 Alejandra Krüger1*
Alejandra Krüger1* Paula M. A. Lucchesi1
Paula M. A. Lucchesi1 A. Mariel Sanso1
A. Mariel Sanso1 Analía I. Etcheverría1
Analía I. Etcheverría1 Ana V. Bustamante1
Ana V. Bustamante1 Julia Burgán1
Julia Burgán1 Luciana Fernández1
Luciana Fernández1 Daniel Fernández1
Daniel Fernández1 Gerardo Leotta2
Gerardo Leotta2 Alexander W. Friedrich3
Alexander W. Friedrich3 Nora L. Padola1*
Nora L. Padola1* John W. A. Rossen3
John W. A. Rossen3- 1Laboratorio de Inmunoquímica y Biotecnología, Facultad de Ciencias Veterinarias, Centro de Investigación Veterinaria de Tandil, Consejo Nacional de Investigaciones Científicas y Técnicas, Comisión de Investigaciones Científicas, Universidad Nacional del Centro de la Provincia de Buenos Aires, Tandil, Argentina
- 2Línea Seguridad Alimentaria, Instituto de Genética Veterinaria Ing. F.N. Dulout, Consejo Nacional de Investigaciones Científicas y Técnicas, La Plata, Argentina
- 3Department of Medical Microbiology, University Medical Center Groningen, University of Groningen, Groningen, Netherlands
The Shiga-toxin producing Escherichia coli (STEC) may cause serious illness in human. Here we analyze O26:H11 strains known to be among the most reported STEC strains causing human infections. Genetic characterization of strains isolated from animal, food, and clinical specimens in Argentina showed that most carried either stx1a or stx2a subtypes. Interestingly, stx2a-positive O26:H11 rarely isolated from cattle in other countries showed to be an important proportion of O26:H11 strains circulating in cattle and food in our region. Seventeen percent of the isolates harbored more than one gene associated with antimicrobial resistance. In addition to stx, all strains contained the virulence genes eae-β, tir, efa, iha, espB, cif, espA, espF, espJ, nleA, nleB, nleC, and iss; and all except one contained ehxA, espP, and cba genes. On the other hand, toxB and espI genes were exclusively observed in stx2-positive isolates, whereas katP was only found in stx1a-positive isolates. Our results show that O26:H11 STEC strains circulating in Argentina, including those isolated from humans, cattle, and meat products, present a high pathogenic potential, and evidence that cattle can be a reservoir of O26:H11 strains harboring stx2a.
Introduction
Shiga toxin-producing Escherichia coli (STEC) are E. coli strains that can cause human diseases, like gastrointestinal illnesses and hemolytic uremic syndrome (HUS). E. coli O157 has been the most commonly reported STEC serogroup since it was identified in the 1980s. However, the number of non-O157 STEC infections has increased substantially in the last years. The O26 is one of the few serogroups frequently reported in non-O157 cases (Rivas et al., 2010; EFSA, 2013; Gould et al., 2013; ISPCH, 2014). In particular, strains of serotype O26:H11/- have been associated with severe human diseases (Gerber et al., 2002; Zimmerhackl et al., 2010; Käppeli et al., 2011).
All STEC strains are characterized by the ability to produce Shiga toxins (Stx). The Stx family consists of two major types: Stx1 and Stx2, which can be further divided into several subtypes. A single STEC strain may carry one or more Shiga toxin-encoding genes (stx) which are generally carried by prophages. Epidemiological studies indicate that different subtypes of stx are related to different clinical manifestations after STEC infection (Krüger and Lucchesi, 2015). Particularly, the stx2a subtype is associated with highly virulent strains and HUS. Characterization of stx genotypes showed that STEC O26 strains isolated from patients can harbor stx1a, stx2a or both, however, strains harboring only stx2a were significantly associated with HUS (Bielaszewska et al., 2013). Furthermore, Bielaszewska et al. (2013) identified a new highly virulent stx2a-positive O26 clone as an emerging cause of HUS in Europe.
The production of Stx seems to be essential but not solely responsible for STEC pathogenicity. Other known and putative virulence factors are usually present in pathogenic STEC strains. Some of them, like the adhesin intimin encoded on a bacterial chromosomal pathogenicity island and the enterohemolysin EhxA encoded on a plasmid have been found in association with severe clinical disease in humans (Boerlin et al., 1999; Aldick et al., 2007).
It is a well-known fact that ruminants are the main reservoir of STEC strains (Naylor et al., 2005). Some studies suggest that domestic animals serve as reservoir for human pathogenic O26 STEC strains (Leomil et al., 2005). The O26 STEC strains also seem to be widely distributed in cattle, since those strains have been isolated from bovines belonging to different animal categories and production systems (Monaghan et al., 2011; Fernández et al., 2012; Paddock et al., 2014; Bonardi et al., 2015; Ison et al., 2015). Moreover, O26:H11 strains have been isolated from beef and dairy products (Bosilevac and Koohmaraie, 2011; Madic et al., 2011; Mohammed et al., 2014). Several studies from Europe and the United States report that O26 strains isolated from food and cattle generally carry stx1 or both stx1 and stx2 genes; moreover, O26 strains harboring only stx2a have been rarely isolated from cattle and food (Pearce et al., 2006; Geue et al., 2009; Bonanno et al., 2015; Ison et al., 2015).
Our aim was to characterize O26:H11 STEC strains isolated from cattle, food and humans to contribute to the global knowledge of virulence profiles and epidemiology of O26 strains circulating in Argentina.
Material and Methods
Bacterial Strains and Growth Conditions
E. coli O26:H11 strains were selected from STEC collections in Argentina. The strains had been isolated between 1995 and 2013, from cattle, meat, human, and farm environment (Table 1). Most of the strains had been previously characterized by PCR regarding the presence of stx1, stx2, eae, ehxA, and saa genes. Strains were stored at −70°C with 20% (v/v) glycerol and when necessary grown in Luria Bertani broth at 37°C overnight.
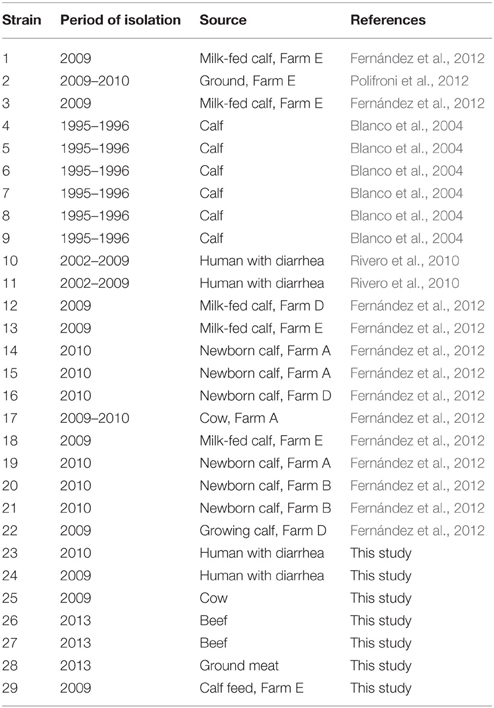
Table 1. Characteristics of O26:H11 STEC isolates tested in this study.
Microarray-based Characterization of Virulence Factors
Bacterial genomic DNA was extracted with the UltraClean Microbial DNA Isolation Kit (Mo Bio) or Wizard Genomic DNA Purification Kit (Promega) according to the manufacturer's instructions. Virulence and antimicrobial resistance genes were evaluated with a commercial oligonucleotide microarray for E. coli according to the manufacturer's protocol (CLONDIAG Escherichia coli combined Assay, Alere Technologies GmbH; Geue et al., 2010). The array contained 87 probes targeting virulence genes and 102 probes targeting antimicrobial resistance associated genes. Visualization of hybridization was achieved using the ArrayMate instrument (CLONDIAG GmbH) and signals were analyzed automatically. The results were converted into a binary numerical format (1– present, 0– absent) and further analyzed using BioNumerics (Version 6.6; Applied Maths).
stx Subtyping
Specific PCR reactions were performed to identify stx1a, stx1b, and stx1c subtypes (Scheutz et al., 2012).
The presence of stx2b, stx2e, stx2f, and stx2g subtypes was evaluated with the oligonucleotide microarray. This assay also detects stx2a, stx2c, and stx2d subtypes but does not discriminate among them. Therefore, strains positive with the probe that detects stx2a, c, d subtypes where further subtyped with specific PCR reactions (Scheutz et al., 2012).
Detection of eae-β Gene
Strains were tested for the presence of the eae-β subtype by specific PCR using the primer set SK1/LP4 (Oswald et al., 2000).
Antibiotic Susceptibility Testing
Isolates carrying resistance genes were tested for resistance to several antimicrobials by the disk diffusion method according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2014). The following antimicrobial disks were used: ampicillin (10 μg), cephalothin (30 μg), cefotaxime (30 μg), cefoxitin (30 μg), amoxicillin/clavulanate (20 μg-10 μg), gentamicin (10 μg), amikacin (30 μg), streptomycin (300 μg), nalidixic acid (30 μg), ciprofloxacin (5 μg), chloramphenicol (30 μg), trimethoprim-sulfamethoxazole (1.25–23.75 μg), nitrofurantoin (300 μg), tetracycline (30 μg), colistin (10 μg), florfenicol (30 μg).
Multiple-locus Variable Number Tandem Repeat Analysis (MLVA)
For typing, an MLVA protocol that comprises nine variable number of tandem repeats (VNTR) loci, seven loci proposed by Lindstedt et al. (2007), i.e., CVN001, 002, 003, 004, 007, 014, 015, and two additional loci proposed by Løbersli et al. (2012), i.e., CVN016 and 017 was used. The seven loci proposed by Lindstedt et al. (2007) were amplified as described by Bustamante et al. (2010) and the two additional ones according to González et al. (2014). PCR products were analyzed in 6% denaturing polyacrylamide gels, and detected by silver staining (GenePrint STR Systems—Silver Stain Detection, Promega).
Allelic variants identified for each VNTR were sequenced with the same primers used to amplify those regions (Macrogen, Inc.). The sequences obtained were analyzed using Chromas 2.32 software (Technelysium Pty. Ltd.) and allele sequences of each VNTR were aligned with the software Clustal W (Larkin et al., 2007) in order to identify the number of tandem repeat units (TR). Alleles were named according to the number of TR. The absence of an amplification product was considered a null allele (−2).
The diversity index (DN), based on Nei's marker diversity, was calculated for each locus using the formula DN = 1–Σ(fra)2, where fra is the allelic frequency (Noller et al., 2003). The discriminatory power of the method was assessed using the Simpson diversity index (DS) (Hunter and Gaston, 1988).
Results
Genetic Characterization of Virulence Factors
Figure 1 shows the genes encoding virulence factors detected in the O26:H11 strains, clustered according to the Bionumerics analysis. Genes that were not found in any isolate are summarized in a footnote in the figure.

Figure 1. Cluster analysis of genes associated with virulence. B, E, H, and M indicate isolates obtained from bovine, environmental, human, and meat sources, respectively. Results for the lpf gene are not included because of the ambiguous signal observed for all strains. Genes not found in the any of the studied strains: bfpA, ccl, cdtB, cfaC, cnf 1, cof A, eaaA, eatA, epeA, espA_C_rodentium, espB_O157, espC, espF, espF_C_rodentium, etpD, fanA, fasA, fedA, fedF, f17-A, f17-G, fim41a, hlyE, ipaD, ipaH, ireA, iroN, K88ab, lngA ltcA, nfaE, nleB O157:H7, nleB Salmonella, perA, pet, pic, prf B, rpeA, saa, sat, senB, sepA, sfaS, sigA, sta1, sta2, stb, stx2b, stx2c, stx2d, stx2e, stx2f, stx2g, subA, tsh, vat, virF.
The stx types identified with the array were in agreement with previous PCR results. Forty five percent of the isolates were stx1-positve, 52% stx2-positive, and one isolate (3%) was stx1 and stx2-positive. By using the PCR subtyping protocol, all stx1 genes corresponded to the stx1a subtype and all stx2 corresponded to the stx2a subtype.
All tested isolates harbored the genes encoding for the adhesin Intimin (eae subtype β), and its receptor Tir (tir). Other adhesion related genes were also identified. The presence of efa1, espB, and iha genes was demonstrated in all isolates, whereas espP was found in all except the stx1 and stx2-positive isolate. The toxB gene was only observed in stx2-positive isolates. The fasA, fedA, fedF, fim41a, nfaE, and saa genes were not found.
Among toxin-encoding genes, ehxA (encoding for a hemolysin) was present in 97% of the isolates and astA (encoding for EAST1, the enteroaggregative Escherichia coli heat-stable enterotoxin 1) was detected in 72% of the isolates. The cba and celB genes associated with colicin activity were identified in 97 and 17% of the isolates, respectively. Also, one bovine isolate was positive for the gene encoding colicin M (cma) and one isolate obtained from human with diarrhea was positive for mchB, mchC, mchF, and mcmA microcin genes.
Several type III secretion system components (translocators and effectors) were identified. All the strains were positive for cif, espA, espfO103, espJ, nleA, nleB, and nleC genes, and 76% of the isolates harbored tccP. The espI gene was only identified in stx2-positive strains. The etpD, a gene encoding for a type II secretion pathway-related protein, was not found.
The iss and hemL genes were identified in 100 and 97% of the isolates, respectively. The katP gene, encoding a catalase peroxidase, was detected in stx1-positive isolates only.
Antimicrobial Resistance
Five O26:H11 isolates (17%) carried more than two antimicrobial resistance genes, being blaTEM, strA-strB, and sul2 the most frequently detected (Table 2). Genes that were not found in any of the isolates are summarized in a footnote in the table. The integron integrase encoding gene (intI1) was detected in isolate 21. The antimicrobial susceptibilities of the five isolates carrying resistance genes were determined using disk diffusion method. Four isolates were resistant to multiple antimicrobial agents (Table 2), and most genetic resistances were phenotypically confirmed. The exception was isolate 23 (human isolate) that carried strA, strB, and sul2 genes but was susceptible for the tested antibiotics.

Table 2. Antibiotic resistance profiles present in O26:H11 STEC isolates.
MLVA Typing
The isolates could be principally discriminated by alleles of loci CVN014 (DN = 0.83) and CVN016 (DN = 0.70), followed by those of CVN017 (DN = 0.63). On the other hand, loci CVN002 and CVN003 showed a null allele in all the isolates. MLVA typing divided the total number of strains into 19 distinct genotypes, 13 of them being unique (Figure 2). The Simpson's index of diversity, calculated for the combined typing set, showed a value of DS = 0.96. Three main clusters were obtained: one group included the stx2- positive isolates and one stx1-positive isolate (isolate 17); a second group, most of the stx1-positive strains and the stx1 and stx2-positive isolate (isolate 23) and a third group included three stx1-positive strains.
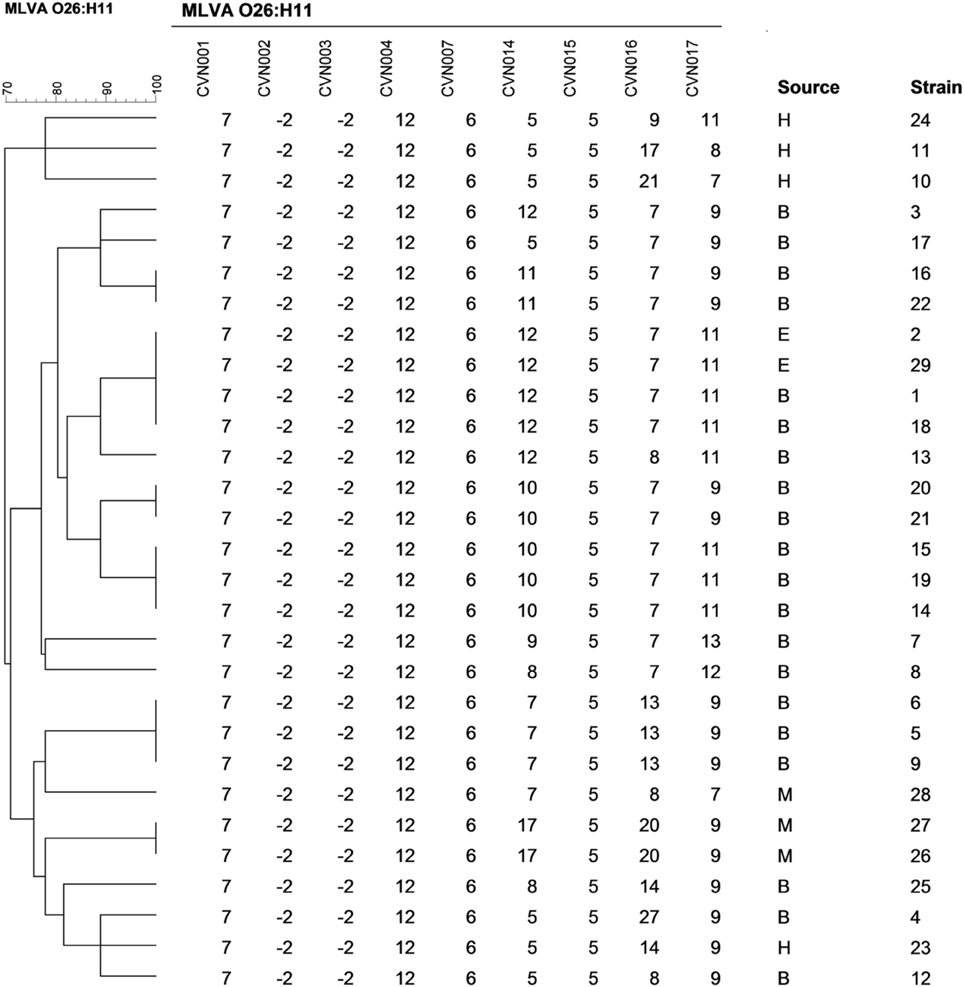
Figure 2. Multiple-locus variable-number tandem repeat analysis–based clustering of STEC O26:H11 isolates investigated in this study. Similarities among MLVA profiles were calculated using categorical coefficients and UPGMA clustering method. B, E, H, and M indicate isolates obtained from bovine, environmental, human, and meat sources, respectively.
Isolates with the same MLVA type may indicate they are clonal. For some isolates this was supported by revealing a time-spatial relationship and by their identical virulence profile (isolates 14 and 19 from dairy farm A; 26 and 27 from a same meat processing plant; 20 and 21 from dairy farm B; 16 and 22 from dairy farm D; 1 and 18 from dairy farm E; and 2 and 29 also from dairy farm E).
Discussion
STEC O26:H11 strains are considered among the most reported STEC strains involved in human infections worldwide. To our knowledge, there is no specific study of O26:H11 STEC strains in Latin America. Our aim was to determine the characteristics of E. coli O26:H11 strains circulating in Argentina in the period 1995–2013 to contribute to the global characterization of these strains.
Subtyping of stx genes indicated that O26:H11 strains in our region mainly present either stx1a or stx2a genotypes, similar to O26:H11 strains reported in other countries (Bielaszewska et al., 2013; Bletz et al., 2013; Bonanno et al., 2015). The stx2d subtype, present in the new pathogenic O26:H11 clone circulating in France (Delannoy et al., 2015), was not detected in our collection.
Genetic characterization showed that in addition to stx genes, O26:H11 strains harbored genes encoding other toxins, adhesins, and components related to the type III secretion system that contribute to their virulence. In particular, eae-β, tir, efa, iha, espB, cif, espA, espF, espJ, nleA, nleB, nleC, and iss genes were detected in all the isolates; and all except one contained ehxA, espP, and cba genes.
The analysis based on the presence/absence of genes associated with virulence identified three main clusters, one containing the stx1a-positive isolates, another the stx2a-positive isolates and a third one the stx1a and stx2a-positive isolate (Figure 1). The toxB, espI, and katP genes were differentially distributed between stx1a-positive and stx2a-positive groups. The toxB and espI genes, previously associated with severe disease (Mundy et al., 2004; Michelacci et al., 2014), were exclusively present in stx2a-positive isolates. Conversely, the katP gene encoding for catalase-peroxidase was only detected in the stx1a-positive isolates.
Heterogeneity in gene composition of STEC O26:H11 plasmids has been detected by Zhang et al. (2000). Considering the plasmidic markers ehxA, katP, espP, and etpD, two major subgroups of stx2a-positive strains were identified in Europe by Bielaszewska et al. (2013), one of them typical for the new virulent German clone. Noticeably, all the stx2a-positive strains of our study present a distinct profile, positive for ehxA and espP, but negative for katP and etpD genes. Thus, stx2a-positive strains of our Argentinian collection differ from the German clone, and also from the human-pathogenic strain recently identified in France (Delannoy et al., 2015), which is negative for all these plasmidic markers. The plasmid profile of stx1a-positive strains of our study (positive for ehxA, katP, espP, and negative for etpD) is the same that Zweifel et al. (2013) identified in the human stx1-positive strains from patients with HUS or bloody diarrhea in Switzerland. On the other hand, the unique stx1 and stx2-positive isolate was negative for all plasmidic genes (toxB, espP, katP, ehxA, etpD, saa, subAB, epeA) tested, suggesting the absence of a virulence plasmid like pO26-Vir, pO157, or pO113.
The MLVA profiles found in the present study do not coincide with any of the profiles identified by Brandal et al. (2012) among ovine and human O26:H11 isolates from Norway. Although loci CVN001, 004, and 007 were monomorphic in both studies, the alleles were different. The loci CVN002 and CVN003 presented a null allele for all tested samples in our study, in agreement with results obtained by Løbersli et al. (2012) for O26 isolates. Available epidemiological information suggests that isolates forming clusters with identical MLVA profiles were derived from the same source and constitute a single clone. However, one cluster contained human stx1-positive isolates not epidemiologically related. In addition, two isolates (isolates 20 and 21) from the same farm had the same virulence and MLVA profiles but only isolate 21 had antimicrobial resistance genes. Although there is a possibility that these isolates are from the same clone and that isolate 21 acquired antimicrobial genes, it is also possible that MLVA typing is not discriminatory enough to distinguish both isolates from each other. Interestingly, major groups obtained by MLVA analysis were similar to those obtained by the analysis of virulence factors.
Antimicrobial resistance genes were detected in five isolates obtained from two calves, two meat samples and one patient with diarrhea. Except for the human isolate, all other isolates showed phenotypic resistance profiles predicted by the corresponding genotypic profile. Among the antibiotics tested, all four isolates were resistant to ampicillin, amoxicillin/clavulanate, and tetracycline and also showed intermediate resistance to cephalotin. The two meat isolates (isolates 26 and 27) were also resistant to streptomycin, nalidixic acid, and trimethropim-sulfamethoxazole. As we commented below, these two isolates also shared the same virulence and MLVA profiles. Considering the epidemiological link, as they were found in meat samples from the same meat processing plant, our results suggest that both isolates correspond to the same circulating strain.
Previous reports showed that integrons can be frequently detected in STEC strains and that most of the integrons can contain the aadA1 gene alone, or in association with the drf A1 gene (Morabito et al., 2002; Cergole-Novella et al., 2011). In our study, the intI1 gene was detected in isolate 21 which was also positive for aadA1 and drf A1 genes. Strikingly, this isolate carrying an integron and showing multiple resistances to antimicrobials was obtained from a newborn calf. Our results highlight the presence of multi-antimicrobial resistant STEC in cattle and meat in agreement with previous studies reporting the emergence and dissemination of antimicrobial resistance among STEC strains (Zhao et al., 2001; Li et al., 2011; Sasaki et al., 2012). Although antibiotic therapy is discouraged for treatment of STEC infections, the presence of antimicrobial resistant STEC strains in animals represent a risk for animal and human health. The genes coding for antimicrobial resistance could be transferred to other pathogens. Moreover, antimicrobial resistant STEC strains may have a selective advantage over other bacteria in intestines of animals under antibiotic treatments (Zhao et al., 2001). Taking into account that the same classes of antimicrobial agents are used both in humans and animals, joint efforts should be made to reduce the inappropriate use of antimicrobial agents in animals (Aidara-Kane, 2014).
In conclusion, we identified three different populations of native O26:H11 strains whose main differences were associated with genes present in mobile genetic elements. Although O26 strains harboring only stx2a subtype have been rarely isolated from cattle and food in Europe and the United States (Pearce et al., 2006; Geue et al., 2009; Chase-Topping et al., 2012; Ison et al., 2015), stx2a-positive strains have been an important proportion of O26:H11 strains circulating in farms in Argentina and showed to carry genes associated with high virulence, representing a potential risk for public health.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was partly supported by the Interreg IVa-funded projects EurSafety Health-net (III-1-02=73) and SafeGuard (III-2-03=025), and by grants from University Medical Center Groningen Healthy Ageing Pilots, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), and Fondo para la Investigación Científica y Tecnológica (FONCYT).
References
Aidara-Kane, A. (2014). Containment of antimicrobial resistance due to use of antimicrobial agents in animals intended for food: WHO perspective. Rev. Sci. Tech. 31, 277–287.
Aldick, T., Bielaszewska, M., Zhang, W., Brockmeyer, J., Schmidt, H., Friedrich, A. W., et al. (2007). Hemolysin from shiga toxin-negative Escherichia coli O26 strains injures microvascular endothelium. Microbes Infect. 9, 282–290. doi: 10.1016/j.micinf.2006.12.001
Bielaszewska, M., Mellmann, A., Bletz, S., Zhang, W., Kock, R., Kossow, A., et al. (2013). Enterohemorrhagic Escherichia coli O26:H11/H-: a new virulent clone emerges in Europe. Clin. Infect. Dis. 56, 1373–1381. doi: 10.1093/cid/cit055
Blanco, M., Padola, N. L., Krüger, A., Sanz, M. E., Blanco, J. E., González, E. A., et al. (2004). Virulence genes and intimin types of shiga-toxin-producing Escherichia coli isolated from cattle and beef products in Argentina. Int. Microbiol. 7, 269–276.
Bletz, S., Bielaszewska, M., Leopold, S. R., Köck, R., Witten, A., Schuldes, J., et al. (2013). Evolution of enterohemorrhagic Escherichia coli O26 based on single-nucleotide polymorphisms. Genome Biol. Evol. 5, 1807–1816. doi: 10.1093/gbe/evt136
Boerlin, P., McEwen, S. A., Boerlin-Petzold, F., Wilson, J. B., Johnson, R. P., and Gyles, C. L. (1999). Associations between virulence factors of Shiga toxin-producing Escherichia coli and disease in humans. J. Clin. Microbiol. 37, 497–503.
Bonanno, L., Loukiadis, E., Mariani-Kurkdjian, P., Oswald, E., Garnier, L., Michel, V., et al. (2015). Diversity of Shiga toxin-producing Escherichia coli (STEC) O26:H11 strains examined via stx subtypes and insertion sites of Stx and EspK Bacteriophages. Appl. Environ. Microbiol. 81, 3712–3721. doi: 10.1128/AEM.00077-15
Bonardi, S., Alpigiani, I., Tozzoli, R., Vismarra, A., Zecca, V., Greppi, C., et al. (2015). Shiga toxin-producing Escherichia coli O157, O26 and O111 in cattle faeces and hides in Italy. Vet. Rec. Open 2:e000061. doi: 10.1136/vetreco-2014-000061
Bosilevac, J. M., and Koohmaraie, M. (2011). Prevalence and characterization of non-O157 shiga toxin-producing Escherichia coli isolates from commercial ground beef in the United States. Appl. Environ. Microbiol. 77, 2103–2112. doi: 10.1128/AEM.02833-10
Brandal, L. T., Sekse, C., Lindstedt, B. A., Sunde, M., Løbersli, I., Urdahl, A. M., et al. (2012). Norwegian sheep are an important reservoir for human-pathogenic Escherichia coli O26:H11. Appl. Environ. Microbiol. 78, 4083–4091. doi: 10.1128/AEM.00186-12
Bustamante, A. V., Sanso, A. M., Lucchesi, P. M., and Parma, A. E. (2010). Genetic diversity of O157:H7 and non-O157 verocytotoxigenic Escherichia coli from Argentina inferred from multiple-locus variable-number tandem repeat analysis (MLVA). Int. J. Med. Microbiol. 300, 212–217. doi: 10.1016/j.ijmm.2009.08.020
Cergole-Novella, M. C., Pignatari, A. C. C., Castanheira, M., and Guth, B. E. C. (2011). Molecular typing of antimicrobial-resistant shiga-toxin-producing Escherichia coli strains (STEC) in Brazil. Res. Microbiol. 162, 117–123. doi: 10.1016/j.resmic.2010.09.022
Chase-Topping, M. E., Rosser, T., Allison, L. J., Courcier, E., Evans, J., Mckendrick, I. J., et al. (2012). Pathogenic potential to humans of bovine Escherichia coli O26, Scotland. Emerg. Infect. Dis. 18, 439–448. doi: 10.3201/eid1803.111236
CLSI (2014). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fourth Informational Supplement. CLSI Document M100–S24.
Delannoy, S., Mariani-Kurkdjian, P., Bonacorsi, S., Liguori, S., and Fach, P. (2015). Characteristics of emerging human-pathogenic Escherichia coli O26:H11 strains isolated in France between 2010 and 2013 and carrying the stx2d gene only. J. Clin. Microbiol. 53, 486–492. doi: 10.1128/JCM.02290-14
EFSA. Panel on Biological Hazards (2013). Scientific opinion on VTEC-seropathotype and scientific criteria regarding pathogenicity assessment. EFSA J. 11:3138. doi: 10.2903/j.efsa.2013.3138
Fernández, D., Sanz, M. E., Parma, A. E., and Padola, N. L. (2012). Short communication: characterization of shiga toxin-producing Escherichia coli isolated from newborn, milk-fed, and growing calves in Argentina. J. Dairy Sci. 95, 5340–5343. doi: 10.3168/jds.2011-5140
Gerber, A., Karch, H., Allerberger, F., Verweyen, H. M., and Zimmerhackl, L. B. (2002). Clinical course and the role of shiga toxin–producing Escherichia coli infection in the hemolytic-uremic syndrome in pediatric patients, 1997–2000, in Germany and Austria: a prospective study. J. Infect. Dis. 186, 493–500. doi: 10.1086/341940
Geue, L., Klare, S., Schnick, C., Mintel, B., Meyer, K., and Conraths, F. J. (2009). Analysis of the clonal relationship of serotype O26:H11 enterohemorrhagic Escherichia coli isolates from cattle. Appl. Environ. Microbiol. 75, 6947–6953. doi: 10.1128/AEM.00605-09
Geue, L., Schares, S., Mintel, B., Conraths, F. J., Müller, E., and Ehricht, R. (2010). Rapid microarray-based genotyping of enterohemorrhagic Escherichia coli serotype O156:H25/H-/Hnt isolates from cattle and clonal relationship analysis. Appl. Environ. Microbiol. 76, 5510–5519. doi: 10.1128/AEM.00743-10
González, J., Sanso, A. M., Lucchesi, P. M., and Bustamante, A. V. (2014). Comparison of 2 proposed MLVA protocols for subtyping non-O157:H7 verotoxigenic Escherichia coli. Diagn. Microbiol. Infect. Dis. 78, 328–332. doi: 10.1016/j.diagmicrobio.2013.10.017
Gould, L. H., Mody, R. K., Ong, K. L., Clogher, P., Cronquist, A. B., Garman, K. N., et al. (2013). Increased recognition of non-O157 shiga toxin-producing Escherichia coli infections in the United States during 2000-2010: epidemiologic features and comparison with E. coli O157 infections. Foodborne Pathog. Dis. 10, 453–460. doi: 10.1089/fpd.2012.1401
Hunter, P. R., and Gaston, M. A. (1988). Numerical index of the discriminatory ability of typing systems: an application of Simpson's index of diversity. J. Clin. Microbiol. 26, 2465–2466.
Ison, S. A., Delannoy, S., Bugarel, M., Nightingale, K. K., Webb, H. E., Renter, D. G., et al. (2015). Genetic diversity and pathogenic potential of attaching and effacing Escherichia coli O26: H11 strains recovered from bovine feces in the United States. Appl. Environ. Microbiol. 81, 3671–3678. doi: 10.1128/AEM.00397-15
ISPCH (2014). Vigilancia de laboratorio de E. coli productora de toxina Shiga. Chile, 2007–2013, Boletín Instituto de Salud Pública de Chile. Available online at: http://www.ispch.cl/vigilancia-ispch/patogenos
Käppeli, U., Hachler, H., Giezendanner, N., Beutin, L., and Stephan, R. (2011). Human infections with non-O157 shiga toxin-producing Escherichia coli, Switzerland, 2000–2009. Emerg. Infect. Dis. 17, 180–185. doi: 10.3201/eid1702.100909
Krüger, A., and Lucchesi, P. M. (2015). Shiga toxins and stx phages: highly diverse entities. Microbiology 161, 451–462. doi: 10.1099/mic.0.000003
Larkin, M. A., Blackshields, G., Brown, N., Chenna, R., McGettigan, P. A., McWilliam, H., et al. (2007). Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–2948. doi: 10.1093/bioinformatics/btm404
Leomil, L., de Castro, A. F. P., Krause, G., Schmidt, H., and Beutin, L. (2005). Characterization of two major groups of diarrheagenic Escherichia coli O26 strains which are globally spread in human patients and domestic animals of different species. FEMS Microbiol. Lett. 249, 335–342. doi: 10.1016/j.femsle.2005.06.030
Li, M.-C., Wang, F., and Li, F. (2011). Identification and molecular characterization of antimicrobial-resistant shiga toxin–producing Escherichia coli isolated from retail meat products. Foodborne Pathog. Dis. 8, 489–493. doi: 10.1089/fpd.2010.0688
Lindstedt, B. A., Brandal, L. T., Aas, L., Vardund, T., and Kapperud, G. (2007). Study of polymorphic variable-number of tandem repeats loci in the ECOR collection and in a set of pathogenic Escherichia coli and Shigella isolates for use in a genotyping assay. J. Microbiol. Methods 69, 197–205. doi: 10.1016/j.mimet.2007.01.001
Løbersli, I., Haugum, K., and Lindstedt, B. A. (2012). Rapid and high resolution genotyping of all Escherichia coli serotypes using 10 genomic repeat-containing loci. J. Microbiol. Methods 88, 134–139. doi: 10.1016/j.mimet.2011.11.003
Madic, J., Vingadassalon, N., de Garam, C. P., Marault, M., Scheutz, F., Brugère, H., et al. (2011). Detection of shiga toxin-producing Escherichia coli serotypes O26:H11, O103:H2, O111:H8, O145:H28, and O157:H7 in raw-milk cheeses by using multiplex real-time PCR. Appl. Environ. Microbiol. 77, 2035–2041. doi: 10.1128/AEM.02089-10
Michelacci, V., Grande, L., Tozzoli, R., Maugliani, A., Caprioli, A., and Morabito, S. (2014). Identification of two allelic variants of toxB gene and investigation of their distribution among Verocytotoxin-producing Escherichia coli. Int. J. Med. Microbiol. 304, 730–734. doi: 10.1016/j.ijmm.2014.05.009
Mohammed, M. A., Sallam, K. I., Eldaly, E. A. Z, Ahdy, A. M., and Tamura, T. (2014). Occurrence, serotypes and virulence genes of non-O157 shiga toxin-producing Escherichia coli in fresh beef, ground beef, and beef burger. Food Control 37, 182–187. doi: 10.1016/j.foodcont.2013.09.035
Monaghan, A., Byrne, B., Fanning, S., Sweeney, T., McDowell, D., and Bolton, D. J. (2011). Serotypes and virulence profiles of non-O157 shiga toxin-producing Escherichia coli isolates from bovine farms. Appl. Environ. Microbiol. 77, 8662–8668. doi: 10.1128/AEM.06190-11
Morabito, S., Tozzoli, R., Caprioli, A., Karch, H., and Carattoli, A. (2002). Detection and characterization of class 1 integrons in enterohemorrhagic Escherichia coli. Microb. Drug Resist. 8, 85–91. doi: 10.1089/107662902760190626
Mundy, R., Jenkins, C., Yu, J., Smith, H., and Frankel, G. (2004). Distribution of espI among clinical enterohaemorrhagic and enteropathogenic Escherichia coli isolates. J. Med. Microbiol. 53, 1145–1149. doi: 10.1099/jmm.0.45684-0
Naylor, S. W., Gally, D. L., and Low, J. C. (2005). Enterohaemorrhagic, E. coli in veterinary medicine. Int. J. Med. Microbiol. 295, 419–441. doi: 10.1016/j.ijmm.2005.07.010
Noller, A. C., McEllistrem, M. C., Pacheco, A. G., Boxrud, D. J., and Harrison, L. H. (2003). Multilocus variable-number tandem repeat analysis distinguishes outbreak and sporadic Escherichia coli O157:H7 isolates. J. Clin. Microbiol. 41, 5389–5397. doi: 10.1128/JCM.41.12.5389-5397.2003
Oswald, E., Schmidt, H., Morabito, S., Karch, H., Marchès, O., and Caprioli, A. (2000). Typing of intimin genes in human and animal enterohemorrhagic and enteropathogenic Escherichia coli: characterization of a new intimin variant. Infect. Immun. 68, 64–71. doi: 10.1128/IAI.68.1.64-71.2000
Paddock, Z. D., Renter, D. G., Cull, C. A., Shi, X., Bai, J., and Nagaraja, T. G. (2014). Escherichia coli O26 in feedlot cattle: fecal prevalence, isolation, characterization, and effects of an E. coli O157 vaccine and a direct-fed microbial. Foodborne Pathog. Dis. 11, 186–193. doi: 10.1089/fpd.2013.1659
Pearce, M., Evans, J., McKendrick, I., Smith, A., Knight, H., Mellor, D., et al. (2006). Prevalence and virulence factors of Escherichia coli serogroups O26, O103, O111, and O145 shed by cattle in Scotland. Appl. Environ. Microbiol. 72, 653–659. doi: 10.1128/AEM.72.1.653-659.2006
Polifroni, R., Etcheverría, A. I., Sanz, M. E., Cepeda, R. E., Krüger, A., Lucchesi, P. M., et al. (2012). Molecular characterization of shiga toxin-producing Escherichia coli isolated from the environment of a dairy farm. Curr. Microbiol. 65, 337–343. doi: 10.1007/s00284-012-0161-0
Rivas, M., Padola, N. L., Lucchesi, P. M., and Masana, M. (2010). “Diarrheagenic Escherichia coli in Argentina,” in Pathogenic Escherichia coli in Latin America, ed A. G. Torres (Oak Park, IL: Bentham Science Publishers), 142–161.
Rivero, M. A., Passucci, J. A., Rodriguez, E. M., and Parma, A. E. (2010). Role and clinical course of verotoxigenic Escherichia coli infections in childhood acute diarrhoea in Argentina. J. Med. Microbiol. 59, 345–352. doi: 10.1099/jmm.0.015560-0
Sasaki, Y., Usui, M., Murakami, M., Haruna, M., Kojima, A., Asai, T., et al. (2012). Antimicrobial resistance in shiga toxin-producing Escherichia coli O157 and O26 isolates from beef cattle. Jpn. J. Infect. Dis. 65, 117–121.
Scheutz, F., Teel, L. D., Beutin, L., Piérard, D., Buvens, G., Karch, H., et al. (2012). Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 50, 2951–2963. doi: 10.1128/JCM.00860-12
Zhang, W.-L., Bielaszewska, M., Liesegang, A., Tschäpe, H., Schmidt, H., Bitzan, M., et al. (2000). Molecular characteristics and epidemiological significance of shiga toxin-producing Escherichia coli O26 strains. J. Clin. Microbiol. 38, 2134–2140.
Zhao, S., White, D. G., Ge, B., Ayers, S., Friedman, S., English, L., et al. (2001). Identification and characterization of integron-mediated antibiotic resistance among shiga toxin-producing Escherichia coli isolates. Appl. Environ. Microbiol. 67, 1558–1564. doi: 10.1128/AEM.67.4.1558-1564.2001
Zimmerhackl, L. B., Rosales, A., Hofer, J., Riedl, M., Jungraithmayr, T., Mellmann, A., et al. (2010). Enterohemorrhagic Escherichia coli O26:H11-associated hemolytic uremic syndrome: bacteriology and clinical presentation. Semin. Thromb. Hemost. 36, 586–593. doi: 10.1055/s-0030-1262880
Keywords: Shiga toxin-producing Escherichia coli, O26:H11, genetic characterization, microarray, MLVA, hemolytic uremic syndrome, cattle
Citation: Krüger A, Lucchesi PMA, Sanso AM, Etcheverría AI, Bustamante AV, Burgán J, Fernández L, Fernández D, Leotta G, Friedrich AW, Padola NL and Rossen JWA (2015) Genetic characterization of Shiga toxin-producing Escherichia coli O26:H11 strains isolated from animal, food, and clinical samples. Front. Cell. Infect. Microbiol. 5:74. doi: 10.3389/fcimb.2015.00074
Received: 31 July 2015; Accepted: 05 October 2015;
Published: 20 October 2015.
Edited by:
Alfredo G. Torres, University of Texas Medical Branch, USAReviewed by:
Catherine Maeve Burgess, Teagasc, IrelandRodney A. Moxley, University of Nebraska-Lincoln, USA
Copyright © 2015 Krüger, Lucchesi, Sanso, Etcheverría, Bustamante, Burgán, Fernández, Fernández, Leotta, Friedrich, Padola and Rossen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alejandra Krüger, YWtydWdlckB2ZXQudW5pY2VuLmVkdS5hcg==;
Nora Lía Padola, bmxwYWRvbGFAdmV0LnVuaWNlbi5lZHUuYXI=