 Haiyong Guo
Haiyong Guo Jeffrey W. Hall
Jeffrey W. Hall Junshu Yang
Junshu Yang Yinduo Ji
Yinduo Ji- 1Department of Biological Science, School of Life Science, Jilin Normal University, Siping, China
- 2Department of Veterinary and Biomedical Sciences, College of Veterinary Medicine, University of Minnesota, St Paul, MN, United States
The SaeRS two-component system plays important roles in regulation of key virulence factors and pathogenicity. In this study, however, we found that the deletion mutation of saeRS enhanced bacterial survival in human blood, whereas complementation of the mutant with SaeRS returned survival to wild-type levels. Moreover, these phenomena were observed in different MRSA genetic background isolates, including HA-MRSA WCUH29, CA-MRSA 923, and MW2. To elucidate which gene(s) regulated by SaeRS contribute to the effect, we conducted a series of complementation studies with selected known SaeRS target genes in trans. We found coagulase complementation abolished the enhanced survival of the SaeRS mutant in human blood. The coa and saeRS deletion mutants exhibited a similar survival phenotype in blood. Intriguingly, heterologous expression of coagulase decreased survival of S. epidermidis in human blood. Further, the addition of recombinant coagulase to blood significantly decreased the survival of S. aureus. Further, analysis revealed staphylococcal resistance to killing by hydrogen peroxide was partially dependent on the presence or absence of coagulase. Furthermore, complementation with coagulase, but not SaeRS, returned saeRS/coa double mutant survival in blood to wild-type levels. These data indicate SaeRS modulates bacterial survival in blood in coagulase-dependent manner. Our results provide new insights into the role of staphylococcal SaeRS and coagulase on bacterial survival in human blood.
Introduction
Staphylococcus aureus is an important pathogen that can cause various infections, including skin and soft tissue infection and systematic infections such as pneumoniae, endocarditis, and toxic shock syndrome (Klevens et al., 2007; Gordon and Lowy, 2008). The pathogenicity of S. aureus is attributable to its ability to produces different virulence factors, including a series of cell wall-associated proteins and a range of extracellular cytotoxins, proteases, DNases, and enterotoxins (Foster and Höök, 1998), which enable the bacteria to evade innate and or adaptive immune systems (Liu et al., 2005; Clauditz et al., 2006; Lacey et al., 2016) and induce disease.
Two-component signal regulatory systems (TCSs) collaborate with transcriptional regulators to regulate the expression of virulence factors, which in turn contribute to the pathogenesis of S. aureus. The well-studied global and TCSs virulence regulators include Agr (Novick, 2003; Montgomery et al., 2010; Thoendel and Horswill, 2013), ArlRS (Fournier et al., 2001; Bronner et al., 2004; Liang et al., 2005), SaeRS (Giraudo et al., 1999; Liang et al., 2006; Voyich et al., 2009; Nygaard et al., 2010; Zurek et al., 2014; Liu et al., 2016), SarA (Chien et al., 1999; Cheung et al., 2008; Li et al., 2016; Loughran et al., 2016), and MgrA (Ballal et al., 2009; Gupta et al., 2015).
The SaeRS TCS controls the expression of critical virulence genes. SaeRS up-regulates the transcription and expression of hla, hlb, hlgABC, lukED, and coa in vitro (Giraudo et al., 1999; Liang et al., 2006; Rogasch et al., 2006; Nygaard et al., 2010), as well as controls hla expression in vivo, as the sae null mutation significantly decreased the expression of α-toxin (hla) during infection (Goerke et al., 2001, 2005). The mutation of sae eliminated the expression of fnbA, but increased the expression of CP5 in S. aureus strain Newman (Steinhuber et al., 2003). As a consequence of SaeRS signaling pathway disruption, the cytotoxicity and ability of S. aureus to adhere to and invade epithelial cells (Liang et al., 2006) and endothelial cells (Steinhuber et al., 2003) is impaired. In addition, it has been demonstrated that the SaeRS system is an important virulence regulator in various animal models of infection (Goerke et al., 2005; Liang et al., 2006; Voyich et al., 2009; Montgomery et al., 2010; Cho et al., 2015; Zhao et al., 2015).
Staphylococcal coagulase is directly regulated by SaeRS (Liu et al., 2016) and is an important factor for distinguishing S. aureus and coagulase negative staphylococcus. Coagulase converts host prothrombin to staphylothrombin, leading to activate the protease activity of thrombin. It was predicted that coagulase could protect bacteria from phagocytic and immune defenses by causing localized clotting, however, there are contradictory reports regarding the role of coagulase in pathogenicity in animal models of infection (Baddour et al., 1994; Moreillon et al., 1995; Cheng et al., 2010).
In this study, we aimed to determine the role of SaeRS in survival capacity of S. aureus in human blood. We utilized two published SaeRS mutant strains, including hospital-acquired methicillin resistant S. aureus (HA-MRSA) WCUH29 and USA400 community-associated (CA)-MRSA MW2 isolates, generated a saeRS deletion mutant in a USA300 CA-MRSA 923 human isolate, and examined the impact of the saeRS mutation on bacterial survival in the blood. Using targeted complementation studies, gene deletions, and purification of recombinant proteins, we determined the SaeRS regulated coa gene, encoding coagulase, was a mediator of enhanced saeRS mutant survival in human blood. Hence, modulation of coa expression by SaeRS may contribute to S. aureus survival in human blood and bacteremia.
Materials and Methods
Bacterial Strains, Plasmids, and Growth Media
The bacterial strains and plasmids used in this study are listed in Table 1. Escherichia coli DC10B (gift of T.J. Foster) served as the host for all in vitro recombinant DNA (Monk et al., 2012). E. coli transformants were selected on Brain Heart Infusion (BHI; Difco) agar containing erythromycin (100 μg/ml) or Luria-Bertani agar containing ampicillin (100 μg/ml). S. aureus was cultured in Trypticase Soy Broth (TSB; Difco) or on TSA agar at 37°C with appropriate antibiotics. All bacterial cell cultures were incubated with shaking at 220 RPM. S. aureus transformants were selected on TSA containing chloramphenicol (10 μg/ml) or erythromycin (5 μg/ml).

Table 1. Bacterial strains and plasmids used in this study.
Construction of the saeRS and coa Gene Deletion Mutants, and the saeRS and coa Gene Complemented Strains
Deletion of saeRS and/or coa was carried out following the pKOR1 allelic exchange protocol as described (Sun et al., 2005; Bae and Schneewind, 2006) and primers sets saeRS-pKOR1 L–For/Rev and coa-pKOR1-R For/Rev listed in Table 2. The R-For primer was synthesized with a 5′ phosphate group. Each PCR fragment was purified and the two fragments were ligated together with T4 DNA ligase (Promega). The ligation product was mixed with BP Clonase, per manufacturer's instructions, and plasmid pKOR1, incubated at 25°C overnight, then transformed into E. coli DC10B. The pKOR1-saeRSKO or pKOR1-coaKO plasmid was subsequently transformed into S. aureus 923, WCUH29, or MW2, respectively. Big colonies were re-streaked to fresh TSA plates and deletion of saeRS and/or coa was confirmed by diagnostic PCR.

Table 2. Oligonucleotides used in this study.
In order to examine whether the expression of saeRS, coa in trans can complement the effect of the mutation of the respective endogenous gene, we constructed recombinant plasmids, including pYH4/saeRS, pYH4/sa1000, pYH4/efb, pYH4/coa by cloning the saeRS, sa1000, efb, fnbAB, or coa coding region (which was obtained by PCR) into the AscI and PmeI sites of pYH4 (Huang et al., 2004), and electroporated it into the saeRS and/or coa knockout mutant, respectively, resulting in Sa371com, Sa371/pYH4-sa1000, Sa371/pYH4-efb, Sa371/pYH4-fnbAB, Sa371/pYH4-coa, and coa complementation strains in Table 1. The recombinant plasmid DNA were isolated from the complementary strains and confirmed by PCR and DNA sequencing (data not shown).
Cloning, Expression, and Purification of Coagulase-His Tagged Fusion Protein in Escherichia Coli
The coa gene was obtained by PCR amplification using the primers listed in Table 2, cloned into pET24b, and resulted in pET24b-coa. Plasmid pET24b-coa was introduced into a BL21(DE3) strain. The resulting strain was grown in LB medium at room temperature; the expression of coagulase was induced when the culture media reached OD600 nm equal to 0.6 by addition of 1 mM IPTG (isopropyl-b-D-thiogalactoside) and incubation pursued for 4 h. The coagulase-his tagged protein was purified using Ni-NTA agarose column (Novagen) and examined using 12% SDS-PAGE and Coomassie Blue staining as described (Yang et al., 2015). The purification of SarZ-6 × His was carried out as described using the previously constructed pET24b-sarZ plasmid (Liang et al., 2011).
Blood Survival Assay
Strains were cultured in TSB with appropriate antibiotics. Inducer anhydrotetracycline (ATc) was added when indicated to overnight cultures. Following 18 h of culturing, the bacteria were washed twice in sterile PBS and suspended to an OD of 0.14 using a Behring photometer in PBS. Fresh venous human whole blood was collected using heparin containing Vacutainer tubes (BD) from outwardly healthy adult donors. The blood was then immediately used in the assay as described (Liu et al., 2005; Hall et al., 2015, 2017). The percentage of surviving bacteria was calculated as (CFUtimepoint/CFUinitialinput)*100. Human blood collection was approved by the University of Minnesota Institutional Review Board.
Hydrogen Peroxide Survival Assay
To determine the contribution of coagulase to S. aureus survival when challenged with oxidative stress, overnight cultures were washed with PBS and ~2 × 108 CFU were incubated at 37°C in a 1.5% hydrogen peroxide (H2O2)/PBS solution for 60 min (Liu et al., 2005). Serial dilutions were plated on TSA for enumeration of surviving CFU. Percent survival was calculated as surviving CFU/ input CFU multiplied by 100, (#CFUf/#CFUi)*100.
Data Analysis
Independent samples were statistically analyzed using a Student's t-test with an alpha level ≤ 0.05 considered significant. For data figures with more than two independent samples, a one-way ANOVA analysis with a post-hoc Tukey HSD-test was used to determine if there was statistical significance between samples with an alpha level ≤ 0.05 considered significant.
Results
The Deletion Mutation of saeRS Enhanced S. aureus Survival in Human Blood
Our previous studies have demonstrated that the AirSR(YhcSR) system contributes S. aureus survival in human blood (Hall et al., 2015, 2017); and our preliminary study found that AirSR probably positively regulates the transcription of saeRS (data not shown). It has been well-documented that the SaeRS system is a key regulator of virulence factors that contribute to evade innate immune system and pathogenicity in a variety of animal models of S. aureus infection (Steinhuber et al., 2003; Liang et al., 2006; Zurek et al., 2014; Cho et al., 2015). These data led us to speculate that SaeRS system is likely involved in the function of AirSR in survival in the blood. To test this possibility, we deleted saeS and examined the impact of the deletion on bacterial survival in human blood. Unexpectedly, we found that the null mutation of saeS in HA-MRSA WCUH29 significantly increased survival in the blood throughout the 3 h experiment (Figure 1A).
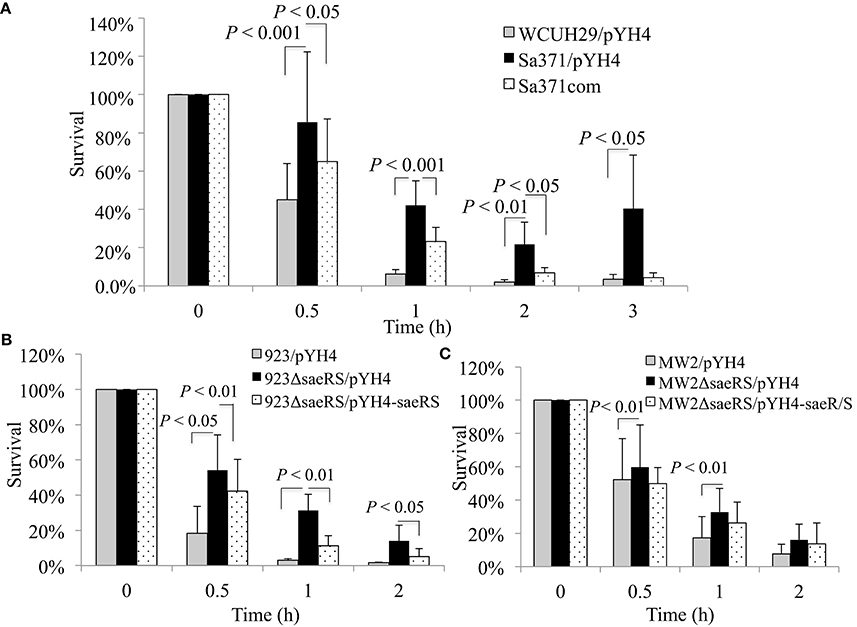
Figure 1. Effect of saeS or saeRS null mutation and complementation on survival of S. aureus in human blood. Percent survival of wild-type S. aureus HA-MRSA WCUH29 (A), USA300 CA-MRSA 923 (B), USA400 CA-MRSA MW2 (C), and saeRS null mutants in freshly collected heparinized human blood with appropriate antibiotics. Bacteria were cultured overnight, diluted, inoculated into blood, and incubated at 37°C in a rotisserie incubator. Percent survival = (#CFUfinal/#CFUinput)*100. The data represents the mean ± SEM of at least six independent experiments.
To determine whether the genetic backgrounds of S. aureus affect SaeRS's mediated bacterial survival in the blood, we created a saeRS deletion mutant in a USA300 CA-MRSA 923 strain and acquired a saeRS allelic replacement mutant of USA400 CA-MRSA MW2 (Voyich et al., 2009). Consistent with the results of WCUH29, the saeRS null mutation remarkably increased the percentage of CFUs that survived in the blood for all strains (Figures 1B,C). Moreover, in trans SaeRS complemented strains had similar survival percentages as the wild-type controls (Figures 1A,B), indicating a specific effect of SaeRS on bacterial survival in human blood.
The Introduction of Constitutive Coagulase Expression System Eliminated the Effect of the saeRS Null Mutation on Bacterial Survival in Human Blood
Previous studies revealed that the SaeRS system is a critical positive regulator of important virulence factors, including fibronectin-binding proteins (fnbB, fnb), fibrinogen-binding proteins (efb), coagulase (coa), and toxins (hla, hlb; Giraudo et al., 1999; Liang et al., 2006; Sun et al., 2010). We hypothesized the deletion of SaeRS prevented the expression of virulence factor with the result being increased bacterial survival in blood. In order to identify which SaeRS regulated gene(s) are involved in this phenomena, we tested the impact of fnbAB, efb, sa1000, or coa overexpression in trans on bacterial survival in blood in the saeRS mutants. The constitutive expression of coagulase restored the percentage of bacteria that survived to wild-type control levels and decreased the survival capacity of S. aureus compared to the saeRS null mutant control (Figures 2A–C). In contrast, the overexpression of FnbAB, Efb, or SA1000 did not eliminate the enhanced survival phenomena of the saeRS knockout mutant (Figure 2A).
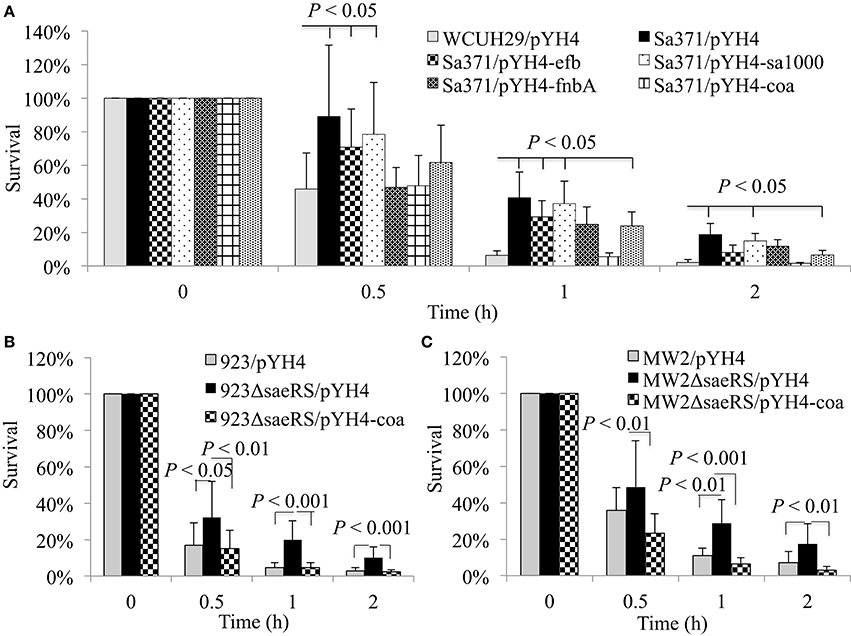
Figure 2. Impact of sa1000, efb, fnbAB, or coa expression in trans on enhanced survival of saeRS null mutants. Percent survival of HA-MRSA WCUH29 (A), USA300 CA-MRSA 923 (B), USA400 CA-MRSA MW2 (C), and the sa1000, efb, fnbAB, or coa expression strain, and saeRS complementation strain in freshly collected heparinized human blood with appropriate antibiotics. Data is the mean and standard error of at least six experiments per strain.
The Deletion Mutation of coa Increased the Survival of S. aureus in Human Blood
To confirm the role of coagulase in SaeRS's involvement in bacterial survival in the blood, we created coa deletion mutants in WCUH29, 923, and MW2 strains. Each new coa mutant was complemented and we tested each strain's ability to coagulate rabbit plasma. The coa deletion mutants of 923 and MW2 strains exhibited negative coagulation, whereas the coa deletion mutant of WCUH29 strain formed partial coagulation (Table 3). The 923 and MW2 wild type, coa null, and complemented strains were examined in our survival assays using human blood. Similar to the results of saeRS null mutants, the deletion of coa enhanced the survival capacity of S. aureus 923 and MW2 after 1 h of infection (Figures 3A,B). The in trans coa complementation restored the survival level to the wild-type control (Figures 3A,B), indicating coagulase production in blood is detrimental to the survival of S. aureus.
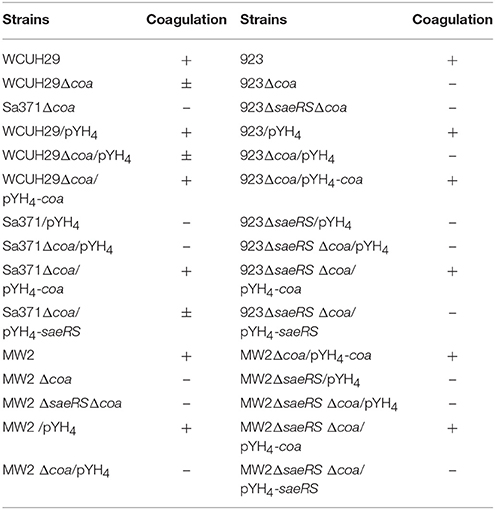
Table 3. Coagulase activity of wild type strains, saeRS or coa null mutants, and saeRS or coa complemented strains.
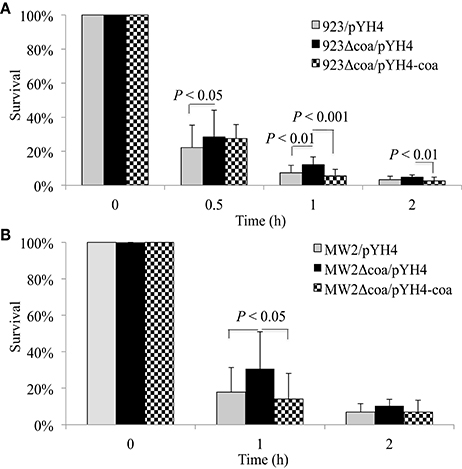
Figure 3. Effect of coa deletion mutation and complementation on survival of S. aureus in human blood. Percent survival of wild-type S. aureus USA300 CA-MRSA 923 (A), USA400 CA-MRSA MW2 (B), and coa deletion mutants and its complementation in freshly collected heparinized human blood with appropriate antibiotics. Bacteria were cultured overnight, diluted, inoculated into blood, and incubated at 37°C in a rotisserie incubator. Percent survival = (#CFUfinal/#CFUinput)*100. The data represents the mean ± SEM of at least six independent experiments.
The Heterologous Expression of Coagulase or Addition of Recombinant Coagulase Inhibited Survival Ability in Human Blood
To further confirm the role of coagulase in survival, we determined the effect of heterologous expressing coagulase in S. epidermidis on survival in the blood. Compared with the control, the induction of coa expression significantly decreased the survival of S. epidermidis in the blood throughout the duration of the experiment (Figures 4A,B). To define the role of coagulase in survival of S. aureus, we cloned, expressed, and purified recombinant coagulase (rCoa). The purity of purified rCoa was examined using SDS-PAGE (Figure 4C); the activity of purified rCoa was confirmed in a coagulation assay using human blood (Figure 4D). The addition of rCoa significantly decreased the survival ability of USA300 CA-MRSA 923 in human blood throughout the period of the experiment in a dose-dependent manner (Figure 4E). In contrast, the addition of control protein, recombinant SarZ, had no impact on survival compared to the negative control (Figure 4E).
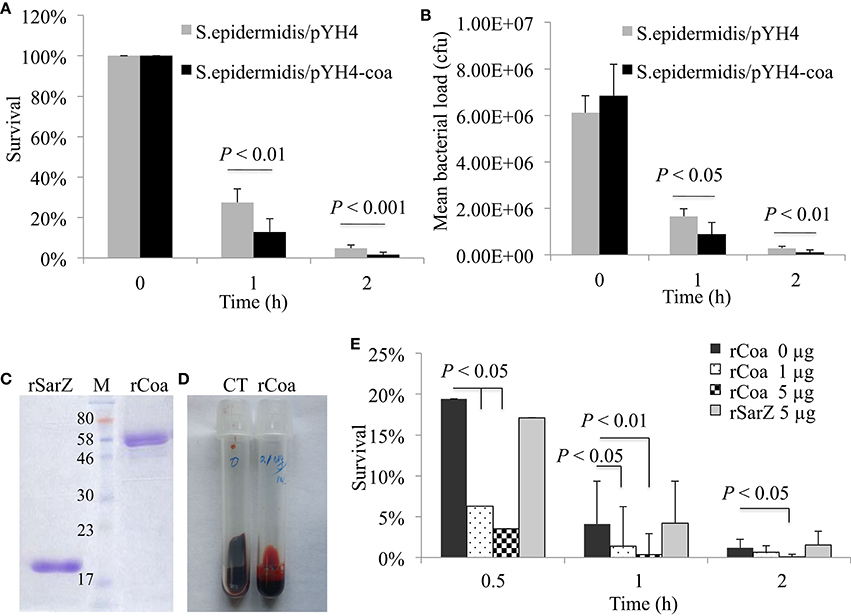
Figure 4. Effect of coa heteroexpression or addition of recombinant coagulase on bacterial survival. (A) Percent survival and (B) bacterial load of wild-type S. epidermidis and coa expression in trans in human blood during induction. (C) SDS-PAGE analysis of purified recombinant Coa (coagulase) and control protein SarZ from E. coli. M, ColorPlus Prestained protein Marker (NEB), the unit of size is kDa. (D) Coagulation activity of purified recombinant Coa (rCoa) from E. coli in human blood. CT, negative control without addition of rCoa. (E) Percent survival of wild-type USA300 CA-MRSA 923 and addition of different concentrations of purified rCoa and control rSarZ protein in human blood. Data is the mean and standard error of at least four experiments per strain.
Coagulase Is Associated with Susceptibility to Hydrogen Peroxide Killing
Coagulase is able to bind host prothrombin to form staphylothrombin, which in turn activates the protease activity of thrombin. Although it has been predicted that coagulase could protect bacteria from phagocytic and immune defenses by causing localized clotting, the role of coagulase in pathogenicity is contradictory (Baddour et al., 1994; Moreillon et al., 1995; Cheng et al., 2010). Reactive oxygen species (ROS) is a key element used by phagocytic cells to kill phagocytosed bacteria (Liu et al., 2005; Clauditz et al., 2006). To explore the potential mechanism of coagulase in survival, we examined whether the deletion mutation of coa alters the susceptibility to the ROS H2O2. The deletion mutation of coa increased the bacterial survival compared to the 923/pYH4 control strain (68 vs. 53%, Figure 5A), whereas the episomally coa complemented strain had significantly decreased survival compared to the coa deletion mutant (68 vs. 51%). Similarly, heterologous expression of coagulase reduced the ability of coagulase negative S. epidermidis to tolerate H2O2—mediated killing compared to the S. epidermidis/pYH4 control (74 vs. 93%, Figure 5B).
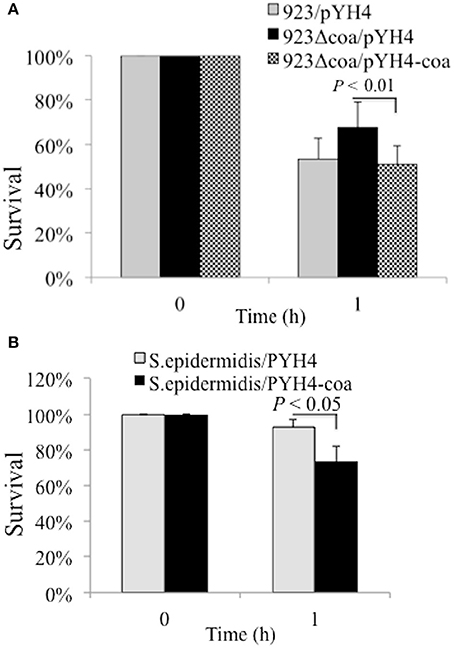
Figure 5. Effect of Coa on bacterial resistance to H2O2. (A) Percent survival of wild-type USA300 CA-MRSA 923, coa deletion mutant and complementation strain after exposure of H2O2. (B) Percent survival of wild-type S. epidermiditis and coa expression strain in trans after exposure of H2O2. The bacterial strains were cultured with appropriate antibiotics. Approximately 5 × 108 CFU were incubated in sterile PBS with 1.5% hydrogen peroxide at 37°C for 1 h. The percent survival was calculated as (#CFUfinal/#CFUinput)*100. The data represents the mean ± SEM of at least four independent experiments.
The Involvement of SaeRS in Survival of S. aureus in Human Blood Is Coagulase Dependent
The above studies demonstrated both SaeRS and coagulase are involved in controlling S. aureus survival in human blood. To further understand whether the role of SaeRS is attributable to its positive regulation of coa, we created saeRS/coa double mutants in 923 and MW2 strains and examined the effect of expression of either coa or saeRS in trans on survival of the double mutant in blood. Similar to the results of coa or saeRS single gene null mutant, the saeRS/coa double null mutation in 923 resulted in a significant increase of bacterial survival in the blood (Figure 6). The expression of coagulase in trans complemented the survival ability of the saeRS/coa double mutants to their wild-type control level, whereas the expression of SaeRS in trans had no impact on the survival ability of the saeRS/coa double mutants (Figure 6). Similar results of saeRS/coa double mutant of MW2 were observed (data not shown).
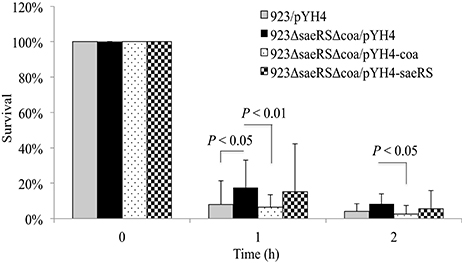
Figure 6. Effect of saeRS and coa double mutation, and complementation on survival of S. aureus in blood. Survival of wild-type USA300 CA-MRSA 923, saeRS/coa double deletion mutant, and coa or saeRS complementation strain in human blood. Data is the mean and standard error of at least four experiments per strain.
Discussion
Our results clearly indicate the roles of SaeRS and coagulase in S. aureus survival in human blood. Despite the fact that SaeRS plays a critical role in bacterial invasion of host cells, cytotoxicity, and pathogenicity of S. aureus in animal models of infection (Liang et al., 2006; Voyich et al., 2009), it did not appear to enhance survival in human blood. Conversely, SaeRS inhibited the survival of S. aureus in human blood, as demonstrated by experiments with the saeRS null mutants and by complementation studies. Surprisingly, the expression of coa in trans could eliminate the enhanced survival of the SaeRS null mutant in blood, which was demonstrated with coa complementation studies in the saeRS single and saeRS/coa double mutants. Consistently, coagulase decreases the bacterial survival in the blood, as supported by experiments with the coa null mutants, coa heterologous expression strains, recombinant coagulase, as well as by complementation experiments. This finding is at odds with the assumption coagulase could protect bacteria from immune defenses by causing localized clotting. Conversely, our data suggest localized clotting mediated by coagulase may be detrimental to the survival of S. aureus in blood. Interestingly, our data indicate that coagulase is somehow associated with S. aureus susceptibility to hydrogen peroxide killing.
Our findings in this study only pinpoint the role of SaeRS in control of S. aureus survival in human blood through regulation of coagulase production. This suggests that SaeRS may play different roles in different stages of host cell-pathogen interactions. Several key steps of infection include bacterial colonization, proliferation, evasion of innate defenses, and cytotoxicity (Ji et al., 1999; Okumura and Nizet, 2014; Janhsen et al., 2016; McGuinness et al., 2016). It was clearly demonstrated that SaeRS is critical for S. aureus to adhere to and invade human epithelial and endothelial cells through regulation of adhesins and invasins (Giraudo et al., 1999; Liang et al., 2006; Sun et al., 2010). Fibronectin-binding proteins (FnBPs) are one kind of major adhesins and invasins, which enable to assemble Fn to connect the host and bacterial cells via binding its receptor α5β1-integrin (Sinha et al., 1999; Dziewanowska et al., 2000; Fowler et al., 2000). Moreover, SaeRS is crucial for S. aureus to mediate the expression of toxins, which are required for cytotoxicity in vitro cell culture (Liang et al., 2006; Liang and Ji, 2007) and pathogenicity in different models of infection (Kernodle et al., 1997; Ji et al., 1999; Schwan et al., 2003).
Our results indicate that the SaeRS system controls the capacity of S. aureus survival in human blood, which contradicts to previous report that the saeRS mutation decreased survival of MW2 strain in human blood (Voyich et al., 2009). To rule out possible effect of genetic background of different S. aureus isolates, we utilized the saeRS null mutant and its parental control MW2 (which are kindly provided by Dr. Voyich) and conducted survival assays. Consistent with the results of the SaeRS mutants of WCUH29 and 923 isolates, the saeRS mutant of MW2 exhibited the similar elevated survival capacity compared to its wild-type control. This contradiction, we believe, is mainly due to the different survival ability of wild type control MW2 in the blood samples. In our studies, <10% of the MW2 cells were able to survive in human blood after 1–3 h; in contrast more than 100% of MW2 cells survived after 3 h of incubation in human blood in the previous report (Voyich et al., 2009). This inconsistence is possibly attributable to the differences in the human blood, as different individuals likely possess different levels of antibodies against S. aureus, which would affect the opsonophagocytic killing process (Pier and Elcock, 1984; Chen et al., 2013; Humphries et al., 2015). Another explanation is that in our experiments S. aureus from the stationary phase were utilized, while Voyich et al used the mid-exponential phase of bacterial cells. It is possible that the temporal regulation of coa by Agr (Novick, 2003) and other regulators that are expressed at different temporal periods may influence the results as the mid-exponential cells grow faster than stationary cells. In addition, it is necessary to further elucidate the influence of antibodies against S. aureus in order to define the role of SaeRS in bacterial survival in the blood in future studies.
Regarding coagulase, our results clearly showed that the addition of rCoa to the blood enhanced the susceptibility of S. aureus to phagocytic killing. Moreover, altering the expression of coagulase affects bacterial sensitivity to hydrogen peroxide killing. It is necessary to further determine the mechanism in future studies. Our finding is supported with previous reports demonstrating that coagulase is not a virulence factor in several infection models as the coa null mutation had no impact on pathogenicity in a rat endocarditis model of infection (Baddour et al., 1994; Moreillon et al., 1995). Moreover, coagulase negative staphylococci (CNS) have become a major nosocomial pathogen (Becker et al., 2014). Additionally, CNS infections cause renal abscesses and lethality in a mouse model of blood stream infection (Wang et al., 2015). On the other hand, our results are inconsistent with recent report that indicated coagulase is a critical virulence factor for abscess formation and bacteremia in a mouse model of blood stream infection (Cheng et al., 2010). This inconsistence is probably due to the difference of animal models of infection and the variation of genetic background for S. aureus isolates in these studies. Moreover, in this study we found that unlike 923 and MW2 isolates, the deletion mutation of coa could not abolish the coagulation activity of WCUH29, suggesting that other factors, such as von Willebrand factor binding protein, are likely involved in coagulation activity as reported (Thomer et al., 2013).
In conclusion, we identified that the SaeRS two-component system plays an important role in survival of S. aureus in human blood via regulation of coagulase production. Coagulase alleviates the survival capacity of S. aureus in human blood. These data suggest that coagulase might be used as an alternative strategy to treat S. aureus-induced bacteremia.
Ethics Statement
This study was carried out in accordance with the recommendations of NIH guidelines with written informed consent from all subjects. All subjects gave written informed consent in accordance with the Declaration of Helsinki. The protocol was approved by the University of Minnesota Institutional Review Board.
Author Contributions
HG, JH, and JY performed the experiments. HG, JH, and YJ designed the experiments. HG, JH, JY, and YJ analyzed the data. HG, JH, and YJ wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Voyich for kindly providing MW2 and MW2/saeRS knockout mutant strains, and Michelle M. Ji for critical reading and editing the manuscript. This study was partially supported by China Scholarship Council No. 201308220113 (HG) and the National Science Foundation of Jilin Province No. 20170101027JC (HG), by a grant AI078951 from the National Institutes of Health (YJ), and by a grant from the USDA/Minnesota agriculture station.
References
Baddour, L. M., Tayidi, M. M., Walker, E., McDevitt, D., and Foster, T. J. (1994). Virulence of coagulase-deficient mutants of Staphylococcus aureus in experimental endocarditis. J. Med. Microbiol. 41, 259–263 doi: 10.1099/00222615-41-4-259
Bae, T., and Schneewind, O. (2006). Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid 55, 58–63. doi: 10.1016/j.plasmid.2005.05.005
Ballal, A., Ray, B., and Manna, A. C. (2009). sarZ, a sarA family gene, is transcriptionally activated by MgrA and is involved in the regulation of genes encoding exoproteins in Staphylococcus aureus. J. Bacteriol. 191, 1656–1665. doi: 10.1128/JB.01555-08
Becker, K., Heilmann, C., and Peters, G. (2014). Coagulase-negative staphylococci. Clin. Microbiol. Rev. 27, 870–926. doi: 10.1128/CMR.00109-13
Bronner, S., Monteil, H., and Prévost, G. (2004). Regulation of virulence determinants in Staphylococcus aureus: complexity and applications. FEMS Microbiol. Rev. 28, 183–200. doi: 10.1016/j.femsre.2003.09.003
Chen, Q., Dintaman, J., Lees, A., Sen, G., Schwartz, D., Shirtliff, M. E., et al. (2013). Novel synthetic (poly)glycerolphosphate-based antistaphylococcal conjugate vaccine. Infect. Immun. 81, 2554–2561. doi: 10.1128/IAI.00271-13
Cheng, A. G., McAdow, M., Kim, H. K., Bae, T., Missiakas, D. M., and Schneewind, O. (2010). Contribution of coagulases towards Staphylococcus aureus disease and protective immunity. PLoS Pathog. 6:e1001036. doi: 10.1371/journal.ppat.1001036
Cheung, A. L., Nishina, K. A., Trotonda, M. P., and Tamber, S. (2008). The SarA protein family of Staphylococcus aureus. Int. J. Biochem. Cell Biol. 40, 355–361. doi: 10.1016/j.biocel.2007.10.032
Chien, Y., Manna, A. C., Projan, S. J., and Cheung, A. L. (1999). SarA, a global regulator of virulence determinants in Staphylococcus aureus, binds to a conserved motif essential for sar-dependent gene regulation. J. Biol. Chem. 274, 37169–37176. doi: 10.1074/jbc.274.52.37169
Cho, H., Jeong, D. W., Liu, Q., Yeo, W. S., Vogl, T., Skaar, E. P., et al. (2015). Calprotectin increases the activity of the SaeRS two component system and murine mortality during Staphylococcus aureus infections. PLoS Pathog. 11:e1005026. doi: 10.1371/journal.ppat.1005026
Clauditz, A., Resch, A., Wieland, K.-P., Peschel, A., and Gotz, F. (2006). Staphyloxanthin plays a role in the fitness of Staphylococcus aureus and its ability to cope with oxidative stress. Infect. Immun. 74, 4950–4953. doi: 10.1128/IAI.00204-06
Dziewanowska, K., Carson, A. R., Patti, J. M., Deobald, C. F., Bayles, K. W., and Bohach, G. A. (2000). Fibronectin binding protein and host cell tyrosine kinase are required for internalization of Staphylococcus aureus by epithelial cells. Infect. Immun. 68, 6321–4678. doi: 10.1128/IAI.68.11.6321-6328.2000
Foster, T. J., and Höök, M. (1998). Surface protein adhesins of Staphylococcus aureus. Trends Microbiol. 6, 484–488. doi: 10.1016/S0966-842X(98)01400-0
Fournier, B., Klier, A., and Rapoport, G. (2001). The two-component system ArlS–ArlR is a regulator of virulence gene expression in Staphylococcus aureus. Mol. Microbiol. 41, 247–261. doi: 10.1046/j.1365-2958.2001.02515.x
Fowler, T., Wann, E. R., Joh, D., Johansson, S., Foster, T. J., and Höök, M. (2000). Cellular invasion by Staphylococcus aureus involves a fibronectin bridge between the bacterial fibronectin-binding MSCRAMMs and host cell β1 integrins. Eur. J. Cell Biol. 79, 672–679. doi: 10.1078/0171-9335-00104
Giraudo, A. T., Calzolari, A., Cataldi, A. A., Bogni, C., and Nagel, R. (1999). The sae locus of Staphylococcus aureus encodes a two-component regulatory system. FEMS Microbiol. Lett. 177, 15–22. doi: 10.1111/j.1574-6968.1999.tb13707.x
Goerke, C., Fluckiger, U., Steinhuber, A., Bisanzio, V., Ulrich, M., Bischoff, M., et al. (2005). The role of Staphylococcus aureus global regulators sae and σB in virulence gene expression during device-related infection. Infect. Immun. 73, 3415–3421. doi: 10.1128/IAI.73.6.3415-3421.2005
Goerke, C., Fluckiger, U., Steinhuber, A., Zimmerli, W., and Wolz, C. (2001). Impact of the regulatory loci agr, sarA and sae of Staphylococcus aureus on the induction of α-toxin during device-related infection resolved by direct quantitative transcript analysis. Mol. Microbiol. 40, 1439–1447. doi: 10.1046/j.1365-2958.2001.02494.x
Gordon, R. J., and Lowy, F. D. (2008). Pathogenesis of methicillin-resistant Staphylococcus aureus infection. Clin. Infect. Dis. 46, S350–S359. doi: 10.1086/533591
Gupta, R. K., Luong, T. T., and Lee, C. Y. (2015). RNAIII of the Staphylococcus aureus agr system activates global regulator MgrA by stabilizing mRNA. Proc. Natl. Acad. Sci. U.S.A. 112, 14036–14041. doi: 10.1073/pnas.1509251112
Hall, J. W., Yang, J., Guo, H., and Ji, Y. (2015). The AirSR two-component system contributes to Staphylococcus aureus survival in human blood and transcriptionally regulates sspABC operon. Front. Microbiol. 6:682. doi: 10.3389/fmicb.2015.00682
Hall, J. W., Yang, J., Guo, H., and Ji, Y. (2017). The Staphylococcus aureus AirSR two-component System mediates reactive oxygen species resistance via transcriptional regulation of staphyloxanthin production. Infect Immun. 85:e00838-16. doi: 10.1128/IAI.00838-16
Herold, B. C., Immergluck, L. C., Maranan, M. C., Lauderdale, D. S., Gaskin, R. E., Boyle-Vavra, S., et al. (1998). Community-acquired methicillin-resistant Staphylococcus aureus in children with no identified predisposing risk. JAMA 279, 593–598. doi: 10.1001/jama.279.8.593
Huang, J., O'Toole, P. W., Shen, W., Amrine-Madsen, H., Jiang, X., Lobo, N., et al. (2004). Novel chromosomally encoded multidrug efflux transporter MdeA in Staphylococcus aureus. Antimicrob. Agents Chemother. 48, 909–917. doi: 10.1128/AAC.48.3.909-917.2004
Humphries, H. E., Brookes, C., Allen, L., Kuisma, E., Gorringe, A., and Taylor, S. (2015). Seroprevalence of antibody-mediated, complement-dependent opsonophagocytic activity against Neisseria meningitidis serogroup B in England. Clin. Vaccine Immunol. 22, 503–509. doi: 10.1128/CVI.00100-15
Janhsen, W. K., Arnold, C., Hentschel, J., Lehmann, T., Pfister, W., Baier, M., et al. (2016). Colonization of CF patients' upper airways with S. aureus contributes more decisively to upper airway inflammation than P. aeruginosa. Med. Microbiol. Immunol. 205, 485–500. doi: 10.1007/s00430-016-0463-0
Ji, Y., Marra, A., Rosenberg, M., and Woodnutt, G. (1999). Regulated antisense RNA eliminates alpha-toxin virulence in Staphylococcus aureus infection. J. Bacteriol. 181, 6585–6590.
Kernodle, D. S., Voladri, R. K., Menzies, B. E., Hager, C. C., and Edwards, K. M. (1997). Expression of an antisense hla fragment in Staphylococcus aureus reduces alpha-toxin production in vitro and attenuates lethal activity in a murine model. Infect. Immun. 65, 179–184.
Klevens, R. M., Morrison, M. A., Nadle, J., Petit, S., Gershman, K., Ray, S., et al. (2007). Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 298, 1763–1771. doi: 10.1001/jama.298.15.1763
Lacey, K. A., Geoghegan, J. A., and McLoughlin, R. M. (2016). The role of Staphylococcus aureus virulence factors in skin infection and their potential as vaccine antigens. Pathogens 5:22. doi: 10.3390/pathogens5010022
Li, L., Cheung, A., Bayer, A. S., Chen, L., Abdelhady, W., Kreiswirth, B. N., et al. (2016). The global regulon sarA regulates β-Lactam antibiotic resistance in methicillin-resistant Staphylococcus aureus in vitro and in endovascular infections. J. Infect. Dis. 214, 1421–1429. doi: 10.1093/infdis/jiw386
Liang, X., Hall, J. W., Yang, J., Yan, M., Doll, K., Bey, R., et al. (2011). Identification of single nucleotide polymorphisms associated with hyperproduction of alpha-toxin in Staphylococcus aureus. PLoS ONE 6:e18428. doi: 10.1371/journal.pone.0018428
Liang, X., and Ji, Y. (2007). Involvement of α5β1-integrin and TNF-α in Staphylococcus aureus alpha-toxin-induced death of epithelial cells. Cell. Microbiol. 9, 1809–1821. doi: 10.1111/j.1462-5822.2007.00917.x
Liang, X., Yu, C., Sun, J., Liu, H., Landwehr, C., Holmes, D., et al. (2006). Inactivation of a two-component signal transduction system, SaeRS, eliminates adherence and attenuates virulence of Staphylococcus aureus. Infect. Immun. 74, 4655–4665. doi: 10.1128/IAI.00322-06
Liang, X., Zheng, L., Landwehr, C., Lunsford, D., Holmes, D., and Ji, Y. (2005). Global regulation of gene expression by ArlRS, a two-component signal transduction regulatory system of Staphylococcus aureus. J. Bacteriol. 187, 5486–5492. doi: 10.1128/JB.187.15.5486-5492.2005
Liu, G. Y., Essex, A., Buchanan, J. T., Datta, V., Hoffman, H. M., Bastian, J. F., et al. (2005). Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. J. Exp. Med. 202, 209–215. doi: 10.1084/jem.20050846
Liu, Q., Yeo, W. S., and Bae, T. (2016). The SaeRS two-component system of Staphylococcus aureus. Genes (Basel) 7:E81. doi: 10.3390/genes7100081
Loughran, A. J., Gaddy, D., Beenken, K. E., Meeker, D. G., Morello, R., Zhao, H., et al. (2016). Impact of sarA and phenol-soluble modulins on the pathogenesis of osteomyelitis in diverse clinical isolates of Staphylococcus aureus. Infect. Immun. 84, 2586–2594. doi: 10.1128/IAI.00152-16
McGuinness, W. A., Kobayashi, S. D., and DeLeo, F. R. (2016). Evasion of neutrophil killing by Staphylococcus aureus. Pathogens 5:32. doi: 10.3390/pathogens5010032
Monk, I. R., Shah, I. M., Xu, M., Tan, M.-W., and Foster, T. J. (2012). Transforming the untransformable: application of direct transformation to manipulate genetically Staphylococcus aureus and Staphylococcus epidermidis. mBio 3:e00277-11. doi: 10.1128/mBio.00277-11
Montgomery, C. P., Boyle-Vavra, S., and Daum, R. S. (2010). Importance of the global regulators Agr and SaeRS in the pathogenesis of CA-MRSA USA300 infection. PLoS ONE 5:e15177. doi: 10.1371/journal.pone.0015177
Moreillon, P., Entenza, J. M., Francioli, P., McDevitt, D., Foster, T. J., François, P., et al. (1995). Role of Staphylococcus aureus coagulase and clumping factor in pathogenesis of experimental endocarditis. Infect. Immun. 63, 4738–4743.
Novick, R. P. (2003). Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol. Microbiol. 48, 1429–1449. doi: 10.1046/j.1365-2958.2003.03526.x
Nygaard, T. K., Pallister, K. B., Ruzevich, P., Griffith, S., Vuong, C., and Voyich, J. M. (2010). SaeR binds a consensus sequence within virulence gene promoters to advance USA300 pathogenesis. J. Infect. Dis. 201, 241–254. doi: 10.1086/649570
Okumura, C. Y., and Nizet, V. (2014). Subterfuge and sabotage: evasion of host innate defenses by invasive gram-positive bacterial pathogens. Annu. Rev. Microbiol. 68, 439–458. doi: 10.1146/annurev-micro-092412-155711
Pier, G. B., and Elcock, M. E. (1984). Nonspecific immunoglobulin synthesis and elevated IgG levels in rabbits immunized with mucoid exopolysaccharide from cystic fibrosis isolates of Pseudomonas aeruginosa. J. Immunol. 133, 734–739.
Rogasch, K., Rühmling, V., Pané-Farré, J., Höper, D., Weinberg, C., Fuchs, S., et al. (2006). Influence of the two-component system SaeRS on global gene expression in two different Staphylococcus aureus Strains. J. Bacteriol. 188, 7742–7758. doi: 10.1128/JB.00555-06
Schwan, W. R., Langhorne, M. H., Ritchie, H. D., and Stover, C. K. (2003). Loss of hemolysin expression in Staphylococcus aureus agr mutants correlates with selective survival during mixed infections in murine abscesses and wounds. FEMS Immunol. Med. Microbiol. 38, 23–28. doi: 10.1016/S0928-8244(03)00098-1
Sinha, B., François, P. P., Nüsse, O., Foti, M., Hartford, O. M., Vaudaux, P., et al. (1999). Fibronectin-binding protein acts as Staphylococcus aureus invasin via fibronectin bridging to integrin-α5β1. Cell. Microbiol. 1, 101–117. doi: 10.1046/j.1462-5822.1999.00011.x
Steinhuber, A., Goerke, C., Bayer, M. G., Döring, G., and Wolz, C. (2003). Molecular architecture of the regulatory locus sae of Staphylococcus aureus and its impact on expression of virulence factors. J. Bacteriol. 185, 6278–6286. doi: 10.1128/JB.185.21.6278-6286.2003
Sun, F., Li, C., Jeong, D., Sohn, C., He, C., and Bae, T. (2010). In the Staphylococcus aureus two-component system sae, the response regulator SaeR binds to a direct repeat sequence and DNA binding requires phosphorylation by the sensor kinase SaeS. J. Bacteriol. 192, 2111–2127. doi: 10.1128/JB.01524-09
Sun, J., Zheng, L., Landwehr, C., Yang, J., and Ji, Y. (2005). Identification of a novel essential two-component signal transduction system, YhcSR, in Staphylococcus aureus. J. Bacteriol. 187:7876. doi: 10.1128/JB.187.22.7876-7880.2005
Thoendel, M., and Horswill, A. R. (2013). Random mutagenesis and topology analysis of the autoinducing peptide biosynthesis proteins in Staphylococcus aureus. Mol. Microbiol. 87, 318–337. doi: 10.1111/mmi.12100
Thomer, L., Schneewind, O., and Missiakas, D. (2013). Multiple ligands of von Willebrand factor-binding protein (vWbp) promote Staphylococcus aureus clot formation in human plasma. J. Biol. Chem. 288, 28283–28292. doi: 10.1074/jbc.M113.493122
Voyich, J. M., Vuong, C., DeWald, M., Nygaard, T. K., Kocianova, S., Griffith, S., et al. (2009). The SaeR/S gene regulatory system is essential for innate immune evasion by Staphylococcus aureus. J. Infect. Dis. 199, 1698–1706. doi: 10.1086/598967
Wang, L., Bi, C., Wang, T., Xiang, H., Chen, F., Hu, J., et al. (2015). A coagulase-negative and non-haemolytic strain of Staphylococcus aureus for investigating the roles of SrtA in a murine model of bloodstream infection. Pathog Dis. 73:ftv042. doi: 10.1093/femspd/ftv042
Yang, J., Liang, X., and Ji, Y. (2015). The novel transcriptional regulator SA1804 is involved in mediating the invasion and cytotoxicity of Staphylococcus aureus. Front. Microbiol. 6:174. doi: 10.3389/fmicb.2015.00174
Zhao, F., Cheng, B. L., Boyle-Vavra, S., Alegre, M. L., Daum, R. S., Chong, A. S., et al. (2015). Proteomic identification of saeRS-dependent targets critical for protective humoral immunity against Staphylococcus aureus skin infection. Infect. Immun. 83, 3712–3721. doi: 10.1128/IAI.00667-15
Keywords: S. aureus, survival, two-component system, SaeRS, coagulase
Citation: Guo H, Hall JW, Yang J and Ji Y (2017) The SaeRS Two-Component System Controls Survival of Staphylococcus aureus in Human Blood through Regulation of Coagulase. Front. Cell. Infect. Microbiol. 7:204. doi: 10.3389/fcimb.2017.00204
Received: 04 March 2017; Accepted: 08 May 2017;
Published: 29 May 2017.
Edited by:
Dongsheng Zhou, Beijing Institute of Microbiology and Epidemiology, ChinaReviewed by:
Weili Liang, National Institute for Communicable Disease Control and Prevention (China CDC), ChinaSanjay K. Shukla, Marshfield Clinic Research Foundation, United States
Hongxia Wang, University of Alabama at Birmingham, United States
Copyright © 2017 Guo, Hall, Yang and Ji. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yinduo Ji, aml4eHgwMDJAdW1uLmVkdQ==