). The chronic inflammatory state and fibrosis in lymphedema are strongly associated with STAT3 and NF-κB (Bowman and Rockson, 2024). A study revealed elevated IL-6 levels in secondary filarial lymphedema (Satapathy et al., 2006). IL-6 is able to hyperactivate downstream STAT3 (Levy and Lee, 2002). In addition, there is a synergistic interaction between STAT3 and NF-κB to produce various inflammatory cytokines. STAT3 and NF-κB may undergo positive feedback amplification to maintain the chronic inflammatory state of lymphedema (Hirano, 2021). NF-κB plays an important role in different types of secondary lymphedema. Zhou Z et al. reported that Substance P promoted macrophage M2 macrophage polarization by regulating the NF-κB/NLRP3 signaling pathway, thus alleviating secondary lymphedema (Zhou et al., 2023). In addition, Yang C reported for the first time that the NF-κB inhibitor bortezomib can relieve limb lymphedema in patients with multiple myeloma (Yang et al., 2021a). In secondary lymphedema associated with lymphatic filariasis infection, Babu S et al. reported that TLR2 and TLR9 are involved in inducing the activation of the MAPK and NF-κB pathways to promote the pathological process of lymphedema (Babu et al., 2011). Interestingly, NF-κB plays an important role in lymphatic vessel formation, which may provide new ideas and directions for the subsequent treatment of lymphedema. Li et al. reported that type II arabinogalactan (CAPW-1) could activate the TLR4/NF-κB/VEGF-C signaling pathway in LECs to promote lymphatic angiogenesis and reduce edema in mice with secondary lymphedema (Li et al., 2024). In summary, inhibition of the inflammatory cell NF-κB pathway is important in the treatment of secondary lymphedema. In addition, the activation of NF-κB in LECs is also important in the development and progression of lymphedema. This seems to be a bidirectional issue that requires further investigation in the future.
3.6.2 EphrinB2/EphB4 signaling pathway
The diminished signaling between EphrinB2 and EphB4 in lymph vessels mirrors the heightened vessel leakage evident in response to bacterial infections and primary lymphedema. This phenomenon is associated with a decrease in CLDN5 levels at LEC junctions, a consequence of the loss of EphrinB2-EphB4 signaling (Frye et al., 2020). The EphrinB2/EphB4 signaling pathway has been identified as a promising target for the treatment of lymphedema and related diseases. The specific signaling pathways of EphrinB2-EphB4 in LECs are not yet fully understood. However, the activation of EphB4 receptors or their downstream signaling pathways, along with the regulation of signaling between these two proteins, may provide new therapeutic approaches through the development of drugs that increase the integrity of LECs.
3.6.3 Dchs1 and Fat4 signaling pathways
Abnormal valve development in mice with defective Dchs1 and Fat4 genes causes blocked lymphatic fluid flow and triggers primary lymphedema (Pujol et al., 2017). This study highlights the possibility that abnormal signaling pathways in related genetic disorders may lead to lymphatic system disorders.
3.6.4 Notch1 signaling pathway
Notch1 has been shown to engage in crosstalk with the VEGF signaling pathway, thereby influencing lymphatic vessel development and function. In their seminal study, Michelini S et al. advanced a compelling proposition regarding the most common genetic variant associated with genetic susceptibility to Notch1 lymphedema (Michelini et al., 2021). LECs lacking Notch1 signaling result in abnormal enlargement and formation of lymphatic vessels. Jannaway M et al. reported that forced activation of Notch1 signaling in the VEGFR3/Notch1 signaling axis in the absence of VEGFR3 restored lymphatic capillary button junction formation and interstitial fluid uptake (Jannaway et al., 2023). Upregulation of Notch1 signaling ameliorates VEGFR3 deficiency and can be used as a potential therapeutic approach for treating lymphedema (Jannaway et al., 2023). Notch1 signaling may play an important role in the prevention or treatment of acquired lymphedema. Tian W et al. reported that patients with acquired lymphedema have elevated levels of the proinflammatory mediator leukotriene B4 (LTB4), which inhibits VEGFR3 and Notch signaling involved in lymphedema. In addition, Notch1 knockout mice are insensitive to the therapeutic effects of LTB4 antagonists, which further supports the important role of Notch1 signaling in LTB4 regulation of lymphedema (Tian et al., 2017). Piezo1 is able to downregulate downstream Notch1. Choi D et al. reported that activation of Piezo1 or the use of the Piezo1 agonist Yoda1 effectively inhibited the development of postoperative lymphedema and enhanced lymphoid sprout growth (Choi et al., 2022). Interestingly, whether Notch1 signaling is downregulated or upregulated in the treatment of lymphedema, which may result from differences in Notch1 upstream signaling, remains a question worth exploring. In conclusion, the Notch1 signaling pathway can be targeted for the treatment of lymphedema. The development of drugs related to the Notch1 signaling pathway may be beneficial for the treatment of lymphedema.
3.6.5 S1P signaling pathway
In a study by Kim D et al., the inhibition of the S1P signaling pathway was confirmed to increase the expression of P-selectin in LECs, which relieved the symptoms of secondary lymphedema (Kim et al., 2023). Currently, research on the pathogenesis of the S1P signaling pathway in lymphedema is still scarce. Focusing on the link between S1P signaling and other pathways and its role in disease progression provides new insights and treatments for lymphedema.
3.6.6 Wnt/β-catenin signaling pathway
FOXC2 and GATA2 are also crucial for the differentiation of lymphatic valve endothelial cells. Oscillatory shear stress (OSS) enhances Wnt/β-catenin signaling in cultured LECs, inducing the expression of lymphedema-associated transcription factors GATA2 and FOXC2 (Choi et al., 2016; Cha et al., 2016).PROX1 is a key transcription factor in LECs differentiation.Further studies by Cha et al. revealed that PROX1 interacts with β-catenin and TCF7L1 to promote the expression of FOXC2 and GATA2 (Cha et al., 2018).Research on the Wnt/β-catenin signaling pathway not only reveals the molecular mechanisms involved in LEC differentiation but also provides new ideas and directions for the molecular-level treatment of lymphedema.
3.6.7 TRPML1/AQPs signaling pathway
Cell permeability is considered one of the potential pathophysiological mechanisms of lymphedema. In particular, the dynamic relocalization of aquaporins (AQPs) within cells significantly affects cell permeability (Conner et al., 2013). A recent study demonstrated that TRPML1 induces the accumulation of AQP2 at the apical surface of cells while disassembling the actin cytoskeleton, significantly increasing cell permeability and enhancing the response of cells to hypoosmotic stimulation in terms of water flux (Scorza et al., 2023). Our lab’s latest research revealed that TRPML1 regulates the localization of AQP3 and AQP5 on the cell membrane in human lymphatic endothelial cells (HLECs), leading to increased cell permeability, promoting the development of lymphedema, and accompanying chronic inflammatory responses (Yang et al., 2024). These findings suggest that TRPML1 may be an important trigger for lymphedema. With further research into the functions of TRPML1 and aquaporins, new molecular therapeutic targets for secondary lymphedema are expected to emerge in the future.
4 Cellular communication in lymphedema
Different subsets of T cells, including Th 1, Th 2, Th 17, Th 22 and Treg cells, play important roles in the pathological process and pathogenesis of lymphedema. The interaction of different subpopulations with LECs is important for a deeper understanding of lymphedema. Moreover, the interaction of neutrophils, macrophages, and DCs with LECs, especially macrophages, is important for the progression of lymphedema. Below, we summarize the previously reported roles of different cell types in lymphedema, including several signaling pathways, therapeutic measures, and pathological processes (Figure 2).
FIGURE 2
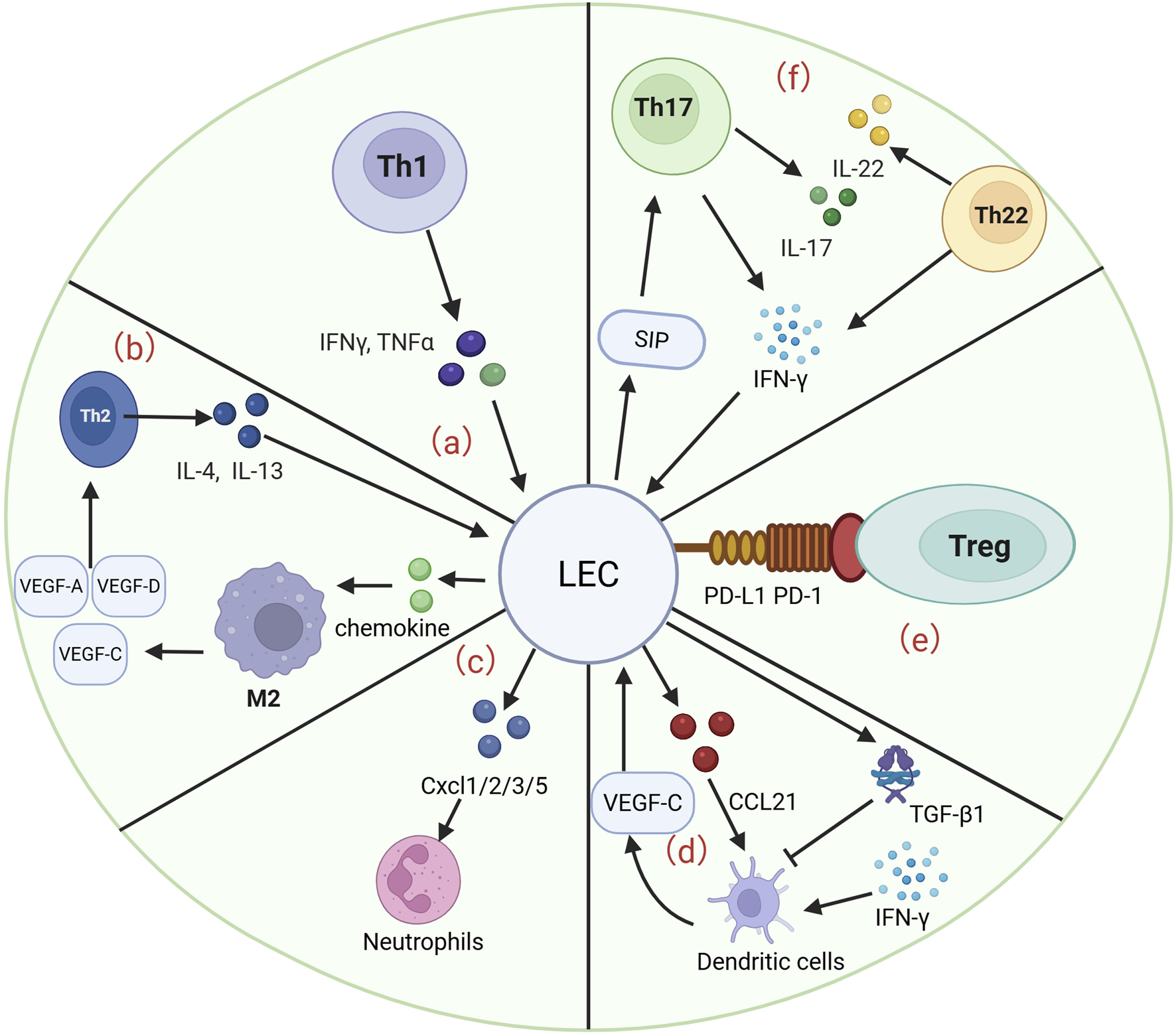
LECs interact with many cells to participate in the development and progression of lymphedema. (a) Th1 cells are able to release IFN-γ and TNF-α to act on LECs, which are involved in the pathological process of lymphedema and the progression of fibrosis. (b) Th2 cells are able to release IL-4 and IL-13 to promote the progression of LEC apoptosis. LECs are able to act on Th2 cells by releasing chemokines that act on M2 macrophages to induce the production of VEGF-A/C/D, enhancing lymphangiogenesis and fibrosis. (c) LECs were able to release CXCL 1/2/3/5 to regulate neutrophil activity and enhance the infiltration of inflammatory cells in a murine lymphedema model. (d) VEGF-C secreted by DCs is able to drive CCL21 secretion by LECs, which guides DCs into lymphatic vessels. LECs are able to release TGF-β1 to inhibit DC maturation and subsequently reduce the inflammatory response. (e) LECs are able to inhibit the inflammatory progression of lymphedema by binding PD-L1 to PD-1 on Tregs. (f) S1P gradient in which LECs are able to recruit Th17 cells. Th17 cells in lymphedema tissue secrete IL-17 and IFN-γ. Similarly, Th22 cells are able to release high levels of IL-22 and IFN-γ, both of which are involved in the progression of lymphedema.
4.1 Th1
Th1 cells play a critical role in the formation and progression of secondary lymphedema through their proinflammatory effects. S1PR1-deficient LECs promote the differentiation of CD4+ T cells into Th1 cells via direct contact (Kim et al., 2023). Similarly, in individuals with filarial lymphedema, BmA significantly increases the production of the Th1-type cytokines IFN-γ and TNF-α (Babu et al., 2009). Paradoxically, in a study of parasitic infection causing lymphedema, Babu S et al. reported a significant reduction in IFN-γ and TNF-α production by Th1 cells, with low expression of T-bet and attenuated Th1 responses (Babu et al., 2006). Loghry HJ et al. reported that Bma-LEC-2, which is secreted by parasites infecting patients with lymphatic filariasis, can selectively induce the apoptosis of Th1 cells and inhibit the host’s Th1 immune response, leading to persistent parasitic infection and the exacerbation of lymphedema disease (Loghry et al., 2022). Studies of Th1 cells in lymphatic filariasis-induced lymphedema still need further investigation. Similarly, Miyazaki T reported that abnormal activation of the calmodulin system in LECs inhibited the generation of TGF-β1, a process that affects the stability of Tregs and may indirectly affect the proinflammatory response of Th1 cells (Miyazaki et al., 2023). The ability of hyaluronidase and P-selectin to improve lymphedema differentiation by enhancing Th1 cells and reducing Th2 cell differentiation is promising (Kim et al., 2023; Cho et al., 2017). It can be speculated that increased Th1 cell differentiation exacerbates the development of lymphedema in the presence of blocked or dysfunctional lymphangiogenesis.
4.2 Th2
Th2 cells, a type of helper T cell, mainly secrete cytokines such as IL-4 and IL-13 during the progression of secondary lymphedema. Compared with Th1 cells, Th2 cells play an equally important role in the pathological process of lymphedema and in antifibrotic therapy. In pathogenesis, Savetsky IL et al. reported that IL-4 and IL-13 secreted by Th2 cells inhibit their proliferation, migration and tubular structure formation by promoting the apoptosis of LECs (Savetsky et al., 2015). In the case of lymphedema, impaired S1P signaling in LECs normally fails to regulate Th2 cell differentiation, further aggravating the symptoms of secondary lymphedema (Kim et al., 2023). Abnormal activation of the calmodulin (calpain) system of LECs leads to impaired stability of Th2 cell subsets, affecting the function of the lymphatic system (Miyazaki et al., 2023). A study by Avraham T et al. and Mehrara BJ et al. revealed that the inhibition of Th2 differentiation with IL-4- or IL-13-blocking agents, such as neutralizing antibodies, significantly relieved the symptoms of secondary lymphedema (Avraham et al., 2013; Mehrara et al., 2021).
4.3 Th17
Th17 cells in lymphedema and elephant cutaneous tumor symptoms due to secondary lymphatic filariasis are closely associated with Th17 cells (Fu and Liu, 2023). The immune response in lymphedema patients progresses in the directions of Th1, Th2, and Th17 (Pfarr et al., 2009). Among them, the levels of the inflammatory cytokines IL-17 and IFN-γ secreted by Th17 cells are significantly greater than those secreted by control cells (Anuradha et al., 2014). Th17 cells often aggravate secondary lymphedema by promoting inflammation and fibrosis progression. Although the relationship between LECs and Th17 cells has not been directly discussed in detail in lymphedema, Th17 cells are also a subset of CD4+ T cells, and it can be speculated that LECs may affect the differentiation or function of Th17 cells through a similar mechanism. In collagen-induced arthritis (CIA), the autophagy process of LECs modulates Th17 efflux from the lymph nodes (LNs), a process associated with SphK1 changes resulting in the S1P gradient (Harlé et al., 2021). Current studies on the interaction between Th17 cells and LECs involved in the pathogenesis of lymphedema are still scarce and warrant further study and discussion.
4.4 Th22
Few studies of Th22 cells in lymphedema are available, and the relationship between LECs and Th22 cells has not been discussed in detail. Only one study reported that Th22 cells from patients with filarial lymphedema coexpressed higher levels of IL-22 and IFN-γ than did those from controls (Anuradha et al., 2014). Th22 and Th17 cells are often involved in chronic inflammatory diseases. Therefore, it is important to explore the effects of Th22 cells on LECs in patients with lymphedema. Moreover, focusing on the role of Th17 and Th22 in patients with lymphedema is highly valuable for further understanding the pathogenesis of lymphedema.
4.5 Tregs
The interaction between Tregs and LECs plays an important role in the drainage of lymphatic fluid and the migration of immune cells. Treg numbers are significantly increased in secondary lymphedema tissues in both mice and humans (Gousopo et al., 2016a). Tregs promote the permeability of LECs and enhance the ability of other immune cells to span LECs through LTα1β2-LTβR signaling (Piao et al., 2020). In contrast, LECs are able to regulate PD-L1 migration and inhibit the inflammatory progression of lymphedema by binding to PD-L1 on Tregs (Piao et al., 2022). Importantly, Tregs are involved in the suppression of Th1/Th2 immune responses and the progression of tissue fibrosis to improve the function of lymphatic vessels (Gousopo et al., 2016b). Gkountidi AO et al. reported that IFN-γ upregulated the expression of MHC II in LECs and that MHC II presented antigens to Tregs to inhibit the activity of effector T cells and promote tumor growth (Gkountidi et al., 2021). Although the above reports were conducted in the context of tumor research, the immunomodulatory function of LECs and the interaction of Tregs may also have some reference value in the pathological process of lymphedema. In particular, the role of LECs in immune regulation may provide indirect implications for further understanding the immunological mechanisms of lymphedema.
4.6 Neutrophil
Secondary lymphedema promotes the infiltration of neutrophils and other inflammatory cells, exacerbating tissue fibrosis and chronic inflammatory responses (Zampell et al., 2012). Similarly, the degree of tissue inflammation in an obesity mouse model of lymphedema is strongly associated with increased infiltration of neutrophils (Savetsky et al., 2014). This process is associated with the possible secretion of chemokines (CXCL1/2/3/5) by LECs to attract neutrophils into lymphatic vessels and eventually into draining lymph nodes (dLNs). The synergistic effect of IL-17A and TNF-α promoted the secretion of chemokines by LECs (Ni et al., 2024). Moreover, the site of edema in patients with secondary lymphedema can be accompanied by neutrophilic skin lesions such as Sweet syndrome, which is also associated with the aggregation and infiltration of neutrophils (Guyot-Caquelin et al., 2010). Frueh FS et al. speculated that neutrophil infiltration in the lymph node area is associated with the pathogenesis of lymphedema after ischemia/reperfusion injury (Frueh et al., 2020). However, the interaction between LECs and neutrophils in lymphedema has not been explored in detail in the literature. Further studies on the interaction between LEC and neutrophils are highly valuable for studying the pathogenesis of lymphedema.
4.7 Macrophage
The interaction of LECs and macrophages is very important in the pathological process of secondary lymphedema (Lala et al., 2018; Ji, 2007). With chemokines secreted by LECs, M2 macrophages secrete VEGF-A, VEGF-C and VEGF-D and regulate CD4+ T-cell accumulation and Th2 differentiation to increase lymphangiogenesis and reduce fibrosis. Therefore, promoting macrophage-to-M2 macrophage polarization in tissues with secondary lymphedema may also be an effective therapeutic modality. (Lala et al., 2018). For example, Zhou Z et al. reported that substance P, a neurotransmitter, downregulates the NF-κB/NLRP3 pathway to polarize bone marrow-derived macrophages (BMDMs) into M2 macrophages, thereby alleviating secondary lymphedema (Zhou et al., 2023). Macrophages also play an important role in the pathogenesis of infectious lymphedema. Weinkopff T et al. reported that lymphedema triggered by filarial infection is associated with excretory secretions such as VEGF-A and IL-8, which stimulate macrophages to produce lymphangiogenic factors capable of causing lymphangiectasia to cause secondary lymphedema (Weinkopff et al., 2014). Research on macrophages in lymphedema has focused mainly on how macrophages act with LECs to participate in the generation of lymphatic vessels and the development of lymphedema. Related studies have focused on the roles of macrophages and LECs in signaling pathways, information molecules, and inflammatory mediators. However, the role of macrophages in the pathogenesis of lymphedema is still unclear and requires further study.
4.8 Dendritic cells
There are no reports on how dendritic cells (DCs) interact with LEC in lymphedema. Almost all studies have focused on how DCs influence LECs to participate in the regulation of lymphoid tissue inflammation, lymphatic vessel development, and lymphatic permeability. On the basis of the role of DCs in the pathological process of abnormal lymphatic vessels, it can be speculated that abnormal development and functional inactivation of the lymphatic system are closely associated with the occurrence of lymphedema. LECs are also able to capture and present exogenous antigens via MHC II to DCs, which are able to cross-present antigens to CD8+ T cells (Santambrogio et al., 2019; Kedl et al., 2017). In contrast, the secretion of factors such as TGF-β1 by LECs suppresses the maturation of DCs to reduce inflammation in lymphoid tissues (Christiansen et al., 2016). DCs play an important role in promoting interactions with LECs during the development of lymphoid tissues. Upon IFN-γ stimulation, DCs significantly increase the release of VEGF-C and act on LECs to promote the generation of lymphatic vessels (Gagliostro et al., 2016). Moreover, VEGF-C secreted by DCs enables LECs to secrete chemokines such as CCL21, and CCL21 binds to receptors on DCs such as CCR7 to guide DCs into lymphatic vessels (Permanyer et al., 2018). Furthermore, LECs support the migration of DCs from peripheral tissues to draining lymph nodes by expressing adhesion molecules such as CD112 (Haghayeg et al., 2024). In terms of regulating permeability, DCs regulate lymphatic vessel permeability through specific signaling pathways, such as the CCR7/IRF4 axis and LECs interaction (Permanyer et al., 2018). In this section, we focus on relevant studies and reports regarding the interaction between DCs and LECs, particularly within the inflammatory tissue of lymphatic vessels, with the goal of providing insights into the pathogenesis of lymphedema and its progression.
4.9 Fibroblasts
The fundamental pathology of secondary lymphedema, a significant complication of cancer treatment, is characterized by an imbalance between TGF-β1-mediated fibrosis and lymphangiogenesis (Baik et al., 2022). On the one hand, significant activation of the Smad2 signaling pathway in lymphedema-associated fibroblasts (LAFs) has been shown to induce epithelial‒mesenchymal transition (EMT) and promote the formation of α-smooth muscle actin-positive stress fibers and collagen deposition under specific conditions (Will et al., 2024a). A recent study revealed the critical role of an interactive network of fibroblasts and LECs in the tumor microenvironment in the progression of lymphedema. In patients with cholangiocarcinoma (CCA) who have lymph node metastasis (LNM), cancer-associated fibroblasts (CAFs) promote lymphangiogenesis by activating the ERK1/2-JNK pathway in LECs through the secretion of PDGF-BB (Yan et al., 2024). Furthermore, the recruitment and 3-D assembly of LECs are induced by fibroblasts stimulated with PDGF-D, resulting in increased LEC monolayer permeability (Cadamuro et al., 2019). Oral CAFs have been shown to promote LEC proliferation, migration, invasion, and lumen formation to a greater extent than normal fibroblasts (NFs) do (Chen et al., 2015). In summary, the interaction between fibroblasts, particularly CAFs, and LECs plays a significant role in the regulation of lymphangiogenesis. This cellular communication significantly influences remodeling of the tumor microenvironment and offers novel insights into the mechanisms underlying secondary lymphedema. While the majority of current studies have focused on CAF‒LEC interactions in the context of tumors, these findings highlight potential therapeutic targets for interventions aimed at regulating fibroblast function to either promote or inhibit lymphangiogenesis, thereby addressing the occurrence and development of lymphedema. Therefore, an in-depth study of the communication mechanisms between fibroblasts and LECs may lead to new strategies and direct therapeutic approaches.
4.10 Adipose-derived stem cells
The pathomechanism of lymphedema involves not only interstitial fluid accumulation, but also multidimensional changes such as chronic inflammation, fibrosis, and abnormal fat deposition (Burton et al., 2023). Notably, adipose tissue has a bidirectional relationship with the lymphatic system, in which obesity has been shown to be an independent risk factor. Recent studies have indicated that adipose-derived stem cells (ADSCs) may hold promise in the treatment of lymphedema, with their therapeutic effects potentially manifesting through the activation of lymphatic vessel neogenesis. In vitro investigations have demonstrated that ADSCs significantly promote the proliferation, migration, and tubular structure formation of human dermal LECs (Saijo et al., 2019). However, the direct effect of ADSCs on LECs remains to be elucidated. A study by Takeda K revealed that factors secreted by ADSCs were more effective than recombinant human vascular endothelial growth factor-C in inducing LECs proliferation, migration, and tube formation (Takeda et al., 2015). However, a significant gap remains in our understanding of the intricate cellular communication between adipocytes and LECs. To address this critical knowledge gap, it is imperative to undertake a systematic analysis of the underlying mechanisms governing the interaction between adipocytes and LECs. This analysis should be facilitated by the establishment of an in vitro model that enables coculture of adipocytes and LECs. This model not only provides novel insights into the complex interactions between these cell types but also serves as a valuable platform for the development of novel therapeutic strategies for lymphedema, with a specific focus on adipose metabolism.
5 Maintenance and dynamic regulation of the lymphatic system microenvironment
5.1 Cell junctions
5.1.1 Connexins
Gap junctions (GJs), which are intercellular channels composed of connexin (Cx) subunits, enable cell-to-cell communication through the direct cytoplasmic exchange of ions and small molecules (Söhl and Willecke, 2004) (Table 3). Among these cell junctions, connexins (Cxs), GJ proteins, have been shown to play a key role in lymphoid development in vertebrates. Many studies suggest that Cx-associated lymphedema is caused by asynchronous contraction of the lymphatic vessels (Castorena-Gonzalez et al., 2018). The absence of different Cxs, including Cx 26, Cx 37, Cx 43, and Cx 47, has also been confirmed in many studies of lymphedema. For example, loss of expression of GJB2, encoding Cx26, suppresses peripheral lymphangiogenesis and causes severe lymphedema in mouse models (Dicke et al., 2011). In addition, Ostergaard P et al. and Ferrell RE et al. reported that mutations in the GJC2 gene encoding Cx47 were associated with hereditary primary lymphedema (Ferrell et al., 2010; Ostergaard et al., 2011). A study by Finegold DN revealed a positive association between the risk of secondary lymphedema after breast cancer treatment (Finegold et al., 2012). Brice G et al. confirmed through Sanger sequencing that a new mutation in the GJA1 gene encoding the Cx43 mutation an leads to oculodentodigital syndrome and primary lymphedema (Brice et al., 2013). Cx43 has a limited role in the lymphatic endothelium, and one study using a knockout mouse model revealed that although Cx43 is widespread in the endothelial layer of lymphatic vessels, its absence does not affect rhythmic contraction or calcium signaling synchronization in lymphatic vessels. It is Cx45, not Cx43, that truly regulates contraction rhythms (Castorena-Gonzalez et al., 2018). Davis MJ and colleagues first reported that the expression of CX45 in mouse lymph endothelial cells is essential for the normal function of the lymphatic system. Unfortunately, there are no reports of human lymphedema associated with Cx45 (GJC1) mutations. This may be because loss-of-function mutations in human Cx45 are potentially lethal and are thus not detected in primary lymphedema clinical screening through whole-exome sequencing (Davis et al., 2024). Kanady JD et al. reported that FOXC2 and Cx37 jointly control lymph valve development (Kanady et al., 2015). In addition, defects in Cx37 and Cx43 have been shown to cause primary lymphedema and chylothorax (Kanady et al., 2011). We currently have limited knowledge of the functional relationships and signaling pathways of Cxs with other proteins. At present, the study of the interaction between Cxs and other proteins can provide a deeper understanding of developmental disorders and the missing functions of LECs, establishing a basis for further exploration of the pathogenesis of lymphedema and therapeutic measures.
TABLE 3
| Types of lymphedema | Category | Mechanisms of action | Data sources |
|---|---|---|---|
| Primary | Cx 26 (GJB2) (Dicke et al., 2011) | Related to lymphatic vessel formation | Mouse model |
| Primary and secondary | Cx 37 (GJA4) (Kanady et al., 2015; Kanady et al., 2011; Hadizadeh et al., 2018) | Related to lymphatic vessel growth and lymphatic valve development Mutations that are associated with secondary lymphedema in breast cancer patients after surgery | Mouse model Patient |
| Primary | Cx 43 (GJA1) (Castorena-Gonzalez et al., 2018; Brice et al., 2013) | Mutations that cause abnormal valve development and/or reduced valve density leading to lymphatic transport disorders | Mouse model Patient |
| Primary and secondary | Cx 47 (GJC2) (Ferrell et al., 2010; Ostergaard et al., 2011) | Mutations that cause abnormal valve development and/or reduced valve density leading to lymphatic transport disorders Mutations that are associated with secondary lymphedema following breast cancer treatment | Mouse model Patient |
| Nothing found | Cx45 (GJC1) (Davis et al., 2024) | Essential for lymphatic valve function Cx45 loss-of-function mutations may be lethal | Mouse model |
| secondary | ROCK2/JAM-A (Lee et al., 2023) | Specific knockdown of ROCK2 in the lymphatic system can reverse lymphedema | Mouse model LECs |
| Primary | Cadherin 4 (FAT4) (Betterman et al., 2020) | Mutations in calmodulin FAT4 cause Hennekam syndrome, one of the characteristics of which is lymphoedema | Patient |
| Primary | VE-cadherin (Yang et al., 2019; Cha et al., 2016) | VE-cadherin can bind to β-catenin, which is essential for the development of lymphatic valves | Mouse model |
| Nothing found | Claudin 5 (Frye et al., 2020; Mahamud et al., 2019) | The localization and stability of CLDN5 may affect lymphatic vascular permeability | Mouse model HLECs |
Cell junctions in lymphedema.
5.1.2 JAM-A
Junctional adhesion molecules (JAMs) are a family of adhesion molecule glycoproteins that are expressed on human LECs. There is a paucity of literature on the association between JAM and lymphedema (Ueki et al., 2008). Lee et al. successfully induced lymphedema by the cytokine-induced ROCK2/JAM-A complex in a 3D bionic model of lymphatic vessels. Lymphatic-specific knockdown of ROCK2 reverses lymphedema in vivo and could serve as a potential therapeutic target for lymphedema (Lee et al., 2023). The role of JAM in the pathogenesis and treatment of lymphedema deserves further investigation and discussion.
5.1.3 Cadherin
VE-cadherin-mediated cellular connectivity and signaling regulatory networks have been demonstrated to play central roles in lymphedema development (Hägerling et al., 2018; Frye et al., 2015). VE-cadherin has been shown to bind to β-catenin, thereby maintaining Prox1, FOXC2, and GATA2 transcriptional regulation, which is required for lymphatic valve development (Yang et al., 2019; Cha et al., 2016). In addition, VE-cadherin was found to regulate valve formation through VEGFR2/3-AKT signaling and to synergistically maintain endothelial junction integrity with uPARAP (Coon et al., 2015; Gucciardo et al., 2025). GATA2, a key transcription factor, not only regulates VE-cadherin and claudin 5 (CLDN5) expression, but also its own expression is regulated by β-catenin signaling, suggesting that a bidirectional regulatory mechanism may be involved in this pathological process (Mahamud et al., 2019). Notably, mutations in the atypical calcineurin protein FAT4 have been identified in patients with Hennekam syndrome, which is characterized by lymphedema as one of its features. This finding underscores the critical role of FAT4 in the lymphovascular system (Betterman et al., 2020). Although the direct relationship between cadherins and lymphedema is not fully understood, recent studies have begun to reveal changes in cadherin localization and molecular expression that may act as potential triggers for lymphedema. For example, VE-cadherin-dependent signaling is critical for lymphatic valve formation and maintenance, and therapies that enhance downstream pathways have the potential to treat lymphedema. Therefore, future studies should investigate the specific role of cadherins in the pathogenesis of lymphedema to identify new intervention targets that could advance the early diagnosis and treatment of this disease.
5.1.4 Claudin
Claudin 5 (CLDN5), a pivotal molecule in cell junctions, is regulated by both GATA2 and EphrinB2/EphB4 signaling, suggesting a potential role in edema formation through its impact on lymphatic endothelial barrier function. GATA2 mutations disrupt cellular junctions by downregulating CLDN5 and VE-cadherin. In contrast, the EphrinB2/EphB4-Rac1/Rho pathway has been demonstrated to dynamically regulate the localization and stability of CLDN5, suggesting that targeting this pathway may selectively regulate lymphatic vessel permeability (Frye et al., 2020; Mahamud et al., 2019). Although there is limited evidence for a direct association between CLDN5 and lymphedema, its aberrant expression has been recognized as a potential pathogenetic factor. In the future, the spatiotemporal regulatory network of CLDN5, as well as the mechanism of synergistic maintenance of the lymphatic barrier with VE-cadherin, must be analyzed, which will provide a basis for precise intervention.
5.2 Oxidative stress in lymphedema
Oxidative stress has been demonstrated to affect normal lymphatic function and exacerbate secondary lymphedema. Aging has been demonstrated to reduce the level of antioxidant enzymes (SODs) and increase superoxide anion levels in lymphatic vessels, thereby promoting chronic inflammation and weakening lymphatic vessel contractility (Pal et al., 2017). The level of oxidative stress in lymphedema is significantly elevated in pathological states. The accumulation of substantial quantities of lymphatic fluid within the affected tissues can result in local tissue hypoxia, which, in turn, can trigger excessive oxidative stress, thereby establishing a self-perpetuating cycle (Siems et al., 2002; Tabibiazar et al., 2006). The inhibition of cyclooxygenase (COX) has been demonstrated to significantly reduce edema and lower free radical levels in a mouse tail lymphedema model, suggesting the potential of antioxidant strategies in lymphedema treatment (Chang et al., 2013). In clinical trials, sodium selenite, an antioxidant, has been shown to reduce ROS production and mitigate symptoms of lymphedema. Furthermore, the observation of enhanced serum markers of oxidative stress in patients undergoing lymphatic venous anastomosis (LVA) and vascularized lymph node flap transfer supports the efficacy of alleviating oxidative stress in reducing lymphatic loading (Yang et al., 2021b). Consequently, interventions that target oxidative stress, such as the utilization of antioxidants and the enhancement of oxidative stress levels after surgical intervention, may offer novel approaches for the management of lymphedema. These interventions are also clinically significant, particularly in reducing the lymphatic load and enhancing lymphatic function.
6 Treatment
The management of lymphedema, a complex chronic disease, continues to face significant challenges, particularly in the context of primary lymphedema due to delayed diagnosis and heterogeneous progression patterns. The clinical management of secondary lymphedema mainly follows a framework based on core strategies, including complete decongestive therapy (CDT) and microsurgical techniques such as lymphovenous anastomosis (Table 4). However, the evidence supporting the long-term efficacy of these interventions remains to be thoroughly substantiated (Senger et al., 2023). In the context of secondary lymphedema, although recombinant VEGF-C and AAV gene therapies have been shown to significantly improve edema in animal models, their clinical application is limited by targeted delivery efficiency and high cost (Szuba et al., 2002; Tammela et al., 2007; Leppäpuska et al., 2022). In contrast, small-molecule drugs such as second-generation tetracyclines exhibit strong clinical translational potential through the dual mechanism of activating LECs proliferation and inducing M2 macrophage polarization (Furlong-Silva et al., 2021). Furthermore, the topical application of pirfenidone (PFD) has been demonstrated to inhibit TGF-β1 signaling, thereby reducing the degree of fibrosis and preventing the development of lymphedema (Baik et al., 2022). However, the utilization of these medications can result in a range of adverse effects, including gastrointestinal reactions and lipid metabolism disorders. Among natural products, Cynanchum atratum extract CAPW-1 promotes lymphangiogenesis through the TLR4/NF-κB pathway and has a synergistic effect with VEGF-C (Li et al., 2024). S R Narahari argued that the “integrative medicine” model promoted in India, which combines traditional and modern approaches and focuses on the quality of life of the patient, is an effective, low-cost way of managing lymphedema and could serve as a model for disability prevention of chronic diseases in developing countries (Narahari and Ryan, 2020). He also provided clear clinical evidence that an integrative medicine program integrating Ayurveda, yoga, anti-infective measures and compression therapy is effective in improving lymphedema, reducing infections, and improving quality of life in patients with lymphatic filariasis and that it is low cost and suitable for gRAS-root dissemination (Narahari et al., 2023). The combination of the ultramicrosurgical technique and ICG lymphangiography has yielded substantial limb volume reduction in postoperative patients. The underlying mechanism of this efficacy is closely associated with oxidative stress modulation (Will et al., 2024b). Future research endeavors must integrate single-cell genomics to elucidate the heterogeneity of LECs, develop engineered biomaterials, and construct AI predictive models to optimize individualized treatments. Ultimately, the aim of these efforts is to achieve a paradigm shift from symptom management to pathological reversal through multitarget regulatory networks.
TABLE 4
| Type of treatment | Specific interventions | Mechanism of action | Type of application | Dominance | Limitations |
|---|---|---|---|---|---|
| Traditional core therapy | Complete CDT (Senger et al., 2023) | Physical reduction (multilayer bandage/massage) | Primary secondary | International gold standard with clear short-term results | Insufficient evidence of long-term efficacy, lifelong maintenance therapy required |
| Microsurgical technique | Lymphatic vascular anastomosis (LVA) (Yang et al., 2021b) | Establishment of lymphatic bypass drainage | Secondary | Precision repair with small incisions | Dependent on operator experience; long-term results to be proven |
| Biological preparation | Recombinant VEGF-C/AAV gene therapy (Szuba et al., 2002; Tammela et al., 2007; Leppäpuska et al., 2022) | Promotes lymphatic neovascularization | Animal model | Significant improvement in edema | Inefficient targeted delivery; costly |
| Small-molecule drug | Second generation tetracyclines (Furlong-Silva et al., 2021) | Activates LEC proliferation and induces M2 macrophage polarization | Secondary | Dual mechanism with strong potential for clinical translation | Gastrointestinal reactions, lipid metabolism disorders and other side effects |
| Topical application of PFD (Baik et al., 2022) | Inhibition of TGF-β1 signaling pathway, anti-fibrosis | Prophylactic | Stopping the progression of fibrosis | Skin irritation, photosensitivity | |
| Natural product | CAPW-1 (Li et al., 2024) | TLR4/NF-κB pathway promotes lymphangiogenesis + synergizes with VEGF-C | Experimental research | Natural source, multitarget synergy | Lack of clinical data |
| Integrative medicine | Ayurvedic herbs/yoga/step pressure therapy (Narahari and Ryan, 2020; Narahari et al., 2023) | Integrated physical and phytotherapy | Resource-limited areas | Low cost, reduced infections, improved quality of life | Difficulty in standardization, wide individual variation in efficacy |
| Technology innovation portfolio | Ultra microscopy + ICG lymphography (Will et al., 2024b) | Precision targeting + oxidative stress regulation | Secondary | Significant limb volume reduction | High technical requirements, high equipment dependency |
The related treatments for lymphedema.
7 Conclusion and discussion
Lymphedema is a complex chronic disease, and its pathological mechanism involves genetic variations, signaling pathway abnormalities, dysregulated intercellular communication, immune microenvironmental disorders, and oxidative stress imbalance at multiple levels. This review methodically summarizes the molecular regulatory network of lymphedema-related signaling pathways and reveals the central roles of key pathways, such as the RAS/MAPK, PI3K/AKT, VEGF-C/VEGFR-3, and HGF/MET pathways, in the development, functional maintenance, and pathological damage of lymphatic vessels. For example, aberrant activation of the RAS/MAPK pathway has been closely associated with hereditary lymphedema, such as NS. Conversely, dysregulation of the VEGF-C/VEGFR-3 axis has been identified as a significant driver of primary lymphedema, including Milroy disease. Furthermore, TGF-β1 signaling has been shown to exacerbate the progression of secondary lymphedema by promoting fibrosis and inflammatory responses. In addition, the vicious cycle of oxidative stress and chronic inflammation has been demonstrated to further deteriorate lymphatic function. Notably, mutations in GJ proteins, such as Cx37 and Cx47, have been shown to result in impaired lymphatic return by interfering with the synchronization of lymphatic vasoconstriction. This provides a novel perspective for understanding abnormal lymphatic vessel dynamics.
At the level of cellular interactions, the dynamic interactions of LECs with immune cells, such as Th1, Th2, Treg, and macrophages, constitute a central feature of the lymphedema microenvironment. For example, IL-4 and IL-13, which are secreted by Th2 cells, have been shown to inhibit lymphangiogenesis by inducing LEC apoptosis. In contrast, M2 macrophage polarization has been shown to promote compensatory lymphangiogenesis through the secretion of VEGF-C. These findings contribute to a more comprehensive understanding of the immunopathological mechanisms underlying lymphedema and establish a theoretical foundation for targeted immunomodulation as a therapeutic approach for this condition.
The existing therapeutic interventions for lymphedema and the current state of research in this domain are inherently limited and encumbered by significant challenges. Clinical interventions, including compression therapy and surgical procedures, are effective in mitigating symptoms. However, they do not address the underlying pathophysiology. Furthermore, there is a paucity of precise treatment regimens that are tailored to specific molecular typing profiles, which is crucial for the management of this condition. In the context of individual signaling pathways, single-target interventions may be inadequate for reversing the pathological process. Consequently, there is an imperative need to explore synergistic regulatory strategies that involve multiple pathways. However, notably, the majority of signaling pathways exhibit cross-talk, a phenomenon exemplified by PI3K/AKT and RAS/MAPK. This observation underscores the existence of substantial research potential and a notable gap in the existing knowledge. Moreover, the majority of extant studies are predicated on animal models, and the heterogeneity of primary versus secondary lymphedema may limit the generalizability of therapeutic strategies. By simulating lymphatic vessel–immune cell interactions, targeting antioxidant–promoting lymphatic regeneration, and remodeling the intralymphatic immune–metabolic microenvironment, further research into the mechanism of lymphedema and exploring therapeutic strategies are highly important. It is anticipated that this study will generate novel research concepts and insights relevant to targeted therapy for lymphedema.
In conclusion, it is imperative that research on lymphedema integrate the exploration of molecular mechanisms with the innovation of therapeutic modalities. In the future, the therapeutic bottlenecks currently experienced by lymphedema patients may be overcome through targeted regulation of oxidative stress, multipathway interventions, and precision medicine strategies. Such strategies may improve lymphatic function and reverse the pathological process, thereby improving the quality of life for these patients.
Statements
Author contributions
QZ: Writing – original draft. ZN: Writing – original draft. YP: Writing – review and editing. YH: Writing – review and editing. YM: Writing – review and editing. JZ: Writing – review and editing. JD: Funding acquisition, Project administration, Writing – review and editing. YY: Conceptualization, Funding acquisition, Supervision, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the Science and Technology Research Project of the Education Department of Jilin Province, grant number JJKH20220973KJ; the Joint Project of China-Japan Union Hospital and Basic Medical College of Jilin University, grant number KYXZ2022JC01.
Acknowledgments
We appreciate the encouragement and helpful comments from other members of the Department of Cell Biology and Medical Genetics, College of Basic Medical Sciences, Jilin University.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
AdamsR. H.AlitaloK. (2007). Molecular regulation of angiogenesis and lymphangiogenesis. Nat. Rev. Mol. Cell Biol.8 (6), 464–478. 10.1038/nrm2183
2
AgollahG. D.Gonzalez-GarayM. L.RasmussenJ. C.TanI. C.AldrichM. B.DarneC.et al (2014). Evidence for SH2 domain-containing 5'-inositol phosphatase-2 (SHIP2) contributing to a lymphatic dysfunction. PLoS One9 (11), e112548. 10.1371/journal.pone.0112548
3
AlpaslanM.FastréE.MestreS.van HaeringenA.RepettoG. M.KeymolenK.et al (2024). Pathogenic variants in HGF give rise to childhood-to-late onset primary lymphoedema by loss of function. Hum. Mol. Genet.33 (14), 1250–1261. 10.1093/hmg/ddae060
4
AnuradhaR.GeorgeP. J.ChandrasekaranV.KumaranP. P.NutmanT. B.BabuS. (2014). Interleukin 1 (IL-1)- and IL-23-mediated expansion of filarial antigen-specific Th17 and Th22 cells in filarial lymphedema. Clin. Vaccine Immunol.21 (7), 960–965. 10.1128/CVI.00257-14
5
AuA. C.HernandezP. A.LieberE.NadrooA. M.ShenY.-M.KelleyK. A.et al (2010). Protein tyrosine phosphatase PTPN14 is a regulator of lymphatic function and choanal development in humans. Am. J. Hum. Genet.87 (3), 436–444. 10.1016/j.ajhg.2010.08.008
6
AvrahamT.DaluvoyS.ZampellJ.YanA.HavivY. S.RocksonS. G.et al (2010). Blockade of transforming growth factor-beta1 accelerates lymphatic regeneration during wound repair. Am. J. Pathol.177 (6), 3202–3214. 10.2353/ajpath.2010.100594
7
AvrahamT.ZampellJ. C.YanA.ElhadadS.WeitmanE. S.RocksonS. G.et al (2013). Th2 differentiation is necessary for soft tissue fibrosis and lymphatic dysfunction resulting from lymphedema. FASEB J.27 (3), 1114–1126. 10.1096/fj.12-222695
8
BabuS.AnuradhaR.KumarN. P.GeorgeP. J.KumaraswamiV.NutmanT. B. (2011). Filarial lymphatic pathology reflects augmented toll-like receptor-mediated, mitogen-activated protein kinase-mediated proinflammatory cytokine production. Infect. Immun.79 (11), 4600–4608. 10.1128/IAI.05419-11
9
BabuS.BhatS. Q.Pavan KumarN.LipiraA. B.KumarS.KarthikC.et al (2009). Filarial lymphedema is characterized by antigen-specific Th1 and th17 proinflammatory responses and a lack of regulatory T cells. PLoS Negl. Trop. Dis.3 (4), e420. 10.1371/journal.pntd.0000420
10
BabuS.BlauveltC. P.KumaraswamiV.NutmanT. B. (2006). Regulatory networks induced by live parasites impair both Th1 and Th2 pathways in patent lymphatic filariasis: implications for parasite persistence. J. Immunol.176 (5), 3248–3256. 10.4049/jimmunol.176.5.3248
11
BaikJ. E.ParkH. J.KataruR. P.SavetskyI. L.LyC. L.ShinJ.et al (2022). TGF-β1 mediates pathologic changes of secondary lymphedema by promoting fibrosis and inflammation. Clin. Transl. Med.12 (6), e758. 10.1002/ctm2.758
12
BalukP.FuxeJ.HashizumeH.RomanoT.LashnitsE.ButzS.et al (2007). Functionally specialized junctions between endothelial cells of lymphatic vessels. J. Exp. Med.204 (10), 2349–2362. 10.1084/jem.20062596
13
BettermanK. L.SuttonD. L.SeckerG. A.KazenwadelJ.OszmianaA.LimL.et al (2020). Atypical cadherin FAT4 orchestrates lymphatic endothelial cell polarity in response to flow. J. Clin. Invest130 (6), 3315–3328. 10.1172/JCI99027
14
BollingerA.PartschH. (1985). The initial lymphatics: new methods and findings: international symposium Zurich 1984.
15
BonettiG.PaolacciS.SamajaM.MalteseP. E.MicheliniS.MicheliniS.et al (2022). Low efficacy of genetic tests for the diagnosis of primary lymphedema prompts novel insights into the underlying molecular pathways. Int. J. Mol. Sci.23 (13), 7414. 10.3390/ijms23137414
16
BosF. L.CauntM.Peterson-MaduroJ.Planas-PazL.KowalskiJ.KarpanenT.et al (2011). CCBE1 is essential for mammalian lymphatic vascular development and enhances the lymphangiogenic effect of vascular endothelial growth factor-C in vivo. Circ. Res.109 (5), 486–491. 10.1161/CIRCRESAHA.111.250738
17
BowmanC.RocksonS. G. (2024). The role of inflammation in lymphedema: a narrative review of pathogenesis and opportunities for therapeutic intervention. Int. J. Mol. Sci.25 (7), 3907. 10.3390/ijms25073907
18
BrandK.KentschH.GlashoffC.RosenbergerG. (2014). RASopathy-associated CBL germline mutations cause aberrant ubiquitylation and trafficking of EGFR. Hum. Mutat.35 (11), 1372–1381. 10.1002/humu.22682
19
BriceG.OstergaardP.JefferyS.GordonK.MortimerP. S.MansourS. (2013). A novel mutation in GJA1 causing oculodentodigital syndrome and primary lymphoedema in a three generation family. Clin. Genet.84 (4), 378–381. 10.1111/cge.12158
20
BrixB.SeryO.OnoratoA.UreC.RoesslerA.GoswamiN.et al (2021). Hemodynamic responses in lower limb lymphedema patients undergoing physical therapy. Biol. (Basel).10 (4), 642. 10.3390/biology10070642
21
BrouillardP.BoonL.VikkulaM. (2014). Genetics of lymphatic anomalies. J. Clin. Invest124 (3), 898–904. 10.1172/JCI71614
22
BrouillardP.DupontL.HelaersR.CoulieR.TillerG. E.PeedenJ.et al (2017). Loss of ADAMTS3 activity causes hennekam lymphangiectasia-lymphedema syndrome 3. Hum. Mol. Genet.26 (21), 4095–4104. 10.1093/hmg/ddx297
23
BuiK.HongY.-K. (2020). Ras pathways on Prox1 and lymphangiogenesis: insights for therapeutics. Front. Cardiovasc Med.7, 597374. 10.3389/fcvm.2020.597374
24
BurtonJ. S.SlettenA. C.MarshE.WoodM. D.SacksJ. M. (2023). Adipose tissue in lymphedema: a central feature of pathology and target for pharmacologic therapy. Lymphat. Res. Biol.21 (1), 2–7. 10.1089/lrb.2022.0003
25
CadamuroM.BrivioS.MertensJ.VismaraM.MoncsekA.MilaniC.et al (2019). Platelet-derived growth factor-D enables liver myofibroblasts to promote tumor lymphangiogenesis in cholangiocarcinoma. J. Hepatol.70 (4), 700–709. 10.1016/j.jhep.2018.12.004
26
CartaC.PantaleoniF.BocchinfusoG.StellaL.VastaI.SarkozyA.et al (2006). Germline missense mutations affecting KRAS isoform B are associated with a severe Noonan syndrome phenotype. Am. J. Hum. Genet.79 (1), 129–135. 10.1086/504394
27
CastelP.ChengA.Cuevas-NavarroA.EvermanD. B.PapageorgeA. G.SimanshuD. K.et al (2019). RIT1 oncoproteins escape LZTR1-mediated proteolysis. Science363 (6432), 1226–1230. 10.1126/science.aav1444
28
CastenholzA. (1984). Morphological characteristics of initial lymphatics in the tongue as shown by scanning electron microscopy. Scan Electron Microsc. (Pt 3), 1343–1352.
29
Castorena-GonzalezJ. A.ZawiejaS. D.LiM.SrinivasanR. S.SimonA. M.de WitC.et al (2018). Mechanisms of connexin-related lymphedema. Circ. Res.123 (8), 964–985. 10.1161/CIRCRESAHA.117.312576
30
ChaB.GengX.MahamudM. R.FuJ.MukherjeeA.KimY.et al (2016). Mechanotransduction activates canonical Wnt/β-catenin signaling to promote lymphatic vascular patterning and the development of lymphatic and lymphovenous valves. Genes Dev.30 (12), 1454–1469. 10.1101/gad.282400.116
31
ChaB.GengX.MahamudM. R.ZhangJ. Y.ChenL.KimW.et al (2018). Complementary wnt sources regulate lymphatic vascular development via PROX1-Dependent Wnt/β-Catenin signaling. Cell Rep.25 (3), 571–584. 10.1016/j.celrep.2018.09.049
32
ChanT. O.RodeckU.ChanA. M.KimmelmanA. C.RittenhouseS. E.PanayotouG.et al (2002). Small GTPases and tyrosine kinases coregulate a molecular switch in the phosphoinositide 3-kinase regulatory subunit. Cancer Cell1 (2), 181–191. 10.1016/s1535-6108(02)00033-8
33
ChangT.-C.UenY.-H.ChouC.-H.SheuJ.-R.ChouD.-S. (2013). The role of cyclooxygenase-derived oxidative stress in surgically induced lymphedema in a mouse tail model. Pharm. Biol.51 (5), 573–580. 10.3109/13880209.2012.749923
34
ChenS.GaoP.ChangZ.XuanM. (2015). Effects of oral cancer-associated fibroblasts on the proliferation, migration, invasion and tube formation to human lymphatic endothelial cells. Hua Xi Kou Qiang Yi Xue Za Zhi33 (5), 524–528. 10.7518/hxkq.2015.05.018
35
CheonH.GelvosaM. N.KimS. A.SongH. Y.JeonJ. Y. (2023). Lymphatic channel sheet of polydimethylsiloxane for preventing secondary lymphedema in the rat upper limb model. Bioeng. Transl. Med.8 (1), e10371. 10.1002/btm2.10371
36
ChoS.RohK.ParkJ.ParkY. S.LeeM.ChoS.et al (2017). Hydrolysis of hyaluronic acid in lymphedematous tissue alleviates fibrogenesis via TH1 cell-mediated cytokine expression. Sci. Rep.7 (1), 35. 10.1038/s41598-017-00085-z
37
ChoiD.ParkE.YuR. P.CooperM. N.ChoI.-T.ChoiJ.et al (2022). Piezo1-Regulated mechanotransduction controls flow-activated lymphatic expansion. Circ. Res.131 (2), e2–e21. 10.1161/CIRCRESAHA.121.320565
38
ChoiD.RamuS.ParkE.JungE.YangS.JungW.et al (2016). Aberrant activation of notch signaling inhibits PROX1 activity to enhance the malignant behavior of thyroid cancer cells. Cancer Res.76 (3), 582–593. 10.1158/0008-5472.CAN-15-1199
39
ChristiansenA. J.DieterichL. C.OhsI.BachmannS. B.BianchiR.ProulxS. T.et al (2016). Lymphatic endothelial cells attenuate inflammation via suppression of dendritic cell maturation. Oncotarget7 (26), 39421–39435. 10.18632/oncotarget.9820
40
CirsteaI. C.KutscheK.DvorskyR.GremerL.CartaC.HornD.et al (2010). A restricted spectrum of NRAS mutations causes Noonan syndrome. Nat. Genet.42 (1), 27–29. 10.1038/ng.497
41
ConnerA. C.BillR. M.ConnerM. T. (2013). An emerging consensus on aquaporin translocation as a regulatory mechanism. Mol. Membr. Biol.30 (1), 1–12. 10.3109/09687688.2012.743194
42
CoonB. G.BaeyensN.HanJ.BudathaM.RossT. D.FangJ. S.et al (2015). Intramembrane binding of VE-cadherin to VEGFR2 and VEGFR3 assembles the endothelial mechanosensory complex. J. Cell Biol.208 (7), 975–986. 10.1083/jcb.201408103
43
CoriddiM.WeeC.MeyersonJ.EifermanD.SkorackiR. (2017). Vascularized jejunal mesenteric lymph node transfer: a novel surgical treatment for extremity lymphedema. J. Am. Coll. Surg.225 (5), 650–657. 10.1016/j.jamcollsurg.2017.08.001
44
CormierJ. N.AskewR. L.MungovanK. S.XingY.RossM. I.ArmerJ. M. (2010). Lymphedema beyond breast cancer: a systematic review and meta-analysis of cancer-related secondary lymphedema. Cancer.116 (22), 5138–5149. 10.1002/cncr.25458
45
CosoS.ZengY.OpeskinK.WilliamsE. D. (2012). Vascular endothelial growth factor receptor-3 directly interacts with phosphatidylinositol 3-kinase to regulate lymphangiogenesis. PLoS One7 (6), e39558. 10.1371/journal.pone.0039558
46
DaiC.WenX.HeW.LiuY. (2011). Inhibition of proinflammatory RANTES expression by TGF-beta1 is mediated by glycogen synthase kinase-3beta-dependent beta-catenin signaling. J. Biol. Chem.286 (9), 7052–7059. 10.1074/jbc.M110.174821
47
DavisM. J.Castorena-GonzalezJ. A.LiM.ZawiejaS. D.SimonA. M.GengX.et al (2024). Connexin-45 is expressed in mouse lymphatic endothelium and required for lymphatic valve function. JCI Insight9 (16), e169931. 10.1172/jci.insight.169931
48
DayanJ. H.LyC. L.KataruR. P.MehraraB. J. (2018). Lymphedema: pathogenesis and novel therapies. Annu. Rev. Med.69, 263–276. 10.1146/annurev-med-060116-022900
49
DiS.ZiyouY.LiuN.-F. (2016). Pathological changes of lymphedematous skin: increased mast cells, related proteases, and activated transforming growth Factor-β1. Lymphat. Res. Biol.14 (3), 162–171. 10.1089/lrb.2016.0010
50
DickeN.PielenstickerN.DegenJ.HeckerJ.TressO.BaldT.et al (2011). Peripheral lymphangiogenesis in mice depends on ectodermal connexin-26 (Gjb2). J. Cell Sci.124 (Pt 16), 2806–2815. 10.1242/jcs.084186
51
DiSipioT.RyeS.NewmanB.HayesS. (2013). Incidence of unilateral arm lymphoedema after breast cancer: a systematic review and meta-analysis. Lancet Oncol.14 (6), 500–515. 10.1016/S1470-2045(13)70076-7
52
DucoliL.DetmarM. (2021). Beyond PROX1: transcriptional, epigenetic, and noncoding RNA regulation of lymphatic identity and function. Dev. Cell56 (4), 406–426. 10.1016/j.devcel.2021.01.018
53
EkvallS.WilbeM.DahlgrenJ.LegiusE.van HaeringenA.WestphalO.et al (2015). Mutation in NRAS in familial noonan syndrome--case report and review of the literature. BMC Med. Genet.16, 95. 10.1186/s12881-015-0239-1
54
EngelJ.KerrJ.Schlesinger-RaabA.SauerH.HölzelD. (2003). Axilla surgery severely affects quality of life: results of a 5-year prospective study in breast cancer patients. Breast Cancer Res. Treat.79 (1), 47–57. 10.1023/a:1023330206021
55
FerrellR. E. (2002). Research perspectives in inherited lymphatic disease. Ann. N. Y. Acad. Sci.979, 39–51. 10.1111/j.1749-6632.2002.tb04866.x
56
FerrellR. E.BatyC. J.KimakM. A.KarlssonJ. M.LawrenceE. C.Franke-SnyderM.et al (2010). GJC2 missense mutations cause human lymphedema. Am. J. Hum. Genet.86 (6), 943–948. 10.1016/j.ajhg.2010.04.010
57
FimboA. M.MinziO. M. S.MmbandoB. P.BarryA.NkayambaA. F.MwamwitwaK. W.et al (2020). Prevalence and correlates of lymphatic filariasis infection and its morbidity following mass ivermectin and albendazole administration in mkinga district, north-eastern Tanzania. J. Clin. Med.9 (5), 1550. 10.3390/jcm9051550
58
FinegoldD. N.BatyC. J.KnickelbeinK. Z.PerschkeS.NoonS. E.CampbellD.et al (2012). Connexin 47 mutations increase risk for secondary lymphedema following breast cancer treatment. Clin. Cancer Res.18 (8), 2382–2390. 10.1158/1078-0432.CCR-11-2303
59
FinegoldD. N.SchachtV.KimakM. A.LawrenceE. C.FoeldiE.KarlssonJ. M.et al (2008). HGF and MET mutations in primary and secondary lymphedema. Lymphat. Res. Biol.6 (2), 65–68. 10.1089/lrb.2008.1524
60
FotiouE.Martin-AlmedinaS.SimpsonM. A.LinS.GordonK.BriceG.et al (2015). Novel mutations in PIEZO1 cause an autosomal recessive generalized lymphatic dysplasia with non-immune Hydrops fetalis. Nat. Commun.6, 8085. 10.1038/ncomms9085
61
Fritz-SixK. L.DunworthW. P.LiM.CaronK. M. (2008). Adrenomedullin signaling is necessary for murine lymphatic vascular development. J. Clin. Invest118 (1), 40–50. 10.1172/JCI33302
62
FruehF. S.JelvaniB.ScheuerC.KörbelC.KimB.-S.GiovanoliP.et al (2020). Short-term molecular and cellular effects of ischemia/reperfusion on vascularized lymph node flaps in rats. PLoS One15 (10), e0239517. 10.1371/journal.pone.0239517
63
FryeM.DierkesM.KüppersV.VockelM.TommJ.ZeuschnerD.et al (2015). Interfering with VE-PTP stabilizes endothelial junctions in vivo via Tie-2 in the absence of VE-cadherin. J. Exp. Med.212 (13), 2267–2287. 10.1084/jem.20150718
64
FryeM.StrittS.OrtsäterH.Hernandez VasquezM.KaakinenM.VicenteA.et al (2020). EphrinB2-EphB4 signalling provides rho-mediated homeostatic control of lymphatic endothelial cell junction integrity. Elife9, e57732. 10.7554/eLife.57732
65
FuA.LiuC. (2023). The function of T cell immunity in lymphedema: a comprehensive review. Lymphat. Res. Biol.21 (6), 556–564. 10.1089/lrb.2023.0002
66
Furlong-SilvaJ.CrossS. D.MarriottA. E.PionnierN.ArcherJ.StevenA.et al (2021). Tetracyclines improve experimental lymphatic filariasis pathology by disrupting interleukin-4 receptor-mediated lymphangiogenesis. J. Clin. Invest131 (5), e140853. 10.1172/JCI140853
67
GagliostroV.SeegerP.GarrafaE.SalviV.BrescianiR.BosisioD.et al (2016). Pro-lymphangiogenic properties of IFN-γ-activated human dendritic cells. Immunol. Lett.173, 26–35. 10.1016/j.imlet.2016.03.008
68
GkountidiA. O.GarnierL.DubrotJ.AngelilloJ.HarléG.BrighouseD.et al (2021). MHC class II antigen presentation by lymphatic endothelial cells in tumors promotes intratumoral regulatory T cell-suppressive functions. Cancer Immunol. Res.9 (7), 748–764. 10.1158/2326-6066.CIR-20-0784
69
GoberdhanD. C. I.WilsonC. (2003). PTEN: tumour suppressor, multifunctional growth regulator and more. Hum. Mol. Genet.12 (Spec No 2), R239–R248. 10.1093/hmg/ddg288
70
GordonK.SpidenS. L.ConnellF. C.BriceG.CottrellS.ShortJ.et al (2013). FLT4/VEGFR3 and milroy disease: novel mutations, a review of published variants and database update. Hum. Mutat.34 (1), 23–31. 10.1002/humu.22223
71
GousopoulosE.ProulxS. T.BachmannS. B.SchollJ.DionyssiouD.DemiriE.et al (2016a). Regulatory T cell transfer ameliorates lymphedema and promotes lymphatic vessel function. JCI Insight1 (16), e89081. 10.1172/jci.insight.89081
72
GousopoulosE.ProulxS. T.SchollJ.UeckerM.DetmarM. (2016b). Prominent lymphatic vessel hyperplasia with progressive dysfunction and distinct immune cell infiltration in lymphedema. Am. J. Pathol.186 (8), 2193–2203. 10.1016/j.ajpath.2016.04.006
73
GradaA. A.PhillipsT. J. (2017). Lymphedema: pathophysiology and clinical manifestations. J. Am. Acad. Dermatol77 (6), 1009–1020. 10.1016/j.jaad.2017.03.022
74
GucciardoF.LebeauA.PirsonS.BuntinxF.IvanovaE.BlacherS.et al (2025). Targeting uPARAP modifies lymphatic vessel architecture and attenuates lymphedema. Circulation151, 1412–1429. 10.1161/CIRCULATIONAHA.124.072093
75
Guyot-CaquelinP.CunyJ. F.DepardieuC.BarbaudA.SchmutzJ. L. (2010). Lymphedema and neutrophilic dermatosis. Ann. Dermatol Venereol.137 (6-7), 477–479. 10.1016/j.annder.2010.03.027
76
HadizadehM.Mohaddes ArdebiliS. M.SalehiM.YoungC.MokarianF.McClellanJ.et al (2018). GJA4/Connexin 37 mutations correlate with secondary lymphedema following surgery in breast cancer patients. Biomedicines6 (1), 23. 10.3390/biomedicines6010023
77
HägerlingR.HoppeE.DierkesC.StehlingM.MakinenT.ButzS.et al (2018). Distinct roles of VE-cadherin for development and maintenance of specific lymph vessel beds. EMBO J.37 (22), e98271. 10.15252/embj.201798271
78
Haghayegh JahromiN.GkountidiA.-O.Collado-DiazV.BlatterK.BauerA.ZambounisL.et al (2024). CD112 supports lymphatic migration of human dermal dendritic cells. Cells13 (5), 424. 10.3390/cells13050424
79
HansonH. L.WilsonM. J.ShortJ. P.ChiozaB. A.CrosbyA. H.NashR. M.et al (2014). Germline CBL mutation associated with a noonan-like syndrome with primary lymphedema and teratoma associated with acquired uniparental isodisomy of chromosome 11q23. Am. J. Med. Genet. A164A (4), 1003–1009. 10.1002/ajmg.a.36375
80
HarléG.KowalskiC.DubrotJ.BrighouseD.ClavelG.PickR.et al (2021). Macroautophagy in lymphatic endothelial cells inhibits T cell-mediated autoimmunity. J. Exp. Med.218 (6), e20201776. 10.1084/jem.20201776
81
HarveyN. L.SrinivasanR. S.DillardM. E.JohnsonN. C.WitteM. H.BoydK.et al (2005). Lymphatic vascular defects promoted by Prox1 haploinsufficiency cause adult-onset obesity. Nat. Genet.37 (10), 1072–1081. 10.1038/ng1642
82
HerradaA. A.Olate-BrionesA.Lazo-AmadorR.LiuC.Hernández-RojasB.RiadiG.et al (2022). Lymph leakage promotes immunosuppression by enhancing anti-inflammatory macrophage polarization. Front. Immunol.13, 841641. 10.3389/fimmu.2022.841641
83
HiranoT. (2021). IL-6 in inflammation, autoimmunity and cancer. Int. Immunol.33 (3), 127–148. 10.1093/intimm/dxaa078
84
IrrthumA.DevriendtK.ChitayatD.MatthijsG.GladeC.SteijlenP. M.et al (2003). Mutations in the transcription factor gene SOX18 underlie recessive and dominant forms of hypotrichosis-lymphedema-telangiectasia. Am. J. Hum. Genet.72 (6), 1470–1478. 10.1086/375614
85
IrrthumA.KarkkainenM. J.DevriendtK.AlitaloK.VikkulaM. (2000). Congenital hereditary lymphedema caused by a mutation that inactivates VEGFR3 tyrosine kinase. Am. J. Hum. Genet.67 (2), 295–301. 10.1086/303019
86
JannawayM.IyerD.MastrogiacomoD. M.LiK.SungD. C.YangY.et al (2023). VEGFR3 is required for button junction formation in lymphatic vessels. Cell Rep.42 (7), 112777. 10.1016/j.celrep.2023.112777
87
JensenC. J.BuchM. B.KragT. O.HemmingsB. A.GammeltoftS.FrödinM. (1999). 90-kDa ribosomal S6 kinase is phosphorylated and activated by 3-phosphoinositide-dependent protein kinase-1. J. Biol. Chem.274 (38), 27168–27176. 10.1074/jbc.274.38.27168
88
JiR. C. (2007). Lymphatic endothelial cells, inflammatory lymphangiogenesis, and prospective players. Curr. Med. Chem.14 (22), 2359–2368. 10.2174/092986707781745541
89
KanadyJ. D.DellingerM. T.MungerS. J.WitteM. H.SimonA. M. (2011). Connexin37 and Connexin43 deficiencies in mice disrupt lymphatic valve development and result in lymphatic disorders including lymphedema and chylothorax. Dev. Biol.354 (2), 253–266. 10.1016/j.ydbio.2011.04.004
90
KanadyJ. D.MungerS. J.WitteM. H.SimonA. M. (2015). Combining Foxc2 and Connexin37 deletions in mice leads to severe defects in lymphatic vascular growth and remodeling. Dev. Biol.405 (1), 33–46. 10.1016/j.ydbio.2015.06.004
91
KarkkainenM. J.FerrellR. E.LawrenceE. C.KimakM. A.LevinsonK. L.McTigueM. A.et al (2000). Missense mutations interfere with VEGFR-3 signalling in primary lymphoedema. Nat. Genet.25 (2), 153–159. 10.1038/75997
92
KarkkainenM. J.SaaristoA.JussilaL.KarilaK. A.LawrenceE. C.PajusolaK.et al (2001). A model for gene therapy of human hereditary lymphedema. Proc. Natl. Acad. Sci. U. S. A.98 (22), 12677–12682. 10.1073/pnas.221449198
93
KartopawiroJ.BowerN. I.KarnezisT.KazenwadelJ.BettermanK. L.LesieurE.et al (2014). Arap3 is dysregulated in a mouse model of hypotrichosis-lymphedema-telangiectasia and regulates lymphatic vascular development. Hum. Mol. Genet.23 (5), 1286–1297. 10.1093/hmg/ddt518
94
KataruR. P.BaikJ. E.ParkH. J.LyC. L.ShinJ.SchwartzN.et al (2021). Lymphatic-specific intracellular modulation of receptor tyrosine kinase signaling improves lymphatic growth and function. Sci. Signal14 (695), eabc0836. 10.1126/scisignal.abc0836
95
KazenwadelJ.BettermanK. L.ChongC.-E.StokesP. H.LeeY. K.SeckerG. A.et al (2015). GATA2 is required for lymphatic vessel valve development and maintenance. J. Clin. Invest125 (8), 2979–2994. 10.1172/JCI78888
96
KedlR. M.LindsayR. S.FinlonJ. M.LucasE. D.FriedmanR. S.TamburiniB. A. J. (2017). Migratory dendritic cells acquire and present lymphatic endothelial cell-archived antigens during lymph node contraction. Nat. Commun.8 (1), 2034. 10.1038/s41467-017-02247-z
97
KhanF.AmatyaB.PallantJ. F.RajapaksaI. (2012). Factors associated with long-term functional outcomes and psychological sequelae in women after breast cancer. Breast21 (3), 314–320. 10.1016/j.breast.2012.01.013
98
KimD.TianW.WuT. T.-H.XiangM.VinhR.ChangJ. L.et al (2023). Abnormal lymphatic Sphingosine-1-Phosphate signaling aggravates lymphatic dysfunction and tissue inflammation. Circulation148 (16), 1231–1249. 10.1161/CIRCULATIONAHA.123.064181
99
KoksharovaG.KokhN.GridinaM.KhapaevR.NimaevV.FishmanV. (2024). A HGF mutation in the familial case of primary lymphedema: a report. Int. J. Mol. Sci.25 (10), 5464. 10.3390/ijms25105464
100
KorhonenE. A.MurtomäkiA.JhaS. K.AnisimovA.PinkA.ZhangY.et al (2022). Lymphangiogenesis requires Ang2/Tie/PI3K signaling for VEGFR3 cell-surface expression. J. Clin. Invest132 (15), e155478. 10.1172/JCI155478
101
KrugmannS.AndersonK. E.RidleyS. H.RissoN.McGregorA.CoadwellJ.et al (2002). Identification of ARAP3, a novel PI3K effector regulating both arf and rho GTPases, by selective capture on phosphoinositide affinity matrices. Mol. Cell.9 (1), 95–108. 10.1016/s1097-2765(02)00434-3
102
KuonquiK.CampbellA.-C.SarkerA.RobertsA.PollackB. L.ParkH. J.et al (2023). Dysregulation of lymphatic endothelial VEGFR3 signaling in disease. Cells13 (1), 68. 10.3390/cells13010068
103
KurotakiN.ImaizumiK.HaradaN.MasunoM.KondohT.NagaiT.et al (2002). Haploinsufficiency of NSD1 causes Sotos syndrome. Nat. Genet.30 (4), 365–366. 10.1038/ng863
104
LalaP. K.NandiP.MajumderM. (2018). Roles of prostaglandins in tumor-associated lymphangiogenesis with special reference to breast cancer. Cancer Metastasis Rev.37 (2-3), 369–384. 10.1007/s10555-018-9734-0
105
LeakL. V. (1976). The structure of lymphatic capillaries in lymph formation. Fed. Proc.35 (8), 1863–1871.
106
LeeE.ChanS.-L.LeeY.PolacheckW. J.KwakS.WenA.et al (2023). A 3D biomimetic model of lymphatics reveals cell-cell junction tightening and lymphedema via a cytokine-induced ROCK2/JAM-A complex. Proc. Natl. Acad. Sci. U. S. A.120 (41), e2308941120. 10.1073/pnas.2308941120
107
LeeS.KangJ.YooJ.GanesanS. K.CookS. C.AguilarB.et al (2009). Prox1 physically and functionally interacts with COUP-TFII to specify lymphatic endothelial cell fate. Blood113 (8), 1856–1859. 10.1182/blood-2008-03-145789
108
LeppäpuskaI.-M.HartialaP.SuominenS.SuominenE.KaartinenI.MäkiM.et al (2022). Phase 1 lymfactin® study: 24-Month efficacy and safety results of combined adenoviral VEGF-C and lymph node transfer treatment for upper extremity lymphedema. J. Plast. Reconstr. Aesthet. Surg.75 (11), 3938–3945. 10.1016/j.bjps.2022.08.011
109
LevyD. E.LeeC.-k. (2002). What does Stat3 do?J. Clin. Invest109 (9), 1143–1148. 10.1172/JCI15650
110
LiN.RuanM.ChenW.HanY.YangK.XuH.et al (2024). An arabinogalactan isolated from Cynanchum atratum promotes lymphangiogenesis and lymphatic vessel remodeling to alleviate secondary lymphedema. Int. J. Biol. Macromol.273 (Pt 2), 133061. 10.1016/j.ijbiomac.2024.133061
111
LiaoS.von der WeidP. Y. (2015). Lymphatic system: an active pathway for immune protection. Semin. Cell Dev. Biol.38, 83–89. 10.1016/j.semcdb.2014.11.012
112
LoghryH. J.SondjajaN. A.MinklerS. J.KimberM. J. (2022). Secreted filarial nematode galectins modulate host immune cells. Front. Immunol.13, 952104. 10.3389/fimmu.2022.952104
113
LuJ. H.HsiaK.SuC. K.PanY. H.MaH.ChiouS. H.et al (2023). A novel dressing composed of adipose stem cells and decellularized wharton's jelly facilitated wound healing and relieved lymphedema by enhancing angiogenesis and lymphangiogenesis in a rat model. J. Funct. Biomater.14 (2), 104. 10.3390/jfb14020104
114
LynchP. M.DelanoF. A.Schmid-SchönbeinG. W. (2007). The primary valves in the initial lymphatics during inflammation. Lymphat. Res. Biol.5 (1), 3–10. 10.1089/lrb.2007.5102
115
MahamudM. R.GengX.HoY.-C.ChaB.KimY.MaJ.et al (2019). GATA2 controls lymphatic endothelial cell junctional integrity and lymphovenous valve morphogenesis through miR-126. Development146 (21), dev184218. 10.1242/dev.184218
116
MaheshwariM.BelmontJ.FernbachS.HoT.MolinariL.YakubI.et al (2002). PTPN11 mutations in Noonan syndrome type I: detection of recurrent mutations in exons 3 and 13. Hum. Mutat.20 (4), 298–304. 10.1002/humu.10129
117
MandelbrotB. B.MandelbrotB. B. (1982). The fractal geometry of nature. New York: W. H. Freeman.
118
MansourS.ConnellF.StewardC.OstergaardP.BriceG.SmithsonS.et al (2010). Emberger syndrome-primary lymphedema with myelodysplasia: report of seven new cases. Am. J. Med. Genet. A152A (9), 2287–2296. 10.1002/ajmg.a.33445
119
MartinelliS.De LucaA.StellacciE.RossiC.ChecquoloS.LepriF.et al (2010). Heterozygous germline mutations in the CBL tumor-suppressor gene cause a noonan syndrome-like phenotype. Am. J. Hum. Genet.87 (2), 250–257. 10.1016/j.ajhg.2010.06.015
120
McClellandJ.BurgessB.CrockP.GoelH. (2016). Sotos syndrome: an unusual presentation with intrauterine growth restriction, generalized lymphedema, and intention tremor. Am. J. Med. Genet. A170a (4), 1064–1069. 10.1002/ajmg.a.37535
121
MehraraB. J.ParkH. J.KataruR. P.BrombergJ.CoriddiM.BaikJ. E.et al (2021). Pilot study of Anti-Th2 immunotherapy for the treatment of breast cancer-related upper extremity lymphedema. Biology10 (9), 934. 10.3390/biology10090934
122
MicheliniS.AmatoB.RicciM.KenanogluS.VeselenyiovaD.KurtiD.et al (2020). Segregation analysis of rare NRP1 and NRP2 variants in families with lymphedema. Genes (Basel)11 (11), 1361. 10.3390/genes11111361
123
MicheliniS.RicciM.SerraniR.BaratiS.KenanogluS.VeselenyiovaD.et al (2021). NOTCH1: review of its role in lymphatic development and study of seven families with rare pathogenic variants. Mol. Genet. Genomic Med.9 (1), e1529. 10.1002/mgg3.1529
124
MiyazakiT.TaketomiY.HigashiT.OhtakiH.TakakiT.OhnishiK.et al (2023). Hypercholesterolemic dysregulation of calpain in lymphatic endothelial cells interferes with regulatory T-Cell stability and trafficking. Arterioscler. Thromb. Vasc. Biol.43 (2), e66–e82. 10.1161/ATVBAHA.122.317781
125
MorcaldiG.BelliniT.RossiC.MaghnieM.BoccardoF.BonioliE.et al (2015). Lymphodysplasia and kras mutation: a case report and literature review. Lymphology48 (3), 121–127.
126
MottaM.FidanM.BellacchioE.PantaleoniF.Schneider-HeieckK.CoppolaS.et al (2019). Dominant noonan syndrome-causing LZTR1 mutations specifically affect the kelch domain substrate-recognition surface and enhance RAS-MAPK signaling. Hum. Mol. Genet.28 (6), 1007–1022. 10.1093/hmg/ddy412
127
NarahariS. R.AggithayaM. G.RyanT. J.MuralidharanK.FranksP. J.MoffattC.et al (2023). Self-care treatment for lymphoedema of lymphatic filariasis using integrative medicine. Br. J. Dermatol190 (1), 94–104. 10.1093/bjd/ljad310
128
NarahariS. R.RyanT. J. (2020). Morbidity management and disability prevention: an agenda for developing nations initiated in India. Lymphology53 (4), 157–161. 10.2458/lymph.4668
129
NiQ.LiG.ChenY.BaoC.WangT.LiY.et al (2024). LECs regulate neutrophil clearance through IL-17RC/CMTM4/NF-κB axis at sites of inflammation or infection. Mucosal Immunol.17 (4), 723–738. 10.1016/j.mucimm.2024.05.003
130
NorrménC.IvanovK. I.ChengJ.ZanggerN.DelorenziM.JaquetM.et al (2009). FOXC2 controls formation and maturation of lymphatic collecting vessels through cooperation with NFATc1. J. Cell Biol.185 (3), 439–457. 10.1083/jcb.200901104
131
OhtaniO. (1987). Three-dimensional organization of lymphatics and its relationship to blood vessels in rat small intestine. Cell Tissue Res.248 (2), 365–374. 10.1007/BF00218204
132
OliverG.AlitaloK. (2005). The lymphatic vasculature: recent progress and paradigms. Annu. Rev. Cell Dev. Biol.21, 457–483. 10.1146/annurev.cellbio.21.012704.132338
133
OstergaardP.SimpsonM. A.BriceG.MansourS.ConnellF. C.OnoufriadisA.et al (2011). Rapid identification of mutations in GJC2 in primary lymphoedema using whole exome sequencing combined with linkage analysis with delineation of the phenotype. J. Med. Genet.48 (4), 251–255. 10.1136/jmg.2010.085563
134
OstergaardP.SimpsonM. A.MendolaA.VasudevanP.ConnellF. C.van ImpelA.et al (2012). Mutations in KIF11 cause autosomal-dominant microcephaly variably associated with congenital lymphedema and chorioretinopathy. Am. J. Hum. Genet.90 (2), 356–362. 10.1016/j.ajhg.2011.12.018
135
PalS.MeiningerC. J.GashevA. A. (2017). Aged lymphatic vessels and mast cells in perilymphatic tissues. Int. J. Mol. Sci.18 (5), 965. 10.3390/ijms18050965
136
PermanyerM.BošnjakB.FörsterR. (2018). Dendritic cells, T cells and lymphatics: dialogues in migration and beyond. Curr. Opin. Immunol.53, 173–179. 10.1016/j.coi.2018.05.004
137
PfarrK. M.DebrahA. Y.SpechtS.HoeraufA. (2009). Filariasis and lymphoedema. Parasite Immunol.31 (11), 664–672. 10.1111/j.1365-3024.2009.01133.x
138
PiaoW.LiL.SaxenaV.IyyathuraiJ.LakhanR.ZhangY.et al (2022). PD-L1 signaling selectively regulates T cell lymphatic transendothelial migration. Nat. Commun.13 (1), 2176. 10.1038/s41467-022-29930-0
139
PiaoW.XiongY.LiL.SaxenaV.SmithK. D.HippenK. L.et al (2020). Regulatory T cells condition lymphatic endothelia for enhanced transendothelial migration. Cell Rep.30 (4), 1052–1062. 10.1016/j.celrep.2019.12.083
140
PintoM.GimiglianoF.TatangeloF.MegnaM.IzzoF.GimiglianoR.et al (2013). Upper limb function and quality of life in breast cancer related lymphedema: a cross-sectional study. Eur. J. Phys. Rehabil. Med.49 (5), 665–673.
141
PujolF.HodgsonT.Martinez-CorralI.PratsA.-C.DevenportD.TakeichiM.et al (2017). Dachsous1-Fat4 signaling controls endothelial cell polarization during lymphatic valve morphogenesis-brief report. Arterioscler. Thromb. Vasc. Biol.37 (9), 1732–1735. 10.1161/ATVBAHA.117.309818
142
RobertsA. E.ArakiT.SwansonK. D.MontgomeryK. T.SchiripoT. A.JoshiV. A.et al (2007). Germline gain-of-function mutations in SOS1 cause Noonan syndrome. Nat. Genet.39 (1), 70–74. 10.1038/ng1926
143
RobertsC. M. L.AngusJ. E.LeachI. H.McDermottE. M.WalkerD. A.RavenscroftJ. C. (2010). A novel NEMO gene mutation causing osteopetrosis, lymphoedema, hypohidrotic ectodermal dysplasia and immunodeficiency (OL-HED-ID). Eur. J. Pediatr.169 (11), 1403–1407. 10.1007/s00431-010-1206-7
144
RocksonS. G.KeeleyV.KilbreathS.SzubaA.TowersA. (2019). Cancer-associated secondary lymphoedema. Nat. Rev. Dis. Prim.5 (1), 22. 10.1038/s41572-019-0072-5
145
Rodriguez-VicianaP.Oses-PrietoJ.BurlingameA.FriedM.McCormickF. (2006). A phosphatase holoenzyme comprised of Shoc2/Sur8 and the catalytic subunit of PP1 functions as an M-Ras effector to modulate raf activity. Mol. Cell22 (2), 217–230. 10.1016/j.molcel.2006.03.027
146
Rodriguez-VicianaP.WarneP. H.DhandR.VanhaesebroeckB.GoutI.FryM. J.et al (1994). Phosphatidylinositol-3-OH kinase as a direct target of ras. Nature370 (6490), 527–532. 10.1038/370527a0
147
SaijoH.SuzukiK.YoshimotoH.ImamuraY.YamashitaS.TanakaK. (2019). Paracrine effects of adipose-derived stem cells promote lymphangiogenesis in irradiated lymphatic endothelial cells. Plast. Reconstr. Surg.143 (6), 1189e–1200e. 10.1097/PRS.0000000000005669
148
SanoM.HirakawaS.SuzukiM.SakabeJ.-I.OgawaM.YamamotoS.et al (2020). Potential role of transforming growth factor-beta 1/Smad signaling in secondary lymphedema after cancer surgery. Cancer Sci.111 (7), 2620–2634. 10.1111/cas.14457
149
SantambrogioL.BerendamS. J.EngelhardV. H. (2019). The antigen processing and presentation machinery in lymphatic endothelial cells. Front. Immunol.10, 1033. 10.3389/fimmu.2019.01033
150
SatapathyA. K.SartonoE.SahooP. K.DentenerM. A.MichaelE.YazdanbakhshM.et al (2006). Human bancroftian filariasis: immunological markers of morbidity and infection. Microbes Infect.8 (9-10), 2414–2423. 10.1016/j.micinf.2006.05.003
151
SavetskyI. L.GhantaS.GardenierJ. C.TorrisiJ. S.García NoresG. D.HespeG. E.et al (2015). Th2 cytokines inhibit lymphangiogenesis. PLoS One10 (6), e0126908. 10.1371/journal.pone.0126908
152
SavetskyI. L.TorrisiJ. S.CuzzoneD. A.GhantaS.AlbanoN. J.GardenierJ. C.et al (2014). Obesity increases inflammation and impairs lymphatic function in a mouse model of lymphedema. Am. J. Physiol. Heart Circ. Physiol.307 (2), H165–H172. 10.1152/ajpheart.00244.2014
153
ScallanJ. P.JannawayM. (2022). Lymphatic vascular permeability. Cold Spring Harb. Perspect. Med.12 (8), a041274. 10.1101/cshperspect.a041274
154
SchubbertS.ShannonK.BollagG. (2007). Hyperactive ras in developmental disorders and cancer. Nat. Rev. Cancer7 (4), 295–308. 10.1038/nrc2109
155
ScorzaS. I.MilanoS.SaponaraI.CertiniM.De ZioR.MolaM. G.et al (2023). TRPML1-Induced lysosomal Ca2+ signals activate AQP2 translocation and water flux in renal collecting duct cells. Int. J. Mol. Sci.24 (2), 1647. 10.3390/ijms24021647
156
SengerJ.-L. B.KadleR. L.SkorackiR. J. (2023). Current concepts in the management of primary lymphedema. Med. Kaunas.59 (5), 894. 10.3390/medicina59050894
157
ShengH.ShaoJ.TownsendC. M.EversB. M. (2003). Phosphatidylinositol 3-kinase mediates proliferative signals in intestinal epithelial cells. Gut52 (10), 1472–1478. 10.1136/gut.52.10.1472
158
SheppardS. E.MarchM. E.SeilerC.MatsuokaL. S.KimS. E.KaoC.et al (2023). Lymphatic disorders caused by mosaic, activating KRAS variants respond to MEK inhibition. JCI Insight8 (9), e155888. 10.1172/jci.insight.155888
159
ShimizuY.CheY.MuroharaT. (2023). Therapeutic lymphangiogenesis is a promising strategy for secondary lymphedema. Int. J. Mol. Sci.24 (9), 7774. 10.3390/ijms24097774
160
ShiouS.-R.YuY.GuoY.WesterhoffM.LuL.PetrofE. O.et al (2013). Oral administration of transforming growth factor-β1 (TGF-β1) protects the immature gut from injury via smad protein-dependent suppression of epithelial nuclear factor κB (NF-κB) signaling and proinflammatory cytokine production. J. Biol. Chem.288 (48), 34757–34766. 10.1074/jbc.M113.503946
161
SiemsW. G.BrenkeR.BeierA.GruneT. (2002). Oxidative stress in chronic lymphoedema. QJM95 (12), 803–809. 10.1093/qjmed/95.12.803
162
SleutjesJ.KleimeierL.LeendersE.KleinW.DraaismaJ. (2022). Lymphatic abnormalities in Noonan syndrome spectrum disorders: a systematic review. Mol. Syndromol.13 (1), 1–11. 10.1159/000517605
163
SöhlG.WilleckeK. (2004). Gap junctions and the connexin protein family. Cardiovasc Res.62 (2), 228–232. 10.1016/j.cardiores.2003.11.013
164
StuiverM. M.ten TusscherM. R.Agasi-IdenburgC. S.LucasC.AaronsonN. K.BossuytP. M. M. (2015). Conservative interventions for preventing clinically detectable upper-limb lymphoedema in patients who are at risk of developing lymphoedema after breast cancer therapy. Cochrane Database Syst. Rev.2015 (2), CD009765. 10.1002/14651858.CD009765.pub2
165
SuzukiJ.ShimizuY.HayashiT.CheY.PuZ.TsuzukiK.et al (2022). Hydrogen sulfide attenuates lymphedema Via the induction of lymphangiogenesis through a PI3K/Akt-Dependent mechanism. J. Am. Heart Assoc.11 (21), e026889. 10.1161/JAHA.122.026889
166
SwartzM. A. (2001). The physiology of the lymphatic system. Adv. Drug Deliv. Rev.50 (1-2), 3–20. 10.1016/s0169-409x(01)00150-8
167
SzubaA.SkobeM.KarkkainenM. J.ShinW. S.BeynetD. P.RocksonN. B.et al (2002). Therapeutic lymphangiogenesis with human recombinant VEGF-C. FASEB J.16 (14), 1985–1987. 10.1096/fj.02-0401fje
168
TabibiazarR.CheungL.HanJ.SwansonJ.BeilhackA.AnA.et al (2006). Inflammatory manifestations of experimental lymphatic insufficiency. PLoS Med.3 (7), e254. 10.1371/journal.pmed.0030254
169
TakedaK.SowaY.NishinoK.ItohK.FushikiS. (2015). Adipose-derived stem cells promote proliferation, migration, and tube formation of lymphatic endothelial cells in vitro by secreting lymphangiogenic factors. Ann. plastic Surg.74 (6), 728–736. 10.1097/SAP.0000000000000084
170
TammelaT.SaaristoA.HolopainenT.LyytikkäJ.KotronenA.PitkonenM.et al (2007). Therapeutic differentiation and maturation of lymphatic vessels after lymph node dissection and transplantation. Nat. Med.13 (12), 1458–1466. 10.1038/nm1689
171
TartagliaM.KalidasK.ShawA.SongX.MusatD. L.van der BurgtI.et al (2002). PTPN11 mutations in Noonan syndrome: molecular spectrum, genotype-phenotype correlation, and phenotypic heterogeneity. Am. J. Hum. Genet.70 (6), 1555–1563. 10.1086/340847
172
TartagliaM.MehlerE. L.GoldbergR.ZampinoG.BrunnerH. G.KremerH.et al (2001). Mutations in PTPN11, encoding the protein tyrosine phosphatase SHP-2, cause Noonan syndrome. Nat. Genet.29 (4), 465–468. 10.1038/ng772
173
TashiroK.FengJ.WuS. H.MashikoT.KanayamaK.NarushimaM.et al (2017). Pathological changes of adipose tissue in secondary lymphoedema. Br. J. Dermatol177 (1), 158–167. 10.1111/bjd.15238
174
TianW.RocksonS. G.JiangX.KimJ.BegayeA.ShuffleE. M.et al (2017). Leukotriene B4 antagonism ameliorates experimental lymphedema. Sci. Transl. Med.9 (389), eaal3920. 10.1126/scitranslmed.aal3920
175
TidymanW. E.RauenK. A. (2008). Noonan, costello and cardio-facio-cutaneous syndromes: dysregulation of the Ras-MAPK pathway. Expert Rev. Mol. Med.10, e37. 10.1017/S1462399408000902
176
TidymanW. E.RauenK. A. (2009). The RASopathies: developmental syndromes of Ras/MAPK pathway dysregulation. Curr. Opin. Genet. Dev.19 (3), 230–236. 10.1016/j.gde.2009.04.001
177
UekiT.IwasawaK.IshikawaH.SawaY. (2008). Expression of junctional adhesion molecules on the human lymphatic endothelium. Microvasc. Res.75 (2), 269–278. 10.1016/j.mvr.2007.07.005
178
VisserR.LandmanE. B. M.GoemanJ.WitJ. M.KarperienM. (2012). Sotos syndrome is associated with deregulation of the MAPK/ERK-signaling pathway. PLoS One7 (11), e49229. 10.1371/journal.pone.0049229
179
von der WeidP. Y. (2013). Lymphatic myogenic constriction - how lymphatic vessels pump lymph uphill. J. Physiol.591 (2), 391–392. 10.1113/jphysiol.2012.247080
180
WalshV.LittleK.WiegandR.RoutJ.FoxL. M. (2016). Evaluating the burden of lymphedema due to lymphatic filariasis in 2005 in khurda district, Odisha state, India. PLoS Negl. Trop. Dis.10 (8), e0004917. 10.1371/journal.pntd.0004917
181
WeinkopffT.MackenzieC.EversoleR.LammieP. J. (2014). Filarial excretory-secretory products induce human monocytes to produce lymphangiogenic mediators. PLoS Negl. Trop. Dis.8 (7), e2893. 10.1371/journal.pntd.0002893
182
WigleJ. T.OliverG. (1999). Prox1 function is required for the development of the murine lymphatic system. Cell98 (6), 769–778. 10.1016/s0092-8674(00)81511-1
183
WillP.DraguA.ZutherJ.HeilJ.ChangD.-H.TraberJ.et al (2024b). Evidence of modern diagnostic, conservative, and surgical therapy of secondary lymphoedema. Handchir Mikr56 (4), 291–300. 10.1055/a-2322-1325
184
WillP. A.KilianK.BiebackK.FrickeF.BernerJ. E.KneserU.et al (2024a). Lymphedema-associated fibroblasts are related to fibrosis and stage progression in patients and a murine microsurgical model. Plast. Reconstr. Surg.154 (4), 688e–700e. 10.1097/PRS.0000000000011141
185
YamamotoT.YoshimatsuH.NarushimaM.YamamotoN.HayashiA.KoshimaI. (2015). Indocyanine green lymphography findings in primary leg lymphedema. Eur. J. Vasc. Endovasc. Surg.49 (1), 95–102. 10.1016/j.ejvs.2014.10.023
186
YanA.AvrahamT.ZampellJ. C.HavivY. S.WeitmanE.MehraraB. J. (2011). Adipose-derived stem cells promote lymphangiogenesis in response to VEGF-C stimulation or TGF-β1 inhibition. Future Oncol.7 (12), 1457–1473. 10.2217/fon.11.121
187
YanJ.XiaoG.YangC.LiuQ.LvC.YuX.et al (2024). Cancer-associated fibroblasts promote lymphatic metastasis in cholangiocarcinoma via the PDGF-BB/PDGFR-β mediated paracrine signaling network. Aging Dis.15 (1), 369–389. 10.14336/AD.2023.0420
188
YangC.YouL.MengH. (2021a). Massive limb lymphedema as the initial presentation of multiple myeloma: a case report. Transl. Cancer Res.10 (3), 1599–1602. 10.21037/tcr-20-3143
189
YangJ. C.-S.HuangL.-H.WuS.-C.KuoP.-J.WuY.-C.WuC.-J.et al (2021b). Lymphaticovenous anastomosis supermicrosurgery decreases oxidative stress and increases antioxidant capacity in the serum of lymphedema patients. J. Clin. Med.10 (7), 1540. 10.3390/jcm10071540
190
YangL.WangG.MaY.ZhaoQ.ZhaoH.WangQ.et al (2024). TRPML1 acts as a predisposing factor in lymphedema development by regulating the subcellular localization of aquaporin-3, -5. PLoS One19 (12), e0310653. 10.1371/journal.pone.0310653
191
YangY.ChaB.MotaweZ. Y.SrinivasanR. S.ScallanJ. P. (2019). VE-Cadherin is required for lymphatic valve formation and maintenance. Cell Rep.28 (9), 2397–2412. 10.1016/j.celrep.2019.07.072
192
YoshimuraA.MutoG. (2011). TGF-β function in immune suppression. Curr. Top. Microbiol. Immunol.350, 127–147. 10.1007/82_2010_87
193
YoungL. C.HartigN.Boned Del RíoI.SariS.Ringham-TerryB.WainwrightJ. R.et al (2018). SHOC2-MRAS-PP1 complex positively regulates RAF activity and contributes to Noonan syndrome pathogenesis. Proc. Natl. Acad. Sci. U. S. A.115 (45), E10576–E10585. 10.1073/pnas.1720352115
194
ZampellJ. C.YanA.ElhadadS.AvrahamT.WeitmanE.MehraraB. J. (2012). CD4(+) cells regulate fibrosis and lymphangiogenesis in response to lymphatic fluid stasis. PLoS One7 (11), e49940. 10.1371/journal.pone.0049940
195
ZawiejaD. C. (2009). Contractile physiology of lymphatics. Lymphat. Res. Biol.7 (2), 87–96. 10.1089/lrb.2009.0007
196
ZhaoS.KonoplevaM.Cabreira-HansenM.XieZ.HuW.MilellaM.et al (2004). Inhibition of phosphatidylinositol 3-kinase dephosphorylates BAD and promotes apoptosis in myeloid leukemias. Leukemia18 (2), 267–275. 10.1038/sj.leu.2403220
197
ZhouZ.SuiX.CaoZ.LiX.QingL.TangJ. (2023). Substance P promote macrophage M2 polarization to attenuate secondary lymphedema by regulating NF-kB/NLRP3 signaling pathway. Peptides168, 171045. 10.1016/j.peptides.2023.171045
198
ZimmermannS.MoellingK. (1999). Phosphorylation and regulation of raf by akt (protein kinase B). Science.286 (5445), 1741–1744. 10.1126/science.286.5445.1741
Summary
Keywords
lymphedema, signaling pathways, inflammation, cell communication, gap junction
Citation
Zhao Q, Niu Z, Pan Y, Hao Y, Ma Y, Zhao J, Du J and Yang Y (2025) Characteristics and advances in signaling pathways, cellular communication, cell junctions, and oxidative stress in lymphedema. Front. Cell Dev. Biol. 13:1521320. doi: 10.3389/fcell.2025.1521320
Received
04 November 2024
Accepted
25 June 2025
Published
22 July 2025
Volume
13 - 2025
Edited by
Sathish Srinivasan, Oklahoma Medical Research Foundation, United States
Reviewed by
Kui Cui, Harvard University, United States
S. R Narahari, Institute of Applied Dermatology, India
Updates
Copyright
© 2025 Zhao, Niu, Pan, Hao, Ma, Zhao, Du and Yang.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yiming Yang, ym_yang@jlu.edu.cn
†These authors have contributed equally to this work
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.