 Eva K. Wirth
Eva K. Wirth Ulrich Schweizer2
Ulrich Schweizer2 Josef Köhrle
Josef Köhrle- 1Institut für Experimentelle Endokrinologie, Charité-Universitätsmedizin Berlin, Berlin, Germany
- 2Institut für Biochemie und Molekularbiologie, Rheinische Friedrich-Wilhelms-Universität Bonn, Bonn, Germany
Thyroid hormone (TH) transport into the brain is not only pivotal for development and differentiation, but also for maintenance and regulation of adult central nervous system (CNS) function. In this review, we highlight some key factors and structures regulating TH uptake and distribution. Serum TH binding proteins play a major role for the availability of TH since only free hormone concentrations may dictate cellular uptake. One of these proteins, transthyretin is also present in the cerebrospinal fluid (CSF) after being secreted by the choroid plexus. Entry routes into the brain like the blood–brain-barrier (BBB) and the blood–CSF-barrier will be explicated regarding fetal and adult status. Recently identified TH transmembrane transporters (THTT) like monocarboxylate transporter 8 (Mct8) play a major role in uptake of TH across the BBB but as well in transport between cells like astrocytes and neurons within the brain. Species differences in transporter expression will be presented and interference of TH transport by endogenous and exogenous compounds including endocrine disruptors and drugs will be discussed.
Molecules Involved in TH Transport in the Brain
The hydrophobic but amphipathic charged amino acid-derived thyroid hormones (TH) are carried and distributed by several binding proteins from their site of production, storage, and secretion, the thyroid gland, to their target tissues including the brain. In human blood, four major proteins, thyroxine-binding globulin (TBG), transthyretin (TTR), albumin, and apolipoprotein B 100 (ApoB100), bind more than 99% of the circulating TH T4, T3, and 3-iodo-thyronamine (3-T1AM). In contrast, only TTR has been found as one of the main proteins in CSF where it is produced and directionally secreted by choroid plexus (CP) epithelial cells into the liquor, which does not contain the high affinity TBG or the high capacity albumin TH binding proteins. Whether 3-T1AM, a TH-derived biogenic amine and its high affinity binding protein ApoB100 occur in CSF, remains to be studied.
Thyroid hormone enters the brain either directly via the blood–brain barrier (BBB) or indirectly via the blood–CSF-barrier (B–CSF-B), with the BBB route as the major entry path for the prohormone T4. T4 is locally metabolized by selenoenzymes to either active T3 via Type 2 deiodinase (Dio2) or inactivated by Type 3 deiodinase (Dio3), to yield reverse T3 (rT3). rT3, devoid of T3-like action, might be involved in developmental regulation of neuronal migration guided by astrocytes and glial cells (1). Dio2 is mostly expressed in astrocytes and tanycytes while Dio3 is mainly found in neurons. Whether Type 1 deiodinase (Dio1), catalyzing both 5′-deiodination (activation of T4 to T3) and 5-deiodination (inactivation of T4 and T3) is species-dependently expressed in brain remains controversial (2, 3). Dual entry paths of TH and cell type-specific expression of functional Dio enzymes in the brain raise the issues of (i) coordinated transport of active TH and TH metabolites between various brain cell types, (ii) organized communication between peripheral, thyroid-derived, and brain TH, and (iii) demands, supply, and disposal of TH precursors, metabolites, and active TH.
Adequate TH supply for the brain is of eminent importance during development but not less relevant in the differentiated adult organism with its changing hormonal requirement for metabolic and environmental adaptation.
Components controlling TH availability and action have been described in brain stem and progenitor cells (4) and TH receptor (TR) expression in the human brain has been demonstrated decades ago (5). Already during the first trimester human brain expresses various TH transporters (see Table 1), Dio enzymes, TR-isoforms, and isotypes in a development- and cell type-specific manner. Later in human pregnancy during weeks 17–20, endothelial cells and astrocytes organize the BBB [see Ref. (6)]. Endothelial cells express the TH transporter organic anion transporter polypeptide 1C1 (OATP1C1), which limits brain access of TH, especially T4. At this time point fetal thyroid already starts producing TH, thus disconnecting the fetal TH responsive system from the maternal source of TH, but still depending on further adequate maternal iodide supply.
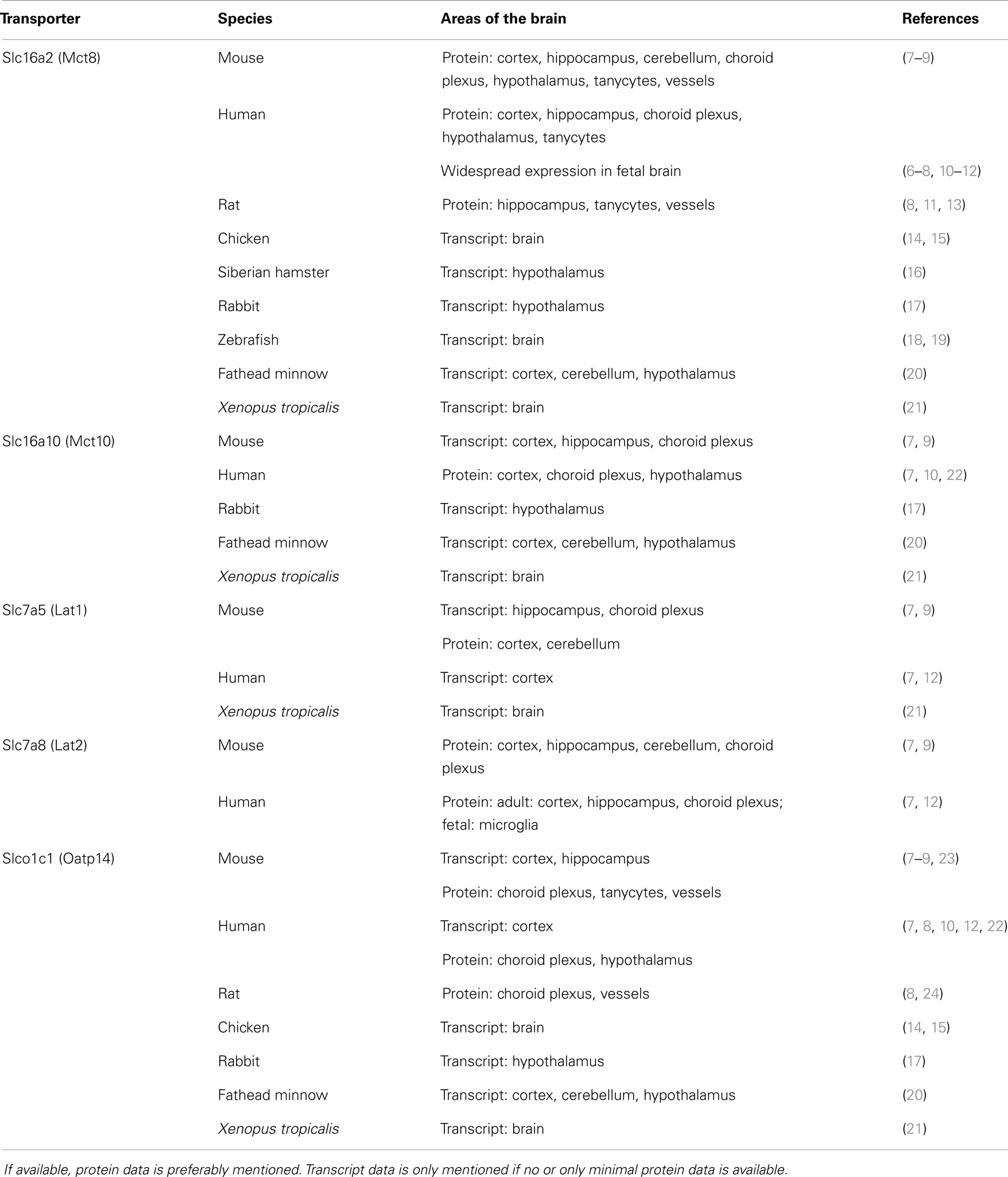
Table 1. Summary of expression profiles of thyroid hormone transmembrane transporters (THTT) in various cell types of the brain of several species.
Facilitated uptake and release of TH by TH transmembrane transporters (THTT) is essential for their intracellular availability. TH have to cross multiple membranes in order to reach their nuclear and mitochondrial receptors. Especially, the entry of TH into the brain via the BBB and their subsequent distribution throughout all brain areas poses challenges in form of membranes of different cell types to be crossed. The complex interaction and communication between astrocytes and neurons, demonstrated for metabolic as well as synaptic processes, is in place regarding TH metabolism and distribution throughout the brain. Therefore, the specific spatio-temporal distribution of TH in different areas and cell types of the brain is required during embryonic development and differentiation, but also for adult maintenance and regulation of brain activity and metabolism.
Role of TH Binding and Distributor Proteins for TH Availability to Brain Cells
Tissue and cellular uptake depends on free TH concentrations in blood and both free T4 and free T3 are available for cellular uptake by THTT, while TH bound with high affinity to serum distributor proteins TBG and TTR is assumed not to be directly available for cellular uptake during organ perfusion by blood (25). In contrast, TH bound to albumin with high capacity but low affinity is easily liberated based on the high TH off-rate constants shown in liver perfusion (26, 27). While peripheral sensory neurons internalize TTR in megalin-dependent manner in context of neuritogenesis in vitro (28), evidence is missing, that TH–TTR ligand–protein complexes are taken up by neurons, astrocytes, or glial cells via receptors for these proteins expressed on brain cells. Such “trojan horse” mechanisms of ligand transmembrane transfer have been demonstrated during fetal development for several protein bound (pro-)hormones and vitamins such as TH, retinol, vitamin D3, and steroid hormones (29–31) for several peripheral target tissues including adult kidney but not yet for the fetal brain. Such mechanisms, mediated by megalin or cubilin receptors, might provide entry routes for hormones bypassing the concept of the “free hormone hypothesis” (32) and use of THTT.
Independent from such a mechanism, the high expression of evolutionary conserved TTR in CP and meninges of the developing and adult brain offers a further exchange compartment for TH, especially T4, a high affinity ligand of TTR. During fetal brain development, the prominent voluminous CP is required for proper CNS growth and differentiation. TTR is the only TH binding protein expressed and directionally secreted into CSF. TTR, first described in CSF and subsequently in plasma, constitutes up to 20% of CSF protein and its secretion into CSF starts already in fetal week 8 before its hepatic production (28). Whether its adequate function as binding and distribution protein for two major morphogen precursors, T4 and RBP-bound retinol, is essential or redundant, remains to be established. Lack of a major phenotype of TTR gene inactivation in the mouse came as a surprise with respect to normal brain development, HPT axis, TH homeostasis, and retinol-dependent functions (33). These mice grew normal, were fertile and had normal tissue T4 levels though plasma TH concentrations were significantly reduced. Apparently, lack of TTR can be compensated by a shift of TH binding to rodent TBG, which has lower TH affinity compared to human TBG (34). TH metabolism and T3-responsive gene expression of HPT axis and liver was unchanged in TTR knock-out mice. These observations either suggest a minor role for TTR-dependent TH binding and directional transport into CSF for proper brain development (35) or indicate existence of still unknown compensatory mechanisms active in absence of TTR during mouse development. At least this mouse model did not support the hypothesis that hormone binding and distribution proteins such as TTR in case of TH directly contribute to cellular hormone uptake as proposed by Pardridge (36) in distinction to several observations by the group of Willnow (31).
In contrast to these observations in the TTR knock-out mouse model are some findings on effects of endogenous or exogenous ligands of TTR, such as (iso-)flavonoids and endocrine disruptors, interfering with TH homeostasis in circulation, in CNS, and during fetal development. Several natural flavonoids, secondary metabolites of plants contained in our regular diet, avidly bind to TTR and displace TH from TTR binding based on their structural resemblance to TH. Resulting elevated free TH blood concentrations increase renal TH loss (transiently), elevate TH tissues levels, and enhance TH transfer via the placenta into fetal circulation including the fetal brain. In vitro as well as rodent animal studies provided evidence for interference of flavonoids and other TTR binding endocrine disruptors with CP-derived TTR-mediated TH transfer into CSF and the brain, resulting in disturbed homeostasis of brain TH levels and bioavailability (37).
More studies are needed analyzing (i) interference by natural and synthetic flavonoids with TH transport, (ii) action of endocrine disrupters such as the flame retardants polybrominated diphenylethers (PBDE) during neuronal stem cell development (38), or (iii) impact of ligands and pharmaceuticals, structurally related to TH (39) and interfering with THTT function (40). Recently, molecular actions and biological functions have been initially characterized for so far “neglected or minor” endogenous TH metabolites such as acetic acid- (Tetrac, Triac) or amine-derivatives (3-T1AM) of TH (41) and 3,5-T2, the latter abundantly present in the CNS (42). This raises the questions, (i) whether they are active players in TH-regulated brain function during development and in the adult organism, (ii) whether and how these metabolites are generated and transported in the brain, and (iii) how their mode of action interferes with classical TH action, which is mainly mediated via T3-liganded TR. New modes of action may be envisaged for these TH metabolites at the plasma membrane, on cytosolic signaling cascades, or on other subcellular compartments of brain cells (43–45).
THTT among Species
Many transporters have been shown to transport TH. The most specific THTT is the monocarboxylate transporter 8 (Mct8; Slc16a2). Up to date, it is the only transporter with TH as the exclusive substrate. All other transporters out of the classes of monocarboxylate transporters (MCT), organic anion transporting polypeptides (OATP), and l-type amino acid transporters (LAT) also transport other substrates like amino acids. Most data about the presence and localization of THTT has been generated in mice and humans. The following table summarizes expression of the most researched THTT Mct8, Mct10, Lat1, Lat2, and Oatp1c1 in various vertebrate species (Table 1).
Research focus has been on the only THTT identified to cause a human disease so far, i.e., Mct8. MCT8 is widely expressed in the human fetal brain in several cell types (Table 1). Strong transcript and protein signals were observed in the cortical plate and subplate, as well as in ventricular and subventricular zones. Throughout fetal development CP epithelial cells and ependymal cells express high levels of MCT8 (6–8).
Comparably high MCT8 expression has been reported for monkey brain, which expresses OATP1C1 and LAT1 albeit at much lower levels (46). MCT8 mutations cause a severe syndrome of psychomotor retardation, the Allan–Herndon–Dudley syndrome (AHDS) (47–50). This syndrome also comprises endocrine manifestations with high circulating T3, low T4, and normal to elevated TSH levels. The mouse model for Mct8-deficiency replicates the endocrine phenotype, but it does not mimic the psychomotor retardation of the human syndrome (7, 51, 52). Since AHDS is not comparable to the classical phenotype of congenital hypothyroidism or cretinism, it is of great importance to identify THTT in brain areas of human brains, as well as in the model organism used for analyzing TH transport to be able to understand phenotypic variations between these models. Other animal models apart from Mct8-deficient mice will be needed to evaluate the involvement of THTT in basic brain development. Recently, zebrafish has been evaluated for developmental effects of Mct8-deficiency. Significant species differences with respect to cell types, time point, dynamics, and regulation of THTT especially in the developing but also the adult brain have been reported for humans, monkey, chicken, rodent, fish, and amphibian brain (Table 1). For example, we demonstrated the expression of an additional transporter, Lat2, in mouse neurons during development, which is not expressed in human developing neurons (7). Research on double knock-out mice of Mct8 with either Mct10 or Oatp1c1 yielded valuable data on the interplay and possible compensation between these transporters (53, 54). Simultaneous deletion of Mct8 and Oatp1c1 lead to the ablation of both T3 and T4 transport across BBB. Symptoms of brain hypothyroidism were intensified underlining the importance of THTT for proper brain development and function. Further research analyzing the developmental expression of THTT and comparing effects of loss of function among different species will yield important information on the temporal effects of these transporters on brain development.
Structural Aspects of THTT
Thyroid hormone transport proteins MCT8, MCT10, OATP1C1, as well as LAT1 and LAT2 belong to different subfamilies within a huge protein superfamily of transport proteins, the major facilitator superfamily, MFS (55). General insights into the function of such transmembrane proteins can be derived from pathogenic mutations, e.g., in MCT8 from patients affected by AHDS (56).
Substrate recognition by transport proteins is fundamentally different from ligand binding in, e.g., nuclear receptors: while receptors are optimized for high affinity binding of their ligands, this would be detrimental for transporters, as these have to release their substrates easily. Most receptors have one binding site, while THTT should have at least two – one accessible from the exterior and one accessible form the interior of the cell. These two binding sites may overlap and differ only according to conformational changes associated with transport. The question is thus, how can a transporter achieve specificity and at the same time prevent tight binding?
With the exception of MCT8, all other THTT transport additional substrates, namely amino acids, bile acids, or conjugated steroids. It should therefore be of particular interest to compare how different protein families have adapted to transport TH and whether the substrate–protein interactions are similar or not. Experimental structures are not available for any of the THTT. Homology models based on experimental structures have been created for OATP1C1 (57), MCT8 (39), and LAT1 (58).
The homology model of rat Oatp1c1 was based on three high resolution crystal structures of bacterial transport proteins, lactose permease LacY, glycerol–phosphate transporter GlpT, and multidrug resistance protein EmrD. Authors achieved similar models with all templates and highlighted sequence identities between Oatp1c1 and templates below 10%. Conserved amino acids between Oatp1c1 and bacterial transporters were known to be functionally important in LacY and GlpT and may therefore not be involved in substrate specificity. While this work nicely shows that Oatp1c1 conforms to the overall structure of the bacterial transporters, there is no information on how specificity is established (57).
Chemical probes reactive with cysteines (p-chloromercury benzenesulfonate, pCMBS) or histidines (diethylpyrocarbonate, DEPC) were used to modify MCT8 and to test its activity afterward (59, 60). This approach suggested that Cys481, Cys497, and His192 may be close to the substrate translocation channel. Mutation to Ala of these critical amino acids rendered MCT8 resistant to pCMBS and DEPC.
We created a MCT8 homology model based on the inward-open conformation of GlpT and identified two charged amino acids within the transmembrane domains, Asp498 and Arg445, which are essential for transport (39). The homology model predicted a salt bridge between both residues. We suggested interaction of TH carboxyl and amino groups with these amino acids during transport, since TH analogs lacking the carboxyl or amino groups are not transported by MCT8 (39). The salt bridge was later independently confirmed by charge reversal mutants (61). MCT8 accepts only L-T3, L-T4, L-rT3, and L-3,3′-T2 as substrates (39). Based on the occurrence of a His–Arg clamp pinching T3 in the T3 receptor β structure, we tested the hypothesis that a His–Arg pair spaced by about 15Å could serve the same purpose in MCT8 (62). Mutation of His192 (which may work together with Arg445 in an outward open conformation) clearly demonstrated His192 participation in substrate recognition (63). Interestingly, His192 corresponds to Gln88 in MCT10, a closely related homolog of MCT8 unable to transport T4 (64). Mutation of His415 and Arg301, conserved in MCT8 and MCT10, affected transport kinetics as expected from substrate interactions (63). These findings corroborate the usefulness of the MCT8 homology model and suggest how the substrate is bound by MCT8 – at least in the inward-open conformation.
Recently, a LAT1 homology model was presented based on the crystal structure of bacterial agmatine antiporter AdiC (58). Iodotyrosines were identified in silico as LAT1 substrates and confirmed experimentally. Interestingly, while carboxy and amino groups are present in all LAT1 substrates, modeling suggests that these functions are sampled by the transporter by backbone polar contacts instead of side chain contacts as predicted in MCT8 (58, 63). Different transporters may have adapted different strategies to recognize iodinated TH substrates.
THTT in Cell Types of the Brain
Analyses of brain regions provide important insight into THTT distribution. Immunohistochemical staining for Mct8 in mouse brains did not only show typical neuronal staining patterns, but also staining in astrocytes, CP, and tanycytes (7). However, most techniques like in situ hybridization or immunohistochemistry do not allow for a real cell type-specific resolution and detection. Effects of TH uptake into neurons vs. astrocytes can not be dissected in complete brains. Primary cultures of mouse brain cells facilitate selection of a single cell type and the evaluation of effects of THTT deletion. Primary cultures of mouse neurons, astrocytes, and microglia can be used to detect THTT expression and functionality, as well as the reaction of specific cells on different conditions of TH access. Genetically engineered mouse lines can be used to create cell cultures originating from wildtype and transporter-deficient mice. Employing these cultures, we detected expression of Mct8, Lat1, and Lat2 transcripts and proteins in neurons and astrocytes, while Lat2 is additionally expressed in microglia (9, 65). Functional uptake studies in Mct8- and Lat2-deficient primary neuronal and astrocyte cell cultures demonstrated involvement of both transporters in T3 and T4 uptake into neurons and astrocytes (7, 9). The fraction of TH uptake mediated by transporters of the Mct, Lat, and Oatp groups can also be monitored by utilizing inhibitors of these transporters. Transport by Mct can be inhibited with bromosulphophtalein (BSP), while 2-aminobicyclo-(2,2,1)-heptane-2-carboxylic acid (BCH) is a Lat inhibitor and probenicid inhibits transport by Oatps. Uptake studies with these inhibitors gave very similar results on the involvement of these groups of transporters in TH uptake to genetic inactivation (7, 9).
Up to date, all concepts of interaction and cooperation between neurons and astrocytes imply astrocytes as providers of energy, communication and T3 for neurons. However, it has been shown that neurons are generally able to carry out all functions needed for energy metabolism by themselves. It is therefore quite possible that neurons are able to convert T4 to T3 by expressing functional Dio2. Cell type-specific animal models and primary cell cultures are of great importance to study the interaction of neurons and astrocytes regarding transfer of TH metabolites and regulation of deiodinases independent of TH uptake at the BBB.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Eva K. Wirth, Ulrich Schweizer, and Josef Köhrle were supported by funding of the Deutsche Forschungsgemeinschaft (WI3768/1-1, WI3768/2-1, KO922/16-1, GRK1208/2 TP3, Schw914/3-1, and SPP 1629 ThyroidTransAct).
References
1. Farwell AP, Dubord-Tomasetti SA, Pietrzykowski AZ, Stachelek SJ, Leonard JL. Regulation of cerebellar neuronal migration and neurite outgrowth by thyroxine and 3,3′,5′-triiodothyronine. Brain Res Dev Brain Res (2005) 154(1):121–35. doi: 10.1016/j.devbrainres.2004.07.016
2. Visser TJ, Leonard JL, Kaplan MM, Larsen PR. Kinetic evidence suggesting two mechanisms for iodothyronine 5′-deiodination in rat cerebral cortex. Proc Natl Acad Sci U S A (1982) 79(16):5080–4. doi:10.1073/pnas.79.16.5080
3. Verhoelst CH, Roelens SA, Darras VM. Role of spatiotemporal expression of iodothyronine deiodinase proteins in cerebellar cell organization. Brain Res Bull (2005) 67(3):196–202. doi:10.1016/j.brainresbull.2005.06.030
4. Lemkine GF, Raj A, Alfama G, Turque N, Hassani Z, Alegria-Prevot O, et al. Adult neural stem cell cycling in vivo requires thyroid hormone and its alpha receptor. FASEB J (2005) 19(7):863–5. doi:10.1096/fj.04-2916fje
5. Bernal J, Pekonen F. Ontogenesis of the nuclear 3,5,3′-triiodothyronine receptor in the human fetal brain. Endocrinology (1984) 114(2):677–9. doi:10.1210/endo-114-2-677
6. Chan SY, Hancox LA, Martin-Santos A, Loubiere LS, Walter MN, Gonzalez AM, et al. MCT8 expression in human fetal cerebral cortex is reduced in severe intrauterine growth restriction. J Endocrinol (2014) 220(2):85–95. doi:10.1530/JOE-13-0400
7. Wirth EK, Roth S, Blechschmidt C, Hölter SM, Becker L, Racz I, et al. Neuronal 3′,3,5-triiodothyronine (T3) uptake and behavioral phenotype of mice deficient in Mct8, the neuronal T3 transporter mutated in Allan-Herndon-Dudley syndrome. J Neurosci (2009) 29(30):9439–49. doi:10.1523/JNEUROSCI.6055-08.2009 Epub 2009/07/31,
8. Roberts LM, Woodford K, Zhou M, Black DS, Haggerty JE, Tate EH, et al. Expression of the thyroid hormone transporters monocarboxylate transporter-8 (SLC16A2) and organic ion transporter-14 (SLCO1C1) at the blood-brain barrier. Endocrinology (2008) 149(12):6251–61. doi:10.1210/en.2008-0378
9. Braun D, Kinne A, Bräuer AU, Sapin R, Klein MO, Köhrle J, et al. Developmental and cell type-specific expression of thyroid hormone transporters in the mouse brain and in primary brain cells. Glia (2011) 59(3):463–71. doi:10.1002/glia.21116 Epub 2011/01/26,
10. Friesema EC, Visser TJ, Borgers AJ, Kalsbeek A, Swaab DF, Fliers E, et al. Thyroid hormone transporters and deiodinases in the developing human hypothalamus. Eur J Endocrinol (2012) 167(3):379–86. doi:10.1530/EJE-12-0177
11. Kallo I, Mohacsik P, Vida B, Zeold A, Bardoczi Z, Zavacki AM, et al. A novel pathway regulates thyroid hormone availability in rat and human hypothalamic neurosecretory neurons. PLoS One (2012) 7(6):e37860. doi:10.1371/journal.pone.0037860
12. Chan SY, Martin-Santos A, Loubiere LS, Gonzalez AM, Stieger B, Logan A, et al. The expression of thyroid hormone transporters in the human fetal cerebral cortex during early development and in N-Tera-2 neurodifferentiation. J Physiol (2011) 589(Pt 11):2827–45. doi:10.1113/jphysiol.2011.207290 Epub 2011/04/14,
13. Sharlin DS, Gilbert ME, Taylor MA, Ferguson DC, Zoeller RT. The nature of the compensatory response to low thyroid hormone in the developing brain. J Neuroendocrinol (2010) 22(3):153–65. doi:10.1111/j.1365-2826.2009.01947.x Epub 2010/01/01,
14. Geysens S, Ferran JL, Van Herck SL, Tylzanowski P, Puelles L, Darras VM. Dynamic mRNA distribution pattern of thyroid hormone transporters and deiodinases during early embryonic chicken brain development. Neuroscience (2012) 221:69–85. doi:10.1016/j.neuroscience.2012.06.057
15. Van Herck SL, Geysens S, Delbaere J, Tylzanowski P, Darras VM. Expression profile and thyroid hormone responsiveness of transporters and deiodinases in early embryonic chicken brain development. Mol Cell Endocrinol (2012) 349(2):289–97. doi:10.1016/j.mce.2011.11.012
16. Herwig A, de Vries EM, Bolborea M, Wilson D, Mercer JG, Ebling FJ, et al. Hypothalamic ventricular ependymal thyroid hormone deiodinases are an important element of circannual timing in the Siberian hamster (Phodopus sungorus). PLoS One (2013) 8(4):e62003. doi:10.1371/journal.pone.0062003
17. Mebis L, Debaveye Y, Ellger B, Derde S, Ververs EJ, Langouche L, et al. Changes in the central component of the hypothalamus-pituitary-thyroid axis in a rabbit model of prolonged critical illness. Crit Care (2009) 13(5):R147. doi:10.1186/cc8043
18. Arjona FJ, de Vrieze E, Visser TJ, Flik G, Klaren PH. Identification and functional characterization of zebrafish solute carrier Slc16a2 (Mct8) as a thyroid hormone membrane transporter. Endocrinology (2011) 152(12):5065–73. doi:10.1210/en.2011-1166
19. Vatine GD, Zada D, Lerer-Goldshtein T, Tovin A, Malkinson G, Yaniv K, et al. Zebrafish as a model for monocarboxyl transporter 8-deficiency. J Biol Chem (2013) 288(1):169–80. doi:10.1074/jbc.M112.413831
20. Muzzio AM, Noyes PD, Stapleton HM, Lema SC. Tissue distribution and thyroid hormone effects on mRNA abundance for membrane transporters Mct8, Mct10, and organic anion-transporting polypeptides (Oatps) in a teleost fish. Comp Biochem Physiol A Mol Integr Physiol (2014) 167:77–89. doi:10.1016/j.cbpa.2013.09.019
21. Connors KA, Korte JJ, Anderson GW, Degitz SJ. Characterization of thyroid hormone transporter expression during tissue-specific metamorphic events in Xenopus tropicalis. Gen Comp Endocrinol (2010) 168(1):149–59. doi:10.1016/j.ygcen.2010.04.015 Epub 2010/04/27,
22. Alkemade A, Friesema EC, Kalsbeek A, Swaab DF, Visser TJ, Fliers E. Expression of thyroid hormone transporters in the human hypothalamus. J Clin Endocrinol Metab (2011) 96(6):E967–71. doi:10.1210/jc.2010-2750
23. Tohyama K, Kusuhara H, Sugiyama Y. Involvement of multispecific organic anion transporter, Oatp14 (Slc21a14), in the transport of thyroxine across the blood-brain barrier. Endocrinology (2004) 145(9):4384–91. doi:10.1210/en.2004-0058
24. Sugiyama D, Kusuhara H, Taniguchi H, Ishikawa S, Nozaki Y, Aburatani H, et al. Functional characterization of rat brain-specific organic anion transporter (Oatp14) at the blood-brain barrier: high affinity transporter for thyroxine. J Biol Chem (2003) 278(44):43489–95. doi:10.1074/jbc.M306933200
25. Richardson SJ. Evolutionary changes to transthyretin: evolution of transthyretin biosynthesis. FEBS J (2009) 276(19):5342–56. doi:10.1111/j.1742-4658.2009.07244.x
26. Mendel CM, Weisiger RA, Jones AL, Cavalieri RR. Thyroid hormone-binding proteins in plasma facilitate uniform distribution of thyroxine within tissues: a perfused rat liver study. Endocrinology (1987) 120(5):1742–9. doi:10.1210/endo-120-5-1742
27. Richardson SJ. Cell and molecular biology of transthyretin and thyroid hormones. Int Rev Cytol (2007) 258:137–93. doi:10.1016/S0074-7696(07)58003-4
28. Fleming CE, Mar FM, Franquinho F, Saraiva MJ, Sousa MM. Transthyretin internalization by sensory neurons is megalin mediated and necessary for its neuritogenic activity. J Neurosci (2009) 29(10):3220–32. doi:10.1523/JNEUROSCI.6012-08.2009
29. Sousa MM, Norden AG, Jacobsen C, Willnow TE, Christensen EI, Thakker RV, et al. Evidence for the role of megalin in renal uptake of transthyretin. J Biol Chem (2000) 275(49):38176–81. doi:10.1074/jbc.M002886200
30. Willnow TE, Hammes A, Nykjaer A. Endocytosis of sex steroids: the hypothesis of free hormones revisited. Ann Endocrinol (Paris) (2008) 69(2):101–2. doi:10.1016/j.ando.2008.02.024
31. Willnow TE, Nykjaer A. Cellular uptake of steroid carrier proteins – mechanisms and implications. Mol Cell Endocrinol (2010) 316(1):93–102. doi:10.1016/j.mce.2009.07.021
32. Mendel CM. The free hormone hypothesis. Distinction from the free hormone transport hypothesis. J Androl (1992) 13(2):107–16.
33. Episkopou V, Maeda S, Nishiguchi S, Shimada K, Gaitanaris GA, Gottesman ME, et al. Disruption of the transthyretin gene results in mice with depressed levels of plasma retinol and thyroid hormone. Proc Natl Acad Sci U S A (1993) 90(6):2375–9. doi:10.1073/pnas.90.6.2375
34. Vranckx R, Savu L, Maya M, Nunez EA. Characterization of a major development-regulated serum thyroxine-binding globulin in the euthyroid mouse. Biochem J (1990) 271(2):373–9.
35. Palha JA. Transthyretin as a thyroid hormone carrier: function revisited. Clin Chem Lab Med (2002) 40(12):1292–300. doi:10.1515/CCLM.2002.223
36. Pardridge WM. Plasma protein-mediated transport of steroid and thyroid hormones. Am J Physiol (1987) 252(2 Pt 1):E157–64.
37. Schröder-van der Elst JP, van der Heide D, Rokos H, Morreale de Escobar G, Köhrle J. Synthetic flavonoids cross the placenta in the rat and are found in fetal brain. Am J Physiol (1998) 274(Pt 1):E253–6.
38. Morvan-Dubois G, Fini JB, Demeneix BA. Is thyroid hormone signaling relevant for vertebrate embryogenesis? Curr Top Dev Biol (2013) 103:365–96. doi:10.1016/B978-0-12-385979-2.00013-7
39. Kinne A, Kleinau G, Hoefig CS, Grüters A, Köhrle J, Krause G, et al. Essential molecular determinants for thyroid hormone transport and first structural implications for monocarboxylate transporter 8. J Biol Chem (2010) 285(36):28054–63. doi:10.1074/jbc.M110.129577
40. Braun D, Kim TD, le Coutre P, Kohrle J, Hershman JM, Schweizer U. Tyrosine kinase inhibitors noncompetitively inhibit MCT8-mediated iodothyronine transport. J Clin Endocrinol Metab (2012) 97(1):E100–5. doi:10.1210/jc.2011-1837
41. Brix K, Fuhrer D, Biebermann H. Molecules important for thyroid hormone synthesis and action – known facts and future perspectives. Thyroid Res (2011) 4(Suppl 1):S9. doi:10.1186/1756-6614-4-S1-S9 Epub 2011/08/13,
42. Pinna G, Meinhold H, Hiedra L, Thoma R, Hoell T, Graf KJ, et al. Elevated 3,5-diiodothyronine concentrations in the sera of patients with nonthyroidal illnesses and brain tumors. J Clin Endocrinol Metab (1997) 82(5):1535–42. doi:10.1210/jcem.82.5.3939
43. Moeller LC, Broecker-Preuss M. Transcriptional regulation by nonclassical action of thyroid hormone. Thyroid Res (2011) 4(Suppl 1):S6. doi:10.1186/1756-6614-4-S1-S6 Epub 2011/08/13,
44. Davis PJ, Lin HY, Tang HY, Davis FB, Mousa SA. Adjunctive input to the nuclear thyroid hormone receptor from the cell surface receptor for the hormone. Thyroid (2013) 23(12):1503–9. doi:10.1089/thy.2013.0280
45. Brent GA. Mechanisms of thyroid hormone action. J Clin Invest (2012) 122(9):3035–43. doi:10.1172/JCI60047
46. Ito K, Uchida Y, Ohtsuki S, Aizawa S, Kawakami H, Katsukura Y, et al. Quantitative membrane protein expression at the blood-brain barrier of adult and younger cynomolgus monkeys. J Pharm Sci (2011) 100(9):3939–50. doi:10.1002/jps.22487 Epub 2011/01/22,
47. Schwartz CE, May MM, Carpenter NJ, Rogers RC, Martin J, Bialer MG, et al. Allan-Herndon-Dudley syndrome and the monocarboxylate transporter 8 (MCT8) gene. Am J Hum Genet (2005) 77(1):41–53. doi:10.1086/431313
48. Biebermann H, Ambrugger P, Tarnow P, von Moers A, Schweizer U, Grueters A. Extended clinical phenotype, endocrine investigations and functional studies of a loss-of-function mutation A150V in the thyroid hormone specific transporter MCT8. Eur J Endocrinol (2005) 153(3):359–66. doi:10.1530/eje.1.01980
49. Dumitrescu AM, Liao XH, Best TB, Brockmann K, Refetoff S. A novel syndrome combining thyroid and neurological abnormalities is associated with mutations in a monocarboxylate transporter gene. Am J Hum Genet (2004) 74(1):168–75. doi:10.1086/380999
50. Friesema EC, Grueters A, Biebermann H, Krude H, von Moers A, Reeser M, et al. Association between mutations in a thyroid hormone transporter and severe X-linked psychomotor retardation. Lancet (2004) 364(9443):1435–7. doi:10.1016/S0140-6736(04)17226-7
51. Dumitrescu AM, Liao XH, Weiss RE, Millen K, Refetoff S. Tissue-specific thyroid hormone deprivation and excess in monocarboxylate transporter (mct) 8-deficient mice. Endocrinology (2006) 147(9):4036–43. doi:10.1210/en.2006-0390
52. Trajkovic M, Visser TJ, Mittag J, Horn S, Lukas J, Darras VM, et al. Abnormal thyroid hormone metabolism in mice lacking the monocarboxylate transporter 8. J Clin Invest (2007) 117(3):627–35. doi:10.1172/JCI28253
53. Müller J, Mayerl S, Visser TJ, Darras VM, Boelen A, Frappart L, et al. Tissue-specific alterations in thyroid hormone homeostasis in combined Mct10 and Mct8 deficiency. Endocrinology (2014) 155(1):315–25. doi:10.1210/en.2013-1800
54. Mayerl S, Müller J, Bauer R, Richert S, Kassmann CM, Darras VM, et al. Transporters MCT8 and OATP1C1 maintain murine brain thyroid hormone homeostasis. J Clin Invest (2014) 124(5):1987–99. doi:10.1172/JCI70324
55. Kinne A, Schulein R, Krause G. Primary and secondary thyroid hormone transporters. Thyroid Res (2011) 4(Suppl 1):S7. doi:10.1186/1756-6614-4-S1-S7 Epub 2011/08/13,
56. Kinne A, Roth S, Biebermann H, Köhrle J, Grüters A, Schweizer U. Surface translocation and tri-iodothyronine uptake of mutant MCT8 proteins are cell type-dependent. J Mol Endocrinol (2009) 43(6):263–71. doi:10.1677/JME-09-0043
57. Westholm DE, Marold JD, Viken KJ, Duerst AH, Anderson GW, Rumbley JN. Evidence of evolutionary conservation of function between the thyroxine transporter Oatp1c1 and major facilitator superfamily members. Endocrinology (2010) 151(12):5941–51. doi:10.1210/en.2010-0640
58. Geier EG, Schlessinger A, Fan H, Gable JE, Irwin JJ, Sali A, et al. Structure-based ligand discovery for the Large-neutral Amino Acid Transporter 1, LAT-1. Proc Natl Acad Sci U S A (2013) 110(14):5480–5. doi:10.1073/pnas.1218165110
59. Lima de Souza EC, Groeneweg S, Visser WE, Peeters RP, Visser TJ. Importance of cysteine residues in the thyroid hormone transporter MCT8. Endocrinology (2013) 154(5):1948–55. doi:10.1210/en.2012-2101
60. Groeneweg S, Lima de Souza EC, Visser WE, Peeters RP, Visser TJ. Importance of His192 in the human thyroid hormone transporter MCT8 for substrate recognition. Endocrinology (2013) 154(7):2525–32. doi:10.1210/en.2012-2225
61. Groeneweg S, Friesema EC, Kersseboom S, Klootwijk W, Visser WE, Peeters RP, et al. The role of Arg445 and Asp498 in the human thyroid hormone transporter MCT8. Endocrinology (2014) 155(2):618–26. doi:10.1210/en.2013-1521
62. Kleinau G, Schweizer U, Kinne A, Köhrle J, Grüters A, Krude H, et al. Insights into molecular properties of the human monocarboxylate transporter 8 by combining functional with structural information. Thyroid Res (2011) 4(Suppl 1):S4. doi:10.1186/1756-6614-4-S1-S4 Epub 2011/08/13,
63. Braun D, Lelios I, Krause G, Schweizer U. Histidines in potential substrate recognition sites affect thyroid hormone transport by monocarboxylate transporter 8 (MCT8). Endocrinology (2013) 154(7):2553–61. doi:10.1210/en.2012-2197
64. Friesema EC, Jansen J, Jachtenberg JW, Visser WE, Kester MH, Visser TJ. Effective cellular uptake and efflux of thyroid hormone by human monocarboxylate transporter 10. Mol Endocrinol (2008) 22(6):1357–69. doi:10.1210/me.2007-0112
Keywords: blood–brain-barrier, transthyretin, deiodinase, flavonoids, endocrine disruptors, Mct8, l-type amino acid transporter, organic anion transporters
Citation: Wirth EK, Schweizer U and Köhrle J (2014) Transport of thyroid hormone in brain. Front. Endocrinol. 5:98. doi: 10.3389/fendo.2014.00098
Received: 10 April 2014; Accepted: 07 June 2014;
Published online: 24 June 2014.
Edited by:
Noriyuki Koibuchi, Gunma University Graduate School of Medicine, JapanReviewed by:
Takashi Yoshimura, Nagoya University, JapanHeike Heuer, Leibniz Institute for Age Research-Fritz Lipmann Institute, Germany
Copyright: © 2014 Wirth, Schweizer and Köhrle. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Josef Köhrle, Institut für Experimentelle Endokrinologie, Charité-Universitätsmedizin Berlin, Augustenburger Platz 1, Berlin 13353, Germany e-mail:am9zZWYua29laHJsZUBjaGFyaXRlLmRl