Abstract
For over a decade there has been general, but not universal, consensus that the earliest known evidence for horse husbandry was at Eneolithic Botai, Kazakhstan, circa 3,500 BCE. Recent ancient genomic analyses, however, indicate that Botai is not the source of modern domestic horse stock (DOM2 lineage), but is instead related to the Przewalski clade of horses. DOM2 appears to instead to have emerged in early Bronze Age (mid 3rd Mill. BCE) in the Pontic-Caspian steppe, and spread quickly replacing other horse lineages after approximately 2,000 BCE. Whilst the specific evidence for earlier husbandry at Botai is not diminished by this evidence, it has broken the consensus regarding the early stages of horse domestication, with some now viewing it as a later event. This paper argues that domestication is rarely an event, but instead a process that is ongoing. The case is made for a “prey pathway” initial phase of domestication in multiple localities during the Eneolithic, which was based around local subsistence pastoralist niche construction. This took different forms due to the presence or absence of ruminant domestic stock in the Eastern European or Central Asian steppes, respectively. Whilst “push” factors likely played a part in the development of horse specialist pastoralism at Botai, it is suggested that “pull” factors accelerated the spread of DOM2 lineages, replacing others, in the later Bronze Age. The DOM2 spread was principally driven, not by local subsistence needs, but wider social, economic and military desirability of equestrianism. The long-term process of horse domestication continues in modernity with major breed changes caused first by the post-medieval agricultural revolution and, more currently, the desire for sporting achievement.
1. Introduction
Humans' early interactions with horses were as the hunters of a significant quarry (Olsen, 1989, 1996; Outram, 2006; West, 2006), but horses were also a favored subject of both parietal and mobiliary art during the Pleistocene (Bahn and Vertut, 1997; Pruvost et al., 2011). Whilst horses evolved in the Americas their interaction with humans there was cut short, with them going extinct upon the onset of the Holocene (Webb and Hemmings, 2006). In Eurasia, as environments became increasingly wooded, horse populations became more fragmented into patchy refugia (Leonardi et al., 2018), favoring the plains of the Iberian Peninsula, North and Central Europe (Benecke, 1994; Bendrey, 2012) and the steppes of Eastern Europe and Northern Central Asia (Kuzmina, 1997). Horse/human relationships of the early Holocene are relatively poorly evidenced or understood.
What is generally accepted is that the domestication of the horse was relatively late compared to other large domestic species in Eurasia such as cattle, sheep, goats and pigs (Clutton-Brock, 1999; Outram, 2014). Once harnessed, domestic horses had a revolutionary effect upon transportation, trade and types of warfare (Anthony, 2007; Kelekna, 2009). As a form of land transport, the horse was unbeaten for speed over the millennia until the steam trains of the industrial revolution. The use of domestic horses has also been long associated with the spread of bronze metallurgy alongside Indo-European languages and associated cultures by steppe pastoralists (Piggott, 1992; Mallory, 1996; Anthony, 2007), though others have favored an Anatolian origin for Indo-European (Renfrew, 1987; Gamkrelidze and Ivanov, 1995; Bouckaert et al., 2012). Aspects of this debate have been intensified by recent ancient genomic evidence. The massive genetic impact of the expansion of early Bronze Age Yamnaya steppe pastoralists across Europe (Allentoft et al., 2015; Haak et al., 2015), circa 3,000 BCE, certainly lends support to the former theory in relation to Indo-European language spread (Anthony and Ringe, 2015). However, the inclusion of domestic horses in that package is now open to question given the timing of the genetic spread of the modern lineage of domestic horses (DOM2) across the same territory, which was up to a 1000 years later (Librado et al., 2021).
This evidence for the spread of DOM2 horses matches well with the very late 3rd Mill. BCE dating of early chariot burials in the Trans-Ural Sintashta Culture (Anthony, 2009; Outram et al., 2011) and the first depictions on seals of equids being ridden in Mesopotamia (Anthony, 1996). This horizon is clear enough and some scholars would prefer to see this, or its immediate run-up, as being the effective date for horse domestication (e.g., Levine, 2004). Indeed, others (Taylor et al., 2020) see this as a phase associated strongly with wheeled vehicles and chariotry, with riding not becoming common until later still. Other scholars (Olsen, 2006a,b; Anthony, 2007; Outram, 2014) have argued for much earlier phases of the horse domestication process, which could potentially have more than one center. One potential such locus for early horse domestication is at the Copper Age Sredny Stog Culture site of Deriivka in Ukraine (c. 4,500–3,500 BCE) (Anthony, 1996) due to the high preponderance of horse bones and the finding of clear “bit wear” on the teeth of a stallion (Anthony and Brown, 1991; Anthony et al., 1991). However, radiocarbon dating revealed that this particular stallion was an intrusive Iron Age animal (Anthony and Brown, 2000). This culture remains of interest in terms of developing horse-human relationships, but conclusive evidence is currently lacking for husbandry. The search for earlier phases of horse domestication shifted eastwards to steppes of Northern Kazakhstan and the Eneolithic Botai Culture (c. 3,500–3,000 BCE), because this culture displayed an extreme economic focus on horses (Zaibert, 2009; Outram, 2014). Bit wear was also discovered on directly-radiocarbon-dated Botai horses (Brown and Anthony, 1998) and an extensive, multi-proxy, contextual argument was made for horse husbandry at Botai itself, and other Botai Cuture sites such as Krasnyi-Yar and Vasilkovka (Olsen, 2006a,b; Olsen et al., 2006). However, horse domestication at Botai was not universally accepted (see Levine, 2000, 2004; Levine et al., 2005), for reasons that will be discussed below, and thus the wider academic community tended to reserve judgement. However, further work provided additional lines of evidence, most notably the identification of lipid residues of both horse adipose fats and, crucially, milk in Botai ceramics (Outram et al., 2009). Whilst acceptance was still not universal, there was, for over a decade, reasonable consensus that Botai held the earliest evidence for horse husbandry, though the existence of other centers in Eastern Europe remained a possibility (Anthony, 2007; Anthony and Brown, 2011). In 2018, the first ancient genomics was carried out on Botai horses that revealed they were the ancestors of modern Przewalki's horses (Gaunitz et al., 2018) and could not have given rise to the modern domestic lineage, which is now known to have formed in the Pontic-Caspian steppe in the mid 3rd Mill. BCE (Librado et al., 2021). Whilst it is utterly clear that the Botai culture was not the source of modern domestic horses, nothing in Gaunitz et al. (2018) negated the evidence for husbandry at Botai. Indeed, new evidence for the presence of corral features at Botai sites was presented in that paper. None-the-less, the fact that Botai was not the source of modern domestic stock has led to new arguments against early domestication at Botai (Taylor and Barrón-Ortiz, 2021). Thus, there is current debate between those who see horse domestication as a late, single event, and those who see it as a multi-centered, multi-stage process. This debate speaks more widely to general understandings of animal domestication and is the topic of the rest of this paper.
2. Archaeological background to the Neolithic, Eneolithic and Bronze Age of Northern Kazakhstan
The Northern region of Kazakhstan is largely “forest steppe” comprising grasslands with stands of birch and pine (see Figure 1, for location of key named sites). The central and southern regions, on the other hand, are semi-arid and largely treeless. The focus of this section is on the North because it is within these less arid “forest steppe” zones that we see the development of particular horse specialization during the Eneolithic (Outram and Bogaard, 2019). The vegetation in the area during prehistory was very similar but with some increase in pine noted from the 4th Mill. BCE until the start of the Iron Age (Kremenetski et al., 1997).
Figure 1
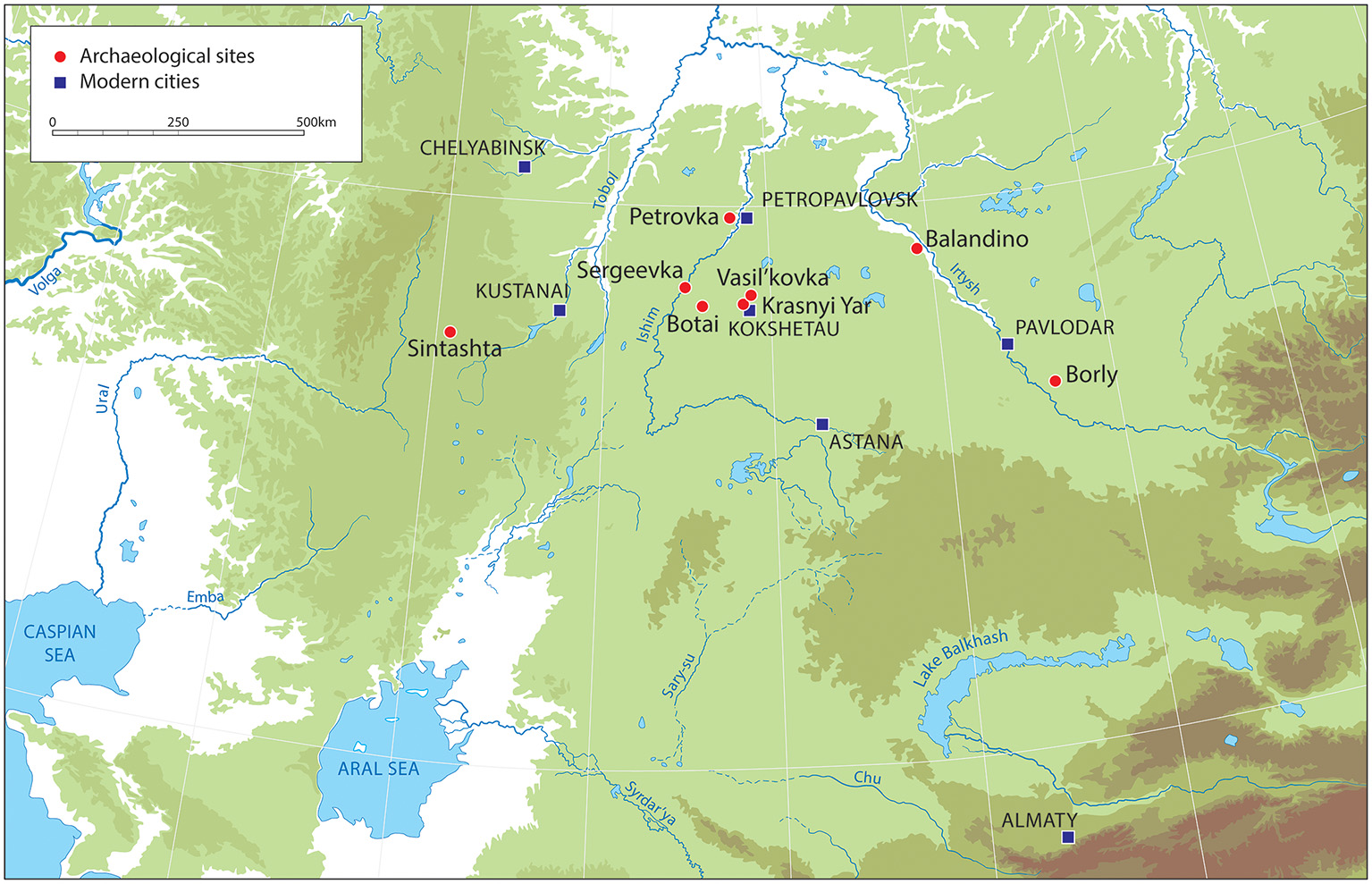
A map of Central Asia showing the locations of key archaeological sites mentioned in the text.
Following an aceramic Mesolithic, the 5th Mill. BCE saw the rise of the pottery using Neolithic Makhandzhar and Atbasar cultures (Kislenko and Tatarintseva, 1999), whose economy is not deeply researched but appears to be based on hunting, gathering and fishing. Large mammal species exploited include bison, aurochs and saiga as well as horses (Kislenko and Tatarintseva, 1999). Thus, the “Neolithic” label relates to possession of ceramics, rather than a farming economy, or indeed lithic tradition, which still focuses upon large blade and micro-blade technologies. Settlements are generally small and rather ephemeral in nature (Outram and Bogaard, 2019). As such, in western European nomenclature, this might be seen as a ceramic Mesolithic. It is worth noting that further North in the Russian forest steppe there were already cultures practicing mixed farming (Frachetti, 2008). It is therefore possible that Neolithic cultures of Northern Kazakhstan were at least cognizant of the exploitation domestic animals by neighboring peoples.
The following Eneolithic period commenced around 3,500 BCE and saw significant changes to settlement structure, economic focus and material culture. Two cultural labels are used for this period. In the central north was the Botai Culture (Zaibert, 2009), and to its west and southwest the Tersek Culture (Kalieva and Logvin, 1997). Sites assigned to these cultures display both intra- and inter-variation, but a focus on the exploitation of the horse, large settlements of pit houses, increased usage of ceramics and a shift toward bifacial and ground stone lithic traditions (Kalieva and Logvin, 1997; Zaibert, 2009). The type site of Botai itself stands out because of its size, well over 100 houses (Zaibert, 2009), and its utter specialization in horses. At Botai horses are not just a major focus but represent well over 99% of the faunal assemblage (Olsen, 2006a; Anthony, 2007). Furthermore, extensive flotation sieving and archaeobotanic analyses at Botai have not identified significant plant food sources (Motuzaite Matuzeviciute et al., 2019). Through recent archaeological fieldwork, it is now clear that this Eneolithic phenomenon extends further east to the catchment of the River Irtysh. In this region the site of Borly 4 (Svyatko et al., 2015; Merts and Merts, 2018) is of similar nature and shares an extreme focus on horses. As such, these Eneolithic horse-specialized societies extended for well over 1,000 km within the forest steppe zones between the South Urals and the foothills of the Altai. Recent ancient genomic work indicates that the Botai people were derived from Ancient Northern Eurasian (ANE) peoples, as part of a previously undescribed genetic cline that existed in northern Central Asia and southern Siberia up until the Bronze Age. The Botai people have no known surviving descendants (Damgaard et al., 2018; Jeong et al., 2019). Given the separation-by-distance structure presented with the cline Botai is situated, it seems most reasonable to conclude that the Eneolithic developments seen are an indigenous cultural adaptation, rather than an external introduction by migrating people. This does not deny the possibility of influence from neighbors.
These Eneolithic Cultures end in the early 3rd Mill. BCE and give way to a relatively poorly understood Early Bronze Age period. Sites from this period that have been subjected to extensive excavation and zooarchaeological analysis are comparatively thin on the ground (Outram and Bogaard, 2019). Sites such as Sergeevka and Balandino display the mixed use of horses, which are still very well represented, alongside domestic ruminants (Benecke and von den Driesch, 2003; Frachetti, 2008; Zaibert, 2009). The earlier 3rd Mill. BCE saw the expansion of Yamnaya-derived peoples from the Pontic-Caspian steppe, who had domestic ruminants, to form an enclave, known as the Afanasievo Culture, to the North of Altai. The Afanasievo share genetics with the Yamnaya, yet it is important to note that an unadmixed, ANE-derived (Botai-like), individual has been sequenced from a culturally Afanasievo-influenced burial from North Eastern Kazakhstan during this period (Damgaard et al., 2018). This highlights that Botai-related ancestries persisted into the early Bronze Age when mixed pastoralism commenced in the region and other external cultural influences are evidenced.
The middle Bronze Sintashta and Petrovka Cultures (c. 2,100–1,800 BCE), however, do represent major changes in population make-up due to migration. During this, and the subsequent late Bronze Age Andronovo Culture, the population of the central steppe displays almost complete genetic turnover to Sintashta-derived peoples characterized by admixture of Yamnaya pastoralist and European farmer lineages (de Barros Damgaard et al., 2018). Simultaneously we see the rapid replacement of local horses with the DOM2 modern domestic lineage (Librado et al., 2021). The Sintashta Culture of the Trans-Ural region of Russia and Kazakhstan displays wealth and status through burials that can contain weapons, chariots and horses (Anthony, 2009). During the Middle and Late Bronze Age, subsistence was, however, dominated by the pastoral exploitation of cattle, sheep and goats (Outram et al., 2012; Outram, 2015; Haruda, 2018). Both lipid residue analyses (Outram et al., 2012) in Kazakhstan and proteomic analyses in adjacent regions (Wilkin et al., 2021a; Scott et al., 2022) indicate very limited evidence for Bronze Age horse milking, with dairying dominated by ruminant exploitation. Focused research upon Mongolia indicates an in increase in horse milking at around 1,200 BCE (Wilkin et al., 2021b), though other regions have not been studied in such detail as allow full regional comparisons.
The Petrovka period sees fewer horse burials and they are very rare in the Andronovo, yet there is an association between increased proportions of horse bones and lipid residues in ceramics with funerary contexts (Outram et al., 2011). Whilst ratios of cattle vs. caprines correlate well with environment and aridity (Bendrey, 2011a), it has been suggested that high proportions of horses during this period might be better explained as an expression of wealth or status (Outram and Bogaard, 2019, p189). Whilst the southeast of Kazakhstan presents evidence for the Bronze Age exploitation of wheat and broomcorn millet (Frachetti et al., 2010; Matuzeviciute et al., 2015) there is scant such evidence in central and northern regions (Lightfoot et al., 2014) which appear to have remained dependent upon pastoralism and occasional hunting. Generally elevated human δ15N values had once been taken to indicate high levels of fishing during Central Asian prehistory (O'Connell et al., 2003). However, it has recently become possible to discern whether such high values have an aquatic or terrestrial dietary cause through compound-specific determinations (Naito et al., 2016), and extensive sampling in Bronze Age Kazakhstan has revealed a largely terrestrial cause (Itahashi et al., 2020).
3. The Botai domestication debate
3.1. Evidence in favor of horse domestication at Botai
It is decidedly dubious whether a clear, dichotomous distinction exists between what is wild and domestic (Vigne et al., 2005). In many respects, when discussing human-animal relations in the past, it would perhaps be more helpful to describe the actual nature of a close relationship in terms of forms of husbandry, economic and social context, and associated cultural values. In so far as “domestic” is a useful term, this paper takes it to mean a shift in focus away from the hunted dead animal toward the live herd and guaranteeing its future progeny (Meadow, 1984; Outram, 2014). This approach is consistent with Zeder (2015, p. 3191) definition that “domestication is a sustained multigenerational, mutualistic relationship in which one organism assumes a significant degree of influence over the reproduction and care of another organism”. Clearly, such a relationship will likely have a raft of genetic effects, whether intended or inadvertent (see MacHugh et al., 2017), but need not lead to complete genetic separation from the wild, as implied by older definitions that fail to match-up to real-life examples (e.g., Blumler and Byrne, 1991). Such a new relationship will also likely affect species abundance, diet, disease and zoonoses, mobility, herd structure, pathology and plastic activity/function-related morphology (Outram, 2015; Zeder, 2015) within animal populations. Clearly, patterns of human economy, settlement structure, technology, mobility and cultural expression will also be impacted in return.
Initial attention was drawn to Botai as a potential locus for early horse domestication because of the sudden specialist focus upon that animal in association with the appearance of substantial settlements (Olsen, 2006a; Anthony, 2007; Outram and Bogaard, 2019) (Figure 2 depicts the interpretation of geophysical survey at Botai). In many respects, as discussed below, when put its proper context, this line of evidence is actually powerful support for some degree of horse husbandry. In terms of “smoking-gun” type evidence, however, the first to be identified at the site was “bit wear” (Brown and Anthony, 1998). It had been demonstrated experimentally that, even when soft bits were used, as likely the case at Botai, such harnessing could cause characteristic bevelling of the lower P2 teeth (Anthony et al., 2006). Bendrey (2007) devised two additional criteria for identifying bitted animals, including a parallel-sided band of wear on the anterior of the P2 (in the absence of any unusual dietary wear on other facets) and exostosis on the mandible's diastema indicative of irritation of the gums by the bit. Such patterning was found not to exist on any specimens studied that were known not to have been harnessed (Bendrey, 2007). Both these new forms of evidence were found later at Botai (Outram et al., 2009). Thus, Botai displayed examples of Type 1 (bevel), Type 2 (parallel band of wear) and Type 3 (diastema pathology), as they became labeled (Anthony and Brown, 2011), representing a reasonable basis to conclude that at least some Botai horses were harnessed and possibly ridden.
Figure 2
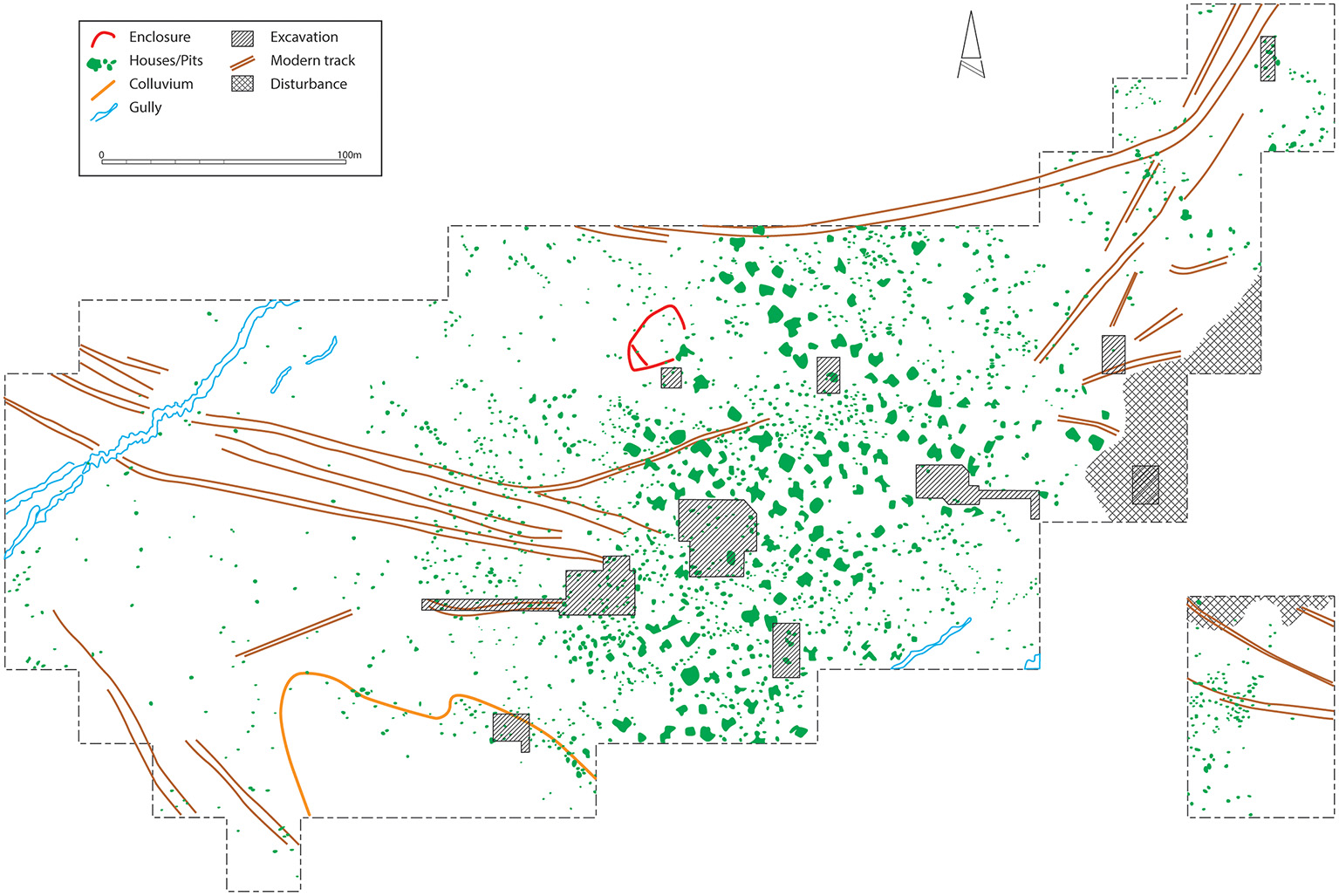
Interpretation diagram of fluxgate gradiometry undertaken at Botai.
Olsen (2006a; 2006b; Olsen et al., 2006) subsequently compiled a strong set of contextual arguments for horse husbandry in the Botai culture, combining consideration of Botai itself as well of other Botai Culture sites called Vasil'kovka and Krasnyi Yar. Horses represent a significant transport challenge to any hunter (Outram and Rowley-Conwy, 1998), so if only human foot transport was available one would expect to see “schlepp effect” (differential skeletal part transport) from the kill-site to the settlement (Perkins and Daly, 1968). Yet skeletal parts are quite evenly represented (Olsen, 2006a) suggesting slaughter near or in the settlement rather than hunting at some distance. Such patterning is not in frequently used as an indicator of human control and domesticity, and at Botai it is accompanied by some apparent cases of cranial injury akin to pole-axing as a potential controlled slaughter method (Olsen, 2006a; Outram and Bogaard, 2019, Figure 8.2). Olsen et al. (2006) also note that the tool assemblage at Botai Culture sites is very heavily biased toward scrapers and bone tools associated with hide working. Points, whilst present in limited numbers, are far less well represented than one would expect for a society living almost exclusively from big game hunting.
A further indicator that horses may have been kept close to, or in, settlements is abundant evidence for the use of horse dung within house construction materials (French and Kousoulakou, 2003; Olsen et al., 2006), as evidenced through soil micromorphology. Of course, the dung of wild horse can be collected from surrounding lands, though one would assume such a prey animal would be wary of grazing in the immediate vicinity of the habitations of it primary predator (a point pertinent to the lack of schlepp effect also). Dung would be abundant if domestic horses here grazed or corralled close to settlements. Indeed, there is evidence for corral-like structures at Botai Culture sites. At Botai itself, to the northwest of the settlement, on the very edge of the spread of pit-houses, there is an enclosure (see Figure 2). This feature was picked up as an anomaly in a fluxgate gradiometer survey, and is trapezoidal in shape (about 25 m long, and 20 m wide). It has been ground-truthed by excavation trenches and a small open-area excavation. There is a continuous trench feature with post settings and additional post holes that contained bone radiocarbon dated to the Botai period (Gaunitz et al., 2018). Magnetic survey at Krasnyi Yar (Olsen et al., 2006, Figure 2) also displays an anomaly consistent with being a trapezoidal enclosure (about 25 m long by 15 m wide) and similar positioned to the immediate northwest of the settlement's housing. It is hard to see the function of such enclosures for horse hunters, but its size and position are ideal for a horse corral.
Two further lines of biomolecular evidence include residue evidence for horse milking and ancient genomic evidence for unusual horse coat colors. Identifying horse milking from absorbed lipid residues in ceramics is more challenging than in ruminants. Whilst ruminant animal milk can be discerned from compound-specific isotope ratios of carbon, due to differing metabolic pathways that create a fractionation effect (Dudd and Evershed, 1998), this does not work for non-ruminant equids. Carbon stable isotope ratios can identify the fat as equine, but deuterium isotope ratios must additionally be employed to distinguish adipose from dairy fats (Outram et al., 2009). Horse milking is principally a summer occupation and milk synthesized in summer produces deuterium isotope ratios that are radically different to adipose fats that represent an integrated climatic signal for the year (Outram et al., 2009). This only works clearly because of Central Asia's extremely continental climate and the method was verified experimentally on modern samples before being applied at Botai. Five potsherds from Botai clearly had the less depleted deuterium isotope ratios to be consistent with milk (Outram et al., 2009). This was seen by many as being a smoking gun for domestication.
Changes in coat color diversity, whether as a result of deliberate selection, founders' effect or inadvertent removal of selective pressure are often seen as potential indicators of domestic control (MacHugh et al., 2017). This line of evidence has been of interest to those studying the ancient genetics of horses (e.g., Ludwig et al., 2015). Gaunitz et al. (2018) found limited evidence for the leopard-spotting complex in Botai horses. The leopard-spotting complex might be attractive but can lead to eye-sight problems (Bellone et al., 2013) and one line of argument is that it should be in very low proportion in the wild, but might increase under human husbandry as selective pressures are removed. More recently, further examples of leopard-spotting and silver coat color associated alleles have been found in Botai samples (Librado et al., 2021) which is tantalizing, but in need of further investigation and sampling within the same genetic context (same clade over time) to reach clear conclusions.
One further line of evidence, which derives from molecular genetics, is the reconstruction of demographic trends. Drift-based models for the size of populations (Gaunitz et al., 2018) suggest that horse populations were seriously crashing in the earlier Holocene. Perhaps this is not surprising given their actual extinction in the New World (Webb and Hemmings, 2006). This more fractured and declining horse population is also backed up by the modeling of archaeological finds of horse bones (Leonardi et al., 2018) and the notably structured-by-distance nature of early Holocene horse genetics (Librado et al., 2021). In this context, it is worth returning to the original reason zooarchaeologists became interested in Botai. If horse populations were patchy and declining, and this general pattern holds true in northern Central Asia, why would what had been mixed hunter-gatherers react by (a) focusing almost exclusively on the resource in decline, whilst (b) apparently reducing mobility and increasing settlement size? Such a reaction would seem to contravene the logic of all optimal foraging models, particularly “diet breadth” models (MacArthur and Pianka, 1966), but also well-known models related to mobility and ecological productivity such as the “economic defensibility” model (Dyson-Hudson and Smith, 1978). Whilst the use of such models is unfashionable, and they should generally not be used deterministically, they are of heuristic value and one must recognize subsistence economies need to be at least tenable (Outram and Bogaard, 2019). To quote Higgs and Jarman (1975, p. 2), “…ultimately all human culture and society is based upon and only made possible by biological and economic viability…however unfashionable the term and ideas behind determination may be, the very existence of natural laws presupposes a degree of determinism.” Is it economically viable for peoples who are purely hunters to have adapted to declining horse numbers in such a way? Nothing similar was seen in the Pleistocene when horse populations were much higher. On the other hand, if the niche construction of local hunter-gatherers involved the evolution of forms of horse husbandry, then the archaeological and genetic patterns one sees are well explained. At present, only Eurasia-wide equine demographic models are available (Gaunitz et al., 2018) and, if more region-specific models become available in the future, the dynamics of horse specialization at Botai could be revisited.
3.2. Challenges to Botai domestication evidence
Whilst early horse domestication need not necessarily relate to uses of horses for transportation, none-the-less, one of the longest standing arguments employed against domestication before c. 2,200 BCE has been the lack of artistic depiction of riding or chariotry (e.g., Taylor et al., 2020). Early artistic depictions of chariotry come from Assyria (c. 2050 BCE) (Owen, 1991), almost synchronously with the earliest physical evidence for actual chariots in burials of the Sintashta Culture (see Figure 3). Whilst this argument has been repeatedly employed, it is worth noting that there is no such figurative depiction of any activities at Botai, with decorations on ceramics and bone being geometric in nature (see Zaibert, 2009). Should we thus conclude the Botai people also did not hunt, as there is no depiction of that activity either? It is worth reflecting on the wider nature of prehistoric societies and their use of figurative art. It is clear that for many prehistoric societies one sees little or no such art, whilst it flourishes in others. An example of this that is very pertinent to the same Bronze Age horizon is the use of seagoing boats in northwest Europe. The appearance of rock art and other depictions of such boats is synchronous with the actual physical survival of waterlogged, sea-going sewn plank boats in the early Bronze Age (Van de Noort, 2006). However, seagoing boats of sufficient size to carry Neolithic settlers and their domestic cattle to the British Isles must have been employed from the earliest Neolithic. This is indisputable, so the evidence base tells us less about the existence of seagoing boats, but more perhaps about the nature of coastal archaeological preservation, developments in artistic traditions and how societies wished to project what was important to them.
Figure 3
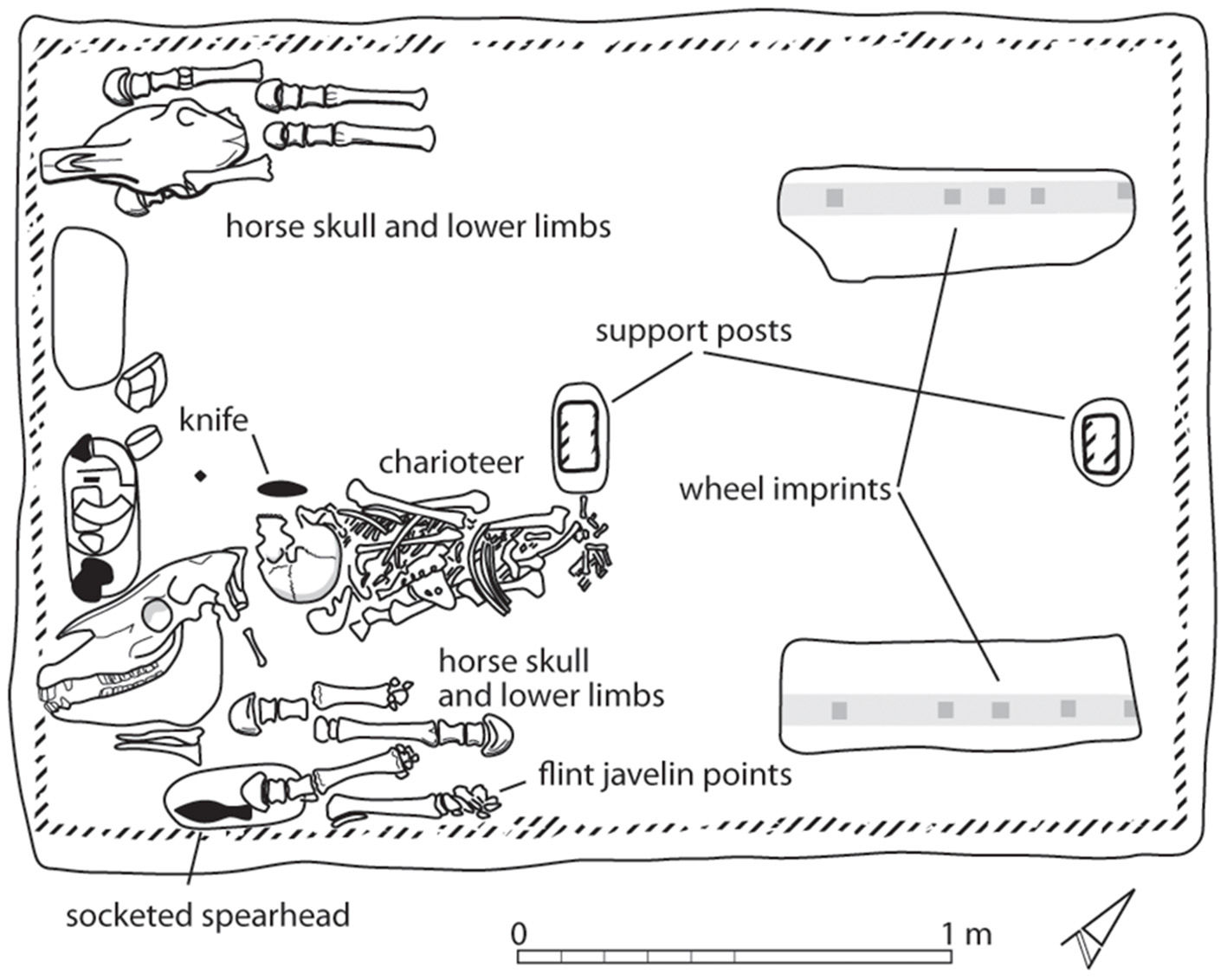
Sintashta Mogila grave 30, which contained an inhumation and two horses represented by heads and feet, alongside weapons, horse tack and the remains of a spoked-wheeled chariot (redrawn, after Anthony, 2009, Figure 4.4).
Whilst the Botai culture has provided no artistic portrayal horse husbandry, horse harnessing is depicted in bronze artifacts found of the Elunino Culture and Seima-Turbinsky complex of the Early Bronze Age in the Ob-Irtysh Region of Kazakhstan and Russia (Molodin and Neskorov, 2010; Kovtun, 2013; Merts, 2016). Horses, with a stocky neck and brushy mane - signature traits of Przewalski (Botai-lineage) horses – are depicted on the pommels of knife hilts often straining against reins. A classic example recently discovered in North East Kazakhstan (Merts, 2016) can be seen in Figure 4. Another example from the Ob-Irtysh interfluve in Russia depicts two horses in parallel, both with reins (Molodin and Neskorov, 2010), whilst one depicts a person on the ground holding and pulling the reins (Kovtun, 2013), removing any doubt that control by harnessing is what is being depicted. The potential significance of these bronzes to horse domestication is only becoming apparent now that the geospatial and temporal dynamics of Botai-lineage and DOM2 horses has become more apparent from genomic work (Gaunitz et al., 2018; Librado et al., 2021). Both the Sintashta Culture and these Ob-Irtysh bronzes date to the late 3rd Mill. BCE, but the Ob-Irtysh is well over 1,000km further East, with no current evidence for DOM2 spread to that region so early. Moreover, unadmixed Ancient Northern Eurasian (ANE) peoples, similar in their genomic composition to earlier Botai groups, were still present in the Ob-Irtysh at this time (Damgaard et al., 2018). Furthermore, Seima-Turbinsky complex bronzes employ superior casting technologies that were yet to be adopted by the Sintashta Culture (Merts, 2016), implying that there was limited contact and exchange between the two. During the Early Bronze Age of the Ob-Irtysh we see early pastoralism involving introduced domestic ruminants plus the presence of horses in the zooarchaeological assemblages. Hence, those horses are either unexpectedly early and unevidenced introductions of DOM2 before associated human migrations from the west, or Botai-lineage domestic horses in continued use from the stock first husbanded in the Eneolithic. This question can be resolved by further genomic research.
Figure 4
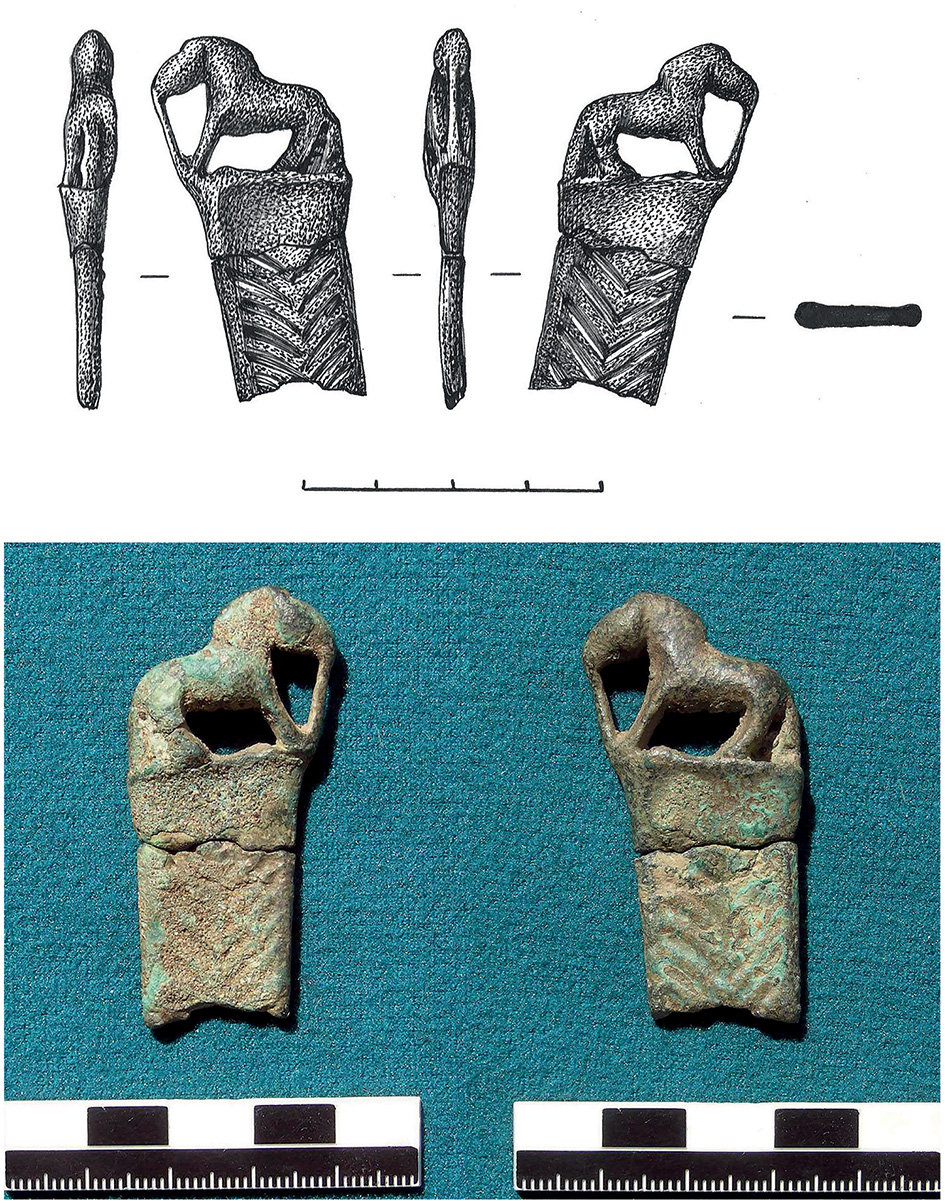
An Early Bronze Age Seima-Turbinsky complex dagger hilt from the Pavlodar Region of Kazakhstan, depicting a horse straining against reins (Merts, 2016).
Bit wear evidence on teeth has been the subject of constant debate. The bevelling noted by Brown and Anthony (1998) is certainly a type of bit wear that is regularly observed and has been experimentally demonstrated (Anthony et al., 2006). However, this method can suffer from some equifinality, as similar bevelling can occur as a result of malocclusion. In such cases the maxilliary P2 tooth hooks over the mandibular P2 (Bendrey, 2007, 2011b). Bevelling has been noted in wild Pleistocene horses (Olsen, 2006a,b). It is possible, however, to distinguish bit wear bevels from hooked malocclusion if up upper and lower jaws are found together, but equifinality remains at Botai. Bendrey's (2007) further research discovered that a different form of bit wear did not suffer from this type of equifinality when tested against a substantial set of horses specimens with known histories. This method identifies a clear parallel-side band of wear down the anterior surface of the mandibular P2, that penetrates to the dentine and is present in the absence of other irregular wear on buccal and lingual sides of the tooth, which can be caused by diet and advanced age (Bendrey, 2007). This type of wear was identified on a Botai stallion (Outram et al., 2009). Taylor and Barrón-Ortiz (2021) studied a very large sample of North American Pleistocene horses and presented a picture of a specimen purporting to display this type of wear. However, the specimen shown did not meet (Bendrey, 2007) criteria because it was a heavily worn tooth with considerable dietary wear on buccal and lingual facets, of a type explicitly not classified as bit wear and unlike the Botai specimen (see Outram et al., 2021 for direct comparison). Thus, if anything, this work demonstrated the absence of this type of bit wear in an even larger reference collection unbitted horses, hence strengthening the empirical basis of Bendrey's (2007) method. There is no question that equine bit and other dental wear can be highly variable. Bit wear does not occur in all bitted horses and can take different forms. This is highly dependent upon the type of bit (if one is used at all), the horses' behavior and the equestrians' styles and competence. It remains the case, however, that there is wear at Botai consistent with bit use that has not been found in unbitted animals.
Taylor and Barrón-Ortiz (2021) also questioned the evidence for horse milking at Botai (Outram et al., 2009). This critique was two-pronged, suggesting an alternative cause for the compound-specific δ2H values and pointing to proteomic results that did not show milking at Botai (Taylor and Barrón-Ortiz, 2021). Their suggested alternative to horse milk residues was summer seasonal hunting for meat. This is a fundamental error of methodological understanding, since in the original paper (Outram et al., 2009) this possibility was specifically tested for in reference samples and summer meat was found to present an integrated δ2H signal for the year and not a summer signal (Outram et al., 2021). This was the basis upon which the original method was founded. The use of recent proteomic evidence is no less problematic. Taylor and Barrón-Ortiz (2021) cite recent proteomic work on human dental calculus from sites in the Pontic-Caspian steppe and Botai that did not find horse milk at Botai, but did find it in the Bronze Age of Pontic-Caspian steppe (Wilkin et al., 2021a). The argument was thus made that there was evidence that horses were not milked at Botai and that was a later phenomenon associated only with DOM2 horses (Taylor and Barrón-Ortiz, 2021). However, they fail to note that the positive result in the Pontic-Caspian was based upon finding horse milk in only 2 of 17 samples, whilst only 2 samples were available from Botai, both negative (Wilkin et al., 2021a). The hit rate in the Pontic-Caspian was only 11.7%, with 15 negative samples. The result for Botai was based upon only 2 specimens, so, if one considers these frequencies, the binomial probability of observing 2 negative drawings is equal to 0.7785. Therefore, Taylor and Barrón-Ortiz's (2021) conclusion is not statistically supported (Outram et al., 2021). Taylor and Barrón-Ortiz (2021) are also unclear about the actual date of the positive Pontic-Caspian samples, hence allowing the reader to assume a much late date in line with their argument. In fact, the date given in the paper (Wilkin et al., 2021b) is early 3rd Mill. BCE, which in fact demonstrates horse milking at a time much closer to Botai dates than the widespread use of DOM2. In fact, it pre-dates clear evidence for DOM2 as understood from the ancient genomics (Librado et al., 2021). Even this date requires further reflection, because the date given in Wilkin et al. (2021a), has undergone a reservoir effect recalibration making it younger. This adjustment is open to question because an extensive study of dietary stable isotope values and radiocarbon dates of both humans and herbivorous animals (that do not suffer reservoir effect) (Hollund et al., 2010) concluded that such reservoir adjustments were unsafe in this region. The likely reason for this is that we now know that aquatic input into pastoralist diets cannot be assumed from δ15N values in the absence of compound-specific determinations (see Itahashi et al., 2020). Without adjustment, the date of these milk residues would be contemporaneous with Botai. In either case, they represent evidence for milking of pre-DOM2 horses. Cutting-edge methods will always be associated with some uncertainty, but there remains valid evidence for milking of horses before DOM2 at Botai and in the Pontic –Caspian steppe.
A final longstanding debate relates to herd structure at Botai. It has been argued that Botai does not have the expected focus on the slaughter of sub-adult males one would expect to fit economic models for meat production (Levine, 2004; Levine et al., 2005). This is true, the majority of animals at Botai are adult, but in many cases younger adult. Is this evidence against domestication at Botai? The expected “meat” pattern derives from well-understood models applied to domestic stock in mixed farming societies (see Payne, 1973). The presence of such a strategy would be good evidence for an economy wishing to maximize its production of meat, whilst economizing on the fodder stocks needed for overwintering. However, if the herd structure was maintained to provide a range of primary (meat) and secondary (milk, transport) products then the herd structure would be different (Anthony and Brown, 2011). It is arguable that Payne (1973) type models are not entirely applicable to small-scale subsistence pastoralists, and maybe specifically not horses. Pastoralists, reliant entirely on their herds, have general reluctance to slaughter animals, often viewing it as “sacrifice” (Ingold, 1980, p. 101; Lavrillier, 2011, p. 216) and selection for slaughter prior to winter is often based upon culling the weak that are unlikely to survive rather than age-specific selection (Fijn, 2011, p. 197). Mongolian pastoralists avoid slaughtering young animals (Fijn, 2011, p. 227). Slaughter is not necessarily either about maximizing production or saving fodder. Herders rely upon their animals totally and their herd is their wealth and security. Thus maintaining herd size and culling (sacrificing) only what is required is thus a key form of economic risk reduction. Sedentary mixed farmers are severely limited by available fodder, hence a strategy that maximizes production in relation to foddering needs. A steppe pastoralist is not necessarily reliant upon fodder stores, but upon moving animals to new grazing land, whether this takes a nomadic form of movement or herder movement tethered around a more permanent settlement (see Wendrich and Barnard, 2008). Furthermore, horses are naturally adapted to year-round grazing and, unlike ruminants, are very capable of clearing the snow to feed (Shishlina, 2003, p. 356). The Botai herd structure may perfectly well fit the needs of horse specialist, mixed use, subsistence pastoralism. Matters may be complicated still further if Botai contains a mixture domestic animals with additionally hunted or more loosely herded horses, as suggested by Olsen (2006a). If there are spatio-temporal seasonal factors in play regarding site of carcass deposition or other taphonomic factors, matters are complicated still further (Outram and Bogaard, 2019, p. 174).
3.3. Summary
There is ample, multi-proxy evidence for the management of horses at Botai and the absence of some forms of evidence, which are particular to specific uses of domestic horses, does not imply they were not husbanded in some form. Artistic depictions of specific activities are frequently absent in prehistory and many past cultures entirely lack that kind of artistic tradition, including Botai. The Early Bronze Age horse bronzes of the Seima-Turbinsky complex in the Ob-Irtysh, do, however, have the potential to represent control of horses in the region prior to the introduction of DOM2 stock. Domestic animal herd structures frequently do not conform to specialized production patterns. The critiques related to “type 2” bit wear and lipid residue analyses contain fundamental misunderstandings of method and thus do not detract from the validity of the original results, which had already taken full account of the issues raised. There remains evidence in favor of bitting and horse milking at Botai. The implications of recent proteomic research into horse milking are misrepresented. If correct, the proteomic results in fact demonstrate horse milking prior to DOM2 selection and do not provide meaningful results in relation to Botai specifically.
Horse specialization in the face of falling horse populations, the presence of likely corrals, ample use of horse dung, the lack of schlepp effect, evidence of poleaxing, the nature of the material culture, alongside still-valid evidence in favor of milking and bitting are all suggestive that there must have been at least some degree of husbandry at Botai, which may well have continued into the Early Bronze Age within Northern and Eastern Kazakhstan in the period before DOM2 horses were introduced.
4. Recent trends in domestication research
When considering the precise nature of the horse-human relationship at Botai and the overall long-term processes of horse domestication, including DOM2 and beyond, it is important to consider a number of recent general insights into the concept of domestication.
One of the most important contributions to thinking on this topic was recently made by Bogaard et al. (2021). They argue powerfully that domestication must be seen as a “dynamic, ongoing process” (Bogaard et al., 2021, p. 61). This seems like a simple enough observation, but if their meaning is fully comprehended this represents a significant critique of how domestication often been presented. Firstly, there is the clear implication that domestication is not an “event”; the “wild” was not made “domestic” by short-term human agency. It was a complex process of niche construction that altered the human-animal relationships in a way that defies simple dichotomous separation as “wild” or “domestic” during its initial phases, even if such a distinction appears in hindsight. Secondly, the process was not something that only happened in the past. It is still happening, not only in relation to new species, but also in relation to those already considered to be domestic. Thirdly, the process it not something that happens to “things” (animals) as a result of human agency, but instead both the animals and humans are themselves processes. Both the animals and humans involved change their nature as the relationship evolves and have agency (Bogaard et al., 2021), though this need not always imply that agency is equal, depending upon circumstances.
A different observation has only recently become possible with the acceleration of ancient genomic research. Much zooarchaeological research, and that based upon modeling from modern DNA, suffers from the fallacy of presentism, assuming that one can simply classify modern species as wild or domestic and model backwards in order to understand domestication in the past. It has become increasingly clear that processes of admixture, mass introgression and extinction make for very different biogeographies in the past, such that representative ancient genetic studies essential (MacHugh et al., 2017). An early example of such a surprise relates to domestic pigs in northwest Europe. Modern European domestic pigs appear closely related to European wild boar, so the presentist fallacy was to assume pigs were locally domesticated in Europe. However, it transpires that European Neolithic farmers kept pigs of Near Eastern ancestry, yet those domestic pigs interactions led to mass introgression from local wild animals to the extent that modern patterns utterly fail to describe the early stages of European pig farming (Larson et al., 2007). Another fascinating example is the relatively recent, but almost total, loss of Native American dog breeds (Ní Leathlobhair et al., 2018). Whilst, dogs are well attested, archaeologically and ethnographically, in the Americas from 9,000 years ago until the recent past, their lineages are almost totally lost, surviving in the modern day mostly in a form contagious canine cancer. A final illustrative example has only just been published. Ancient genomics has revealed a previously unknown goat lineage, now labeled the “Taurasian Tur” (Capra taurensis), which appears to have made some genetic contribution to Western Eurasian Neolithic domestic goats, but not those East of the Zagros, and is now extinct (Daly et al., 2022). This reveals regional complexity and additional lineages involved in the processes of goat domestication, not visible in modern genetics, and in need of further investigation. Hence, it is not safe (e.g., Taylor and Barrón-Ortiz, 2021), to use the modern lineage of domestic horses as a line of argument against the involvement of another lineage within the domestication process. Redding (2005) noted that animal domestication processes might involve “failed experiments”. This is true, but it also now clear they can be successful but subsequently lost.
Zeder (2012, 2015) laid out a number of “pathways” to animal domestication. The “commensal pathway” refers to animal species attracted into anthropogenic settings, often by food waste or stores, that go on to develop a closer relationship, e.g., dogs. The “prey pathway” relates to a shift from hunting an animal, though increasing game management, to husbandry. The “directed pathway” implies a deliberate human goal to domestic a new species (Zeder, 2012). It is also possible to identify that human motivations within such processes could perhaps be classes as having either “push” or “pull” causal factors (Zeder, 2015). “Push” factors relate to need and are usually models relating to subsistence stress, whilst “pull” factors are based less around necessity but a desire for improved circumstances whether economic, a matter of social competition or improved lifeways in general. The above discussions do not negate the value of considering such pathways or causal factors as part of domestication processes. However, it is clear that in an ongoing process these may shift over time. Furthermore, if domestication did involve multiple lineages in multiple loci, different pathways and causes might pertain and, indeed, interact over time. How does this apply to process of horse domestication?
5. Discussion of the nature of a multi-center, multi-stage horse domestication process
Any consideration of the complex process of horse domestication is likely to oversimplify, but it is possible to evidence three key phases where there was a rapid and marked change in human-equine relationships. The first pre-dates the emergence of the modern domestic lineage (DOM2) and involves an economic intensification in the pastoralist exploitation of horses for subsistence. Whilst most clearly seen in the Eneolithic of the forest steppes of Central Asia, similar processes were likely underway, with regional variation in Eastern Europe and maybe elsewhere. The second phase is the emergence of DOM2 in the Pontic-Caspian steppe, followed thereafter by its rapid spread throughout Eurasia (Librado et al., 2021). This was closely linked with developed equestrianism. Many aspects of horse-human relationships changed in the following centuries, including the spread of domestic horses to the wider world. However, a third key phase is relatively recent and linked to the post-medieval agricultural revolution and subsequent modern interest in equine sporting performance.
5.1. Local subsistence pastoralism
Eneolithic and early Bronze Age developments in the horse-human relationship are apparent in several regions of Eastern Europe and Central Asia (see general summary in Anthony, 2007). However, it is clear that these were localized developments that did not significantly disrupt the geographical structure of horse genetics (Librado et al., 2021). In the case of Botai the result was near total economic focus upon the horse at the same time that settlement size radically increases. This author would argue that, in the context of generally declining horse populations, specialized hunting would not be viable and is not consistent with the evidence seen at Botai. Horse husbandry at Botai included at least some physical control, including corralling, of at least some of the horses present, and there is evidence for milking and harnessing (Outram et al., 2009; Outram and Bogaard, 2019). The geographical structure of Botai human (de Barros Damgaard et al., 2018) and horse genetics (Librado et al., 2021) suggests this was most probably a local economic transition. As such, this was likely a “prey pathway” adaptation where hunters increased the management of their equine prey until that became husbandry and a specialist form of horse pastoralism. Given the trajectory and pressure on horse populations (Gaunitz et al., 2018), it can be argued that a “push” factor was in play. Neolithic origins of this process remain an unevidenced possibility. By the Eneolithic the shift had significantly changed faunal patterning, settlement structure and the material culture seen. Whilst there was extreme focus on horses across the entirety of northern Kazakhstan at this time, there is subtle regional variation, with Botai itself being both the largest and most specialized site.
It is possible to debate a range of possible economic scenarios at Botai. Chechushkov and Kosintsev (2020) favor specialized horse hunting alongside the use of a number of tamed/domestic animals, a possibility that has also been mooted by Olsen (2006a). Such a scenario may well have had a role in the process of moving from hunting to herding. However, it perhaps only partially explains the full range of evidence at Botai and there are two further arguments against it. Firstly, the use of a few tamed animals would not have solved hunting pressure on declining wild horse populations. Secondly, it is not clear why a shift toward using some tamed horses would result in a change in frequency of hunted animal species (Rowley-Conwy, pers. comm.). Having a number of tamed horses would not obviate the value of hunting aurochs, bison, saiga and deer alongside horses. However, if the activities of horse husbandry guarantee the subsistence base, then the need for hunting, any species, is reduced, and the problem of declining population is solved. The exploitation of secondary products (mare”s milk), would have further added to the subsistence base and provided nutritious and storable foodstuffs.
There are no ethnographic sources to inform us about specialized, subsistence, equine pastoralism. The sources we have relate mixed equine/ruminant pastoralists or modern forms of horse ranching, where the ranching is used to produce foodstuffs for market (see Ingold, 1980), not direct subsistence. However, it may be possible to learn something about the range of potential lifeways at Botai from the consideration of reindeer herders. Reindeer herders are often also heavily specialized in that single animal species in the absence of significant plant food supplies, and reindeer can also be hunted, tamed, herded, milked, ridden and used for traction (Ingold, 1980, p. 10; Outram, 2014). Because of these multiple functions the extent or closeness of the reindeer-human relationship (or extent of domesticity) will vary depending upon the role of the animal. Animals that are ridden, for instance, may be much more controlled, kept separately, and even develop a closer bond with humans. Females that are milked, will likely have much closer association with humans than animals herded for their meat, as a group. In the case of Botai is possible to imagine a limited number horses being ridden and used to aid herding, which could be either relatively loose or tightly controlled. The level of physical control or breeding exerted may well be considerably different, thus producing differing effects upon genetics, activity-related morphology, mobility patterns or age at death, for instance. Distinguishing such complex combined patterning is not necessarily beyond cutting-edge methodologies in the future.
The Evenkis reindeer pastoralists of Siberia keep “domestic” reindeer, as significant herds that are moved to ensure sufficient pasture. As well as the general domestic herd, they also employ “transport” reindeer (Lavrillier, 2011, p. 217). They maintain a concept of wild and domestic landscapes. The wild species they hunt, including such species as deer, elk and “wild” reindeer, tend to stay away from human activity and areas trampled and associated with the keeping of domestic stock (Lavrillier, 2011, p. 220). This practice creates a tripartite division of animals of the same species. Such a model might well be appropriate for Botai, since there are a very small number of mixed wild large mammal species present and that might include a small number of wild horses encountered on hunting trips in an otherwise pastoral subsistence economy aided by limited riding to aid herding.
Whilst horses overwhelmingly dominate at all Botai Culture settlements, the proportions of large, wild, non-equine species at Krasnyi Yar and Vasil'kovka are slightly higher (Olsen et al., 2006). It is also notable that, through geophysics at those sites (Olsen et al., 2006), that the settlement structure comprises clear linear rows of pit-houses without overlap. This is not true of Botai, where there are many more houses, and apparently both lines of pit houses as well as sub-circular clusters (see Figure 5). It is also possible to discern overlaps between houses and alignments suggesting multiple phases with potentially different spatial arrangements. This raises the possibility that at Botai, itself, lifeways changed over time, raising the possibility that forms of husbandry also did. This issue can only be tackled by a new campaign of targeted excavation with an advanced dating programme. Thus, whilst there is a strong argument for a prey pathway domestication leading to specialized horse herding, there is much yet to decipher about the exact form that took and how it changed over time.
Figure 5
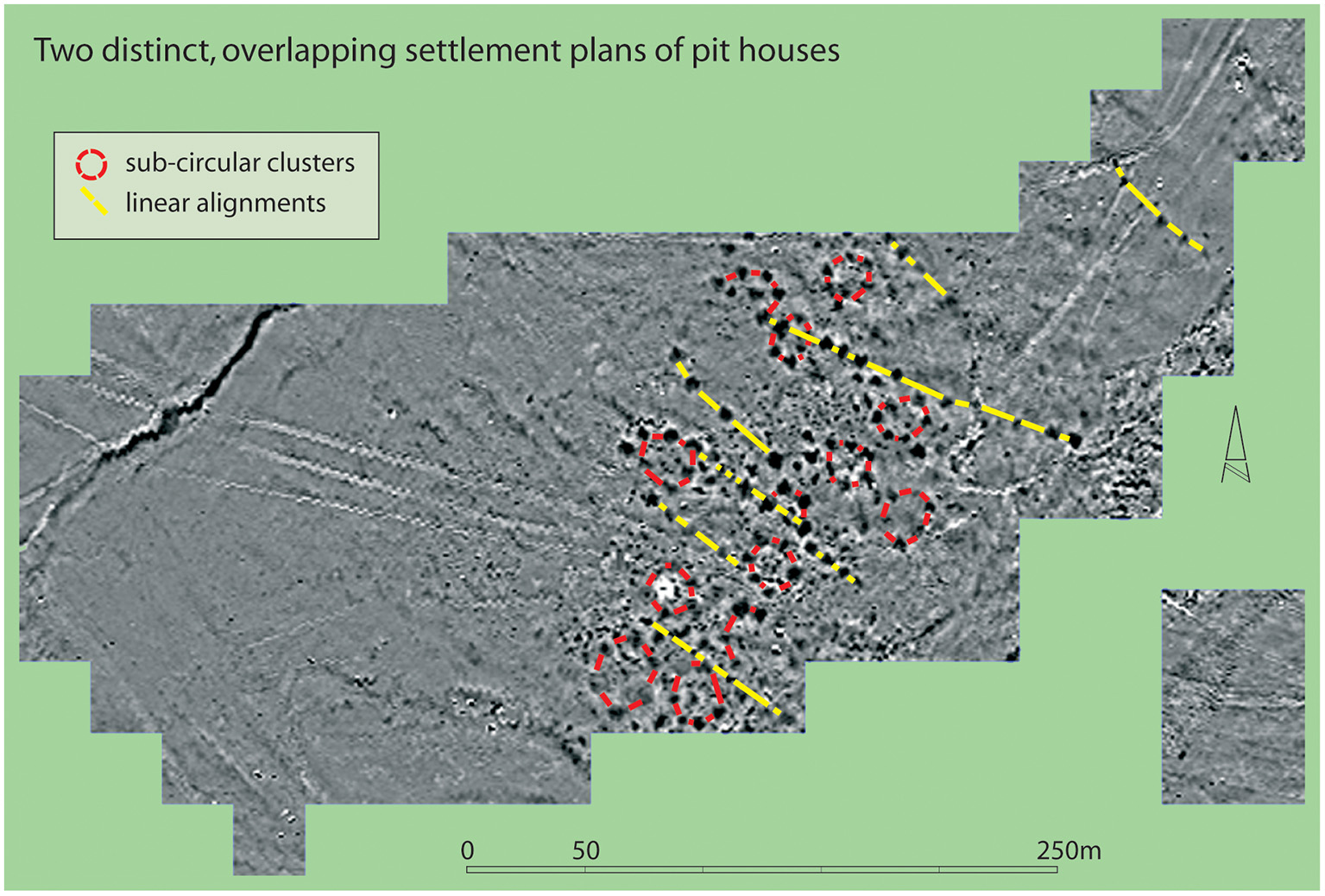
Fluxgate gradiometry at Botai showing different alignments of pit houses and the presence of areas of overlap suggestive of multiple phases of settlement structure.
Their must also have been a local pastoral phase of horse domestication in the Pontic Caspian region in the run-up to selection for DOM2 horses. The exact dating of the first phases is still open for debate. Whilst the famous stallion at Deriivka was later and intrusive, the Copper Age Sredny Stog Culture still presents considerable interest in horses and remains worthy of further consideration (Anthony, 1996, 2007). Lipid residue analysis undertaken at Deriivka (Mileto et al., 2017) demonstrates a preponderance of horse fats in pottery to match the strong representation in faunal assemblages. However, horse milk was not evidenced. Unfortunately, the climate of the region may not produce sufficient seasonal separation in δ2H values to make that method viable at Deriivka (Mileto et al., 2017). The Khvalynsk Culture of the Volga region also presents significant potential for middle Eneolithic horse husbandry in a region where horses are likely ancestral to DOM2 (Anthony et al., 2022). Horse milking is, however, first evidenced in Eastern Europe in two human dental calculus proteomic samples from a Yamnaya site of Pontic-Caspian steppe (Wilkin et al., 2021a), the dating of which is either late Eneolithic or early Bronze Age, depending on aforementioned debates about reservoir effects. Either way, this is in the lead up to DOM2 genetic selection (Librado et al., 2021), likely evidencing an earlier pastoral phase. A further proteomic study in the region (Scott et al., 2022) did not pick up any further examples of such early horse milking, but instead evidenced a focus on sheep milk.
The major difference between the Pontic-Caspian and Northern Kazakhstan at this time is that the Yamnaya had domestic ruminant animals in abundance, and whilst horses are a significant component at many sites (Shishlina, 2003; see Kaiser, 2010), ruminants products would have dominated food supply and associated residues. Since the Yamnaya had very early, heavy-wheeled carts (Anthony, 2007) it is possible horses were involved in traction, though, with slow heavy, heavy vehicles, cattle are a distinct alternative (Librado et al., 2021; Scott et al., 2022). Horses may have been used in much the same way as mixed pastoralists do today, to ride and help herd, provide a limited contribution to diet and clear snow for ruminant animals, which struggle in those conditions (Shishlina, 2003). Significant new evidence has, in fact, just emerged, based upon human entheseal and pathological modifications on a number Yamnaya human skeletons, for the practice of at least some horse riding in the early 3rd. Mill. BCE in Romania, Bulgaria, and Hungary (Trautmann et al., 2023). The difference in emphasis in the Pontic-Caspian likely set the scene for later developments.
5.2. Single center selection for equestrianism and subsequent spread of DOM2
DOM2 horses have their ancestral origins in the Pontic-Caspian steppe (Librado et al., 2021). The ancient genomic evidence suggests that, during the 3rd Mill. BCE, horses ancestral to DOM2 underwent selection for two different phenotypic traits (Librado et al., 2021). Selection associated with the GSDMC gene likely reduced back pathologies, whilst further selection related to the ZFPM1 gene is associated with reduced anxiety and aggressive behavior (Librado et al., 2021). Such selection is suggestive of adaptation to increased use in equestrianism. DOM2 spread rapidly and widely in Eurasia after about 2,000 BCE (Librado et al., 2021), a time synchronous with Trans-Ural Sintashta Culture and the first physical evidence for chariots (Anthony, 2009).
There is considerable ongoing debate (see Kanne, 2022) over whether the spread of DOM2 horses was associated purely with their use with chariots (e.g., Taylor et al., 2020) or whether riding was a regular component of non-elite as well as elite peoples in the early Bronze Age (Kanne, 2022). It is important to note that the genetic selections apparently related to improved use for equestrianism commence prior to any evidence for chariotry (Librado et al., 2021), and arguably the GSDMC selection is perhaps more pertinent to weight bearing on the back than traction. Furthermore, the widespread evidence for DOM2 use in the Bronze Age (Librado et al., 2021) extends into culture and regions where there is no evidence for chariot use, or indeed where the terrain would be less appropriate. Thus, the use of DOM2 for riding from its earliest stages seems most likely, though elite use with chariots likely plays a significant social role in DOM2 spread.
This author would argue for an initial pastoral phase using local horses, which were ancestral to DOM2, in the Pontic-Caspian steppe during the Eneolithic and early Bronze Age. Because of the dominance of domestic ruminant animals in food production, the role of horses was more related to riding to aid herding of ruminant domestic stock, which led to selection for improved equestrian utility. Initial interest in horses in the region may have also been “prey route”, but later selection for equestrianism could have been more “directed” and related to “pull factors”. The elite use of horses, and chariotry, and displays of wealth in funerary monuments speaks to a new cultural role of horses in the Bronze Age world following the Sintashta horizon. Horses were likely involved in the display of wealth and status with increasing involvement in warfare. Evidence for the milking of DOM2 horses is largely absent from later Bronze Age contexts, based upon both lipid residue evidence in Kazakhstan (Outram et al., 2012) or proteomic evidence in the Caucasus or Pontic-Caspian regions (Scott et al., 2022). Use of horses for meat was not a primary subsistence concern in most cases, apart from when consumed at special occasions such as funerary rites (Outram et al., 2011). Horse representation during this period is variable and not correlated well with environmental factors, unlike cattle/caprine ratios, and may relate more to wealth (Outram and Bogaard, 2019, p. 189).
5.3. Ongoing horse domestication processes: the post-medieval agricultural revolution and sport
One could identify a myriad of important developments in the human-equine relationship over time (see Kelekna, 2009). These would include the worldwide diaspora of Eurasian domestic horses to the rest of the world and their impact on native communities (see Mitchell, 2015). A significant consideration in domestication process is hydridization. The hybridization of horses with donkeys produces mules (mare with male donkey) or hinnies (stallion with female donkey). Recent genomic evidence from French material suggests such hybridization was not common in the Iron Age, but the breeding of mules (not hinnies) in the Roman world must have been a significant and highly organized activity, that largely ceases in the early middle ages when unhybridized donkeys become common (Lepetz et al., 2021). Whilst mule production in the Roman period demonstrates highly organized breeding programmes, we also know that in pre-modern, historical contexts there were stud networks, such as those seen for breeding medieval warhorses (Ameen et al., 2021a). From the early medieval period we start to also see the wider introduction of Middle Eastern horse lineages into other regions (Fages et al., 2019), perhaps encouraged through such stud networks.
However, if one is to select a third major turning point in the horse domestication continuum it is in the modern period. Major changes in sizes of horse bones and in genetics are evidenced in the last few 100 years, broadly corresponding with the “agricultural revolution”. This revolution, initially centered in England and other areas of Europe, was approximately concurrent with the industrial revolution and saw major changes in land use strategies, increasingly mechanized farming and a significant interest in animal stock improvement (Allen, 1999; Russell, 2007). Selective breeding at this time surpasses anything seen previously in terms of its physical effect upon the animals and their lineages. Taking as an example a sequence of horse bone metrical analyses from England (Ameen et al., 2021b), one sees radical increase in average horse stature in the post-medieval period, alongside an increased total range of stature. Over the same time period, ancient genomics shows considerable loss of y-chromosome lineages, decreased heterozygosity and strongly increased genetic load (Fages et al., 2019). The effects are all the result of highly selective breeding for desired traits but at the cost of a significant loss of diversity and increased concentration of deleterious genes that can cause disease and malady (Orlando and Librado, 2019). This stage of the domestication process is undoubtedly “directed” and driven by the “pull” factor of increased economic production and the prestige attached to producing prize animals. Such emphasis only increased with breeding programmes for performance sport horses (Todd et al., 2018).
6. Conclusion
Much remains to be debated regarding the detailed sequence of horse domestication, and some geographic regions and time periods possess stronger evidence bases than others. Some aspects of this paper are in need of further empirical research and testing, but are currently viable hypotheses. There remains multi-faceted evidence at Botai, and a range of similar sites in northern Kazakhstan, for specialized horse pastoralism including corralling and use of secondary products. The situation Eastern Europe is more complex due to the additional presence ruminant domestic animals, but there is a good contextual case for changing horse human relationships in the Eneolithic plains of Ukraine, the Volga and Pontic-Caspian steppes of Russia. By the early Bronze Age Yamnaya Culture it appears that horses are in use within a mixed pastoralist regime. Whilst there is now some proteomic evidence horse milking in this region by this date, the significance of horses in this region may have been more focussed upon herding other stock. This greater emphasis on riding likely led to the phenotypic selections seen in the rise of DOM2 horses in this region. There is no evidence for spread of DOM2 lineages within wider Yamnaya expansions, however. The phenotypic selection significantly post-dates the introduction heavy wheeled carts, most likely pulled by oxen, whilst it pre-dates chariots (Librado et al., 2021). The phenotypic selection for reduced susceptibility to back pathology is, perhaps, more suggestive of DOM2 being associated with early riding. The first evidenceable phase of horse domestication is multi-regional and related to local pastoral adaptations. At least at Botai, it is likely a “prey route” process given impetus by subsistence “push” factors.
The selection for DOM2 horses is propelled by the “pull” factor of improved equestrian capability in the Pontic-Caspian region. Its subsequent Eurasian diaspora after c. 2,000 BCE likely involved both riding and chariotry, with the latter speaking to wealth and power. Bronze Age society at this time involved further significant migrations, but also many societal “pull” factors that would have made DOM2 acquisition desirable. These would include trade and warfare advantages as well as display of prestige and status. The horse-human relationship continues to evolve, with some of its most recent effects upon horse lineages and phenotypes being the most dramatic.
Statements
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
The author confirms being the sole contributor of this work and has approved it for publication.
Acknowledgments
I would like to thank Victor Zaibert, who discovered and investigated Botai for over 40 years but sadly passed away while this article was being written. Thanks to Peter Rowley-Conwy for allowing me to cite his personal communication, and every colleague with whom I have discussed horse domestication. Seán Goddard assisted in the creation of the Figure 3 and Viktor Merts kindly granted use of Figure 4. Two reviewers provided very useful comments.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
Allen R. C. (1999). Tracking the agricultural revolution in England. Econ. History Rev. LII, 209–235. 10.1111/1468-0289.00123
2
Allentoft M. E. Sikora M. Sjögren K. G. Rasmussen S. Rasmussen M. Stenderup J. et al . (2015). Population genomics of bronze age eurasia. Nature.522, 167–172. 10.1038/nature14507
3
Ameen C. Baker G. P. Benkert H. Van Qui C. V. Webley R. Liddiard R. et al . (2021a). Interdisciplinary Approaches to the Medieval Warhorse. Cheiron Int J Equine Equestrian History. 1, 233005. 10.22618/TP.Cheiron.20211.1.233005
4
Ameen C. Benkert H. Fraser T. Gordon R. Holmes M. Johnson W. et al . (2021b). In search of the “great horse”: a zooarchaeological assessment of horses from England (AD 300–1650). Int. J. Osteoarchaeol.31, 1247–1257. 10.1002/oa.3038
5
Anthony D. Telegin D. Y. Brown D. (1991). The origin of horseback riding. Sci Am.265, 94–100. 10.1038/scientificamerican1291-94
6
Anthony D. W. (1996). “Bridling horse power: the domestication of the horse”, in Horses through Time, ed. S. Olsen (Boulder: Roberts Rinehart), p. 57–82.
7
Anthony D. W. (2007). The Horse, the Wheel, and Language: How Bronze-Age Riders from the Eurasian Steppes Shaped the Modern World. Princeton: Princeton University Press. 10.1515/9781400831104
8
Anthony D. W. (2009). “The Sintashta Genesis: the roles of climate change, warfare, and long-distance trade,” in Social Complexity in Prehistoric Eurasia. eds B.K. Hanks, K.M. Linduff (Cambridge: Cambridge University Press), p. 47–73. 10.1017/CBO9780511605376.005
9
Anthony D. W. Brown D. R. (1991). The origins of horseback riding. Antiquity.65, 22–38. 10.1017/S0003598X00079278
10
Anthony D. W. Brown D. R. (2000). Eneolithic horse exploitation in the Eurasian steppes: diet, ritual and riding. Antiquity.74, 75–86. 10.1017/S0003598X00066163
11
Anthony D. W. Brown D. R. (2011). The secondary products revolution, horse-riding, and mounted warfare. J. World Prehistory24, 131–160. 10.1007/s10963-011-9051-9
12
Anthony D. W. Brown D. R. George C. (2006). “Early horseback riding and warfare: the importance of the magpie around the neck”, in Horses and humans: the evolution of human-equine relationships, eds S.L. Olsen, S. Grant, A.M. Choyke, L. Bartosiewicz (Oxford Archaeopress), p. 137–156.
13
Anthony D. W. Khokhlov A. A. Agapov S. A. Agapov D. S. Schulting R. Olalde I. et al . (2022). The Eneolithic cemetery at Khvalynsk on the Volga River. Praehistorische Zeitschrift. 97:22–67. 10.1515/pz-2022-2034
14
Anthony D. W. Ringe D. (2015). The Indo-European homeland from linguistic and archaeological perspectives. Ann. Rev. Linguistics1, 199–219. 10.1146/annurev-linguist-030514-124812
15
Bahn P. G. Vertut J. (1997). Journey Through the Ice Age. Berkeley: University of California Press.
16
Bellone R. R. Holl H. Setaluri V. Devi S. Maddodi N. Archer S. et al . (2013). Evidence for a retroviral insertion in TRPM1 as the cause of congenital stationary night blindness and leopard complex spotting in the horse. PLoS ONE8, e78280. 10.1371/journal.pone.0078280
17
Bendrey R. (2007). New methods for the identification of evidence for bitting on horse remains from archaeological sites. J Archaeol Sci.34, 1036–1050. 10.1016/j.jas.2006.09.010
18
Bendrey R. (2011a). Some like it hot: environmental determinism and the pastoral economies of the later prehistoric Eurasian steppe. Pastoralism Res. Policy Practice1, 1–16. 10.1186/2041-7136-1-8
19
Bendrey R. (2011b). Identification of metal residues associated with bit-use on prehistoric horse teeth by scanning electron microscopy with energy dispersive X-ray microanalysis. J Archaeol Sci.38, 2989–2994. 10.1016/j.jas.2011.06.016
20
Bendrey R. (2012). From wild horses to domestic horses: a European perspective. World Archaeol.44, 135–157. 10.1080/00438243.2012.647571
21
Benecke N. (1994). “Zur Domestikation des Pferdes in Mittel- und Osteuropa: Einige neue archäozoologische Befunde,” in Die Indogermanen und das pferd, eds B. Hansel, S. Zimmer S. (Budapest: Archaeolingua Alapitvány), p. 123–44.
22
Benecke N. von den Driesch A. (2003). “Horse exploitation in the Kazakh steppes during the Eneolithic and Bronze Age”, in Prehistoric Steppe Adaptation and the Horse, eds M. Levine, C. Renfrew, K. Boyle (Cambridge: McDonald Institute), p. 69–82.
23
Blumler M. A. Byrne R. (1991). The ecological genetics of domestication and the origins of agriculture. Curr Anthropol.32, 23–35. 10.1086/203912
24
Bogaard A. Allaby R. Arbuckle B. S. Bendrey R. Crowley S. Cucchi T. et al . (2021). Reconsidering domestication from a process archaeology perspective. World Archaeol.53, 56–77. 10.1080/00438243.2021.1954990
25
Bouckaert R. Lemey P. Dunn M. Greenhill S. J. Alekseyenko A. V. et al . (2012). Mapping the origins and expansion of the Indo-European language family. Science.337, 957–60. 10.1126/science.1219669
26
Brown D. Anthony D. (1998). Bit wear, horseback riding and the Botai site in Kazakhstan. J Archaeol Sci.25, 331–347. 10.1006/jasc.1997.0242
27
Chechushkov I. V. Kosintsev P. A. (2020). The Botai horse practices represent the neolithization process in the central Eurasian steppes: Important findings from a new study on ancient horse DNA. J. Archaeol. Sci. Rep.32, 102426. 10.1016/j.jasrep.2020.102426
28
Clutton-Brock J. (1999). A Natural History of Domestic Mammals (2nd edition). Cambridge: Cambridge University Press.
29
Daly K. G. Arbuckle B. S. Rossi C. Mattiangeli V. Lawlor P. A. Mashkour M. et al . (2022). A novel lineage of the Capra genus discovered in the Taurus Mountains of Turkey using ancient genomics. bioRxiv. https://doi.org/10.1101/04,08.48761910.1101/2022.04.08.487619
30
Damgaard P. D. B. Marchi N. Rasmussen S. Peyrot M. Renaud G. Korneliussen T. et al . (2018). 137 ancient human genomes from across the Eurasian steppes. Nature.557, 369–374. 10.1038/s41586-018-0094-2
31
de Barros Damgaard P. Martiniano R. Kamm J. Moreno-Mayar J. V. Kroonen G. Peyrot M. et al . (2018). The first horse herders and the impact of early Bronze Age steppe expansions into Asia. Science. 360, eaar7711. 10.1126/science.aar7711
32
Dudd S. N. Evershed R. P. (1998). Direct demonstration of milk as an element of archaeological economies. Science.282, 1478–1481. 10.1126/science.282.5393.1478
33
Dyson-Hudson R. Smith E. A. (1978). Human territoriality: an ecological reassessment. Am Anthropol.80, 21–41. 10.1525/aa.1978.80.1.02a00020
34
Fages A. Hanghøj K. Khan N. Gaunitz C. Seguin-Orlando A. Leonardi M. et al . (2019). Tracking five millennia of horse management with extensive ancient genome time series. Cell.177, 1419–1435. 10.1016/j.cell.2019.03.049
35
Fijn N. (2011). Living with herds: human-animal coexistence in Mongolia. Cambridge: Cambridge University Press. 10.1017/CBO9780511976513
36
Frachetti M. D. (2008). Pastoralist Landscapes and Social Interaction in Bronze Age Eurasia. Berkeley: University of California Press. 10.1525/9780520942691
37
Frachetti M. D. Spengler R. N. Fritz G. J. Mar'yashev A. N. (2010). Earliest direct evidence for broomcorn millet and wheat in the central Eurasian steppe region. Antiquity.84, 993–1010. 10.1017/S0003598X0006703X
38
French C. Kousoulakou M. (2003). “Geomorphological and micromorphological investigations of palaeosols, valley sediments and a sunken floored dwelling at Botai, Kazakhstan”, in Prehistoric Steppe Adaptation and the Horse, eds M. A. Levine, C. Renfrew, K. V. Boyle (Cambridge: McDonald Institute), p. 105–114.
39
Gamkrelidze T. V. Ivanov V. (1995). Indo-European and the Indo-Europeans: A Reconstruction and Historical Analysis of a Proto-Language and a Proto-Culture, Vol. 1. Berlin: Mouton de Gruyter. 10.1515/9783110815030
40
Gaunitz C. Fages A. Hanghøj K. Albrechtsen A. Khan N. Schubert M. et al . (2018). Ancient genomes revisit the ancestry of domestic and Przewalski's horses. Science360, 111–114. 10.1126/science.aao3297
41
Haak W. Lazaridis I. Patterson N. Rohland N. Mallick S. Llamas B. et al . (2015). Massive migration from the steppe was a source for Indo-European languages in Europe. Nature.522, 207–211. 10.1038/nature14317
42
Haruda A. (2018). Regional pastoral practice in central and southeastern Kazakhstan in the Final Bronze Age (1300–900 BCE). Archaeol. Res. Asia15, 146–156. 10.1016/j.ara.2017.09.004
43
Higgs E. S. Jarman M. R. (1975). “Palaeoeconomy”, in Palaeoeconomy: Being the second volume of Papers in Economic Prehistory by members and associates of the British Academy major research project in the early history of agriculture, eds. E.S. Higgs (Cambridge: Cambridge University Press), p. 1–8.
44
Hollund H. I. Higham T. Belinskij A. Korenevskij S. et al . (2010). Investigation of palaeodiet in the North Caucasus (South Russia) Bronze Age using stable isotope analysis and AMS dating of human and animal bones. J Archaeol Sci.37, 2971–83. 10.1016/j.jas.2010.08.009
45
Ingold T. (1980). Hunter, Pastoralists and Ranchers. Cambridge: Cambridge University Press. 10.1017/CBO9780511558047
46
Itahashi Y. Ananyevskaya E. Yoneda M. Miller A. R. V. Nishiaki Y. Matuzeviciute G. M. et al . (2020). Dietary diversity of Bronze-Iron Age populations of Kazakhstan quantitatively estimated through the compound-specific nitrogen analysis of amino acids. J. Archaeol. Sci. Rep.33, 102565. 10.1016/j.jasrep.2020.102565
47
Jeong C. Balanovsky O. Lukianova E. Kahbatkyzy N. Flegontov P. Zaporozhchenko V. et al . (2019). The genetic history of admixture across inner Eurasia. Nat. Ecol. Evol.3, 966–976. 10.1038/s41559-019-0878-2
48
Kaiser E. (2010). Der Übergang zur Rinderzucht im nördlichen Schwarzmeerraum. Godišnjak Centra za balkanološka ispitivanja39, 23–34.
49
Kalieva S. S. Logvin V. N. (1997). Skotovody Turgaya v Tret'em Tysyacheletii do Nashej Ehry. Kustanai: Kustanai University.
50
Kanne K. (2022). Riding, ruling, and resistance equestrianism and political authority in the hungarian bronze age. Curr Anthropol.63, 239–366. 10.1086/720271
51
Kelekna P. (2009). The Horse in Human History. Cambridge: Cambridge University Press.
52
Kislenko A. Tatarintseva N. (1999). “The eastern Ural steppe at the end of the Stone Age”, in Late Prehistoric Exploitation of the Eurasian Stepp, eds M. Levine, Y. Rassamakin, A. Kislenko, N. Tatarintseva (Cambridge: McDonald Institute), p. 183–216.
53
Kovtun I. V. (2013). Predystoriya indoariyskoy mifologii Rossiyskaya akad. nauk, Sibirskoye otd-niye, In-t Ekologii Cheloveka. Kemerovo: Aziya-Print.
54
Kremenetski C. V. Tarasov P. E. Cherkinsky A. E. (1997). Postglacial development of Kazakhstan pine forests. Geographie physique et Quaternaire51, 391–404. 10.7202/033138ar
55
Kuzmina I. E. (1997). Horses of North Eurasia from the Pliocene till the present time. St. Petersburg: Zoological Institute of the Russian Academy of Sciences (In Russian).
56
Larson G. Albarella U. Dobney K. Rowley-Conwy P. Schibler J. et al . (2007). Ancient DNA, pig domestication, and the spread of the Neolithic into Europe. Proc. Natl. Acad. Sci. USA104, 15276–15281. 10.1073/pnas.0703411104
57
Lavrillier A. (2011). “The creation and persistence of cultural landscapes among Siberian Evenkis: two concepts of sacred space”, in Landscape and Culture in Northern Eurasia, eds. P. Jorden (Walnut Creek: Left Coast Press), p. 215–34.
58
Leonardi M. Boschin F. Giampoudakis K. Beyer R. M. Krapp M. Bendrey R. et al . (2018). Late Quaternary horses in Eurasia in the face of climate and vegetation change. Sci. Adv. 4, eaar5589. 10.1126/sciadv.aar5589
59
Lepetz S. Clavel B. Alioglu D. Chauvey L. Schiavinato S. Tonasso-Calvière L. et al . (2021). Historical management of equine resources in France from the Iron Age to the Modern Period. J. Archaeol. Sci. Rep.40, 103250. 10.1016/j.jasrep.2021.103250
60
Levine M. (2004). “Exploring the criteria for early horse domestication”, in Traces of Ancestry: studies in honour of Colin Renfrew, ed M. Jones (Cambridge, McDonald Institute), p. 115–26. 10.3828/bfarm.2004.3.6
61
Levine M. A. Whitwell K. E. Jeffcott L. B. (2005). Abnormal thoracic vertebrae and the evolution of horse husbandry. Archaeofauna Int. J. Archaeozool.14, 93–109.
62
Levine M. A. (2000). “Domestication and early history of the horse”, in The Domestic Horse: The Origins, Development and Management of its Behavior, eds D. Mills, S. McDonnell, (Cambridge: Cambridge University Press), p. 5–22.
63
Librado P. Khan N. Fages A. Kusliy M. A. Suchan T. Tonasso-Calvière L. et al . (2021). The origins and spread of domestic horses from the Western Eurasian steppes. Nature., 598, 634–640. 10.1038/s41586-021-04018-9
64
Lightfoot E. Motuzaite-Matuzeviciute G. O'Connell T. C. Kukushkin I. A. Loman V. Varfolomeev V. et al . (2014). How ‘pastoral' is pastoralism? Dietary diversity in bronze age communities in the central kazakhstan steppes. Archaeometry. 57, 232–249. 10.1111/arcm.12123
65
Ludwig A. Reissmann M. Benecke N. Bellone R. Sandoval-Castellanos E. Cieslak M. et al . (2015). Twenty-five thousand years of fluctuating selection on leopard complex spotting and congenital night blindness in horses. Phil. Trans. R. Soc. B370, 20130386. 10.1098/rstb.2013.0386
66
MacArthur R. H. Pianka E. R. (1966). On optimal use of a patchy environment. Am. Nat.100, 603–609. 10.1086/282454
67
MacHugh D. E. Larson G. Orlando L. (2017). Taming the past: ancient DNA and the study of animal domestication. Ann. Rev. Animal Biosci.5, 329–351. 10.1146/annurev-animal-022516-022747
68
Mallory J. P. (1996). In Search of the Indo-Europeans: Language, Archaeology and Myth. London: Thames and Hudson.
69
Matuzeviciute G. M. Lightfoot E. O'Connell T. C. Voyakin D. Liu X. Loman V. et al . (2015). The extent of cereal cultivation among the Bronze Age to Turkic period societies of Kazakhstan determined using stable isotope analysis of bone collagen. J. Archaeol. Sci.59, 23–34. 10.1016/j.jas.2015.03.029
70
Meadow R. H. (1984). “Animal domestication in the Middle East: a view from the eastern margin”, in Animals in archaeology. Volume 3. Early herders and their flocks, eds J. Clutton-Brock, C. Grigson (Oxford: British Archaeological Reports), p. 309–37.
71
Merts I. V. Merts V. K. (2018). Eneoliticheskiy kompleks poseleniya Borly 4, Chelovek i Sever: Antropologiya, arkheologiya, ekologiya: materialy vserossiyskoy nauchnoy konferentsii, g. Tyumen', 2-6 aprelya, Tyumen': FITS TyumNTS SO RAN, Vyp. 4. S. 160-164.
72
Merts V. K. (2016). Novaya nakhodka navershiya Seyminsko-Turbinskogo nozha iz Vostochnogo Kazakhstana. Mir Bol'shogo Altaya2, 673–78.
73
Mileto S. Kaiser E. Rassamakin Y. Evershed R. P. (2017). New insights into the subsistence economy of the Eneolithic Dereivka culture of the Ukrainian North-Pontic region through lipid residues analysis of pottery vessels. J. Archaeol. Sci. Rep.13, 67–74. 10.1016/j.jasrep.2017.03.028
74
Mitchell P. (2015). Horse Nations: The Worldwide Impact of the Horse on Indigenous Societies Post-1492. Oxford: Oxford University Press. 10.1093/oso/9780198703839.001.0001
75
Molodin V. I. Neskorov A.V. (2010). Kollektsiya Seyminsko-Turbinskikh Bronz iz Priirtysh'ya (tragediya unikal'nogo pamyatnika-posledstviya bugrovshchichestva XXI veka). Arkheologiya, Etnografiya i Antropologiya Yevrazii3, 58–71. 10.1016/j.aeae.2010.10.006
76
Motuzaite Matuzeviciute G. Lightfoot E. Liu X. Jacob J. Outram A. K. Zaibert V. F. et al . (2019). Archaeobotanical investigations at the earliest horse herder site of Botai in Kazakhstan. Archaeol. Anthropol. Sci.11, 6243–6258. 10.1007/s12520-019-00924-2
77
Naito Y. I. Bocherens H. Chikaraishi Y. Drucker D. G. Wißing C. Yoneda M. et al . (2016). An overview of methods used for the detection of aquatic resource consumption by humans: compound-specific delta N-15 analysis of amino acids in archaeological materials. J. Archaeol. Sci. Rep.6, 720–732. 10.1016/j.jasrep.2015.11.025
78
Ní Leathlobhair M. Perri A. R. Irving-Pease E. K. Witt K. E. Linderholm A. Haile J. et al . (2018). The evolutionary history of dogs in the Americas. Science.361, 81–85. 10.1126/science.aao4776
79
O'Connell T. C. Levine M. A. Hedges R. E. M. (2003). “The importance of fish in the diet of central Eurasian peoples from the Mesolithic to the Early Iron Age”, in Prehistoric Steppe Adaptation and the Horse, eds M. Levine, C. Renfrew, K. Boyle (Cambridge: McDonald Institute), p. 253–68.
80
Olsen S. L. (1989). Solutré: a theoretical approach to the reconstruction of Upper Palaeolithic hunting strategies. J Hum Evol.18, 295–327. 10.1016/0047-2484(89)90034-1
81
Olsen S. L. (1996). “Horse hunters of the Ice Age”, in Horses through Time, ed. Olsen, S. (Boulder: Roberts Rinehart), p. 35–56.
82
Olsen S. L. (2006a). “Early horse domestication: weighing the evidence”, in Horses and humans: the evolution of human-equine relationships, eds S.L. Olsen, S. Grant, A.M. Choyke, L. Bartosiewicz (Oxford: Archaeopress), p. 81–113. 10.30861/9781841719900
83
Olsen S. L. (2006b). “Early horse domestication on the Eurasian Steppe”, in Documenting Domestication: New Genetic and Archaeological Paradigms, eds M.A. Zeder, D.G. Bradley, E. Emschwiller, B.D. Smith (Berkley: University of California Press), p. 245–69. 10.1525/9780520932425-020
84
Olsen S. L. Bradley B. Maki D. Outram A. (2006). “Community organization among Copper Age sedentary horse pastoralists of Kazakhstan”, in Beyond the steppe and the sown: Proceedings of the 2002 University of Chicago Conference on Eurasian Archaeology, eds D.L. Peterson, L. M. Popova, A. T Smith (Leiden: Brill), p. 89–111. 10.1163/9789047408215_009
85
Orlando L. Librado P. (2019). Origin and evolution of deleterious mutations in horses. Genes., 10, 649. 10.3390/genes10090649
86
Outram A. Rowley-Conwy P. (1998). Meat and marrow utility indices for horse (Equus). J. Archaeol. Sci.25, 839–849. 10.1006/jasc.1997.0229
87
Outram A. K. (2006). “Juggling with indices: a review of the evidence and interpretations regarding Upper Palaeolithic horse skeletal part abundances”, in Horses and humans: the evolution of human-equine relationships, eds S. L. Olsen, S. Grant A. M. Choyke, L. Bartosiewicz (Oxford: Archaeopress), p. 49–60.
88
Outram A. K. (2014). “Animal Domestications”, in Oxford Handbook of the Archaeology and Anthropology of Hunter-Gatherers, eds V. Cummings, P. Jordan, M. Zvelebil, M. (Oxford: Oxford University Press), p. 749–763.
89
Outram A. K. (2015). “Pastoralism”, in The Cambridge World History, Volume II: A World with Agriculture, 12, 000. BCE - 500 CE, eds G. Barker, C. Goucher (Cambridge: Cambridge University Press), p. 161–185. 10.1017/CBO9780511978807.007
90
Outram A. K. Bendrey R. Evershed R. P. Orlando L. Zaibert V. (2021). Rebuttal of Taylor and Barron-Ortiz 2021 Rethinking the evidence for early horse domestication at Botai. Zenodo. 10.5281./zenodo.5142604
91
Outram A. K. Bogaard A. (2019). Subsistence and Society in Prehistory: New Directions in Economic Archaeology. Cambridge: Cambridge University Press. 10.1017/9781316415177
92
Outram A. K. Kasparov A. Stear N. A. Varfolomeev V. Usmanova E. Evershed R. P. et al . (2012). Patterns of pastoralism in later Bronze Age Kazakhstan: new evidence from faunal and lipid residue analyses. J. Archaeol. Sci.39, 2424–2435. 10.1016/j.jas.2012.02.009
93
Outram A. K. Stear N. A. Bendrey R. Olsen S. Kasparov A. Zaibert V. et al . (2009). The earliest horse harnessing and milking. Science.323, 1332–1335. 10.1126/science.1168594
94
Outram A. K. Stear N. A. Kasparov A. Usmanova E. Varfolomeev V. Evershed R. P. et al . (2011). Horses for the dead: funerary foodways in Bronze Age Kazakhstan. Antiquity.85, 116–128. 10.1017/S0003598X00067478
95
Owen D. I. (1991). The first equestrian: an Ur III glyptic scene. Acta Sumerologica. 13, 259–273.
96
Payne S. (1973). Kill off patterns in sheep and goats: the mandibles from Asvan Kale. Anatolian Stud.23, 281–303. 10.2307/3642547
97
Perkins D. Daly P. (1968). A hunters' village in Neolithic Turkey. Sci Am.219, 96–109. 10.1038/scientificamerican1168-96
98
Piggott S. (1992). Wagon, Chariot and Carriage. London: Thames and Hudson.
99
Pruvost M. Bellone R. Benecke N. Sandoval-Castellanos E. Cieslak M. Kuznetsova T. et al . (2011). Genotypes of predomestic horses match phenotypes painted in Paleolithic works of cave art. Proc Nat Acad Sci.108, 18626–630. 10.1073/pnas.1108982108
100
Redding R. W. (2005). “Breaking the mold: a consideration of variation in the evolution of animal domestication”, in The first steps of animal domestication, eds J.-D Vigne, J. Peters, D. Helmer (Oxford: Oxbow), p. 41–8.
101
Renfrew C. (1987). Archaeology and Language: The Puzzle of Indo-European Origins. London: Jonathan Cape.
102
Russell N. (2007). Like engend'ring like: heredity and animal breeding in early modern England. Cambridge: Cambridge University Press.
103
Scott A. Reinhold S. Hermes T. Kalmykov A. A. Belinskiy A. Buzhilova A. et al . (2022). Emergence and intensification of dairying in the Caucasus and Eurasian steppes. Nat. Ecol. Evol.6, 813–822. 10.1038/s41559-022-01701-6
104
Shishlina N. (2003). “Yamnaya culture pastoral exploitation: a local sequence”, in Prehistoric Steppe Adaptation and the Horse, eds M. Levine, C. Renfrew, K. Boyle (Cambridge: McDonald Institute), p. 353–65.
105
Svyatko S. V. Mertz I. V. Reimer P. J. (2015). Reservoir effect on re-dating of eurasian steppe cultures: first results for eneolithic and early bronze Age North-East Kazakhstan. Radiocarbon.57, 625–644. 10.2458/azu_rc.57.18431
106
Taylor W. T. T. Barrón-Ortiz C. I. (2021). Rethinking the evidence for early horse domestication at Botai. Sci Rep.11, 1–11. 10.1038/s41598-021-86832-9
107
Taylor W. T. T. Clark J. Bayarsaikhan J. Tuvshinjargal T. Jobe J. T. Fitzhugh W. et al . (2020). Early pastoral economies and herding transitions in Eastern Eurasia. Sci Rep.10, 1–15. 10.1038/s41598-020-57735-y
108
Todd E. T. Ho S. Y. Thomson P. C. Ang R. A. Velie B. D. Hamilton N. A. et al . (2018). Founder-specific inbreeding depression affects racing performance in Thoroughbred horses. Sci Rep.8, 1–10. 10.1038/s41598-018-24663-x
109
Trautmann M. Frînculeasa A. Preda-Bălănică B. Petruneac M. Focşăneanu M. Alexandrov S. et al . (2023). First bioanthropological evidence for Yamnaya horsemanship. Sci. Adv. 9, eade2451. 10.1126/sciadv.ade2451
110
Van de Noort R. (2006). Argonauts of the North Sea-a social maritime archaeology for the 2nd millennium BC. Proceed. Prehistoric Soc.72, 267–287. 10.1017/S0079497X00000852
111
Vigne J-. D. Helmer D. Peters J. (2005). “New archaeozoological approaches to trace the first steps of animal domestication: general presentation, reflections and proposals”, in The first Steps of Animal Domestication, eds J.-D. Vigne, J. Peters, D. Helmer (Oxford: Oxbow), p. 1–16.
112
Webb S. D. Hemmings C. A. (2006). “Last horses and first humans in North America”, in Horses and Humans: The Evolution of Human-Equine Relationships, eds S.L. Olsen, S. Grant A.M. Choyke, L. Bartosiewicz (Oxford: Archaeopress), p. 11–23.
113
Wendrich W. Barnard H. (2008). “The archaeology of mobility: definitions and research approaches”, in The Archaeology of Mobility: Old World and New World Nomadism, eds H. Barnard, W. Wendrich (Los Angeles: Cotsen Institute, University of California), p. 1–24. 10.2307/j.ctvdjrq8t.3
114
West D. (2006). “Horse hunting in Central Europe at the end of the Pleistocene”, in Horses and Humans: the Evolution of Human-Equine Relationships, eds S.L. Olsen, S. Grant A.M. Choyke, L. Bartosiewicz (Oxford: Archaeopress), p. 25–47.
115
Wilkin S. Ventresca Miller A. Fernandes R. Spengler R. Taylor W. T. T. Brown D. R. et al . (2021a). Dairying enabled early bronze age yamnaya steppe expansions. Nature.598, 629–633. 10.1038/s41586-021-03798-4
116
Wilkin S. Ventresca Miller A. Taylor W. T. Miller B. K. Hagan R. W. Bleasdale M. et al . (2021b). Dairy pastoralism sustained eastern Eurasian steppe populations for 5,000 years. Nat. Ecol. Evol.4, 346–355. 10.1038/s41559-020-1120-y
117
Zaibert V. F. (2009). Botaiskaya Kultura. Almaty: KazAkparat.
118
Zeder M. A. (2012). The domestication of animals. J. Anthropol. Res.68, 161–190. 10.3998/jar.0521004.0068.201
119
Zeder M. A. (2015). Core questions in domestication research. Proc. Nat. Acad. Sci.112, 3191–3198. 10.1073/pnas.1501711112
Summary
Keywords
horse, Botai, domestication, pastoralism, equestrianism, Kazakhstan
Citation
Outram AK (2023) Horse domestication as a multi-centered, multi-stage process: Botai and the role of specialized Eneolithic horse pastoralism in the development of human-equine relationships. Front. Environ. Archaeol. 2:1134068. doi: 10.3389/fearc.2023.1134068
Received
29 December 2022
Accepted
04 April 2023
Published
24 April 2023
Volume
2 - 2023
Edited by
Cheryl Makarewicz, University of Kiel, Germany
Reviewed by
Benjamin Arbuckle, University of North Carolina at Chapel Hill, United States; Yimin Yang, University of Chinese Academy of Sciences, China
Updates
Copyright
© 2023 Outram.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alan K. Outram a.k.outram@exeter.ac.uk
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.