 Keith Dadson1†
Keith Dadson1†
- 1 Department of Biology, York University, Toronto, ON, Canada
- 2 Institut Pasteur Korea, Seoul, South Korea
The widespread physiological actions of adiponectin have now been well characterized as clinical studies and works in animal models have established strong correlations between circulating adiponectin level and various disease-related outcomes. Thus, conventional thinking attributes many of adiponectin’s beneficial effects to endocrine actions of adipose-derived adiponectin. However, it is now clear that several tissues can themselves produce adiponectin and there is growing evidence that locally produced adiponectin can mediate functionally important autocrine or paracrine effects. In this review article we discuss regulation of adiponectin production, its mechanism of action via receptor isoforms and signaling pathways, and its principal physiological effects (i.e., metabolic and cardiovascular). The role of endocrine actions of adiponectin and changes in local production of adiponectin or its receptors in whole body physiology is discussed.
Introduction
Adiponectin: Regulation of its Expression and Post-Translational Modification
Adiponectin was discovered as an adipocyte-derived 30 kDa secretory protein, which consists of an amino-terminal signal sequence followed by a collagenous domain and a carboxyl-terminal globular domain (Scherer et al., 1995; Hu et al., 1996; Maeda et al., 1996; Nakano et al., 1996). Adiponectin is transcriptionally regulated by peroxisome proliferator-activated receptor γ (PPARγ), C/EBP, SREBP, E47, and Id3 protein (Fajas et al., 1998; Osborne, 2000; Motoshima et al., 2002; Yilmaz et al., 2004; Doran et al., 2008; Figure 1). Drugs like rosiglitazone and pioglitazone, belonging to the thiazolidinedione (TZD) class of PPARγ agonists, have been clinically and experimentally proven to be potent inducers of adiponectin expression (Tsuchida et al., 2005; Phillips et al., 2008; Liu et al., 2009) and indeed many of the metabolic and cardioprotective effects of rosiglitazone or pioglitazone are absent/decreased in mice lacking adiponectin (Li et al., 2010; Tao et al., 2010; Zhou et al., 2010). Therefore, elevated adiponectin expression is a critical mechanism of action in mediating beneficial effects of this drug class. Regulation of SREBP-1c is another well known mechanism activating adiponectin transcription while more recently Id3 and E47 were demonstrated as novel regulators of this SREBP-1c-mediated adiponectin expression in adipocytes. E47 potentiates SREBP-1c-mediated adiponectin promoter activation and this is inhibited upon interaction with Id3. Decreased Id3 levels increased adiponectin expression and Id3-null mice had increased adiponectin expression in visceral fat tissue and serum (Doran et al., 2008).
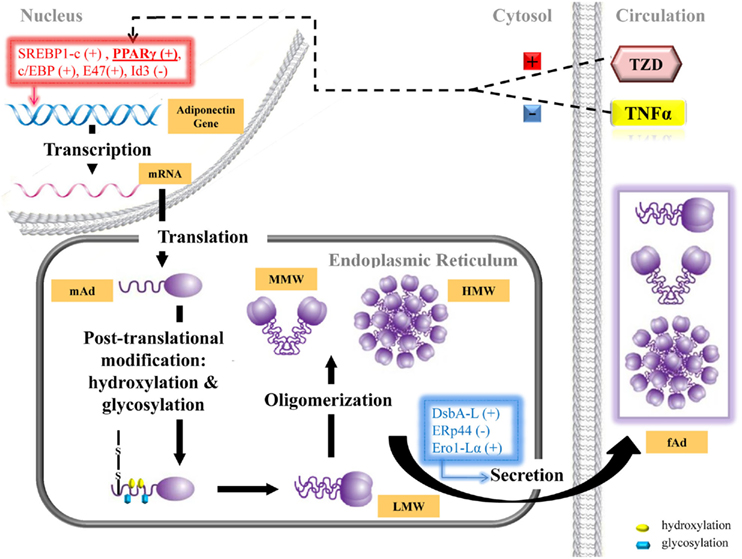
Figure 1. Schematic representation of the steps involved in transcription, translation, post-translational modification, oligomerization, and secretion of adiponectin. Several transcription factors (top left) which mediate adiponectin gene transcription are regulated to increase (thiazolidinedione, TZD) or decrease (tumor necrosis factor-alpha, TNF-α) adiponectin expression. Monomeric adiponectin (mAd) is posttranslationally modified and further oligomerized to form trimers (low molecular weight, LMW), hexamers (medium, MMW) and oligomeric (high, HMW) forms. Various mechanisms (bottom right) mediate this oligomerization and secretion resulting in secretion of HMW, MMW, and LMW forms.
Extensive post-translational modification plays a vital role for assembling adiponectin to form its functional oligomeric complexes (Wang et al., 2008; Simpson and Whitehead, 2010). The initiation step of adiponectin multimerization involves the non-collagenous globular domain forming trimers (Waki et al., 2003). Subsequently, the disulfide bond formed via Cys39 (mouse) or Cys36 (human) is critical for adiponectin to form higher molecular weight multimers based on its trimeric form (Tsao et al., 2003). Post-translational modification including hydroxylation and glycolysation of the four conserved lysine residues (lys68, lys71, lys80, lys104) within the collagenous domain of adiponectin are required for the formation of HMW oligomeric complex (Wang et al., 2002, 2006). The disulfide bond A oxidoreductase-like protein (DsbA-L) was found to positively regulate the process of adiponectin multimerization. The secretion of adiponectin is specifically regulated by endoplasmic reticulum (ER) proteins ERp44 and Ero1-Lα. The covalent bond formed between ERp44 and the thiol group of Cys39 on adiponectin retains adiponectin in ER while the disulfide bond formed between ERp44 and Ero1-Lα releases adiponectin (Anelli et al., 2003; Wang et al., 2007; Schraw et al., 2008). Adiponectin exists abundantly in the plasma and circulates in its HMW (oligomer), MMW (hexamer), and LMW (trimer) oligomeric forms (Waki et al., 2003). The combination of these oligomeric forms is often referred to as full-length adiponectin (fAd). An additional circulating form of adiponectin, the albumin binding LMW, has been identified subsequently (Ebinuma et al., 2006). Moreover, upon protease cleavage the globular domain of fAd (referred to hereafter as gAd) can be librated. Although significant circulating levels are not observed, gAd is proposed to be cleaved locally by specific tissues or at sites of inflammation (Fruebis et al., 2001; Waki et al., 2005).
Adiponectin Receptors and Receptor Adaptor Proteins
Adiponectin exerts many of its cellular effects principally through binding to two receptor isoforms with seven putative transmembrane domains. These adiponectin receptor 1 (AdipoR1) and adiponectin receptor 2 (AdipoR2) isoforms have distinct distribution patterns in various tissues (Yamauchi et al., 2003a, 2007; Kadowaki et al., 2007, 2008). It was shown that skeletal muscle cells bound gAd more avidly than fAd and suppression of AdipoR1 expression with siRNA reduced high-affinity gAd binding (Yamauchi et al., 2003b). Conversely, suppression of AdipoR2 expression with siRNA largely reduced fAd binding, but only modestly reduced globular adiponectin binding. Collectively, available data indicates that AdipoR1 is a high-affinity receptor for gAd and a low-affinity receptor for fAd, whereas AdioR2 is an intermediate-affinity receptor for fAd and gAd. Since AdipoR1 is the predominant form expressed in skeletal muscle, while AdipoR2 is predominantly expressed in liver, this correlated with the fact that gAd exerts its insulin mimetic and insulin-sensitizing effect more effectively compared to fAd in skeletal muscle and vice versa (Yamauchi et al., 2002). T-cadherin, was also found to competitively bind only the hexameric and HMW forms of adiponectin (Hug et al., 2004; Asada et al., 2007; Chan et al., 2008). Although T-cadherin lacks an intracellular domain (Hug et al., 2004), various studies have suggested the involvement of this protein in mediating functional effects of adiponectin. These include cardioprotective effects (Denzel et al., 2010), anti-atherosclerotic effects in vasculature (Takeuchi et al., 2007; Andreeva et al., 2010), anti-diabetic effects in skeletal muscle (Hug et al., 2004) as well as anti-fibrotic effects in liver (Asada et al., 2007).
Several adiponectin receptor binding proteins have now been identified (Buechler et al., 2010; Heiker et al., 2010). The first and best characterized is adaptor protein containing pleckstrin homology domain, phosphotyrosine binding (PTB) domain, and leucine zipper motif (APPL1; Mao et al., 2006a). Other more recently identified adaptor proteins which have been suggested to be involved in adiponectin’s intracellular signal transduction include activated protein kinase C1 (RACK1; Xu et al., 2009), ER protein 46 (ERp46; Charlton et al., 2010), and protein kinase CK2β (Heiker et al., 2009). Among the adaptor proteins, only APPL1 associates with both AdipoR1 and AdipoR2 while RACK1, ERp46, and CK2β bind to AdipoR1. APPL1 interacts with the intracellular region of adiponectin receptors through its PTB domain (Mao et al., 2006a) and sequentially activates downstream signaling. It has now been shown that APPL1 plays an important role in mediating many of adiponectin’s effects, including metabolic effects in liver, muscle, and endothelial cells (Kobayashi et al., 2004; Mao et al., 2006a; Cheng et al., 2007, 2009; Chandrasekar et al., 2008; Wang et al., 2009a; Zhou et al., 2009; Cleasby et al., 2011; Xin et al., 2011) as well as cardioprotective effects (Fang et al., 2010; Park et al., 2011). The interaction between RACK1 or CK2β and AdipoR1 was indicated by yeast two-hybrid studies and confirmed in cells by colocalization and coimmunoprecipitation (Heiker et al., 2009; Xu et al., 2009). RNAi-mediated RACK1 knockdown prevented adiponectin regulated glucose uptake in HepG2 cells (Xu et al., 2009). Pharmacological inhibition of CK2β attenuated adiponectin signaling in skeletal muscle cells (Heiker et al., 2009). Coimmunoprecipitation also confirmed the interaction between ERp46 and AdipoR1, but not AdipoR2, and interestingly the suppression of ERp46 expression resulted in increased cell surface AdipoR1 levels and enhanced adiponectin stimulated phosphorylation of AMP kinase (AMPK) but reduced phosphorylation of p38-mitogen-activated protein kinase (MAPK; Charlton et al., 2010).
Circulating Adiponectin: Regulation and Changes in Normal and Disease States
Adiponectin Isoforms, Their Circadian Rhythmn and Clearance
Adiponectin circulates in the concentration range of ∼3–30 μg/ml in healthy individuals, with a lower level in male compare to female which is mainly attributed to lower amounts of hexameric HMW form (Xu et al., 2005; Wang et al., 2008). Studies on the metabolism and clearance of adiponectin have yielded variable results with a half-life of approximately 75 min reported recently based on tracking fluorescently labeled recombinant adiponectin in the circulation (Halberg et al., 2009) while a previous study (as the study performed in rabbit using recombinant human adiponectin) found HMW adiponectin had a half-life of 13 h and LMW a half-life of 17.5 h (Peake et al., 2005). A recent study indicated the important role of posttranslational modifications, including sialylation which modifies the O-linked glycans on Thr residues with sialic acid, in the regulation of adponectin’s half-life (Richards et al., 2010). Indeed, enzymatic removal of the sialic acid from adiponectin accelerates its clearance from circulation (Richards et al., 2010). Like many other metabolic hormones adiponectin, especially its HMW form, is regulated by the biological clock and shows circadian rhythms with a reduction occurring during the night (Froy, 2007; Cano et al., 2009; Gomez-Abellan et al., 2010; Scheer et al., 2010; Tan et al., 2011). The endocrine effects of adipose-derived adiponectin are conventionally believed to regulate many physiological processes, and many studies have established strong correlations between circulating adiponectin levels and various disease states.
Adiponectin in Obesity and Diabetes
A decreased plasma adiponectin level has been found in patients with obesity and type 2 diabetes despite the increasing mass of adipose tissue (Arita et al., 1999; Ouchi et al., 2001; Weyer et al., 2001; Matsuda et al., 2002; Daimon et al., 2003; Spranger et al., 2003; Ryo et al., 2004; Liu et al., 2007). Moreover, many studies have shown that instead of the absolute total circulating level of adiponectin, the ratio between HMW and total adiponectin can more accurately predict insulin resistance and development of features of the metabolic syndrome (Araki et al., 2006; Hara et al., 2006; Katsuki et al., 2006; Liu et al., 2007; Hamilton et al., 2011). However, although HMW is often referred to as most biologically active for this reason, there remains a lack of direct metabolic studies have been conducted using only the HMW form of adiponectin. Proinflammatory cytokines, in particular tumor necrosis factor α (TNFα), are considered to be a principal cause of the reduction in circulating adiponectin seen in obese/diabetic patients (Ouchi et al., 2003a,b; Takemura et al., 2007). Nevertheless, it is important to balance these strong clinical correlations with consideration of whether alterations in adiponectin are always a cause or, in some cases, a consequence of disease states and this will be highlighted below.
Adiponectin in Cardiovascular Disease
A substantial amount of evidence indicates a potential pathophysiological contribution of adiponectin in cardiovascular disease (Shinmura, 2010; Xu et al., 2010; Hui et al., 2011; Okamoto, 2011). Clinical studies have generally identified negative correlations between plasma adiponectin levels and various aspects of cardiovascular disease such as atherosclerosis, myocardial infarction, heart failure, endothelial dysfunction and hypertension, and established as an independent risk factor for these. The HMW form of adiponectin has been generally regarded as the best predictor of cardiovascular outcome yet, interestingly, a recent study also identifies trimeric LMW adiponectin as a potentially useful biomarker in cardiovascular disease (Hamilton et al., 2011). Adiponectin knockout mice have been particularly informative in terms of elucidating the interaction between adiponectin and cardiovascular injury, with exaggerated degrees of induced cardiovascular defects typically observed in these mice which can be corrected by adiponectin replenishment. Additionally, several single nucleotide polymorphisms (SNPs) have been identified in the adiponectin gene that consequently cause a reduction of its serum concentration (Menzaghi et al., 2002, 2007; Tanko et al., 2005; Yang and Chuang, 2006). For example, clinical data has shown evidence of the correlation between adiponectin gene +45 and +276G SNPs with obesity, type 2 diabetes, insulin resistance, cardiovascular disease, and hypertension (Mousavinasab et al., 2006; Loos et al., 2007; Yang et al., 2007; Dolley et al., 2008). Adiponectin exerts its beneficial cardiovascular influence via targeting many cell types and inducing antioxidative, metabolic, anti-fibrotic, anti-apoptotic, anti-inflammatory, and vasodilator activities (Xu et al., 2010; Hui et al., 2011). The concept of adiponectin resistance, potentially due to changes in expression of AdipoRs and APPL1/2, may be an underestimated factor in the development of CVD (Lau et al., 2011).
Additional Disease Processes Related to Changes in Adiponectin Levels
An extremely wide variety of physiological processes have been found to be regulated by adiponectin and, as a consequence, changes in circulating adiponectin have been implicated in various clinical settings (Kadowaki et al., 2008; Wang and Scherer, 2008; Yamauchi and Kadowaki, 2008; Shetty et al., 2009; Brochu-Gaudreau et al., 2010; Chiarugi and Fiaschi, 2010). Correlations have been established between changes in circulating adiponectin levels and cancer (Kelesidis et al., 2006; Jarde et al., 2011; Paz-Filho et al., 2011), hepatic fibrosis (Dogru et al., 2010), reproductive events (Michalakis and Segars, 2010), bone mass density (BMD; Napoli et al., 2010), and inflammation (Fantuzzi, 2008). Some of these are discussed in more detail below.
Adiponectin Physiology and Evidence for Contribution of Endocrine and Local Effects
Skeletal Muscle
Adiponectin has clearly been shown to regulate glucose and fatty acid metabolism in skeletal muscle, principally via studies which have used animal models with enhanced or suppressed circulating adiponectin or used recombinant forms of the protein to treat cells in vitro or skeletal muscle ex vivo. Cell based in vitro studies show adiponectin can increase both basal and insulin-stimulated glucose uptake by promoting GLUT4 translocation to the cell membrane (Ceddia et al., 2005; Fang et al., 2005, 2009; Mao et al., 2006a) and increase fatty acid uptake and oxidation (Tomas et al., 2002; Yoon et al., 2006) through the activation of AMPK, p38-MAPK, and PPARα pathways (Yamauchi et al., 2002, 2003b; Yoon et al., 2006). Animal model studies in vivo correlate well with these observations as systemic infusion, adenoviral-based delivery or genetic overexpression of adiponectin can successfully correct high-fat diet-induced insulin resistance in skeletal muscle and decrease serum TG and FFA levels (Yamauchi et al., 2001, 2003c; Maeda et al., 2002; Combs et al., 2004). Although it is very well accepted that adiponectin mediates beneficial metabolic effects in skeletal muscle, the precise underlying molecular mechanisms were uncovered in more detail recently. Adiponectin can increase skeletal muscle mitochondrial mass and oxidative capacity, at least in part via inducing extracellular Ca2+ influx and subsequently activating the Ca2+/calmodulin-dependent protein kinase kinase beta (CaMKKβ)–AMPK–Sirt1–peroxisome proliferator-activated receptor gamma coactivator-1alpha (PGC1α) pathway (Iwabu et al., 2010). Previous studies also showed transgenic mice overexpressing adiponectin had improved insulin sensitivity whereas adiponectin knockout mice exhibit some degree of insulin resistance and decreased expression of PGC1α and PPARγ (Civitarese et al., 2006; Kadowaki et al., 2006). Most recently, Scherer’s group identified another important mechanism underlying adiponectins beneficial metabolic effects, namely enhanced ceramide catabolism in skeletal muscle (Holland et al., 2011).
The prevailing assumption was that the metabolic effects of adiponectin in skeletal muscle were due to endocrine effects of adipocyte-derived adiponectin (Figure 2). Importantly, emerging evidence (Delaigle et al., 2004, 2006; Krause et al., 2008; Liu et al., 2009; Amin et al., 2010; Jortay et al., 2010; Van Berendoncks et al., 2010) suggests that adiponectin can also be expressed and secreted by skeletal muscle and thus may also be classified as a myokine (Pedersen and Febbraio, 2008) which exerts its effect locally. We have shown an increased level of adiponectin mRNA, intracellular and secreted protein in response to rosiglitazone treatment in vitro; and subsequently verified that this skeletal muscle produced adiponectin exerted functional metabolic effects including enhanced insulin-stimulated Akt phosphorylation and glucose uptake (Liu et al., 2009). In agreement with this, Leff’s group also demonstrated PPARγ-mediated skeletal muscle adiponectin production which mediated autocrine effects to improve insulin sensitivity and could protect against high-fat diet-induced insulin resistance in vivo (Amin et al., 2010). It has been suggested that skeletal muscle adiponectin content increases in response to certain inflammatory conditions and obesity in an attempt at providing local anti-inflammatory and antioxidative protection (Delaigle et al., 2004, 2006; Jortay et al., 2010). Indeed, the globular form of adiponectin mediates potent metabolic effects in skeletal muscle (Fruebis et al., 2001; Tomas et al., 2002; Ceddia et al., 2005; Chen et al., 2005; Fang et al., 2005; Mao et al., 2006a) and it is conceivable that elevated local amounts of gAd are produced in inflamed tissue by elastase enzyme derived from infiltrating inflammatory cells (Waki et al., 2005). Finally, autocrine effects of gAd have recently been identified in the regulation of skeletal muscle cell differentiation (Fiaschi et al., 2009, 2010).
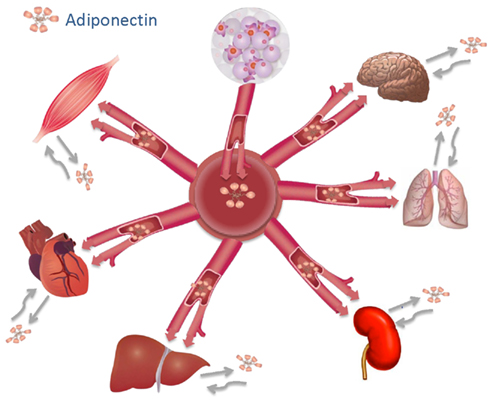
Figure 2. Endocrine and autocrine/paracrine effects of adiponectin. The figure indicates that adiponectin in circulation (blood vessel, center) is derived primarily from adipose tissue (top). Circulating adiponectin can travel to numerous tissues and mediate endocrine effects. In addition, several tissues can also produce adiponectin (solid gray arrow) which can then act locally (twisted gray arrow) to mediate functional autocrine or paracrine effect.
Local effects of adiponectin may also be determined by changes in expression of its receptor isoforms and signaling intermediates, although relatively little is known on this topic to date. Weight loss induced by either exercise or diet together with exercise enhance the expression of adiponectin receptor mRNA in skeletal muscle of humans and animal models (Vu et al., 2007; Christiansen et al., 2010). A study in non-diabetic Mexican Americans with or without a family history of Type 2 diabetes concluded that skeletal muscle expression levels of both AdipoR1 and AdipoR2 correlated positively with insulin sensitivity (Civitarese et al., 2004). Hyperglycemia and hyperinsulinemia can both alter AdipoR expression in muscle cells and consequently adiponectin sensitivity (Fang et al., 2005). Although it is generally believed that enhancing AdipoR–APPL1 signaling is beneficial (Mao et al., 2006a; Cheng et al., 2007, 2009; Saito et al., 2007; Zhou et al., 2009; Fang et al., 2010; Cleasby et al., 2011), recent studies have identified that the abundance of APPL1 mRNA is significantly higher in muscle of type 2 diabetic individuals (Holmes et al., 2011). Bariatric surgery corrected hyperglycemia and this was correlated with increased circulating adiponectin and skeletal muscle AdipoR1 expression with reduced APPL1 content (Holmes et al., 2011).
Cardiovascular
As described above, many studies have established correlations between circulating adiponectin levels and various cardiovascular outcomes and the underlying mechanisms are now well understood. For example, adiponectin is now established as a cardioprotective adipokine as it mediates beneficial effects on cardiac remodeling events such as energy metabolism, hypertrophy, fibrosis, and apoptosis (Shibata et al., 2004, 2005, 2007; Liao et al., 2005; Palanivel et al., 2007; Fang et al., 2010; Li et al., 2010; Tao et al., 2010; Shimano et al., 2011). Anti-inflammatory, vasodilator, and anti-atherosclerotic effects confer further beneficial influences of adiponectin on the cardiovascular system (Fantuzzi, 2008; Brochu-Gaudreau et al., 2010; Xu et al., 2010; Hui et al., 2011).
Mice lacking adiponectin have been particularly informative in establishing the cardioprotective role of adiponectin, with numerous studies in these mice demonstrating an exaggerated response of the heart to cardiac stress (Shibata et al., 2004, 2005, 2007; Liao et al., 2005; Li et al., 2010; Tao et al., 2010; Shimano et al., 2011), which was attenuated upon restoration of circulating adiponectin. The ability of adiponectin to counteract deterioration in cardiac function was mediated by metabolic, anti-apoptotic, anti-fibrotic, and anti-hypertrophic effects (Shibata et al., 2004, 2005, 2007; Tao et al., 2007; Wang et al., 2009b; Fang et al., 2011; Park et al., 2011). Adiponectin has been shown to regulate fatty acid β-oxidation in the heart. In cell based in vitro studies of isolated cardiomyocytes, adiponectin was shown to stimulate the phosphorylation of AMPK, IRS1, and Akt (T308 and S473) correlating with the regulation of glucose and fatty acid uptake and metabolism (Palanivel et al., 2007), and to target cofilin to mediate remodeling of the actin cytoskeleton leading to the translocation of lipoprotein lipase (LPL) to the cell surface (Ganguly et al., 2011). Adiponectin was also shown to stimulate the phosphorylation of acetyl coenzyme A carboxylase (ACC), as well as to induce CPT-1 expression and activation through AMPK (Li et al., 2007a). Recently (Fang et al., 2010), we demonstrated that adiponectin increases fatty acid uptake, CD36 translocation, and insulin-stimulated glucose transport as well as Akt phosphorylation in isolated adult cardiomyocytes, and enhances fatty acid oxidation in conjunction with AMPK and ACC phosphorylation in the isolated working heart. However, despite an increase in fatty acid oxidation and myocardial oxygen consumption, adiponectin increased hydraulic work, and maintained cardiac efficiency (Fang et al., 2010).
The phosphorylation of AMPK was shown to attenuate norepinepherine induced cardiomyocyte hypertrophy and ERK phosphorylation (Shibata et al., 2004), angiotensin II induced NF-κB activation and hypertrophy (Wang et al., 2011), and also shown to fully (Shibata et al., 2005), or minimally (Wang et al., 2009a) attenuate hypoxia–reoxygenation induced apoptosis. Adiponectin was shown to attenuate hypoxia–reoxygenation induced apoptosis in H9C2 cells through the AdipoR1/APPL1 signaling pathway (Park et al., 2011). Cardiac fibrosis is associated with impaired cardiac function, and there are numerous studies demonstrating the exaggerated fibrotic response of the heart to cardiac stress in the absence of adiponectin (Shibata et al., 2004, 2005, 2007; Liao et al., 2005; Li et al., 2010; Tao et al., 2010; Shimano et al., 2011). Very few studies have directly investigated regulation of extracellular matrix components by adiponectin in vitro. Cardiac fibroblasts express AdipoR1 (Huang et al., 2009) and treatment of adult rat fibroblasts with gAd was shown to increase IL-6 expression and secretion via the activation of AMPK, p38-MAPK, and ERK1/2 (Fan et al., 2011).
Although an overwhelming amount of data indicates numerous beneficial effects of adiponectin, there is also some contradictory evidence from clinical and experimental studies on the cardioprotective role of adiponectin. For example, recent clinical data have positively correlated high levels of adiponectin with mortality and severity in patients with congestive heart failure (Shinmura, 2010). Adiponectin knockout mice subjected to long-term pressure overload suggested that under chronic stress, adiponectin deficiency preserves oxidative capacity and cardiac function despite an increase in cardiac hypertrophy compared to wild-type mice, suggesting that adiponectin may in fact be playing a permissive role in long-term cardiac dysfunction (O’Shea et al., 2010).
Both cardiomyocytes (Pineiro et al., 2005; Ding et al., 2007) and epicardial adipose tissue (Gormez et al., 2011; Hirata et al., 2011) can produce adiponectin and thus increase autocrine/paracrine bioavailability. Indeed, epicardial adipose-derived adiponectin was recently identified as a predictor of positive outcome following cardiac surgery (Kourliouros et al., 2011), although it should be noted that expression of adiponectin from epicardial adipose tissue has been shown to be lower than that from subcutaneous adipose tissue (Bambace et al., 2011). Similarly, cardiomyocytes produce relatively small amounts of adiponectin (Pineiro et al., 2005; Ding et al., 2007), yet these are likely to be sufficient for locally mediated effects. A number of studies have found that cardiac adiponectin levels are altered in various cardiomyopathies. For example, a recent study has shown in patients with dilated cardiomyopathy that adiponectin expression was decreased sixfold and this was mirrored in immunohistochemical analysis of endomyocardial biopsies (Skurk et al., 2008). Additionally, the accumulation of adiponectin within the myocardial tissue following stress through leakage from the vascular compartment could also serve to increase the local supply of bioavailable adiponectin and serve to compensate for the inflammatory induced downregulation of both local and systemic adiponectin expression (Ouchi et al., 2000; Shibata et al., 2007; Fujita et al., 2008). Interestingly, adiponectin has also been shown to accumulate in atherosclerotic plaques and whether this is causative or protective against progression of atherosclerosis is still incompletely resolved, although the latter seems most likely (Li et al., 2007b; Cai et al., 2010; Reynolds et al., 2010).
Myocardial adiponectin resistance (Saito et al., 2007; Kollias et al., 2011; Ma et al., 2011) may necessarily be the first target in developing adiponectin-based therapeutics in the treatment of cardiovascular disease. Overall, circulating or local adiponectin levels tend to correlate negatively with cardiovascular disease incidence and prognosis, however since many cardiovascular events are progressive in nature there may in some cases be a temporal compensatory increase in adiponectin expression, particularly within the affected tissue. Thus, several paradoxical observations have been reported in the literature and it is likely that the timing of targeting adiponectin therapeutically will be vital to its success in the cardiovascular arena (Dembinski, 2010).
Liver
Regulation of hepatic glucose and fatty acid metabolism plays an important role in the ability of adiponectin to improve whole body energy homeostasis (Kadowaki and Yamauchi, 2005; Fang and Sweeney, 2006; Kadowaki et al., 2007). For example, low levels or defects in adiponectin action correlate with steatosis, hepatomegaly, and local inflammation associated with various liver diseases. The intracellular signaling mechanisms via which adiponectin mediates effects in hepatocytes are similar to those in muscle (Wang et al., 2009c), with one apparent exception being a more important role of the AdipoR2 isoform in mediating the effects of adiponectin in liver (Yamauchi et al., 2003b, 2007; Yamauchi and Kadowaki, 2008). Adiponectin is known to exert its effects in the liver primarily through the activation of the AMPK and PPARα pathways. In addition to the well characterized insulin-sensitizing and insulin-like effects of adiponectin in the liver, low serum adiponectin levels have been associated with high TNFα levels and the presence of non-alcoholic fatty liver disease (NAFLD) independent of insulin resistance (Hui et al., 2004; Aygun et al., 2006; Polyzos et al., 2011). In particular, adiponectin has been shown to mediate anti-fibrotic effects through the activation of AMPK in hepatic stellate cells (Adachi and Brenner, 2008), anti-apoptotic effects via PI3K and AMPK activation in hepatocytes (Jung et al., 2009), and to be anti-inflammatory through the inhibition of TNFα induced hepatotoxicity (Sennello et al., 2005). A role for T-cadherin has also been proposed in mediating the effects of adiponectin on liver fibrosis (Asada et al., 2007). Furthermore, circulating adiponectin levels have been found to be downregulated in morbidly obese patients with non-alcoholic steatohepatitis (NASH) compared to individuals with simple steatosis (Uribe et al., 2008; Ma et al., 2009), while a paradoxical increase in serum adiponectin levels were detected in patients with cirrhosis, independent of insulin resistance (Tietge et al., 2004; Kaser et al., 2005). It is worth bearing in mind that increased adiponectin levels in liver cirrhosis may reflect reduced hepatic clearance of adiponectin and/or a compensatory increase toward the overwhelming production of proinflammatory cytokines in cirrhosis. Thus, a potentially detrimental contribution of adiponectin as NAFLD progresses to cirrhosis must be considered (Polyzos et al., 2010).
Hepatic AdipoR1 and AdipoR2 mRNA expression levels were higher in insulin-resistant subjects, perhaps reflecting a compensatory mechanism for reduced plasma adiponectin (Felder et al., 2010). Liver fibrosis in individuals infected with hepatitis C virus was associated with hyperadiponectinemia and, interestingly, reduced AdipoR1 expression (Corbetta et al., 2011). A study in humans showed no change in AdipoR expression in patients with NASH (Uribe et al., 2008), but an important role for changes in AdipoR expression was also shown in a study using a high-fat and high-cholesterol diet in obese fa/fa Zucker rats to induce NASH was associated with decreased AdipoR1 and AdipoR2 expression (Matsunami et al., 2011). In addition, liver expression and localization of adiponectin were increased in wild-type mice in response to carbon tetrachloride induced hepatofibrosis (Yoda-Murakami et al., 2001).
Lung
Respiratory complications are often observed in obese individuals (Ford, 2005; Shore, 2010). Importantly, decreased serum adiponectin levels correlate with poor lung function in asthma and chronic obstructive pulmonary disease (COPD, independent of adiposity (Sood et al., 2008; Stanciu et al., 2009; Sutherland et al., 2009; Chan et al., 2010; Thyagarajan et al., 2010). Indeed, clinical treatment of COPD with corticosteroids and antibiotics improved lung function concomitant with elevated circulating adiponectin levels and a decrease in systemic inflammatory markers such as IL-6 and TNFα (Krommidas et al., 2010). Note, it is possible that the increased adiponectin levels may result from diminished IL-6 and TNFα after treatment. Adiponectin knockout mice exhibited progressive alveolar enlargement and endothelial cell apoptosis, which was attenuated by adenoviral administration of adiponectin (Nakanishi et al., 2011). Continuous infusion of adiponectin via subcutaneously implanted osmotic pumps to replenish decreased levels was found to attenuate ovalbumin-induced airway inflammation in mice through the attenuation of inflammatory cell influx, corresponding with a reduction in IL-13 and IL-5 (Shore et al., 2006). Furthermore, chronic allergic airway inflammation and pulmonary vascular remodeling are also exacerbated in adiponectin deficient mice (Medoff et al., 2009; Nakagawa et al., 2009; Summer et al., 2009). Nevertheless, a double-blind randomized clinical trial found that asthmatic patients exhibited only a modestly beneficial effect in the late asthmatic response to inhaled allergen challenge after 28 days of rosiglitazone treatment to increase serum adiponectin levels (Richards et al., 2010).
Although the studies described above focused on endocrine effects of adiponectin, it is again important to consider the potential effects of locally produced adiponectin in the lung. Adiponectin was found to be localized to the murine pulmonary vascular endothelium under normal (Summer et al., 2009) or hypoxic (Nakagawa et al., 2009) conditions and bronchoalveolar fluid contained low levels of adiponectin (Summer et al., 2008). Adiponectin was overexpressed in the bronchoalveolar lavage (BAL) fluid of COPD patients and in a multimeric distribution profile differing from that found in serum (Zhu et al., 2010). Interestingly, these findings correlated with the increased localization of AdipoR1 to the airway epithelial cells of COPD patients (Miller et al., 2009).
Others
Numerous effects of adiponectin have been established in other peripheral tissues (Kadowaki et al., 2008), besides the obvious autocrine effects on adipocytes themselves (Wu et al., 2003). For example, the longstanding complication of nephropathy in obesity and diabetes has naturally led to studies on the pathophysiological role of adiponectin in this process (Chen et al., 2004; Srivastava, 2006; Stenvinkel, 2011). Increased circulating adiponectin levels are found in predialysis patients with end stage renal disease (ESRD; Shen et al., 2007; Stenvinkel, 2011) and adiponectin suggested to be a predictive factor for the progression of chronic kidney disease in men (Kollerits et al., 2007). General consensus based on available literature indicates that adiponectin is renoprotective (Abe et al., 2010), for example via attenuating pathological progression toward renal fibrosis and glomerular hypertrophy (Ohashi et al., 2007). Even in the absence of a stressor, adiponectin deficient mice exhibited segmentally fused podocyte processes, increased albumin leakage into the urine (albuminuria), and kidney oxidant stress when compared to wild-type controls, while treatment with adiponectin reduced the degree of albumin permeability of a podocyte monolayer in vitro (Sharma et al., 2008).
Obesity is strongly associated with increased BMD due not only to the increased mechanical load, but also to adipocyte-derived hormonal factors mediating the cross-talk between adipocytes and bone (Confavreux et al., 2009). As such, there is accumulating and contradictory evidence indicating that adiponectin plays a role in bone maintenance and metabolism (Lenchik et al., 2003; Bozic et al., 2010; Barbour et al., 2011). Specifically, in vitro, adiponectin has been found to decrease osteoclast differentiation and bone-resorption activity (Oshima et al., 2005) via APPL1-mediated Akt1 suppression (Tu et al., 2011), while increasing osteoblast proliferation and differentiation via an AdipoR1 dependent p38-MAPK/JNK signaling pathway (Luo et al., 2005), suggesting that adiponectin positively influences bone growth. However, additional studies in adiponectin deficient or hyperadiponectinemic examining bone mass and fragility yielded some paradoxical observations (Oshima et al., 2005; Williams et al., 2009; Mitsui et al., 2011). Indeed, another study reported no abnormality in bone mass or turnover in adiponectin knockout mice or adiponectin transgenic mice overexpressing globular adiponectin (Shinoda et al., 2006). Interestingly, adiponectin has been found to be expressed by bone forming osteoblasts (Berner et al., 2004; Shinoda et al., 2006) indicating a potential complex autocrine, paracrine, and endocrine role of adiponectin in mediating bone density.
Adiponectin also has centrally mediated effects, such as regulation of food intake and energy expenditure (Qi et al., 2004; Kadowaki et al., 2008). Although adiponectin was reportedly unable to cross the blood brain barrier (Spranger et al., 2006), it was found in the cerebrospinal fluid of rats (Qi et al., 2004) and humans (Neumeier et al., 2007) although at significantly lower levels and with different oligomeric profile than that in peripheral circulation (Ebinuma et al., 2007). Adiponectin mRNA expression and localization within the CNS has now been shown (Rodriguez-Pacheco et al., 2007; Psilopanagioti et al., 2009). Several studies have documented the functionality of adiponectin (Rodriguez-Pacheco et al., 2007) in vitro, and through intracerebral injection (Hoyda et al., 2009a; Iwama et al., 2009; Park et al., 2011), and also central expression of adiponectin receptors (Hoyda et al., 2009b).
Adiponectin Action as a Therapeutic Target
The rationale for targeting adiponectin is based on the well documented beneficial physiological actions of adiponectin spanning diabetes, inflammation, cardiovascular diseases, and cancer and it is expected that studies in animal models will translate well to human physiology in the case of adiponectin (Mao et al., 2006b; Zhu et al., 2008; Shetty et al., 2009; Wang et al., 2009c; Marette and Sweeney, 2011). Adiponectin-based therapeutics would have potentially wide-ranging applications in markets with widespread demographics. In diabetes alone there are still significant unmet therapeutic needs despite an annual global market value of around $30 billion. Synthesis and administration of recombinant forms of adiponectin is generally not a viable therapeutic approach due to the cost of synthesizing correctly posttranslationally modified bioactive forms and the disadvantage of the route of administration, although the recombinant globular domain of adiponectin is in pre-clinical trials for Merck and Protemix have a highly glycosylated form of adiponectin in pre-clinical trials. There are several reports of compounds which increase adiponectin expression and secretion, although these increases tend to be very modest at non-supra-physiological doses (Zhu et al., 2008). The commonly prescribed TZD class of anti-diabetic agents act at least in part via elevating adiponectin (Pajvani et al., 2004; Kubota et al., 2006; Li et al., 2010). A more attractive therapeutic option would be the discovery of small molecule compounds which mimic or enhance adiponectin action. Presently, a small molecule adiponectin-mimetic developed by Rigel Pharmaceuticals that improves insulin sensitivity in a diabetic mouse model is in pre-clinical development and Kadowaki’s group have also identified such a compound. However there are no potent and specific adiponectin-based therapeutics which are clinically available yet. Based on the information in this review article it is interesting to consider the future possibility of combining adiponectin-based therapeutics with tissue- or cell-specific delivery approaches.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Related work in the authors laboratory has been supported by Canadian Diabetes Association, Heart & Stroke Foundation of Canada, and Canadian Institutes of Health Research. Unfortunately it was not possible to quote all relevant literature and we apologize for any omissions of pertinent research papers.
References
Abe, Y., Eto, S., Matsumae, T., Ogahara, S., Murata, T., Watanabe, M., Nakashima, H., and Saito, T. (2010). The proportion and metabolic effects of adiponectin multimeric isoforms in patients with chronic kidney disease on maintenance hemodialysis. Ren. Fail. 32, 849–854.
Adachi, M., and Brenner, D. A. (2008). High molecular weight adiponectin inhibits proliferation of hepatic stellate cells via activation of adenosine monophosphate-activated protein kinase. Hepatology 47, 677–685.
Amin, R. H., Mathews, S. T., Camp, H. S., Ding, L., and Leff, T. (2010). Selective activation of PPARgamma in skeletal muscle induces endogenous production of adiponectin and protects mice from diet-induced insulin resistance. Am. J. Physiol. Endocrinol. Metab. 298, E28–E37.
Andreeva, A. V., Han, J., Kutuzov, M. A., Profirovic, J., Tkachuk, V. A., and Voyno-Yasenetskaya, T. A. (2010). T-cadherin modulates endothelial barrier function. J. Cell Physiol. 223, 94–102.
Anelli, T., Alessio, M., Bachi, A., Bergamelli, L., Bertoli, G., Camerini, S., Mezghrani, A., Ruffato, E., Simmen, T., and Sitia, R. (2003). Thiol-mediated protein retention in the endoplasmic reticulum: the role of ERp44. EMBO J. 22, 5015–5022.
Araki, S., Dobashi, K., Kubo, K., Asayama, K., and Shirahata, A. (2006). High molecular weight, rather than total, adiponectin levels better reflect metabolic abnormalities associated with childhood obesity. J. Clin. Endocrinol. Metab. 91, 5113–5116.
Arita, Y., Kihara, S., Ouchi, N., Takahashi, M., Maeda, K., Miyagawa, J., Hotta, K., Shimomura, I., Nakamura, T., Miyaoka, K., Kuriyama, H., Nishida, M., Yamashita, S., Okubo, K., Matsubara, K., Muraguchi, M., Ohmoto, Y., Funahashi, T., and Matsuzawa, Y. (1999). Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem. Biophys. Res. Commun. 257, 79–83.
Asada, K., Yoshiji, H., Noguchi, R., Ikenaka, Y., Kitade, M., Kaji, K., Yoshii, J., Yanase, K., Namisaki, T., Yamazaki, M., Tsujimoto, T., Akahane, T., Uemura, M., and Fukui, H. (2007). Crosstalk between high-molecular-weight adiponectin and T-cadherin during liver fibrosis development in rats. Int. J. Mol. Med. 20, 725–729.
Aygun, C., Senturk, O., Hulagu, S., Uraz, S., Celebi, A., Konduk, T., Mutlu, B., and Canturk, Z. (2006). Serum levels of hepatoprotective peptide adiponectin in non-alcoholic fatty liver disease. Eur. J. Gastroenterol. Hepatol. 18, 175–180.
Bambace, C., Telesca, M., Zoico, E., Sepe, A., Olioso, D., Rossi, A., Corzato, F., Di Francesco, V., Mazzucco, A., Santini, F., and Zamboni, M. (2011). Adiponectin gene expression and adipocyte diameter: a comparison between epicardial and subcutaneous adipose tissue in men. Cardiovasc. Pathol. 20, e153–e156.
Barbour, K. E., Zmuda, J. M., Boudreau, R., Strotmeyer, E. S., Horwitz, M. J., Evans, R. W., Kanaya, A. M., Harris, T. B., Bauer, D. C., and Cauley, J. A. (2011). Adipokines and the risk of fracture in older adults. J. Bone Miner. Res. 26, 1568–1576.
Berner, H. S., Lyngstadaas, S. P., Spahr, A., Monjo, M., Thommesen, L., Drevon, C. A., Syversen, U., and Reseland, J. E. (2004). Adiponectin and its receptors are expressed in bone-forming cells. Bone 35, 842–849.
Bozic, B., Loncar, G., Prodanovic, N., Radojicic, Z., Cvorovic, V., Dimkovic, S., and Popovic-Brkic, V. (2010). Relationship between high circulating adiponectin with bone mineral density and bone metabolism in elderly males with chronic heart failure. J. Card. Fail. 16, 301–307.
Brochu-Gaudreau, K., Rehfeldt, C., Blouin, R., Bordignon, V., Murphy, B. D., and Palin, M. F. (2010). Adiponectin action from head to toe. Endocrine 37, 11–32.
Buechler, C., Wanninger, J., and Neumeier, M. (2010). Adiponectin receptor binding proteins – recent advances in elucidating adiponectin signalling pathways. FEBS Lett. 584, 4280–4286.
Cai, X. J., Chen, L., Li, L., Feng, M., Li, X., Zhang, K., Rong, Y. Y., Hu, X. B., Zhang, M. X., Zhang, Y., and Zhang, M. (2010). Adiponectin inhibits lipopolysaccharide-induced adventitial fibroblast migration and transition to myofibroblasts via AdipoR1-AMPK-iNOS pathway. Mol. Endocrinol. 24, 218–228.
Cano, P., Cardinali, D. P., Rios-Lugo, M. J., Fernandez-Mateos, M. P., Reyes Toso, C. F., and Esquifino, A. I. (2009). Effect of a high-fat diet on 24-hour pattern of circulating adipocytokines in rats. Obesity (Silver Spring) 17, 1866–1871.
Ceddia, R. B., Somwar, R., Maida, A., Fang, X., Bikopoulos, G., and Sweeney, G. (2005). Globular adiponectin increases GLUT4 translocation and glucose uptake but reduces glycogen synthesis in rat skeletal muscle cells. Diabetologia 48, 132–139.
Chan, D. W., Lee, J. M., Chan, P. C., and Ng, I. O. (2008). Genetic and epigenetic inactivation of T-cadherin in human hepatocellular carcinoma cells. Int. J. Cancer 123, 1043–1052.
Chan, K. H., Yeung, S. C., Yao, T. J., Ip, M. S., Cheung, A. H., Chan-Yeung, M. M., and Mak, J. C. (2010). Elevated plasma adiponectin levels in patients with chronic obstructive pulmonary disease. Int. J. Tuberc. Lung Dis. 14, 1193–1200.
Chandrasekar, B., Boylston, W. H., Venkatachalam, K., Webster, N. J., Prabhu, S. D., and Valente, A. J. (2008). Adiponectin blocks interleukin-18-mediated endothelial cell death via APPL1-dependent AMP-activated protein kinase (AMPK) activation and IKK/NF-kappaB/PTEN suppression. J. Biol. Chem. 283, 24889–24898.
Charlton, H. K., Webster, J., Kruger, S., Simpson, F., Richards, A. A., and Whitehead, J. P. (2010). ERp46 binds to AdipoR1, but not AdipoR2, and modulates adiponectin signalling. Biochem. Biophys. Res. Commun. 392, 234–239.
Chen, J., Muntner, P., Hamm, L. L., Jones, D. W., Batuman, V., Fonseca, V., Whelton, P. K., and He, J. (2004). The metabolic syndrome and chronic kidney disease in U.S. adults. Ann. Intern. Med. 140, 167–174.
Chen, M. B., McAinch, A. J., Macaulay, S. L., Castelli, L. A., O’Brien, P. E., Dixon, J. B., Cameron-Smith, D., Kemp, B. E., and Steinberg, G. R. (2005). Impaired activation of AMP-kinase and fatty acid oxidation by globular adiponectin in cultured human skeletal muscle of obese type 2 diabetics. J. Clin. Endocrinol. Metab. 90, 3665–3672.
Cheng, K. K., Iglesias, M. A., Lam, K. S., Wang, Y., Sweeney, G., Zhu, W., Vanhoutte, P. M., Kraegen, E. W., and Xu, A. (2009). APPL1 potentiates insulin-mediated inhibition of hepatic glucose production and alleviates diabetes via Akt activation in mice. Cell Metab. 9, 417–427.
Cheng, K. K., Lam, K. S., Wang, Y., Huang, Y., Carling, D., Wu, D., Wong, C., and Xu, A. (2007). Adiponectin-induced endothelial nitric oxide synthase activation and nitric oxide production are mediated by APPL1 in endothelial cells. Diabetes 56, 1387–1394.
Chiarugi, P., and Fiaschi, T. (2010). Adiponectin in health and diseases: from metabolic syndrome to tissue regeneration. Expert Opin. Ther. Targets 14, 193–206.
Christiansen, T., Paulsen, S. K., Bruun, J. M., Ploug, T., Pedersen, S. B., and Richelsen, B. (2010). Diet-induced weight loss and exercise alone and in combination enhance the expression of adiponectin receptors in adipose tissue and skeletal muscle, but only diet-induced weight loss enhanced circulating adiponectin. J. Clin. Endocrinol. Metab. 95, 911–919.
Civitarese, A. E., Jenkinson, C. P., Richardson, D., Bajaj, M., Cusi, K., Kashyap, S., Berria, R., Belfort, R., DeFronzo, R. A., Mandarino, L. J., and Ravussin, E. (2004). Adiponectin receptors gene expression and insulin sensitivity in non-diabetic Mexican Americans with or without a family history of Type 2 diabetes. Diabetologia 47, 816–820.
Civitarese, A. E., Ukropcova, B., Carling, S., Hulver, M., DeFronzo, R. A., Mandarino, L., Ravussin, E., and Smith, S. R. (2006). Role of adiponectin in human skeletal muscle bioenergetics. Cell Metab. 4, 75–87.
Cleasby, M. E., Lau, Q., Polkinghorne, E., Patel, S. A., Leslie, S. J., Turner, N., Cooney, G., Xu, A., and Kraegen, E. (2011). The adaptor protein APPL1 increases glycogen accumulation in rat skeletal muscle through activation of the PI3-kinase signalling pathway. J. Endocrinol. 210, 81–92.
Combs, T. P., Pajvani, U. B., Berg, A. H., Lin, Y., Jelicks, L. A., Laplante, M., Nawrocki, A. R., Rajala, M. W., Parlow, A. F., Cheeseboro, L., Ding, Y. Y., Russell, R. G., Lindemann, D., Hartley, A., Baker, G. R., Obici, S., Deshaies, Y., Ludgate, M., Rossetti, L., and Scherer, P. E. (2004). A transgenic mouse with a deletion in the collagenous domain of adiponectin displays elevated circulating adiponectin and improved insulin sensitivity. Endocrinology 145, 367–383.
Confavreux, C. B., Levine, R. L., and Karsenty, G. (2009). A paradigm of integrative physiology, the crosstalk between bone and energy metabolisms. Mol. Cell. Endocrinol. 310, 21–29.
Corbetta, S., Redaelli, A., Pozzi, M., Bovo, G., Ratti, L., Redaelli, E., Pellegrini, C., Beck-Peccoz, P., and Spada, A. (2011). Fibrosis is associated with adiponectin resistance in chronic hepatitis C virus infection. Eur. J. Clin. Invest. 41, 898–905.
Daimon, M., Oizumi, T., Saitoh, T., Kameda, W., Hirata, A., Yamaguchi, H., Ohnuma, H., Igarashi, M., Tominaga, M., and Kato, T. (2003). Decreased serum levels of adiponectin are a risk factor for the progression to type 2 diabetes in the Japanese Population: the Funagata study. Diabetes Care 26, 2015–2020.
Delaigle, A. M., Jonas, J. C., Bauche, I. B., Cornu, O., and Brichard, S. M. (2004). Induction of adiponectin in skeletal muscle by inflammatory cytokines: in vivo and in vitro studies. Endocrinology 145, 5589–5597.
Delaigle, A. M., Senou, M., Guiot, Y., Many, M. C., and Brichard, S. M. (2006). Induction of adiponectin in skeletal muscle of type 2 diabetic mice: In vivo and in vitro studies. Diabetologia 49, 1311–1323.
Dembinski, R. (2010). Adiponectin in critically ill patients: more questions than answers? Crit. Care Med. 38, 2415–2416.
Denzel, M. S., Scimia, M. C., Zumstein, P. M., Walsh, K., Ruiz-Lozano, P., and Ranscht, B. (2010). T-cadherin is critical for adiponectin-mediated cardioprotection in mice. J. Clin. Invest. 120, 4342–4352.
Ding, G., Qin, Q., He, N., Francis-David, S. C., Hou, J., Liu, J., Ricks, E., and Yang, Q. (2007). Adiponectin and its receptors are expressed in adult ventricular cardiomyocytes and upregulated by activation of peroxisome proliferator-activated receptor gamma. J. Mol. Cell. Cardiol. 43, 73–84.
Dogru, T., Karadurmus, N., Kilciler, G., Tapan, S., and Ercin, C. N. (2010). The relationship of plasma adiponectin with liver fibrosis in morbid obesity. Obes. Surg. 20, 1462; author reply 1463–1464.
Dolley, G., Bertrais, S., Frochot, V., Bebel, J. F., Guerre-Millo, M., Tores, F., Rousseau, F., Hager, J., Basdevant, A., Hercberg, S., Galan, P., Oppert, J. M., Lacorte, J. M., and Clement, K. (2008). Promoter adiponectin polymorphisms and waist/hip ratio variation in a prospective French adults study. Int. J. Obes. (Lond.) 32, 669–675.
Doran, A. C., Meller, N., Cutchins, A., Deliri, H., Slayton, R. P., Oldham, S. N., Kim, J. B., Keller, S. R., and McNamara, C. A. (2008). The helix-loop-helix factors Id3 and E47 are novel regulators of adiponectin. Circ. Res. 103, 624–634.
Ebinuma, H., Miida, T., Yamauchi, T., Hada, Y., Hara, K., Kubota, N., and Kadowaki, T. (2007). Improved ELISA for selective measurement of adiponectin multimers and identification of adiponectin in human cerebrospinal fluid. Clin. Chem. 53, 1541–1544.
Ebinuma, H., Miyazaki, O., Yago, H., Hara, K., Yamauchi, T., and Kadowaki, T. (2006). A novel ELISA system for selective measurement of human adiponectin multimers by using proteases. Clin. Chim. Acta 372, 47–53.
Fajas, L., Fruchart, J. C., and Auwerx, J. (1998). Transcriptional control of adipogenesis. Curr. Opin. Cell Biol. 10, 165–173.
Fan, D., Li, L., Wang, C., Cui, X. B., Zhou, Y., and Wu, L. L. (2011). Adiponectin induces interleukin-6 production and its underlying mechanism in adult rat cardiac fibroblasts. J. Cell. Physiol. 226, 1793–1802.
Fang, X., Fetros, J., Dadson, K. E., Xu, A., and Sweeney, G. (2009). Leptin prevents the metabolic effects of adiponectin in L6 myotubes. Diabetologia 52, 2190–2200.
Fang, X., Palanivel, R., Cresser, J., Schram, K., Ganguly, R., Thong, F. S., Tuinei, J., Xu, A., Abel, E. D., and Sweeney, G. (2010). An APPL1-AMPK signaling axis mediates beneficial metabolic effects of adiponectin in the heart. Am. J. Physiol. Endocrinol. Metab. 299, E721–E759.
Fang, X., Palanivel, R., Cresser, J., Schram, K., Ganguly, R., Thong, F. S., Tuinei, J., Xu, A., Abel, E. D., and Sweeney, G. (2011). An APPL1-AMPK signaling axis mediates beneficial metabolic effects of adiponectin in the heart. Am. J. Physiol. Endocrinol. Metab. 299, E721–E729.
Fang, X., Palanivel, R., Zhou, X., Liu, Y., Xu, A., Wang, Y., and Sweeney, G. (2005). Hyperglycemia- and hyperinsulinemia-induced alteration of adiponectin receptor expression and adiponectin effects in L6 myoblasts. J. Mol. Endocrinol. 35, 465–476.
Fang, X., and Sweeney, G. (2006). Mechanisms regulating energy metabolism by adiponectin in obesity and diabetes. Biochem. Soc. Trans. 34, 798–801.
Fantuzzi, G. (2008). Adiponectin and inflammation: consensus and controversy. J. Allergy Clin. Immunol. 121, 326–330.
Felder, T. K., Hahne, P., Soyal, S. M., Miller, K., Hoffinger, H., Oberkofler, H., Krempler, F., and Patsch, W. (2010). Hepatic adiponectin receptors (ADIPOR) 1 and 2 mRNA and their relation to insulin resistance in obese humans. Int. J. Obes. (Lond.) 34, 846–851.
Fiaschi, T., Cirelli, D., Comito, G., Gelmini, S., Ramponi, G., Serio, M., and Chiarugi, P. (2009). Globular adiponectin induces differentiation and fusion of skeletal muscle cells. Cell Res. 19, 584–597.
Fiaschi, T., Tedesco, F. S., Giannoni, E., Diaz-Manera, J., Parri, M., Cossu, G., and Chiarugi, P. (2010). Globular adiponectin as a complete mesoangioblast regulator: role in proliferation, survival, motility, and skeletal muscle differentiation. Mol. Biol. Cell 21, 848–859.
Ford, E. S. (2005). The epidemiology of obesity and asthma. J. Allergy Clin. Immunol. 115, 897–909; quiz 910.
Froy, O. (2007). The relationship between nutrition and circadian rhythms in mammals. Front. Neuroendocrinol. 28, 61–71.
Fruebis, J., Tsao, T. S., Javorschi, S., Ebbets-Reed, D., Erickson, M. R., Yen, F. T., Bihain, B. E., and Lodish, H. F. (2001). Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc. Natl. Acad. Sci. U.S.A. 98, 2005–2010.
Fujita, K., Maeda, N., Sonoda, M., Ohashi, K., Hibuse, T., Nishizawa, H., Nishida, M., Hiuge, A., Kurata, A., Kihara, S., Shimomura, I., and Funahashi, T. (2008). Adiponectin protects against angiotensin II-induced cardiac fibrosis through activation of PPAR-alpha. Arterioscler. Thromb. Vasc. Biol. 28, 863–870.
Ganguly, R., Schram, K., Fang, X., Kim, M., Rodrigues, B., Thong, F. S., and Sweeney, G. (2011). Adiponectin increases LPL activity via RhoA/ROCK-mediated actin remodelling in adult rat cardiomyocytes. Endocrinology 152, 247–254.
Gomez-Abellan, P., Gomez-Santos, C., Madrid, J. A., Milagro, F. I., Campion, J., Martinez, J. A., Ordovas, J. M., and Garaulet, M. (2010). Circadian expression of adiponectin and its receptors in human adipose tissue. Endocrinology 151, 115–122.
Gormez, S., Demirkan, A., Atalar, F., Caynak, B., Erdim, R., Sozer, V., Gunay, D., Akpinar, B., Ozbek, U., and Buyukdevrim, A. S. (2011). Adipose tissue gene expression of adiponectin, tumor necrosis factor-alpha and leptin in metabolic syndrome patients with coronary artery disease. Intern. Med. 50, 805–810.
Halberg, N., Schraw, T. D., Wang, Z. V., Kim, J. Y., Yi, J., Hamilton, M. P., Luby-Phelps, K., and Scherer, P. E. (2009). Systemic fate of the adipocyte-derived factor adiponectin. Diabetes 58, 1961–1970.
Hamilton, M. P., Gore, M. O., Ayers, C. R., Wu, X., McGuire, D. K., and Scherer, P. (2011). Adiponectin and cardiovascular risk profile in patients with type 2 diabetes mellitus: parameters associated with adiponectin complex distribution. Diab. Vasc. Dis. Res. 8, 190–194.
Hara, K., Horikoshi, M., Yamauchi, T., Yago, H., Miyazaki, O., Ebinuma, H., Imai, Y., Nagai, R., and Kadowaki, T. (2006). Measurement of the high-molecular weight form of adiponectin in plasma is useful for the prediction of insulin resistance and metabolic syndrome. Diabetes Care 29, 1357–1362.
Heiker, J. T., Kosel, D., and Beck-Sickinger, A. G. (2010). Molecular mechanisms of signal transduction via adiponectin and adiponectin receptors. Biol. Chem. 391, 1005–1018.
Heiker, J. T., Wottawah, C. M., Juhl, C., Kosel, D., Morl, K., and Beck-Sickinger, A. G. (2009). Protein kinase CK2 interacts with adiponectin receptor 1 and participates in adiponectin signaling. Cell. Signal. 21, 936–942.
Hirata, Y., Kurobe, H., Akaike, M., Chikugo, F., Hori, T., Bando, Y., Nishio, C., Higashida, M., Nakaya, Y., Kitagawa, T., and Sata, M. (2011). Enhanced inflammation in epicardial fat in patients with coronary artery disease. Int. Heart J. 52, 139–142.
Holland, W. L., Miller, R. A., Wang, Z. V., Sun, K., Barth, B. M., Bui, H. H., Davis, K. E., Bikman, B. T., Halberg, N., Rutkowski, J. M., Wade, M. R., Tenorio, V. M., Kuo, M. S., Brozinick, J. T., Zhang, B. B., Birnbaum, M. J., Summers, S. A., and Scherer, P. E. (2011). Receptor-mediated activation of ceramidase activity initiates the pleiotropic actions of adiponectin. Nat. Med. 17, 55–63.
Holmes, R. M., Yi, Z., De Filippis, E., Berria, R., Shahani, S., Sathyanarayana, P., Sherman, V., Fujiwara, K., Meyer, C., Christ-Roberts, C., Hwang, H., Finlayson, J., Dong, L. Q., Mandarino, L. J., and Bajaj, M. (2011). Increased abundance of the adaptor protein containing pleckstrin homology domain, phosphotyrosine binding domain and leucine zipper motif (APPL1) in patients with obesity and type 2 diabetes: evidence for altered adiponectin signalling. Diabetologia. 54, 2122–2131.
Hoyda, T. D., Samson, W. K., and Ferguson, A. V. (2009a). Adiponectin depolarizes parvocellular paraventricular nucleus neurons controlling neuroendocrine and autonomic function. Endocrinology 150, 832–840.
Hoyda, T. D., Smith, P. M., and Ferguson, A. V. (2009b). Adiponectin acts in the nucleus of the solitary tract to decrease blood pressure by modulating the excitability of neuropeptide Y neurons. Brain Res. 1256, 76–84.
Hu, E., Liang, P., and Spiegelman, B. M. (1996). AdipoQ is a novel adipose-specific gene dysregulated in obesity. J. Biol. Chem. 271, 10697–10703.
Huang, D., Yang, C., Wang, Y., Liao, Y., and Huang, K. (2009). PARP-1 suppresses adiponectin expression through poly(ADP-ribosyl)ation of PPAR gamma in cardiac fibroblasts. Cardiovasc. Res. 81, 98–107.
Hug, C., Wang, J., Ahmad, N. S., Bogan, J. S., Tsao, T. S., and Lodish, H. F. (2004). T-cadherin is a receptor for hexameric and high-molecular-weight forms of Acrp30/adiponectin. Proc. Natl. Acad. Sci. U.S.A. 101, 10308–10313.
Hui, J. M., Hodge, A., Farrell, G. C., Kench, J. G., Kriketos, A., and George, J. (2004). Beyond insulin resistance in NASH: TNF-alpha or adiponectin? Hepatology 40, 46–54.
Hui, X., Lam, K. S., Vanhoutte, P. M., and Xu, A. (2011). Adiponectin and cardiovascular health: an update. Br. J. Pharmacol. doi: 10.1111/j.1476-5381.2011.01395.x. [Epub ahead of print].
Iwabu, M., Yamauchi, T., Okada-Iwabu, M., Sato, K., Nakagawa, T., Funata, M., Yamaguchi, M., Namiki, S., Nakayama, R., Tabata, M., Ogata, H., Kubota, N., Takamoto, I., Hayashi, Y. K., Yamauchi, N., Waki, H., Fukayama, M., Nishino, I., Tokuyama, K., Ueki, K., Oike, Y., Ishii, S., Hirose, K., Shimizu, T., Touhara, K., and Kadowaki, T. (2010). Adiponectin and AdipoR1 regulate PGC-1alpha and mitochondria by Ca(2+) and AMPK/SIRT1. Nature.
Iwama, S., Sugimura, Y., Murase, T., Hiroi, M., Goto, M., Hayashi, M., Arima, H., and Oiso, Y. (2009). Central adiponectin functions to inhibit arginine vasopressin release in conscious rats. J. Neuroendocrinol. 21,
Jarde, T., Perrier, S., Vasson, M. P., and Caldefie-Chezet, F. (2011). Molecular mechanisms of leptin and adiponectin in breast cancer. Eur. J. Cancer 47, 33–43.
Jortay, J., Senou, M., Delaigle, A., Noel, L., Funahashi, T., Maeda, N., Many, M. C., and Brichard, S. M. (2010). Local induction of adiponectin reduces lipopolysaccharide-triggered skeletal muscle damage. Endocrinology 151, 4840–4851.
Jung, T. W., Lee, Y. J., Lee, M. W., and Kim, S. M. (2009). Full-length adiponectin protects hepatocytes from palmitate-induced apoptosis via inhibition of c-Jun NH2 terminal kinase. FEBS J. 276, 2278–2284.
Kadowaki, T., and Yamauchi, T. (2005). Adiponectin and adiponectin receptors. Endocr. Rev. 26, 439–451.
Kadowaki, T., Yamauchi, T., and Kubota, N. (2008). The physiological and pathophysiological role of adiponectin and adiponectin receptors in the peripheral tissues and CNS. FEBS Lett. 582, 74–80.
Kadowaki, T., Yamauchi, T., Kubota, N., Hara, K., and Ueki, K. (2007). Adiponectin and adiponectin receptors in obesity-linked insulin resistance. Novartis Found. Symp. 286, 164–176; discussion 176–182, 200–203.
Kadowaki, T., Yamauchi, T., Kubota, N., Hara, K., Ueki, K., and Tobe, K. (2006). Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J. Clin. Invest. 116, 1784–1792.
Kaser, S., Moschen, A., Cayon, A., Kaser, A., Crespo, J., Pons-Romero, F., Ebenbichler, C. F., Patsch, J. R., and Tilg, H. (2005). Adiponectin and its receptors in non-alcoholic steatohepatitis. Gut 54, 117–121.
Katsuki, A., Suematsu, M., Gabazza, E. C., Murashima, S., Nakatani, K., Togashi, K., Yano, Y., and Sumida, Y. (2006). Decreased high-molecular weight adiponectin-to-total adiponectin ratio in sera is associated with insulin resistance in Japanese metabolically obese, normal-weight men with normal glucose tolerance. Diabetes Care 29, 2327–2328.
Kelesidis, I., Kelesidis, T., and Mantzoros, C. S. (2006). Adiponectin and cancer: a systematic review. Br. J. Cancer 94, 1221–1225.
Kobayashi, H., Ouchi, N., Kihara, S., Walsh, K., Kumada, M., Abe, Y., Funahashi, T., and Matsuzawa, Y. (2004). Selective suppression of endothelial cell apoptosis by the high molecular weight form of adiponectin. Circ. Res. 94, e27–e31.
Kollerits, B., Fliser, D., Heid, I. M., Ritz, E., and Kronenberg, F. (2007). Gender-specific association of adiponectin as a predictor of progression of chronic kidney disease: the Mild to Moderate Kidney Disease Study. Kidney Int. 71, 1279–1286.
Kollias, A., Tsiotra, P. C., Ikonomidis, I., Maratou, E., Mitrou, P., Kyriazi, E., Boutati, E., Lekakis, J., Economopoulos, T., Kremastinos, D. T., Dimitriadis, G., and Raptis, S. A. (2011). Adiponectin levels and expression of adiponectin receptors in isolated monocytes from overweight patients with coronary artery disease. Cardiovasc. Diabetol. 10, 14.
Kourliouros, A., Karastergiou, K., Nowell, J., Gukop, P., Tavakkoli Hosseini, M., Valencia, O., Mohamed Ali, V., and Jahangiri, M. (2011). Protective effect of epicardial adiponectin on atrial fibrillation following cardiac surgery. Eur. J. Cardiothorac. Surg. 39, 228–232.
Krause, M. P., Liu, Y., Vu, V., Chan, L., Xu, A., Riddell, M. C., Sweeney, G., and Hawke, T. J. (2008). Adiponectin is expressed by skeletal muscle fibers and influences muscle phenotype and function. Am. J. Physiol. Cell Physiol. 295, C203–C212.
Krommidas, G., Kostikas, K., Papatheodorou, G., Koutsokera, A., Gourgoulianis, K. I., Roussos, C., Koulouris, N. G., and Loukides, S. (2010). Plasma leptin and adiponectin in COPD exacerbations: associations with inflammatory biomarkers. Respir. Med. 104, 40–46.
Kubota, N., Terauchi, Y., Kubota, T., Kumagai, H., Itoh, S., Satoh, H., Yano, W., Ogata, H., Tokuyama, K., Takamoto, I., Mineyama, T., Ishikawa, M., Moroi, M., Sugi, K., Yamauchi, T., Ueki, K., Tobe, K., Noda, T., Nagai, R., and Kadowaki, T. (2006). Pioglitazone ameliorates insulin resistance and diabetes by both adiponectin-dependent and -independent pathways. J. Biol. Chem. 281, 8748–8755.
Lau, W. B., Tao, L., Wang, Y., Li, R., and Ma, X. L. (2011). Systemic adiponectin malfunction as a risk factor for cardiovascular disease. Antioxid. Redox Signal. 15, 1863–1873.
Lenchik, L., Register, T. C., Hsu, F. C., Lohman, K., Nicklas, B. J., Freedman, B. I., Langefeld, C. D., Carr, J. J., and Bowden, D. W. (2003). Adiponectin as a novel determinant of bone mineral density and visceral fat. Bone 33, 646–651.
Li, L., Wu, L., Wang, C., Liu, L., and Zhao, Y. (2007a). Adiponectin modulates carnitine palmitoyltransferase-1 through AMPK signaling cascade in rat cardiomyocytes. Regul. Pept. 139, 72–79.
Li, C. J., Sun, H. W., Zhu, F. L., Chen, L., Rong, Y. Y., Zhang, Y., and Zhang, M. (2007b). Local adiponectin treatment reduces atherosclerotic plaque size in rabbits. J. Endocrinol. 193, 137–145.
Li, P., Shibata, R., Unno, K., Shimano, M., Furukawa, M., Ohashi, T., Cheng, X., Nagata, K., Ouchi, N., and Murohara, T. (2010). Evidence for the importance of adiponectin in the cardioprotective effects of pioglitazone. Hypertension 55, 69–75.
Liao, Y., Takashima, S., Maeda, N., Ouchi, N., Komamura, K., Shimomura, I., Hori, M., Matsuzawa, Y., Funahashi, T., and Kitakaze, M. (2005). Exacerbation of heart failure in adiponectin-deficient mice due to impaired regulation of AMPK and glucose metabolism. Cardiovasc. Res. 67, 705–713.
Liu, Y., Chewchuk, S., Lavigne, C., Brule, S., Pilon, G., Houde, V., Xu, A., Marette, A., and Sweeney, G. (2009). Functional significance of skeletal muscle adiponectin production, changes in animal models of obesity and diabetes, and regulation by rosiglitazone treatment. Am. J. Physiol. Endocrinol. Metab. 297, E657–E664.
Liu, Y., Retnakaran, R., Hanley, A., Tungtrongchitr, R., Shaw, C., and Sweeney, G. (2007). Total and high molecular weight but not trimeric or hexameric forms of adiponectin correlate with markers of the metabolic syndrome and liver injury in Thai subjects. J. Clin. Endocrinol. Metab. 92, 4313–4318.
Loos, R. J., Ruchat, S., Rankinen, T., Tremblay, A., Perusse, L., and Bouchard, C. (2007). Adiponectin and adiponectin receptor gene variants in relation to resting metabolic rate, respiratory quotient, and adiposity-related phenotypes in the Quebec Family Study. Am. J. Clin. Nutr. 85, 26–34.
Luo, X. H., Guo, L. J., Yuan, L. Q., Xie, H., Zhou, H. D., Wu, X. P., and Liao, E. Y. (2005). Adiponectin stimulates human osteoblasts proliferation and differentiation via the MAPK signaling pathway. Exp. Cell Res. 309, 99–109.
Ma, H., Gomez, V., Lu, L., Yang, X., Wu, X., and Xiao, S. Y. (2009). Expression of adiponectin and its receptors in livers of morbidly obese patients with non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 24, 233–237.
Ma, Y., Liu, Y., Liu, S., Qu, Y., Wang, R., Xia, C., Pei, H., Lian, K., Yin, T., Lu, X., Sun, L., Yang, L., Cao, Y., Lau, W. B., Gao, E., Wang, H., and Tao, L. (2011). Dynamic alteration of adiponectin/adiponectin receptor expression and its impact on myocardial ischemia/reperfusion in type 1 diabetic mice. Am. J. Physiol. Endocrinol. Metab. 301, E447–E455.
Maeda, K., Okubo, K., Shimomura, I., Funahashi, T., Matsuzawa, Y., and Matsubara, K. (1996). cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem. Biophys. Res. Commun. 221, 286–289.
Maeda, N., Shimomura, I., Kishida, K., Nishizawa, H., Matsuda, M., Nagaretani, H., Furuyama, N., Kondo, H., Takahashi, M., Arita, Y., Komuro, R., Ouchi, N., Kihara, S., Tochino, Y., Okutomi, K., Horie, M., Takeda, S., Aoyama, T., Funahashi, T., and Matsuzawa, Y. (2002). Diet-induced insulin resistance in mice lacking adiponectin/ACRP30. Nat. Med. 8, 731–737.
Mao, X., Kikani, C. K., Riojas, R. A., Langlais, P., Wang, L., Ramos, F. J., Fang, Q., Christ-Roberts, C. Y., Hong, J. Y., Kim, R. Y., Liu, F., and Dong, L. Q. (2006a). APPL1 binds to adiponectin receptors and mediates adiponectin signalling and function. Nat. Cell Biol. 8, 516–523.
Mao, X., Hong, J. Y., and Dong, L. Q. (2006b). The adiponectin signaling pathway as a novel pharmacological target. Mini Rev. Med. Chem. 6, 1331–1340.
Marette, A., and Sweeney, G. (2011). Cardiovascular complications of diabetes: recent insights in pathophysiology and therapeutics. Expert Rev. Endocrinol. Metab. 6, 689–696.
Matsuda, M., Shimomura, I., Sata, M., Arita, Y., Nishida, M., Maeda, N., Kumada, M., Okamoto, Y., Nagaretani, H., Nishizawa, H., Kishida, K., Komuro, R., Ouchi, N., Kihara, S., Nagai, R., Funahashi, T., and Matsuzawa, Y. (2002). Role of adiponectin in preventing vascular stenosis. The missing link of adipo-vascular axis. J. Biol. Chem. 277, 37487–37491.
Matsunami, T., Sato, Y., Ariga, S., Sato, T., Shimomura, T., Kashimura, H., Hasegawa, Y., and Yukawa, M. (2011). Regulation of synthesis and oxidation of fatty acids by adiponectin receptors (AdipoR1/R2) and insulin receptor substrate isoforms (IRS-1/-2) of the liver in a nonalcoholic steatohepatitis animal model. Metab. Clin. Exp. 60, 805–814.
Medoff, B. D., Okamoto, Y., Leyton, P., Weng, M., Sandall, B. P., Raher, M. J., Kihara, S., Bloch, K. D., Libby, P., and Luster, A. D. (2009). Adiponectin deficiency increases allergic airway inflammation and pulmonary vascular remodeling. Am. J. Respir. Cell Mol. Biol. 41, 397–406.
Menzaghi, C., Ercolino, T., Di Paola, R., Berg, A. H., Warram, J. H., Scherer, P. E., Trischitta, V., and Doria, A. (2002). A haplotype at the adiponectin locus is associated with obesity and other features of the insulin resistance syndrome. Diabetes 51, 2306–2312.
Menzaghi, C., Trischitta, V., and Doria, A. (2007). Genetic influences of adiponectin on insulin resistance, type 2 diabetes, and cardiovascular disease. Diabetes 56, 1198–1209.
Michalakis, K. G., and Segars, J. H. (2010). The role of adiponectin in reproduction: from polycystic ovary syndrome to assisted reproduction. Fertil. Steril. 94, 1949–1957.
Miller, M., Cho, J. Y., Pham, A., Ramsdell, J., and Broide, D. H. (2009). Adiponectin and functional adiponectin receptor 1 are expressed by airway epithelial cells in chronic obstructive pulmonary disease. J. Immunol. 182, 684–691.
Mitsui, Y., Gotoh, M., Fukushima, N., Shirachi, I., Otabe, S., Yuan, X., Hashinaga, T., Wada, N., Mitsui, A., Yoshida, T., Yoshida, S., Yamada, K., and Nagata, K. (2011). Hyperadiponectinemia enhances bone formation in mice. BMC Musculoskelet. Disord. 12, 18. doi:10.1186/1471-2474-12-18
Motoshima, H., Wu, X., Sinha, M. K., Hardy, V. E., Rosato, E. L., Barbot, D. J., Rosato, F. E., and Goldstein, B. J. (2002). Differential regulation of adiponectin secretion from cultured human omental and subcutaneous adipocytes: effects of insulin and rosiglitazone. J. Clin. Endocrinol. Metab. 87, 5662–5667.
Mousavinasab, F., Tahtinen, T., Jokelainen, J., Koskela, P., Vanhala, M., Oikarinen, J., Keinanen-Kiukaanniemi, S., and Laakso, M. (2006). Common polymorphisms (single-nucleotide polymorphisms SNP+45 and SNP+276) of the adiponectin gene regulate serum adiponectin concentrations and blood pressure in young Finnish men. Mol. Genet. Metab. 87, 147–151.
Nakagawa, Y., Kishida, K., Kihara, S., Funahashi, T., and Shimomura, I. (2009). Adiponectin ameliorates hypoxia-induced pulmonary arterial remodeling. Biochem. Biophys. Res. Commun. 382, 183–188.
Nakanishi, K., Takeda, Y., Tetsumoto, S., Iwasaki, T., Tsujino, K., Kuhara, H., Jin, Y., Nagatomo, I., Kida, H., Goya, S., Kijima, T., Maeda, N., Funahashi, T., Shimomura, I., Tachibana, I., and Kawase, I. (2011). Involvement of endothelial apoptosis underlying COPD-like phenotype in adiponectin-null mice: implications for therapy. Am. J. Respir. Crit. Care Med. 183, 1164–1175.
Nakano, Y., Tobe, T., Choi-Miura, N. H., Mazda, T., and Tomita, M. (1996). Isolation and characterization of GBP28, a novel gelatin-binding protein purified from human plasma. J. Biochem. 120, 803–812.
Napoli, N., Pedone, C., Pozzilli, P., Lauretani, F., Ferrucci, L., and Incalzi, R. A. (2010). Adiponectin and bone mass density: the InCHIANTI study. Bone 47, 1001–1005.
Neumeier, M., Weigert, J., Buettner, R., Wanninger, J., Schaffler, A., Muller, A. M., Killian, S., Sauerbruch, S., Schlachetzki, F., Steinbrecher, A., Aslanidis, C., Scholmerich, J., and Buechler, C. (2007). Detection of adiponectin in cerebrospinal fluid in humans. Am. J. Physiol. Endocrinol. Metab. 293, E965–E969.
Ohashi, K., Iwatani, H., Kihara, S., Nakagawa, Y., Komura, N., Fujita, K., Maeda, N., Nishida, M., Katsube, F., Shimomura, I., Ito, T., and Funahashi, T. (2007). Exacerbation of albuminuria and renal fibrosis in subtotal renal ablation model of adiponectin-knockout mice. Arterioscler. Thromb. Vasc. Biol. 27, 1910–1917.
Okamoto, Y. (2011). Adiponectin provides cardiovascular protection in metabolic syndrome. Cardiol. Res. Pract. 2011, 313179.
Osborne, T. F. (2000). Sterol regulatory element-binding proteins (SREBPs): key regulators of nutritional homeostasis and insulin action. J. Biol. Chem. 275, 32379–32382.
O’Shea, K. M., Chess, D. J., Khairallah, R. J., Rastogi, S., Hecker, P. A., Sabbah, H. N., Walsh, K., and Stanley, W. C. (2010). Effects of adiponectin deficiency on structural and metabolic remodeling in mice subjected to pressure overload. Am. J. Physiol. Heart Circ. Physiol. 298, H1639–H1645.
Oshima, K., Nampei, A., Matsuda, M., Iwaki, M., Fukuhara, A., Hashimoto, J., Yoshikawa, H., and Shimomura, I. (2005). Adiponectin increases bone mass by suppressing osteoclast and activating osteoblast. Biochem. Biophys. Res. Commun. 331, 520–526.
Ouchi, N., Kihara, S., Arita, Y., Nishida, M., Matsuyama, A., Okamoto, Y., Ishigami, M., Kuriyama, H., Kishida, K., Nishizawa, H., Hotta, K., Muraguchi, M., Ohmoto, Y., Yamashita, S., Funahashi, T., and Matsuzawa, Y. (2001). Adipocyte-derived plasma protein, adiponectin, suppresses lipid accumulation and class A scavenger receptor expression in human monocyte-derived macrophages. Circulation 103, 1057–1063.
Ouchi, N., Kihara, S., Arita, Y., Okamoto, Y., Maeda, K., Kuriyama, H., Hotta, K., Nishida, M., Takahashi, M., Muraguchi, M., Ohmoto, Y., Nakamura, T., Yamashita, S., Funahashi, T., and Matsuzawa, Y. (2000). Adiponectin, an adipocyte-derived plasma protein, inhibits endothelial NF-kappaB signaling through a cAMP-dependent pathway. Circulation 102, 1296–1301.
Ouchi, N., Kihara, S., Funahashi, T., Matsuzawa, Y., and Walsh, K. (2003a). Obesity, adiponectin and vascular inflammatory disease. Curr. Opin. Lipidol. 14, 561–566.
Ouchi, N., Kihara, S., Funahashi, T., Nakamura, T., Nishida, M., Kumada, M., Okamoto, Y., Ohashi, K., Nagaretani, H., Kishida, K., Nishizawa, H., Maeda, N., Kobayashi, H., Hiraoka, H., and Matsuzawa, Y. (2003b). Reciprocal association of C-reactive protein with adiponectin in blood stream and adipose tissue. Circulation 107, 671–674.
Pajvani, U. B., Hawkins, M., Combs, T. P., Rajala, M. W., Doebber, T., Berger, J. P., Wagner, J. A., Wu, M., Knopps, A., Xiang, A. H., Utzschneider, K. M., Kahn, S. E., Olefsky, J. M., Buchanan, T. A., and Scherer, P. E. (2004). Complex distribution, not absolute amount of adiponectin, correlates with thiazolidinedione-mediated improvement in insulin sensitivity. J. Biol. Chem. 279, 12152–12162.
Palanivel, R., Fang, X., Park, M., Eguchi, M., Pallan, S., De Girolamo, S., Liu, Y., Wang, Y., Xu, A., and Sweeney, G. (2007). Globular and full-length forms of adiponectin mediate specific changes in glucose and fatty acid uptake and metabolism in cardiomyocytes. Cardiovasc. Res. 75, 148–157.
Park, M., Youn, B., Zheng, X. L., Wu, D., Xu, A., and Sweeney, G. (2011). Globular adiponectin, acting via AdipoR1/APPL1, protects H9c2 cells from hypoxia/reoxygenation-induced apoptosis. PLoS ONE 6, e19143. doi:10.1371/journal.pone.0019143
Paz-Filho, G., Lim, E. L., Wong, M. L., and Licinio, J. (2011). Associations between adipokines and obesity-related cancer. Front. Biosci. 16, 1634–1650.
Peake, P. W., Kriketos, A. D., Campbell, L. V., Shen, Y., and Charlesworth, J. A. (2005). The metabolism of isoforms of human adiponectin: studies in human subjects and in experimental animals. Eur. J. Endocrinol. 153, 409–417.
Pedersen, B. K., and Febbraio, M. A. (2008). Muscle as an endocrine organ: focus on muscle-derived interleukin-6. Physiol. Rev. 88, 1379–1406.
Phillips, S. A., Ciaraldi, T. P., Oh, D. K., Savu, M. K., and Henry, R. R. (2008). Adiponectin secretion and response to pioglitazone is depot dependent in cultured human adipose tissue. Am. J. Physiol. Endocrinol. Metab. 295, E842–E850.
Pineiro, R., Iglesias, M. J., Gallego, R., Raghay, K., Eiras, S., Rubio, J., Dieguez, C., Gualillo, O., Gonzalez-Juanatey, J. R., and Lago, F. (2005). Adiponectin is synthesized and secreted by human and murine cardiomyocytes. FEBS Lett. 579, 5163–5169.
Polyzos, S. A., Kountouras, J., Zavos, C., and Stergiopoulos, C. (2010). Adipocytokines in insulin resistance and non-alcoholic fatty liver disease: the two sides of the same coin. Med. Hypotheses 74, 1089–1090.
Polyzos, S. A., Toulis, K. A., Goulis, D. G., Zavos, C., and Kountouras, J. (2011). Serum total adiponectin in nonalcoholic fatty liver disease: a systematic review and meta-analysis. Metabolism 60, 313–326.
Psilopanagioti, A., Papadaki, H., Kranioti, E. F., Alexandrides, T. K., and Varakis, J. N. (2009). Expression of adiponectin and adiponectin receptors in human pituitary gland and brain. Neuroendocrinology 89, 38–47.
Qi, Y., Takahashi, N., Hileman, S. M., Patel, H. R., Berg, A. H., Pajvani, U. B., Scherer, P. E., and Ahima, R. S. (2004). Adiponectin acts in the brain to decrease body weight. Nat. Med. 10, 524–529.
Reynolds, H. R., Buyon, J., Kim, M., Rivera, T. L., Izmirly, P., Tunick, P., and Clancy, R. M. (2010). Association of plasma soluble E-selectin and adiponectin with carotid plaque in patients with systemic lupus erythematosus. Atherosclerosis 210, 569–574.
Richards, D. B., Bareille, P., Lindo, E. L., Quinn, D., and Farrow, S. N. (2010). Treatment with a peroxisomal proliferator activated receptor gamma agonist has a modest effect in the allergen challenge model in asthma: a randomised controlled trial. Respir. Med. 104, 668–674.
Rodriguez-Pacheco, F., Martinez-Fuentes, A. J., Tovar, S., Pinilla, L., Tena-Sempere, M., Dieguez, C., Castano, J. P., and Malagon, M. M. (2007). Regulation of pituitary cell function by adiponectin. Endocrinology 148, 401–410.
Ryo, M., Nakamura, T., Kihara, S., Kumada, M., Shibazaki, S., Takahashi, M., Nagai, M., Matsuzawa, Y., and Funahashi, T. (2004). Adiponectin as a biomarker of the metabolic syndrome. Circ. J. 68, 975–981.
Saito, T., Jones, C. C., Huang, S., Czech, M. P., and Pilch, P. F. (2007). The interaction of Akt with APPL1 is required for insulin-stimulated Glut4 translocation. J. Biol. Chem. 282, 32280–32287.
Scheer, F. A., Chan, J. L., Fargnoli, J., Chamberland, J., Arampatzi, K., Shea, S. A., Blackburn, G. L., and Mantzoros, C. S. (2010). Day/night variations of high-molecular-weight adiponectin and lipocalin-2 in healthy men studied under fed and fasted conditions. Diabetologia 53, 2401–2405.
Scherer, P. E., Williams, S., Fogliano, M., Baldini, G., and Lodish, H. F. (1995). A novel serum protein similar to C1q, produced exclusively in adipocytes. J. Biol. Chem. 270, 26746–26749.
Schraw, T., Wang, Z. V., Halberg, N., Hawkins, M., and Scherer, P. E. (2008). Plasma adiponectin complexes have distinct biochemical characteristics. Endocrinology 149, 2270–2282.
Sennello, J. A., Fayad, R., Morris, A. M., Eckel, R. H., Asilmaz, E., Montez, J., Friedman, J. M., Dinarello, C. A., and Fantuzzi, G. (2005). Regulation of T cell-mediated hepatic inflammation by adiponectin and leptin. Endocrinology 146, 2157–2164.
Sharma, K., Ramachandrarao, S., Qiu, G., Usui, H. K., Zhu, Y., Dunn, S. R., Ouedraogo, R., Hough, K., McCue, P., Chan, L., Falkner, B., and Goldstein, B. J. (2008). Adiponectin regulates albuminuria and podocyte function in mice. J. Clin. Invest. 118, 1645–1656.
Shen, Y. Y., Charlesworth, J. A., Kelly, J. J., Loi, K. W., and Peake, P. W. (2007). Up-regulation of adiponectin, its isoforms and receptors in end-stage kidney disease. Nephrol. Dial. Transplant. 22, 171–178.
Shetty, S., Kusminski, C. M., and Scherer, P. E. (2009). Adiponectin in health and disease: evaluation of adiponectin-targeted drug development strategies. Trends Pharmacol. Sci. 30, 234–239.
Shibata, R., Izumiya, Y., Sato, K., Papanicolaou, K., Kihara, S., Colucci, W. S., Sam, F., Ouchi, N., and Walsh, K. (2007). Adiponectin protects against the development of systolic dysfunction following myocardial infarction. J. Mol. Cell. Cardiol. 42, 1065–1074.
Shibata, R., Ouchi, N., Ito, M., Kihara, S., Shiojima, I., Pimentel, D. R., Kumada, M., Sato, K., Schiekofer, S., Ohashi, K., Funahashi, T., Colucci, W. S., and Walsh, K. (2004). Adiponectin-mediated modulation of hypertrophic signals in the heart. Nat. Med. 10, 1384–1389.
Shibata, R., Sato, K., Pimentel, D. R., Takemura, Y., Kihara, S., Ohashi, K., Funahashi, T., Ouchi, N., and Walsh, K. (2005). Adiponectin protects against myocardial ischemia-reperfusion injury through AMPK- and COX-2-dependent mechanisms. Nat. Med. 11, 1096–1103.
Shimano, M., Ouchi, N., Shibata, R., Ohashi, K., Pimentel, D. R., Murohara, T., and Walsh, K. (2011). Adiponectin deficiency exacerbates cardiac dysfunction following pressure overload through disruption of an AMPK-dependent angiogenic response. J. Mol. Cell. Cardiol. 49, 210–220.
Shinmura, K. (2010). Is adiponectin a bystander or a mediator in heart failure? The tangled thread of a good-natured adipokine in aging and cardiovascular disease. Heart Fail. Rev. 15, 457–466.
Shinoda, Y., Yamaguchi, M., Ogata, N., Akune, T., Kubota, N., Yamauchi, T., Terauchi, Y., Kadowaki, T., Takeuchi, Y., Fukumoto, S., Ikeda, T., Hoshi, K., Chung, U. I., Nakamura, K., and Kawaguchi, H. (2006). Regulation of bone formation by adiponectin through autocrine/paracrine and endocrine pathways. J. Cell. Biochem. 99, 196–208.
Shore, S. A. (2010). Obesity, airway hyperresponsiveness, and inflammation. J. Appl. Physiol. 108, 735–743.
Shore, S. A., Terry, R. D., Flynt, L., Xu, A., and Hug, C. (2006). Adiponectin attenuates allergen-induced airway inflammation and hyperresponsiveness in mice. J. Allergy Clin. Immunol. 118, 389–395.
Simpson, F., and Whitehead, J. P. (2010). Adiponectin – it’s all about the modifications. Int. J. Biochem. Cell Biol. 42, 785–788.
Skurk, C., Wittchen, F., Suckau, L., Witt, H., Noutsias, M., Fechner, H., Schultheiss, H. P., and Poller, W. (2008). Description of a local cardiac adiponectin system and its deregulation in dilated cardiomyopathy. Eur. Heart J. 29, 1168–1180.
Sood, A., Cui, X., Qualls, C., Beckett, W. S., Gross, M. D., Steffes, M. W., Smith, L. J., and Jacobs, D. R. Jr. (2008). Association between asthma and serum adiponectin concentration in women. Thorax 63, 877–882.
Spranger, J., Kroke, A., Mohlig, M., Bergmann, M. M., Ristow, M., Boeing, H., and Pfeiffer, A. F. (2003). Adiponectin and protection against type 2 diabetes mellitus. Lancet 361, 226–228.
Spranger, J., Verma, S., Gohring, I., Bobbert, T., Seifert, J., Sindler, A. L., Pfeiffer, A., Hileman, S. M., Tschop, M., and Banks, W. A. (2006). Adiponectin does not cross the blood-brain barrier but modifies cytokine expression of brain endothelial cells. Diabetes 55, 141–147.
Srivastava, T. (2006). Nondiabetic consequences of obesity on kidney. Pediatr. Nephrol. 21, 463–470.
Stanciu, S., Marinescu, R., Iordache, M., Dumitrescu, S., Muresan, M., and Bogdan, M. A. (2009). Are systemic inflammatory profiles different in patients with COPD and metabolic syndrome as compared to those with COPD alone? Rom. J. Intern. Med. 47, 381–386.
Stenvinkel, P. (2011). Adiponectin in chronic kidney disease: a complex and context sensitive clinical situation. J. Ren. Nutr. 21, 82–86.
Summer, R., Fiack, C. A., Ikeda, Y., Sato, K., Dwyer, D., Ouchi, N., Fine, A., Farber, H. W., and Walsh, K. (2009). Adiponectin deficiency: a model of pulmonary hypertension associated with pulmonary vascular disease. Am. J. Physiol. Lung Cell Mol. Physiol. 297, L432–L438.
Summer, R., Little, F. F., Ouchi, N., Takemura, Y., Aprahamian, T., Dwyer, D., Fitzsimmons, K., Suki, B., Parameswaran, H., Fine, A., and Walsh, K. (2008). Alveolar macrophage activation and an emphysema-like phenotype in adiponectin-deficient mice. Am. J. Physiol. Lung Cell Mol. Physiol. 294, L1035–L1042.
Sutherland, T. J., Sears, M. R., McLachlan, C. R., Poulton, R., and Hancox, R. J. (2009). Leptin, adiponectin, and asthma: findings from a population-based cohort study. Ann. Allergy Asthma Immunol. 103, 101–107.
Takemura, Y., Ouchi, N., Shibata, R., Aprahamian, T., Kirber, M. T., Summer, R. S., Kihara, S., and Walsh, K. (2007). Adiponectin modulates inflammatory reactions via calreticulin receptor-dependent clearance of early apoptotic bodies. J. Clin. Invest. 117, 375–386.
Takeuchi, T., Adachi, Y., Ohtsuki, Y., and Furihata, M. (2007). Adiponectin receptors, with special focus on the role of the third receptor, T-cadherin, in vascular disease. Med. Mol. Morphol. 40, 115–120.
Tan, B. K., Adya, R., Lewandowski, K. C., O’Hare, J. P., and Randeva, H. S. (2011). Diurnal variation and effect of insulin on circulating high molecular weight (HMW) adiponectin and NF-kappaB activity in human endothelial cells. Atherosclerosis 214, 174–177.
Tanko, L. B., Siddiq, A., Lecoeur, C., Larsen, P. J., Christiansen, C., Walley, A., and Froguel, P. (2005). ACDC/adiponectin and PPAR-gamma gene polymorphisms: implications for features of obesity. Obes. Res. 13, 2113–2121.
Tao, L., Gao, E., Jiao, X., Yuan, Y., Li, S., Christopher, T. A., Lopez, B. L., Koch, W., Chan, L., Goldstein, B. J., and Ma, X. L. (2007). Adiponectin cardioprotection after myocardial ischemia/reperfusion involves the reduction of oxidative/nitrative stress. Circulation 115, 1408–1416.
Tao, L., Wang, Y., Gao, E., Zhang, H., Yuan, Y., Lau, W. B., Chan, L., Koch, W. J., and Ma, X. L. (2010). Adiponectin: an indispensable molecule in rosiglitazone cardioprotection following myocardial infarction. Circ. Res. 106, 409–417.
Thyagarajan, B., Jacobs, D. R. Jr., Smith, L. J., Kalhan, R., Gross, M. D., and Sood, A. (2010). Serum adiponectin is positively associated with lung function in young adults, independent of obesity: the CARDIA study. Respir. Res. 11, 176.
Tietge, U. J., Boker, K. H., Manns, M. P., and Bahr, M. J. (2004). Elevated circulating adiponectin levels in liver cirrhosis are associated with reduced liver function and altered hepatic hemodynamics. Am. J. Physiol. Endocrinol. Metab. 287, E82–E89.
Tomas, E., Tsao, T. S., Saha, A. K., Murrey, H. E., Zhang Cc, C., Itani, S. I., Lodish, H. F., and Ruderman, N. B. (2002). Enhanced muscle fat oxidation and glucose transport by ACRP30 globular domain: acetyl-CoA carboxylase inhibition and AMP-activated protein kinase activation. Proc. Natl. Acad. Sci. U.S.A. 99, 16309–16313.
Tsao, T. S., Tomas, E., Murrey, H. E., Hug, C., Lee, D. H., Ruderman, N. B., Heuser, J. E., and Lodish, H. F. (2003). Role of disulfide bonds in Acrp30/adiponectin structure and signaling specificity. Different oligomers activate different signal transduction pathways. J. Biol. Chem. 278, 50810–50817.
Tsuchida, A., Yamauchi, T., and Kadowaki, T. (2005). Nuclear receptors as targets for drug development: molecular mechanisms for regulation of obesity and insulin resistance by peroxisome proliferator-activated receptor gamma, CREB-binding protein, and adiponectin. J. Pharmacol. Sci. 97, 164–170.
Tu, Q., Zhang, J., Dong, L. Q., Saunders, E., Luo, E., Tang, J., and Chen, J. (2011). Adiponectin inhibits osteoclastogenesis and bone resorption via APPL1-mediated suppression of Akt1. J. Biol. Chem. 286, 12542–12553.
Uribe, M., Zamora-Valdes, D., Moreno-Portillo, M., Bermejo-Martinez, L., Pichardo-Bahena, R., Baptista-Gonzalez, H. A., Ponciano-Rodriguez, G., Uribe, M. H., Medina-Santillan, R., and Mendez-Sanchez, N. (2008). Hepatic expression of ghrelin and adiponectin and their receptors in patients with nonalcoholic fatty liver disease. Ann. Hepatol. 7, 67–71.
Van Berendoncks, A. M., Garnier, A., Beckers, P., Hoymans, V. Y., Possemiers, N., Fortin, D., Martinet, W., Van Hoof, V., Vrints, C. J., Ventura-Clapier, R., and Conraads, V. M. (2010). Functional adiponectin resistance at the level of the skeletal muscle in mild to moderate chronic heart failure. Circ. Heart Fail. 3, 185–194.
Vu, V., Riddell, M. C., and Sweeney, G. (2007). Circulating adiponectin and adiponectin receptor expression in skeletal muscle: effects of exercise. Diabetes Metab. Res. Rev. 23, 600–611.
Waki, H., Yamauchi, T., Kamon, J., Ito, Y., Uchida, S., Kita, S., Hara, K., Hada, Y., Vasseur, F., Froguel, P., Kimura, S., Nagai, R., and Kadowaki, T. (2003). Impaired multimerization of human adiponectin mutants associated with diabetes. Molecular structure and multimer formation of adiponectin. J. Biol. Chem. 278, 40352–40363.
Waki, H., Yamauchi, T., Kamon, J., Kita, S., Ito, Y., Hada, Y., Uchida, S., Tsuchida, A., Takekawa, S., and Kadowaki, T. (2005). Generation of globular fragment of adiponectin by leukocyte elastase secreted by monocytic cell line THP-1. Endocrinology 146, 790–796.
Wang, A., Liu, M., Liu, X., Dong, L. Q., Glickman, R. D., Slaga, T. J., Zhou, Z., and Liu, F. (2011). Up-regulation of adiponectin by resveratrol: the essential roles of the Akt/FOXO1 and AMP-activated protein kinase signaling pathways and DsbA-L. J. Biol. Chem. 286, 60–66.
Wang, C., Xin, X., Xiang, R., Ramos, F. J., Liu, M., Lee, H. J., Chen, H., Mao, X., Kikani, C. K., Liu, F., and Dong, L. Q. (2009a). Yin-Yang regulation of adiponectin signaling by APPL isoforms in muscle cells. J. Biol. Chem. 284, 31608–31615.
Wang, Y., Gao, E., Tao, L., Lau, W. B., Yuan, Y., Goldstein, B. J., Lopez, B. L., Christopher, T. A., Tian, R., Koch, W., and Ma, X. L. (2009b). AMP-activated protein kinase deficiency enhances myocardial ischemia/reperfusion injury but has minimal effect on the antioxidant/antinitrative protection of adiponectin. Circulation 119, 835–844.
Wang, Y., Zhou, M., Lam, K. S., and Xu, A. (2009c). Protective roles of adiponectin in obesity-related fatty liver diseases: mechanisms and therapeutic implications. Arq. Bras. Endocrinol. Metabol. 53, 201–212.
Wang, Y., Lam, K. S., Chan, L., Chan, K. W., Lam, J. B., Lam, M. C., Hoo, R. C., Mak, W. W., Cooper, G. J., and Xu, A. (2006). Post-translational modifications of the four conserved lysine residues within the collagenous domain of adiponectin are required for the formation of its high molecular weight oligomeric complex. J. Biol. Chem. 281, 16391–16400.
Wang, Y., Lam, K. S., Yau, M. H., and Xu, A. (2008). Post-translational modifications of adiponectin: mechanisms and functional implications. Biochem. J. 409, 623–633.
Wang, Y., Xu, A., Knight, C., Xu, L. Y., and Cooper, G. J. (2002). Hydroxylation and glycosylation of the four conserved lysine residues in the collagenous domain of adiponectin. Potential role in the modulation of its insulin-sensitizing activity. J. Biol. Chem. 277, 19521–19529.
Wang, Z. V., and Scherer, P. E. (2008). Adiponectin, cardiovascular function, and hypertension. Hypertension 51, 8–14.
Wang, Z. V., Schraw, T. D., Kim, J. Y., Khan, T., Rajala, M. W., Follenzi, A., and Scherer, P. E. (2007). Secretion of the adipocyte-specific secretory protein adiponectin critically depends on thiol-mediated protein retention. Mol. Cell. Biol. 27, 3716–3731.
Weyer, C., Funahashi, T., Tanaka, S., Hotta, K., Matsuzawa, Y., Pratley, R. E., and Tataranni, P. A. (2001). Hypoadiponectinemia in obesity and type 2 diabetes: close association with insulin resistance and hyperinsulinemia. J. Clin. Endocrinol. Metab. 86, 1930–1935.
Williams, G. A., Wang, Y., Callon, K. E., Watson, M., Lin, J. M., Lam, J. B., Costa, J. L., Orpe, A., Broom, N., Naot, D., Reid, I. R., and Cornish, J. (2009). In vitro and in vivo effects of adiponectin on bone. Endocrinology 150, 3603–3610.
Wu, X., Motoshima, H., Mahadev, K., Stalker, T. J., Scalia, R., and Goldstein, B. J. (2003). Involvement of AMP-activated protein kinase in glucose uptake stimulated by the globular domain of adiponectin in primary rat adipocytes. Diabetes 52, 1355–1363.
Xin, X., Zhou, L., Reyes, C. M., Liu, F., and Dong, L. Q. (2011). APPL1 mediates adiponectin-stimulated p38 MAPK activation by scaffolding the TAK1-MKK3-p38 MAPK pathway. Am. J. Physiol. Endocrinol. Metab. 300, E103–E110.
Xu, A., Chan, K. W., Hoo, R. L., Wang, Y., Tan, K. C., Zhang, J., Chen, B., Lam, M. C., Tse, C., Cooper, G. J., and Lam, K. S. (2005). Testosterone selectively reduces the high molecular weight form of adiponectin by inhibiting its secretion from adipocytes. J. Biol. Chem. 280, 18073–18080.
Xu, A., Wang, Y., Lam, K. S., and Vanhoutte, P. M. (2010). Vascular actions of adipokines molecular mechanisms and therapeutic implications. Adv. Pharmacol. 60, 229–255.
Xu, Y., Wang, N., Ling, F., Li, P., and Gao, Y. (2009). Receptor for activated C-kinase 1, a novel binding partner of adiponectin receptor 1. Biochem. Biophys. Res. Commun. 378, 95–98.
Yamauchi, T., Hara, K., Kubota, N., Terauchi, Y., Tobe, K., Froguel, P., Nagai, R., and Kadowaki, T. (2003a). Dual roles of adiponectin/Acrp30 in vivo as an anti-diabetic and anti-atherogenic adipokine. Curr. Drug Targets Immune Endocr. Metabol. Disord. 3, 243–254.
Yamauchi, T., Kamon, J., Ito, Y., Tsuchida, A., Yokomizo, T., Kita, S., Sugiyama, T., Miyagishi, M., Hara, K., Tsunoda, M., Murakami, K., Ohteki, T., Uchida, S., Takekawa, S., Waki, H., Tsuno, N. H., Shibata, Y., Terauchi, Y., Froguel, P., Tobe, K., Koyasu, S., Taira, K., Kitamura, T., Shimizu, T., Nagai, R., and Kadowaki, T. (2003b). Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature 423, 762–769.
Yamauchi, T., Kamon, J., Waki, H., Imai, Y., Shimozawa, N., Hioki, K., Uchida, S., Ito, Y., Takakuwa, K., Matsui, J., Takata, M., Eto, K., Terauchi, Y., Komeda, K., Tsunoda, M., Murakami, K., Ohnishi, Y., Naitoh, T., Yamamura, K., Ueyama, Y., Froguel, P., Kimura, S., Nagai, R., and Kadowaki, T. (2003c). Globular adiponectin protected ob/ob mice from diabetes and ApoE-deficient mice from atherosclerosis. J. Biol. Chem. 278, 2461–2468.
Yamauchi, T., and Kadowaki, T. (2008). Physiological and pathophysiological roles of adiponectin and adiponectin receptors in the integrated regulation of metabolic and cardiovascular diseases. Int. J. Obes. (Lond.) 32(Suppl 7), S13–S18.
Yamauchi, T., Kamon, J., Minokoshi, Y., Ito, Y., Waki, H., Uchida, S., Yamashita, S., Noda, M., Kita, S., Ueki, K., Eto, K., Akanuma, Y., Froguel, P., Foufelle, F., Ferre, P., Carling, D., Kimura, S., Nagai, R., Kahn, B. B., and Kadowaki, T. (2002). Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 8, 1288–1295.
Yamauchi, T., Kamon, J., Waki, H., Terauchi, Y., Kubota, N., Hara, K., Mori, Y., Ide, T., Murakami, K., Tsuboyama-Kasaoka, N., Ezaki, O., Akanuma, Y., Gavrilova, O., Vinson, C., Reitman, M. L., Kagechika, H., Shudo, K., Yoda, M., Nakano, Y., Tobe, K., Nagai, R., Kimura, S., Tomita, M., Froguel, P., and Kadowaki, T. (2001). The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 7, 941–946.
Yamauchi, T., Nio, Y., Maki, T., Kobayashi, M., Takazawa, T., Iwabu, M., Okada-Iwabu, M., Kawamoto, S., Kubota, N., Kubota, T., Ito, Y., Kamon, J., Tsuchida, A., Kumagai, K., Kozono, H., Hada, Y., Ogata, H., Tokuyama, K., Tsunoda, M., Ide, T., Murakami, K., Awazawa, M., Takamoto, I., Froguel, P., Hara, K., Tobe, K., Nagai, R., Ueki, K., and Kadowaki, T. (2007). Targeted disruption of AdipoR1 and AdipoR2 causes abrogation of adiponectin binding and metabolic actions. Nat. Med. 13, 332–339.
Yang, W. S., and Chuang, L. M. (2006). Human genetics of adiponectin in the metabolic syndrome. J. Mol. Med. 84, 112–121.
Yang, W. S., Yang, Y. C., Chen, C. L., Wu, I. L., Lu, J. Y., Lu, F. H., Tai, T. Y., and Chang, C. J. (2007). Adiponectin SNP276 is associated with obesity, the metabolic syndrome, and diabetes in the elderly. Am. J. Clin. Nutr. 86, 509–513.
Yilmaz, M. I., Sonmez, A., Caglar, K., Gok, D. E., Eyileten, T., Yenicesu, M., Acikel, C., Bingol, N., Kilic, S., Oguz, Y., and Vural, A. (2004). Peroxisome proliferator-activated receptor gamma (PPAR-gamma) agonist increases plasma adiponectin levels in type 2 diabetic patients with proteinuria. Endocrine 25, 207–214.
Yoda-Murakami, M., Taniguchi, M., Takahashi, K., Kawamata, S., Saito, K., Choi-Miura, N. H., and Tomita, M. (2001). Change in expression of GBP28/adiponectin in carbon tetrachloride-administrated mouse liver. Biochem. Biophys. Res. Commun. 285, 372–377.
Yoon, M. J., Lee, G. Y., Chung, J. J., Ahn, Y. H., Hong, S. H., and Kim, J. B. (2006). Adiponectin increases fatty acid oxidation in skeletal muscle cells by sequential activation of AMP-activated protein kinase, p38 mitogen-activated protein kinase, and peroxisome proliferator-activated receptor alpha. Diabetes 55, 2562–2570.
Zhou, L., Deepa, S. S., Etzler, J. C., Ryu, J., Mao, X., Fang, Q., Liu, D. D., Torres, J. M., Jia, W., Lechleiter, J. D., Liu, F., and Dong, L. Q. (2009). Adiponectin activates AMP-activated protein kinase in muscle cells via APPL1/LKB1-dependent and phospholipase C/Ca2+/Ca2+/calmodulin-dependent protein kinase kinase-dependent pathways. J. Biol. Chem. 284, 22426–22435.
Zhou, M., Xu, A., Lam, K. S., Tam, P. K., Che, C. M., Chan, L., Lee, I. K., Wu, D., and Wang, Y. (2010). Rosiglitazone promotes fatty acyl CoA accumulation and excessive glycogen storage in livers of mice without adiponectin. J. Hepatol. 53, 1108–1116.
Zhu, M., Hug, C., Kasahara, D. I., Johnston, R. A., Williams, A. S., Verbout, N. G., Si, H., Jastrab, J., Srivastava, A., Williams, E. S., Ranscht, B., and Shore, S. A. (2010). Impact of adiponectin deficiency on pulmonary responses to acute ozone exposure in mice. Am. J. Respir. Cell Mol. Biol. 43, 487–497.
Keywords: adiponectin, metabolic syndrome, cardiovascular disease
Citation: Dadson K, Liu Y and Sweeney G (2011) Adiponectin action: a combination of endocrine and autocrine/paracrine effects. Front. Endocrin. 2:62. doi: 10.3389/fendo.2011.00062
Received: 23 June 2011;
Paper pending published: 14 July 2011;
Accepted: 10 October 2011;
Published online: 08 November 2011.
Edited by:
Paul Thomas Pfluger, University of Cincinnati, USAReviewed by:
Daniel Konrad, University Children’s Hospital, SwitzerlandStergios A. Polyzos, Aristotle University of Thessaloniki, Greece
Copyright: © 2011 Dadson, Liu and Sweeney. This is an open-access article subject to a non-exclusive license between the authors and Frontiers Media SA, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and other Frontiers conditions are complied with.
*Correspondence: Gary Sweeney, Department of Biology, York University, Toronto, ON, Canada M3J 1P3. e-mail:Z3N3ZWVuZXlAeW9ya3UuY2E=
†Keith Dadson and Ying Liu have contributed equally to this work.