Abstract
The Late Bronze Age saw the rise of a new site type in southern Britain, commonly termed a midden. These vast monumental mounds, some comprising tens of thousands of artefacts/ecofacts dominated by animal bone and ceramics, signal a societal refocus towards agricultural intensification and communal feasting on a grand scale. These sites point to agricultural produce being the mainstay of the economy, with bronze having a reduced social and economic importance. This likely created new regimes of managing landscapes and livestock and new networks and agricultural catchments anchored on the sites. Major questions remain surrounding the strategies employed to enhance agricultural productivity, how landscapes and livestock were managed, and how different sites and regions met these challenges at a time of climatic deterioration. This article employs multi-isotope analyses on domestic and wild fauna to address these questions. It presents carbon, nitrogen, and sulfur (δ13C, δ15N, and δ34S, respectively) isotope analyses on 235 animals from 6 midden sites in 2 major regions of midden accumulation: the Vale of Pewsey in Wiltshire and the Thames Valley. The results demonstrate distinct differences in approaches to maximising agricultural productivity, with varied strategies apparent at a site, species, and regional level. Some sites, such as Potterne and Runnymede, clearly drew domestic fauna from a wide catchment where wide-ranging management and foddering regimes were employed. Other sites, such as East Chisenbury, had a more restricted catchment but a tightly controlled, intensive management regime. These data provide new insights into regional responses to the reorganisation of the economy, landscape use, and developing agricultural networks during the Bronze Age–Iron Age transition in Britain, revealing dynamic and evolving societal change.
1. Introduction
During the Late Bronze Age and Early Iron Age (ca. 900–500 bce), major shifts in European society occurred. Long-standing networks surrounding copper extraction, bronze production, and artefact manufacture and exchange started to dissolve at a time of climatic deterioration, with higher precipitation (Needham, 2007; Armit et al., 2014; Turney et al., 2016). This has been suggested to constitute an economic crisis (Needham, 2007; Sharples, 2010) with the status of bronze changing (Roberts et al., 2015). In southern Britain, society shifted its focus to agricultural intensification and communal feasting on a grand scale, producing a new site type commonly termed a midden (Needham and Spence, 1996). These middens constitute vast mounds of material culture, sometimes comprising hundreds of thousands of artefacts dominated by animal bone and pottery. Previous research has proposed structure in depositional practices rather than mere waste accumulation (McOmish, 1996; Madgwick and Mulville, 2015a,b; Madgwick, 2016). Defining what constitutes a midden of this type is not straightforward, but broadly comparable accumulations have been identified as far north as Lincolnshire and as far west as south Wales and Devon. Two main epicentres of activity are Wiltshire, centring on the Vale of Pewsey and the Thames Valley. These new sites of deposition seem to replace the previous focus on depositions of bronze objects (Tubb, 2011), with an accumulation of debris that signals communal consumption, and social mobilisation on a scale that is arguably unprecedented in British prehistory.
The rise of midden sites indicates a shift in societal focus away from metalwork and towards agricultural production, suggestive of intensification and a desire to maximise productivity to support ostentatious feasts (Tubb, 2011). That said, researchers have also argued that middens were intimately connected to the production and circulation of metalwork (Waddington and Sharples, 2011). The suggestion of agricultural intensification, however, remains a hypothesis, and very little research has been conducted on how the agricultural economy changed and how animals were managed. Valuable zooarchaeological research has provided some information on husbandry regimes (e.g., Serjeantson, 1996, 2007; Locker, 2000; Serjeantson et al., 2010; Madgwick, 2011), and taphonomic work on faunal remains has helped elucidate the scale, rhythm, and frequency of feasting events (Madgwick, 2014, 2016; Madgwick and Mulville, 2015b). The use of carbon (δ13C) and nitrogen (δ15N) isotope analysis of animal bone collagen can provide valuable information for better understanding husbandry practices in this transitional phase, and these data are even more informative when combined with sulfur (δ34S) isotopes. However, very little stable and radiogenic isotope research has been published to explore animal management, with only limited studies focusing on carbon and nitrogen isotopes at the sites of Potterne, Llanmaes, and East Chisenbury, including a very small number of strontium values (Madgwick et al., 2012a,b; Madgwick and Mulville, 2015a; Andrews et al., 2021). Agricultural intensification may have necessitated novel and diverse strategies at a regional level to maximise landscape resources. Alternatively, standardised approaches may have been employed, with animals having to have been raised in a certain way to be deemed suitable for these events, just as they were processed in a prescribed manner in some instances (Madgwick and Mulville, 2015a). As part of the Arts and Humanities Research Council (AHRC)-funded FEASTNET (Feasting Networks and Resilience at the end of the British Bronze Age) project, this research explores approaches to husbandry and animal management at an inter-site and inter-regional level focusing on Wiltshire and the Thames Valley. This is achieved by employing three isotope approaches on faunal bone (and occasionally dentine) collagen: carbon, nitrogen, and sulfur.
1.1. Midden sites in Wiltshire and the Thames Valley
Sites with similar characteristics, including comparable dark, organic soils, and extensive deposits, including substantial ceramic and animal bone assemblages, emerged in southern Britain from around the 9th century bce. The exact timing of their development and use remains poorly understood due to the so-called Hallstatt plateau in the radiocarbon calibration curve, although Bayesian modelling has improved chronological resolution (Waddington et al., 2019). These sites appear to represent a similar phenomenon, which emphasises the social arena of feasting through significant deposition of animal bones, objects, and artefacts representative of conspicuous consumption (Tubb, 2011). Deciphering the symbolic meaning of these accumulations and the role of the feasts that created them is complex, but suggestions have included galvanising and reorganising social relations at a time of crisis (Sharples, 2010; Madgwick and Mulville, 2015a) and a close connection to fertility and the success of the agricultural and pastoral cycle (McOmish, 1996). The continuity of practice and dark anthropogenic soils are common features of middens, but the composition, character, and nature of deposition are diverse both within and between regions. This research explores six sites across two major regions of midden accumulation. The locations of sites that are analysed in this study are presented in Figure 1. The largest concentration of middens is in the Vale of Pewsey, Wiltshire, in central southern England. Two of the largest and richest middens in England are analysed from this region—East Chisenbury and Potterne, along with two further sites that have produced more modest assemblages: All Cannings Cross and Stanton St Bernard. These sites are in close proximity, all within 15 km of each other. Two sites are also analysed from the Thames Valley, although these are approximately 45 km apart. Runnymede in the county of Surrey in south-east England, is exceptionally artefact-rich. Whitecross Farm, Wallingford, Oxfordshire, in central southern England is more modest in terms of its artefactual assemblages. Summaries of the excavation histories, artefactual and fauna assemblages, and the general character of each midden are provided in the Supplementary material along with a summary of comparative chronology.
Figure 1

Map of the midden sites analysed in this article. In Wiltshire: All Cannings Cross, East Chisenbury, Potterne, and Stanton St Bernard. Midden sites in the Thames Valley: Runnymede and Whitecross Farm, Wallingford.
1.2. Carbon, nitrogen, and sulfur isotope analysis
Carbon (δ13C) and nitrogen (δ15N) stable isotope analyses have long been used for reconstructing diet, faunal management regimes, and landscape use (e.g., Hamilton and Thomas, 2012; Gillis et al., 2013; Madgwick et al., 2013; Stevens et al., 2013). Although less common, sulfur isotope analysis has become more regularly employed for studying past ecosystems, paleodiets, and mobility in recent years (Hamilton et al., 2019; Madgwick et al., 2019) and is useful for disentangling issues of equifinality (Madgwick et al., 2021). The principles of these methods have been addressed in detail elsewhere (Ambrose, 1993; Sealy, 2001; Lee–Thorp, 2008; Nehlich, 2015) and are therefore only briefly recounted here.
The isotopic composition of bone collagen provides information on dietary protein (Ambrose and Norr, 1993). Carbon (δ13C) is useful for discriminating between C4 (e.g., millet and sorghum) and C3 plants (e.g., wheat, root crops, legumes, and vegetables), which have different photosynthetic pathways. Environmental factors (Hamilton and Thomas, 2012) and the consumption of fungi also impact δ13C (Hamilton et al., 2009; O'Regan et al., 2016). However, the consumption of marine resources (seaweed, molluscs, fish, and marine mammals) has arguably the greatest impact in increasing δ13C values in a temperate Northern European context, where C4 plants are rare (e.g., Little and Schoeninger, 1995; Balasse et al., 2006; Mulville et al., 2009). Nitrogen (δ15N) principally reflects the trophic position, distinguishing herbivorous, omnivorous, and carnivorous diets, with a trophic-level enrichment between 3 and 6‰ (Hamilton and Thomas, 2012; O'Connell et al., 2012). δ15N values also vary due to manuring, where soil and, consequently, crop and faunal δ15N increase (Hamilton and Thomas, 2012; Bogaard and Outram, 2013). Marine diets also produce higher δ15N values due to elongated marine food chains (Schoeninger and DeNiro, 1984), as does nursing (Nehlich et al., 2011) and nutritional stress (Fuller et al., 2005). Landscape baselines have an important impact on faunal δ15N and δ13C (see Stevens et al., 2013), and these are dictated by multifarious and poorly understood variables, including lithology, hydrology, soil chemistry, and floral biota. Therefore, there is a locational, as well as dietary, driver in δ15N and δ13C isotope values.
Sulfur (δ34S) is less abundant in collagen and is present only in the amino acid methionine, and therefore, only advances in mass spectrometry have brought this into the mainstream of biomolecular archaeology (e.g., Sayle et al., 2019). Originating in dietary protein, the drivers of variation are diverse, with strong landscape, and dietary components (Nehlich, 2015). Sulfur isotope values in the biosphere and in organisms are much more variable than carbon and nitrogen, ranging from approximately −30 to 30‰ (Nehlich, 2015). Marine influence is commonly evident in archaeological sulfur isotope values, with marine sulfate having a homogeneous δ34S value of approximately 20‰ (Lee–Thorp, 2008), and marine diets and coastal origins reflected in data (Richards et al., 2001; Craig et al., 2006; Madgwick et al., 2013). Recent research has also demonstrated the value of sulfur isotopes for identifying animals husbanded on wetlands, which produce distinctive negative sulfur isotope values (Guiry et al., 2022). Trophic shifts cause minimal fractionation in sulfur (Richards et al., 2001; Lee–Thorp, 2008).
The aim of this study is to explore strategies in the management of animals and landscapes in response to climatic, social, and economic change at the end of the Bronze Age using carbon (δ13C), nitrogen (δ15N), and sulfur (δ34S) isotopes on faunal remains. Themes to be investigated include local and regional approaches to maximising productivity to support feasts and whether prescribed modes of husbandry were enacted for feasting animals. In addition, foddering strategies (e.g., browse, pannage, marine resources, and animal protein for pigs) are explored along with the exploitation and management of different landscapes for raising animals (e.g., salt marsh/wetland grazing and manuring). Existing zooarchaeological data, particularly in relation to demography, are considered in terms of regional management strategies and economic foci (e.g., milk, meat, wool, and traction).
2. Materials and methods
A total of 235 faunal remains, principally caprine (n = 84), cattle (n = 52), and pig (n = 93), augmented by a small number of deer (n = 4) and dogs (n = 2), from six midden sites were analysed for carbon (δ13C), nitrogen (δ15N), and sulfur (δ34S) isotopes of bone collagen. This represents one of the largest faunal studies using these three isotope systems yet delivered in archaeology. A summary of the taxonomic composition of samples from each site is provided in Table 1. Details of the taxa, elements, and contexts sampled are in Supplementary Table 1.
Table 1
| Caprine | Pig | Cattle | Cervid | Dog | Total | |
|---|---|---|---|---|---|---|
| All Cannings Cross | 10 | 5 | 1 | 1 | – | 17 |
| East Chisenbury | 20 | 9 | 6 | 2 | 1 | 38 |
| Potterne | 12 | 28 | 20 | – | – | 60 |
| Stanton St Bernard | 15 | 15 | 6 | – | 1 | 37 |
| Runnymede | 13 | 25 | 11 | 1 | – | 50 |
| Wallingford | 14 | 11 | 8 | – | – | 33 |
| Total | 84 | 93 | 52 | 4 | 2 | 235 |
Samples analysed for δ34S, δ13C, and δ15N isotopes by taxon and site (including those that failed quality control criteria).
Mandibles with teeth in situ were preferentially selected so that collagen analyses could be paired with mobility isotopes (strontium and oxygen) in dental enamel, which will be described elsewhere. Sheep and goats were not differentiated morphologically due to concerns surrounding the reliability of dental criteria (Zeder and Pilaar, 2010), but it is highly likely that the vast majority are sheep based on the paucity of evidence for goats in prehistoric Britain. When possible, repeat sampling of the same-sided elements was undertaken to ensure that each sample represented a distinct individual. In many instances, flexibility was required in the sampling strategy, and maxillae and permanent loose teeth were also sampled. Given the richness and highly fragmented nature of these midden sites, it is highly unlikely that repeat sampling of the same individual occurred, and different contexts were targeted to further reduce the risk. Bone collagen is likely to represent an averaged dietary signal for the lifetime of the animal, whereas the two dentine samples will provide information on a shorter phase of early life.
Collagen extraction was undertaken at the Cardiff University BioArchaeology laboratory following a modified Longin method (Brown et al., 1988). Bone (approximately 0.5 g) and occasionally dentine (approximately 0.2 g) were cut using a Strong 209 precision drill with a diamond wheel attachment and surfaces abraded with a diamond burr to remove adhering contaminants and >5 μm of the cortex. Samples were demineralised in 0.5 M HCl and stored at 4°C. HCl was changed weekly until full demineralisation (approximately 2 weeks). The samples were then rinsed with deionised water and gelatinised in pH3 solution (acidified with HCl) in a hot block at 70°C for 48 h. The samples were then filtered using Ezee™ filters (8 μm Ezee-filter, Elkay, Basingstoke), transferred to polypropylene test tubes, and frozen overnight before lyophilisation and weighing.
Ratios of δ13C, δ15N, and δ34S are reported as per mille (‰) relative to Vienna Pee Dee Belemnite (VPDB), Air, and Vienna Canyon Diablo Troilite (VCDT), respectively. Analyses were undertaken in duplicate (with a few exceptions where collagen yields were poor) at the National Environmental Isotope Facility, British Geological Survey, Nottinghamshire, United Kingdom. The instrumentation comprises a ThermoFinnigan EA IsoLink coupled to a Delta V Plus isotope ratio mass spectrometer via a Conflo IV interface. δ13C analyses were normalised to the VPDB scale using gelatine B2215 (Elemental Microanalysis–certified value of −22.91‰) and caffeine IAEA600 (−27.771‰), δ15N analyses were normalised to the AIR scale using gelatine B2215 (Elemental Microanalysis–certified value of 4.26‰) and caffeine IAEA600 (1.0‰), and δ34S values were normalised to the VCDT scale using gelatine B2215 (Elemental Microanalysis–certified value of 1.21‰) and International Atomic Energy Agency (IAEA) S-2 (22.7‰). A further two laboratory check standards were used, a modern cattle bone calibrated against N1 and N2 = 4.4 ± 0.16 (N), USGS 40 and 41 = −22.6 ± 0.12 (C), S1 and S2 = 12.71 ± 0.29, and commercial collagen (M1360P) calibrated against N1 and N2 = 8.12 (N), USGS 40 and 41 = −20.45 (C), and S1 and S2 = 2.99 (S). The 1σ reproducibility of repeat analyses of B2215 across the runs was better than 0.03 for δ13C, 0.03 for δ15N, and 0.12 for δ34S. B2215 was used to calculate %C, %N, and %S (certified N% = 16.55, C% = 43.78, S% = 0.51). Only the samples that adhered to published quality control indicators were considered (DeNiro, 1985; Nehlich and Richards, 2009; see Supplementary Table 1).
3. Results
Full details of the data set are reported in Supplementary Table 1, and summary statistics are provided in Supplementary Table 2. Of the 235 individuals analysed, 14 samples did not meet quality control criteria and are highlighted in grey but excluded from further consideration (Supplementary Table 2). Outliers described in the following sub-sections are defined according to Tukey's interquartile range (1.5 × IQR beyond Q1 or Q3; Lightfoot and O'Connell, 2016). Summary result descriptions pertaining to all data from a site include cervids and dogs.
3.1. δ13C isotope results
When combining taxa, δ13C isotope results show relatively limited variation across sites, with medians ranging from −22.0 to −21.7‰ (Figure 2). Caprines were relatively homogeneous across sites, especially in Wiltshire, where site medians ranged between −21.8 and −21.5‰, and only two outliers (at Potterne and All Cannings Cross) were observed. The Thames Valley sites had greater variability, with Runnymede having the widest range and both having lower medians than all Wiltshire sites. Pigs were more variable but not markedly so considering they are omnivores. Runnymede and Potterne are noteworthy in having the widest ranges (−20.2 to −23.2‰ and −20.5 to −22.5‰, respectively), but this could be an artefact of sample size (PTN, n = 28; RMD, n = 24). The cattle sample was the smallest, with only Potterne and Runnymede having more than six samples and unsurprisingly having the widest ranges. Runnymede was notable in having a lower median (−22.6‰) than all other sites (between −21.1‰ and −22.1‰, including the single All Cannings Cross sample).
Figure 2
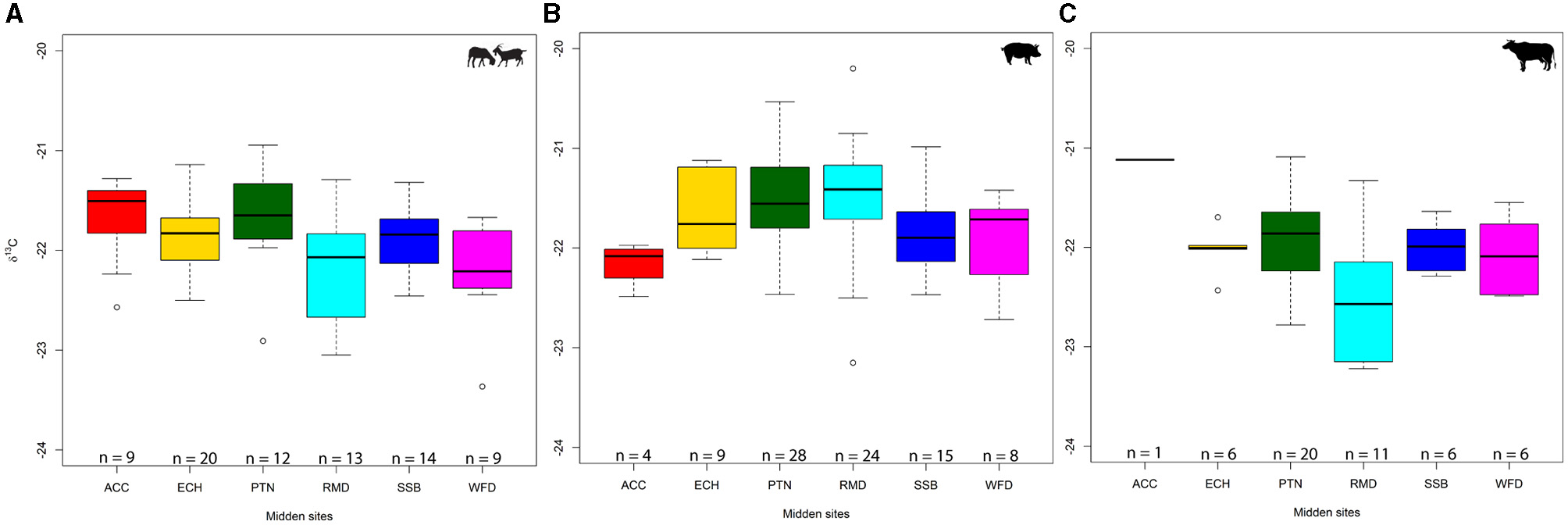
Box-and-whisker plots representing δ13C values: caprine (A), pig (B), and cattle (C). Midden sites: ACC, All Cannings Cross; ECH, East Chisenbury; PTN, Potterne; RMD, Runnymede; SSB, Stanton St Bernard; WFD, Wallingford (BGS©UKRI).
3.2. δ15N isotope results
δ15N values showed greater variation at both an inter- and intra-site level (Figure 3). Runnymede and Potterne, the largest two samples, had the highest median values when taxa were pooled (6.2‰), with East Chisenbury having the lowest (4.9‰; see Supplementary Table 2). Somewhat surprisingly, it was the relatively small sample (n = 36) of Stanton St Bernard that had the largest range (3.4–10.1‰) and the even smaller sample (n = 15) from All Cannings Cross that had the largest interquartile range (2.9‰).
Figure 3
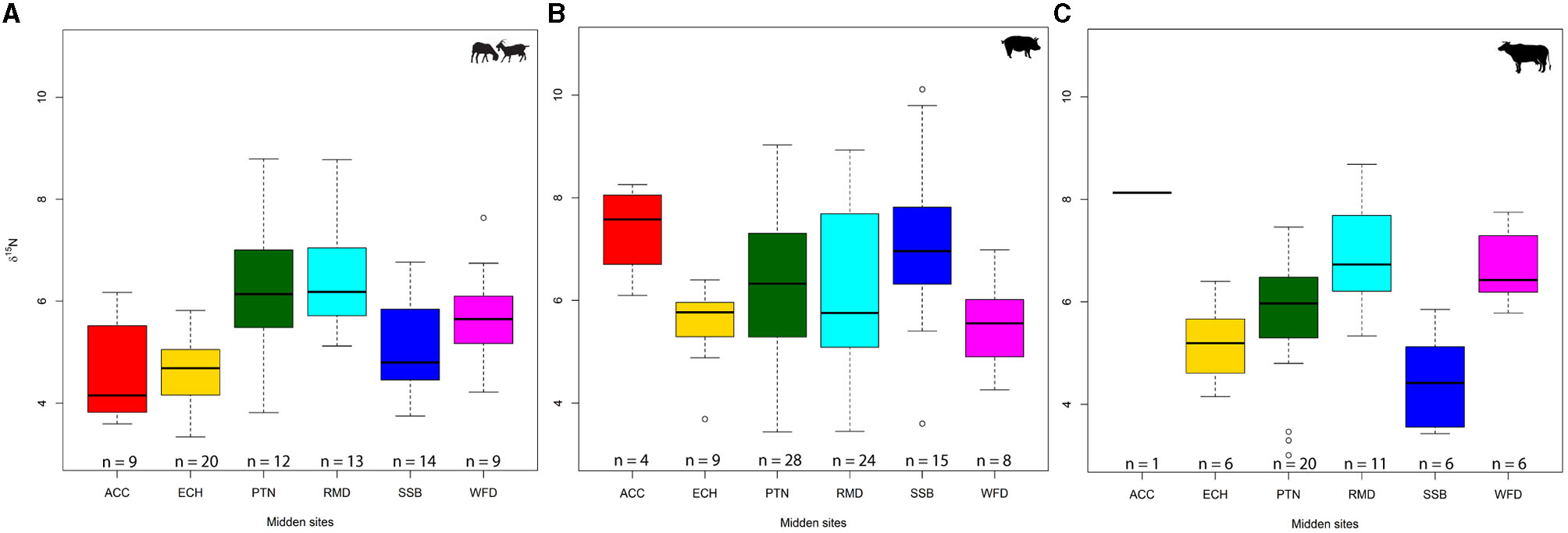
Box-and-whisker plots representing δ15N values: caprine (A), pig (B), and cattle (C). Midden sites: ACC, All Cannings Cross; ECH, East Chisenbury; PTN, Potterne; RMD, Runnymede; SSB, Stanton St Bernard; WFD, Wallingford (BGS©UKRI).
Caprines showed the least inter-site variation in δ15N, with medians ranging from 4.2‰ (All Cannings Cross) to 6.2‰ (Runnymede) and only one outlier at Wallingford (7.6‰). Potterne had the largest range (3.8–8.8‰) despite having a small caprine sample (n = 12). Runnymede (6.2‰) and Potterne (6.1‰) had the highest medians and absolute values (both 8.8‰). A high cervid value (8.3‰) suggests a δ15N-rich baseline around Runnymede. Surprisingly, the largest sample, from East Chisenbury (n = 20), had the smallest range (3.3–5.8‰). Animals from chalk sites often have low δ15N values, and it is therefore unsurprising that East Chisenbury, All Cannings Cross, and Stanton St Bernard produced the lowest medians. However, it is noteworthy that this is only the case for caprines (which align with the sites' cervids), suggesting that cattle and pigs were husbanded differently or raised elsewhere.
Pigs were markedly more diverse, reflecting their capacity for an omnivorous diet. Medians and maxima are higher for all Wiltshire sites when compared to cattle and caprines. At Runnymede, medians and maxima were comparable across taxa, but Wallingford was exceptional, with both herbivore taxa showing higher medians and maxima than pigs (5.6 and 7.0‰, respectively). As with caprines, East Chisenbury pigs had the smallest range (3.7–6.4‰) and interquartile range (0.7‰), although one low outlier (3.7‰) was present. The only other outliers were at Stanton St Bernard (3.6 and 10.1‰), which had the largest range. The two largest samples, from Potterne (n = 28) and Runnymede (n = 24), presented remarkably similar patterns of δ15N variation. Although the All Cannings Cross sample was small (n = 4), it had the highest median (7.6‰) despite having the lowest for caprines. Cattle showed marked inter-site variability and were the only taxon for which there was a notable difference between the Wiltshire and Thames Valley regions. Except for the single sample from All Cannings Cross, the Thames Valley sites had higher δ15N medians and maxima than those of the Wiltshire sites. Stanton St Bernard is noteworthy in having the lowest cattle values (median 4.4‰), in contrast to the site's pig sample and the single high (8.1‰) cattle sample from nearby All Cannings Cross. Potterne again had a large δ15N range (3.0–7.5‰), but unlike other taxa, this was extended by three low outliers, rather than being a spectrum of values.
3.3. δ34S isotope results
Sulfur represents a much more variable isotope system, reflecting both origins and diet (Figure 4). The degree of variation in the data set at both inter- and intra-site levels is vast. Potterne is the most variable in each taxon and has the widest overall range (−17.1 to +16.7‰). This is among the widest ranges published so far for an archaeological site globally. It is noteworthy that there are no outliers at the site; this range represents a spectrum of values. However, as more δ34S data sets are produced, such ranges may become more common. East Chisenbury again showed limited variability, notwithstanding two low outliers, with the smallest interquartile range (3.3‰). For caprines, Runnymede (−16.1 to 13.8‰) and Potterne (−16.9 to 11.1‰) had the widest ranges, although the Runnymede range was extended by outliers. Wallingford, a site that has shown limited variability in other proxies and tight clusters in δ34S for other taxa, shows great variation in the small sample of nine caprines (−16.9 to 10.8‰). The largest caprine sample, from East Chisenbury (n = 20), had the smallest range (8.3–17.5‰) but had a dominance of high values, with the highest median (13.8‰). All Cannings Cross and Stanton St Bernard caprines also had a dominance of positive values, with only one negative outlier at All Cannings Cross (−4.2‰). This emphasises the degree to which Potterne differed from other Wiltshire sites.
Figure 4
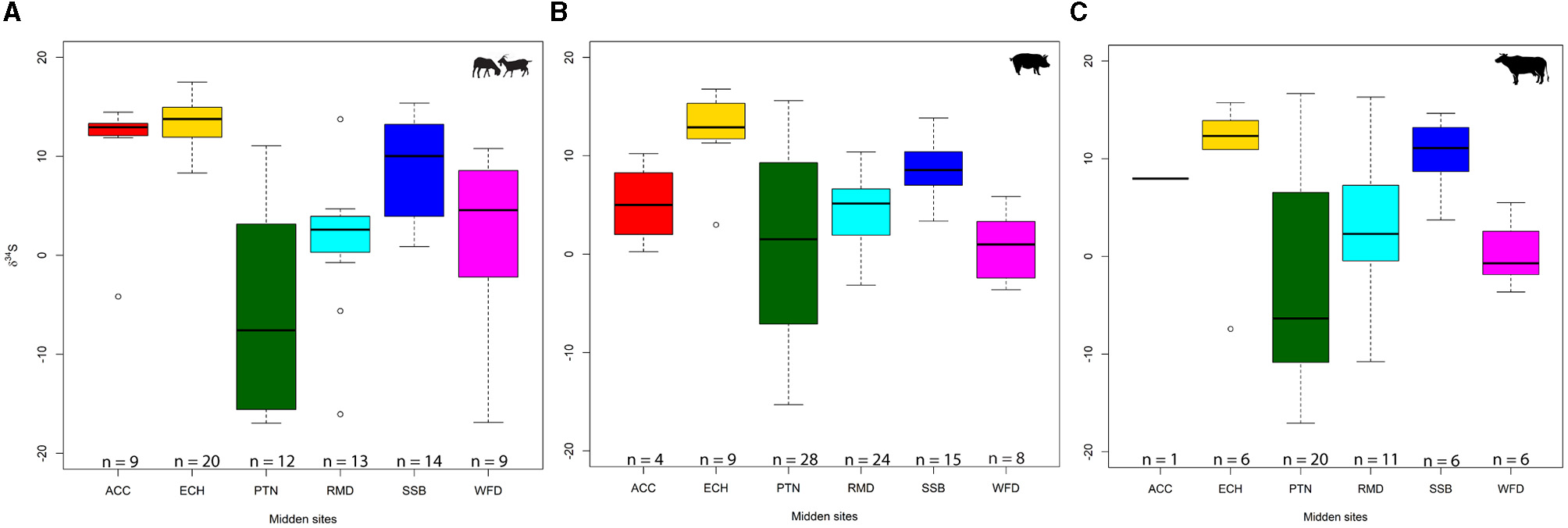
Box-and-whisker plots representing δ34S values: caprine (A), pig (B), and cattle (C). Midden sites: ACC, All Cannings Cross; ECH, East Chisenbury; PTN, Potterne; RMD, Runnymede; SSB, Stanton St Bernard; WFD, Wallingford (BGS©UKRI).
Potterne was, by far, the most wide-ranging site in pig δ34S (−15.3 to 15.6‰). By contrast, the three other Wiltshire sites were again dominated by high values with no negative results. East Chisenbury again had the highest median (12.9‰) but had one low outlier (3.0‰). Both Runnymede (−3.2 to 10.4‰) and Wallingford (−3.6 to 5.9‰) had much smaller ranges and few negative values when compared to caprines. The cattle data set presents some similar patterns. The Potterne sample is again the most diverse (−17.1 to 16.7‰), with a very wide range in only 20 individuals (interquartile range: 16.8‰). As with caprines, but in contrast to pigs, Runnymede cattle produced wide-ranging values (−10.8 to 16.3‰). By contrast, Wallingford had a limited range (−3.6 to 5.5‰), aligning with the pig sample but differing with the caprines sample. The homogeneity in the East Chisenbury samples persists in cattle (interquartile range: 2.4‰), with a dominance of high values (median: 12.3‰, with a single negative outlier: −7.4‰). Animals from Stanton St Bernard (median: 11.1‰) and the single cattle sample from All Cannings Cross (8.0‰) all produced positive values. These three middens are similar across taxa in terms of δ34S, with Potterne again representing a major outlier among Wiltshire middens. Overall, Potterne and Runnymede show the broadest δ34S ranges, but these are more regularly distributed at Potterne while in Runnymede the range is extended by outliers (see Supplementary Figure 1).
Scatter plots with δ13C and δ15N values divided per species (Figure 5) highlight the degree of bivariate variability at the inter-site and inter-taxon levels. It is noteworthy that Potterne and Runnymede generally have the most wide-ranging values across taxa and isotope systems. The consistently diverse values suggest that this is not an artefact of sample size. Considering the midden's size, East Chisenbury has surprisingly limited ranges across taxa and isotope systems, compared to other middens. For example, All Cannings Cross exhibits a surprisingly high δ15N in pigs and cattle for a chalk site yet has the lowest mean δ15N for caprines. Wallingford typically presents small ranges that broadly align with the interquartile range of the other Thames Valley midden of Runnymede, although caprine δ34S contrasts greatly, with a wide spectrum of values. Animals from Stanton St Bernard generally have a limited range, although, in some instances, show a different pattern from other Wiltshire middens, most notably in δ15N, with very low values in cattle and the greatest range in pigs. Scatter plots for δ34S and δ13C (Figure 6) reinforce the point that Potterne δ34S values are very widespread, with mean values contrasting to other Wiltshire sites, considering all taxa. The other Wiltshire sites show similar, predominantly positive values, particularly for sheep and cattle.
Figure 5

Scatter plots with δ13C and δ15N values divided per species with error bars representing the mean and one standard deviation (SD): caprine (A), pig (B), and cattle (C). Midden sites: ACC, All Cannings Cross; ECH, East Chisenbury; PTN, Potterne; RMD, Runnymede; SSB, Stanton St Bernard; WFD, Wallingford.
Figure 6

Scatter plots with δ13C and δ34S values divided per species with error bars representing the mean and one standard deviation (SD): caprine (A), pig (B), and cattle (C). Midden sites: ACC, All Cannings Cross; ECH, East Chisenbury; PTN, Potterne; RMD, Runnymede; SSB, Stanton St Bernard; WFD, Wallingford.
4. Discussion
Such a large and diverse data set presents a complex picture in terms of husbandry regimes and landscape use. There is marked variation across sites, taxa, and isotope systems. To explore patterns further, the discussion is structured by taxon, with broader interpretations offered thereafter. The focus of this article is the management, rather than the origins, of the animals. However, the data lend themselves to consideration of local or wider management. Local can be broadly defined as the biosphere zone of the site, with the same or similar lithology, sedimentology, and hydrology. This is a diffuse definition but is likely to represent a larger area in the more homogeneous chalklands of East Chisenbury, All Cannings Cross, and Stanton St Bernard and a smaller area in the more varied landscapes around Potterne, Runnymede, and Wallingford.
4.1. Caprines
Generally, caprines exhibited less variation than other domesticates in δ15N and δ13C at both an intra-and inter-site level. Whilst there is still marked variation, as expected in such a substantial data set, it may be considered suggestive of a greater degree of local management, including grazing on manured and unimproved pasturage in the vicinity of sites. At all sites, caprines are the most common domesticate (by NISP), and it is plausible that they were the mainstay of the faunal economy, supporting the local populace year-round, rather than being brought from farther afield for feasting events. However, the δ34S results cast doubt on this, with Potterne, Runnymede, and Wallingford showing exceptionally large ranges. Understanding of the degree of δ34S variation in single landscapes remains poor, and there is increasing evidence that certain hydrological processes can cause wide-ranging sulfur in limited geographical areas, especially in negative ranges (Stevens et al., 2023). However, the greater variation in caprines for this proxy, which is more strongly driven by mobility, points to varied origins of the caprines at the three aforementioned sites.
Caprines were clearly key to the economy at East Chisenbury, where there was unusual evidence of sheep dairying (Serjeantson, 2007). The narrow range of values across all three isotope proxies, despite being the largest sample, suggests that these animals were principally husbanded locally (supported by the presence of many young individuals) in a relatively formalised manner. Evidence suggests East Chisenbury could represent the centre for sheep husbandry, with tightly controlled management supporting a mixed dairy, meat, and wool economy, the latter evidenced by a large number of spindle whorls, representing 16% of small finds, although the older, wool-producing sheep were principally slaughtered elsewhere (Serjeantson et al., 2010). This interpretation is not, however, straightforward. The presence of three individuals with δ34S values above 15‰ would conventionally be considered to be derived from a coastal area, likely in western Britain (but see Hamilton et al., 2019). Primary bioavailable δ34S mapping is required to clarify this, although the three cervids and dogs from the site (14.2–16.2‰) suggest high values could be obtained locally. Nonetheless, the limited range when compared to other sites suggests controlled management regimes, raising large stocks, predominantly in local landscapes. These findings align with All Cannings Cross and Stanton St Bernard, although the latter has a broader δ34S range, likely indicating some animals raised farther afield. The three sites, all founded on the chalkland east of Potterne, have similar ranges for caprines. This and the common occurrence of young animals suggest they employed similar management regimes, perhaps under the same system of control to maximise dairy and meat production.
By contrast, Potterne clearly exhibited very different approaches to caprine management and procurement despite being nearby. The range in all three proxies is markedly larger and, interestingly, the majority of caprines from Potterne have δ34S values that do not overlap with those from other Wiltshire middens (Figure 4). This suggests a broader catchment at Potterne, with animals procured from a wider range but few coming from the same areas (presumably the Wiltshire chalklands) that dominate the sample from All Cannings Cross, Stanton St Bernard, and East Chisenbury. There is also a lack of coastal values (>15‰) in Potterne caprines compared to cattle and pigs, suggesting that catchments were different between taxa. The δ15N and δ13C results firmly support this, with different landscape baselines in evidence and animals raised on both manured and unimproved pasturage. The animal with the highest δ15N value (8.8‰) also has a high δ13C value (−21.1‰), meaning salt-marsh origins are plausible (Müldner et al., 2014), but the absence of sulfur values above 12‰ indicates that coastally raised caprines were rare at Potterne. This cannot, however, be entirely ruled out as more sheltered coastal zones can produce significantly lower δ34S biosphere values (averaging 8.8‰ in humans from coastal south-east Wales) within the coastal zone in Britain (Hemer et al., 2017). The preponderance of negative sulfur values (including the individual posited to be salt marsh–raised) strongly suggests that animals were raised in wetland environments (Guiry et al., 2022; Stevens et al., 2023). It is unclear where the closest suitable environment is, but the Somerset Levels, more than 50 km away, provides a possible origin, although primary biosphere mapping may well identify closer possibilities. The difference between Potterne's approach to caprine management compared to other Wiltshire sites is also supported by the common occurrence of older, as well as prime meat animals, suggesting a mixed utility (Locker, 2000) and the recovery of weaving artefacts (Lawson, 2000).
Runnymede caprines showed similarities to Potterne in producing wide-ranging values across isotope proxies. One notable difference is that the δ34S values are generally much more constrained. This suggests that the majority of the caprines were likely raised locally (although primary biosphere mapping is needed), with some coming from farther afield, two of which (with negative values) were likely raised on wetlands or locally gleyed soils. Western parts of the Thames, in Oxfordshire, are known to produce similar negative values (Nehlich et al., 2011). However, the locally raised animals exhibit wide-ranging δ15N and δ13C results, suggestive of varied husbandry regimes, potentially including foddering, in landscapes that were both manured and unimproved, quite unlike the controlled management of three of the Wiltshire sites. Wallingford caprines also showed more variation in δ34S, unlike cattle and pigs, indicating that caprines were likely drawn from a wider catchment, rather than the site locality having a diverse biosphere range. Unlike at Potterne, this does not seem to relate to wool production, as animals were culled younger at Wallingford (Powell and Clark, 2006). The range is comparable to Runnymede, suggesting that caprines were raised in wetlands but not in coastal zones. The presence of outliers in both δ15N and δ13C supports the interpretation of a wide catchment. Overall, the evidence suggests wider catchments and more diverse management regimes at the Thames Valley sites and Potterne, with the three other Wiltshire sites showing formalised, intensive management but at a local level.
4.2. Pigs
Pigs showed marked variation at both an inter- and intra-site level across data sets, clearly demonstrating different catchments and patterns of management. However, considering the pig sample is the largest and that pigs can be subject to wide-ranging husbandry regimes as omnivores, the results are not as diverse as might be expected.
Unlike the patterns for herbivores, the Wiltshire chalkland sites of All Cannings Cross, Stanton St Bernard, and East Chisenbury showed different catchments and patterns of management in pigs. All Cannings Cross had the smallest sample (n = 4) and smallest δ13C and δ15N range. There is little to suggest that these animals were not locally raised, although high δ15N, with all animals >6‰, is consistent with animals being fed meal scraps, perhaps raised as sty pigs adjacent to settlement. The low δ13C values ( ≤ -22‰) suggest that forest fungi and pannage were not important at All Canning Cross but were relied on to a greater degree at East Chisenbury and Stanton St Bernard, although the effect of seasonal consumption of forest forage such as fungi is variable (Hamilton et al., 2009; O'Regan et al., 2016). The low δ15N overall and the limited δ15N range (except for one low outlier) at East Chisenbury suggest that pig husbandry may have been organised to utilise forest pannage but to avoid animal protein and meal scraps. Therefore, these pigs are unlikely to have been free-ranging but more controlled in terms of the resources they could exploit, aligning with the tightly controlled management in other taxa at East Chisenbury. Stanton St Bernard pigs show the widest range of all sites in δ15N, demonstrating a more flexible approach to pig foddering. Here, animal protein, likely in the form of domestic waste, was an important dietary component.
This fluid approach to husbandry at Stanton St Bernard is not necessarily evidence of a wide catchment, as the extent of the δ34S range is comparable to the site's herbivore taxa and to All Cannings Cross pigs. Whilst it is likely different landscapes were utilised in pig rearing, there is no evidence for coastal or wetland environments at either site. The main difference is the wider range of resources used in foddering at Stanton St Bernard (albeit in a larger sample). Interestingly, East Chisenbury δ34S shows limited overlap with the other Wiltshire chalkland sites, with generally higher δ34S, suggestive of coastal origins, although this aligns with herbivores and therefore may point to a higher-than-expected local δ34S baseline at East Chisenbury (supported by high cervid values). In short, these sites are characterised by limited catchments but more diverse husbandry regimes between sites than for other domesticates. Faunal assemblages at the three sites have only small proportions of pigs and site-specific approaches to husbandry appear to have been employed, perhaps tailored to locally available resources and waste management strategies.
Despite the greater inter-site variation of the Wiltshire chalkland pigs, Potterne is a much wider outlier. The broadly comparable δ13C range to Stanton St Bernard and East Chisenbury hints at similar landscapes being exploited for porcine foddering (including seasonal exploitation of forest forage), but the vast δ34S range suggests a wider catchment. It is markedly wider than all other sites and, as with Potterne cattle, points to animals raised in wetland (likely including salt marsh), inland, and coastal environments. The very wide δ15N range supports not only a broad catchment and the exploitation of varied landscapes but also different modes of husbandry. Some appear almost entirely herbivorous and raised in a 15N-depleted, unmanured landscape. Others are likely to have a substantial amount of animal protein in their diet, perhaps fed on meal scraps or midden waste. However, the high δ15N in some caprines from the site precludes confident interpretation, and they could be herbivorous pigs raised in heavily manured 15N-rich landscapes. The wide range in δ15N and δ34S indicates a wide catchment and diverse management regimes. These strategies were employed to maximise the production of pork, a key component of the feasting events, accounting for a markedly high proportion of remains than at other Wiltshire sites.
Runnymede has a similarly large pig sample, but evidence for a wide catchment and the exploitation of varied landscapes is weaker, with a more restricted δ34S range than for other domesticates (although wide enough to suggest exploitation beyond a single location). Two δ13C outliers may point to different landscapes being exploited, but evidence for this is weak. Diverse husbandry and foddering regimes are much clearer, with a δ15N range comparable to Potterne and far wider than the site's other domesticates. No formalised regimes are apparent, with some being almost entirely herbivorous and others having varied quantities of animal protein in feeding, as well as some high δ13C values suggesting forest forage. The site would have plentiful forage for pigs with both woodland and riverside resources. This could also have encouraged pigs to stay away from arable land, which they would be likely to damage (Serjeantson, 1996). Equifinality hinders confident interpretation, and it is likely that the δ15N range also relates to animals being raised in landscapes with different δ15N baselines. Some caprines and cattle have high δ15N, so pig values cannot be automatically explained by animal protein consumption. The δ13C and δ15N scatter plot (Figure 5) shows very comparable signatures between Runnymede and Potterne, indicating similarly varied approaches to raising pigs—exploiting landscape resources flexibly to maximise meat yields. It is noteworthy that the δ15N values are overall lower in pigs than herbivores at Runnymede, suggestive of herbivorous diets in a δ15N-depleted landscape (that differs from where the cattle and caprines were grazed) for some pigs. The large-scale consumption of certain fungi that produce low δ15N values is also possible (see O'Regan et al., 2016).
The more limited Wallingford pig sample (n = 8) sits well within the Runnymede data set in all proxies. This suggests a smaller catchment for pigs, with a limited δ13C spread, the δ34S range being smaller than all other sites and far less variable than in Wallingford caprines. Low δ15N indicates predominantly herbivorous animals with resources drawn mainly from unimproved landscapes, in contrast to the evidence for manuring in cattle at the site. This suggests fairly local pig husbandry regimes, although management may have been intensive given the prescribed foddering on plant foods (albeit in a small data set) and the high proportion of pig bones in the faunal assemblage. The area has been highlighted as suited to pig husbandry, with secondary woodland and the river edge providing ample fodder, such as waterlilies, for pigs (Powell and Clark, 2006).
Overall evidence suggests pigs were fed diverse diets and drawn from a wide catchment to maximise stocks for feasting events at Potterne. Due to a lack of secondary products, pigs are frequently considered a sign of wealth (Serjeantson, 1996) and their being drawn from a wide catchment would emphasise this further. Other Wiltshire sites had more limited catchments but employed different approaches to raising pigs, with variable use of animal protein and forest resources. Runnymede also had a more limited catchment for sourcing pigs, but pork production was maximised through utilising diverse dietary sources. Wallingford appears to have employed a local, plant-based management regime.
4.3. Cattle
The markedly wider range of δ13C and δ15N values in cattle compared to caprines (the other herbivore taxon), particularly at an inter-site level, is noteworthy, especially considering the smaller number of cattle samples. It is plausible that cattle were frequently grazed on varied pasturage and sourced from wider catchments to support feasts. There were also differences between regions with overall lower δ13C and higher δ15N at the Thames Valley sites. This indicates that sites (and regions) employed different approaches to cattle husbandry and aspects such as grazing land and foddering regimes varied between several sites. The lower variation in caprines suggests that this pattern is not purely an issue of local baselines but approaches to animal management. The δ34S range was quite similar to caprines (although smaller at Wallingford), suggesting similar catchments at each site, although equifinality precludes confident interpretation. Zooarchaeological data indicate that cattle were present in smaller numbers than caprines, but their dietary contribution would still have been major at all sites. It is noteworthy that their representation oscillated over time, increasing at Wallingford and decreasing at Runnymede and East Chisenbury. This suggests that their role may have been more than purely as an economic staple, perhaps changing in relation to the frequency of (or their importance in) feasting.
Some aspects are comparable to caprines, with Stanton St Bernard and East Chisenbury showing similar patterns. These sites have overall lower δ15N values than other sites and limited variability in both δ15N and δ13C, suggesting local husbandry with little manuring in the naturally 15N-depleted Wiltshire chalklands. These sites also have a limited δ34S range (excepting a negative outlier at East Chisenbury), comparable to caprines and relatively high for the inland location, based on the current understanding of the sulfur biosphere. At least one individual (<15‰, from East Chisenbury) would conventionally be considered of coastal origin, although baseline analysis is required and the presence of a cervid with an even higher value points to a higher-than-expected local range. The single sample from All Cannings Cross is comparable in terms of δ34S but very different in being among the highest values in both δ15N and δ13C. Whether this is an outlier from another region or a contrasting approach to husbandry at the site is impossible to say. Potterne, which comprised the largest cattle sample, has a much wider range in all proxies. The extreme δ34S range (from −17.1 to 16.7‰), points to different wetland, inland, and coastal zones being utilised. The δ15N and δ13C ranges are fairly large, with some animals raised in unimproved 15N-depleted environments (potentially Wiltshire chalklands). Whilst δ15N values are overall higher at Potterne than East Chisenbury and Stanton St Bernard, there remains only limited evidence for manuring.
The Thames Valley sites contrast markedly, particularly in having higher δ15N and lower δ13C overall. This is likely to relate to a combination of different landscape baselines and varying husbandry and foddering regimes between regions. Caprines generally show less variation than cattle across sites and isotope proxies despite having larger samples. The opposite is true at Wallingford, with cattle having a smaller range than caprines in all data sets, with the difference being most pronounced in δ34S. This strongly suggests more local cattle management at Wallingford, consistent with the low δ34S values, potentially under relatively tightly controlled management regimes. The higher δ15N values compared to Wiltshire (and Wallingford caprines) suggest more manuring in cattle pasturage. Runnymede cattle had a far greater range in all proxies, with Wallingford samples broadly centring on the interquartile range at Runnymede. This provides further evidence for more local husbandry at Wallingford and a wider catchment, utilising a broader range of landscapes and foddering practices at Runnymede. The δ34S was exceptionally wide-ranging, considering only 11 cattle were analysed, and indicates cattle raised in inland, wetland, and coastal landscapes. The other isotope proxies are comparable to Runnymede caprines, suggesting that the two herbivore taxa were managed in a similar way (and derived from similar catchments). The very low δ13C values in Runnymede cattle, lower than for any taxonomic group for any site, suggests some particularly 13C-depleted landscapes favoured for cattle husbandry.
The marked inter-site (and inter-regional) variation in cattle suggests that particular modes of cattle management were employed at different sites. East Chisenbury and Stanton St Bernard again show a pattern of tightly controlled management regimes, although the evidence is weaker than for caprines due to smaller sample sizes. In contrast to caprines, Wallingford cattle point to similarly local use of landscapes, with Potterne and Runnymede again showing more diverse management and a markedly wider catchment, although this may be partially explained by the sample size.
4.4. Summary
This large and multifaceted data set presents a complex picture of diverse approaches to animal management. Both Wiltshire and Thames Valley middens display marked variation in husbandry practices and landscape use between sites, although similar strategies were employed at certain sites (Figure 7). Evidence suggests some sites employed an intensive, specialist management regime to boost mixed economy production for certain animals, whereas wide-ranging, flexible strategies maximised productivity by utilising diverse landscape resources at other sites. A range of strategies for maximising production is in evidence, including the use of manured, forested, salt-marsh, and wetland landscapes.
Figure 7

Bivariate scatter plots comparing δ13C and δ15N (A) and δ13C and δ34S (B) values across taxa at each site. ACC, All Cannings Cross; ECH, East Chisenbury; PTN, Potterne; RMD, Runnymede; SSB, Stanton St Bernard; WFD, Wallingford. Large circles represent taxon means (BGS©UKRI).
Maximising meat production seems to have been a priority at Potterne. A wide catchment and flexibility in husbandry were used to meet the supply demands of the ostentatious feasts. This likely represents household production on a large scale. Caprines show more rigid management strategies at many sites but not at Potterne, where diverse approaches supported the meat and wool economy. Nearby, East Chisenbury, another midden of monumental proportions, showed a very different pattern. Animal management was tightly controlled, with prescribed regimes adhered to and a similar range of landscape resources exploited, predominantly in the vicinity of the site. Both isotope evidence and archaeological evidence point to East Chisenbury being a centre for sheep husbandry with choreographed management strategies supporting a mixed meat, wool, and dairy economy. The nearby middens of All Cannings Cross and Stanton St Bernard show similar patterns of caprine husbandry as East Chisenbury, potentially representing an integrated, specialist caprine management regime that supplied feasts and the local populace. This is not the case for cattle and pigs; it seems that site-specific regimes and, in some instances, more flexible approaches to raising stocks were employed. This could represent animals coming from different or wider catchments and/or husbandry occurring at a household level, drawing on different resources.
Runnymede and Wallingford show a similar pattern of caprine management to Potterne, with the large stocks being raised through diverse husbandry strategies, likely from a wide catchment. As the most prevalent domesticate (by NISP), it is interesting that three sites employed an intensive, local, tightly controlled regime and that three were more flexible, using different landscape resources to increase flocks. Diverse resources, principally from the local area were used to raise the large numbers of pigs at Runnymede, whereas Wallingford pigs likely had more restricted, local plant-based diets. Local landscapes also supported cattle husbandry at Wallingford, in contrast to Runnymede, where diverse landscapes and foddering resources were utilised. As at Potterne, flexibility and a wide catchment were central to providing cattle to Runnymede.
No other δ34S isotope data have been produced from middens or directly contemporaneous settlements in southern Britain. The most fitting comparative data set is from Middle to Late Iron Age fauna from Danebury environs (Hamilton et al., 2019). δ34S ranged from 0.8 to 19.9‰ (n = 71). It is noteworthy that there were no negative values, whereas these are present in all middens in this study except Stanton St Bernard. Using cluster analysis of the faunal data, a local range for the chalkland around the site is assigned at 12.9 to 18.8‰, markedly higher than indicated by British Geological Survey (BGS) mapping (Evans et al., 2022). This aligns more closely to East Chisenbury, All Cannings Cross, and Stanton St Bernard values, particularly for herbivores, hinting that many of these may be local rather than coastally raised. Distant origins are suggested for some animals, supporting the idea of a wide catchment for Runnymede and Potterne, given their markedly wider sulfur ranges than the Danebury fauna. Conventionally, such large δ34S ranges (as evidenced at Potterne) would be assumed to indicate varied origins and wide resourcing. However, given that this range extends across all three domesticates, it is just possible that the immediate vicinity of Potterne has bioavailable δ34S that covers almost as much variation as the whole of Britain. Understanding of δ34S variation in the biosphere, how this changes over time, and the best approaches to mapping is, however, still evolving (Guiry et al., 2021, 2022; Evans et al., 2022; Stevens et al., 2022). When the scale of the site and the volume of deposited material are considered (with limited settlement evidence), a wide catchment and diverse animal origins seem a more parsimonious explanation, a conclusion only viable through a multi-isotope approach.
δ13C and δ15N data have been produced for Runnymede (Hamilton, 2015), for Potterne, Llanmaes (Madgwick et al., 2012b), and the enclosure bordering the East Chisenbury midden (Andrews et al., 2021; Table 2). Differences between these data sets and those presented in this study are subtle, with existing and new Potterne and Runnymede data sets in close alignment and Llanmaes sitting well within the range of other middens. The only noteworthy taxon-specific difference is lower δ15N in cattle from the East Chisenbury enclosure compared to the midden. This hints that improved pasturage was more frequently used for animals feasted on at the midden, compared to in the domestic context of the enclosure. Stevens et al. (2013) study of fauna from the Danebury environs chalkland sites that span the Iron Age in Hampshire is also worthy of comparison. Values overlap with middens, especially the Wiltshire sites founded on chalk, although it is noteworthy that herbivore δ15N values in Danebury environs tend to be lower in terms of mean and absolute range.
Table 2
| Caprine | Pig | Cattle | |||||||
|---|---|---|---|---|---|---|---|---|---|
| δ13C | n | mean | SD | n | mean | SD | n | mean | SD |
| East Chisenbury Enc. | 12 | −21.6 | 0.2 | 12 | −21.3 | 0.3 | 12 | −21.7 | 0.3 |
| Llanmaes (2012) | 12 | −21.8 | 0.4 | 57 | −21.5 | 0.4 | 12 | −22.1 | 0.3 |
| Potterne (2012) | 10 | −21.5 | 0.4 | 36 | −21.2 | 0.5 | 10 | −22.0 | 0.3 |
| Runnymede (2015) | 19 | −21.6 | 0.5 | 20 | −21.0 | 0.5 | 17 | −21.8 | 0.4 |
| Potterne (2023) | 12 | −21.6 | 0.5 | 28 | −21.5 | 0.5 | 20 | −21.9 | 0.5 |
| East Chisenbury (2023) | 20 | −21.9 | 0.4 | 9 | −21.7 | 0.4 | 6 | −22.0 | 0.2 |
| All Cannings Cross (2023) | 9 | −21.7 | 0.4 | 4 | −22.2 | 0.2 | 1 | −21.2 | – |
| Stanton St Bernard (2023) | 14 | −21.9 | 0.4 | 15 | −21.9 | 0.4 | 6 | −22.0 | 0.3 |
| Runnymede (2023) | 13 | −22.1 | 0.6 | 24 | −21.5 | 0.6 | 11 | −22.6 | 0.6 |
| Wallingford (2023) | 9 | −22.2 | 0.5 | 8 | −21.9 | 0.5 | 6 | −22.1 | 0.5 |
| δ15N | n | mean | SD | n | mean | SD | n | mean | SD |
| East Chisenbury Enc. | 12 | 3.9 | 0.7 | 12 | 5.4 | 1.5 | 12 | 3.7 | 1.0 |
| Llanmaes (2012) | 12 | 6.3 | 1.0 | 57 | 6.6 | 1.2 | 12 | 5.5 | 0.9 |
| Potterne (2012) | 10 | 5.5 | 1.4 | 36 | 5.6 | 1.2 | 10 | 5.4 | 0.8 |
| Runnymede (2015) | 19 | 7.1 | 1.0 | 20 | 5.9 | 1.4 | 17 | 5.8 | 1.4 |
| Potterne (2023) | 12 | 6.2 | 1.3 | 28 | 6.3 | 1.3 | 20 | 4.5 | 1.0 |
| East Chisenbury (2023) | 20 | 4.6 | 0.6 | 9 | 5.5 | 0.8 | 6 | 5.7 | 1.2 |
| All Cannings Cross (2023) | 9 | 4.6 | 1.0 | 4 | 7.4 | 0.9 | 1 | 8.1 | – |
| Stanton St Bernard (2023) | 14 | 5.1 | 0.9 | 15 | 7.0 | 1.6 | 6 | 4.5 | 1.0 |
| Runnymede (2023) | 13 | 6.4 | 1.2 | 24 | 6.2 | 1.6 | 11 | 6.9 | 1.1 |
| Wallingford (2023) | 9 | 5.8 | 1.0 | 8 | 5.5 | 0.9 | 6 | 5.9 | 0.7 |
5. Conclusion
This study presents a complex picture of management regimes and catchments at middens. Different sites clearly took distinct approaches to the intensification of agricultural production and the supply of feasts. Some of these patterns of inter-site variation may be underpinned by differences in chronology, though this remains relatively poorly understood (see Supplementary material). East Chisenbury instigated a tightly controlled, specialist caprine economy, with animals intensively managed locally to support a mixed dairy, meat, and wool economy. All Cannings Cross and Stanton St Bernard are perhaps part of the same regime—managing animals, especially caprines, on the surrounding chalklands, although at Stanton St Bernard more diverse management is evidenced to maximise pork production. Potterne contrasts greatly with the other Wiltshire sites and, along with Runnymede, has a very different approach. These sites show wide catchments and diverse husbandry regimes across species. They may have represented regional hubs, drawing in people and animals from across the region and sometimes beyond to support feasting events and the agricultural economy. There were no prescribed modes of husbandry—a wide catchment, flexible management, and diverse pasturage were keys to boosting productivity at these sites. Wallingford pigs and cattle were raised more locally, although caprines were drawn from a wider catchment and raised on varying pasturage.
This study makes a valuable addition to the limited corpus of faunal δ34S data from prehistoric Britain, but it is important to state that the interpretation of δ34S data remains challenging. This research demonstrates the value of combining multi-isotope approaches for exploring animal management. Issues of equifinality, particularly in relation to whether variable data relate to a different catchment or a different husbandry regime, can often be disentangled to some degree with the addition of δ34S data. Further insights will be gained with the contribution of 87Sr/86Sr and δ18O isotope data.
The research provides major new insights into social and economic change during the Bronze Age–Iron Age transition in southern Britain. The high degree of variability across middens demonstrates regional and local responses, using different economic strategies to maximise productivity and support feasting events. This was not, however, a time of agricultural intensification by any means. It is clear that prescribed species-specific approaches were employed at various sites, and just as feasting was often socially circumscribed (Madgwick and Mulville, 2015a), so was animal management. This further supports the view that animals and food were being used much more holistically in the construction of social relations and that they were much more than just a heightened focus for the economy (Barrett, 1989; Needham, 2007). They provided the focus for arenas of social competition, although likely on a much more local and regional scale than during the peak of the bronze trade network.
Statements
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
RM designed the research, acquired funding, selected samples, formulated interpretations, and wrote the manuscript. CE selected samples, undertook all sample preparation, analysed data, formulated interpretations, and wrote the manuscript. AL undertook all mass spectrometry of samples, formulated interpretations, and contributed to the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by the Arts and Humanities Research Council (AH/T006528/1) and UKRI (CoA scheme). The Marie Skłodowska-Curie Actions-European Commission provided a research grant to CE (Grant HORIZON-MSCA-2021-PF-01-101065320) during manuscript writing and revision stages.
Acknowledgments
We are very grateful to Lisa Brown (The Wiltshire Museum), Angie Bolton (Oxfordshire Museums Service), and Neil Wilkin and Sophie Crump (British Museum) for providing access to collections and sampling support. Thanks also go to David McOmish, John Barrett, and Andrew Simms for archive support with East Chisenbury, All Cannings Cross, and Stanton St Bernard and to Rhiannon Stevens, Derek Hamilton, and Eric Guiry for insightful discussion surrounding δ34S variation. We are grateful to Luca Bondioli for commenting on a draft and to the reviewers whose comments have markedly improved the manuscript. Jerome Hancock, Hugh Nianias, and Nathaniel Harrop-Pender assisted with sample processing as part of their MSc studies. We are grateful to the Arts and Humanities Research Council (AH/T006528/1) and UKRI (CoA scheme) for funding the project.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fearc.2023.1221581/full#supplementary-material
References
1
Ambrose S. H. (1993). “Isotopic analysis of paleodiets: methodological and interpretive considerations,” in Investigations of ancient human tissue: chemical analyses in anthropology, ed. M. K. Sandford (Philadelphia: Gordon and Breach) 59–130.
2
Ambrose S. H. Norr L. (1993). “Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate,” in Prehistoric human bone—archaeology at the molecular level, eds. J. B. Lambert and G. Grupe (Berlin: Springer-Verlag Berlin Heidelberg) 1–37. 10.1007/978-3-662-02894-0_1
3
Andrews P. Booth T. Crabb N. Egging Dinwiddy K. Foster J. Garland K. et al . (2021). East Chisenbury midden 2015–17: Further investigations of the late prehistoric midden deposits, enclosure and associated settlement. Wiltshire Archaeol. Nat. History Mag.114, 84–121.
4
Armit I. Swindles G. T. Becker K. Plunkett G. Blaauw M. (2014). Rapid climate change did not cause population collapse at the end of the European Bronze Age. Proc. Nat. Acad. Sci.111, 17045–17049. 10.1073/pnas.1408028111
5
Balasse M. Tresset A. Ambrose S. H. (2006). Stable isotope evidence (δ13C, δ18O) for winter feeding on seaweed by Neolithic sheep of Scotland. J. Zool. 270, 170–176. 10.1111/j.1469-7998.2006.00104.x
6
Barrett J. (1989). “Food, gender and metal: Questions of social reproduction,” in The Bronze Age - Iron Age transition in Europe: Aspects of continuity and change in European Societies 1200-500 BC, eds. M. L. S. Sorensen and R. Thomas (Oxford: BAR).
7
Bogaard A. Outram A. K. (2013). Palaeodiet and beyond: stable isotopes in bioarchaeology. World Archaeol.45, 333–337. 10.1080/00438243.2013.829272
8
Brown T. A. Nelson D. E. Vogel J. S. Southon J. R. (1988). Improved collagen extraction by modified Longin method. Radiocarbon30, 171–177. 10.1017/S0033822200044118
9
Craig O. E. Ross R. Andersen S. H. Milner N. Bailey G. N. (2006). Focus: sulphur isotope variation in archaeological marine fauna from northern Europe. J. Archaeol. Sci.33, 1642–1646. 10.1016/j.jas.2006.05.006
10
DeNiro M. J. (1985). Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature317, 806–809. 10.1038/317806a0
11
Evans J. A. Chenery C. A. Mee K. Marchant A. P. (2022). Biosphere Isotope Domains GB (V2): Interactive Website. British Geological Survey. 10.5285/2ce/7fc22-1b6e-4679-968f-42058c0120fb
12
Fuller B. T. Fuller J. L. Sage N. E. Harris D. A. O'Connell T. C. Hedges R. E. M. (2005). Nitrogen balance and δ15N: why you're not what you eat during nutritional stress. Rapid. Commun. Mass Spectrom.19, 2497–2506. 10.1002/rcm.2090
13
Gillis R. Bréhard S. Bălăşescu A. Ughetto-Monfrin J. Popovici D. Vigne J.-D. et al . (2013). Sophisticated cattle dairy husbandry at Borduşani-Popină (Romania, fifth millennium BC): the evidence from complementary analysis of mortality profiles and stable isotopes. World Archaeol.45, 447–472. 10.1080/00438243.2013.820652
14
Guiry E. Noël S. Fowler J. (2021). Archaeological herbivore δ13C and δ34S provide a marker for saltmarsh use and new insights into the process of 15N-enrichment in coastal plants. J. Archaeol. Sci.125, 105295. 10.1016/j.jas.2020.105295
15
Guiry E. J. Orchard T. J. Needs-Howarth S. Szpak P. (2022). Freshwater wetland–driven variation in sulfur isotope compositions: Implications for human paleodiet and ecological research. Front. Ecol. Evol.10, 953042. 10.3389/fevo.2022.953042
16
Hamilton J. (2015). Carbon and nitrogen isotopes of domestic animals and their application to the archaeology of the Upper and Middle Thames Valley in the Neolithic to Roman periods (Unpublished DPhil). University of Oxford.
17
Hamilton J. Hedges R. E. M. Robinson M. (2009). Rooting for pigfruit: pig feeding in Neolithic and Iron Age Britain compared. Antiquity83, 998–1011. 10.1017/S0003598X00099300
18
Hamilton J. Thomas R. (2012). Pannage, pulses and pigs: Isotopic and zooarchaeological evidence for changing pig management practices in later medieval England. Mediev. Archaeol.56, 234–259. 10.1179/0076609712Z.0000000008
19
Hamilton W. D. Sayle K. L. Boyd M. O. E. Haselgrove C. C. Cook G. T. (2019). ‘Celtic cowboys' reborn: Application of multi-isotopic analysis (δ13C, δ15N, and δ34S) to examine mobility and movement of animals within an Iron Age British society. J. Archaeol. Sci.101, 189–198. 10.1016/j.jas.2018.04.006
20
Hemer K. A. Lamb A. L. Chenery C. A. Evans J. A. (2017). A multi-isotope investigation of diet and subsistence amongst island and mainland populations from early medieval western Britain. Am. J. Phys. Anthropol.162, 423–440. 10.1002/ajpa.23127
21
Lawson A. (2000). Potterne 1982-1985. Animal husbandry in later prehistoric Wiltshire.Salisbury: Trust for Wessex Archaeology.
22
Lee–Thorp J. A. (2008). On isotopes and old bones. Archaeometry50, 925–950. 10.1111/j.1475-4754.2008.00441.x
23
Lightfoot E. O'Connell T. (2016). On the use of biomineral oxygen isotope data to identify human migrants in the archaeological record: intra-sample variation, statistical methods and geographical considerations. PLoS ONE11, e0153850. 10.1371/journal.pone.0153850
24
Little E. A. Schoeninger M. J. (1995). The late woodland diet on Nantucket Island and the problem of maize in coastal New England. Am. Antiq.60, 351–368. 10.2307/282145
25
Locker A. (2000). “Animal bone,” in Potterne 1982-5: Animal husbandry in later prehistoric Wiltshire, ed. A. Lawson (Salisbury: Trust for Wessex Archaeology) 101–117.
26
Madgwick R. (2011). “The faunal assemblage,” in The Whitchurch excavations 2006-2009: An interim report, eds. K. Waddington and N. Sharples (Cardiff: Cardiff University Press) 53–55.
27
Madgwick R. (2014). What makes bones shiny? Investigating trampling as a cause of bone abrasion. Archaeol, Anthropol, Sci, 6, 163–173. 10.1007/s12520-013-0165-0
28
Madgwick R. (2016). New light on feasting and deposition: exploring accumulation history through taphonomic analysis at later prehistoric middens in Britain. Archaeol. Anthropol. Sci.8, 329–341. 10.1007/s12520-015-0271-2
29
Madgwick R. Lamb A. L. Sloane H. Nederbragt A. J. Albarella U. Pearson M. P. et al . (2019). Multi-isotope analysis reveals that feasts in the Stonehenge environs and across Wessex drew people and animals from throughout Britain. Sci. Adv. 5, eaau6078. 10.1126/sciadv.aau6078
30
Madgwick R. Lamb A. L. Sloane H. Nederbragt A. J. Albarella U. Pearson M. P. et al . (2021). A veritable confusion: use and abuse of isotope analysis in archaeology. Arch. J.178, 361–385. 10.1080/00665983.2021.1911099
31
Madgwick R. Mulville J. (2015a). Feasting on fore-limbs: conspicuous consumption and identity in later prehistoric Britain. Antiquity89, 629–644. 10.15184/aqy.2015.24
32
Madgwick R. Mulville J. (2015b). Reconstructing depositional histories through bone taphonomy: extending the potential of faunal data. J. Archaeol. Sci.53, 255–263. 10.1016/j.jas.2014.10.015
33
Madgwick R. Mulville J. Evans J. (2012a). Investigating diagenesis and the suitability of porcine enamel for strontium (87Sr/86Sr) isotope analysis. J. Anal. At. Spectrom.27, 733. 10.1039/c2ja10356g
34
Madgwick R. Mulville J. Stevens R. E. (2012b). Diversity in foddering strategy and herd management in late Bronze Age Britain: An isotopic investigation of pigs and other fauna from two midden sites. Environ. Archaeol.17, 126–140. 10.1179/1461410312Z.00000000011
35
Madgwick R. Sykes N. Miller H. Symmons R. Morris J. Lamb A. (2013). Fallow deer (Dama dama dama) management in Roman South-East Britain. Archaeol. Anthropol. Sci.5, 111–122. 10.1007/s12520-013-0120-0
36
McOmish D. (1996). East Chisenbury: ritual and rubbish at the British Bronze Age—Iron Age transition. Antiquity70, 68–76. 10.1017/S0003598X00082892
37
Müldner G. Britton K. Ervynck A. (2014). Inferring animal husbandry strategies in coastal zones through stable isotope analysis: new evidence from the Flemish coastal plain (Belgium, 1st−15th century AD). J. Archaeol. Sci.41, 322–332. 10.1016/j.jas.2013.08.010
38
Mulville J. Madgwick R. Stevens R. O'Connell T. Craig O. Powell A. et al . (2009). Isotopic analysis of faunal material from South Uist, Western Isles, Scotland. J. North At.2, 51–59. 10.3721/037.002.0106
39
Needham S. (2007). “800BC, The great divide,” in The Earlier Iron Age in Britain and the near continent, eds. C. Haselgrove and R. Pope (Oxford: Oxbow) 39–63. 10.2307/j.ctvh1dwqj.5
40
Needham S. P. Spence T. (1996). Refuse and Disposal at Area 16 East Runnymede: Runnymede Bridge Research Excavations. London: British Museum.
41
Nehlich O. (2015). The application of sulphur isotope analyses in archaeological research: A review. Earth Sci. Rev.142, 1–17. 10.1016/j.earscirev.2014.12.002
42
Nehlich O. Fuller B. T. Jay M. Mora A. Nicholson R. A. Smith C. I. et al . (2011). Application of sulphur isotope ratios to examine weaning patterns and freshwater fish consumption in Roman Oxfordshire, UK. Geochim. Cosmochim. Acta75, 4963–4977. 10.1016/j.gca.2011.06.009
43
Nehlich O. Richards M. P. (2009). Establishing collagen quality criteria for sulphur isotope analysis of archaeological bone collagen. Archaeol. Anthropol. Sci.1, 59–75. 10.1007/s12520-009-0003-6
44
O'Connell T. C. Kneale C. J. Tasevska N. Kuhnle G. G. C. (2012). The diet-body offset in human nitrogen isotopic values: A controlled dietary study. Am. J. Phys. Anthropol.149, 426–434. 10.1002/ajpa.22140
45
O'Regan H. J. Lamb A. L. Wilkinson D. M. (2016). The missing mushrooms: Searching for fungi in ancient human dietary analysis. J. Archaeol. Sci.75, 139–143. 10.1016/j.jas.2016.09.009
46
Powell A. Clark K. M. (2006). “Animal bone,” in Archaeology of the Wallingford Bypass, 1986-92: Late Bronze Age ritual and habitation on a Thames eyot at Whitecross Farm, Wallingford, eds. A. M. Cromarty, A. Barclay, G. Lambrick, and M. Robinson (Oxford: Oxford Archaeology) 105–110.
47
Richards M. P. Fuller B. T. Hedges R. E. M. (2001). Sulphur isotopic variation in ancient bone collagen from Europe: implications for human palaeodiet, residence mobility, and modern pollutant studies. Earth Planet Sci. Lett.191, 185–190. 10.1016/S0012-821X(01)00427-7
48
Roberts B. W. Boughton D. Dinwiddy M. Doshi N. Fitzpatrick A. P. Hook D. et al . (2015). Collapsing commodities or lavish offerings? Understanding massive metalwork deposition at Langton Matravers, Dorset during the Bronze Age-Iron Age transition. Oxford J. Archaeol.34, 365–395. 10.1111/ojoa.12064
49
Sayle K. L. Brodie C. R. Cook G. T. Hamilton W. D. (2019). Sequential measurement of δ 15 N, δ 13 C and δ 34 S values in archaeological bone collagen at the Scottish Universities Environmental Research Centre (SUERC): A new analytical frontier. Rapid Commun. Mass Spectr.33, 1258–1266. 10.1002/rcm.8462
50
Schoeninger M. J. DeNiro M. J. (1984). Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim. Cosmochim. Acta48, 625–639. 10.1016/0016-7037(84)90091-7
51
Sealy J. C. (2001). “Body tissue chemistry and paleodiet,” in Handbook of archaeological Science, eds. D. R. Brothwell and A. M. Pollard (Sussex: John Wiley and Sons Ltd.) 269–281.
52
Serjeantson D. (1996). “The animal bones,” in Runnymede Bridge research excavations. Volume 2, refuse and disposal at Area 16 East, Runnymede, eds. S. Needham and T. Spence (London: British Museum Press) 194–223.
53
Serjeantson D. (2007). “Intensification of animal husbandry in the Late Bronze Age? The contribution of sheep and pigs,” in The Earlier Iron Age in Britain and the Near Continent, eds. C. Haselgrove and R. Pope (Oxford: Oxbow) 80–93. 10.2307/j.ctvh1dwqj.7
54
Serjeantson D. Bagust J. Jenkins C. (2010). Animal bone. The Late Bronze Age and early Iron Age midden site at East Chisenbury, Wiltshire. Wiltshire Archaeol. Nat. Hist. Magaz.103, 35–101.
55
Sharples N. M. (2010). Social Relations in later Prehistory: Wessex in the First Millennium BC. Oxford: Oxford University Press. 10.1093/oso/9780199577712.001.0001
56
Stevens R. E. Lightfoot E. Hamilton J. Cunliffe B. W. Hedges R. E. M. (2013). One for the master and one for the dame: stable isotope investigations of Iron Age animal husbandry in the Danebury Environs. Archaeol. Anthropol. Sci.5, 95–109. 10.1007/s12520-012-0114-3
57
Stevens R. E. Reade H. Read D. S. Bottrell S. H. Frémondeau D. Wexler S. (2022). Iso-Wetlands: unlocking wetland ecologies and agriculture in prehistory through sulfur isotopes. Archaeol. Int.25, 168. 10.14324/111.444.ai.2022.11
58
Stevens R. E. Reade H. Sayle K. L. Tripp J. A. Frémondeau D. Lister A. et al . (2023). Major excursions in sulfur isotopes linked to permafrost change in Eurasia during the last 50,000 years. Res. Sq.10.21203/rs.3.rs-2556240/v1
59
Tubb P. C. (2011). Late Bronze Age/Early Iron Age transition sites in the Vale of Pewsey: the East Chisenbury midden in its regional context. Wiltshire Archaeol. Nat. Hist. Magaz.104, 44–61. 10.30861/9781407308418
60
Turney C. S. M. Jones R. T. Thomas Z. A. Palmer J. G. Brown D. (2016). Extreme wet conditions coincident with Bronze Age abandonment of upland areas in Britain. Anthropocene13, 69–79. 10.1016/j.ancene.2016.02.002
61
Waddington K. Bayliss A. Higham T. Madgwick R. Sharples N. (2019). Histories of deposition: creating chronologies for the Late Bronze Age–Early Iron Age transition in Southern Britain. Archaeol. J.176, 84–133. 10.1080/00665983.2018.1504859
62
Waddington K. Sharples N. (2011). The Excavations at Whitchurch 2006-2009: An Interim Report. Cardiff: Cardiff Studies in Archaeology.
63
Zeder M. Pilaar S. (2010). Assessing the reliability of criteria used to identify mandibles and mandibular teeth in sheep, Ovis, and goats, Capra. J. Archaeol. Sci.37, 225–242. 10.1016/j.jas.2009.10.002
Summary
Keywords
Bronze Age, Iron Age, feasting, animal management, carbon, nitrogen, sulfur, isotopes
Citation
Madgwick R, Esposito C and Lamb AL (2023) Farming and feasting during the Bronze Age–Iron Age transition in Britain (ca. 900–500 bce): multi-isotope evidence for societal change. Front. Environ. Archaeol. 2:1221581. doi: 10.3389/fearc.2023.1221581
Received
12 May 2023
Accepted
18 August 2023
Published
26 September 2023
Volume
2 - 2023
Edited by
Patrick Roberts, Max Planck Institute for the Science of Human History (MPI-SHH), Germany
Reviewed by
Peter Rowley-Conwy, Durham University, United Kingdom; Marcello A. Mannino, Aarhus University, Denmark
Updates
Copyright
© 2023 Madgwick, Esposito and Lamb.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Richard Madgwick madgwickrd3@cardiff.ac.uk
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.