 Alfonso Miranda†
Alfonso Miranda† Bárbara Ávila†Patricia Díaz
Bárbara Ávila†Patricia Díaz Lina RivasKaren Bravo
Lina RivasKaren Bravo Javier AstudilloConstanza BuenoMaría T. UlloaGermán Hermosilla
Javier AstudilloConstanza BuenoMaría T. UlloaGermán Hermosilla Felipe Del Canto
Felipe Del Canto Juan C. Salazar
Juan C. Salazar Cecilia S. Toro*
Cecilia S. Toro*- Programa de Microbiología y Micología, Facultad de Medicina, Instituto de Ciencias Biomédicas, Universidad de Chile, Santiago, Chile
The most common mechanism of trimethoprim (TMP)-resistance is the acquisition of dihydrofolate reductase enzyme resistant to this drug. Previous molecular characterization of TMP-genes resistance in Chilean isolates of Shigella sonnei searching for dfrA1 and dfrA8, showed solely the presence of dfrA8 (formerly dhfrIIIc). However, these genetic markers were absent in S. sonnei strains further isolated during an outbreak in 2009. To identify the TMP-resistance gene in these strains, a genomic DNA library from a TMP-resistant (TMPR) S. sonnei representative strain for the outbreak was used to clone, select and identify a TMP-resistance marker. The TMPR clone was sequenced by primer walking, identifying the presence of the dfrA14 gene in the sul2-strA'-dfrA14-‘strA-strB gene arrangement, harbored in a native 6779-bp plasmid. The same plasmid was isolated by transforming with a ~4.2 MDa plasmid extracted from several TMPR S. sonnei strains into Escherichia coli. This plasmid, named pABC-3, was present only in dfrA14-positive strains and was homologous to a previously described pCERC-1, but different due to the absence of an 11-bp repetitive unit. The distribution of dfrA1, dfrA8, and dfrA14 TMP-resistance genes was determined in 126 TMPR S. sonnei isolates. Most of the strains (96%) carried only one of the three TMP-resistance genes assessed. Thus, all strains obtained during the 2009-outbreak harbored only dfrA14, whereas, dfrA8 was the most abundant gene marker before outbreak and, after the outbreak dfrA1 seems have appeared in circulating strains. According to PFGE, dfrA14-positive strains were clustered in a genetically related group including some dfrA1- and dfrA8-positive strains; meanwhile other genetic group included most of the dfrA8-positive strains. This distribution also correlated with the isolation period, showing a dynamics of trimethoprim genetic markers prevalent in Chilean S. sonnei strains. To our knowledge, dfrA14 gene associated to a small non-conjugative plasmid was detected for the first time in Shigella. Apparently, the strain causing the outbreak must have been introduced, changing drastically the genetic distribution of trimethoprim resistance in Chilean S. sonnei strains.
Introduction
Shigella sonnei has become a major problem of public health due to the increasing multidrug-resistance to antibiotic (MDR) worldwide (Ashkenazi et al., 2003; De Lappe et al., 2003; Seol et al., 2006; Vrints et al., 2009). This pathogen, one of the most frequent etiologic agents of foodborne diseases in industrialized countries, is responsible for shigellosis, an acute enteric disease for which antimicrobial therapy is usually recommended to manage infection and reduce fecal excretion of the bacterium to prevent further dissemination. Consequently, a dramatic increase in the rate of resistance to commonly used drugs is observed. One example is the resistance to sulfamethoxazole and trimethoprim (TMP) that has increased since they were introduced as antimicrobial therapy (Huovinen et al., 1995; Huovinen, 1997), especially in Escherichia coli and Shigella.
Molecular mechanisms of cotrimoxazole (sulfamethoxazole/trimethoprim) resistance could be explained by resistance to trimethoprim and/or to sulfonamides. Specifically for TMP-resistance, several mechanisms have been described (Huovinen et al., 1995; Huovinen, 1997, 2001); however, the most common mechanism is the acquisition of dihydrofolate reductase (DFR) enzyme resistant to this drug. More than 30 different genes are identified encoding TMP-resistant enzymes. They can categorize in two major families based on the length of the N termini of the enzymes and the resistance level they confer: Family A coded by dfrA genes and B coded by dfrB genes (Recchia and Hall, 1995; Seputiené et al., 2010). The most common dfr genes described in Enterobacteriaceae are encoded by plasmids, transposons or integrons (Seputiené et al., 2010; Ke et al., 2011; Labar et al., 2012; Cavicchio et al., 2015; Shin et al., 2015). In Chile, between 40 and 60% of the strains isolated from shigellosis cases in the last 5 years corresponds to S. sonnei. Noteworthy, most of these isolates are resistant to multiple antimicrobial agents. Surveillance of in vitro susceptibility to antimicrobials demonstrated that S. sonnei have evolved to MDR isolates (Marcoleta et al., 2013). Particularly, it is notable that resistance to cotrimoxazole increased from 50 to 100%, in strains isolated since 1995 to 2009, year in which an outbreak occurred. The outbreak (739 cases) mainly affected children under 10 years from Región Metropolitana, and the source of infection was not clearly established (Instituto de Salud Pública, 2009).
Previous molecular characterization of TMP-gene resistance in Chilean isolates of S. sonnei, searching for dfrA1 and dfrA8, showed solely the presence of dfrA8 (formerly dhfrIIIc) (White and Rawlinson, 2001), linked to a conjugative plasmid harboring also the blaTEM gene (Toro et al., 2005). However, S. sonnei strains isolated during the outbreak in 2009 had been negative in detection of dfrA1 and dfrA8 TMP-resistance genetic markers. Therefore, in this study we identified and characterized the genetic determinants of TMP-resistance present in S. sonnei strains isolated during this outbreak and the distribution of those markers in strains isolated from 1995 to 2013.
Materials and Methods
Bacterial Strains and Culture Conditions
One hundred twenty-six TMPR S. sonnei strains obtained in Chile from 1995 to 2013, from stool samples of patients suffering acute diarrhea were studied. These strains were identified by conventional and automated biochemical methods (VITEK-2, Biomérieux), and serotyped by agglutination with type-specific antisera (Denka-Seiken, Tokyo, Japan). In addition, 51 foreign S. sonnei strains, kindly provided by Dr. FX. Weill from the Institut Pasteur collection, were included in this study (Table 1). E. coli DH5α nalidixic acid resistant strain (NALR) was used as the recipient in transformation and conjugation experiments. All bacterial strains were routinely cultured at 37°C on LB broth or agar, or trypticase soy, supplemented with ampicillin (AMP) 100 mg L−1, NAL or TMP 30 mg L−1 when it was required. E. coli V517 and E. coli 39R861 were used as plasmid size standards.
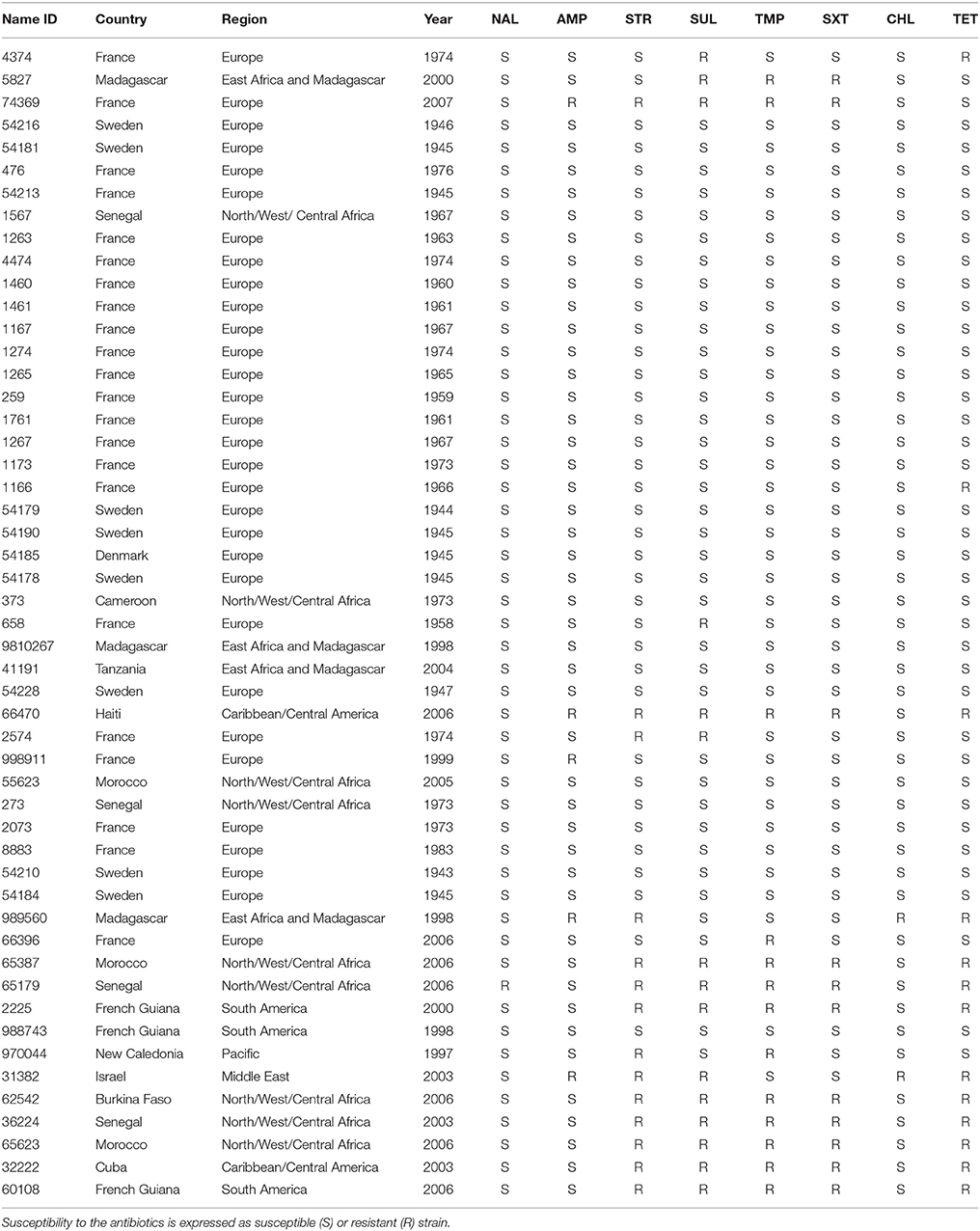
Table 1. Antimicrobial susceptibility characterization of 51 Shigella sonnei strains isolated since 1943 to 2006 from different origins worldwide.
Antimicrobial Susceptibility
Antimicrobial susceptibility was determined by disk diffusion and microdilution methods by following Clinical and Laboratory Standards Institute guidelines (Clinical Laboratory Standars Institute, 2013). AMP, 10 μg; NAL, 30 μg; ciprofloxacin (CIP), 5 μg; chloramphenicol (CHL), 30 μg; streptomycin (STR), 10 μg; sulfamethoxazole/trimethoprim (SXT), 23.75/1.25 μg; tetracycline (TET), 30 μg and trimethoprim (TMP), 30 μg, were used for disk diffusion test. The reference strain E. coli ATCC 25922 was included as a quality control. For analysis purposes, intermediate and resistant isolates were considered together.
Construction of S. sonnei DNA Library
Genomic DNA was obtained from a selected S. sonnei TMPR strain, grown overnight in LB media at 37°C, using the kit E.Z.N.A Bacterial® DNA Kit as described by the provider (Omega Bio-Tek, USA). Further, partially HindIII-digested DNA was separated in 1% agarose gel and the fragments ranging from 2 to 10 kbp were purified using QIAquick gel extraction kit (Qiagen). Meanwhile, the pUC19 plasmid was digested with HindIII, dephosphorylated with the thermo labile alkaline phosphatase FastAP as described by the vendor (Thermo-Scientific, USA), and gel-purified. Both, the genomic DNA fragments and the digested plasmid were incubated with T4 DNA ligase (Thermo-Scientific, USA) for 24 h at 4°C in a ratio of 3:1 (insert: vector). The ligation mixture was transformed into E. coli DH5α cells, and recombinants were recovered overnight at 37°C in presence of TMP 50 mg L−1. The recombinant plasmid obtained (pCLON3) was purified from the isolated TMPR E. coli, using QIAprep Spin Miniprep Kit (QIAGEN) and sequenced (Macrogen, Korea). The 6779-bp fragment cloned was released from the recombinant plasmid pCLON3 by HindIII digestion; then gel-purified, self-ligated and transformed into E. coli DH5α. Finally, E. coli transformants selected in TMP were confirmed to carry the reconstituted native plasmid, named pABC-3. The TMPR genetic marker harbored in this plasmid was dfrA14.
Detection of Trimethoprim Resistance Markers
DNA templates were obtained from individual colonies by bacterial lysis or by purifying total genomic DNA. Primers used are indicated in Table 2. PCR reactions for dfrA1, dfrA8, dfrA14 were performed as follow: an initial 2 min denaturation cycle at 95°C followed by 30 cycles at 95°C for 1 min, 56°C for 30 s, and 72°C for 1 min for dfr primers respectively, with a final extension at 72°C for 10 min. PCR products were analyzed by electrophoresis in 2% agarose gels and stained with ethidium bromide.
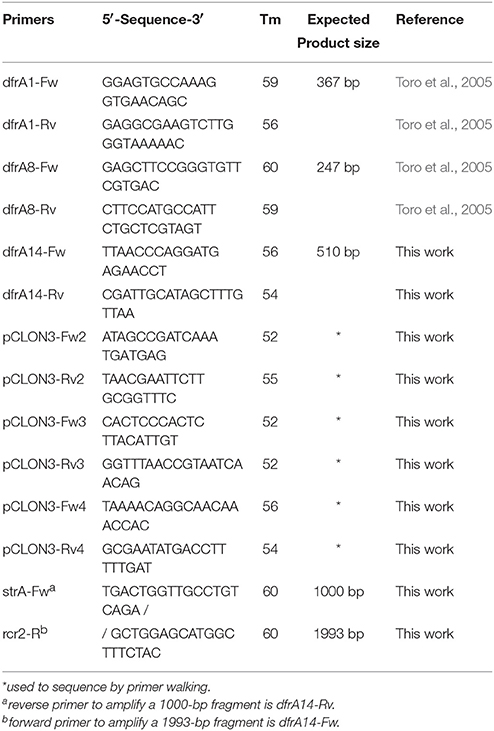
Table 2. PCR primers used in this study.
Purification and Isolation of Native Plasmid Containing the dfrA14 Gene
The native plasmids were obtained from a representative S. sonnei strain using E.Z.N.A. plasmid DNA mini kit II (OMEGA Bio-Tek) or by alkaline extraction, separated in a 1% agarose gel electrophoresis, and visualized with ethidium bromide. Two plasmids of ~4.2 MDa were purified using QIAquick gel extraction kit (Qiagen). Chemically competent E. coli DH5α (Inoue et al., 1990) were transformed with 250 ng of the plasmid gel-isolated and selected in presence of TMP 30 mg L−1. Finally, plasmid extracted from the TMPR transformant bacteria, was sequenced and compared with the reconstituted plasmid, pABC-3, showing 100% identity. pABC-3 restriction analysis in silico was performed using NebCutter software. SspI was selected for restriction fragment long polymorphism (RFLP) analysis producing 3 bands of 2920, 2343, and 1516 bp. Restriction was made using 5 μg of plasmid and SspI 1U μg−1 in 20 μL of mix reaction during 2 h at 37°C.
Conjugation Analysis
Conjugations were carried out with 10 TMPR- S. sonnei strains using NALR- E. coli DH5α as recipient. The overnight culture of each purified donor and recipient cells were diluted 100-fold in 2 mL of fresh LB broth and incubated at 37°C until OD600 reached about 0.6. Then 0.2 mL each of the donor and recipient cultures was mixed. After incubation at 37°C without shaking for 3–4 h, the mixture was plated on LB agar supplemented with TMP 30 mg L−1 and NAL 30 mg L−1. Plasmid extraction and PCR reactions for the dfrA14 gene were applied to confirm the transconjugants that acquired plasmids and resistance genetic marker. Changes of the susceptibility to antimicrobial agents were measured by the disk diffusion method as described above.
Sequence Analysis
Comparative analysis of nucleotide sequences was performed using BLAST at the National Center for Biotechnology Information (NCBI) (site www.ncbi.nlm.nih.gov/BLAST/). The pABC-3 plasmid sequence was deposited at the GenBank database under accession number KT988306. Graphic representation of the loci alignment was performed using EasyFig v2.1 (Sullivan et al., 2011), with the blastn algorithm. Plasmid sequences incorporated in this comparison were: Klebsiella pneumoniae pKDO1 (GenBank accession number NC_019389.1), E. coli pPGRT46 (KM023153.1), E. coli pSTOJO1 (AJ313522.1), Yersinia ruckeri pYR1521 (NG_041026.1), E. coli pCERC1 (NC_019070.1), S. sonnei pKKTET7 (NC_008439.1), S. sonnei pSS4 (AF534183.1), Salmonella Typhimurium pSRC15 (NC_013104.1), Shigella flexneri pSFxv_3 (CP001386.1), E. coli pCN061p3 (CP006639.1), Pasteurella multocida pVM111 (NG_035903.1). More information about these plasmids is detailed in the Supplementary Table.
Pulsed Field Gel Electrophoresis (PFGE)
Genomic DNA from the selected strains was included into agarose plugs and digested with endonuclease XbaI (Thermos Scientific). According to PulseNet protocol (CDC), the macrorestriction fragments were separated by pulsed-field gel electrophoresis on a CHEF-DRIII Chiller system (Bio-Rad Laboratories, Richmond, CA), in 1% agarose gel using 0.5X TBE buffer at 6 V/cm and 14°C, with ramped pulse times of 2.2 to 54.2 s for 21 h. Salmonella enterica serovar Braenderup was included as molecular size standard. The DNA band profiles were analyzed with GelCompar software (version 3.0; Applied Maths, Sint-Martens-Latem, Belgium). The cluster analysis and generation of dendrograms was performed using UPGMA with a 1.5% band tolerance. The similarity between DNA profiles was determined using Dice's correlation coefficient. We defined arbitrarily pulsotypes and pulsogroups with similarities of >91.1% and 75%, respectively.
Results
Identification of dfrA14 TMP-Resistance Genetic Marker in S. sonnei Strains
Resistance to TMP has been increasing rapidly worldwide, since this antibiotic was introduced. Chilean S. sonnei strains isolated from 1995 to 2013 also displayed increased level of TMP-resistance as measured by disk diffusion, from 49 up to 81% and exceptionally 100% during the outbreak in the summer of 2008–2009 (Figure 1A). Seventy-five percent, 264 out of the total amount of 350 S. sonnei strains, were TMPR. To characterize the molecular determinants of TMP-resistance in these Chilean S. sonnei strains, we randomly chose 126 TMPR strains obtained from different periods, divided as 3 groups: 45 strains before the outbreak (isolated since 1995–1997 and 2004–2006 period), the 41 strains during the outbreak (2008–2009), and 40 strains isolated after the outbreak (2010–2013). Previously, we demonstrated that S. sonnei strains isolated in 1995 harbored solely dfrA8 gene (Toro et al., 2005). Detection of dfrA1 and dfrA8 by PCR was negative in the 41 S. sonnei strains collected during the 2008-2009 outbreak.
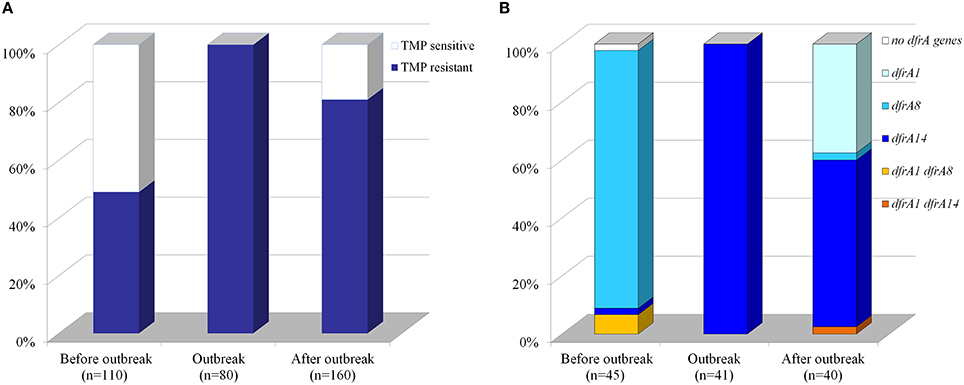
Figure 1. Distribution of trimethoprim-resistant strains and trimethoprim-genetic markers among Chilean Shigella sonnei isolates. (A) Percentage of TMP-susceptible and–resistant strains. Susceptibility to TMP was measured by disk diffusion in a total of 350 strains isolated before, during or after the 2008–2009 outbreak. A total of 264 strains were TMPR including the 80 strains obtained during the outbreak. (B) Percentage of strains harboring TMP-resistance genetic markers. Presence of dfrA1, dfrA8, and dfrA14 was assessed by PCR in a smaller group (126 strains) randomly selected from TMPR S. sonnei strains.
To identify the TMP-resistance gene in these strains, a DNA library was done in pUC19 (AmpR) using HindIII-digested genomic DNA from the TMPR S. sonnei C8225 as representative strain for the outbreak and E. coli DH5α as host. Recombinant plasmids from TMPR clones were selected, purified, and transformed into a new E. coli DH5α, isolating the TMP-genetic marker in the plasmid named pCLON3. The dfrA14 gene was identified as the TMP-resistance marker by sequencing the insert of this recombinant plasmid and comparing with available data in the GenBank database.
Distribution of TMP-Resistance Gene Markers in S. sonnei Strains
To determine the presence of dfrA14 gene in Chilean S. sonnei strains, specific primers for this marker were designed (Table 2). Thus, only 1 out 45 strains isolated before the outbreak harbored dfrA14, meanwhile 100% of strains belonged to the outbreak displayed only the dfrA14 TMP-genetic marker and 24 out 40 isolated after the outbreak carried this gene (Figure 1B). Most of the strains isolated before the outbreak (96%) displayed the dfrA8 gene marker. In contrast, dfrA1 was detected in 40% after the outbreak. The dfrA1/dfrA8 combination was detected only in three strains isolated before the outbreak and dfrA1/dfrA14 in one strain isolated after the outbreak. The dfrA8/dfrA14 combination was not present in this Chilean group of strains (Figure 1B).
Considering the 126 strains, most of them (96%) carried one of the three TMP-resistance gene assessed; only one strain (0.8%), isolated before the outbreak was negative for the three genes, and just 4 strains (3%) displayed two resistance genes. Fifty-two percent of all strains harbored only dfrA14, whereas 32% of strains displayed dfrA8 and 12% dfrA1.
Characterization of the Plasmid-Borne dfrA14 Gene in S. sonnei Strains
The recombinant plasmid harboring the dfrA14 TMP-marker (pCLON3) sequenced by primer walking, was bioinformatically analyzed and sequences revealed an insert of a 6779-bp fragment harboring the dfrA14 cassette in the sul2-strA'-dfrA14-‘strA-strB arrangement. Alignment of the complete 6779-bp fragment showed most likely identity with the pCERC-1 plasmid (Anantham and Hall, 2012; accession number JN012467.1). Only one specific difference was detected compared to pCERC-1 plasmid: a deletion of an 11-bp short repeat at orf3.
According to this analysis, the cloned sequence corresponded to a whole native plasmid. To validate this finding, the insert was released from the recombinant plasmid pCLON3 by HindIII digestion (Figure 2, lane 2); then, the 6779-bp fragment was gel-purified, self-ligated and transformed into E. coli DH5α. Finally, transformants selected in TMP were confirmed to carry the reconstituted native plasmid with the TMPR genetic marker, dfrA14 (Figure 2, lane 3). This plasmid was named pABC-3 (GenBank accession number KT988306). Wild type and transformants strains were checked for their antimicrobial susceptibility profile to 8 antibiotics. E. coli transformant strains with pCLON3 and pABC-3 were resistant to NAL, SXT, and TMP but susceptible to tetracycline and streptomycin, measured by disk diffusion, indicating that sul2 present in the cloned plasmid is functional and the str genes are mutated by dfrA14 insertion. pCLON3 coded additionally the Amp-resistance harbored in the pUC19 vector. MIC to TMP was measured in S. sonnei C8225, DH5α/pCLON3 and DH5α/pABC-3. All of them raised the MIC over 1000 mg L−1, as expected for a dfr type A coded enzyme as dfrA14.
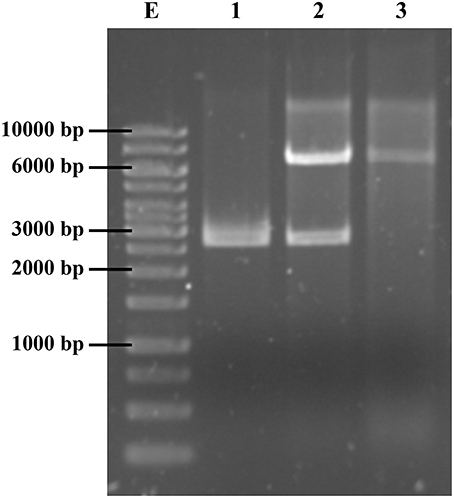
Figure 2. Isolation of plasmid pABC-3, harboring the dfrA14 gene. Lane 1, pUC19 digested with HindIII. Lane 2, pCLON3, recombinant pUC19 harboring a ~7 kb fragment TMP-resistance marker, digested with HindIII. Lane 3, the native pABC-3 plasmid reconstituted by self-ligation and recovered from DH5α TMPR transformants, digested with HindIII.
In parallel, the plasmid electrophoretic profiles for S. sonnei TMPR dfrA14-positive strains isolated in 2008-2009 was analyzed. As Figure 3A shows, a pattern including a double band of ~4.2 MDa, was found in all of them. Size estimation of plasmids was done by comparison with E. coli V517 and E. coli 39R861 standards plasmids. On the other hand, the double band was absent in S. sonnei TMPR strains isolated before the outbreak (Figure 3B). To determine whether one of those plasmids harbor the dfrA14 gene, double band gel-isolated of several dfrA14–positive S. sonnei strains belonged to the outbreak was used to transform E. coli DH5α and to select TMP-resistance. By this way, it was possible to isolate one plasmid, whose size was similar to the smaller band (Figure 3C). The detection of dfrA14 gene was positive only for TMPR transformants. RFLP of this plasmid was conducted using plasmid DNA extracted from 10 different E. coli transformant strains, using SspI restriction enzyme; thus, it was verified that isolated plasmids harbored the same restriction pattern, as expected for pABC-3 (Figure 3D), suggesting the presence of the same plasmid. Complete sequences of both TMPR plasmids, the one isolated by direct transformation and the other one reconstituted from the TMPR clone, pABC-3, were compared and it was confirmed that they were identical.
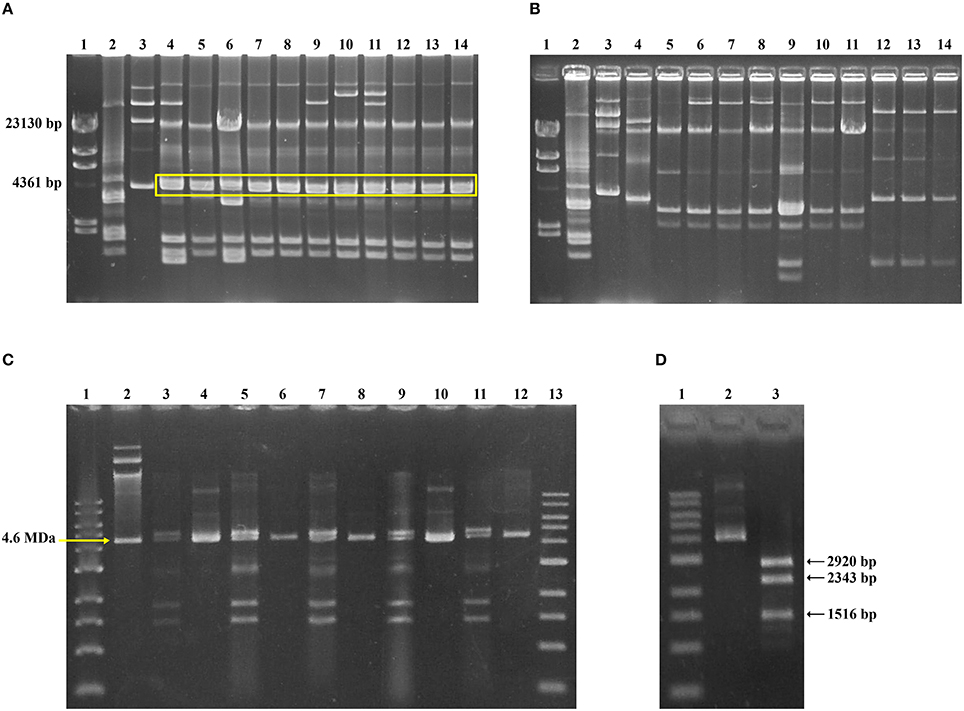
Figure 3. Characterization of pABC-3 plasmid in Chilean TMP-resistant Shigella sonnei strains. Plasmid profiles of representative TMPR S. sonnei strains isolated during the outbreak (A, lane 4 – 14) and before the outbreak (B, lane 4 – 14), obtained by alkaline extraction and separation in 1% agarose gel electrophoresis. Molecular size markers in (A,B): lane 1, Lambda HindIII DNA marker; lane 2, E. coli V517; lane 3, E. coli 39R861. A duplex band migrating near to 4.2 MDa is highlighted by a yellow line in panel A, which is absent in (B). (C) Involvement of pABC-3 plasmid in TMP-resistance. Plasmid profiles from representative TMPR S. sonnei strains next to the corresponding TMPR E. coli DH5α transformant are shown. Lane 1 and 13, 1 kb molecular size marker (New England Biolabs®); lane 2, E. coli 39R861; lane 3 – 4, S. sonnei strain c0027 and the respective TMPRE. coli DH5α transformant; lane 5 – 6, S. sonnei strain c0700 and the TMPR transformant; lane 7 – 8, S. sonnei strain c0719 and the TMPR transformant; lane 9 – 10, S. sonnei strain c8072 and the TMPR transformant; lane 11 – 12, S. sonnei strain c8205 and the TMPR transformant. (D) A representative SspI digestion pattern of pABC-3, in concordance with the in silico predicted size. Lane 1, 1 kb molecular size marker (New England Biolabs®); lane 2, pABC-3 plasmid undigested; lane 3, pABC-3 plasmid digested with SspI.
To link the dfrA14 gene to the presence of the sul2-strA'-dfrA14-‘strA-strB arrangement and the 6779-bp plasmid in the rest of the strains, two overlapping PCR reactions were designed to detect the neighborhood of dfrA14 gene (Table 2). All of the TMPR strains were positive for both reactions, indicating the dfrA14 marker was harbored in this plasmid (data not shown).
In addition, the presence of dfrA14 was evaluated in 51 foreign S. sonnei strains, isolated since 1943 to 2006 from different origins worldwide (Table 1). Thirteen were TMPR, and they did not harbor this marker, however, 10 of them display only dfrA1.
Conjugation experiments were done to determine whether the native pABC-3 plasmid was transferable. However, no transconjugants cells were obtained in TMP when dfrA14–positive S. sonnei strains were used as donors. As control, dfrA8 and blaTEM could be conjugated in the same experimental conditions (Toro et al., 2005). Consequently, no mobilization genes were identified in the sequence of pABC-3. Further in silico analysis detecting incompatibility group based on PCR-based replicon typing (PBRT) showed no similarity with the 18 major incompatibility (Inc) groups of Enterobacteriaceae species (Carattoli et al., 2005). Moreover, using plasmid finder to identify replication origins (http://www.genomicepidemiology.org), using a threshold of 80% we detected 89% identity only with ColRNAI (accession number DQ298019.1), suggesting the presence of an undescribed variant plasmid (Carattoli et al., 2014).
Clonal Analysis of TMP-Resistant S. sonnei Strains
PFGE was performed for all the 126 TMPR S. sonnei strains using XbaI digestion. Figure 4 shows two pulsogroups: A is the minor group included 34 strains, 32 of them belonged to the 45 strains isolated before the outbreak (71%); pulsogroup B is a more heterogenous group (n = 92 strains), including most of the strains isolated during and after the outbreak (97%, n = 79/81). Considering the distribution of dfrA genes, within the pulsogroup A, 32 of them were dfrA8-positive strains and separated in two pulsotypes A1 and A2. In the pulsogroup B, the most frequent profiles are clustered in three pulsotypes. The pulsotype B1 contained 43 strains, from which 40 (93%) harbored the dfrA14 gene; the B2 grouped 12 strains, 11 (92%) harbored the dfrA14 gene. Noteworthy, the pulsotype B3 (11 strains), contained the only strain negative for the three TMP-resistance genes studied, 6 dfrA8-positive strains isolated before the outbreak, 3 dfrA14-positive strains isolated after the outbreak and one dfra1-positive strain isolated after the outbreak. From the 26 remaining strains from pulsogroup B, most of them presented individual pulsotypes and contained dfrA1 or dfrA14 genes.
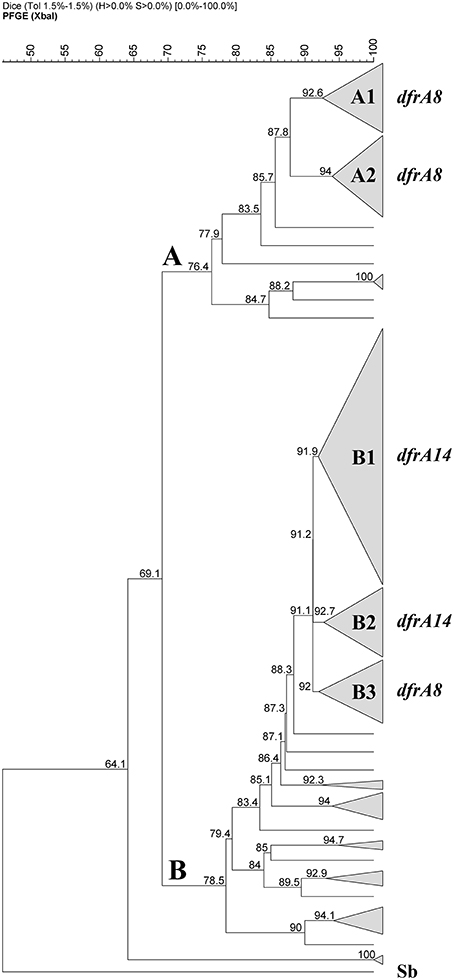
Figure 4. Dendrogram based on Dice coefficients of similarity for 126 TMP-resistant Shigella sonnei strains. The dendrogram was constructed by UPGMA (GelCompar II). Figure shows two pulsogroups (A,B) with a similarity of 69.1%. Twenty-three pulsotypes were defined with a similarity >91%. Within the pulsogroup A, most of the strains were grouped in A1 and A2 pulsotypes, both containing dfrA8 as TMP-resistance marker. Within the pulsogroup B, most of the strains were clustered in three pulsotypes, B1 and B2 include the dfrA14-positive strains and B3 dfrA8-positive strains isolated before the outbreak. Sb, strain of Salmonella serotype Braenderup, digested with XbaI as a size marker.
Together, these results shows a drastic change of the genetic distribution of trimethoprim resistance in Chilean S. sonnei strains.
Discussion
The high increase of TMP-resistance is a worldwide event especially in Gram-negative bacteria (Huovinen et al., 1995; Huovinen, 1997, 2001). In this study, we sought to identify TMP-resistance genes in S. sonnei strains that lacked dfrA1 and dfrA8, the previously described dfr genes (Toro et al., 2005).
Firstly, characterizing TMPR S. sonnei strains belonged to the 2009-oubreak, we cloned the TMP-resistance genetic determinant identified as dfrA14, formerly named dhfrlb (Recchia and Hall, 1995) and harbored in a small plasmid (6779 bp). Sequence of this plasmid was homologous to the pCERC-1, previously found in commensal E. coli (Anantham and Hall, 2012). Both plasmids display the same particular arrangement including the sul2-strA'-dfrA14-‘strA-strB genes (Figure 5, and Supplementary Table), which has also been described in plasmids isolated from other enterobacteria such as uropathogenic E. coli (Ojo et al., 2002) and more recently in a larger plasmid (8.9 kb) from the fish pathogen Y. ruckeri (Huang et al., 2014). Interestingly, the strA'-dfrA14-‘strA insertion was also reported in the pM3224T plasmid including the same arrangement with sul2 and strB cassettes (6050 bp) and with a different architecture for strA'-dfrA14-‘strA and sul2 genes in the mobilizable pM3389T plasmid (6101 bp). Both of these plasmids are harbored by Actinobacillus pleuropneumoniae, a distantly related species belonging to the Pasteurellaceae family that causes respiratory infections in pigs (Bossé et al., 2015). Recently, a novel plasmid (83 kb) isolated in commensal E. coli carrying blaCTX-M-15, sul2-strA-strB cluster and dfrA14 was described; however, the TMP-resistance gene was not inserted in sul2-strA-strB arrangement (Fortini et al., 2015).
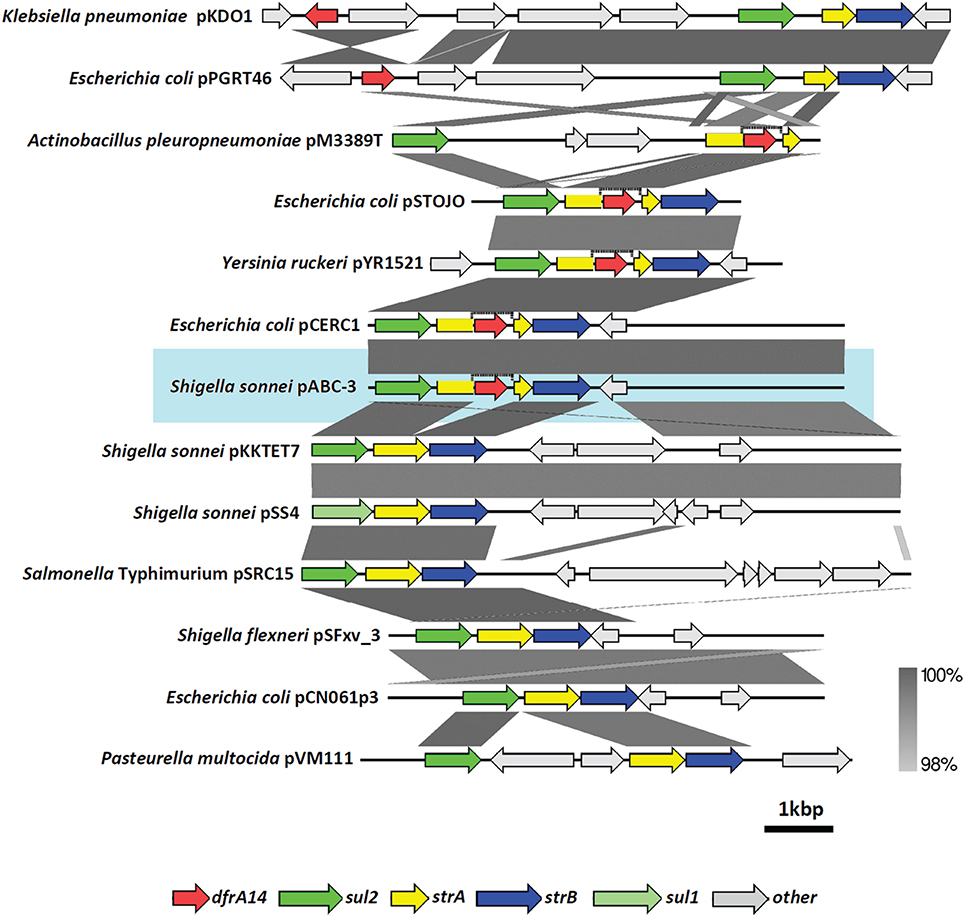
Figure 5. Comparison of different plasmids harboring dfrA14 gene. Alignment of Shigella sonnei pABC-3 with other antibiotic resistance gene clusters contained in plasmids of Gram negative bacteria, which harbor dfrA14 and/or the sul2-strA-strB arrangement. dfrA14 gene is represented by the red arrow. The graphic was built with Easyfig using the blastn algorithm. Vertical bar at the right bottom indicates the threshold of identity percentages in the alignments. More details about plasmids are described in Supplementary Table.
The antibiotic resistance gene cluster containing sul2, strA, and strB genes is also widespread among Gram-negative bacteria and this array can display different combinations with other resistance genes different from dfrA14, such as catA3 or tet genes, giving rise to new resistance gene clusters (Kehrenberg and Schwarz, 2002; Kehrenberg et al., 2003). In S. sonnei, two plasmids similar to pABC-3, pKKTET7 (8401 bp) and pSS4 (8384 bp), have been previously found in strains isolated in Korea, (accession number AF497970-1 and AF534183 respectively). By comparing the pABC-3 (from Chilean strains) with pKKTET7 and pSS4 sequences, it seems that the core is shared among them and the only difference is the presence of tetR and tetA genes instead of dfrA14 in the latter two plasmids. This suggests the exchange of antimicrobial resistance cassettes within a common backbone (Figure 5).
On the other hand, Salmonella Typhimurium isolated in Australia is an example of bacteria harboring this sul2-strA-strB cluster in the pSRC15 plasmid (Figure 5; Yau et al., 2010). In S. flexneri, a plasmid named pSFxv_3 (6200 bp) shares almost 100% of sequence identity with pKKTET-7, including the sul2-strA-strB region. However, pSFxv_3 did not display tet nor dfr markers (Ye et al., 2010). Another example in S. flexneri is the 4.3-MDa plasmid harboring sul2 gene but not dfr gene markers, suggesting that sul2-strA-strB gene cluster is present (Iqbal et al., 2014). This plasmid isolated in samples from Bangladesh was not sequenced but it seems to be the same as pSFxv_3. Thus, to our knowledge, the sul2-strA'-dfrA14-‘strA-strB cluster found in the S. sonnei pABC-3 is described for the first time in Shigella (Figure 5).
Earlier reports established that Shigella TMP-resistance was mediated by transposable genetic elements inserted in a conjugative, multiple antibiotic-resistance plasmid (Tonin and Grant, 1987). Later, the dfrA14 gene was described as the dominant gene among TMPR-plasmids isolated from uropathogenic bacteria in Scotland. This genetic determinant was sequenced and found to be present within a Tn7-like structure (Young and Hillyear, 1994; Young et al., 1994). These features were not similar to the dfrA14 harbored by pABC-3, which is a small plasmid without transposon-like elements.
Antimicrobial resistance phenotype in enterobacteria is often associated with the presence of gene cassettes harbored in integrons. Thus, most of the reports have described TMP-resistance linked to class 1 or class 2 integrons. In this scenario, dfrA1, dfrA12, dfrA15, and dfr17 cassettes are the principal genetic marker displayed at integrons in enterobacteria (Pan et al., 2006; Dubois et al., 2007; Gassama-Sow et al., 2010; Ke et al., 2011; Zhu et al., 2011; Shin et al., 2015). Although the majority of dfr genes seem to be coded within integrons, just some clusters carrying dfrA14 are associated to class 1 or 2 integrons (Kadlec and Schwarz, 2008; Wei et al., 2014). Bioinformatic analysis from this work demonstrated that dfrA14 is harbored in pABC-3 without integron-like elements (data not shown). The presence of dfrA1 and dfrA8 genes coded in integrons has being analyzed.
Recently, dfrA14 was found for the first time in S. flexneri strains by whole genome sequencing. Integration of the genetic information with geographical and temporal data showed that this gene could be detected since 1990; however, no information about the genetic localization or genetic context is provided (Connor et al., 2015).
Interestingly, comparing the orf3 sequence described in pCERC-1 among Shigella plasmids, we detected a variable region of a 11-bp unit (GATGTAAAAGT) repeated five times in pCERC-1, pKKTET7 and pSS046; however, pSFxv_3 and pSS4 displayed only three out these five repetitions. pABC-3, described in this work, had four repetitions, suggesting that this region may discriminate different plasmids.
According to PFGE, all dfrA14-positive strains were clustered in pulsogroup B, grouping 93% of them in pulsotypes B1 and B2, isolated during and after the outbreak. This temporal coincidence suggests the appearance of strains genetically different during the outbreak, harboring the dfrA14-coded plasmid. Apparently, the strains causing the outbreak might have been introduced to the country or they were locally selected by horizontal gene transfer; however, our data do not allow us to discriminate between both hypothesis. Moreover, in neighbor countries have no described isolates similar to Chilean strains. Most of reports from Argentine, Peru, and Brazil described the antibiotic resistance profiles of circulating strains, highlighting in some cases the high level of TMP-resistance; however they do not search for dfrA14 (Merino et al., 2004; Lluque et al., 2015; Seribelli et al., 2016).
To conclude, to our knowledge, this is the first time that the dfrA14 TMP-resistance gene has been found in S. sonnei isolates linked to a small plasmid, becoming one of the most common in Chilean TMPR S. sonnei strains, besides dfrA1 and dfrA8 genes. Apparently, the strain causing the outbreak must have been acquired, changing drastically the temporal dynamics of trimethoprim resistance in Chilean S. sonnei strains and highlighting the urgency to maintain permanent surveillance of antimicrobial resistance profiles and the molecular mechanisms of resistance to improve both prevention and treatment of shigellosis.
Author Contributions
AM: data acquisition, data analysis, data interpretation; BA: data acquisition, data analysis, data interpretation; PD: data analysis, data interpretation, revising of the manuscript; LR: data acquisition, data analysis; JA: data acquisition, data analysis, data interpretation; KB: data acquisition, data analysis, data interpretation; CB: data acquisition, data analysis, data interpretation; MU: study design, data interpretation, revising of the manuscript; GH: data interpretation, revising of the manuscript; FC: data analysis, data interpretation, revising of the manuscript; JS: data analysis, data interpretation, writing of the manuscript, revising of the manuscript; CT: study design, data analysis, data interpretation, writing of the manuscript, revising of the manuscript. MU, GH, and CT are principal investigator at the FONDECYT grant that funded this work.
Funding
This work was supported by Fondo Nacional de Desarrollo Científico y Tecnológico, FONDECYT grant 1130394.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. FX Weill, from Institut Pasteur, Unité des Bactéries Pathogènes Entérique, Institut Pasteur, F-75724, Paris Cedex 15, France, for kindly provide foreign S. sonnei strains. We thank also Dr R. Camponovo, from Integramedica and Dr. Juan Silva from Universidad de Antofagasta, for providing strains belonged to the 2008-2009 outbreak. Finally, we thank Instituto de Salud Pública, Chile, for giving S. sonnei strains isolated during 2010-2013 period.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fcimb.2016.00077
References
Anantham, S., and Hall, R. M. (2012). pCERC1, a small, globally disseminated plasmid carrying the dfrA14 cassette in the strA gene of the sul2-strA-strB gene cluster. Microb. Drug Resist. 18, 364–371. doi: 10.1089/mdr.2012.0008
Ashkenazi, S., Levy, I., Kazaronovski, V., and Samra, Z. (2003). Growing antimicrobial resistance of Shigella isolates. J. Antimicrob. Chemother. 51, 427–429. doi: 10.1093/jac/dkg080
Bossé, J. T., Li, Y., Walker, S., Atherton, T., Fernandez Crespo, R., et al. (2015). Identification of dfrA14 in two distinct plasmids conferring trimethoprim resistance in Actinobacillus pleuropneumoniae. J. Antimicrob. Chemother. 70, 2217–2222. doi: 10.1093/jac/dkv121
Carattoli, A., Bertini, A., Villa, L., Falbo, V., Hopkins, K. L., and Threlfall, E. J. (2005). Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 63, 219–228. doi: 10.1016/j.mimet.2005.03.018
Carattoli, A., Zankari, E., García-Fernández, A., Voldby Larsen, M., Lund, O., Villa, L., et al. (2014). In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 58, 3895–3903. doi: 10.1128/AAC.02412-14
Cavicchio, L., Dotto, G., Giacomelli, M., Giovanardi, D., Grilli, G., Franciosini, M. P., et al. (2015). Class 1 and class 2 integrons in avian pathogenic Escherichia coli from poultry in Italy. Poult. Sci. 94, 1202–1208. doi: 10.3382/ps/pev095
Clinical Laboratory Standars Institute (2013). Performance Standards for Antimicrobial Disk Susceptibility Testing; Twenty-Second Informational Supplement. m100–s23. Wayne, PA: Clinical and Laboratory Standards Institute.
Connor, T., Barker, C., Baker, K., Weill, F. X., Talukder, K., Smith, A., et al. (2015). Species-wide whole genome sequencing reveals historical global spread and recent local persistence in Shigella flexneri. Elife 4:e07335. doi: 10.7554/eLife.07335
De Lappe, N., O'Halloran, F., Fanning, S., Corbett-Feeney, G., Cheasty, T., and Cormican, M. (2003). Antimicrobial resistance and genetic diversity of Shigella sonnei isolates from Western Ireland, an area of low incidence of infection. J. Clin. Microbiol. 41, 1919–1924. doi: 10.1128/JCM.41.5.1919-1924.2003
Dubois, V., Parizano, M. P., Arpin, C., Coulange, L., Bezian, M.-C., and Quentin, C. (2007). High genetic stability of integrons in clinical isolates of Shigella spp. of worldwide origin. Antimicrob. Agents Chemother. 51, 1333–1340. doi: 10.1128/AAC.01109-06
Fortini, D., Fashae, K., Villa, L., Feudi, C., García-Fernández, A., and Carattoli, A. (2015). A novel plasmid carrying blaCTX-M-15 identified in comensal Escherichia coli from healthy pregnant women in Ibadan, Nigeria. J. Global Antimicrob. Resist. 3, 9–12. doi: 10.1016/j.jgar.2014.12.002
Gassama-Sow, A., Aïdara-Kane, A., Barraud, O., Gatet, M., Denis, F., and Ploy, M. C. (2010). High prevalence of trimethoprim-resistance cassettes in class 1 and 2 integrons in Senegalese Shigella spp isolates. J. Infect. Dev. Ctries. 4, 207–212. doi: 10.3855/jidc.583
Huang, Y., Michael, G. B., Becker, R., Kaspar, H., Mankertz, J., Schwarz, S., et al. (2014). Pheno- and genotypic analysis of antimicrobial resistance properties of Yersinia ruckeri from fish. Vet. Microbiol. 171, 406–412. doi: 10.1016/j.vetmic.2013.10.026
Huovinen, P. (1997). Increases in rates of resistance to trimethoprim. Clin. Infect. Dis. 24, S63–S66. doi: 10.1093/clinids/24.Supplement_1.S63
Huovinen, P. (2001). Resistance to trimethoprim-sulfamethoxazole. Clin. Infect. Dis. 32, 1608–1614. doi: 10.1086/320532
Huovinen, P., Sundström, L., Swedberg, G., and Skold, O. (1995). Trimethoprim and sulfonamide resistance. Antimicrob. Agents Chemother. 39, 279–289. doi: 10.1128/AAC.39.2.279
Inoue, H., Nojima, H., and Okayama, H. (1990). High efficiency transformation of Escherichia coli with plasmids. Gene 96, 23–28. doi: 10.1016/0378-1119(90)90336-P
Instituto de Salud Pública (2009). Informe de Brotes de Diarrea por Shigella sonnei, Diciembre – Mayo del 2009. Ministerio de Salud, Departamento de Epidemiología. Available online at: http://epi.minsal.cl/epi/html/Actualidad/BrotesShigellasonnei.pdf
Iqbal, M. S., Rahman, M., Islam, R., Banik, A., Amin, M. B., Akter, F., et al. (2014). Plasmid-mediated sulfamethoxazole resistance encoded by the sul2 gene in the multidrug-resistant Shigella flexneri 2a isolated from patients with acute diarrhea in Dhaka, Bangladesh. PLoS ONE 9:e85338. doi: 10.1371/journal.pone.0085338
Kadlec, K., and Schwarz, S. (2008). Analysis and distribution of class 1 and class 2 integrons and associated gene cassettes among Escherichia coli isolates from swine, horses, cats and dogs collected in the BfT-GermVet monitoring study. J. Antimicrob. Chemother. 62, 469–473. doi: 10.1093/jac/dkn233
Ke, X., Gu, B., Pan, S., and Tong, M. (2011). Epidemiology and molecular mechanism of integron-mediated antibiotic resistance in Shigella. Arch. Microbiol. 193, 767–774. doi: 10.1007/s00203-011-0744-3
Kehrenberg, C., and Schwarz, S. (2002). Nucleotide sequence and organization of plasmid pMVSCS1 from Mannheimia varigena: identification of a multiresistance gene cluster. J. Antimicrob. Chemother. 49, 383–386. doi: 10.1093/jac/49.2.383
Kehrenberg, C., Tham, N. T. T., and Schwarz, S. (2003). New plasmid-borne antibiotic resistance gene cluster in Pasteurella multocida. Antimicrob. Agents Chemother. 47, 2978–2980. doi: 10.1128/AAC.47.9.2978-2980.2003
Labar, A. S., Millman, J. S., Ruebush, E., Opintan, J. A., Bishar, R. A., Aboderin, A. O., et al. (2012). Regional dissemination of a trimethoprim-resistance gene cassette via a successful transposable element. PLoS ONE 7:e38142. doi: 10.1371/journal.pone.0038142
Liu, C., Zheng, H., Yang, M., Xu, Z., Wang, X., Wei, L., et al. (2015). Genome analysis and in vivo virulence of porcine extraintestinal pathogenic Escherichia coli strain PCN033. BMC Genomics 16:717. doi: 10.1186/s12864-015-1890-9
Lluque, A., Mosquito, S., Gomes, C., Riveros, M., Durand, D., Tilley, D. H., et al. (2015). Virulence factors and mechanisms of antimicrobial resistance in Shigella strains from periurban areas of Lima (Peru). Int. J. Med. Microbiol. 305, 480–490. doi: 10.1016/j.ijmm.2015.04.005
Marcoleta, A., Toro, C., Prado, V., Serrano, M., Fernández, P., Benadof, D., et al. (2013). Antibiotic susceptibility patterns among Shigella sonnei, isolated during three different periods in Región Metropolitana, Chile. Rev. Chilena Infectol. 30, 616–621. doi: 10.4067/S0716-10182013000600007
Merino, L. A., Hreñuk, G. E., Ronconi, M. C., and Alonso, J. M. (2004). Resistencia a antibióticos y epidemiología molecular de Shigella spp. en el nordeste argentino. Rev. Panam. Salud Publica 15, 219–224. doi: 10.1590/S1020-49892004000400001
Ojo, K. K., Kehrenberg, C., Schwarz, S., and Odelola, H. A. (2002). Identification of a complete dfrA14 gene cassette integrated at a secondary site in a resistance plasmid of uropathogenic Escherichia coli from Nigeria. Antimicrob. Agents Chemother. 46, 2054–2055. doi: 10.1128/AAC.46.6.2054-2055.2002
Pan, J. C., Ye, R., Meng, D. M., Zhang, W., Wang, H. Q., and Liu, K. Z. (2006). Molecular characteristics of class 1 and class 2 integrons and their relationships to antibiotic resistance in clinical isolates of Shigella sonnei and Shigella flexneri. J. Antimicrob. Chemother. 58, 288–296. doi: 10.1093/jac/dkl228
Recchia, G. D., and Hall, R. M. (1995). Gene cassettes: a new class of mobile element. Microbiology 141, 3015–3027. doi: 10.1099/13500872-141-12-3015
San Millan, A., Escudero, J. A., Gutierrez, B., Hidalgo, L., García, N., Llagostera, M., et al. (2009). Multiresistance in Pasteurella multocida is mediated by coexistence of small plasmids. Antimicrob. Agents Chemother. 53, 3399–3404. doi: 10.1128/AAC.01522-08
Seol, S. Y., Kim, Y. T., Jeong, Y. S., Oh, J. Y., Kang, H. Y., Moon, D. C., et al. (2006). Molecular characterization of antimicrobial resistance in Shigella sonnei isolates in Korea. J. Med. Microbiol. 55, 871–877. doi: 10.1099/jmm.0.46441-0
Seputiené, V., Povilonis, J., Ruzauskas, M., Pavilonis, A., and Suziedéliené, E. (2010). Prevalence of trimethoprim resistance genes in Escherichia coli isolates of human and animal origin in Lithuania. J. Med. Microbiol. 59, 315–322. doi: 10.1099/jmm.0.015008-0
Seribelli, A. A., Frazão, M. R., Medeiros, M. I., and Falcão, J. P. (2016). Molecular and phenotypic characterization of Shigella sonnei isolated over 31 years suggest the circulation of two prevalent subtypes in the São Paulo State in Brazil. J. Med. Microbiol. doi: 10.1099/jmm.0.000290. [Epub ahead of print].
Shin, H. W., Lim, J., Kim, S., Kim, J., Kwon, G. C., and Koo, S. H. (2015). Characterization of trimethoprim-sulfamethoxazole resistance genes and their relatedness to class 1 integron and insertion sequence common region in Gram-negative bacilli. J. Microbiol. Biotechnol. 25, 137–142. doi: 10.4014/jmb.1409.09041
Sullivan, M. J., Petty, N. K., and Beatson, S. A. (2011). Easyfig: a genome comparison visualizer. Bioinformatics 27, 1009–1010. doi: 10.1093/bioinformatics/btr039
Tonin, P. N., and Grant, R. B. (1987). Genetic and physical characterization of trimethoprim resistance plasmids from Shigella sonnei and Shigella flexneri. Can. J. Microbiol. 33, 905–913. doi: 10.1139/m87-157
Toro, C. S., Farfán, M., Contreras, I., Flores, O., Navarro, N., Mora, G. C., et al. (2005). Genetic analysis of antibiotic-resistance determinants in multidrug-resistant Shigella strains isolated from Chilean children. Epidemiol. Infect. 133, 81–86. doi: 10.1017/S0950268804003048
Vrints, M., Mairiaux, E., Van Meervenne, E., Collard, J. M., and Bertrand, S. (2009). Surveillance of antibiotic susceptibility patterns among Shigella sonnei strains isolated in Belgium during the 18-Year period 1990 to 2007. J. Clin. Microbiol. 47, 1379–1385. doi: 10.1128/JCM.02460-08
Wei, Q., Hu, Q., Li, S., Lu, H., Chen, G., Shen, B., et al. (2014). A novel functional class 2 integron in clinical Proteus mirabilis isolates. J. Antimicrob. Chemother. 69, 973–976. doi: 10.1093/jac/dkt456
White, P. A., and Rawlinson, W. D. (2001). Current status of the aadA and dfr gene cassette families. J. Antimicrob. Chemother. 47, 495–496. doi: 10.1093/jac/47.4.495
Yau, S., Liu, X., Djordjevic, S. P., and Hall, R. M. (2010). RSF1010-like plasmids in Australian Salmonella enterica serovar Typhimurium and origin of their sul2-strA-strB antibiotic resistance gene cluster. Microb. Drug Resist. 16, 249–252. doi: 10.1089/mdr.2010.0033
Ye, C. H., Lan, R., Xia, S., Zhang, J., Sun, Q., Zhang, S., et al. (2010). Emergence of a new multidrug-resistant serotype X variant in an epidemic clone of Shigella flexneri. J. Clin. Microbiol. 48, 419–426. doi: 10.1128/JCM.00614-09
Young, H. K., and Hillyear, J. K. (1994). Trimethoprim resistance in urinary pathogens in northern Scotland: epidemic spread of a resistance plasmid encoding the type I b trimethoprim-resistant dihydrofolate reductase. J. Med. Microbiol. 41, 343–348. doi: 10.1099/00222615-41-5-343
Young, H. K., Qumsieh, M. J., and Mclntosh, M. L. (1994). Nucleotide sequence and genetic analysis of the type I b trimethoprim-resistant, Tn4/32-encoded dihydrofolate reductase. J. Antimicrob. Chemother. 34, 715–725. doi: 10.1093/jac/34.5.715
Keywords: Shigella sonnei, trimethoprim resistance genes, dfrA14, molecular epidemiology, MDR plasmid, antimicrobial resistance mechanisms
Citation: Miranda A, Ávila B, Díaz P, Rivas L, Bravo K, Astudillo J, Bueno C, Ulloa MT, Hermosilla G, Del Canto F, Salazar JC and Toro CS (2016) Emergence of Plasmid-Borne dfrA14 Trimethoprim Resistance Gene in Shigella sonnei. Front. Cell. Infect. Microbiol. 6:77. doi: 10.3389/fcimb.2016.00077
Received: 10 May 2016; Accepted: 05 July 2016;
Published: 20 July 2016.
Edited by:
Nora Lía Padola, Universidad Nacional del Centro de la Provincia de Buenos Aires, ArgentinaReviewed by:
William D. Picking, University of Kansas, USAJosé Alejandro Di Conza, UBA/UNL, Argentina
Copyright © 2016 Miranda, Ávila, Díaz, Rivas, Bravo, Astudillo, Bueno, Ulloa, Hermosilla, Del Canto, Salazar and Toro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cecilia S. Toro, Y2V0b3JvQG1lZC51Y2hpbGUuY2w=
†These authors have contributed equally to this work.