 Christopher S. Jazwa
Christopher S. Jazwa Tyler R. Molter
Tyler R. Molter Christopher T. Morgan
Christopher T. Morgan- 1Department of Anthropology, University of Nevada, Reno, NV, United States
- 2SWCA Environmental Consultants, Reno, NV, United States
Chronological and archaeofaunal data indicate that settlement of the earliest, low-density populations on California's Northern Channel Islands was conditioned by variables other than those affecting later, high-density populations. We use a variant of the Ideal Free Distribution (IFD) with considerations for low population densities to model early settlement on Santa Rosa Island (SRI). Early in time, individuals could have maximized their per-capita resource return at the mouth of any of SRI's 19 major drainages, so it was not necessary to distribute themselves in only those habitats with the highest potential return rate. Instead, while some individuals targeted high-ranked habitats, others settled at low-ranked habitats along the south coast that traditional IFD model variants predict would be first settled later. These habitats may have been targeted for other, less often considered environmental characteristics that might have been less important during periods characterized by higher population density or resource stress, perhaps including protection from prevailing northwesterly storms. During the relatively dry Middle Holocene, when population density increased and there was a greater focus on the high-ranked northwest coast, settlement intensity on the south coast did not increase and may have decreased. Later, as settlement at high-ranked habitats in-filled to the point that traditional IFD models predict the lowest-ranked habitats should be settled, there is evidence of population growth and reoccupation on the south coast. This study has implications for understanding initial colonization of new geographic areas, including larger regions in which the settlers did not have complete knowledge of all potential settlement locations.
1 Introduction
Human settlement can be influenced by a wide variety of factors, including resource distribution, seasonality, technological orientation, territoriality, defense, trade, cosmology, and social organization (e.g., Billman and Feinman, 1999; Kennett, 2005; Kowalewski, 2008; Winterhalder et al., 2010). Because of this causal complexity, a variety of models have been developed to isolate the drivers of settlement behavior by generating hypotheses about settlement distribution that can be tested with archaeological data in an effort to better understand these relationships (Jochim, 1976; Bettinger, 1980; Pilgram, 1987; Krist, 2001; Codding and Jones, 2013; Jazwa, 2015). Simple models, while eschewing realism, have the benefit of being clear, explicit, and generally applicable to a wide variety of contexts, whether patterns of change in individual geographic/cultural groups or broad cross-cultural/cross-regional variation (Winterhalder, 2002), with deviations from model predictions frequently yielding insight into the system in question (Clarke, 1972). For example, while resource maximization may govern the majority of economic decisions in some models, there may be exceptions in which this is not the case. At very low population densities associated with a settling population, people may be unable or uninterested in living in places where resource return is maximized. Instead, they need only to find a place that is good enough to provide more resources than they can consume (e.g., Jazwa et al., 2016b; Haas and Kuhn, 2019; Molter, 2021). Later, as population grows, settlement distribution should shift to one matching the maximizing conditions of the model. These deviations can be explained qualitatively or addressed by modifying the existing model or offering a competing model. A direct application of such a model variant, during a period of initial colonization of a large geographic area, would allow archaeologists to identify settlements and explain their distribution, while still maintaining the flexibility to understand settlement expansion later in time.
Fretwell and Lucas's (1969) Ideal Free Distribution (IFD) and Ideal Despotic Distribution (IDD) models have been applied with increasing frequency to archaeological and anthropological contexts (Weitzel and Codding, 2022). In these models, a single variable for habitat quality, “suitability,” is plotted against population density in a series of different habitats. In practice, suitability is often calculated using the availability of key resources like water, forage, game, net primary productivity, or by some other proxy for environmental yield (e.g., Kennett et al., 2009; Codding and Jones, 2013; Moritz et al., 2014; Yaworsky and Codding, 2018; Hanna and Giovas, 2022; Vernon et al., 2022; Jazwa and Jazwa, 2022). In general, as population density increases in a habitat, suitability decreases because of affiliated per-capita declines in resource availability. The difference between the IFD and IDD depends on how free individuals are to move between habitats (Winterhalder et al., 2010). In the IDD, individuals' freedom to move to preferred habitats is constrained by territorial defensive behaviors. The net effect of this distinction is that under IFD conditions, we would expect the highest population densities in the highest ranked habitats; in the IDD, we would expect lower population densities in high-ranked habitats and earlier occupation of lower-ranked habitats. Critically, the simplest formulations of these models do not allow for population-independent changes in habitat suitability, although recent models have expanded to include this eventuality. For example, environmental change may lead to a vertical shift in the suitability curve for some habitats (e.g., Jazwa et al., 2013). Still, modeling how habitat ranks might change over time introduces more complexity to the IFD, which makes applying these models in diachronic contexts challenging.
In this study, we apply the IFD to California's northern Channel Islands (NCI) to assess the degrees to which the initial, low population density settlement of the islands was affected by factors different than the environmental variables that govern settlement distribution later in time (Figure 1). The NCI were among the first settings where archaeologists applied the IFD (Kennett, 2005; Kennett et al., 2009). Since then, there has been a growing body of literature in which the IFD/IDD has been applied to cases around the world (e.g., Kennett et al., 2006; Allen and O'Connell, 2008; O'Connell and Allen, 2012; Giovas and Fitzpatrick, 2014; Prufer et al., 2017; Jazwa and Jazwa, 2017, 2022; Harvey, 2018, 2019; Yaworsky and Codding, 2018; Jazwa and Collins-Elliott, 2021; Collins-Elliott and Jazwa, 2022; Hanna and Giovas, 2022; Plekhov and Levine, 2022; Weitzel and Codding, 2022). Following the initial approaches on the NCI, variants of the model have been developed and their predictions have been tested with archaeological data, including radiocarbon dates and faunal data (Winterhalder et al., 2010; Jazwa et al., 2013, 2016b, 2019; Molter, 2021). In most cases, these data matched IFD and/or IDD predictions, indicating that settlement on the NCI was largely consistent with them, with some deviations. Jazwa et al. (2016b; also Jazwa, 2015), for instance, observed that relatively low-ranked habitats (i.e., those with low initial suitability) along the southern coast of Santa Rosa Island were settled earlier than expected, in some cases earlier than some of the highest-ranked habitats on the island. This is a clear deviation from the early settlement predictions of both the IFD and IDD. They offered the possibility that at low population density, it may not be necessary for individuals to target areas with the most abundant resources. Instead, other, less quantifiable factors like protection from prevailing storm systems by topographic features or relative propensity for warm, sunny days may influence settlement more than they would later during periods of higher population density and affiliated increases in resource stress. In this study, we move beyond this observation to alter assumptions and predictions of the IFD for the NCI and test an alternative version of the model that allows for a different settlement distribution at low-population density. Broadly, in this study, we seek to explain under what circumstances one might expect deviations from standard IFD predictions at low population densities, which has important implications for understanding settlement decisions among founding populations in a new area.
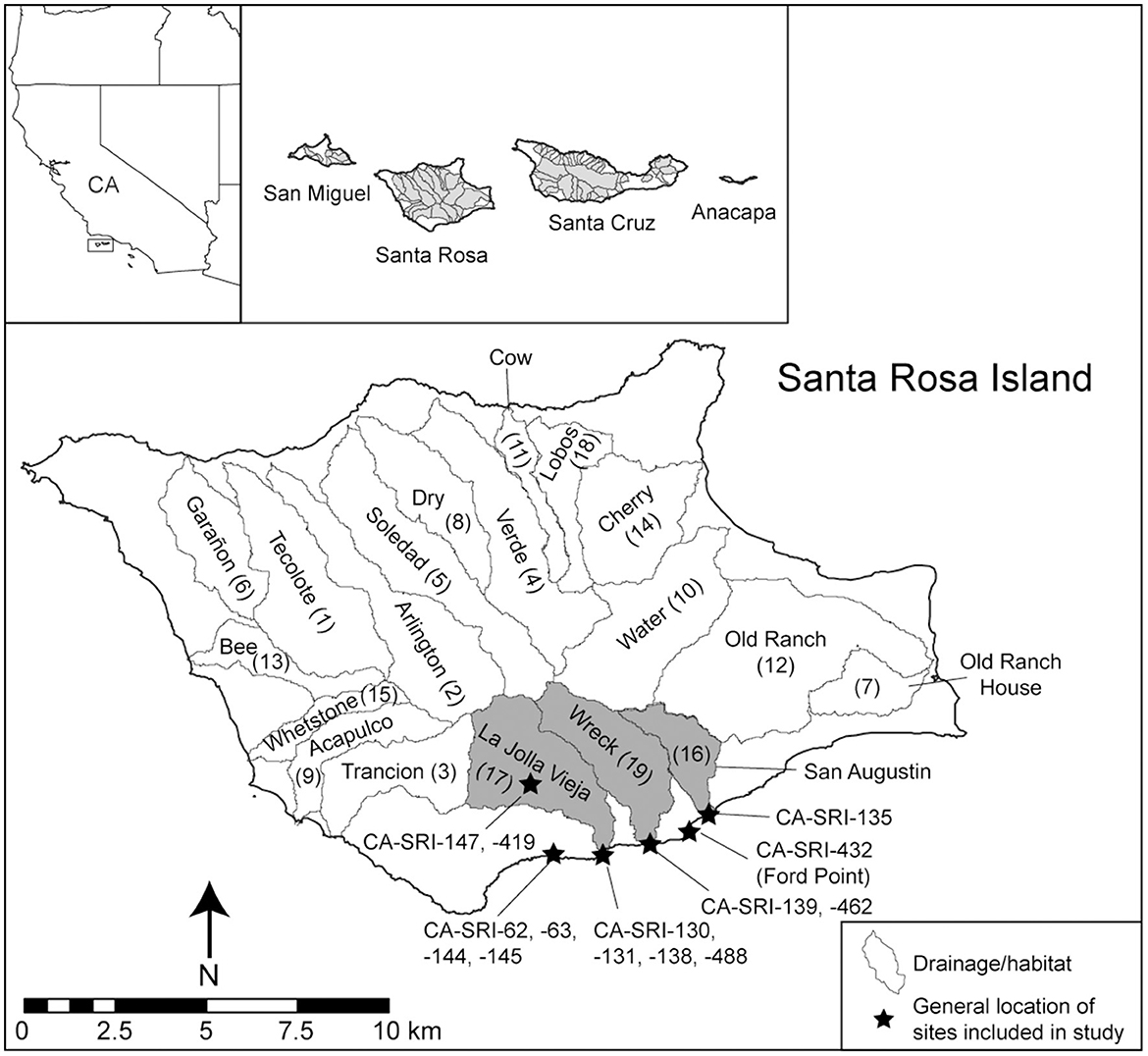
Figure 1. Santa Rosa Island. The 19 major drainages are outlined, labeled, and numbered in parentheses based on their IFD ranking from Jazwa et al. (2016b). The three low ranked drainages discussed in this paper are highlighted in gray. The general locations for the sites investigated as a part of this study are marked with stars.
1.1 Environment and culture history of the NCI
The NCI are a series of four islands located off the coast of California in the Santa Barbara Channel. From west to east, they are San Miguel, Santa Rosa, Santa Cruz, and Anacapa Islands (Figure 1). Sea level rise, beginning after the Last Glacial Maximum and continuing through the initial periods of human occupation of the islands, led to an ~85% reduction in land mass from the date of the first evidence of people on the NCI around 13,000 until ~6,000 cal BP, when marine transgression slowed (Kennett et al., 2008; Reeder-Myers et al., 2015). At the time of earliest occupation, lower sea levels would have exposed land connecting the four NCI into a single larger island, Santarosae (Shamalaḿa; Rick et al., 2023). The NCI and the surrounding marine ecosystem have provided islanders with a great abundance and diversity of subsistence resources throughout their history. This is especially true along what is today the northwest coast of Santa Rosa Island (SRI), which contains large, well-watered drainages that are especially resilient to periods of drought, as well as extensive rocky intertidal coastlines where shellfish can be collected en masse. It is for this reason that previous models have indicated that the habitats there would have been high-ranked, and therefore preferred settlement locations, through time (Kennett et al., 2009; Winterhalder et al., 2010; Jazwa et al., 2016a,b). This is likely among the reasons that the oldest dated site on the NCI, Arlington Springs (CA-SRI-173), is located there (Johnson et al., 2002).
However, the earliest sites on the NCI, those dating to the Paleocoastal Period (13,000–8,200 cal BP; terminal Pleistocene and Early Holocene), are not consistent with what Kennett (2005) and others (e.g., Jazwa, 2015) have described as permanent settlement sites, namely those containing dense, accumulated midden reflecting year-round occupation or consistent reoccupation. Instead, most early sites on the NCI that still survive tend to be more ephemeral occupations located along the edges of elevated coastal terraces (e.g., Erlandson et al., 2011, 2020; Rick et al., 2013), although some prominent sites containing characteristic bifacially flaked artifacts and points (crescents, Channel Islands Barbed, and Amol) have been identified closer to the coast (Erlandson et al., 2011; Erlandson, 2013). Most of the coastal regions that would likely have been targeted for early settlement, however, are now underwater, submerged by rising sea levels, making developing habitat suitability models for the terminal Pleistocene and early Holocene difficult (Kennett et al., 2008; Reeder-Myers et al., 2015). One recently investigated site at the mouth of Trancion Canyon, CA-SRI-347, contains midden deposits consistent with permanent occupation dating between ~11,000 and 8,500 years ago (unpublished radiocarbon dates obtained by Jazwa). This site is unique because it is located adjacent to an area of steep bathymetry where the shorelines have likely not moved far, providing further evidence that additional early permanent settlement sites likely exist in locations that have been submerged. Nevertheless, according to the IFD, habitats that were occupied earliest should continue to be occupied as settlement expands, so even if the earliest phases of occupation have been lost to rising sea levels, the earliest available data on settlement locations should be indicative of preferences in population distribution.
For this reason, the earliest clear evidence for sustained occupation of the coast of SRI dates to the beginning of the middle Holocene (8,200–4,200 cal BP). The model developed by Jazwa et al. (2016b: Figure 9) includes speculation as to what settlement distribution may have looked like before this time, but it was not possible to directly test model predictions for the very earliest settlement on the island. Other than CA-SRI-347, among the earliest dated permanent coastal residential sites on SRI are CA-SRI-187 at the mouth of Old Ranch and Old Ranch House Canyons (8,050–7,040 cal BP), CA-SRI-138 at the mouth of La Jolla Vieja Canyon (7,545–7,415 cal BP), CA-SRI-3 at the mouth of Tecolote Canyon (7,890–7,520 cal BP), and CA-SRI-4 at the mouth of Arlington Canyon (7,470–6,920 cal BP; Kennett, 1998, 2005; Winterhalder et al., 2010; Jazwa et al., 2013, 2016b; Jazwa, 2015). While marine transgression continued after the earliest occupation of these sites, it was limited enough that they would still have been adjacent to or within several 100 meters of the coast at any period for which there is evidence of occupation and could therefore be considered coastal settlement sites. Furthermore, with the exception of CA-SRI-3 and CA-SRI-4, which are associated with adjacent drainages, these sites are located relatively far apart from each other (7.5–10 km from each one to the next closest) and adjacent to different environmental zones, including drainages expected to be high-, middle-, and low-ranked for settlement (Jazwa et al., 2016b). Still, compared to some locations on the NCI, especially parts of northern Santa Cruz Island, all habitats on SRI are relatively high-ranked and able to support small populations (Kennett et al., 2009; Winterhalder et al., 2010).
During the Middle Holocene, populations increased and more permanent villages were established on SRI. These primary villages were mostly located on productive coastal habitats with access to fresh water from springs or streams (Kennett, 2005; Winterhalder et al., 2010; Jazwa et al., 2016a). In some cases, these water sources were seasonal. While the Early Holocene was generally a wet period in California and the Great Basin (Benson et al., 2001, 2002, 2013), climate records from throughout the region suggest that the middle Holocene was overall a relatively dry interval (e.g., LaMarche, 1973, 1974; Lindström, 1990; Thompson, 1992; Quade et al., 1998; Hughes and Graumlich, 2000; Benson et al., 2002; Kennett, 2005; Kennett et al., 2007). During drier periods, the large, resilient drainages of the northwest coast of SRI would have been especially valued locations for human occupation (Jazwa et al., 2016a). As the island population increased during the Middle Holocene, settlement focused on that part of the island, with people becoming more sedentary to take advantage of the northwest coast to access fresh water and its highly productive coastline. The archaeological record is consistent with this prediction, with evidence for Middle Holocene settlement at the mouth of Tecolote, Arlington, Soledad, Dry, Verde, Cow, and Lobos Canyons along the north coast (Kennett, 1998; Winterhalder et al., 2010; Jazwa et al., 2016b). This increased focus on north coast settlement is associated with a greater diversity of site types, with patterns of interior residential bases accompanying coastal settlements and special purpose shellfish harvesting and shucking sites (Kennett and Clifford, 2004; Kennett, 2005; Jazwa et al., 2015; Jazwa and Zoellner, 2024). Movement to the interior during the summer and fall may have also been related to fog water collection or terrestrial plant harvesting (Jazwa et al., 2015, 2016a).
During the Middle Holocene, red abalone middens come to characterize many sites across the NCI (Glassow, 2013). Red abalone (Haliotis rufescens) is a large marine gastropod that is among the highest return food species on the NCI (Braje et al., 2007). In southern California, they tend to live in subtidal environments, but may live in shallower, intertidal regions during periods of cooler water (e.g., Raab, 1992; Glassow et al., 2012). There is a debate over the reason for the appearance of these sites during this period. Glassow (1993; Glassow, 2013; Glassow et al., 2012) has argued for an environmental explanation, in which red abalones were more readily available because sea surface temperature (SST) was colder at this time (Kennett and Kennett, 2000; Kennett et al., 2007). Conversely, Braje et al. (2009; Erlandson et al., 2004, 2005, 2008) argue that red abalones were more common during the Middle Holocene because they were released from predation pressure from otters, who were increasingly hunted by humans. Both may have influenced the population of accessible abalone at this time, but notably, the location of red abalone middens on Santa Rosa Island do not necessarily follow the predictions of settlement rank models (Jazwa et al., 2015, 2016b; Jazwa and Zoellner, 2024). Put another way, the factors that influence where abalone are present may or may not track the variables that influence the location of permanent settlement sites. As Gamble (2017) points out, however, there is great diversity in the nature and use of the red abalone middens, including residential, special purpose, and potentially even feasting sties. Jazwa et al. (2015) found the highest concentration of red abalone in an otherwise relatively low-ranked habitat, Bee Canyon, suggesting that sites like CA-SRI-338 were used primarily for abalone processing before transportation back to permanent settlements elsewhere on the island.
From 4,200 to 1,500 cal BP, sea surface temperature became more variable (Kennett and Kennett, 2000). McFarland et al. (2025) have argued that this led to a change in the relative proportions of marine invertebrate taxa on the NCI, especially an increase in the average proportion of urchins compared to other kelp-dependent invertebrates. Furthermore, people living in the Santa Barbara Channel region developed the single-piece fishhook by around 2,500 cal BP (Rick et al., 2002), immediately preceding a dramatic increase in fish in the archaeological record (Glassow, 1993; Kennett, 2005; Jazwa et al., 2016b, 2020). Both of these changes are consistent with changes in the faunal record from the NCI. The general pattern of climatic amelioration during the Late Holocene (Kennett and Kennett, 2000; Kennett et al., 2007) coincided with increases in population density, the number of settlement sites, and the range of habitats occupied on the islands (Kennett, 2005; Kennett et al., 2009; Winterhalder et al., 2010; Jazwa et al., 2016b, 2019). By the start of the Medieval Climatic Anomaly (MCA; ~1,150–650 cal BP), for example, settlement had expanded to the mouths of all 19 large drainages on SRI (Jazwa et al., 2019). During the MCA, a period of extreme and persistent droughts occurred in the southwestern United States and NCI settlement structure changed dramatically. Overall island population densities continued to increase, though these appear to have been concentrated in a smaller number of large coastal village sites. These village sites were often in new locations, typically across the drainage mouths from preexisting settlement sites. These sites are often the large villages named in early 19th century mission records (Johnson, 1993; Glassow et al., 2010). The villages generally had high population densities and their political economy has been described as that of a complex chiefdom with an economy characterized by sophisticated sewn plank canoes called tomols and purple olive shell (Callianax biplicata) beads produced by craft specialists (e.g., Arnold, 1992, 2001; Kennett, 2005).
1.2 Settlement patterns and the ideal free distribution
The NCI have proven to be a productive test area for applying and refining applications of IFD/IDD models. Anthropological applications of the IFD first appear in the literature in Kennett's (2005) book in theoretical terms, as speculation as to how they could be applied to settlement on the NCI. After a similarly theoretical application for the settlement of Oceania (Kennett et al., 2006; Allen and O'Connell, 2008; revisited in O'Connell, 2020), the first formal test of the predictions of the model using ecological and chronological data was in Kennett et al. (2009). Winterhalder et al. (2010) followed that up by using a Bayesian statistical approach applying the chronology of initial settlement for drainages with dated sites to assess whether the suitability calculations by Kennett et al. (2009) appropriately weighted natural resource availability for the different habitats. Since that time, the IFD/IDD have been applied to a great diversity of contexts around the world, including elsewhere in North America, the Caribbean, Mesoamerica, Greece, and northwest Africa, among other places (e.g., Giovas and Fitzpatrick, 2014; Prufer et al., 2017; Jazwa and Jazwa, 2017, 2022; Harvey, 2018, 2019; Yaworsky and Codding, 2018; Jazwa and Collins-Elliott, 2021; Collins-Elliott and Jazwa, 2022; Hanna and Giovas, 2022; Plekhov and Levine, 2022; Weitzel and Codding, 2022). However, despite the initial studies on the NCI demonstrating that settlement distributions are largely consistent with IFD predictions, further studies have used the archaeological record there to refine nuances of the model. The NCI are particularly suitable for this purpose because they are bounded by water and therefore we can assess a complete set of potential habitats, settlement locations can generally be assumed to be individual points at the mouths of prominent drainages based on our knowledge of settlement on the coast, subsistence resources are limited in range (but not quantity) compared to mainland contexts, and there is a long history of archaeological research with good information about settlement history of most of the settlement locations. Since the initial applications, further research has been conducted to better understand the effects of population-independent environmental change (Jazwa et al., 2013), apply the zooarchaeological record as a proxy for population density (Jazwa, 2015; Jazwa et al., 2016b), and assess the transition from the IFD to IDD under periods of resource stress and population restructuring (Jazwa et al., 2019).
In this study, we use the NCI archaeological record to better understand how the IFD can be used to model settlement at very low population densities. A fundamental prediction of the IFD is that initial settlement will be in those habitats ranked highest (e.g., Jazwa et al., 2016b). As population density increases, settlement will expand to progressively lower ranked habitats, with successively lower-ranked habitats settled and “filling in” with human population until the ratio of people to resources in each habitat makes per capita returns in each habitat effectively equal. However, Jazwa et al. (2016b) found that the first prediction regarding initial settlement was only partially supported on the NCI. Some lower-ranked habitats, namely those along the south coast of SRI, were first settled much earlier than predicted by the model. In fact, settlement distribution best matched the IFD only beginning during the Middle Holocene when intensive settlement began to focus on the high-ranked, northwest coast of SRI. Jazwa et al. (2016b) attempted to explain this in two ways. First, they argued that the earliest settlements on the island were not necessarily the earliest permanent settlements; those were likely submerged by Pleistocene and Early Holocene sea level transgressions. Jazwa has obtained radiocarbon dates reflecting earlier evidence for permanent settlement at the mouth of Trancion Canyon, the third highest ranked drainage overall, supporting this idea. More importantly, however, Jazwa (2015) speculated that at low population densities, settlement decisions may be governed by different considerations than those used in previous iterations of IFD modeling. Population density was low enough that individuals could maximize their return at a wide variety of habitats. Unlike later in time, there was no need for people to position themselves in habitats that would maximize overall resource return. Rather, they could have chosen from a large number of habitats that all could have provided more subsistence resources than what population at that size can exploit. In the case of the NCI, when considering all potential habitats for Santa Cruz, San Miguel, and SRI, there are no SRI habitats among the lowest ranked 1/3 of all drainages across the islands. Low-ranked drainages that may be resource limiting, even at low population densities, are confined to Santa Cruz and San Miguel Islands (Winterhalder et al., 2010).
Therefore, it may be the case that settlement decisions at low population densities may not be guided by efforts to maximize resource return. Instead, in those cases, the components of the model associated with calculating habitat suitability may only be relevant to the extent that resources meet a certain threshold. In those cases, settlers may choose between any of those habitats based on a different set of criteria that are otherwise not considered of high importance once population density increases to the point that it is beneficial to maximize resource return. This decision may be based on less quantifiable considerations as simple as propensity for warm, sunny days and/or less frequent storms. Alternatively, a common assumption of the IFD is one of complete knowledge of the region under consideration, including sufficient information to accurately appraise and compare relative suitabilities for all habitats (Winterhalder et al., 2010: 471; Assumption f). This may not necessarily be the case, especially in a large geographic area, and a settling population may be willing to settle down in one of the first habitats that they encounter that are good enough to provide the subsistence resources they need in quantities that are more than they can consume, even if other, nearby habitats allow for greater resource maximization (Haas and Kuhn, 2019).
1.2.1 A low density, target-oriented IFD variant for SRI
To more realistically model how a low-density hunter-gatherer population might be expected to target different habitats, we build on the logic of target-oriented rather than maximizing economic models (Bordley and LiCalzi, 2000; Schmidtz, 2004; Bettinger, 2009; Morgan, 2009). In target-oriented models, economic decisions are based on meeting some minimum threshold of production, return, or yield. For hunter-gatherer populations, yield is often measured in calories, with the target being the minimum number of calories a population requires over a give timespan. As an example, consider a population of 20 individuals: though caloric requirements vary by age, sex, body size and activity level (Ainsworth et al., 1993), such a population would require, on average, ~2,000 calories per person per day to stay healthy, or 40,000 calories per day for the entire population. If a given habitat can produce enough resources so this target can be met and population density remains constant, then such a habitat can be said to satisfy the minimum subsistence requirement for the population. In a generally highly productive environment like the NCI, we posit that even relatively low-ranked habitats could have met such targets and therefore been just as attractive to early NCI occupants as more productive, ostensibly high-ranked habitats.
In practice, we maintain the same habitat rankings through time, but we relax two relevant assumptions of the model, namely complete knowledge of the landscape at the time of initial settlement and that individuals attempt to maximize resource return (opting instead to meet resource return targets). We also add predictions for low population densities and modify the existing predictions for initial settlement. After settlement reaches an unspecified population density threshold, the distribution should look more like the traditional formulation of the IFD. Importantly, this transition is likely to be gradual. Early settlement sites at lower ranked habitats may persist as settlement intensity increases at high-ranked habitats. This may continue at low levels temporarily or until settlement at the low-ranked habitat is eventually predicted to be settled following system-wide increases in population density.
Here, we adopt the hypothesis that at low population density on SRI, settlement should not be organized to maximize overall economic return, but instead to take advantage of other benefits to habitats reaching an unspecified resource return threshold. For consistency with the existing NCI literature, we modify the model predictions given in Jazwa et al. (2016b; following Kennett et al., 2009; Winterhalder et al., 2010; Jazwa et al., 2013) to follow from this hypothesis. We also use the same resource weight calculations given by Jazwa et al. (2016b) as they have been shown to be largely appropriate for characterizing habitat suitability, at least once overall population gets sufficiently large. In our new formulation, the predictions are: (1) initial settlement can be at any habitat as long as it meets a minimum suitability threshold; (2) there should be a transition associated with increasing population density to a focus on the higher ranked habitats, although early occupations of low-ranked habitats may have persisted resulting from a founder effect (Brantingham and Perreault, 2010; Jazwa et al., 2016b: prediction 1 or J16, P1; Haas and Kuhn, 2019); (3) over time, there should be evidence for resource depression in high-ranked habitats, reflecting increased population density, while sites from low-ranked habitats that continue to be occupied may or may not show a similar pattern, as population density increase is less likely there (J16, P2); (4) dates of settlement, resettlement, or increase in population density for lower ranked locations should coincide with periods of resource depression at higher ranked locations (J16, P3); (5) settlement at the highest ranked locations should persist even as settlement expands to other sites (J16, P4); (6) extrinsic environmental change can depress suitability (J16, P5); and (7) technological developments can influence habitat suitability and settlement rankings (J16, P6).
In this study, we focus on the south coast of SRI (Figure 1), an area that contains the 16th (San Augustin Canyon), 17th (La Jolla Vieja Canyon), and 19th (Wreck Canyon) ranked drainages of 19 overall on the island in the calculations presented by Jazwa et al. (2016b). We focus on these habitats precisely because they are low-ranked and should be first settled later than radiocarbon dates indicate they are. These drainages contain less water than those of the northwest coast and are less resilient to periods of drought because they are relatively small, steep, and prevailing northwesterly weather systems are blocked by the central ridgeline of the island, creating something of a rain shadow (Jazwa et al., 2016a). Additionally, while productive rocky intertidal coastline and offshore kelp forests are present near these habitats, they are more spatially limited than elsewhere on the island (Kennett et al., 2009; Jazwa et al., 2016b). La Jolla Vieja Canyon is the one that Jazwa (2015) recognized as being settled anomalously early based on standard IFD predictions. Notably, in rankings by Kennett et al. (2009) and Winterhalder et al. (2010) including all the NCI, these three drainages are not near the bottom of the 46 ranked drainages total, with the lowest of the three, San Augustin, ranked 27, and all likely meeting the minimum threshold for resource availability. We rely on chronological and faunal data presented by Jazwa et al. (2016b, 2019) for comparisons with other habitats on the island to test model predictions, but we present new or updated data from sites along the south coast to address the new formulation of the model.
2 Materials and methods
2.1 Field sampling
Shell midden deposits included in this study were excavated in two phases. In 1996 and 1999, Douglas Kennett and Don Morris excavated four 25 × 25 cm units from CA-SRI-130, two 25 × 25 cm units and one 50 × 50 cm unit (Unit 3) from CA-SRI-131, and four 25 × 25 cm units from CA-SRI-1351 (Figure 1). In August 2019, we excavated one 25 × 25 cm unit from CA-SRI-139 from an eroding cliff-face with Richard-Patrick Cromwell and Gilbert Unzueta. We excavated in arbitrary 10 cm levels, separating materials from clearly defined strata and collecting radiocarbon samples in the field from the wall profile after excavation. We also included previously published comparison data for one 25 × 25 cm unit each from CA-SRI-131 and−488, and two 25 × 25 cm units from CA-SRI-138 (Jazwa, 2015; Jazwa et al., 2016b). We excavated using the same methods as CA-SRI-139. Kennett and Morris also obtained radiocarbon dates that we include in this analysis from further excavation units at CA-SRI-147 (Braje et al., 2007), CA-SRI-62, and CA-SRI-432, and field sampling from eroding escarpments at CA-SRI-419 and CA-SRI-462. We collected samples from eroding escarpments at CA-SRI-63, CA-SRI-144, and CA-SRI-145.
2.2 Radiocarbon chronology
We include 38 radiocarbon dates from the south coast of Santa Rosa Island in this analysis, 12 of which are new dates obtained for this study. Of those dates, 24 are from sites for which we are including excavation data and the remaining 14 are from nearby sites. For the 12 dates new to this study, we cleaned the samples, subjected them to a 50% hydrochloric acid etch, hydrolyzed them to generate carbon dioxide at the UNR Human Paleoecology and Archaeometry Laboratory, and submitted them for measurement for radiocarbon at the Pennsylvania State University AMS Laboratory. Original references for the 26 previously published dates are provided in Table 1.
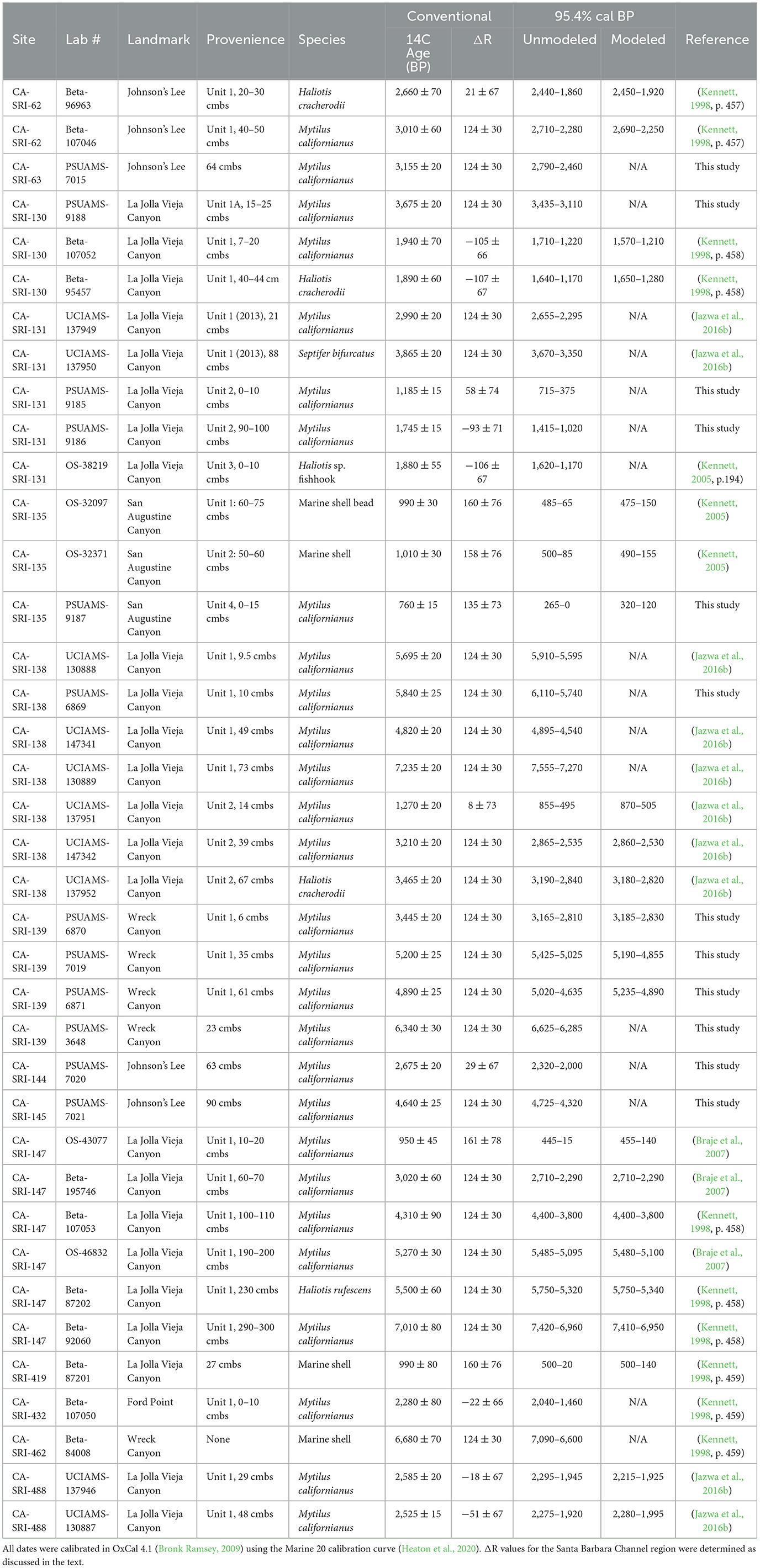
Table 1. Radiocarbon dates from the sites in this study, including excavated and unexcavated sites.
We calibrated all the available dates for the south coast of Santa Rosa Island in OxCal 4.4 (Bronk Ramsey, 2009) using the Marine20 calibration curve (Heaton et al., 2020). We used a variable ΔR value for the Santa Barbara Channel region for samples with dates of < 2,500 radiocarbon years before present (Hendy et al., 2013) and an estimated ΔR value of 124 ± 30 for older samples obtained from six historical, pre-bomb Callianax biplicata shells. All ΔR values have been corrected for the Marine 20 calibration curve (Sean Hixon, Brendan Culleton, and Douglas Kennett, personal communication, 2021).
For our analysis, we assess settlement expansion following a modified version of Arnold's (1992) chronological framework to better distinguish important periods of change during the Late Holocene: Early Period (7450–2550 cal BP), Middle Period (2550–800 cal BP), Middle to Late Period Transition (MLT; 800–650 cal BP), and Late Period (650–168 cal BP). Notably, the droughts of the MCA occurred at the end of the Middle Period through the MLT (i.e., Late Middle/MLT). The Early Period was not uniform, so we distinguish the periods before and after the Middle and Late Holocene boundary at 4200 cal BP for the purposes of this study.
2.3 Faunal analysis
After excavation, we size sorted the midden through nested 1/2”, 1/4”, and 1/8” screens. Materials from sites excavated in the 1990s were preliminarily sorted at University of California, Santa Barbara and California State University, Long Beach by trained undergraduate students under Kennett's (1998; 2005) supervision. We similarly size-separated the material from CA-SRI-139 prior to sorting the midden to taxa. We sorted all the 1/2” material to artifact type or taxa, in most cases assigning shellfish to the species level. For the purposes of this analysis, we do not more precisely identify fish and sea mammal bone. For the smaller material, we similarly sorted subsamples, 100 g for 1/4” and 15 g for 1/8”, excluding non-cultural rock. For residual bulk midden samples, we sorted for diagnostic artifacts, bone, charcoal, ochre, Mytilus californianus hinges, and Callianax biplicata shell. We used the total weight of bulk midden and proportions of the subsamples to estimate overall taxonomic contributions for each level. We obtained the data published in Jazwa (2015) and Jazwa et al. (2016b) following the same protocols. We combined data from stratigraphic levels of the same units into associated contexts, with a single excavation unit comprising either one or two contexts depending on radiocarbon date sequences and similarities in faunal constituents between levels. For comparison, we normalized faunal weights in each context to one cubic meter. We calculated the Shannon-Weaver diversity index (Shannon and Weaver, 1949), evenness, and taxonomic richness for shellfish in each context using these compiled shell weights.
We use meat weight calculations summarized by Rick (2004; see also Colten, 2001; Jazwa et al., 2019) to estimate the relative contribution of shellfish, fish, sea mammal, and bird to the diet for each of these components. Meat weight calculations have important limitations, especially for sea mammals that are likely to be field processed. Not all elements have the same meat to bone ratio, which might bias the contribution of sea mammal to the diet (Mason et al., 1998; Gifford-Gonzalez and Hildebrandt, 2012). Nevertheless, the great difference between the ratio of bone/shell to meat for fish, sea mammal, shell, and bird makes it difficult to compare them based on faunal weight, so we cautiously apply meat weights for this comparison. Furthermore, the fact that all the sites considered in this analysis are located on the coast likely reduces the extent of field processing.
Finally, we estimated mussel size for those incomplete shell fragments with the umbo present using the equation provided in Singh and McKechnie (2015)2. For complete shells larger than 20 mm long or fragments of shells estimated to be >20 mm, we obtained the average shell length by context. To avoid counting individual mussels twice, we only included values from those shells of the side with a higher count from each stratigraphic level (i.e., left or right valves). We also calculated the relative proportion of mussels within each assemblage that were < 20 mm long.
3 Results
3.1 Chronology
Based on the new and previously published radiocarbon dates, it is clear that permanent settlements were established at the mouths of both La Jolla Vieja and Wreck Canyons earlier than would be predicted by the IFD. All dates are given as 95.4% calibrated ranges. Early occupation at CA-SRI-138, Unit 1 dates to 7555–7270 cal BP (Table 1; Figure 2). Among permanent settlement sites, only CA-SRI-187 at the mouth of Old Ranch and Old Ranch House Canyons (Jazwa et al., 2013), CA-SRI-3 at the mouth of Tecolote Canyon (Winterhalder et al., 2010), and CA-SRI-347 at the mouth of Trancion Canyon (Jazwa, unpublished radiocarbon dates) date earlier. All three of those drainages are much higher ranked than La Jolla Vieja. There is a reversal in the radiocarbon dates associated with the terminal occupation of CA-SRI-138, Unit 1, with the sample from 49 cmbs dating to 4895–4540 cal BP, but samples from 9.5 and 10 cmbs date to 5910–5595 and 6110–5740 cal BP, respectively. This suggests some level of mixing in the upper component of this unit, perhaps resulting from erosion from an upslope, adjacent area of the site. However, settlement at Wreck Canyon is also early. A radiocarbon date from CA-SRI-462 places occupation at 7090–6600 cal BP, and one from CA-SRI-139 dates to 6625–6285 cal BP. This date is not from the excavation unit at the site. The bulk of occupation from that unit was from late in the Middle Holocene, dating from 5235–4890 to 5190–4855 cal BP. A lower density of deposits indicates persistent occupation at the site as late as 3185–2830 cal BP.
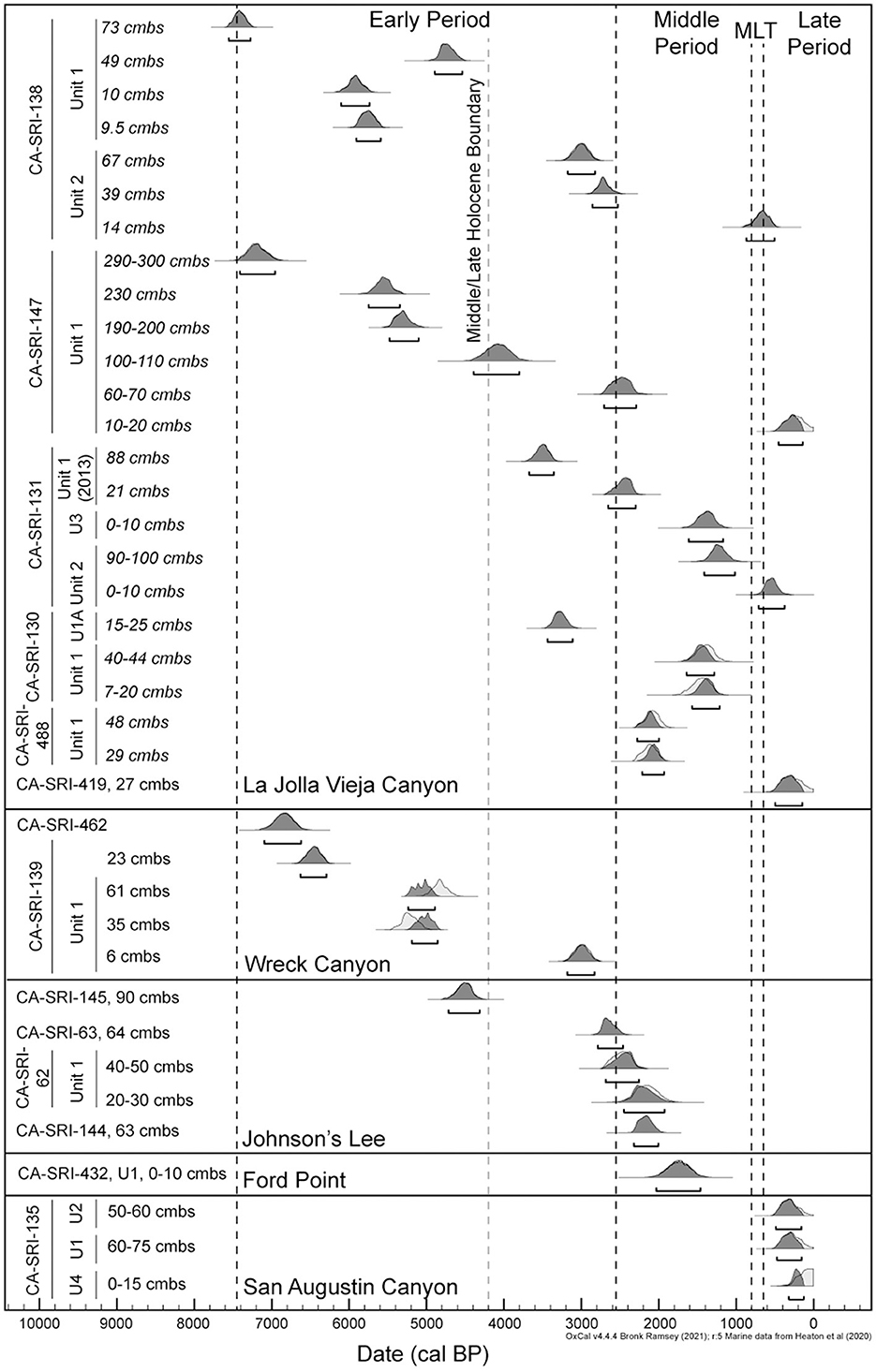
Figure 2. 95.4% calibrated ranges for the radiocarbon dates from sites included in this analysis. All dates were calibrated in OxCal 4.1 (Bronk Ramsey, 2009) using the Marine 20 calibration curve (Heaton et al., 2020) and the ΔR values as discussed in the text. When possible, the dates are modeled in OxCal to account for stratigraphy within excavation units.
Of the three drainages considered in this study, La Jolla Vieja exhibits the greatest evidence of persistent occupation through the Middle and Late Holocene. CA-SRI-147, the radiocarbon and faunal record of which was reported by Braje et al. (2007), was occupied as early as 7410–6950 cal BP and persisted until 455–140 cal BP. This site is a rockshelter with at least three meters of dense cultural deposits, but it is ~1 km up the drainage, meaning it is not a coastal settlement. Nevertheless, it was clearly a focus of occupation for a long time. CA-SRI-130 and CA-SRI-131 are located on either side of the drainage mouth, and both have evidence for occupation dating to the beginning of the Late Holocene, with CA-SRI-131 occupied as early as 3670–3350 cal BP and CA-SRI-130 as early as 3435–3110 cal BP. Occupation of CA-SRI-130 ended before the start of the MCA, but it persisted at CA-SRI-131 until 715–375 cal BP. There is no evidence for occupation of Wreck Canyon after the low density of occupation at CA-SRI-139 dating to the beginning of the Late Holocene.
Though not directly associated with any of the three drainages, there is a Middle Holocene presence at Johnson's Lee to the west, with CA-SRI-145 dating as early as 4725–4320 cal BP. The other dates currently available from Johnson's Lee correspond to the Late Holocene, with the latest from CA-SRI-144 dating to 2320–2000 cal BP. However, Johnson's Lee is the likely location of the historically-named village of Nilal'uy, perhaps at CA-SRI-62, and therefore was likely occupied until the time of Spanish contact (Kennett, 2005; Glassow et al., 2010). Unfortunately, that part of the island and that site were heavily impacted by the construction, use, and razing of an Air Force Base that was in use from 1952 to 1963, before modern archaeological investigations were possible. CA-SRI-432, at Ford Point between San Augustin and Wreck Canyons, was occupied from 2040 to 1460 cal BP. Finally, CA-SRI-135 at the mouth of San Augustin Canyon, is the potential location of the historically-named village of Helewashkuy (Kennett, 2005; Glassow et al., 2010). The radiocarbon dates are consistent with this, with the earliest dating to 490–155 cal BP. Dates from the four units from the site are all within the last 500 years before present and the units vary in depth but are highly dense. It is possible other sites near the mouth of San Augustin may date earlier, but we have not yet been able to obtain earlier dates from any of them.
3.2 Faunal data
We present data from 19 distinct components from six different sites originally collected by Jazwa (2015) and Molter (2021). Over time, there is a general decrease in the relative contribution of shellfish to the diet along with an increase in sea mammal and especially fish (Table 2; Figure 3; Supplementary Table 1), a pattern common on the NCI (see Glassow, 1993; Colten, 2001; Jazwa et al., 2016b, 2020; Hagerty, 2024). Overall, the densest estimated meat weights are from the latest deposits, the Late Middle/MLT occupation of CA-SRI-131, Unit 2 and the Late Period village components from CA-SRI-135, along with the Middle Period component from CA-SRI-138, Unit 2. This is consistent with the late expansion of settlement to low ranked habitats discussed in Jazwa et al. (2016b; also Winterhalder et al., 2010), and the development of a limited number of high-density villages during the Late Period, with locations influenced by territorial behavior (Jazwa et al., 2019). The large amount of fish at CA-SRI-131, Unit 2 (73.2%) and at CA-SRI-135 (more than 85% of total meat weight in each of Units 1–3) is consistent with late settlement sites elsewhere on the island that relied on fish to support higher population densities (e.g., Jazwa et al., 2020).
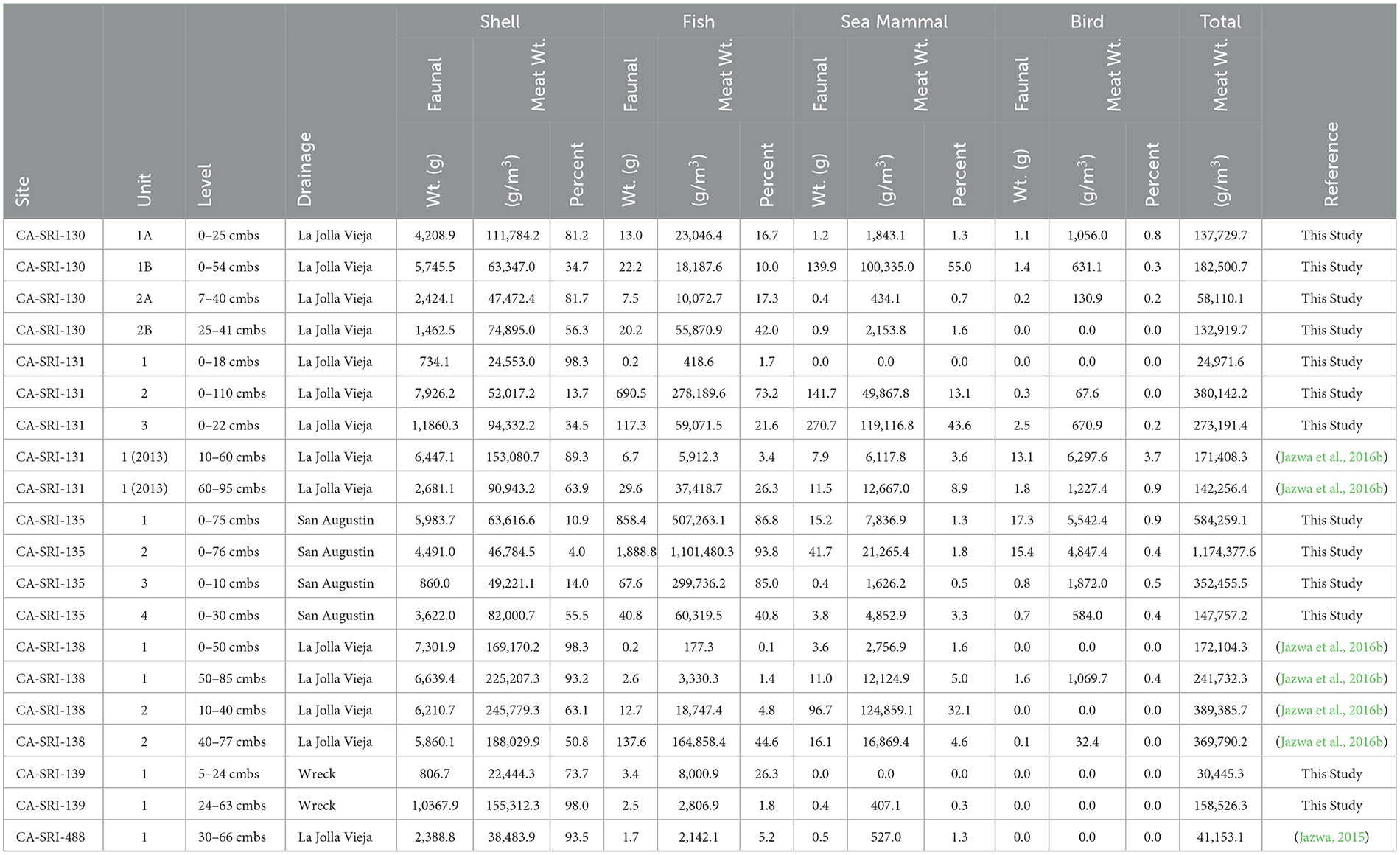
Table 2. Calculated faunal weight, meat weight density, and percent of total meat weight by major faunal component for all excavated sites.
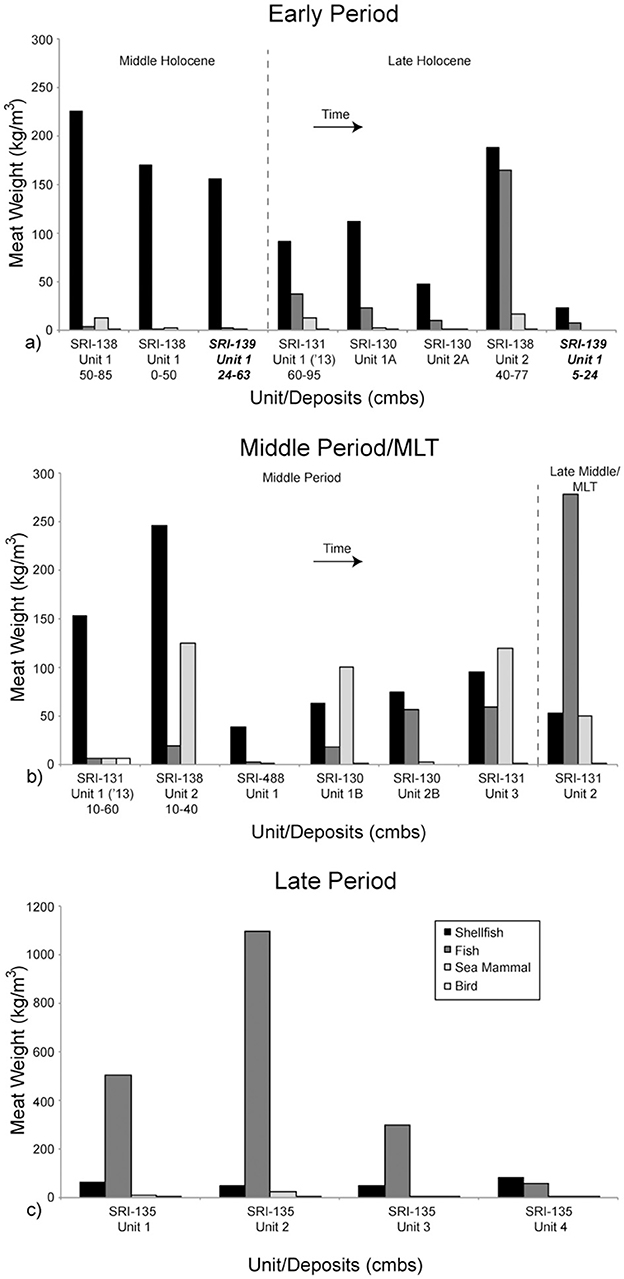
Figure 3. Meat weight density of the distinct components of each excavation unit. Time is generally represented as left to right, and top to bottom from (a) to (c). All components in this figure (a, b) are from La Jolla Vieja Canyon except the two from CA-SRI-139, which are in bold and italicized. They are from Wreck Canyon. All components in this figure (c) are from San Augustin Canyon.
During the earliest occupation dating to the beginning of the Early Period, however, there is nevertheless dense occupation at CA-SRI-138, Unit 1 near the mouth of La Jolla Vieja Canyon, with total estimated meat weight density of 241.7 kg/m3 in the deeper component and 172.1 kg/m3 in the upper component. These components have the 8th and 10th highest meat weight densities of all 19 dated components we have for the south coast of the island, meaning that meat weight density does not appear to increase in a unilinear trajectory over time. The later deposits at CA-SRI-138, Unit 2 increase in density into the later part of the Early Period and the Middle Period, suggesting an increase in population density at that location over time. However, there is a gap in occupation between the latest date of occupation at Unit 1 (4895–4540 cal BP) and the earliest date of occupation at Unit 2 (3180–2020 cal BP). This may indicate that the site was temporarily depopulated and later reoccupied in a manner consistent with settlement expansion to lower-ranked habitats over time, per the IFD.
CA-SRI-139, at the mouth of Wreck Canyon, was also occupied relatively early, with its most intensive occupation phase during the Middle Holocene. While its deposit was not as dense as the earliest occupation at La Jolla Vieja Canyon, the overall meat weight density of the denser, earlier phase at the site was 12th overall at 158.5 kg/m3. However, both CA-SRI-139 and CA-SRI-138, Unit 1 indicate an almost complete reliance on shellfish, with very little fish or sea mammal in the record. In fact, the three Middle Holocene occupation phases associated with these sites are among the five densest in terms of shellfish meat weight density (2nd, 4th, and 5th) among those considered here. Clearly, then, there is an important focus on these habitats early in time that persists, but only grows slowly through time during the Middle Period, MLT, and Late Period.
When focusing on the particular shellfish species present at the components considered here, it is clear that California mussel (Mytilus californianus) is the dominant species through time (Table 3; Figure 4). This is not unexpected as it follows the pattern commonly observed in faunal analysis of most sites on the NCI (Colten, 2001; Rick, 2007, 2011; Braje et al., 2007; Erlandson et al., 2009; Jazwa et al., 2015, 2016b, 2019; Jazwa and Zoellner, 2024). The Middle Holocene component of CA-SRI-139 has an especially high density of California mussel (251.2 kg/m3), second only to the Middle Period occupation of CA-SRI-138, Unit 2. Black turban snails (Tegula funebralis) are also especially prominent at CA-SRI-139, far more abundant than at any other site in the study area (118.2 kg/m3), consistent with some contemporaneous sites on the north coast of SRI (Jazwa et al., 2015). Purple urchin (Strongylocentrotus purpuratus) and platform mussel (Mytilisepta bifurcata) are among the other prominent species in the study area.
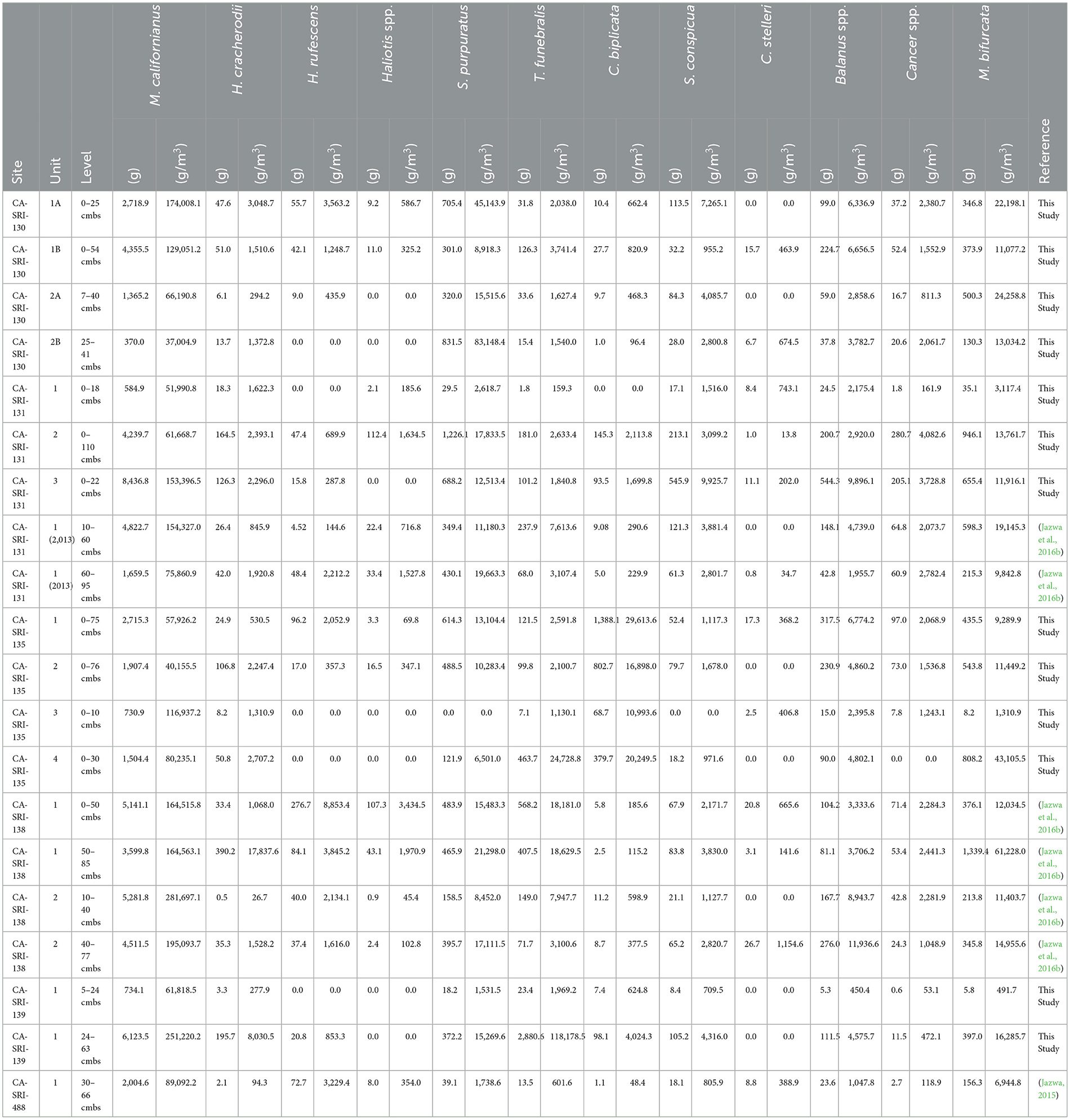
Table 3. Calculated shell weight and shell weight density of major species for all excavated sites.
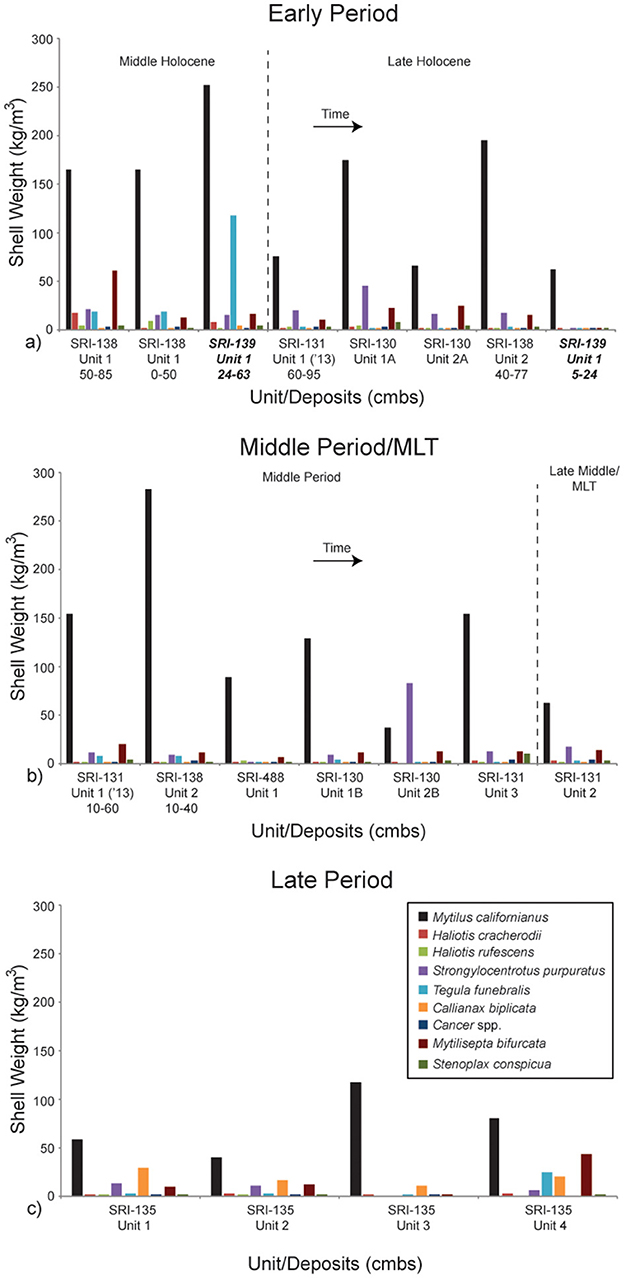
Figure 4. Shell weight density of the nine most important dietary constituents. Components are separated and the graphs represent time as in Figure 3. Time is generally represented as left to right, and top to bottom from (a) to (c). All components in this figure (a, b) are from La Jolla Vieja Canyon except the two from CA-SRI-139, which are in bold and italicized. They are from Wreck Canyon. All components in this figure (c) are from San Augustin Canyon.
Diversity of shellfish taxa tells an interesting story of dietary change on the south coast of SRI (Table 4; Figure 5). Diversity is highest during the Late Period, which makes sense considering that was the time when populations were highest (greatest at CA-SRI-135, Unit 2; 1.8). However, diversity was also relatively high early in time during the Early Period. The decrease in shellfish diversity during the Middle Period does make sense in the context of an increasing focus on fish and sea mammal at the time. However, the high diversity at the earliest occupation of CA-SRI-138, Unit 1 (1.4), indicates that the population targeted a wide range of species in moderate amounts early in time. This is indicative of a substantial population, although not nearly as large as the density of people present later in time. Taxonomic richness is relatively high early, with 16 taxa present at the Middle Holocene deposits from CA-SRI-139. There is variability in richness later toward the end of the Early Period and throughout the Middle Period, despite overall lower diversity. This suggests that while even though people at some sites like CA-SRI-488 targeted a relatively high number of different taxa (19), the focus was on a relatively small number of species, as reflected by the diversity score of 0.7. During the Late Period, as represented by CA-SRI-135, there is generally a high taxonomic richness, 20 at Unit 1 and 17 at Unit 2, although it is lower at the relatively shallow Units 3 and 4.
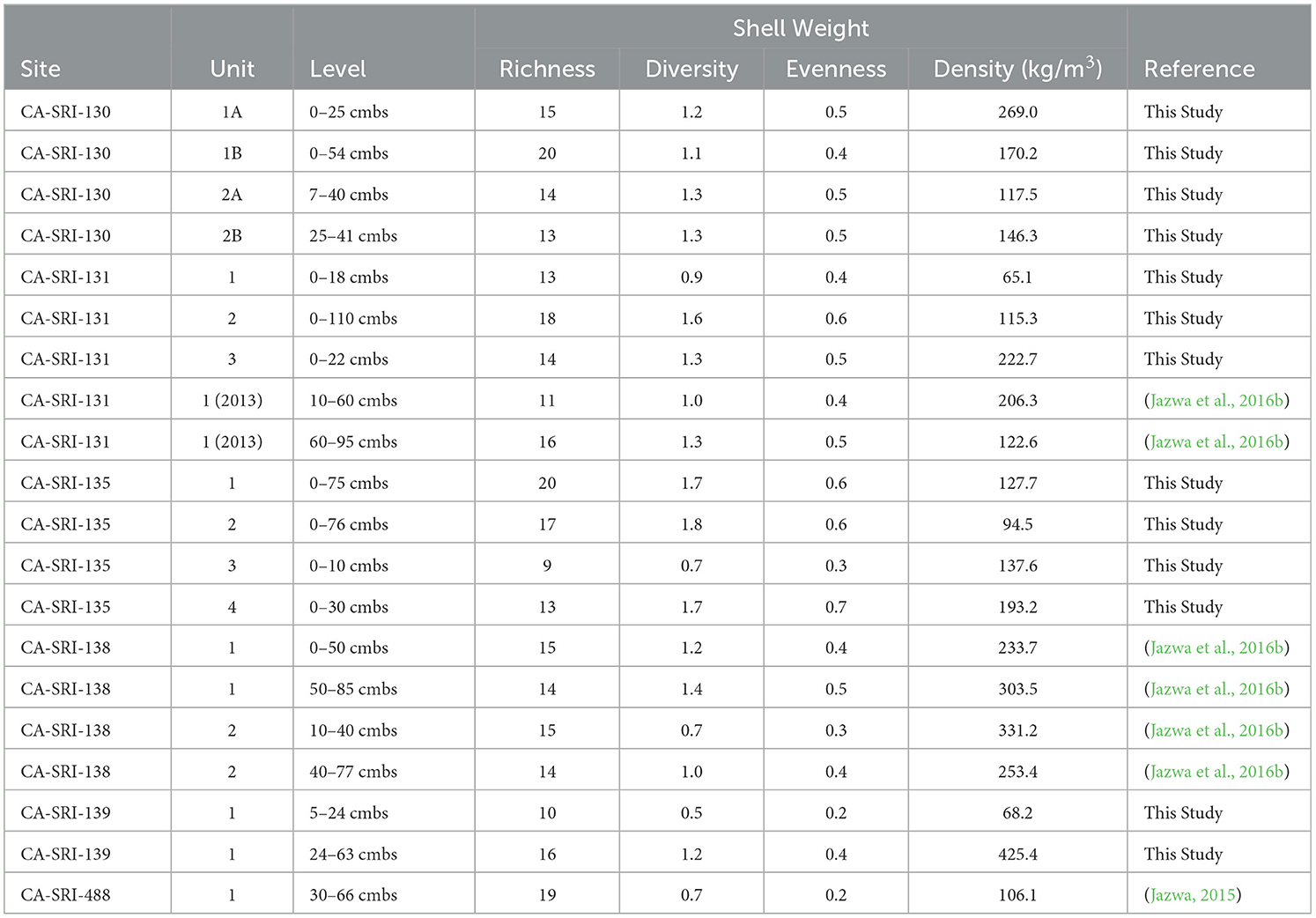
Table 4. Taphonomic richness, Shannon-Weaver diversity index, evenness for shellfish weights for all excavated sites.
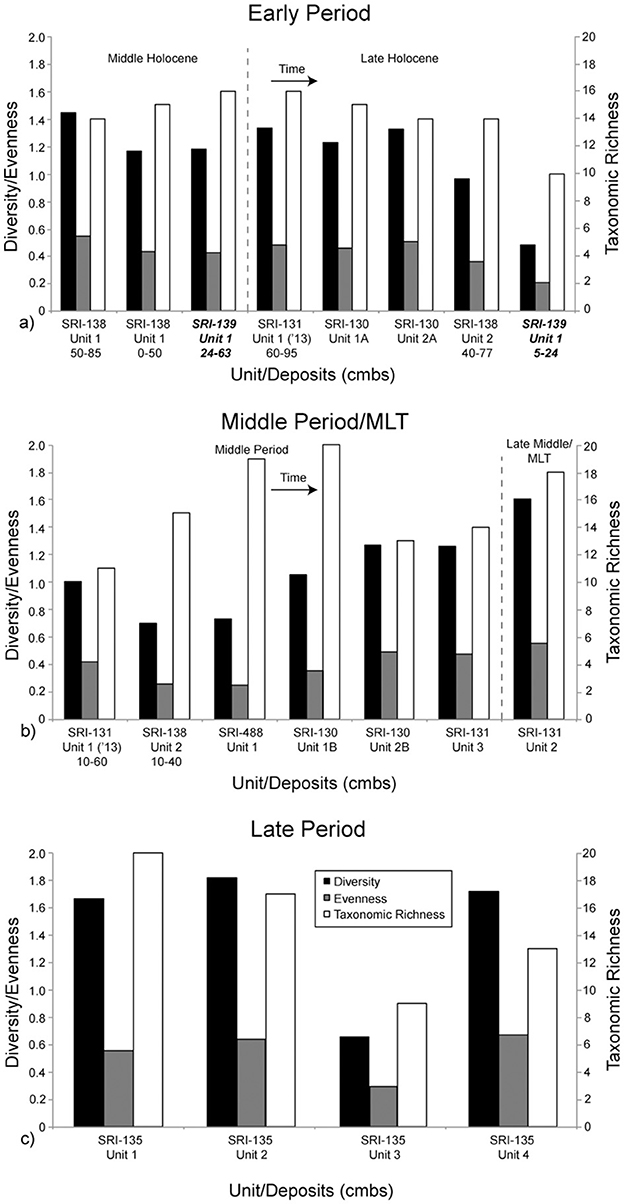
Figure 5. The Shannon-Weaver diversity index, evenness, and taxonomic richness. Components are separated and the graphs represent time as in Figure 3. Time is generally represented as left to right, and top to bottom from (a) to (c). All components in this figure (a, b) are from La Jolla Vieja Canyon except the two from CA-SRI-139, which are in bold and italicized. They are from Wreck Canyon. All components in this figure (c) are from San Augustin Canyon.
Finally, we provide the estimated average mussel size by context and relative proportion of mussels within each assemblage that were < 20 mm long (Table 5; Figure 6). This provides a proxy for the likelihood that mussels were collected by plucking individual shells or stripping them from the rock. The latter is more likely to occur when fewer large, high-return mussels are present (Jones and Richman, 1995; Bettinger et al., 1997). In general, average mussel size is highest at the earliest occupied components, including CA-SRI-138, Unit 1 (49.9) and CA-SRI-139 (54.3). These are consistent with average sizes for components with high average mussel size elsewhere on the island (Jazwa et al., 2016b). Overall, mussel size decreases later in the Early Period and into the Middle Period, with the highest proportion of small mussels during the Middle Period. This suggests evidence for overpredation on mussels around the time that people adopt more fish and sea mammal in their diet. As these alternatives become more prominent, mussel size recovers, although never to the average size reflected early in time. Average size at CA-SRI-135 varies from 43.1 to 46.6 mm.
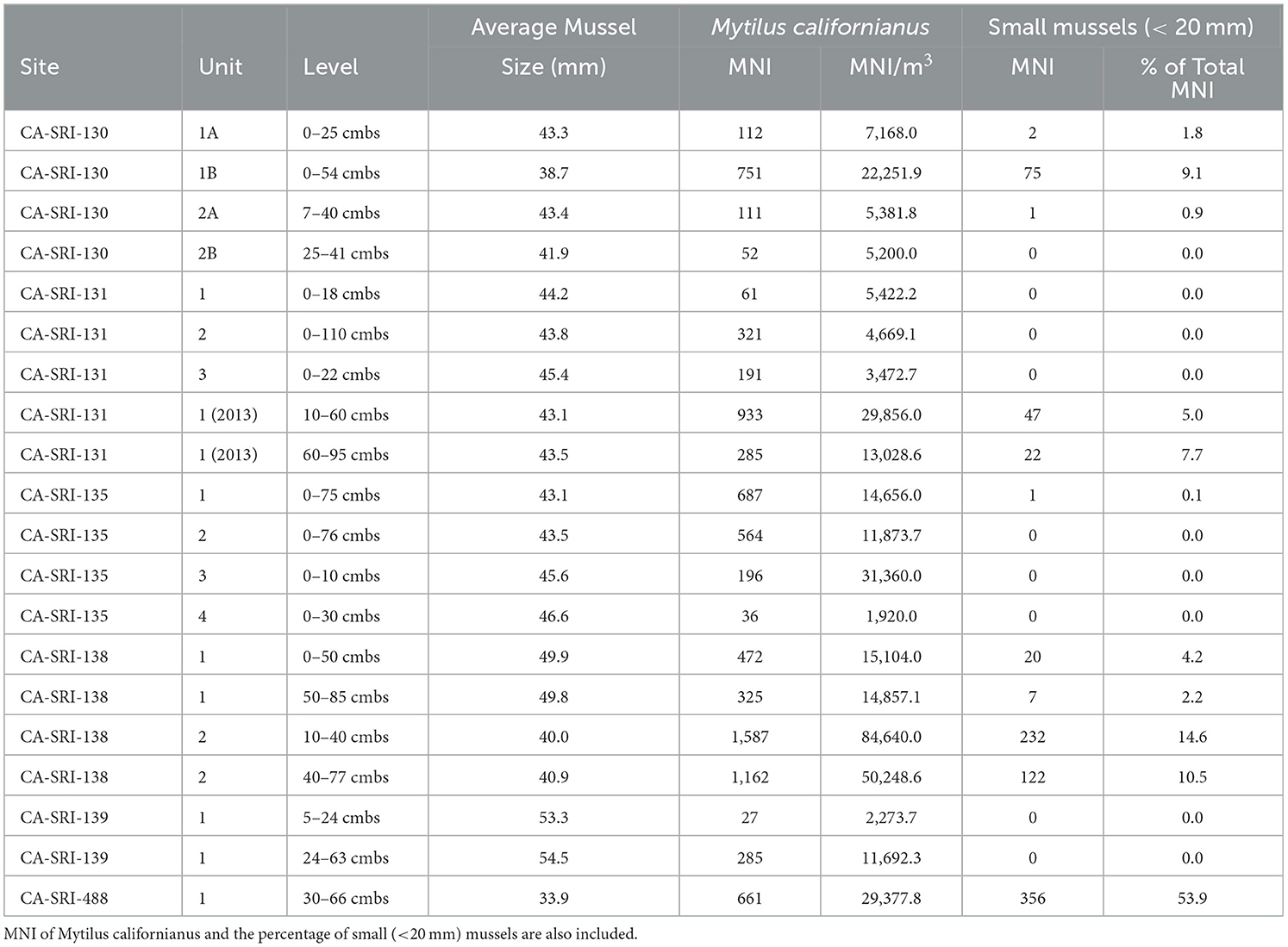
Table 5. Mussel size, estimated for the relationship between umbo size and shell length given in Singh and McKechnie (2015).
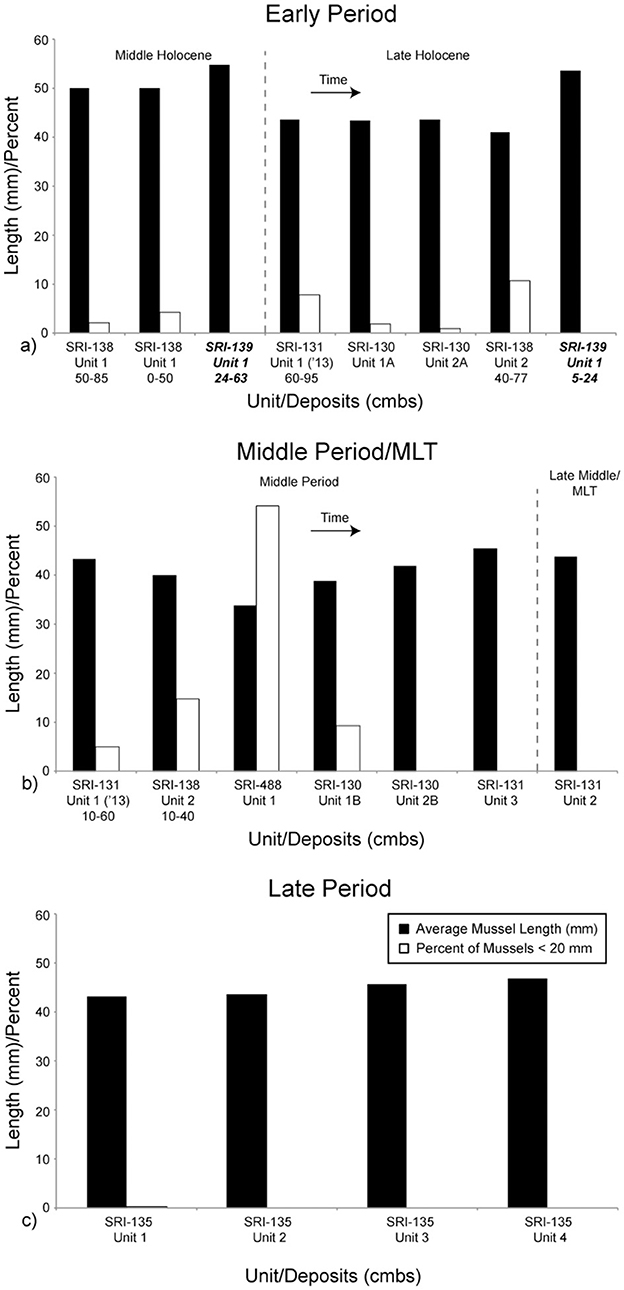
Figure 6. Average mussel size and percent of small (<20 mm) mussels. Note that the y-axis represents both average mussel length (mm) and percent of small mussels (%). Components are separated and the graphs represent time as in Figure 3. Time is generally represented as left to right, and top to bottom from (a) to (c). All components in this figure (a, b) are from La Jolla Vieja Canyon except the two from CA-SRI-139, which are in bold and italicized. They are from Wreck Canyon. All components in this figure (c) are from San Augustin Canyon.
4 Discussion
The chronology of human population distribution on SRI indicates that there is a difference in those factors guiding settlement decisions between the initial settlers through the beginning of the Middle Holocene and those people who occupied the island after the period of population aggregation and growth during relatively dry conditions of the Middle Holocene (Kennett, 2005; Jazwa et al., 2015). During the Middle Holocene, relatively dry conditions prompted islanders to target locations near some of the wettest drainages on SRI that were most resilient to drought (Jazwa et al., 2016a,b). At that time, there was a restructuring of settlement patterns to include more, high population density residential sites, along with seasonal or otherwise short-term special purpose sites (Kennett, 2005; Jazwa et al., 2015). Beginning during this period, settlement patterns closely fit predictions of the IFD until the MCA. Afterward, settlement patterns transition to more closely fit the IDD, a mark of increasing territoriality (Kennett et al., 2009; Jazwa et al., 2019).
Initial settlement, in this case including the Paleocoastal period and beginning of the Middle Holocene, appears to be governed by a different set of decisions. Rather than a focus on just the highest ranked habitats as predicted by the IFD, there is an early phase in which settlement is distributed among both high- and low-ranked habitats. This requires an updated set of target-oriented as opposed to maximizing predictions for the IFD (Table 6). Using SRI as an example, there are five known habitats with settlement sites that predate 7000 years ago. Of these sites, CA-SRI-3 (Tecolote Canyon; Drainage Rank 1), CA-SRI-4 (Arlington Canyon; Drainage Rank 2), CA-SRI-347 (Trancion Canyon; Drainage Rank 3), and CA-SRI-187 (Old Ranch House Canyon; Drainage Rank 7), are in high or medium-high ranked habitats (Kennett, 1998; Winterhalder et al., 2010; Jazwa et al., 2013). However, the final site, CA-SRI-138, is anomalous to this pattern and located at the mouth of La Jolla Vieja Canyon, which is ranked 17th out of 19 drainages. Furthermore, as settlement expands during the Middle Holocene, there is evidence for occupation at CA-SRI-139 and CA-SRI-462, at the mouth of Wreck Canyon (ranked 19), earlier than many other drainages, most notably Cañada Verde, which was ranked fourth in the formulation by Jazwa et al. (2016b) and first by Winterhalder et al. (2010). Additionally, faunal density at these early deposits from both CA-SRI-138 and−139 is high enough to reflect permanent settlement (following Kennett, 2005; Kennett et al., 2009) rather than ephemeral occupation. This is consistent with prediction 1 of our updated IFD model. Additionally, with the exception of CA-SRI-3 and−4, which are located at adjacent drainages, all other early settlement sites we have dated are spaced around the island, at least 7.5 km from each other, suggesting that there is no evidence for concentrated occupation of the island in any one location at this time.
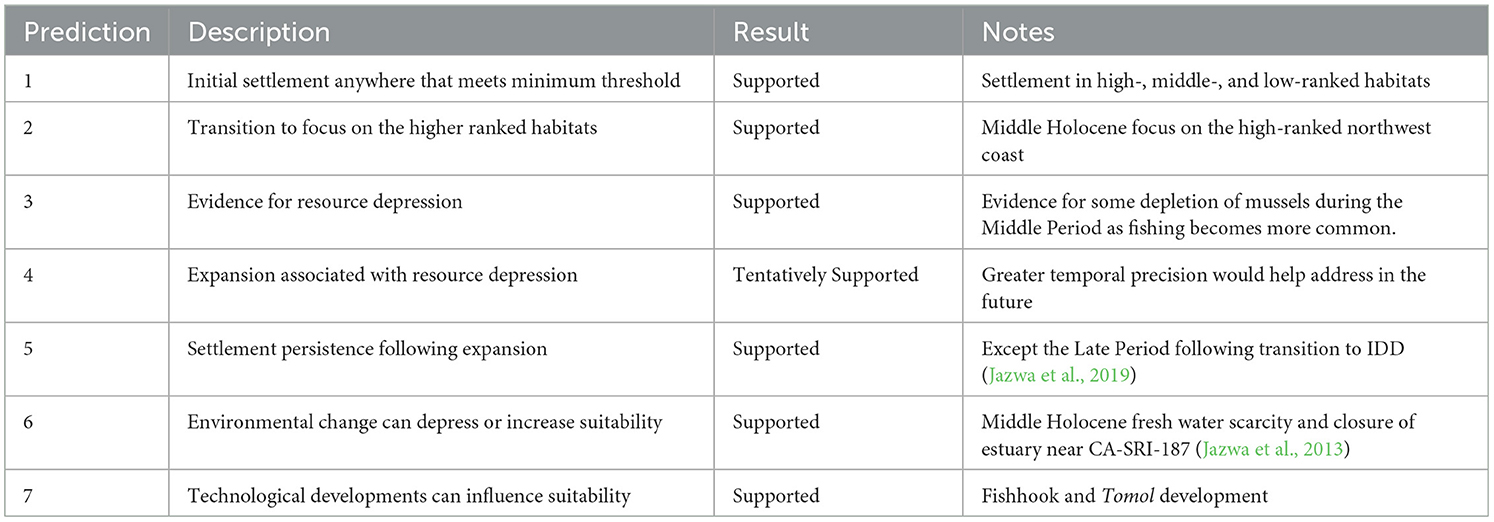
Table 6. Model predictions and assessments for the updated IFD variant considering settlement decisions at low overall population density.
Those early occupation phases at CA-SRI-138 and −139 contain a higher density of shellfish than nearly all the subsequent components along the south coast of Santa Rosa Island for which we have excavation data (Figures 3, 4). While some later phases have higher overall meat weight density when considering all faunal constituents, they are more reliant on fish and sea mammal than during the earlier period. This suggests a restructuring of diet beginning during the Middle Period, consistent with the introduction of the shell fishhook. In the second half of the Middle Holocene, however, settlement expanded greatly along the north coast of SRI, both to new, relatively high-ranked habitats, and within high-ranked habitats. On the south coast, settlement did not expand to the same degree as on the north coast, indicating a transition to focus more on higher ranked habitats that had a higher potential for maximizing resource return (Prediction 2). Furthermore, there is an apparent gap in occupation at the mouth of La Jolla Vieja Canyon after 4985–4540 cal BP until a different area of the site was occupied at the very end of the Early Period. Other than a much lower density component associated with the upper strata at CA-SRI-139, there is no available evidence for occupation of Wreck Canyon after this time.
Island-wide, Jazwa et al. (2016b) argued that there is evidence for resource depression over time between habitats, but evidence within habitats was more equivocal. At Bee Canyon, a relatively low-ranked habitat on the west coast of the island, there is evidence for increase in shellfish diversity and variation in mussel size over time. As important, there is a general increase in fish and sea mammal over time at each of the drainages that were considered. Both of these patterns indicate increasing population density. However, on the south coast of SRI, our current study, which includes data from a greater number of excavated units, sheds additional light on local patterns. California mussels are the most abundant species at most sites on the NCI, and therefore can reflect patterns of overpredation. On the south coast of SRI, there is a decrease in mussel size from the Early Period to the Middle Period, before increasing slightly into the Late Period. However, shellfish diversity also decreases during the Middle Period. Typically, one would expect diversity to increase over time, as there is stress on the highest ranked species, namely mussels (Jazwa et al., 2016b). In the lower-ranked habitats considered here, however, a transitional phase in which resources become stressed before adopting a greater focus on fishing and sea mammal hunting may not have occurred. Put another way, as settlement distribution on SRI adopted a distribution more consistent with a traditional IFD toward the end of the Early Period, population may have diminished in the habitats considered here. Then, as population in-filled at higher ranked habitats, eventually people refocused on the lower-ranked habitats like the ones on the south coast (Prediction 4). Aside from a heavy focus on collecting mussels at CA-SRI-138, the diet of the growing population at that time was more reliant on fishing and sea mammal hunting (Prediction 3). This new array of dietary taxa was supported by the development of the single-piece shell fishhook and later by the tomol plank canoe (Prediction 7).
Jazwa et al. (2016b, 2019) argued that during the Middle Period, settlement eventually expanded to include nearly all of the 19 major drainages on the island. The possible exception to this is Wreck Canyon, which does not have any available radiocarbon dates after 3165–2810 cal BP. This suggests that once settlement included each habitat, settlement persisted there (Prediction 5). There are three exceptions to this. First, occupation of the early-settled habitats in low-ranked La Jolla Vieja and Wreck Canyons decreased or gradually terminated during the period of transition from the low-population conditions considered in this paper to those more consistent with traditional IFD formulations (Prediction 2). Second, at CA-SRI-187 at the mouth of Old Ranch and Old Ranch House Canyons on the east end of SRI, the closure of the productive Abalone Rocks Estuary by 5000 cal BP caused a decrease in suitability of those habitats relative to others on the islands and led to a temporary depopulation of the site (Jazwa et al., 2013; Prediction 6). Finally, beginning during the MCA, there was a major restructuring of settlement distribution on the island consistent with a transition to IDD settlement patterns (Jazwa et al., 2019; Prediction 5). On the south coast, this translated to a focus on settlement at major village sites at the mouth of San Augustin Canyon (CA-SRI-135) and Johnson's Lee (CA-SRI-62). The faunal record from the Late Period village of Helewashkuy at CA-SRI-135 is consistent with other Late Period villages on the island. It is overwhelmingly dominated by fish and shellfish diversity is relatively high, both of which likely indicate high human population density.
4.1 Settlement distribution at low population density
This study is directly applicable to understanding settlement distributions associated with initial movement into a new geographic region. The IFD has been shown to be effective at understanding settlement distribution and expansion over time (Codding and Jones, 2013; Yaworsky and Codding, 2018; Harvey, 2019; Weitzel and Codding, 2022; Jazwa and Collins-Elliott, 2021; Jazwa and Jazwa, 2022; Miller and Carmody, 2022). However, Jazwa et al. (2016b) highlighted limitations to the IFD for low population density. In the example given here, we argue that there is no benefit to small groups of people targeting the best habitats with the greatest potential for resource return. They can still individually act in a way that maximizes their return within their habitat (i.e., by targeting California mussels), but will have a comparable return at all habitats that meet a minimum threshold rank. In doing so, they might choose to distribute themselves on the landscape in such a way as to prioritize other traits typically excluded in ranking habitat suitability per the IFD. On SRI, prevailing winds and storms generally come from the northwest, and the central ridgeline of the relatively rugged island blocks the south side from them. Even during calm weather periods in the summer, the authors have numerous anecdotal observations of days in which the north coast is overcast and cold, but the south side is warm and sunny. Such conditions are less important during periods of resource stress when it is necessary to target habitats with the most abundant resources, but all else being equal, such factors may be considered. Considerations like viewshed (e.g., Lake et al., 1998; Jones, 2006) may also be relevant.
As important, a common assumption of the IFD is one of complete knowledge of the region under consideration, including sufficient information to accurately appraise and compare relative suitabilities for all habitats (Winterhalder et al., 2010: 471; Assumption f). This may not necessarily be the case, especially in a large geographic area, and a settling population may be willing to settle down in one of the first habitats that they encounter that is good enough to provide the subsistence resources they need in quantities that are more than they can consume, even if other, nearby habitats allow for greater resource maximization (Haas and Kuhn, 2019). In such cases, early settlement decisions may have been conditioned by factors such as proximity to previously settled regions. On the NCI, this is less likely to be relevant as the islands are relatively small and it is likely that settlers would be able to quickly assess all potential habitats. However, when considering the initial peopling of the Americas south of the Cordilleran Ice Sheet, for example, people would have first encountered habitats along their route rather than more distant interior habitats that may have had higher resource return. This requires a relaxation of the assumption of complete knowledge in cases in which such information is not available.
While we have chosen to model human settlement patterns for low population densities as a variant of the IFD with modified predictions, the behavior we describe here is consistent with satisficing rather than maximizing models (Bordley and LiCalzi, 2000; Schmidtz, 2004). Satisficing economic models are target- as opposed to maximization-oriented and have been shown to mediate decision making problems relating to risk, uncertainty, and incomplete knowledge (Halstead and O‘Shea, 1989; Low, 1990; Goland, 1991; Winterhalder and Leslie, 2002; Morgan, 2009), as may have been the case for early settlers on the NCI. The use of a formal satisficing model at low population densities that gives way to a standard IFD model once a population threshold has been met could provide insight that our approach might not.
Finally, our study is limited in that much of the land that would have been on the coast of Santarosae during the Paleocoastal period has since been submerged by marine transgression. While an increasing number of Paleocoastal sites have been found across the NCI (Erlandson et al., 2011, 2015, 2016, 2019, 2020; Rick et al., 2013; Jazwa and Rosencrance, 2019; Braje et al., 2020), most are inland. While several coastal sites have been found, CA-SRI-347 at the mouth of Trancion Canyon provides the best evidence for permanent settlement. We cannot say for certain that the patterns we observed in this study are characteristic of the full phase of early occupation, but rather the end of the early, low-density phase of human presence on the NCI. Regardless, we can say that population density was low enough at the very beginning of the Early Period that settlement should be focused on the highest-ranked habitats under the standard formulation of the IFD, which it clearly was not. Our study consequently provides insight into early settlement decisions at low population density when there is little or no resource stress, regardless of whether or not it represents the precise settlement distribution of the very earliest settlers of the NCI.
5 Conclusion
Periods of initial settlement of a geographic region are of particular interest in archaeology. This includes movement into previously uncolonized areas like the initial peopling of the Americas (e.g., Anderson and Gillam, 2000; Braje et al., 2020; Meltzer, 2021). It stands to reason that this initial phase of occupation might be governed by different considerations than later in time when population density is higher and people have a more complete understanding of the landscape, the resources available there, and risks and variation that might affect them on a yearly or longer-term timeline. Ideal distribution models have become prominent in archaeology as tools to understand settlement expansion (e.g., Kennett et al., 2006; Winterhalder et al., 2010; Codding and Jones, 2013; Jazwa et al., 2013, 2016b; Giovas and Fitzpatrick, 2014; Codding and Bird, 2015; Prufer et al., 2017; Yaworsky and Codding, 2018; Harvey, 2019; Jazwa and Collins-Elliott, 2021; Jazwa and Jazwa, 2022; Weitzel and Codding, 2022). One of the greatest benefits of these models is their ability to apply proxies to understand a variety of relevant factors. These include population density and growth, migration, settlement, resource selection and exploitation, environmental change, subsistence efficiency and economic returns, hierarchy, and competition (e.g., Smith and Winterhalder, 1992; Tregenza et al., 1996; Winterhalder, 2002; Kennett, 2005; Bird and O'Connell, 2006; Jazwa et al., 2013, 2016b; Nettle et al., 2013; Codding and Bird, 2015). The IFD and IDD are flexible. For example, they adapt well to different geographic scales and allow for consideration of what might happen when the population has grown such that it benefits some individuals to control the settlement of others (IDD). Here, we consider the opposite, when population is sufficiently small that there is not a risk of others diminishing habitat suitability appreciably, but also that settlers would benefit equally from settling in a variety of habitats.
We use the example of early settlement of SRI because its habitats are all relatively productive compared to other potential locations, even on neighboring Santa Cruz and Santa Rosa Islands (Kennett et al., 2009). Additionally, the island is geographically bounded by the Santa Barbara Channel, resource distribution and use are well understood, and archaeological research over more than the past century has generated a comprehensive dataset of site distribution and chronology. It is for these reason that the NCI have served as a test case for archaeological theory, including in the development of ideal distribution models. We have shown that on SRI, population density during the Paleocoastal period and the beginning of the Early Period was sufficiently small that people settled in both high- and low-ranked habitats, while not living in some other high- and middle-ranked habitats. We focused our analysis on the south coast specifically because the habitats there are notably unremarkable (i.e., low-ranked), but nevertheless contain evidence of early permanent settlement. During the Early Period, prior to the Middle to Late Holocene boundary, there was a transitional phase associated with relatively dry conditions in which population began to focus more on the northwest coast of the island and develop a system of settlement and mobility including permanent settlement sites and special purpose sites that were used seasonally (Kennett, 2005; Jazwa et al., 2015). At this time, settlement at low-ranked habitats continued, but either did not grow or diminished. Later, during the Middle Period, as population density grew across the island, eventually settlement at the low-ranked south coast habitats expanded consistent with IFD predictions, and during the Late Period, it was the location of two of the eight village sites documented in Spanish mission records (Glassow et al., 2010). This provides a framework for understanding the processes behind settlement distribution during the earliest phases of occupation of a new area and could be applied elsewhere to better predict and understand the decisions made by small groups as they expand into new places.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
CJ: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Validation, Writing – original draft, Writing – review & editing. TM: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Writing – original draft. CM: Conceptualization, Formal analysis, Methodology, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. US National Science Foundation provided funding for fieldwork and data collection. This project was supported by CHIS (P11AC30805, P19AC01221, Jazwa; 1443CA8120-96-003, Kennett), the NSF (BCS-1338350, BCS-1623514, BCS-1724639, Jazwa; SBR-9521974, BCS-1460369, Kennett), the Society for California Archaeology Orphaned Collections Grant (Molter), UCSB, CSULB, UO, PSU, and UNR.
Acknowledgments
We would like to thank Channel Islands National Park, including Kristin Hoppa, Laura Kirn, Lulis Cuevas, Drew Adams, Don Morris, and Kelly Minas. Gil Unzueta and Richard-Patrick Cromwell assisted with excavation. Douglas Kennett generously provided collections from his earlier excavations for analysis. We would like to thank Kevin Shoemaker for his comments on Molter's MA thesis, from which this manuscript was created. Thanks also to Mary Oster, Alexandra Zoellner, Ayla Burger, Gabrielle Chaplin, Erika Cory, Katherine Regan, Shelby Burton, and Nathan Seeliger for assistance with laboratory analysis. We also appreciate the invitation to participate in this special edition by Patrick Faulkner, Katherine Woo, and Iain McKechnie, as well as the comments on the manuscript by two reviewers.
Conflict of interest
TM was employed by SWCA Environmental Consultants.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fearc.2025.1535110/full#supplementary-material
Supplementary Table 1 | Compiled faunal data for all the new faunal data generated for this study.
Footnotes
1. ^The large village site to the east of the mouth of San Augustin Canyon has been misassigned in previous publications as CA-SRI-436 (e.g., Kennett, 1998, 2005; Glassow et al., 2010). For consistency with the site database for Channel Islands National Park, we use the originally assigned trinomial CA-SRI-135 in this paper.
2. ^We use the method and equation established by Singh and McKechnie (2015). While a more precise method was recently published by McFarland et al. (2023), the data for our analysis were collected before this method was developed and we no longer have access to the original collections.
References
Ainsworth, B. E., Haskell, W. L., Loen, A. S. Jr., Jacobs, D. R., Montoye, H. J., Sallis, J. F., et al. (1993). Compendium of physical activities: classification of energy costs of human physical activities. Med. Sci. Sports Exerc. 25, 71–80. doi: 10.1249/00005768-199301000-00011
Allen, J., and O'Connell, J. F. (2008). “Getting from Sunda to Sahul,” in Islands of Inquiry: Colonization, Seafaring and the Archaeology of Maritime Landscapes, eds. G. Clarke, F. Leach, and S. O‘Connor (Canberra, Australia: ANU E Press), 31–46. doi: 10.22459/TA29.06.2008.02
Anderson, D. G., and Gillam, J. C. (2000). Paleoindian colonization of the Americas: implications from an examination of physiography, demography, and artifact distribution. Am. Antiq. 65, 43–66. doi: 10.2307/2694807
Arnold, J. E. (1992). Complex hunter-gatherer-fishers of prehistoric California: chiefs, specialists, and maritime adaptations of the Channel Islands. Am. Antiq. 57, 60–84. doi: 10.2307/2694835
Arnold, J. E. (2001). “The Chumash in world and regional perspectives,” in The Origins of a Pacific Coast Chiefdom: The Chumash of the Channel Islands, ed. J.E. Arnold (Salt Lake City, UT: The University of Utah Press), 1–20.
Benson, L., Kashgarian, M., Rye, R., Lund, S., Paillet, F., Smoot, J., et al. (2002). Holocene multidecadal and multicentennial droughts affecting Northern California and Nevada. Quat. Sci. Rev. 21, 659–682. doi: 10.1016/S0277-3791(01)00048-8
Benson, L. V., Hattori, E. M., Southon, J., and Aleck, B. (2013). Dating North America's oldest petroglyphs, Winnemucca Lake Subbasin, Nevada. J. Archaeol. Sci. 40, 4466–4476. doi: 10.1016/j.jas.2013.06.022
Benson, L. V., Lund, S. P., Smoot, J. P., Kashgarian, M., and Burdett, J. W. (2001). “Records of climate change from the Owens Lake Basin, California,” in Smithsonian Contributions to Earth Sciences, eds. R. Hershler, D. Madsen, and D. Currey (Washington, DC: Smithsonian).
Bettinger, R. L. (1980). Explanatory/predictive models of hunter-gatherer adaptation. Adv. Archaeol. Method Theory 3, 189–245. doi: 10.1016/B978-0-12-003103-0.50010-1
Bettinger, R. L. (2009). Hunter-Gatherer Foraging: Five Simple Models. Clinton Corners, New York: Eliot Werner Publications. doi: 10.2307/j.ctvzgb80x
Bettinger, R. L., Malhi, R., and McCarthy, H. (1997). Central place models of acorn and mussel processing. J. Archaeol. Sci. 24, 887–899. doi: 10.1006/jasc.1996.0168
Billman, B. R., and Feinman, G. M. (1999). Settlement Pattern Studies in the Americas: Fifty years Since Viru'. Washington, DC: Smithsonian Institution Press.
Bird, D. W., and O'Connell, J. F. (2006). Behavioral ecology and archaeology. J. Archaeol. Res. 14, 143–188. doi: 10.1007/s10814-006-9003-6
Bordley, R., and LiCalzi, M. (2000). Decision analysis using targets instead of utility functions. Decis. Econ. Finance 23, 53–74. doi: 10.1007/s102030050005
Braje, T. J., Erlandson, J. M., Rick, T. C., Davis, L., Dillehay, T., Fedje, D. W., et al. (2020). Fladmark + 40: what have we learned about a potential Pacific Coast peopling of the Americas? Am. Antiq. 85, 1–21. doi: 10.1017/aaq.2019.80
Braje, T. J., Erlandson, J. M., Rick, T. C., Dayton, P. K., and Hatch, M. B. (2009). Fishing from past to present: continuity and resilience of red abalone fisheries on the Channel Islands, California. Ecol. Appl. 19, 906–919. doi: 10.1890/08-0135.1
Braje, T. J., Kennett, D. J., Erlandson, J. M., and Culleton, B. J. (2007). Human impacts on nearshore shellfish taxa: a 7,000 year record from Santa Rosa Island, California. Am. Antiq. 72, 735–756. doi: 10.2307/25470443
Brantingham, P. J., and Perreault, C. (2010). Detecting the effects of selection and stochastic forces in archaeological assemblages. J. Archaeol. Sci. 37, 3211–3225. doi: 10.1016/j.jas.2010.07.021
Bronk Ramsey, C. (2009). Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360. doi: 10.1017/S0033822200033865
Codding, B. F., and Bird, D. W. (2015). Behavioral ecology and the future of archaeological science. J. Archaeol. Sci. 56, 9–20. doi: 10.1016/j.jas.2015.02.027
Codding, B. F., and Jones, T. L. (2013). Environmental productivity predicts migration, demographic, and linguistic patterns in prehistoric California. Proc. Nat. Acad. Sci. 110, 14569–14573. doi: 10.1073/pnas.1302008110
Collins-Elliott, S. A., and Jazwa, C. S. (2022). Dynamic modeling of the effects of site placement on environmental suitability: a theoretical example from northwest Morocco. Environ. Archaeol. 27, 447–460. doi: 10.1080/14614103.2020.1763020
Colten, R. H. (2001). “Ecological and economic analysis of faunal remains from Santa Cruz Island,” in The Origins of a Pacific Coast Chiefdom: The Chumash of the Channel Islands, ed. J.E. Arnold (Salt Lake City, UT: The University of Utah Press), 199–220.
Erlandson, J. M. (2013). Channel Island amol points: a stemmed paleocoastal type from Santarosae Island, Alta California. Calif. Archaeol. 5, 105–121. doi: 10.1179/1947461X13Z.0000000006
Erlandson, J. M., Braje, T. J., Ainis, A. F., Culleton, B. J., Gill, K. M., Hoffman, C. A., et al. (2020). Maritime paleoindian technology, subsistence, and ecology at an ~11,700 year old paleocoastal site on California's Northern Channel Islands, USA. PLoS ONE. 15:e0238866. doi: 10.1371/journal.pone.0238866
Erlandson, J. M., Braje, T. J., and Gill, K. M. (2019). A paleocoastal site complex from Santarosae Island, California. PaleoAmerica 5, 300–305. doi: 10.1080/20555563.2019.1653157
Erlandson, J. M., Gill, K. M., Glassow, M. A., and Gusick, A. E. (2016). Three paleocoastal lithic sites on Santa Cruz Island, California. PaleoAmerica 2, 52–55. doi: 10.1080/20555563.2016.1141741
Erlandson, J. M., Gill, K. M., Rick, T. C., and Reeder-Myers, L. A. (2015). Three late paleocoastal shell middens on Santa Cruz Island, California. PaleoAmerica 1, 113–115. doi: 10.1179/2055556314Z.00000000011
Erlandson, J. M., Rick, T. C., and Braje, T. J. (2009). Fishing up the food web?: 12,000 years of maritime subsistence and adaptive adjustments on California's Channel Islands. Pac. Sci. 63, 711–724. doi: 10.2984/049.063.0411
Erlandson, J. M., Rick, T. C., Braje, T. J., Casperson, M., Culleton, B., Fulfrost, B., et al. (2011). Paleoindian seafaring, maritime technologies, and coastal foraging on California's Channel Islands Science 331, 1181–1185. doi: 10.1126/science.1201477
Erlandson, J. M., Rick, T. C., Braje, T. J., Steinberg, A., and Vellanoweth, R. L. (2008). Human impacts on ancient shellfish: a 10,000 year record from San Miguel, California. J. Archaeol. Sci. 35, 2144–2152. doi: 10.1016/j.jas.2008.01.014
Erlandson, J. M., Rick, T. C., Estes, J. A., Graham, M. H., Braje, T. J., and Vellanoweth, R. L. (2005). “Sea otters, shellfish, and humans: 10,000 years of ecological interaction on San Miguel Island, California,” in Proceedings of the Sixth California Islands Symposium, ed. D. K. Garcelon and C. A. Schwemm (Arcata, CA: Institute for Wildlife Studies and National Park Service), 58–69.
Erlandson, J. M., Rick, T. C., and Vellanoweth, R. L. (2004). “Human impacts on ancient environments: a case study from California's Northern Channel Islands,” in Voyages of Discovery: The Archaeology of Islands, ed. S. Fitzpatrick (Westport, CT: Praeger), 51–83.
Fretwell, S. D., and Lucas, H. L. Jr. (1969). On territorial behavior and other factors influencing habitat distribution in birds. I. Theoretical development. Acta Biotheor. 19, 16–36. doi: 10.1007/BF01601953
Gamble, L. H. (2017). Feasting, ritual practices, social memory, and persistent places: new interpretations of shell mounds in southern California. Am. Antiq. 82, 427–451. doi: 10.1017/aaq.2017.5
Gifford-Gonzalez, D., and Hildebrandt, W. (2012). “If mussels weighed a ton: problems with quantifying Santa Barbara Channel archaeofaunas,” in Exploring Methods of Faunal Analysis: Insights from California Archaeology, ed. M. Glassow, and T. Joslin (Los Angeles, CA: Cotsen Institute of Archaeology Press at UCLA), 97–108. doi: 10.2307/j.ctvdjrqzg.10
Giovas, C. M., and Fitzpatrick, S. M. (2014). Prehistoric migration in the Caribbean: past perspectives, new models and the ideal free distribution in West Indian colonization. World Archaeol. 46, 569–589. doi: 10.1080/00438243.2014.933123
Glassow, M. A. (1993). “Changes in subsistence on marine resources through 7,000 years of prehistory on Santa Cruz Island,” in Archaeology on the Northern Channel Islands of California, ed. M. A. Glassow (Salinas, CA: Coyote Press), 75–94.
Glassow, M. A. (2013). “Settlement systems on Santa Cruz Island between 6300 and 5300 BP,” in California's Channel Islands: The Archaeology of Human-Environment Archaeology, ed. C.S. Jazwa, and J.E. Perry (Salt Lake City, UT: University of Utah Press), 60–74.
Glassow, M. A., Braje, T. J., Costello, J. G., Erlandson, J. M., Johnson, J. R., Morris, D. P., et al. (2010). Channel Islands National Park Archaeological Overview and Assessment. Ventura, CA: Cultural Resources Division, Channel Islands National Park.
Glassow, M. A., Thakar, H. B., and Kennett, D. J. (2012). Red abalone collecting and marine water temperature during the Middle Holocene occupation of Santa Cruz Island, California. J. Archaeol. Sci. 39, 2574–2582. doi: 10.1016/j.jas.2012.03.017
Goland, C. (1991). “The ecological context of hunter-gatherer storage: environmental predictability and environmental risk.” in Foragers in Context: Long Term, Regional, and Historical Perspectives in Hunter-Gatherer Studies, Vol 10, ed. P. T. Miracle, L. E. Fisher, and J. Brown (Ann Arbor, MI: Michigan Discussions in Anthropology, Hunter-Gatherer Studies), 107–125.
Haas, R., and Kuhn, S. L. (2019). Forager mobility in constructed environments. Curr. Anthropol. 60, 499–535. doi: 10.1086/704710
Hagerty, S. A. (2024). Dietary shifts and cultural complexity: exploring Late Holocene faunal remains and marine mammal hunting on the Northern Channel Islands. Master's thesis. Reno (NV): University of Nevada, Reno.
Halstead, P., and O‘Shea, J. (1989). “Introduction: cultural responses to risk and uncertainty,” in Bad Year Economics, Cultural Responses to Risk and Uncertainty, eds. P. Halstead, and J. O'Shea (Cambridge, UK: Cambridge University Press), 1–7 doi: 10.1017/CBO9780511521218.002
Hanna, J. A., and Giovas, C. M. (2022). An islandscape IFD: using the ideal free distribution to predict pre-Columbian settlements from Grenada to St. Vincent, Eastern Caribbean. Environ. Archaeol. 27, 402–419. doi: 10.1080/14614103.2019.1689895
Harvey, D.C. (2018). Core-periphery dynamics in the Kern River watershed: Implications for Tubatulabal territorial maintenance during the Late Holocene. Hunter Gatherer Res. 4, 557–587. doi: 10.3828/hgr.2018.33
Harvey, D. C. (2019). Habitat distribution, settlement systems, and territorial maintenance in the southern Sierra Nevada, California. PhD dissertation. Reno (NV): University of Nevada, Reno.
Heaton, T. J., Köhler, P., Butzin, M., Bard, E., Reimer, R. W., Austin, W. E., et al. (2020). Marine20—the marine radiocarbon age calibration curve (0–55,000 cal BP). Radiocarbon 62, 779–820. doi: 10.1017/RDC.2020.68
Hendy, I. L., Dunn, L., Schimmelmann, A., and Pak, D. K. (2013). Resolving varve and radiocarbon chronology differences during the last 2000 years in the Santa Barbara sedimentary record, California. Quat. Int. 310, 155–168. doi: 10.1016/j.quaint.2012.09.006
Hughes, M. K., and Graumlich, L. J. (2000). Multi-Millennial Nevada Precipitation Reconstruction. International Tree-Ring Data Bank. IGBP PAGES/World Data Center-A for Paleoclimatology Data Contribution Series #2000-049. Boulder, CO: NOAA/NGDC Paleoclimatology Program.
Jazwa, C. S. (2015). A dynamic ecological model for human settlement on California's northern Channel Islands. PhD. dissertation. University Park (PA): Pennsylvania State.
Jazwa, C. S., Braje, T. J., Erlandson, J. E., and Kennett, D. J. (2015). Central place foraging and shellfish processing on California's Northern Channel Islands. J. Anthropol. Archaeol. 40, 33–47. doi: 10.1016/j.jaa.2015.05.005
Jazwa, C. S., and Collins-Elliott, S. A. (2021). An ecological model of settlement expansion in northwestern Morocco. Quatern. Int. 597, 103–117. doi: 10.1016/j.quaint.2020.12.037
Jazwa, C. S., Duffy, C. J., Leonard, L., and Kennett, D. J. (2016a). Hydrological modeling and prehistoric settlement on Santa Rosa Island, California, USA. Geoarchaeology 31, 101–120. doi: 10.1002/gea.21532
Jazwa, C. S., and Jazwa, K. A. (2017). Settlement ecology in Bronze Age Messenia. J. Anthropol. Archaeol. 45, 157–169. doi: 10.1016/j.jaa.2016.12.003
Jazwa, C. S., and Jazwa, K. A. (2022). Resource competition and settlement distribution in Bronze Age Greece. Hum. Ecol. 50, 399–418. doi: 10.1007/s10745-022-00324-6
Jazwa, C. S., Joslin, T. L., and Kennett, D. J. (2020). Fishing, subsistence change, and foraging strategies on western Santa Rosa Island, California. Am. Antiq. 85, 591–608. doi: 10.1017/aaq.2020.18
Jazwa, C. S., Kennett, D., Winterhalder, B., and Joslin, T. L. (2019). Territoriality and the rise of despotic social organization on western Santa Rosa Island, California. Quat. Int. 518, 41–56. doi: 10.1016/j.quaint.2017.11.009
Jazwa, C. S., Kennett, D. J., and Winterhalder, B. (2013). “The ideal free distribution and settlement history at Old Ranch Canyon, Santa Rosa Island,” in California's Channel Islands: The Archaeology of Human-Environment Interactions, eds. C. S. Jazwa and J. E. Perry (Salt Lake City, UT: University of Utah Press), 75–96.
Jazwa, C. S., Kennett, D. J., and Winterhalder, B. (2016b). A Test of ideal free distribution predictions using targeted survey and excavation on California's Northern Channel Islands. J. Archaeol. Method Theory 23, 1242–1284. doi: 10.1007/s10816-015-9267-6
Jazwa, C. S., and Rosencrance, R. L. (2019). Technological change and interior settlement on western Santa Rosa Island, California. J. Anthropol. Archaeol. 54, 235–253. doi: 10.1016/j.jaa.2019.04.007
Jazwa, C. S., and Zoellner, A. (2024). Middle Holocene residential mobility in western Santa Rosa Island, California. Quat. Int. 689–690, 16–29. doi: 10.1016/j.quaint.2023.10.011
Jochim, M. (1976). Hunter-Gatherer Subsistence and Settlement, A Predictive Model. New York: Academic Press.
Johnson, J. R. (1993). “A geographic analysis of island Chumash marriage networks,” in Archaeology on the Northern Channel Islands of California: Studies of Subsistence, Economics, and Social Organization, eds. M.A. Glassow (Salinas, CA: Coyote Press), 19–46.
Johnson, J. R., Stafford, T. W. Jr., Ajie, H. O., and Morris, D. P. (2002). Arlington Springs revisited,” in The Fifth California Islands Symposium, ed. D. R. Brown, K. C. Mitchell, and H. W. Chaney (Santa Barbara, CA: Santa Barbara Museum of Natural History), 541–545.
Jones, E. (2006). Using viewshed analysis to explore settlement choice: a case study of the Onondoga Iroquois. Am. Antiq. 71, 523–538. doi: 10.2307/40035363
Jones, T. L., and Richman, J. R. (1995). On mussels: Mytilus californianus as a prehistoric resource. North Am. Archaeol. 16, 33–58. doi: 10.2190/G5TT-YFHP-JE6A-P2TX
Kennett, D., Anderson, A., and Winterhalder, B. (2006). “The ideal free distribution, food production, and the colonization of Oceania,” in Behavioral Ecology and the Transition to Agriculture, ed. D. J. Kennett and B. Winterhalder (Berkeley, CA: University of California Press), 265–288. doi: 10.1525/9780520932456-015
Kennett, D. J. (1998). Behavioral ecology and the evolution of hunter-gatherer societies on the northern Channel Islands, California. PhD dissertation. Santa Barbara (CA): University of California, Santa Barbara.
Kennett, D. J. (2005). The Island Chumash: Behavioral Ecology of a Maritime Society. Berkeley, CA: University of California Press. doi: 10.1525/9780520931435
Kennett, D. J., and Clifford, R. A. (2004). “Flexible strategies for resource defense on the northern Channel Islands of California: an agent-based model,” in Voyages of Discovery: the Archaeology of Islands, ed. S. M. Fitzpatrick (Westport, CA: Praeger), 21–50.
Kennett, D. J., and Kennett, J. P. (2000). Competitive and cooperative responses to climatic instability in coastal southern California. Am. Antiq. 65, 379–395. doi: 10.2307/2694065
Kennett, D. J., Kennett, J. P., Erlandson, J. M., and Cannariato, K. G. (2007). Human responses to Middle Holocene climate change on California's Channel Islands. Quat. Sci. Rev. 26, 351–367. doi: 10.1016/j.quascirev.2006.07.019
Kennett, D. J., Kennett, J. P., West, G. J., Erlandson, J. M., Johnson, J. R., Hendy, I. L., et al. (2008). Wildfire and abrupt ecosystem disruption on California's Northern Channel Islands at the Ållerød–Younger Dryas boundary (13.0–12.9 ka). Quat. Sci. Rev. 27, 2530–2545. doi: 10.1016/j.quascirev.2008.09.006
Kennett, D. J., Winterhalder, B., Bartruff, J., and Erlandson, J. M. (2009). “An ecological model for the emergence of institutionalized social hierarchies on California's northern Channel Islands,” in Pattern and Process in Cultural Evolution, ed. S. Shennan (Berkeley: University of California Press), 297–314. doi: 10.1525/9780520943360-021
Kowalewski, S. A. (2008). Regional settlement pattern studies. J. Archaeol. Res. 16, 225–285. doi: 10.1007/s10814-008-9020-8
Krist, F. J. (2001). A predictive model of paleo-Indian subsistence and settlement. PhD dissertation. East Lansing (MI): Michigan State University.
Lake, M. W., Woodman, P. E., and Mithen, S. J. (1998). Tailoring GIS software for archaeological applications: an example concerning viewshed analysis. J. Archaeol. Sci. 25, 27–38. doi: 10.1006/jasc.1997.0197
LaMarche, V. C. (1973). Holocene climatic variations inferred from treeline fluctuations in the White Mountains, California. Quat. Res. 3, 632–660. doi: 10.1016/0033-5894(73)90035-5
LaMarche, V. C. (1974). Paleoclimatic inferences from long tree-ring records. Science 183, 1043–1048. doi: 10.1126/science.183.4129.1043
Lindström, S. (1990). Submerged tree stumps as indicators of Mid-Holocene aridity in the Lake Tahoe Basin. J. Calif. Gt. Basin Anthropol. 12, 146–157.
Low, B. S. (1990). “Human responses to environmental extremeness and uncertainty: a cross-cultural perspective,” in Risk and Uncertainty in Tribal and Peasant Economies, ed. E. Cashdan (Boulder, CO: Westview Press), 229–256.
Mason, R. D., Peterson, M. L., and Tiffany, J. A. (1998). Weighing vs. counting: measurement reliability and the California school of midden analysis. Am. Antiq. 63, 303–324. doi: 10.2307/2694700
McFarland, J. D., Ainis, A. F., and Jazwa, C. S. (2025). Multiscale analysis of zooarchaeological data to reconstruct past kelp forest productivity for the Northern Channel Islands, California U.S.A. Estuar. Coast. Shelf Sci. 316:109178. doi: 10.1016/j.ecss.2025.109178
McFarland, J. D., Sbei, A. N., Jazwa, C. S., and Stull, K. E. (2023). Using multiple linear regression for estimating total mussel (Mytilus californianus) shell size to accommodate morphological variability related to diverse environments. J. Archaeol. Sci. Rep. 49:104004. doi: 10.1016/j.jasrep.2023.104004
Meltzer, D. J. (2021). First Peoples in a New World: Populating Ice Age America (2nd Edition). Cambridge, UK: Cambridge University Press. doi: 10.1017/9781108632867
Miller, S. D., and Carmody, S. B. (2022). Fire on the mountain: the ideal free distribution and early hunter-gatherer demography in the Tennessee River Drainage, USA. Environ. Archaeol. 27, 357–371. doi: 10.1080/14614103.2020.1777054
Molter, T. (2021). Early settlement in marginal habitats on Santa Rosa Island, California. Master's thesis. Reno (NV): University of Nevada, Reno.
Morgan, C. (2009). Climate change, uncertainty and prehistoric hunter-gatherer mobility. J. Anthropol. Archaeol. 28, 382–396. doi: 10.1016/j.jaa.2009.07.004
Moritz, M., Hamilton, I. M., Chen, Y., and Scholte, P. (2014). Mobile pastoralists in the Logone floodplain distribute themselves in an ideal free distribution. Curr. Anthropol. 55, 115–122. doi: 10.1086/674717
Nettle, D., Gibson, M. A., Lawson, D. W., and Sear, R. (2013). Human behavioral ecology: current research and future prospects. Behav. Ecol. 24, 1031–1040. doi: 10.1093/beheco/ars222
O'Connell, J. F. (2020). Aboriginal fires modify an ideal free distribution. Proc. Nat. Acad. Sci. 117, 13873–13875. doi: 10.1073/pnas.2008944117
O'Connell, J. F., and Allen, F. (2012). The restaurant at the end of the universe: modelling the colonization of Sahul. Aust. Archaeol. 74, 5–17. doi: 10.1080/03122417.2012.11681932
Pilgram, T. K. (1987). Predicting archaeological sites from environmental variables: a mathematical model for the Sierra Nevada foothills, California. Oxford, UK: BAR International. doi: 10.30861/9780860544098
Plekhov, D., and Levine, E. I. (2022). Defining suitability in mixed agropastoral societies: a case study from Bactria in northern Afghanistan. Environ. Archaeol. 27, 372–387. doi: 10.1080/14614103.2020.1777055
Prufer, K. M., Thompson, A. E., Meredith, C. R., Culleton, B. J., Jordan, J. M., Ebert, C. E., et al. (2017). The Classic Period Maya transition from and ideal free to an ideal despotic settlement system at the polity of Uxbenká. J. Anthropol. Archaeol. 45, 53–68. doi: 10.1016/j.jaa.2016.11.003
Quade, J., Forester, R. M., Pratt, W. L., and Carter, C. (1998). Black mats, spring-fed streams, and late-glacial-age recharge in the southern Great Basin. Quat. Res. 49, 129–148. doi: 10.1006/qres.1997.1959
Raab, L. M. (1992). An optimal foraging analysis of prehistoric shellfish collecting on San Clemente Island, California. J. Ethnobiol. 12, 63–80.
Reeder-Myers, L., Erlandson, J. M., Muhs, D. R., and Rick, T. C. (2015). Sea level, paleogeography, and archaeology on California's Northern Channel Islands. Quat. Res. 83, 263–272. doi: 10.1016/j.yqres.2015.01.002
Rick, T. C. (2004). Daily activities, community dynamics, and historical ecology on California's northern Channel Islands. PhD dissertation. Eugene (OR): University of Oregon.
Rick, T. C. (2007). The Archaeology and Historical Ecology of Late Holocene San Miguel Island, California. Los Angeles, CA: Cotsen Institute of Archaeology. doi: 10.2307/j.ctvdjrrhn
Rick, T. C. (2011). Weathering the storm: coastal subsistence and ecological resilience on Late Holocene Santa Rosa Island, California. Quat. Res. 239, 135–146. doi: 10.1016/j.quaint.2010.06.008
Rick, T. C., Duguid, D., Sholts, S. B., Spitzer, M. D., Wake, T. A., and Erlandson, J. M. (2023). Investigations at an eroded shell midden reveal direct evidence for fur seal hunting and diverse subsistence strategies on Late Holocene Santa Rosa (Wima) Island, California. J. Island Coast. Archaeol. 20, 1–15. doi: 10.1080/15564894.2023.2183439
Rick, T. C., Erlandson, J. M., Jew, N. P., and Reeder-Myers, L. A. (2013). Archaeological survey, paleogeography, and the search for late Pleistocene paleocoastal peoples of Santa Rosa Island, California. J. Field Archaeol. 38, 324–331. doi: 10.1179/0093469013Z.00000000065
Rick, T. C., Vellanoweth, R. L., Erlandson, J. M., and Kennett, D. J. (2002). On the antiquity of a single-piece shell fishhook: AMS radiocarbon evidence from the southern California coast. J. Archaeol. Sci. 29, 933–942. doi: 10.1006/jasc.2001.0788
Schmidtz, D. (2004). “Satisficing as a humanly rational strategy,” in Satisficing and Maximizing: Moral Theorists on Practical Reason, ed. M. Byron (Cambridge, UK: Cambridge University Press), 30–58. doi: 10.1017/CBO9780511617058.003
Shannon, C. E., and Weaver, W. (1949). The Mathematical Theory of Communication. Urbana, IL: University of Illinois Press.
Singh, G. G., and McKechnie, I. (2015). Making the most of fragments: a method for estimating shell length from fragmentary mussels (Mytilus californianus and Mytilus trossulus) on the Pacific coast of North America. J. Archaeol. Sci. 58, 175–183. doi: 10.1016/j.jas.2015.02.029
Smith, E. A., and Winterhalder, B. (1992). Evolutionary Ecology and Human Behavior. New York: Aldine de Gruyter.
Thompson, R. S. (1992). Late Quaternary environments in Ruby Valley, Nevada. Quat. Res. 37, 1–15. doi: 10.1016/0033-5894(92)90002-Z
Tregenza, T., Parker, G. A., and Thompson, D. J. (1996). Interference and the ideal free distribution: models and tests. Behav. Ecol. 7, 379–386. doi: 10.1093/beheco/7.4.379
Vernon, K. B., Yaworsky, P. M., Spangler, J., Brewer, S., and Codding, B. F. (2022). Decomposing habitat suitability across the forager to farmer transition. Environ. Archaeol. 27, 420–433. doi: 10.1080/14614103.2020.1746880
Weitzel, E. M., and Codding, B. F. (2022). The ideal distribution model and archaeological settlement patterning. Environ. Archaeol. 27, 349–356. doi: 10.1080/14614103.2020.1803015
Winterhalder, B. (2002). “Models,” in Darwin and Archaeology: A Handbook of Key Concepts, ed. J. P. Hart and J. E. Terrell (Westport, CT: Bergin and Garvey), 201–223 doi: 10.5040/9798400637810.0015
Winterhalder, B., Kennett, D. J., Grote, M. N., and Bartruff, J. (2010). Ideal free settlement of California's Northern Channel Islands. J. Anthropol. Archaeol. 29, 469–490. doi: 10.1016/j.jaa.2010.07.001
Winterhalder, B., and Leslie, P. (2002). Risk-sensitive fertility, the variance compensation hypothesis. Evol. Hum. Behav. 23, 59–82. doi: 10.1016/S1090-5138(01)00089-7
Keywords: ideal free distribution, Chumash, California Channel Islands, settlement patterns, archaeology
Citation: Jazwa CS, Molter TR and Morgan CT (2025) Low population density settlement patterns on California's Northern Channel Islands. Front. Environ. Archaeol. 4:1535110. doi: 10.3389/fearc.2025.1535110
Received: 26 November 2024; Accepted: 30 June 2025;
Published: 24 July 2025.
Edited by:
Patrick Faulkner, The University of Sydney, AustraliaReviewed by:
Mirani Litster, James Cook University, AustraliaHeather Thakar, Texas A&M University, United States
Copyright © 2025 Jazwa, Molter and Morgan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christopher S. Jazwa, Y2phendhQHVuci5lZHU=