 Matthew Campbell1,2*
Matthew Campbell1,2* Richard Walter3,4
Richard Walter3,4 Armagan Sabetian5
Armagan Sabetian5 Louise Furey6
Louise Furey6 Emma Ash7
Emma Ash7 Andrew McAlister1
Andrew McAlister1 Jingjing Zhang5,8
Jingjing Zhang5,8 Caitlin Haylock1,2
Caitlin Haylock1,2- 1Anthropology Department, School of Social Sciences, University of Auckland Waipapa Taumata Rau, Auckland, New Zealand
- 2CFG Heritage Ltd., Auckland, New Zealand
- 3Southern Pacific Archaeological Research, School of Social Sciences, University of Otago Otākou Whakaihu Waka, Dunedin, New Zealand
- 4School of Social Sciences, University of Queensland, Brisbane, QLD, Australia
- 5School of Science, Auckland University of Technology Te Wānaga Aronui o Tāmaki Makau Rau, Auckland, New Zealand
- 6Independent Researcher, Auckland, New Zealand
- 7Auckland War Memorial Museum Tāmaki Paenga Hira, Auckland, New Zealand
- 8Plant and Food Research Rangahau Ahumāra Kai, Auckland, New Zealand
The stratified midden on Ōtata Island, in New Zealand's Hauraki Gulf, dates from the fourteenth century CE, soon after the East Polynesian settlement of New Zealand, to the eighteenth century, just prior to the arrival of Europeans. Analysis of the fishbone assemblages shows that the main targeted species throughout the sequence was snapper (Chrysophrys auratus), accounting for between 50 and 88% of each assemblage. Cranial bones of snapper significantly outnumbered vertebrae, demonstrating that snapper bodies were being preserved for off-site consumption. This pattern is not apparent for other fish species. An early focus on individual fish capture, with benthic or reef species more common than in later assemblages, was superseded by an emphasis on pelagic schooling species, indicating increased use of netting both in response to demographic pressures and as netting technology developed. Snapper size reconstructions for Occupation 3 were significantly skewed toward smaller fish as the environment recovered from the eruption of the nearby Rangitoto volcano and deposition of tephra on the seabed. Otolith trace element analysis showed that juvenile snapper behaviors remained consistent throughout the sequence, but that there were subtle environmental changes. These are attributed to anthropogenic terrestrial changes, but otherwise there were no measurable anthropogenic changes to marine ecosystems.
Introduction
New Zealand was the last major landmass to be settled by humans, with the arrival of East Polynesian voyagers, the ancestors of the Māori, around 1,300 CE (Anderson, 1991; Anderson et al., 2014; Walter et al., 2017). Pre-European Māori were aceramic horticulturalists but a limited suite of cultigens meant they were reliant on wild foods, and marine resources were a major contributor to diet. The giant flightless moa (Dinornithiformes) were hunted to extinction by 1,450 CE, within 150 years of first settlement (Holdaway and Jacomb, 2000; Anderson, 2000), and seals were extirpated throughout much of their range at the same time (Smith, 2002; Collins et al., 2014). Moa and seal were less common in the warmer North Island, where kūmara (sweet potato, Ipomoea batatas) cultivation was central to the economy. While kūri (Māori dog, Canis familiaris) and kiore (Polynesian rat, Rattus exulans), along with a range of birds, were available, fish and shellfish were always of greater importance, contributing vital proteins and fats that were not available from plant foods.
The impact of settlement on terrestrial environments has been a major focus of scholarship in New Zealand since the nineteenth century (e.g., von Haast, 1871; Anderson, 2013). For instance, as moa, a keystone species, were hunted to extinction, other bird species followed, such as their main predator, the Haast's eagle (Hieraaetus moorei) (Worthy and Holdaway, 2002; Tennyson and Martinson, 2006). Expanding human populations were accompanied by extensive forest clearance for kūmara cultivation and by incidental burning (McWethy et al., 2014). While this is reasonably well understood, Māori impacts on marine environments have received much less attention. Several researchers have concluded that cultural and natural effects are difficult to disentangle and that human impacts, if any, were minor (e.g., Shawcross, 1967; Anderson, 1997; Leach and Davidson, 2000, 2001; Leach, 2006). Māori populations were never large and it is uncertain whether Māori fishing practices would have had measurable effects on fish stocks. Leach (2006) and Davidson (2011) suggest that indirect pressures may have been more important. Land clearance for horticulture may have resulted in increased sediment loads in harbors and estuaries, leading to ecosystem changes of nearshore habitats. Wells et al. (2019) show that growth rates of the bivalve tuangi (Austrovenus stutchburyi), an intertidal soft shore species, declined at several archaeological sites following increased sediment influx into estuaries due to land clearance, in some cases by as much as 50% between 1,400 and 1,800 CE.
Terrestrial ecosystems are relatively static and humans interact with them directly. Conversely, marine ecosystems are dynamic and experience human interactions more indirectly. It is challenging to unravel the complexity of human impacts on marine ecosystems. In this paper we examine these questions by analyzing a midden from Ōtata Island, near Auckland. This represents a rare opportunity to study the fishbone assemblage from a stratified midden spanning the full range of pre-European Māori occupation, particularly as Ōtata is a small island with a restricted environment unsuitable for horticulture or permanent habitation. People visited the island to fish and little else, so any changes in the fishery should be evident.
The Ōtata midden
Ōtata is a small island (~600 × 500 m with a maximum elevation of ~65 m) in the Noises Group at the margin of the inner Hauraki Gulf, where the bathymetry drops away to the cooler waters of the outer gulf (Figure 1). Archaeological site R10/139 was originally recorded as two layers of midden, but in 2018 a storm and accompanying swells eroded ~5 m of the coastal scarp and scoured out the pebble beach, exposing further stratigraphic layers for ~50 m along the scarp. Seven excavation squares (Squares A–G) were excavated in the face of the midden and behind it in 2020 and 2021. Five occupation layers and three natural layers were recorded, although not all layers were present in all squares (Figure 2). The general stratigraphy is as follows:
A storm deposit from the 2018 storm consisting of water rolled beach cobbles and shell.
Occupation 5, which probably consists of several similar occupations or events superimposed and mixed. The deposit consists of mixed fragmented and burnt shell and fire-cracked rock in a dark charcoal stained matrix. In Square A an intact hāngi (deep earth oven) containing large intact and fragmented beach boulders was dug into and covered by this matrix. The upper 100 mm of the layer was a topsoil containing modern plastic and glass as well as faunal material.
Occupation 4 was present in Squares D, E and F and could be distinguished from Occupation 5 immediately above it. This deposit had more loam, less charcoal, shell and fire-cracked rock but lenses of oyster shell and fish at the base. It seems probable that Occupation 4 is the first of several late events that are distinguished as Occupation 5. The two occupations have indistinguishable dates and for the sake of analysis and discussion Occupations 4 and 5 are treated as one (Occupation 4/5).
Below Occupation 4 was a storm deposit of fine beach gravel and shell fragments 130–200 mm thick, with some obsidian and basalt flakes mixed into it.
Occupation 3 was a relatively thin deposit containing shell, fish bone and fire-cracked rock but with notably less charcoal staining than Occupations 4 and 5.
A thick deposit, 200–300 mm deep of redeposited tephra was present below Occupation 3. This appears to be a slopewash deposit and contains lenses of soil and clay but no cultural material below the upper few mm. This tephra relates to the Rangitoto A and B eruptions dating to 1,400–1,450 CE (Needham et al., 2011).
Occupation 2 was only present in Square E and consisted of an ash-filled earth oven with rakeout cut into Occupation 1, and occasional obsidian and basalt mixed into a surrounding wave deposited pebble layer. This pebble layer was also visible in Squares D, F and G.
Occupation 1 was visible in all squares and was deepest, ~100 mm, in Squares E and F. It contained fish, bird and sea mammal bone, obsidian and basalt flakes, and a few bone and shell fishhooks in a fine, clean pebbly beach matrix, but significantly less shell or charcoal. It appears to have been disturbed by waves or storms with small beach pebbles mixed in to the upper parts of the layer by wave action.
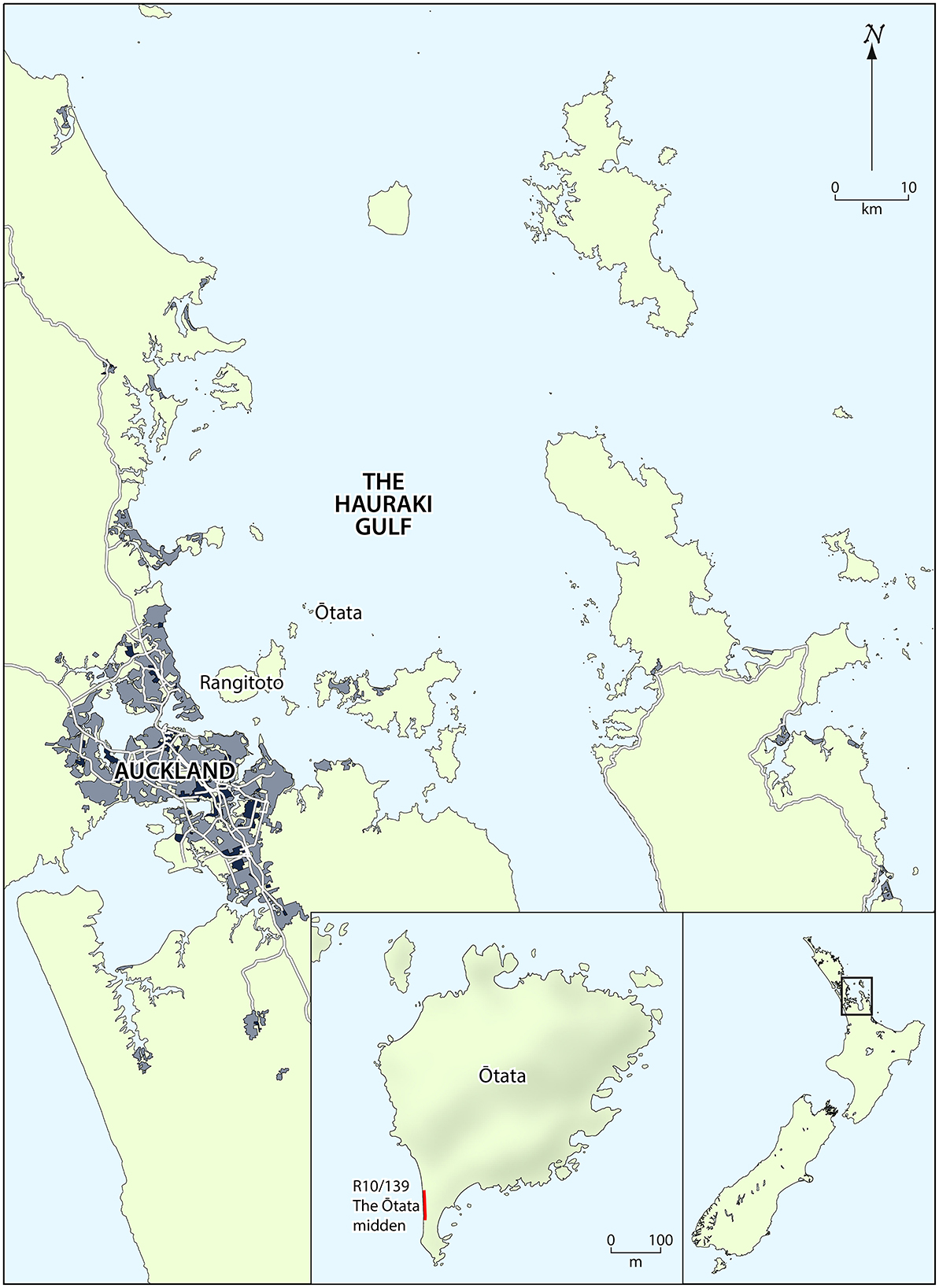
Figure 1. The Hauraki Gulf, showing the location of Ōtata.

Figure 2. Square E (1,000 mm wide) and Square F (500 mm wide) were excavated 700–800 mm into the face of the exposed midden. They demonstrate the typical stratigraphy for the site. Photo and drawing are not to the same scale.
Only a preliminary dating program has been carried out so far. Four terrestrial and three marine samples were submitted to the Radiocarbon Dating Laboratory at the University of Waikato for AMS dating: two each from Occupations 1, 3, and 5, and one from Occupation 4. The results (Table 1) show that Occupation 1 dates to the fourteenth century CE, not as early as the earliest dates for East Polynesian settlement for New Zealand, but soon after; Occupation 3 dates of the mid-fifteenth to early seventeenth centuries, after the Rangitoto eruption; and Layers 4 and 5 date to the early eighteenth century up to modern times, and that these two layers are indistinguishable. A preliminary Bayesian modeling of the dates, using 1,400–1,450 CE for the Rangitoto tephra (Needham et al., 2011) as an upper boundary for Occupation 1 and lower boundary for Occupation 3, and 1,800 CE as a reasonable date for the probable incursion of European material into the site and an upper boundary for Occupations 4 and 5, does not alter the overall periodization of the occupations, but considerably reduces the uncertainty associated with the marine dates (Figure 3).
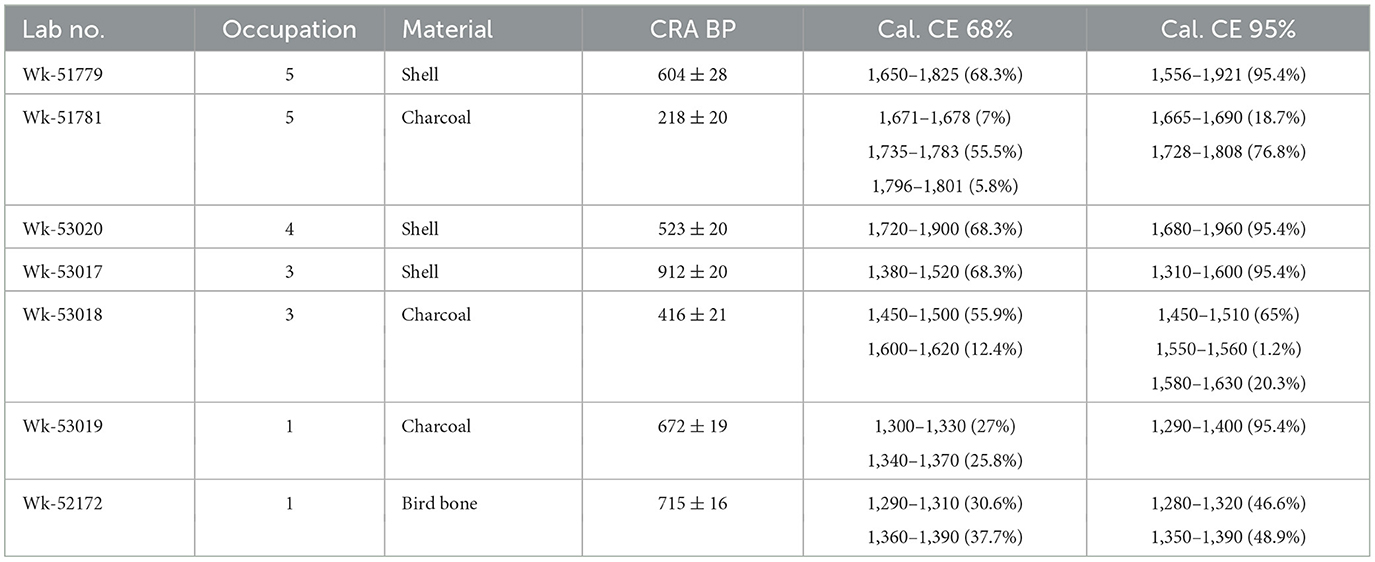
Table 1. Unmodelled radiocarbon results.
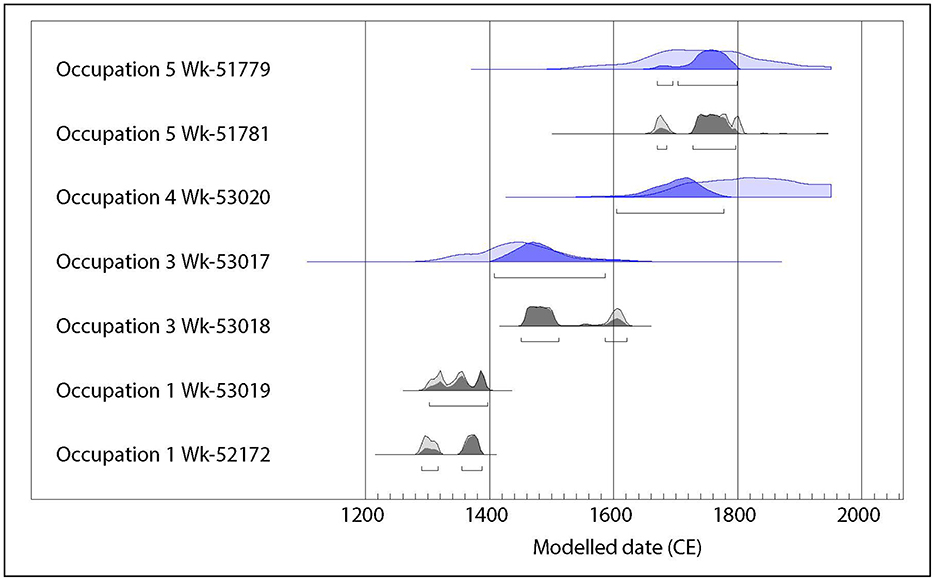
Figure 3. Bayesian age model for the occupations. The lighter shade distributions are the unmodelled calibrated dates, darker shade distributions represent the results after Bayesian modeling. Terrestrial dates are in gray, marine dates in blue. Distribution bars represent the 95% confidence interval.
The Ōtata fish
Fishbone was analyzed by Caitlin Haylock using comparative collections at CFG Heritage and the Anthropology Department, University of Auckland Waipapa Taumata Rau, following the method developed by Campbell (2016). The results of the analysis are presented in Table 2 (Linnean taxonomy and Māori names are given in Table 3). This includes fish from the highly disturbed, redeposited tephra and sand layers, which are not discussed further. Fishbone in these layers may have originated in the occupation layers above and below, although there is no way of assigning them; or they may have originated in occupations now badly disturbed and no longer in situ. Fish are only present in Occupation 2 in small numbers, and this also is not discussed further in any detail.
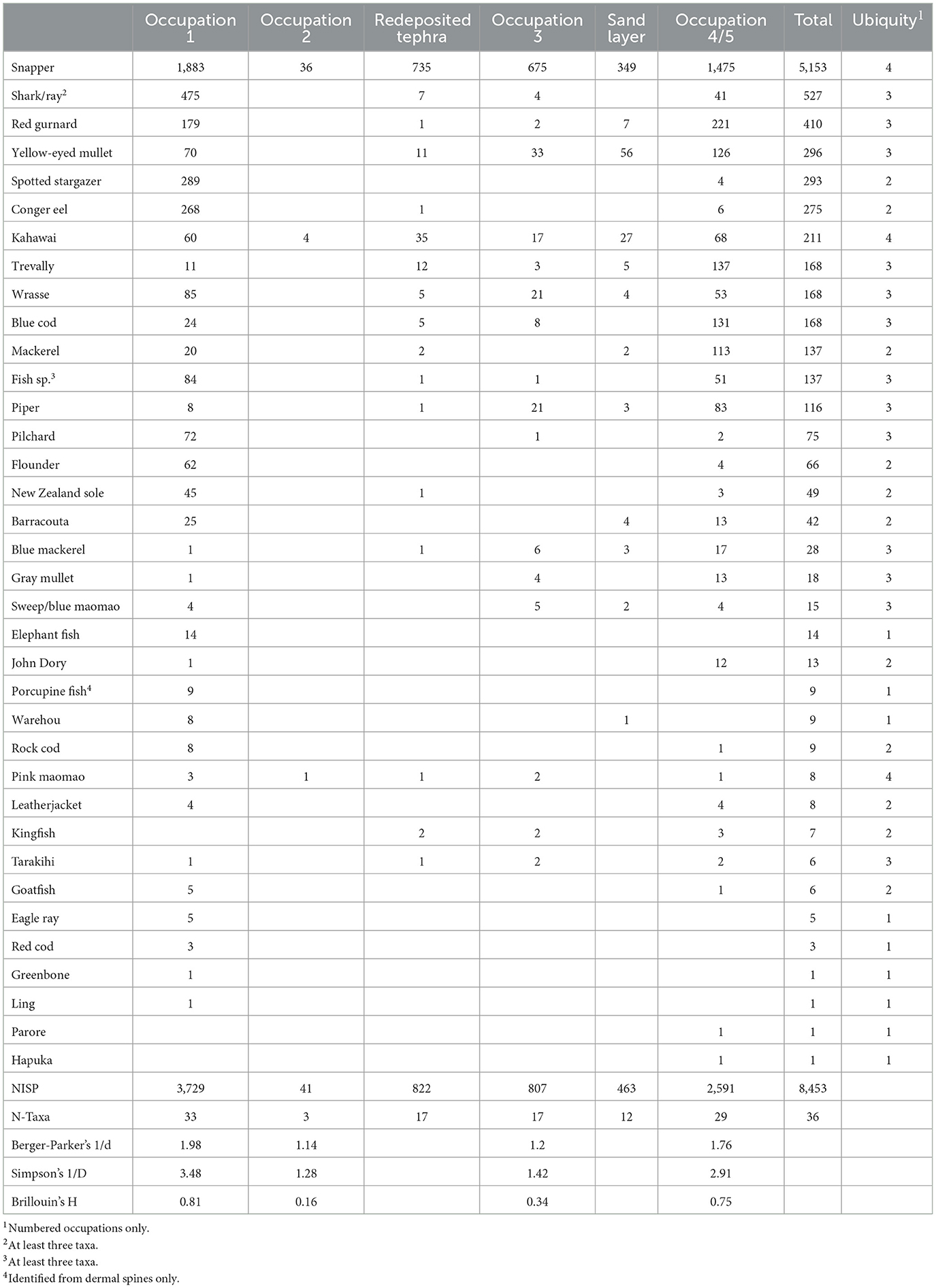
Table 2. Fish by NISP from all contexts.
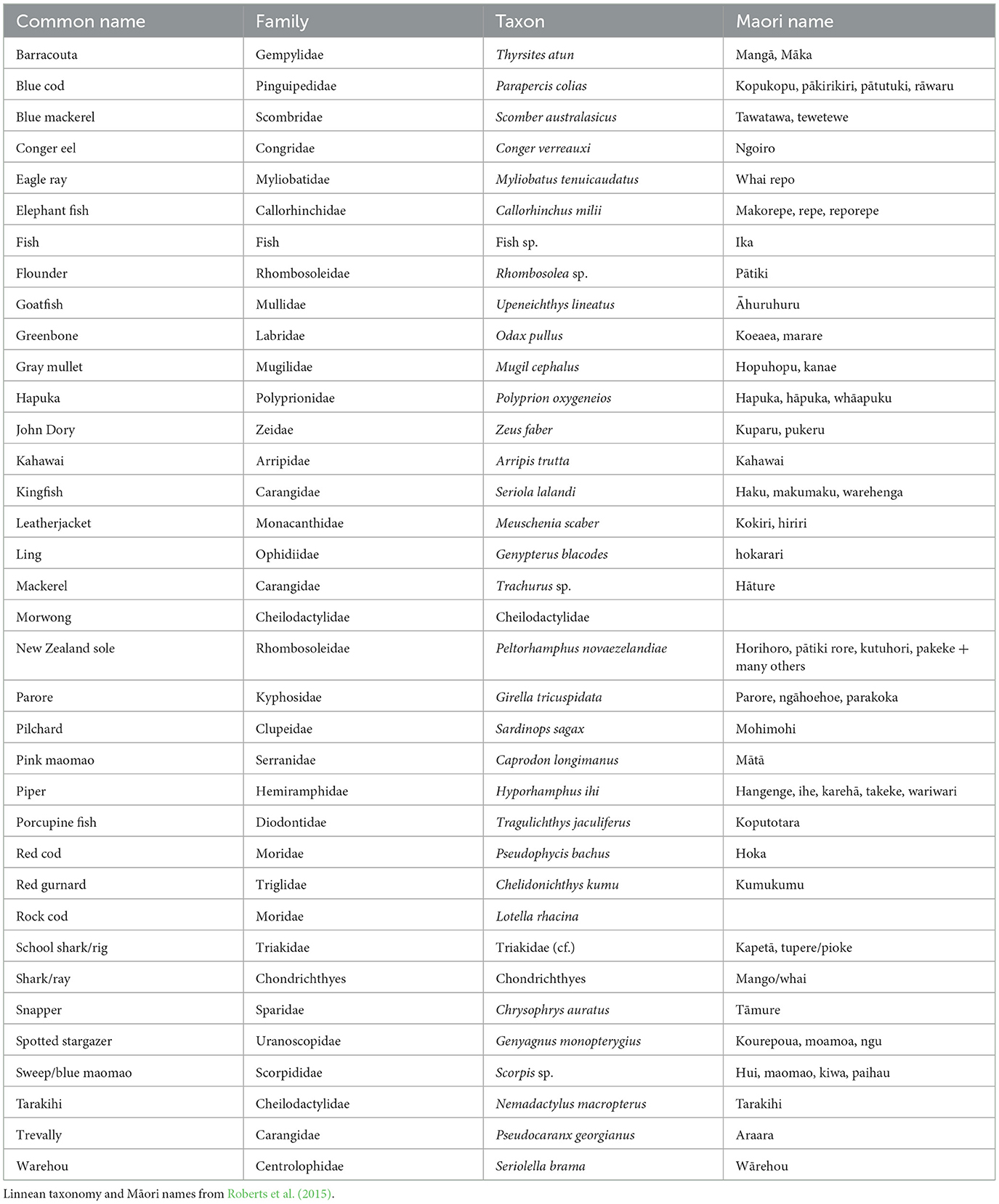
Table 3. Fish taxa identified at Ōtata.
In addition to the 34 fish taxa identified to low taxonomic level, several taxa were present in small numbers that could not be identified with the available comparative collections. Based on vertebrae, there were at least 3 species of bony fish in this category. All these are collectively listed in Table 2 as Fish sp. Wrasses are a collective category for the subfamily Pseudolabrini, which contains several indistinguishable species with very similar lifestyles. Based again on vertebrae there were at least three shark taxa, the majority of which were identified as cf. Triakidae [either school shark (Galeorhinus galeus) or rig (Mustelus lenticulatis)] with at least two other larger bodied taxa also present (Shepherd and Campbell, 2021). All these are collectively listed as Chondrichthyes. Ray vertebrae are generally distinctive (Campbell et al., 2022) and none of the Chondrichthyes vertebrae appeared to be rays. Elephant fish (Callorhinchus milii), a ghost shark (Chimaeriformes), was the only cartilaginous fish that could be identified to low taxonomic level, from its distinctive tooth plates.
Elephant fish were found only in Square C from Occupation 1, and only in small numbers (from both the undisturbed and disturbed deposits, which gives us some confidence that the disturbed upper deposit relates to Occupation 1). Elephant fish are mostly recorded in cooler South Island waters, though some have been recorded as far north as the East Cape on the lower North Island east coast (Anderson et al., 1998), and there is a record as far north as Muriwai on the west coast (Tom Trnski, pers. comm. October 3, 2024). Elephant fish are a benthic species to 200 m that aggregate in spring or early summer to spawn (Roberts et al., 2015, p. 39; Francis, 2024, p. 47), so could have come from the inner or outer Hauraki Gulf. They are not known from the gulf today. Gulf waters are warmer than at Muriwai or East Cape (Greig et al., 1988; Uddstrom and Oien, 1999) and would probably not have been an ideal habitat. It is possible that these specimens were preserved fish brought on site, though firstly it seems unlikely that fish would be brought to a fishing camp, and secondly if preserved they would probably not have had their heads still attached. It is more likely that they represent a vagrant or relic population that was easily extirpated.
The assemblages are dominated by snapper (Chrysophrys auratus) (Figure 4) (a sparid or sea bream, not to be confused with the lutjanids or tropical snapper), accounting for between 50 and 88% for each occupation. Snapper is often the most common fish in upper North Island assemblages (Leach, 2006, p. 79; Nims, 2022) and the Ōtata results are in line with this. Snapper are abundant in northern waters and are an important keystone species. They are a popular sport fish today as well as forming an economically important fishery. In early summer they form spawning aggregations in open waters. They take a baited hook and can be netted. They are thought to form relatively distinct regional sub-populations, with limited mixing (Paul, 2000; Leach, 2006; Francis, 2024).
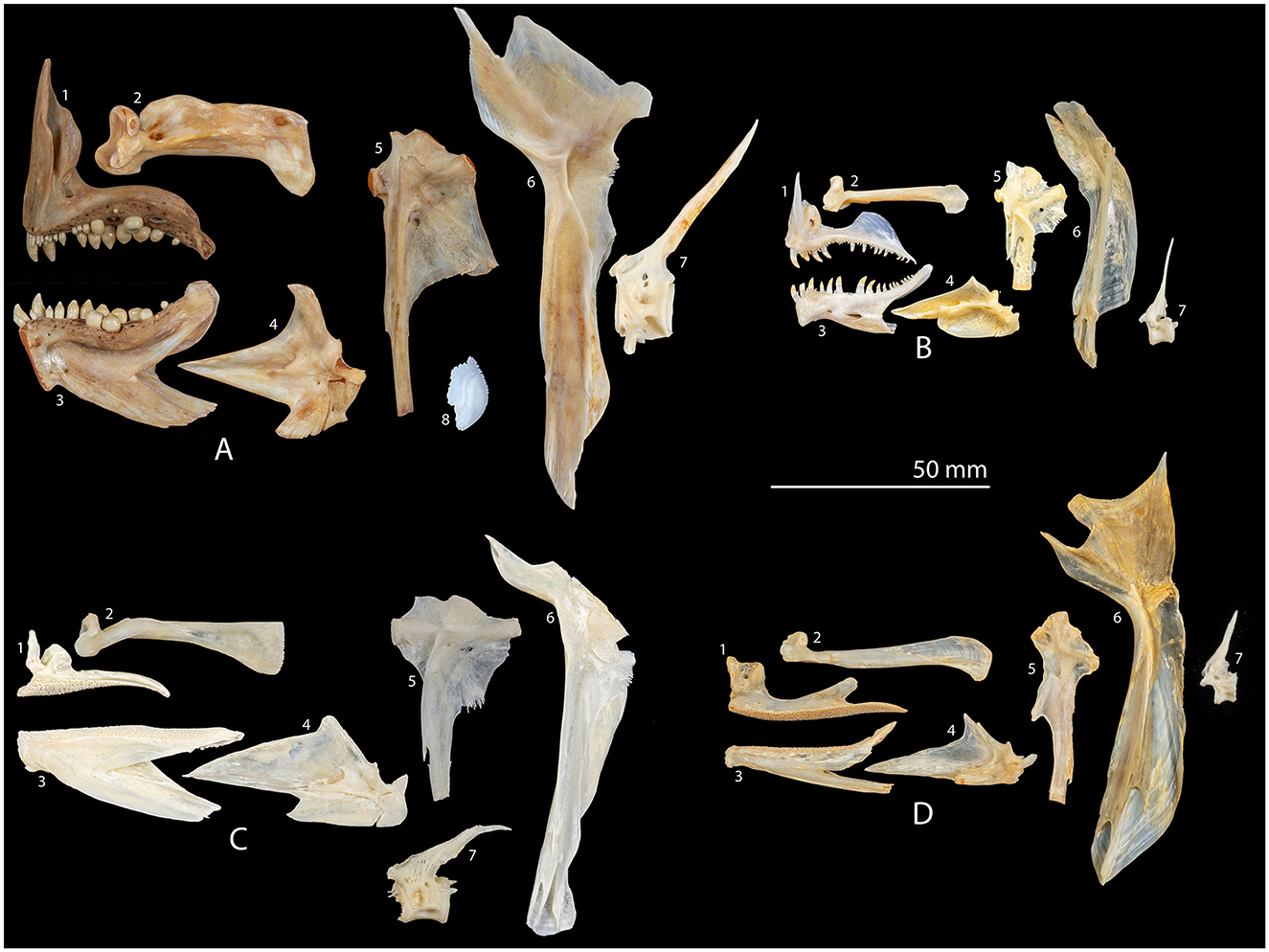
Figure 4. Mesial views of right comparative specimens of: 1 premaxilla, 2 maxilla, 3 dentary, 4 articular, 5 hyomandibular, 6 cleithrum, 7 3rd vertebra (left lateral view), 8 otolith (snapper only); for (A) snapper (Chrysophrys auratus) fork length (fl) = 525 mm, (B) blue cod (Parapercis colias) fl = 325 mm, (C) kahawai (Arripis trutta) fl = 530 mm, (D) red gurnard (Chelidonichthys kumu) fl = 365 mm.
Table 2 includes five diversity indices. NISP and N-Taxa are two that are most familiar to archaeologists, the total number of identified specimens and the total number of taxa identified. Occupation 1 has both the highest total NISP and the highest number of identified taxa, followed by Occupations 4/5, 3, and 2.
The other three diversity indices are derived from the NISP and N-Taxa counts, and are calculated for the numbered occupations only. Although developed to measure biodiversity these can be readily adopted to measure the diversity of archaeological assemblages (Magurran, 2004). The Berger-Parker index is an index of dominance, that is, how much the most common taxon dominates the assemblage. It is easy to calculate, a simple count of the total NISP divided by the NISP of the most common taxon, in each case, snapper. Simpson's index is another measure of dominance, in this case taking the structure, or evenness, of the whole assemblage into consideration. While snapper are dominant, other taxa are also present in reasonable numbers and so contribute to Simpson's D. Both Berger-Parker's d and Simpson's D are presented as their inverse, so that diversity increases as the measure increases. Brillouin's H is an appropriate diversity index when the randomness of the sample cannot be guaranteed (Magurran, 2004, p. 113), which is the case for archaeological assemblages. It is a measure of the evenness of the assemblage. All three indexes in Table 2 correlate and show that the Occupation 1 assemblage is the most even, or diverse, followed by Occupation 4/5. In fact these measures all correlate with total NISP, suggesting that if all assemblages had NISPs of 3,700 they would all be closer to each other in evenness and diversity. Total NISP, in the case of Ōtata, predicts the other diversity statistics, although this is not always the case with other assemblages (e.g., Campbell and Nims, 2019: Table 1). This probably reflects the dominance of snapper in the assemblages.
These diversity measures can be shown graphically using rank abundance, or Whittaker, plots (Magurran, 2004, p. 21). Figure 5 shows the dominance of snapper, ranked 1, then a steep slope to the next most common taxon, after which the slopes tend to be quite even (note that the y axis is a log10 scale). This suggests that, while snapper were the primary target in all occupations, other fish were not taken preferentially but rather as an incidental catch of the snapper fishery. The contents of each assemblage are essentially consistent and the occupants were undertaking the same, or at least comparable, activities on the island in each occupation.
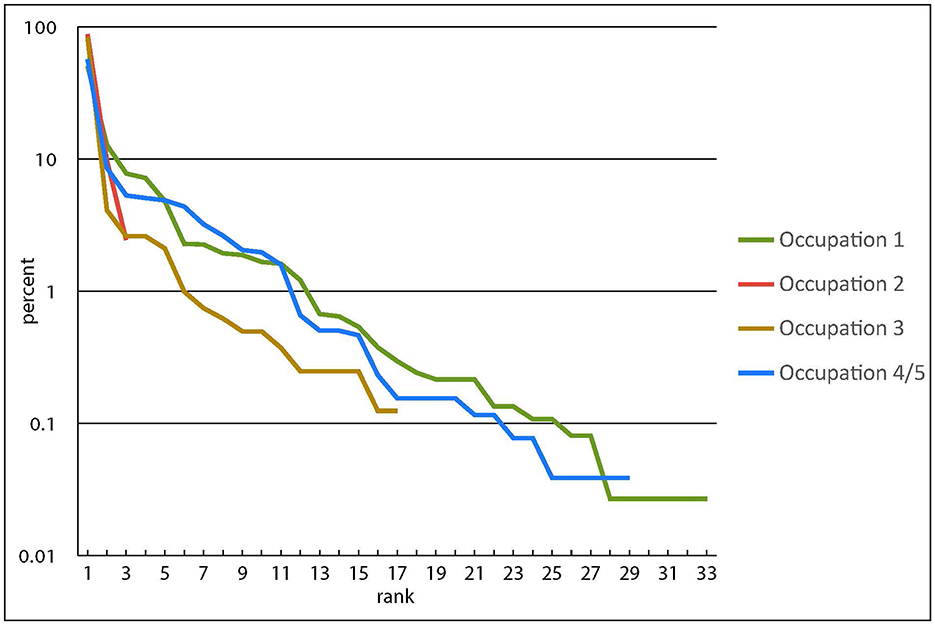
Figure 5. Rank abundance (Whittaker) plots for each occupation (the y axis is a log10 scale).
The snapper fishery
First among these activities was catching snapper. Comparing the counts of snapper cranial bones to vertebra from the assemblages shows that cranial bones far outnumber vertebrae (Figure 6). We identified 34 cranial bones including otoliths for most taxa, and snapper have 24 vertebrae. All snapper bones are robust and we would expect them to survive in roughly equal proportions. Vertebrae consistently account for only 20–30% of snapper bone for all occupations. The results are highly statistically significant (Table 4). Snapper, then, were being targeted and their bodies, including vertebrae, preserved and taken elsewhere for later, probably winter, consumption. Early explorers and ethnographers describe preservation by sun-drying, steaming and drying, or drying on hot rocks (e.g., Anon, 1837, p. 90; Best, 1977 [1929]; Beattie, 1994, p. 116). All the evidence indicates that Ōtata represents a seasonal fishing camp probably occupied in early summer, and that snapper were the primary target.
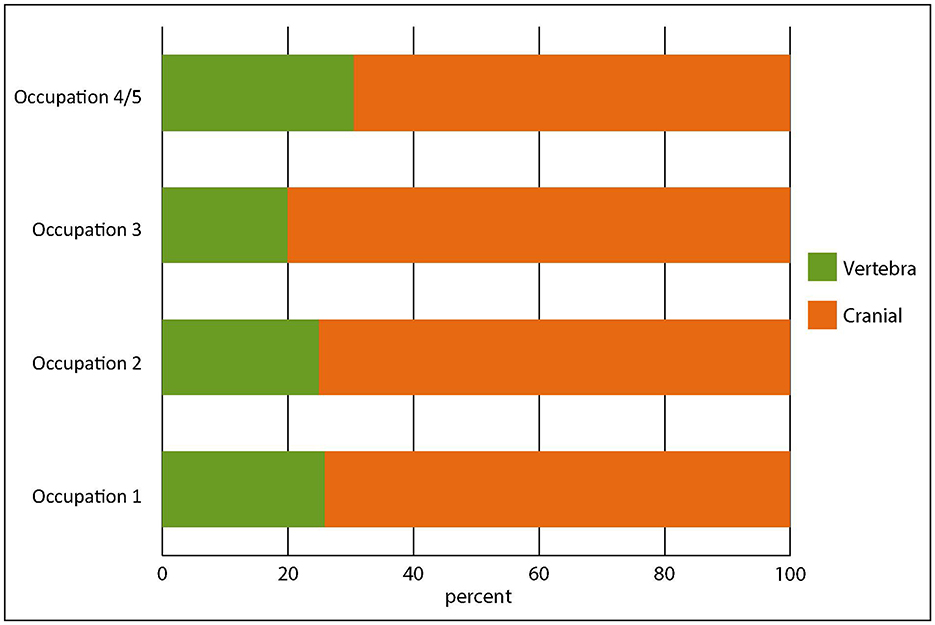
Figure 6. Proportions of snapper cranial bones and vertebra from each occupation.
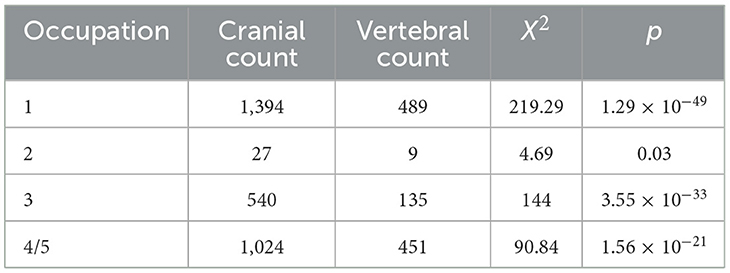
Table 4. Chi-squared scores for snapper cranial bones and vertebrae from each occupation—we counted 32 cranial bones while snapper have 24 vertebrae (Roberts et al., 2015, p. 1288).
Sharks and rays were also commonly targeted for preservation by Māori and the shark at the site, particularly in Occupation 1, may represent a shark fishery alongside the more clearly visible snapper fishery. A school shark fishery was described by numerous nineteenth century ethnographers and explorers and school shark and rig vertebrae have been identified in abundance by aDNA analysis at the Auckland Airport NRD site (R11/859), although this was not a shark fishery, rather the shark were brought on site for consumption at funerary feats (Shepherd and Campbell, 2021; Campbell et al., 2022). Cartilaginous vertebrae are vulnerable to diagenetic processes and cranial elements of cartilaginous fish do not survive, while very fine screens would be required to find Triakidae teeth or dermal denticles, so shark and ray numbers are not really comparable to bony fish numbers. Even so, the pattern observed for snapper does not hold for other common species, with vertebrae often more common than cranial bones (Figure 7)—taxa most frequently identified from vertebrae, such as yellow-eyed mullet (Aldrichetta forsteri), pilchard (Sardinops sagax) and piper (Hyporhamphus ihi), which have small, fragile cranial bones, or conger eel (Conger verreauxi), which have 150–160 vertebrae (Roberts et al., 2015, p. 278), are not included.
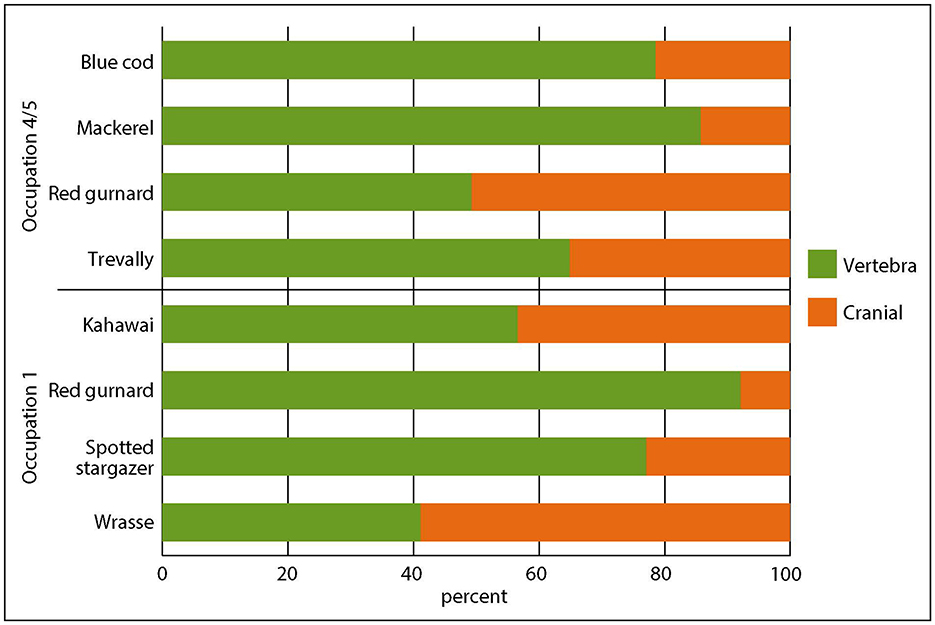
Figure 7. Proportions of cranial bones and vertebra for selected taxa from Occupations 1 and 4/5 where NISP > 30.
Vertebrae far outnumber cranial bones for spotted stargazers (Genyagnus monopterygius) in Occupation 1. Stargazers (Uranoscopidae) are benthic ambush predators with armored heads, so while their head bones should survive, perhaps the fish were headed off site and only the bodies brought back. More surprising is that the same pattern is evident for red gurnard (Chelidonichthys kumu) in Occupation 1, and for blue cod (Parapercis colias) and mackerel (Trachurus sp.) in Occupation 4/5. While some of this may be due to taphonomic processes, it is more probable that cultural or even individual preferences are at play. Either way, none of these taxa were targeted for preservation in the same way that snapper evidently were (Figure 7).
Individual vs. mass capture
Other taxa mainly found in Occupation 1 are also of interest. Fish like congers and stargazers are benthic species, not easily netted though they will readily take a baited hook. Leatherjackets (Meuschenia scaber) and wrasses tend to live on reefs, where they would also not be netted. Wrasses take a baited hook but leatherjackets have small mouths and were presumably speared or taken by hand. Demersal species, such as flatfish or gurnards can be taken in nets on clean bottoms, though flatfish are commonly speared.
These taxa are less common in later occupations where fish like yellow-eyed mullet, mackerel, trevally (Pseudocaranx georgianus) or piper become more common (Figure 8). These are pelagic schooling taxa, often with small mouths, that are more likely to be taken by mass capture methods, particularly netting. Fish can live in a wide range of depths and can move up and down the water column, often diurnally, and schools of some species can aggregate or disaggregate depending on seasonal or environmental factors, especially for spawning, so by “pelagic schooling”, we mean generally epipelagic, generally schooling. An exception to this pattern is pilchard, a pelagic schooling species that is mainly present in Occupation 1 and not later occupations. A change in focus to pelagic schooling fish does not mean that these were not caught in Occupation 1, or that individually captured benthic or reef species were not caught in later occupations. At Ōtata snapper continued to be the main focus of the fishery, and these could be caught by both baited hook and net.
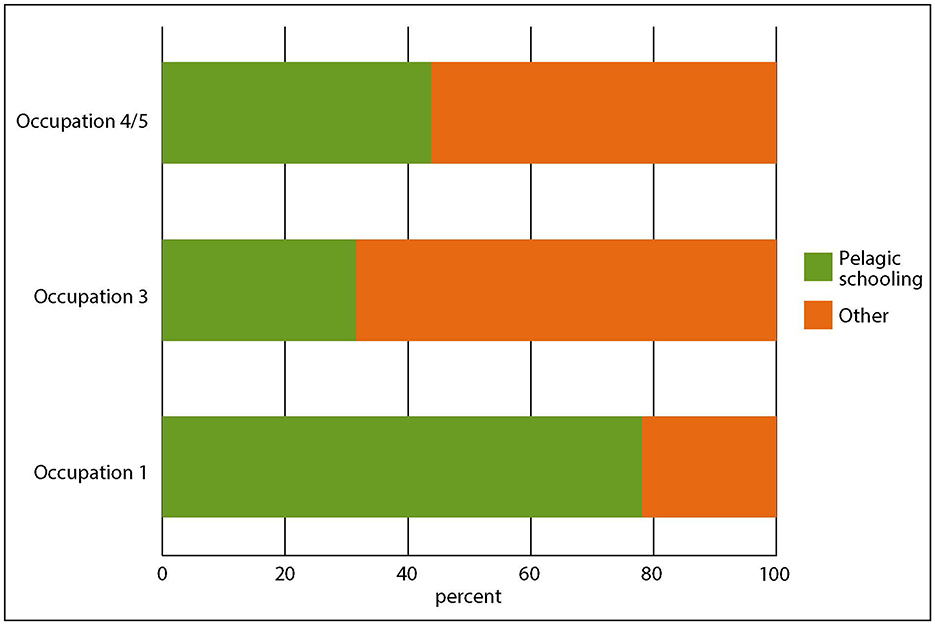
Figure 8. Changing proportions of pelagic schooling and “other” taxa, except for snapper and Chondrichthyes.
Nims (2022) noted the same general pattern throughout the northern North Island—a focus on snapper alongside rocky reef and soft substrate species prior to around 1,500 CE followed by a shift to pelagic schooling species. He proposed that loss of megafauna, anthropogenic landscape transformations and population growth put pressure on inshore fisheries, with previously dependable coastal fisheries suffering resource depression. Alongside this, social changes and population growth led Māori to expand settlement and lay claim to new territories and resources and defend these places with fortified pā. This may have necessitated more reliable and sustainable fishing practices.
These are not the only possible causes for a change of focus to pelagic schooling fish. Leach (2006, p. 250) quotes several ethnographic sources showing that Māori craved fat and oil that was often lacking in their diet. With the extinction of moa and extirpation of seals in the upper North Island, this shortage must have become pronounced. Of the fish identified from Ōtata, pelagic schooling fish often have a higher fat content than other species, which may have been another reason to target them. Vlieg (1988) provides the proximate composition of several commercial New Zealand fish species, which are reproduced in Table 5 for the fish from Ōtata that he provides data for. These figures are for filets, although Vlieg's data show that oil is distributed unevenly in head, viscera, frame and skin as well as filets, and parts of the fish other than just filets would have been eaten (Leach, 2006). The figures in Table 5 are useful for discussing relative oil content, but are not useful for calculating absolute dietary contributions.
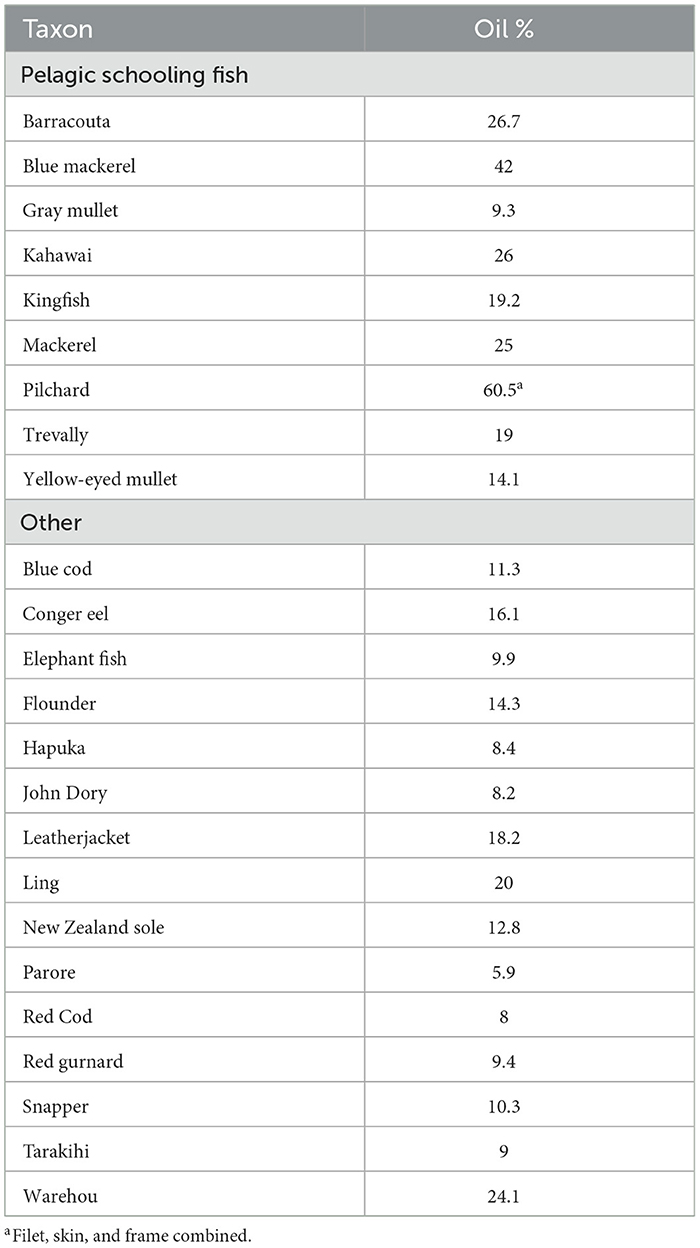
Table 5. Percentages of oil in filets of several fish species (from Vlieg, 1988).
Net technology for the mass capture of fish seems to have been particularly well developed in New Zealand. Joseph Banks, aboard the Endeavor in 1769, recorded a net that was “5 fathom [9 m] deep and its length we could only guess, as it was not stretchd out, but it could not from its bulk be less than 4 or 500 hundred fathom [730–910 m] … almost every house you go into has netts in it making” (Beaglehole, 1962, p. 444). Cruise (1974 [1824], p. 314) observed that British navy nets were “contemptible when compared with those of the New Zealanders” (see also Best, 1977 [1929]). Mair (1923, p. 19) at Maketu in 1885 described a net 95 chains (1,910 m) long that caught at least 37,000 large fish. Mair lists the fish that were counted, including: kahawai, snapper, trevally, gurnard, moki, tarakihi, parore, kingfish, school shark, other large and small sharks, rays and “uncounted numbers of smaller fish, such as … mackerel … sand fish [perhaps flounders], and some new species” (Mair, 1923, p. 22). Of these, only kahawai (Arripis trutta) and the carangids—mackerel, trevally and kingfish (Seriola lalandi)—are pelagic schooling fish. The event witnessed by Mair may not be a good model for interpreting archaeological assemblages, or it may indicate that our interpretations of what happened to fish once they were landed are flawed.
Shaped stone net weights or pumice floats are recovered archaeologically but not as commonly as a reliance on netting would suggest. Most weights and floats would have been expedient, with suitable stone cobbles sewn into the feet of nets and wooden floats tied to the heads (Witter, 2007). Wooden floats would not survive while unworked cobbles would not be recognized as net weights—the beach at Ōtata is made up of such cobbles. Hooks and two-piece hook points are more common in the archaeological record. At Ōtata several one- and two-piece hooks and points of shell and bone were found, along with evidence of manufacture on site, but only in Occupations 1 and 2 and no recognizable net gear was recovered.
A reliance on netting schooling fish is not unique to New Zealand among Polynesian societies. Nims et al. (2024) have shown that at Hakea Beach on Nuku Hiva mass capture of fish took place from early in the sequence. Hakea is a deeply indented bay with a sandy beach and limited reefs. Small bodied and small mouthed fish caught by netting included Mullidae (goat fish), Mugilidae (mullet) and small Lutjanidae, the latter in particular pointing to extensive netting inshore as larger lutjanids live in deeper waters where they are more likely to be caught by angling.
Net technology in the Marquesas is not as well described as for New Zealand. The accounts of early explorers (collated by Ferdon, 1993) describe night fishing for flying fish with hand nets, as well as “seines and straight nets, the latter up to one hundred feet in length” and a variety of hand nets (Ferdon, 1993, p. 98). Handy (1971 [1923], p. 169) describes specialized large, deep-sea nets 30 feet square, handled by four canoes, as well as nets of all sizes grading down to hand nets. In the Cook Islands Hiroa (1944) provides good descriptions of nets from the early twentieth century. Off-shore fishing, by which he presumably means outside the reef, was confined to hook and line, and nets to catch flying fish. He describes the use of long nets in the lagoon, often as stationary fish traps, but also as seines handled by two or more canoes. Short set nets were between 5 and 10 yards long, but gives no dimensions for long nets. Nonetheless, none of these nets are as large as those described by Banks, Cruise or Mair. The reason for this is simple, as Hiroa (1944, p. 222) put it: “The unevenness of the lagoon bed, studded with holes, rocks, and coral upgrowths, is not conducive to the hauling of long seine nets”. In contrast, wide bays and sandy beaches well suited to large nets are common in New Zealand. Similarly, Nims et al. (2024) pointed to the Hakea environment as enabling a reliance on mass capture with nets—they also listed reliability of catch and avoidance of ciguatera poisoning from larger reef fishes as other, non-mutually exclusive explanations for the patterns they observed.
While not all fish would take a hook, all fish can potentially be netted. Netting, however, was not the only mass capture technique used by Māori. For instance, John Boultbee in 1827 described a specialized lure technique for taking barracouta (Thyrsites atun): “sometimes they will load a canoe in two hours with fish” (Boultbee, 1986, p. 75; see also Anderson, 1981; Beattie, 1994). Barracouta were an important fish resource throughout the sequence on the South Island east coast. Lure points are well described, for instance, from the early site of Shag River Mouth, where three wooden lure shanks were recovered from wet contexts and barracouta dominate the fish assemblages (Anderson, 1981; Anderson and Gumbley, 1996; Anderson and Smith, 1996). Specialized barracouta techniques, though derived from tropical Polynesian lure forms, were clearly an early innovation in New Zealand. Similarly, pā kahawai, or kahawai trolling lures, are known in museum collections from the early nineteenth century. These were two piece lures with a shell or bone point and a wooden shank with iridescent pāua (Haliotis sp.) shell inlay. Bone and shell points are common archaeologically but cannot be attributed to a particular hook or lure form. Pāua inlays are only known from late period sites (pāua shell delaminates easily in archaeological contexts and does not survive well) so the antiquity of this specific form is unknown (Smith, 2007; see also Davidson, 1984, p. 66).
The net described by Mair was made at the behest of Te Pokiha, who was determined that it “should outrival the famous historic one made by his remote ancestor, Taramainuku” (Mair, 1923, p. 19). Fishing was frequently a competitive activity and many of the patterns that can be seen in the archaeological record are not solely attributable to economic necessity, even if competition can rarely be seen in the data.
Snapper size
Snapper mouth parts were measured and live fork lengths estimated following the method in Leach and Boocock (1995), measuring dentary, articular, maxilla and premaxilla, but not quadrate or otolith. While the data from Occupations 1 and 4/5 follow a roughly normal distribution and the fish are all essentially the same size and in the same length range, the Occupation 3 data are highly skewed toward smaller fish and the average and median lengths are significantly smaller (Table 4, Figure 8). Other than a single fish from Occupation 3, none are particularly large, with average lengths not much greater than the current allowable size limit of 300 mm (https://www.mpi.govt.nz/fishing-aquaculture/recreational-fishing/fishing-rules/), and many considerably smaller (Table 6, Figure 9).
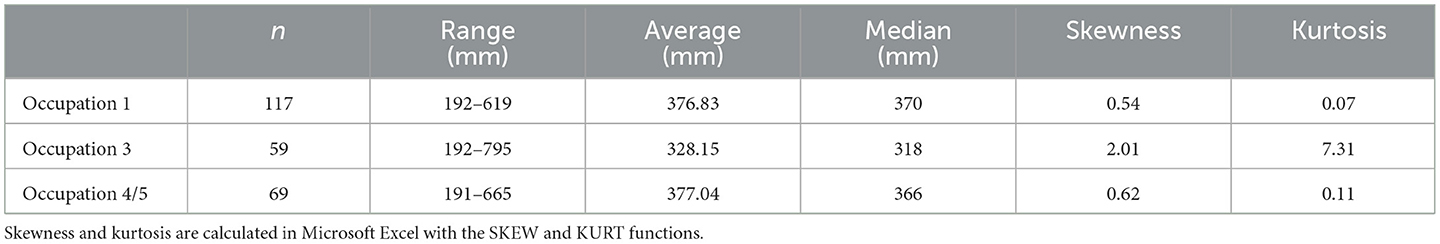
Table 6. Snapper size distribution data.
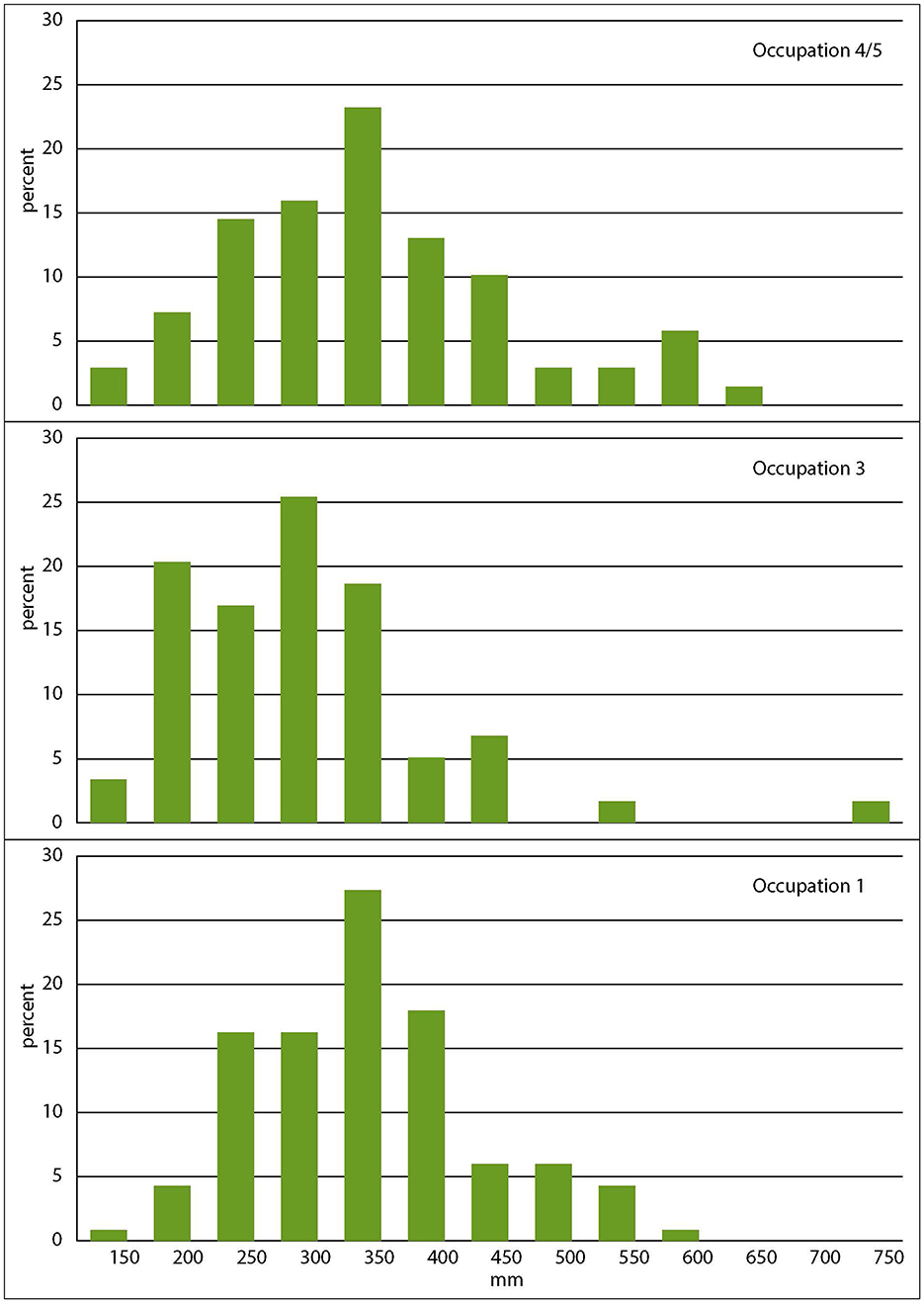
Figure 9. Snapper size distributions.
The layer beneath Occupation 3 consists of redeposited Rangitoto tephra mixed with fine clays. It is assumed to have been washed off the nearby shallow slopes soon after the eruption, before the airfall deposit had time to consolidate and become vegetated. A thin topsoil then built up on the redeposited tephra, indicating a brief time lag before Occupation 3. We interpret the Occupation 3 snapper size distribution as a direct result of the eruption. The deposition of tephra would have smothered the seafloor, disrupting the local ecosystem. Any fish that survived would have moved elsewhere in the gulf, and the waters around Ōtata and the local snapper population would have taken time to recover. The Occupation 3 snapper show this process in action, as younger fish were recruited from other parts of the gulf, skewing the catch toward smaller fish. By Occupation 4/5 the population had recovered to a typical size and age distribution.
Otolith chemistry
Much recent archaeofaunal research in New Zealand, and the Pacific more widely, has begun to focus more closely on human-environmental interactions in marine ecosystems, benefitting from multi-technical and cross-disciplinary collaborative approaches (Allen and Kahn, 2024). Such studies can provide fine-grained data on changes in fish populations and environments. Archaeological assemblages offer a unique perspective on former ecosystems and can be used to examine environmental baselines on longer timescales than are otherwise available to the life sciences.
We analyzed the changing proportions of barium (Ba) and strontium (Sr) of snapper otoliths from the Ōtata midden (Campbell et al., 2025). The otoliths were sectioned and analyzed with laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS) along a linear path from the core of the otolith to the proximal tip. We analyzed the results for the first 1,500 μm of the laser transect, roughly equivalent to the first 2 years of the fish's life. We then applied Behavioral Change Point Analysis (BCPA), a statistical method used to resolve fine-scale patterns of animal behavior and movement, tracking this life history. This identified three distinct Behavioral States (BS) characterized by distinctive proportions of Ba and Sr: BS 1, with low Sr and low Ba values; BS2, with moderate Sr and high Ba values; and BS 3, with high Sr and low Ba values (Table 7). The BCPA results for all occupations showed that from soon after the East Polynesian settlement of New Zealand (Occupation 1, fourteenth century CE) to just prior to the arrival of Europeans and their technologies (Occupation 4/5, eighteenth century), juvenile snapper exhibited consistent patterns of early habitat residency and movement. Snapper larvae settle in structured sponge and seagrass nurseries in estuarine and sheltered environments, which provide protection from predators and allow them to conserve energy, and as they grow they move to increasingly open, oceanic waters (Parsons et al., 2014, 2020; Sabetian et al., 2021). The BCPA results reinforce this, showing that in their first year they primarily reside in lower salinity nearshore environments (BS 1) and in their second year they increasingly shift to higher salinity habitats (BS 2 and 3). These findings must be interpreted with caution. While relative barium and strontium concentrations are well-established proxies for freshwater and saltwater sources (de Vries et al., 2005; Izzo et al., 2018; Macdonald and Crook, 2010; Elsdon and Gillanders, 2005), they can also be influenced by various environmental and physiological factors. That said, the confidence in our novel time-series approach lies in its ability to counterbalance the conventional point-analysis, multi-tracer methodology which primarily rely on absolute (or averaged) values to establish habitat residency at specific time points in a fish's life. In contrast, our approach provides a continuous, high-resolution reconstruction of habitat transitions, offering a more nuanced perspective on environmental history.
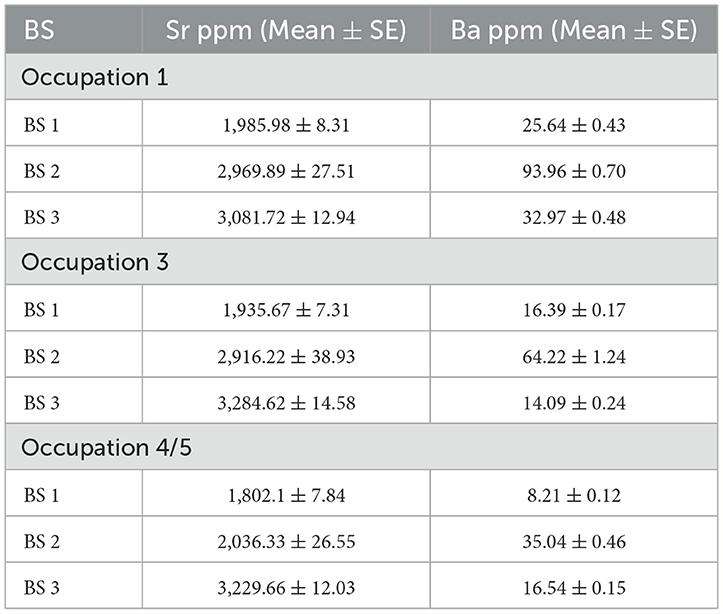
Table 7. Sr and Ba mean concentrations for each behavioral state for each occupation.
Movement patterns of juvenile snapper did not change during the pre-European period in the Hauraki Gulf, further reinforcing the conclusion that Māori fishing pressures had no measurable effect on snapper populations and behaviors. While behaviors may not have changed, the BCPA analysis did shows that habitat characteristics did, with the same patterns observed across the occupations. BS 1 was represented by the lowest Sr and Ba concentrations for all occupations (Table 5), indicating the consistent use of low salinity sheltered nearshore nursery habitats away from any major freshwater influence. Another pattern was that BS 2 was characterized by mid-level Sr and the highest Ba concentrations for all occupations. This chemical signature is typical of shallow mixing zones in coastal areas where freshwater and seawater converge (Wilson, 2005). This suggests that this area also served as a transitional habitat for juvenile snapper as they began to move from sheltered nurseries to open waters. This is supported by the observation that all specimens spent the least amount of time in BS 2: Occupation 1 (22.4%), Occupations 3 (7.2%), and Occupation 4/5 (12.8%). The third and most interesting pattern was in Ba concentrations in BS 2 which were, on average, 1.5 times higher in Occupation 1 than in Occupation 3, and 2.5 times higher than in Occupation 4/5.
Ba concentrations are expected to vary in a dynamic environment such as the Hauraki Gulf (Alibert et al., 2003; Hamer et al., 2003, 2006; Elsdon and Gillanders, 2006) but a 60% decline of Ba concentrations between the fourteenth and eighteenth centuries requires explanation. Ba and Sr are both in group 2 of the periodic table, which they share with Ca, and their carbonates are absorbed into otoliths as they substitute for CaCO3. In the marine environment Ba is influenced by a broad range of environmental and biological factors. Ba from freshwater sources is in a soluble, bioavailable form, BaCO3, while as it is flushed into saline environments and comes into contact with sulfate-rich sea floor sediments it converts to mineral, non-bioavailable BaSO4, which is why it is such a useful marker of freshwater influences in otoliths. Ba concentrations are also strongly influenced by freshwater inputs, fluvial runoff, volcanic activity, sediment resuspension, and biological processes such as adsorption onto particles (Cao et al., 2016). In the case of the Hauraki Gulf, the Rangitoto volcano was active for at least 150 years, and probably longer, before its eruption ceased around 1,450 CE (Shane et al., 2013; Allington et al., 2023). At Ōtata this is marked by the tephra layer between Occupations 1 and 3. Occupation 1, which has the highest Ba concentration in BS 2, was during a period of active volcanism that had ceased by the time of Occupation 3. Weathering and erosion of fresh volcanic deposits potentially contributed to further enrichment of Ba in gulf waters (Lein and Kravchishina, 2021). The decline in Ba values in Occupations 3 and 4/5 could, then, be due to the cessation of volcanic activity. However, this Ba would seem not to have been in a bioavailable form as the elevated Ba signature does not show up in BS 3 for Occupation 1 when the fish were moving into more open waters closer to the ostensible source of volcanic Ba. We cannot entirely rule volcanism out as an explanation, but it seems unlikely.
A more probable explanation lies in altered hydrodynamics of the Hauraki Gulf's mixing zones, which in Occupation 1 are also transitional zones for juvenile snapper, leading to shifts in Sr and Ba concentrations in later occupations. High Ba levels show that Ba remains enriched in these mixing zones throughout the pre-European sequence, but this enrichment diminished over time. Decreasing Ba levels may be linked to a transition in the Gulf's freshwater inputs or shifting fluvial sediment dynamics, with lower concentrations of Ba in the later occupations suggesting altered freshwater input into the coastal ecosystem (Sinclair and McCulloch, 2014). One possible cause is deforestation initiated by Māori for activities such as garden clearance (McWethy et al., 2010, 2014), leading to increased sedimentation and changing amount, frequency and variability of freshwater inputs. There are several possible ways in which this could be reflected in changing Ba levels. The fluvial mixing zone may move further offshore so that it no longer coincides so neatly with the juvenile snapper transitional zone; or sedimentation may directly impact the transitional zone, shrinking the extent of nursery habitats so that their margin moves inshore away for the Ba rich mixing zone; or the amount and distribution of Ba in marine waters may change; or the mechanics of the mixing zones may be altered. Any or all of these may be the case, along with others that we have not considered. We can't provide a definitive answer, but juvenile snapper in transitional zones are no longer swimming in the same Ba rich waters in later occupations as they were in Occupation 1.
Discussion and conclusion
This paper has examined change and continuity in the pre-European Māori fishery at Ōtata. Anthropogenic impacts on terrestrial ecosystems in pre-European New Zealand, such as extinctions, extirpations and deforestation, are well understood. But are there parallel processes taking place in marine ecosystems? Because of the different ways humans interact with terrestrial and marine environments, evidence of change in the latter is likely to be subtle. Wells et al. (2019) demonstrate changes in growth rates of tuangi, a filter-feeding intertidal bivalve, probably as a result of increased sedimentation following land clearance. There is no evidence that tuangi were over-harvested. Pre-European Māori population densities were never high and the period between first settlement and the advent of European technology is barely 500 years. Māori fishing technology was sophisticated but it is unclear that it could have had any measurable impact on fish populations in that period. Snapper stocks, for instance, were resilient in the face of European fishing until the removal of restrictions on the fishery and the development of industrial scale fishing in the 1960s (Mossman, 2008).
Ōtata is an ideal location for an analysis of fishing, a clearly stratified midden on a small island easily accessed from the mainland, but also a location where the range of settlement and subsistence activities was limited. Ōtata is not suited to horticulture or permanent settlement and has no useful lithic resources. Fishing and shellfishing, and some birding, were the reasons for visiting the island. A “conventional” analysis of the fish from the Ōtata midden—that is, identifying and counting a subset of fish bones to the lowest possible taxonomic level, a consideration of the structure of the assemblages and of environments and fishing technologies—has not demonstrated any changes that we can attribute to anthropogenic environmental change or fishing pressures. A change from individual to mass capture after Occupation 1 follows the development of netting technologies, probably accompanied by demographic pressures, a trend observed throughout the northern North Island. The population structure of the Occupation 3 snapper, skewed toward smaller individuals, is best explained as a result of a natural environmental event, the Rangitoto eruption. Overall, the activities represented in the fishbone assemblages were consistent across all occupations—targeting snapper for preservation and taking other fish as an incidental or expedient catch. Pre-European Māori fishing activities would seem to have had no discernible impact on fish populations in the gulf.
Similarly, BCPA has not shown any changes in otolith chemistry that demonstrate changes in juvenile (approximately the first 2 years of life) snapper behavior or movement patterns. Larvae consistently settled in structured nearshore nursery habitats and as they matured they gradually moved to more open, oceanic waters. What the analysis demonstrated was that the environments the fish were swimming in did change in the ~400 years between Occupation 1 and Occupation 4/5, particularly the transitional zone where juvenile snapper begin to move away from the shelter of sponge and seagrass nurseries. We attribute this to anthropogenic land-based changes—deforestation and increased sedimentation altered the hydrodynamics and chemistry of the zone where fresh and salt water mix, and altered the balance between this mixing zone and the transitional snapper zone. A 60% change in Ba concentrations does not, however, mean that the marine environment as a whole underwent a comparable alteration. Instead, these changes were probably quite subtle and had no long-term effect on Māori fishing.
Our analysis supports previous research that suggests changes in the marine environment and effects on marine organisms in the pre-European period were restricted to the nearshore environment and that there were no substantial changes to fish populations or population structures (e.g., Leach, 2006; Davidson, 2011; Wells et al., 2019). The apparent extirpation of a vulnerable population of elephant fish may appear notable, but it is as likely that this would not have been a viable population even without Māori fishing.
New Zealand's short history provides unique opportunities to examine pre-human and relatively unaltered early human environments. It also offers the opportunity to examine how a pre-modern people settled into new environments, how they changed their environments and how their environments changed them. As Groube (1967, p. 11) wrote nearly 60 years ago: “the first people who came here (East Polynesian) were a neolithic, fishing, agricultural people… When Cook came to these shores the New Zealand Maori were still a neolithic, fishing, agricultural people”. There were changes in demography, technology, subsistence, art, settlement patterns and social structures, and Māori effects on their terrestrial environments were often extensive and significant. In contrast, effects on the marine environment were localized and low-impact, with no observable effects on fish stocks.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Author contributions
MC: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Supervision, Validation, Writing – original draft. RW: Writing – review & editing. AS: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Writing – review & editing. LF: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Writing – review & editing. EA: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Writing – review & editing. AM: Formal analysis, Investigation, Writing – review & editing. JZ: Formal analysis, Investigation, Methodology, Writing – review & editing. CH: Data curation, Formal analysis, Investigation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The excavation was funded by the Auckland War Memorial Museum Tāmaki Paenga Hira through the Edward Earle Vaile Fund. Analysis was funded by the Green Foundation for Polynesian Anthropology and The Noises Charitable Trust. The LA-ICP-MS analysis was funded by the Faculty of Health and Environmental Sciences, Auckland University of Technology Te Wānaga Aronui o Tāmaki Makau Rau.
Acknowledgments
Our thanks to our project partners Ngāi Tai ki Tāmaki and The Noises Charitable Trust.
Conflict of interest
Authors MC and CH were employed by CFG Heritage Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fearc.2025.1565503/full#supplementary-material
References
Alibert, C., Kinsley, L., Fallon, S. J., McCulloch, M. T., Berkelmans, R., and McAllister, F. (2003). Source of trace element variability in Great Barrier Reef corals affected by the Burdekin flood plumes. Geochim. Cosmochim. Acta 67, 231–246. doi: 10.1016/S0016-7037(02)01055-4
Allen, M. S., and Kahn, J. G. (2024). Advances in East Polynesian zooarchaeology: special issue introduction, review (2016–2024), and assessment. Archaeol. Oceania 59, 157–175. doi: 10.1002/arco.5329
Allington, M. L., Nilsson, A., Hill, M. J., Suttie, N., Daniil, D., Hjorth, I., et al. (2023). Constraining the eruption history of Rangitoto volcano, New Zealand, using palaeomagnetic data. Quater. Geochronol. 78, 101459. doi: 10.1016/j.quageo.2023.101459
Anderson, A. J. (1981). Barracouta fishing in prehistoric and early historic New Zealand. J. Soc. Océan. 37, 145–158. doi: 10.3406/jso.1981.3056
Anderson, A. J. (1991). The chronology of colonization in New Zealand. Antiquity 65, 767–795. doi: 10.1017/S0003598X00080510
Anderson, A. J. (1997). Uniformity and regional variation in marine fish catches from prehistoric New Zealand. Asian Perspect. 36, 1–26.
Anderson, A. J. (2013). “A fragile plenty: pre-European Māori and the New Zealand environment,” in Making a New Land: Environmental Histories of New Zealand, eds E. Pawson, and T. Brooking (Dunedin: Otago University Press), 35–51.
Anderson, A. J., Binney, J., and Harris, A. (2014). Tangata Whenua: An Illustrated History. Wellington: Bridget Williams Books.
Anderson, A. J., and Gumbley, W. (1996). “Fishing gear,” in Shag River Mouth: The Archaeology of an Early Southern Maori Village, eds A. J. Anderson, B. Allingham, and I. W. G. Smith (Canberra: ANH Publications), 148–160.
Anderson, A. J., and Smith, I. W. G. (1996). “Fish remains,” in Shag River Mouth: The Archaeology of an Early Southern Maori Village, eds A. J. Anderson, B. Allingham, and I. W. G. Smith (Canberra: ANH Publications), 237–244.
Anderson, O. F., Bagley, N. W., Hurst, R. J., Francis, M. P., Clark, M. R., and McMillan, P. J. (1998). Atlas of New Zealand Fish and Squid Distributions from Research Bottom Trawls. NIWA Technical Report, 42. Wellington: NIWA.
Anon (1837). The British Colonization of New Zealand; Being an Account of the Principles, Objects, and Plans of the New Zealand Association; Together with Particulars Concerning the Position, Extent, Soil and Climate, Natural Productions, and Native Inhabitants of New Zealand. London: John W. Parker.
Beaglehole, J. C., (ed.). (1962). The Endeavour Journal of Joseph Banks 1768-1771, Volume 1. Sydney: Trustees of the Public Library of New South Wales in Association with Angus and Robertson.
Best, E. (1977). [1929]. Fishing Methods and Devices of the Maori. Wellington: Keating EC, Government Printer.
Boultbee, J. (1986). Journal of a Rambler: The Journal of John Boultbee. Auckland: Oxford University Press.
Campbell, M. (2016). Body part representation and the extended analysis of New Zealand fishbone. Archaeol. Ocean. 51, 18–30. doi: 10.1002/arco.5079
Campbell, M., and Nims, R. (2019). Small screens, small fish and the diversity of pre-European Māori fish catches. J. Pac. Archaeol. 10, 43–54. doi: 10.70460/jpa.v10i2.290
Campbell, M., Shepherd, L., Kellett, M., and Brassey, R. (2022). A highly fragrant comestible: the cartilaginous fish (Chondrichthyes) in pre-European Māori New Zealand. Archaeol. Ocean. 57, 1–15. doi: 10.1002/arco.5248
Campbell, M., Walter, R., Furey, L., Ash, E., McAlister, A., Tromp, M., et al. (2025). Reconstructing pre-European marine habitats using archaeological assemblages: a case-study from the Otata midden, New Zealand. J. Archaeol. Sci. Rep. 64, 105160. doi: 10.1016/j.jasrep.2025.105160
Cao, Z., Siebert, C., Hathorne, E. C., Dai, M., and Frank, M. (2016). Constraining the oceanic barium cycle with stable barium isotopes. Earth Planet. Sci. Lett. 434, 1–9. doi: 10.1016/j.epsl.2015.11.017
Collins, C. J., Rawlence, N. J., Worthy, T. H., Scofield, R. P., Tennyson, A. J. D., Smith, I. W. G., et al. (2014). Pre-human New Zealand sea lion (Phocarctos hookeri) rookeries on mainland New Zealand. J. R. Soc. New Zeal. 44, 1–37. doi: 10.1080/03036758.2013.828761
Cruise R. A. (1974) [1824]. Journal of a Ten Month's Residence in New Zealand. 2nd Edn. Christchurch [Longman, Hurst, Rees, Orme, Brown, and Green, London]: Capper Press..
Davidson, J. M. (2011). Archaeological investigations at Maungarei: a large Māori settlement on a volcanic cone in Auckland, New Zealand. Tuhinga 22, 19–100.
de Vries, M. C., Gillanders, B. M., and Elsdon, T. S. (2005). Facilitation of barium uptake into fish otoliths: influence of strontium concentration and salinity. Geochim. Cosmochim. Acta 69, 4061–4072. doi: 10.1016/j.gca.2005.03.052
Elsdon, T. S., and Gillanders, B. M. (2005). Alternative life-history patterns of estuarine fish: Barium in otoliths elucidates freshwater residency. Can. J. Fish. Aquatic Sci. 62, 1143–1152. doi: 10.1139/f05-029
Elsdon, T. S., and Gillanders, B. M. (2006). Identifying migratory contingents of fish by combining otolith Sr:Ca with temporal collections of ambient Sr:Ca concentrations. J. Fish Biol. 69, 643–657. doi: 10.1111/j.1095-8649.2006.01136.x
Ferdon, E. N. (1993). Early Observations of Marquesan Culture, 1593–1813. Tucson: The University of Arizona Press.
Francis, M. P. (2024). Coastal Fishes of New Zealand: A Comprehensive Guide to Identification and Behaviour. Nelson: Potton and Burton.
Greig, M. J., Ridgway, N. M., and Shakespeare, B. S. (1988). Sea surface temperature variations at coastal sites around New Zealand. New Zeal. J. Marine Freshw. Res. 22, 391–400. doi: 10.1080/00288330.1988.9516310
Groube, L. M. (1967). Models in prehistory: a consideration of the New Zealand evidence. Archaeol. Phys. Anthropol. Ocean. 2, 1–27.
Hamer, P. A., Jenkins, G. P., and Coutin, P. (2006). Barium variation in Pagrus auratus (Sparidae) otoliths: a potential indicator of migration between an embayment and ocean waters in south-eastern Australia. Estuar. Coast. Shelf Sci. 68, 686–702. doi: 10.1016/j.ecss.2006.03.017
Hamer, P. A., Jenkins, G. P., and Gillanders, B. M. (2003). Otolith chemistry of juvenile snapper Pagrus auratus in Victorian waters: natural chemical tags and their temporal variation. Marine Ecol. Prog. Series 263, 261–273. doi: 10.3354/meps263261
Handy E. S. C. (1971) [1923]. The Native Culture in the Marquesas. Bernice P. Bishop Museum Bulletin, 9. Kraus Reprint Co. New York NY: [Bernice P. Bishop Museum, Honolulu].
Hiroa, T. R. (1944). Arts and Crafts of the Cook Islands. Bernice P. Bishop Museum Bulletin, 179. Bernice P. Honolulu: Bishop Museum Bulletin.
Holdaway, R., and Jacomb, C. (2000). Rapid extinction of the moas (Aves: Dinornithiformes): model, test and implications. Science 287, 2250–2254. doi: 10.1126/science.287.5461.2250
Izzo, C., Reis-Santos, P., and Gillanders, B. M. (2018). Otolith chemistry does not just reflect environmental conditions: a meta-analytic evaluation. Fish Fish. 19, 441–454. doi: 10.1111/faf.12264
Leach, B. F. (2006). Fishing in Pre-European New Zealand. Wellington: New Zealand Journal of Archaeology Special Publication. New Zealand Archaeological Association.
Leach, B. F., and Boocock, A. (1995). Estimating live fish catches from archaeological bone fragments of snapper, Pagrus auratus. Tuhinga Rec. Mus. New Zeal. Te Papa Tongarewa 3, 1–28. doi: 10.3897/tuhinga.3.e34121
Leach, B. F., and Davidson, J. M. (2000). Pre-European catches of snapper (Pagrus auratus) in Northern New Zealand. J. Archaeol. Sci. 27, 509–522. doi: 10.1006/jasc.1999.0474
Leach, B. F., and Davidson, J. M. (2001). The use of size–frequency diagrams to characterize prehistoric fish catches and to assess human impact on inshore fisheries. Int. J. Osteoarchaeol. 11, 150–162. doi: 10.1002/oa.553
Lein, A. Y., and Kravchishina, M. D. (2021). Barium geochemical cycle in the ocean. Lithol. Min. Resour. 56, 293–308. doi: 10.1134/S0024490221040052
Macdonald, J. I., and Crook, D. A. (2010). Variability in Sr: Ca and Ba: Ca ratios in water and fish otoliths across an estuarine salinity gradient. Marine Ecol. Prog. Series 413, 147–161. doi: 10.3354/meps08703
McWethy, D. B., Whitlock, C., Wilmshurst, J. M., McGlone, M. S., Fromont, M., Li, X., et al. (2010). Rapid landscape transformation in South Island, New Zealand, following initial Polynesian settlement Proc. Natl. Acad. Sci. 107, 21343–21348. doi: 10.1073/pnas.1011801107
McWethy, D. B., Wilmshurst, J. M., Whitlock, C., Wood, J. R., and McGlone, M. S. (2014). A high-resolution chronology of rapid forest transitions following Polynesian arrival in New Zealand. PLoS ONE 9, e0111328. doi: 10.1371/journal.pone.0111328
Mossman, S. (2008). Snapper: New Zealand's Greatest Fish Te Ika Rangatira o Aoteraoa. Auckland: AUT Media.
Needham, A. J., Lindsay, J. M., Smith, I. E. M., Augustinus, P., and Shane, P. A. (2011). Sequential eruption of alkaline and sub-alkaline magmas from a small monogenetic volcano in the Auckland Volcanic Field, New Zealand. J. Volcanol. Geotherm. Res. 201, 126–142. doi: 10.1016/j.jvolgeores.2010.07.017
Nims, R. (2022). The archaeological ecodynamics of Māori fisheries: climate change, harvest effects, and mass capture strategies in northern Aotearoa (PhD thesis). University of Auckland, Auckland, New Zealand.
Nims, R., Pillay, P., and Allen, M. S. (2024). Mass capture fishing in the Marquesas Islands. Archaeol. Ocean. 59, 234–250. doi: 10.1002/arco.5320
Parsons, D. M., Buckthought, D., Edhouse, S., and Lohrer, A. M. (2020). The paradox of the Hauraki Gulf snapper population: testing the nursery habitat concept. Marine Ecol. 41, 1–11. doi: 10.1111/maec.12582
Parsons, D. M., Sim-Smith, C. J., Cryer, M., Francis, M. P., Hartill, B., Jones, E. G., et al. (2014). Snapper (Chrysophrys auratus): a review of life history and key vulnerabilities in New Zealand. New Zeal. J. Marine Freshw. Res. 48, 256–283. doi: 10.1080/00288330.2014.892013
Roberts, C. D., Stewart, A. L., and Struthers, C. D. (eds.), (2015). The Fishes of New Zealand. Wellington: Te Papa Press.
Sabetian, A., Zhang, J., Campbell, M., Walter, R., Allen, H., Reid, M., et al. (2021). Fish nearshore habitat-use patterns as ecological indicators of nursery quality. Ecol. Indicat. 131, e108225. doi: 10.1016/j.ecolind.2021.108225
Shane, P., Gehrels, M., Zawalna-Geer, A., Augustinus, P., Lindsay, J., and Chaillou, I. (2013). Longevity of a small shield volcano revealed by crypto-tephra studies (Rangitoto volcano, New Zealand): change in eruptive behavior of a basaltic field. J. Volcanol. Geother. Res. 257, 174–183. doi: 10.1016/j.jvolgeores.2013.03.026
Shawcross, W. (1967). An evaluation of the theoretical capacity of a New Zealand harbour to carry a human population. Tane 13, 3–11.
Shepherd, L. D., and Campbell, M. (2021). Ancient DNA analysis of an archaeological assemblage of Chondrichthyes vertebrae from South Auckland, New Zealand. J. Archaeol. Sci. Rep. 36, 102830. doi: 10.1016/j.jasrep.2021.102830
Sinclair, D. J., and McCulloch, M. T. (2014). Corals record low mobile barium concentrations in the Burdekin River during the 1974 flood: evidence for limited Ba supply to rivers? Palaeogeogr. Palaeoclimatol. Palaeoecol. 214, 155–174. doi: 10.1016/S0031-0182(04)00398-0
Smith, I. W. G. (2002). “Retreat and resilience: fur seals and human settlement in New Zealand,” in The Exploitation and Cultural Importance of Sea Mammals. Proceedings of the 9th Conference of the International Council of Archaeozoology, Durham, August 2002, ed G. G. Monks (Oxford: Oxbow Books), 6–18.
Smith, I. W. G. (2007). “Metal pa kahawai—a post-contact fishing lure form in northern New Zealand.” in Vastly Ingenious: The Archaeology of Pacific Material Culture: In Honour of Janet Davidson M, eds A. Anderson, K. Green, and B. F. Leach (Dunedin: Otago University Press), 69–78.
Uddstrom, M. J., and Oien, N. A. (1999). On the use of high-resolution satellite data to describe the spatial and temporal variability of sea surface temperatures in the New Zealand region. J. Geophys. Res. 104, 20729–20751. doi: 10.1029/1999JC900167
Vlieg, P. (1988). Proximate Composition of New Zealand Marine Finsfish and Shellfish. Palmerston North: Biotechnology Division, Department of Scientific and Industrial Research.
von Haast, J. (1871). Moas and moa hunters: address to the Philosophical Institute of Canterbury. Trans. Proc. New Zeal. Inst. 4, 66–107.
Walter, R., Buckley, H., Jacomb, C., and Matisoo-Smith, L. (2017). Mass migration and the Polynesian settlement of Aotearoa/New Zealand. J. World Prehist. 30, 351–376. doi: 10.1007/s10963-017-9110-y
Wells, S. R., Wing, L. C., Smith, A. M., and Smith, I. W. G. (2019). Historical changes in bivalve growth rates indicate ecological consequences of human occupation in estuaries. Aquatic Conserv. Marine Freshw. Ecosyst. 29, 1452–1465. doi: 10.1002/aqc.3039
Wilson, A. M. (2005). Fresh and saline groundwater discharge to the ocean: a regional perspective. Water Resour. Res. 41, 3399. doi: 10.1029/2004WR003399
Witter, A. (2007). Stone net weights at Pegasus Town, North Canterbury. Archaeol. New Zeal. 50, 175–185.
Keywords: New Zealand, Hauraki Gulf, Pre-European Māori, fish, snapper (Chrysophrys auratus)
Citation: Campbell M, Walter R, Sabetian A, Furey L, Ash E, McAlister A, Zhang J and Haylock C (2025) Fish from the Ōtata midden: change and continuity in pre-European Māori fisheries. Front. Environ. Archaeol. 4:1565503. doi: 10.3389/fearc.2025.1565503
Received: 23 January 2025; Accepted: 30 April 2025;
Published: 02 July 2025.
Edited by:
Clara Boulanger, University College London, United KingdomReviewed by:
Jen Harland, University of the Highlands and Islands, United KingdomAbhayan GS, University of Kerala, India
Copyright © 2025 Campbell, Walter, Sabetian, Furey, Ash, McAlister, Zhang and Haylock. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Matthew Campbell, bWNhbTAxMUBhdWNrbGFuZHVuaS5hYy5ueg==