 Paul J. Davis1,2*
Paul J. Davis1,2* Gennadi V. Glinsky3Hung-Yun Lin4John T. Leith5
Gennadi V. Glinsky3Hung-Yun Lin4John T. Leith5 Aleck Hercbergs6Heng-Yuan Tang2
Aleck Hercbergs6Heng-Yuan Tang2 Osnat Ashur-Fabian7,8
Osnat Ashur-Fabian7,8 Sandra Incerpi9Shaker A. Mousa2
Sandra Incerpi9Shaker A. Mousa2
- 1Department of Medicine, Albany Medical College, Albany, NY, USA
- 2Pharmaceutical Research Institute, Albany College of Pharmacy and Health Sciences, Rensselaer, NY, USA
- 3Stanford University, Palo Alto, CA, USA
- 4Taipei Medical University, Taipei, Taiwan
- 5Rhode Island Nuclear Science Center, Narragansett, RI, USA
- 6Cleveland Clinic, Cleveland, OH, USA
- 7Hematology Institute and Blood Bank, Meir Medical Center, Kfar-Saba, Israel
- 8Department of Medicine, Sackler Faculty of Medicine, Tel Aviv University, Tel Aviv, Israel
- 9Department of Sciences, University Roma Tre, Rome, Italy
Integrin αvβ3 is generously expressed by cancer cells and rapidly dividing endothelial cells. The principal ligands of the integrin are extracellular matrix proteins, but we have described a cell surface small molecule receptor on αvβ3 that specifically binds thyroid hormone and thyroid hormone analogs. From this receptor, thyroid hormone (l-thyroxine, T4; 3,5,3′-triiodo-l-thyronine, T3) and tetraiodothyroacetic acid (tetrac) regulate expression of specific genes by a mechanism that is initiated non-genomically. At the integrin, T4 and T3 at physiological concentrations are pro-angiogenic by multiple mechanisms that include gene expression, and T4 supports tumor cell proliferation. Tetrac blocks the transcriptional activities directed by T4 and T3 at αvβ3, but, independently of T4 and T3, tetrac modulates transcription of cancer cell genes that are important to cell survival pathways, control of the cell cycle, angiogenesis, apoptosis, cell export of chemotherapeutic agents, and repair of double-strand DNA breaks. We have covalently bound tetrac to a 200 nm biodegradable nanoparticle that prohibits cell entry of tetrac and limits its action to the hormone receptor on the extracellular domain of plasma membrane αvβ3. This reformulation has greater potency than unmodified tetrac at the integrin and affects a broader range of cancer-relevant genes. In addition to these actions on intra-cellular kinase-mediated regulation of gene expression, hormone analogs at αvβ3 have additional effects on intra-cellular protein-trafficking (cytosol compartment to nucleus), nucleoprotein phosphorylation, and generation of nuclear coactivator complexes that are relevant to traditional genomic actions of T3. Thus, previously unrecognized cell surface-initiated actions of thyroid hormone and tetrac formulations at αvβ3 offer opportunities to regulate angiogenesis and multiple aspects of cancer cell behavior.
Introduction
Integrins are heterodimeric structural proteins of the plasma membrane and are principally involved in cell–cell relationships in tissues and cell–extracellular matrix (ECM) protein interactions (1). The extracellular domain of an integrin binds specific ECM proteins and outside-in transmission of the occurrence of liganding results in the generation of specific signals by the intra-cellular domain of the integrin. These signals, usually involving various kinases, may result in cellular changes in actin (2, 3) and cell motility (4), modulate endocytosis (5), and affect transcription of specific genes (6–8).
Amply expressed by and activated in cancer cells, integrin αvβ3 interacts with ECM proteins, but has recently been shown to have a panel of specific receptors for non-protein, small molecule ligands (9). Among these are sites for the binding of thyroid hormone (10, 11), dihydrotestosterone (12), and resveratrol (13). The thyroid hormone receptor (TR) on αvβ3 has been well-studied (11, 14). What is now apparent is that this receptor has more complex and coherent effects on cancer-relevant gene expression than had been apparent in analyses of the impact of large molecule (protein) interactions with the integrin. The multiple genes whose expression is modulated from the extracellular domain of αvβ3 by thyroid hormone or its derivative, tetraiodothyroacetic acid (tetrac), relate to angiogenesis, cancer cell proliferation, metastasis, and cancer cell defense pathways (15). The latter include genes relevant to anti-apoptosis, anti-angiogenesis, chemoresistance (MDR1), and repair of double-strand DNA breaks induced by radiation. Within the cell, unmodified tetrac mimics certain actions of thyroid hormone. At the extracellular domain of αvβ3, in contrast, tetrac blocks binding of l-thyroxine (T4) and 3,5,3′-triiodo-l-thyronine (T3) – that is, it is a thyroid hormone antagonist. Covalent bonding of tetrac to a nanoparticle prevents cell entry of tetrac and, compared to unmodified tetrac, broadens the spectrum of defensive cancer cell genes whose expression can be desirably regulated from the integrin. This expanded panel includes pro-apoptotic genes and epidermal growth factor receptor (EGFR) gene (see subsequent sections). In addition, the potency of nanoparticulate tetrac as a thyroid hormone antagonist at αvβ3 is greater than that of unmodified tetrac. Thus, without entering the cancer or endothelial cell, thyroid hormone analogs non-genomically initiate important actions on tumor cell and blood vessel cell gene expression. In this review, we survey αvβ3-mediated effects of thyroid hormone and analogs on gene expression in human cancer cells, analyzed by RT-PCR. We also point out that, from its receptor on the integrin, thyroid hormone has adjunctive effects on nuclear receptors for thyroid hormone and for estrogen, regulating the state of phosphorylation or acetylation of such receptors and controlling the formation of complexes within the nucleus of coactivators and receptors.
Early Evidence That Thyroid Hormone Could Modulate Gene Expression from the Cell Exterior: Protooncogene Expression; Angiogenesis
Prior to the discovery of the plasma membrane receptor for thyroid hormone and hormone analogs on integrin αvβ3, agarose-T4 had been shown to regulate protooncogene expression (16, 17). Agarose-T4 is a prototypic nanoparticulate formulation of l-thyroxine in which T4 is covalently bound to a linear polysaccharide polymer; the product is excluded from the cell interior. The thyroid hormone effect on gene expression in these studies was mitogen-activated protein kinase (MAPK)-dependent and was reproduced in cells that lacked the nuclear TR.
Studied in the chick chorioallantoic membrane (CAM) model and also prior to recognition of the hormone receptor on αvβ3, T4 at physiological free concentrations and T3 at concentrations that were supraphysiologic were shown to increase vascularity threefold in 72 h (18). The degree of activity was comparable to that of fibroblast growth factor 2 (FGF2; bFGF). Agarose-T4 also reproduced the pro-angiogenic effect of thyroid hormone. The effects of unmodified thyroid hormone and of agarose-T4 on angiogenesis were found to be inhibited by tetrac, the hormone analog subsequently shown to block the iodothyronine receptor site on the cell surface. Pharmacologic inhibitors of MAPK (ERK1/2) and of protein kinase C also eliminated thyroid hormone-induced angiogenesis. RT-PCR studies revealed that the hormone-induced transcription of FGF2 within 6 h, and measurement of FGF2 protein in the medium showed increased release of the angiogenic factor. Thus, the promotion of vascular sprouting (19) and new vessel formation by thyroid hormone was attributable to initiation at the plasma membrane of a non-genomic effect culminating in expression of a specific vascular growth factor gene, manufacture of the gene product and release of the latter protein into the medium.
The cell surface receptor for thyroid hormone and tetrac was shortly thereafter defined on the extracellular domain of integrin αvβ3 and functionally described in the context of angiogenesis (10). Other thyroid hormone agonist analogs, such as GC-1 (20) and diiodothyropropionic acid (DITPA) (21) were also shown to be pro-angiogenic, and tetrac blocked the activity of these analogs at the integrin. However, the anti-angiogenic properties of tetrac expressed at the integrin extend beyond the blockade of binding of T4 and T3 to αvβ3. As discussed in the next section, tetrac or its reformulation as a nanoparticulate may affect expression of blood vessel-relevant genes beyond FGF2 independently of T4 and T3. Tetrac and Nanotetrac may also disrupt crosstalk between αvβ3 and adjacent receptors for other vascular growth factors, such as vascular endothelial growth factor (VEGF) and FGF2 (22), and platelet-derived growth factor (PDGF) (Shaker A. Mousa, unpublished observations). However, these effects on crosstalk are unrelated to gene transcription.
Tetrac, Nanotetrac, and Gene and microRNA Expression That is Relevant to Angiogenesis
As indicated above, unmodified tetrac is taken up by cells and expresses low-grade T4-like activity and may be converted to triiodothyroacetic acid (triac), which is also thyromimetic (23, 24). To limit the action of tetrac exclusively to integrin αvβ3, we covalently bonded tetrac to a nanoparticle of sufficient size (~200 nm) to preclude cell uptake of the complex (25), thus mimicking agarose-T4. The polymer we used was biodegradable poly(lactic-co-glycolic acid), in contrast to the physiologically inert agarose. The nanoparticulate formulation involved a stable ether bond of the outer ring hydroxyl group of tetrac to a 6-carbon linker and amide-bonding of the latter to PLGA (25). The amide bond was imbedded in the nanoparticle and thus not readily accessible to circulating or tissue peptidases. The resulting Nanotetrac indeed was restricted to the extracellular space and preserved the previously known actions of tetrac, but it was also found to have desirable additional biologic activities not previously obtained with tetrac.
Microarray studies of two human cancer cell lines showed that tetrac and Nanotetrac downregulated expression of VEGFA (26), the gene product of which is a principal inducer of the porous blood vessels associated with cancers (27). These effects are initiated at plasma membrane αvβ3. Tetrac and Nanotetrac also increased transcription of thrombospondin 1 (THBS1, TSP1). TSP1 protein is an endogenous suppressor of angiogenesis and is invariably suppressed in cancer cells. Nanotetrac also decreased expression of EGFR, the gene product of which mediates actions of EGF on angiogenesis. Tetrac lacked this action. Nanotetrac, but not tetrac, downregulates expression of NFκB via the integrin and NFκB de-activation is an anti-angiogenic target (28, 29). Finally, thyroid hormone may regulate transcription of the monomeric αv gene (30), but it is not known whether this action is initiated at the αvβ3 protein or requires the nuclear TR.
In recent studies of microRNA (miR), we have shown that Nanotetrac increases cellular abundance of miR-15A in breast cancer cells by 10-fold (31) and decreases miR-21 by 50%. miR-21 is pro-angiogenic in certain tumor cells (32) and miR-15A decreases angiogenesis by a VEGF-dependent mechanism (33).
Transcriptional mechanisms involved in the anti-angiogenic activity of Nanotetrac at αvβ3 are summarized in Table 1.
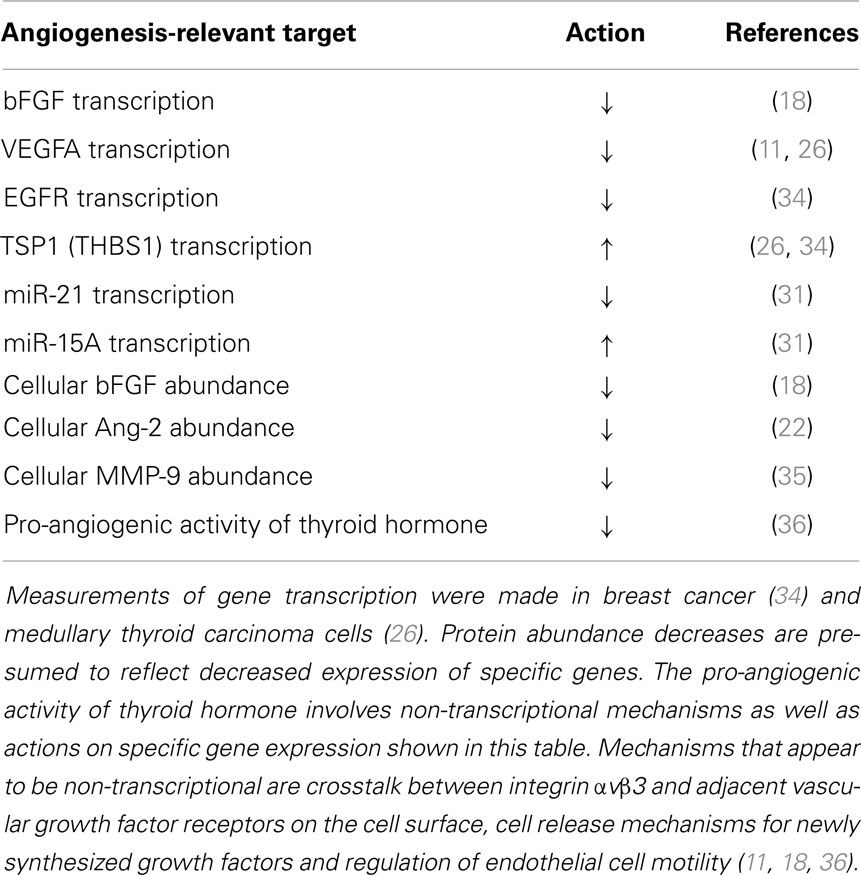
Table 1. Transcriptional mechanisms by which Nanotetrac/tetrac is anti-angiogenic.
Thyroid Hormone Supports Cancer Cell Proliferation and is Anti-Apoptotic; Tetrac–Nanotetrac Transcriptionally Inhibits Cancer Cell Proliferation, is Pro-Apoptotic and Disrupts Cell Defense Pathway Gene Expression
A number of laboratories have described the stimulatory effect of thyroid hormone on tumor cells (37–43) and clinical studies have defined thyroid hormone dependence of cancers, in that spontaneous or medically induced hypothyroidism has improved outcomes (44–49). Trophic actions of thyroid hormone on tumor cells were presumed to require a TR isoform and to be genomic in mechanism – that is, to require physical interaction of a TR protein and T3 – until recognition of the existence in 2005 (10) of the cell surface receptor for thyroid hormone and tetrac on αvβ3, described above (11, 50). Existence of this receptor offered a discrete, non-genomic mechanism for initiation of tumor cell proliferation. TRβ may be involved in certain cancer cell proliferative responses to thyroid hormone (51, 52), but work by Cheng and co-workers indicates that TRβ is a tumor suppressor that, when mutated in the thyroid gland, may be oncogenic (53).
The demonstration that T4 – including the agarose-T4 formulation – was a proliferative factor for certain human tumor (breast, thyroid cancer) (42, 54) and animal cells (C6, F98, GL261 glioma cell lines) (43) was accompanied by evidence that unmodified tetrac inhibited the T4 effect. We had shown that unmodified tetrac blocked non-genomic actions of thyroid hormone on plasma membrane functions (11, 14, 55). The proliferative effect was MAPK-dependent. Interestingly, in human breast cancer (MCF-7) cells, tetrac-inhibitable enhancement of proliferation by thyroid hormone involved Ser-118 phosphorylation of nuclear estrogen receptor-α (ERα); this pathway is identical to that by which estrogen stimulates MCF-7 cell proliferation (54).
In a model of resveratrol-induced apoptosis that involved MAPK phosphorylation of p53 at Ser-15, we showed that T4 was anti-apoptotic. The hormone prevented the p53 phosphorylation step in several tumor cell lines (42, 56). Tetrac blocked this anti-apoptotic activity of T4. Additional evidence of the anti-apoptotic activity of T4 included inhibition of nucleosome liberation by resveratrol, as well as cellular accumulation of the pro-apoptotic BcLxs protein (56, 57). The hormone did not, however, affect cell accumulation of survival protein BcLxl. The action of thyroid hormone on nucleosome liberation and BcLxs in tumor cells was prevented by tetrac (58).
Subsequent microarray studies conducted with Nanotetrac in human breast cancer (MDA-MB-231) cells revealed a coherent pro-apoptosis pattern of gene expression. That is, transcription of the X-linked inhibitor of apoptosis (XIAP) gene was downregulated and transcription of a set of pro-apoptotic genes – CASP2, CAP8AP2, DFFA, and BCL2L14 – was stimulated (11, 34).
We would also note that Nanotetrac downregulates expression of 8 of 9 cyclin genes and 1 cyclin-dependent kinase gene (34) and more than 20 oncogenes.
Tetrac–Nanotetrac Actions on Expression of Genes Relevant to Tumor Invasiveness
Catenins are proteins involved in cell–cell adhesion. β-catenin also has transcriptional functions in the nucleus. Mutation and overexpression of the β-catenin gene occurs in a variety of cancers, including colorectal carcinoma, breast, and ovarian cancer (59, 60). Nanotetrac increases transcription of the CBY1 gene (34), the gene product of which is an inhibitor of nuclear functions of β-catenins. This is a desirable action of Nanotetrac at αvβ3 in cancer cells. The action would be deleterious in non-cancer cells, but the latter when not undergoing cell division express little αvβ3. Like β-catenins, integrin αvβ3 participates in cellular adhesion complexes.
Nanotetrac also affects α-catenins, downregulating expression of the CTNNA1 and CTNNA2 genes. Mutation of CTNNA2 is associated with tumor invasiveness and thus inhibition of transcription of the gene is desirable, as is downregulation of the non-mutated gene in cancers. The non-mutated gene product of CTNNA1 can function as a tumor invasion suppressor (61), but mutation is associated with gastrointestinal tract and other cancers (62).
As mentioned above, MMP-9 expression is induced by thyroid hormone (35). The observations were recently made in myeloma cells and were inhibited by tetrac, thus implicating αvβ3 in this action of T4. This action of the hormone may contribute to local extension of myeloma in bone and, if documented to be present in solid tumor cells, may presage metastasis. MMP-2 transcription may also be subject to control by thyroid hormone (63, 64). Several mechanisms may be involved in the hormonal action on MMP-2, and it is not yet known whether this effect of the hormone is initiated at integrin αvβ3. The importance of this is that an intact metalloproteinase axis interferes with cell–cell interaction, resulting in tissue destabilization and support of cancer cell invasiveness and metastasis (65).
Overview of Anti-Cancer Properties of Nanotetrac, Acting as a Single Modality
The anti-cancer actions of Nanotetrac are broadly based in terms of mechanisms, despite initiation at a single target receptor on integrin αvβ3, and in this regard resemble the pluralistic anti-angiogenic actions of the drug. As noted above, the coherence of the effects of the agent on expression of differentially regulated cancer cell genes is remarkable. It is possible that there are effects of Nanotetrac at αvβ3 that may involve integrity of the actin cytoskeleton in cancer cells, and that the drug may influence interactions of the integrin with ECM proteins that may disorient tumor cell movement or interfere with defensive responses (see Conjunctive Radiation and Tetrac/Nanotetrac Treatment of Cancer Cells: Radiosensitization below). However, these possibilities have not yet been examined.
Nanotetrac promotes apoptosis, antagonizes anti-apoptotic (survival) defenses, disrupts control of the cell cycle, and interferes with function of the frequently mutated catenins (11, 26, 34). As noted above in the review of angiogenesis, thyroid hormone and tetrac or its Nanotetrac formulation affect matrix metalloproteinase gene expression. We would also note that thyroid hormone (T4) has protein-trafficking action on integrin αvβ3, directing internalization of the membrane protein – without the hormone ligand – and nuclear uptake of the αv monomer, but not of β3. In the nuclear compartment, αv is a coactivator (66) involved in transcription of a number of important cancer-relevant genes (see below, Adjunctive Modifications of Nuclear Hormone Receptors that Originate at the Hormone Receptor on αvβ3; Nuclear Uptake of αv Monomer).
Some of these actions of Nanotetrac/tetrac are summarized in Table 2.

Table 2. Mechanisms of selected cancer chemotherapeutic actions of Nanotetrac.
Chemosensitization by Tetrac of Cancer Cells Resistant to Other Cancer Chemotherapeutic Agents
P-glycoprotein (P-gp; MDR1; ABCB1) is a plasma membrane efflux pump whose ligands include a number of cancer chemotherapeutic agents (72). The pump is a principal component of cancer cell chemoresistance. Thyroid hormone causes transcription of MDR1 (73–75) and increases function of the P-gp protein (75). Thus, ambient thyroid hormone may be viewed as a support mechanism for chemoresistance (76). It is not known what the molecular basis is for regulation by iodothyronines of P-gp function or MDR1 gene expression, i.e., microarray studies have not been conducted to establish whether the induction of MDR1 gene expression is dependent upon the hormone receptor on integrin αvβ3. However, tetrac increases the intra-cellular retention time of doxorubicin by doxorubicin-resistant breast cancer cells (67), an effect attributed to action of tetrac–Nanotetrac on pump function of P-gp or on gene expression (76).
Conjunctive Radiation and Tetrac/Nanotetrac Treatment of Cancer Cells: Radiosensitization
Hercbergs and co-workers have defined the potentiation of radiation exposure by tetrac in animal glioma (C6) cells (68) and human glioblastoma (U87MG) cells (69), and Nanotetrac in human prostate cancer (PC3, LNCaP) cells (70). In vitro studies revealed that at a 4 Gy x-radiation dose 1 h after exposure to tetrac, there is a 60% reduction in cell survival, compared to control (68). The mechanism of action of tetrac and Nanotetrac is interference with cancer cell repair of double-strand DNA breaks (neutral comet assay/mean tail moment) (69). What components of the DNA break repair process – and, specifically, transcription of what specific genes – are affected by tetrac/Nanotetrac is not yet known.
Adjunctive Modifications of Nuclear Hormone Receptors That Originate at the Hormone Receptor on αvβ3; Nuclear Uptake of αv Monomer
The above discussion relates to regulation of transcription of specific cancer cell genes by thyroid hormone analogs that act at the cell surface via integrin αvβ3. Relevant additionally to the end result of modulation of transcription of specific genes from integrin αvβ3 is the adjunctive input from the integrin to the state of nuclear TRs. We have recently reviewed this subject (77). In brief, trafficking of cytoplasmic TRβ1 to the cell nucleus is directed by T4 at the integrin via MAPK, and the importing by the nucleus of TRα1 is promoted by T3 via activation of phosphatidylinositol 3-kinase (78). Two discrete binding domains exist at the TR site on αvβ3: the S1 site binds T3 exclusively and S2 binds both T4 and T3. Tetrac–Nanotetrac interferes with hormone binding at both domains. In the case of TRβ1 trafficking, translocation of the receptor into the nucleus occurs as a complex with activated MAPK; specific phosphorylation of the receptor (activation) is a consequence (79, 80). An example of specific gene transcription that occurs as a result of this trafficking/phosphorylation is expression of hypoxia-inducible factor-1α (HIF1α) in response to T3 at the S1 site (78). The complex process of stimulating cancer cell or endothelial cell proliferation occurs via the S2 domain.
Integrin αvβ3 may be internalized by cells as a result of the protein’s liganding of T4 (13). The αv monomer is imported by the nucleus as a result of this process and has been shown to be a coactivator protein that binds to the promoter region of a number of genes, including ERα, HIF-1α, cyclooxygenase-2 (COX-2), and TRβ1. ERα protein is important to breast, ovarian, and certain lung cancers. We have implicated nuclear COX-2 protein in the pharmacologic induction of apoptosis (58). HIF-1α protein is a cell survival factor that triggers angiogenesis and cellular conversion to anaerobic metabolism (81). The αv monomer does not import thyroid hormone and the β3 monomer is not taken up by the nucleus. This remarkable process was an unexpected consequence of studies of small molecule actions at the integrin and offers a novel mechanism for regulation of gene expression from the cell surface and integrin.
Conclusion
Integrin αvβ3 controls a variety of intra-cellular and transcellular functions. It is a transmembrane structural protein that is differentially expressed/activated in tumor cells and dividing blood vessel cells. The definition of the specific thyroid hormone-tetrac receptor site on αvβ3 (10, 11, 14) enabled recognition of the existence of control from a single locus of expression of differentially regulated, angiogenesis-relevant genes as well as modulation of function of adjacent vascular growth factor receptors. Nanotetrac is a prototypic anti-angiogenic and anti-cancer agent focused on a single, specific small molecule receptor site on the extracellular domain of αvβ3. From this site, Nanotetrac blocks actions of VEGF, FGF2, and PDGF at their plasma membrane receptors, inhibits expression of VEGFA and EGFR, stimulates transcription of TSP1, decreases endothelial cell abundance of Ang-2 without affecting Ang-1, selectively regulates miRNAs that control angiogenesis and decreases endothelial cell motility (Table 1).
From the standpoint of anti-cancer activity, Nanotetrac desirably disrupts gene expression critical to cell cycling in αvβ3-bearing tumor cells and dividing endothelial cells and interferes with a substantial group of cell survival pathways so that apoptosis is advanced, and defensive anti-apoptosis pathways are disordered (Table 2). Nanotetrac reverses chemoresistance and confers radiosensitivity. This novel and extensive spectrum of actions makes Nanotetrac an attractive anti-angiogenic and anti-cancer agent for further development. The agent has been shown to be an effective anti-proliferative, pro-apoptotic agent in a variety of human cancer cell lines (25, 31, 40–43, 71), to be effective against human cancer xenografts (26, 57, 70, 82–84) and to include important downregulation of tumor-associated angiogenesis (22, 26, 36, 57, 82, 84).
In the absence of an agent such as Nanotetrac with anti-thyroid hormone activity at integrin αvβ3, a reduction in circulating thyroid hormone, notably T4, that is either spontaneous or medically induced appears to be effective in slowing clinical growth of certain solid tumors. These include breast (45), glioblastoma multiforme (44), head-and-neck cancers (47), and renal cell carcinoma (46). We can postulate that such reductions in systemic levels of T4 largely affect tumors via the examples of gene expression reviewed above. Several of the current authors have recently confirmed clinically that systematic reduction in circulating T4 (euthyroid hypothyroxinemia) may arrest growth of certain cancers (85).
Conflict of Interest Statement
Paul J. Davis and Shaker A. Mousa are co-inventors of Nanotetrac. The issued and pending patents for Nanotetrac are held by NanoPharmaceuticals LLC, a company from which the inventors receive no reimbursement. The other co-authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Arnaout MA, Mahalingam B, Xiong JP. Integrin structure, allostery, and bidirectional signaling. Annu Rev Cell Dev Biol (2005) 21:381–410. doi: 10.1146/annurev.cellbio.21.090704.151217
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
2. Havaki S, Kouloukoussa M, Amawi K, Drosos Y, Arvanitis LD, Goutas N, et al. Altered expression pattern of integrin alphavbeta3 correlates with actin cytoskeleton in primary cultures of human breast cancer. Cancer Cell Int (2007) 7:16. doi:10.1186/1475-2867-7-16
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
3. Saltel F, Chabadel A, Bonnelye E, Jurdic P. Actin cytoskeletal organisation in osteoclasts: a model to decipher transmigration and matrix degradation. Eur J Cell Biol (2008) 87:459–68. doi:10.1016/j.ejcb.2008.01.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
4. Koutsioumpa M, Polytarchou C, Courty J, Zhang Y, Kieffer N, Mikelis C, et al. Interplay between alphavbeta3 integrin and nucleolin regulates human endothelial and glioma cell migration. J Biol Chem (2013) 288:343–54. doi:10.1074/jbc.M112.387076
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
5. Colin D, Limagne E, Jeanningros S, Jacquel A, Lizard G, Athias A, et al. Endocytosis of resveratrol via lipid rafts and activation of downstream signaling pathways in cancer cells. Cancer Prev Res (Phila) (2011) 4:1095–106. doi:10.1158/1940-6207.CAPR-10-0274
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
6. Reuning U. Integrin alphavbeta3 promotes vitronectin gene expression in human ovarian cancer cells by implicating rel transcription factors. J Cell Biochem (2011) 112:1909–19. doi:10.1002/jcb.23111
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
7. Umemoto T, Yamato M, Ishihara J, Shiratsuchi Y, Utsumi M, Morita Y, et al. Integrin-alphavbeta3 regulates thrombopoietin-mediated maintenance of hematopoietic stem cells. Blood (2012) 119:83–94. doi:10.1182/blood-2011-02-335430
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
8. Roth P, Silginer M, Goodman SL, Hasenbach K, Thies S, Maurer G, et al. Integrin control of the transforming growth factor-beta pathway in glioblastoma. Brain (2013) 136:564–76. doi:10.1093/brain/aws351
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
9. Davis PJ, Mousa SA, Cody V, Tang HY, Lin HY. Small molecule hormone or hormone-like ligands of integrin alphavbeta3: implications for cancer cell behavior. Horm Cancer (2013) 4:335–42. doi:10.1007/s12672-013-0156-8
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
10. Bergh JJ, Lin HY, Lansing L, Mohamed SN, Davis FB, Mousa S, et al. Integrin alphaVbeta3 contains a cell surface receptor site for thyroid hormone that is linked to activation of mitogen-activated protein kinase and induction of angiogenesis. Endocrinology (2005) 146:2864–71. doi:10.1210/en.2005-0102
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
11. Davis PJ, Davis FB, Mousa SA, Luidens MK, Lin HY. Membrane receptor for thyroid hormone: physiologic and pharmacologic implications. Annu Rev Pharmacol Toxicol (2011) 51:99–115. doi:10.1146/annurev-pharmtox-010510-100512
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
12. Lin HY, Sun M, Lin C, Tang HY, London D, Shih A, et al. Androgen-induced human breast cancer cell proliferation is mediated by discrete mechanisms in estrogen receptor-alpha-positive and -negative breast cancer cells. J Steroid Biochem Mol Biol (2009) 113:182–8. doi:10.1016/j.jsbmb.2008.12.010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
13. Lin HY, Lansing L, Merillon JM, Davis FB, Tang HY, Shih A, et al. Integrin alphaVbeta3 contains a receptor site for resveratrol. FASEB J (2006) 20:1742–4. doi:10.1096/fj.06-5743fje
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
14. Cheng SY, Leonard JL, Davis PJ. Molecular aspects of thyroid hormone actions. Endocr Rev (2010) 31:139–70. doi:10.1210/er.2009-0007
15. Davis PJ, Lin HY, Sudha T, Yalcin M, Tang HY, Hercbergs A, et al. Nanotetrac targets integrin alphavbeta3 on tumor cells to disorder cell defense pathways and block angiogenesis. Onco Targets Ther (2014) 7:1619–24. doi:10.2147/OTT.S67393
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
16. Lin HY, Shih A, Davis FB, Davis PJ. Thyroid hormone promotes the phosphorylation of STAT3 and potentiates the action of epidermal growth factor in cultured cells. Biochem J (1999) 338(Pt 2):427–32. doi:10.1042/0264-6021:3380427
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
17. Shih A, Lin HY, Davis FB, Davis PJ. Thyroid hormone promotes serine phosphorylation of p53 by mitogen-activated protein kinase. Biochemistry (2001) 40:2870–8. doi:10.1021/bi001978b
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
18. Davis FB, Mousa SA, O’Connor L, Mohamed S, Lin HY, Cao HJ, et al. Proangiogenic action of thyroid hormone is fibroblast growth factor-dependent and is initiated at the cell surface. Circ Res (2004) 94:1500–6. doi:10.1161/01.RES.0000130784.90237.4a
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
19. Mousa SA, Davis FB, Mohamed S, Davis PJ, Feng X. Pro-angiogenesis action of thyroid hormone and analogs in a three-dimensional in vitro microvascular endothelial sprouting model. Int Angiol (2006) 25:407–13.
20. Mousa SA, O’Connor LJ, Bergh JJ, Davis FB, Scanlan TS, Davis PJ. The proangiogenic action of thyroid hormone analogue GC-1 is initiated at an integrin. J Cardiovasc Pharmacol (2005) 46:356–60. doi:10.1097/01.fjc.0000175438.94906.a0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
21. Mousa SA, O’Connor L, Davis FB, Davis PJ. Proangiogenesis action of the thyroid hormone analog 3,5-diiodothyropropionic acid (DITPA) is initiated at the cell surface and is integrin mediated. Endocrinology (2006) 147:1602–7. doi:10.1210/en.2005-1390
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
22. Mousa SA, Bergh JJ, Dier E, Rebbaa A, O’Connor LJ, Yalcin M, et al. Tetraiodothyroacetic acid, a small molecule integrin ligand, blocks angiogenesis induced by vascular endothelial growth factor and basic fibroblast growth factor. Angiogenesis (2008) 11:183–90. doi:10.1007/s10456-007-9088-7
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
23. Moreno M, de Lange P, Lombardi A, Silvestri E, Lanni A, Goglia F. Metabolic effects of thyroid hormone derivatives. Thyroid (2008) 18:239–53. doi:10.1089/thy.2007.0248
24. Agrawal NK, Goyal R, Rastogi A, Naik D, Singh SK. Thyroid hormone resistance. Postgrad Med J (2008) 84:473–7. doi:10.1136/pgmj.2008.069740
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
25. Bharali DJ, Yalcin M, Davis PJ, Mousa SA. Tetraiodothyroacetic acid-conjugated PLGA nanoparticles: a nanomedicine approach to treat drug-resistant breast cancer. Nanomedicine (Lond) (2013) 8:1943–54. doi:10.2217/nnm.12.200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
26. Yalcin M, Dyskin E, Lansing L, Bharali DJ, Mousa SS, Bridoux A, et al. Tetraiodothyroacetic acid (tetrac) and nanoparticulate tetrac arrest growth of medullary carcinoma of the thyroid. J Clin Endocrinol Metab (2010) 95:1972–80. doi:10.1210/jc.2009-1926
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
27. Nagy JA, Dvorak AM, Dvorak HF. Vascular hyperpermeability, angiogenesis, and stroma generation. Cold Spring Harb Perspect Med (2012) 2:a006544. doi:10.1101/cshperspect.a006544
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
28. Palenski TL, Gurel Z, Sorenson CM, Hankenson KD, Sheibani N. Cyp1B1 expression promotes angiogenesis by suppressing NF-kappaB activity. Am J Physiol Cell Physiol (2013) 305:C1170–84. doi:10.1152/ajpcell.00139.2013
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
29. Omar HA, Arafa el SA, Salama SA, Arab HH, Wu CH, Weng JR. OSU-A9 inhibits angiogenesis in human umbilical vein endothelial cells via disrupting Akt-NF-kappaB and MAPK signaling pathways. Toxicol Appl Pharmacol (2013) 272:616–24. doi:10.1016/j.taap.2013.07.014
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
30. Pathak A, Sinha RA, Mohan V, Mitra K, Godbole MM. Maternal thyroid hormone before the onset of fetal thyroid function regulates reelin and downstream signaling cascade affecting neocortical neuronal migration. Cereb Cortex (2011) 21:11–21. doi:10.1093/cercor/bhq052
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
31. Mousa SA, Thangirala S, Lin HY, Tang HY, Glinsky GV, Davis PJ. MicroRNA-21 and microRNA-15A expression in human breast cancer (MDA-MB-231) cells exposed to nanoparticulate tetraiodothyroacetic acid (Nanotetrac). Meeting of the Endocrine Society. (2014). Abstract SUN-0472.
32. Zhao D, Tu Y, Wan L, Bu L, Huang T, Sun X, et al. In vivo monitoring of angiogenesis inhibition via down-regulation of mir-21 in a VEGFR2-luc murine breast cancer model using bioluminescent imaging. PLoS One (2013) 8:e71472. doi:10.1371/journal.pone.0071472
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
33. Sun CY, She XM, Qin Y, Chu ZB, Chen L, Ai LS, et al. miR-15a and miR-16 affect the angiogenesis of multiple myeloma by targeting VEGF. Carcinogenesis (2013) 34:426–35. doi:10.1093/carcin/bgs333
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
34. Glinskii AB, Glinsky GV, Lin HY, Tang HY, Sun M, Davis FB, et al. Modification of survival pathway gene expression in human breast cancer cells by tetraiodothyroacetic acid (tetrac). Cell Cycle (2009) 8:3562–70. doi:10.4161/cc.8.21.9963
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
35. Cohen K, Flint N, Shalev S, Erez D, Baharal T, Davis PJ, et al. Thyroid hormone regulates adhesion, migration and matrix metalloproteinase 9 activity via alphavbeta3 integrin in myeloma cells. Oncotarget (2014) 5:6312–22.
36. Mousa SA, Lin HY, Tang HY, Hercbergs A, Luidens MK, Davis PJ. Modulation of angiogenesis by thyroid hormone and hormone analogues: implications for cancer management. Angiogenesis (2014) 17:463–9. doi:10.1007/s10456-014-9418-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
37. Goodman AD, Hoekstra SJ, Marsh PS. Effects of hypothyroidism on the induction and growth of mammary cancer induced by 7,12-dimethylbenz(a)anthracene in the rat. Cancer Res (1980) 40:2336–42.
38. Borek C, Guernsey DL, Ong A, Edelman IS. Critical role played by thyroid hormone in induction of neoplastic transformation by chemical carcinogens in tissue culture. Proc Natl Acad Sci U S A (1983) 80:5749–52. doi:10.1073/pnas.80.18.5749
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
39. Hall LC, Salazar EP, Kane SR, Liu N. Effects of thyroid hormones on human breast cancer cell proliferation. J Steroid Biochem Mol Biol (2008) 109:57–66. doi:10.1016/j.jsbmb.2007.12.008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
40. Cohen K, Ellis M, Khoury S, Davis PJ, Hercbergs A, Ashur-Fabian O. Thyroid hormone is a MAPK-dependent growth factor for human myeloma cells acting via alphavbeta3 integrin. Mol Cancer Res (2011) 9:1385–94. doi:10.1158/1541-7786.MCR-11-0187
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
41. Meng R, Tang HY, Westfall J, London D, Cao JH, Mousa SA, et al. Crosstalk between integrin alphavbeta3 and estrogen receptor-alpha is involved in thyroid hormone-induced proliferation in human lung carcinoma cells. PLoS One (2011) 6:e27547. doi:10.1371/journal.pone.0027547
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
42. Lin HY, Tang HY, Shih A, Keating T, Cao G, Davis PJ, et al. Thyroid hormone is a MAPK-dependent growth factor for thyroid cancer cells and is anti-apoptotic. Steroids (2007) 72:180–7. doi:10.1016/j.steroids.2006.11.014
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
43. Davis FB, Tang HY, Shih A, Keating T, Lansing L, Hercbergs A, et al. Acting via a cell surface receptor, thyroid hormone is a growth factor for glioma cells. Cancer Res (2006) 66:7270–5. doi:10.1158/0008-5472.CAN-05-4365
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
44. Hercbergs AA, Goyal LK, Suh JH, Lee S, Reddy CA, Cohen BH, et al. Propylthiouracil-induced chemical hypothyroidism with high-dose tamoxifen prolongs survival in recurrent high grade glioma: a phase I/II study. Anticancer Res (2003) 23:617–26.
45. Cristofanilli M, Yamamura Y, Kau SW, Bevers T, Strom S, Patangan M, et al. Thyroid hormone and breast carcinoma. Primary hypothyroidism is associated with a reduced incidence of primary breast carcinoma. Cancer (2005) 103:1122–8. doi:10.1002/cncr.20881
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
46. Schmidinger M, Vogl UM, Bojic M, Lamm W, Heinzl H, Haitel A, et al. Hypothyroidism in patients with renal cell carcinoma: blessing or curse? Cancer (2011) 117:534–44. doi:10.1002/cncr.25422
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
47. Nelson M, Hercbergs A, Rybicki L, Strome M. Association between development of hypothyroidism and improved survival in patients with head and neck cancer. Arch Otolaryngol Head Neck Surg (2006) 132:1041–6. doi:10.1001/archotol.132.10.1041
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
48. Hercbergs AH, Ashur-Fabian O, Garfield D. Thyroid hormones and cancer: clinical studies of hypothyroidism in oncology. Curr Opin Endocrinol Diabetes Obes (2010) 17:432–6. doi:10.1097/MED.0b013e32833d9710
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
49. Mondul AM, Weinstein SJ, Bosworth T, Remaley AT, Virtamo J, Albanes D. Circulating thyroxine, thyroid-stimulating hormone, and hypothyroid status and the risk of prostate cancer. PLoS One (2012) 7:e47730. doi:10.1371/journal.pone.0047730
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
50. Lin HY, Cody V, Davis FB, Hercbergs AA, Luidens MK, Mousa SA, et al. Identification and functions of the plasma membrane receptor for thyroid hormone analogues. Discov Med (2011) 11:337–47.
51. Perri A, Catalano S, Bonofiglio D, Vizza D, Rovito D, Qi H, et al. T3 enhances thyroid cancer cell proliferation through TRbeta1/Oct-1-mediated cyclin D1 activation. Mol Cell Endocrinol (2014) 382:205–17. doi:10.1016/j.mce.2013.10.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
52. Wu SM, Cheng WL, Lin CD, Lin KH. Thyroid hormone actions in liver cancer. Cell Mol Life Sci (2013) 70:1915–36. doi:10.1007/s00018-012-1146-7
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
53. Kim WG, Cheng SY. Thyroid hormone receptors and cancer. Biochim Biophys Acta (2013) 1830:3928–36. doi:10.1016/j.bbagen.2012.04.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
54. Tang HY, Lin HY, Zhang S, Davis FB, Davis PJ. Thyroid hormone causes mitogen-activated protein kinase-dependent phosphorylation of the nuclear estrogen receptor. Endocrinology (2004) 145:3265–72. doi:10.1210/en.2004-0308
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
55. Lin HY, Tang HY, Davis FB, Mousa SA, Incerpi S, Luidens MK, et al. Nongenomic regulation by thyroid hormone of plasma membrane ion and small molecule pumps. Discov Med (2012) 14:199–206.
56. Lin HY, Tang HY, Keating T, Wu YH, Shih A, Hammond D, et al. Resveratrol is pro-apoptotic and thyroid hormone is anti-apoptotic in glioma cells: both actions are integrin and ERK mediated. Carcinogenesis (2008) 29:62–9. doi:10.1093/carcin/bgm239
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
57. Yalcin M, Lin HY, Sudha T, Bharali DJ, Meng R, Tang HY, et al. Response of human pancreatic cancer cell xenografts to tetraiodothyroacetic acid nanoparticles. Horm Cancer (2013) 4:176–85. doi:10.1007/s12672-013-0137-y
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
58. Lin HY, Davis PJ, Tang HY, Mousa SA, Luidens MK, Hercbergs AH, et al. The pro-apoptotic action of stilbene-induced COX-2 in cancer cells: convergence with the anti-apoptotic effect of thyroid hormone. Cell Cycle (2009) 8:1877–82. doi:10.4161/cc.8.12.8747
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
59. King TD, Suto MJ, Li Y. The Wnt/beta-catenin signaling pathway: a potential therapeutic target in the treatment of triple negative breast cancer. J Cell Biochem (2012) 113:13–8. doi:10.1002/jcb.23350
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
60. White BD, Chien AJ, Dawson DW. Dysregulation of Wnt/beta-catenin signaling in gastrointestinal cancers. Gastroenterology (2012) 142:219–32. doi:10.1053/j.gastro.2011.12.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
61. Vermeulen SJ, Nollet F, Teugels E, Vennekens KM, Malfait F, Philippe J, et al. The alphaE-catenin gene (CTNNA1) acts as an invasion-suppressor gene in human colon cancer cells. Oncogene (1999) 18:905–15. doi:10.1038/sj.onc.1202348
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
62. Debruyne P, Vermeulen S, Mareel M. The role of the E-cadherin/catenin complex in gastrointestinal cancer. Acta Gastroenterol Belg (1999) 62:393–402.
63. Huang YH, Lin YH, Chi HC, Liao CH, Liao CJ, Wu SM, et al. Thyroid hormone regulation of miR-21 enhances migration and invasion of hepatoma. Cancer Res (2013) 73:2505–17. doi:10.1158/0008-5472.CAN-12-2218
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
64. Chen RN, Huang YH, Lin YC, Yeh CT, Liang Y, Chen SL, et al. Thyroid hormone promotes cell invasion through activation of furin expression in human hepatoma cell lines. Endocrinology (2008) 149:3817–31. doi:10.1210/en.2007-0989
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
65. Hadler-Olsen E, Winberg JO, Uhlin-Hansen L. Matrix metalloproteinases in cancer: their value as diagnostic and prognostic markers and therapeutic targets. Tumour Biol (2013) 34:2041–51. doi:10.1007/s13277-013-0842-8
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
66. Lin HY, Su YF, Hsieh MT, Lin S, Meng R, London D, et al. Nuclear monomeric integrin alphav in cancer cells is a coactivator regulated by thyroid hormone. FASEB J (2013) 27:3209–16. doi:10.1096/fj.12-227132
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
67. Rebbaa A, Chu F, Davis FB, Davis PJ, Mousa SA. Novel function of the thyroid hormone analog tetraiodothyroacetic acid: a cancer chemosensitizing and anti-cancer agent. Angiogenesis (2008) 11:269–76. doi:10.1007/s10456-008-9110-8
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
68. Hercbergs A, Davis PJ, Davis FB, Ciesielski MJ, Leith JT. Radiosensitization of GL261 glioma cells by tetraiodothyroacetic acid (tetrac). Cell Cycle (2009) 8:2586–91. doi:10.4161/cc.8.16.9328
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
69. Hercbergs AH, Lin HY, Davis FB, Davis PJ, Leith JT. Radiosensitization and production of DNA double-strand breaks in U87MG brain tumor cells induced by tetraiodothyroacetic acid (tetrac). Cell Cycle (2011) 10:352–7. doi:10.4161/cc.10.2.14641
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
70. Mousa SA, Sudha T, Yalcin M, Leith JT, Hercbergs A, Lin HY, et al. Actions of nanoparticulate tetraiodothyroacetic acid (Nanotetrac) on human prostate carcinoma xenograft growth, vascularity and integrin response to radiation. Annual Meeting of the American Association for Cancer Research. San Diego, CA (2014). Abstract 5378.
71. Lin HY, Landersdorfer CB, London D, Meng R, Lim CU, Lin C, et al. Pharmacodynamic modeling of anti-cancer activity of tetraiodothyroacetic acid in a perfused cell culture system. PLoS Comput Biol (2011) 7(2):e1001073. doi:10.1371/journal.pcbi.1001073
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
72. Zinzi L, Capparelli E, Cantore M, Contino M, Leopoldo M, Colabufo NA. Small and innovative molecules as new strategy to revert MDR. Front Oncol (2014) 4:2. doi:10.3389/fonc.2014.00002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
73. Nishio N, Katsura T, Inui K. Thyroid hormone regulates the expression and function of P-glycoprotein in Caco-2 cells. Pharm Res (2008) 25:1037–42. doi:10.1007/s11095-007-9495-x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
74. Kurose K, Saeki M, Tohkin M, Hasegawa R. Thyroid hormone receptor mediates human MDR1 gene expression-Identification of the response region essential for gene expression. Arch Biochem Biophys (2008) 474:82–90. doi:10.1016/j.abb.2008.03.020
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
75. Burk O, Brenner SS, Hofmann U, Tegude H, Igel S, Schwab M, et al. The impact of thyroid disease on the regulation, expression, and function of ABCB1 (MDR1/P glycoprotein) and consequences for the disposition of digoxin. Clin Pharmacol Ther (2010) 88:685–94. doi:10.1038/clpt.2010.176
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
76. Davis PJ, Incerpi S, Lin HY, Tang HY, Sudha T, Mousa SA. Thyroid hormone and P-glycoprotein in tumor cells. Biomed Res Int (2014) 2014:168427.
77. Davis PJ, Lin HY, Tang HY, Davis FB, Mousa SA. Adjunctive input to the nuclear thyroid hormone receptor from the cell surface receptor for the hormone. Thyroid (2013) 23:1503–9. doi:10.1089/thy.2013.0280
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
78. Lin HY, Sun M, Tang HY, Lin C, Luidens MK, Mousa SA, et al. L-Thyroxine vs. 3,5,3’-triiodo-L-thyronine and cell proliferation: activation of mitogen-activated protein kinase and phosphatidylinositol 3-kinase. Am J Physiol Cell Physiol (2009) 296:C980–91. doi:10.1152/ajpcell.00305.2008
79. Davis PJ, Shih A, Lin HY, Martino LJ, Davis FB. Thyroxine promotes association of mitogen-activated protein kinase and nuclear thyroid hormone receptor (TR) and causes serine phosphorylation of TR. J Biol Chem (2000) 275:38032–9. doi:10.1074/jbc.M002560200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
80. Lin HY, Zhang S, West BL, Tang HY, Passaretti T, Davis FB, et al. Identification of the putative MAP kinase docking site in the thyroid hormone receptor-beta1 DNA-binding domain: functional consequences of mutations at the docking site. Biochemistry (2003) 42:7571–9. doi:10.1021/bi0273967
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
81. Koh MY, Spivak-Kroizman TR, Powis G. HIF-1alpha and cancer therapy. Recent Results Cancer Res (2010) 180:15–34. doi:10.1007/978-3-540-78281-0_3
82. Mousa SA, Yalcin M, Bharali DJ, Meng R, Tang HY, Lin HY, et al. Tetraiodothyroacetic acid and its nanoformulation inhibit thyroid hormone stimulation of non-small cell lung cancer cells in vitro and growth in xenografts. Lung Cancer (2012) 76:39–45. doi:10.1016/j.lungcan.2011.10.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
83. Yalcin M, Bharali DJ, Dyskin E, Dier E, Lansing L, Mousa SS, et al. Tetraiodothyroacetic acid and tetraiodothyroacetic acid nanoparticle effectively inhibit the growth of human follicular thyroid cell carcinoma. Thyroid (2010) 20:281–6. doi:10.1089/thy.2009.0249
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
84. Yalcin M, Bharali DJ, Lansing L, Dyskin E, Mousa SS, Hercbergs A, et al. Tetraiodothyroacetic acid and tetrac nanoparticles inhibit growth of human renal cell carcinoma xenografts. Anticancer Res (2009) 29:3825–31.
85. Hercbergs A, Johnson RE, Ashur-Fabian O, Garfield DH, Davis PJ. Medically induced euthyroid hypothyroxinemia may extend survival in compassionate need cancer patients: an observational study. Oncologist (2014). doi:10.1634/theoncologist.2014-0308
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: integrin, thyroid hormone, tetraiodothyroacetic acid, nanoparticle, gene transcription
Citation: Davis PJ, Glinsky GV, Lin H-Y, Leith JT, Hercbergs A, Tang H-Y, Ashur-Fabian O, Incerpi S and Mousa SA (2015) Cancer cell gene expression modulated from plasma membrane integrin αvβ3 by thyroid hormone and nanoparticulate tetrac. Front. Endocrinol. 5:240. doi: 10.3389/fendo.2014.00240
Received: 10 November 2014; Paper pending published: 25 November 2014;
Accepted: 19 December 2014; Published online: 12 January 2015.
Edited by:
Wen Zhou, Columbia University, USAReviewed by:
Jean-Yves Scoazec, Université Lyon 1, FranceYun Zhu, Medical University of South Carolina, USA
Fei Teng, University of Arizona, USA
Copyright: © 2015 Davis, Glinsky, Lin, Leith, Hercbergs, Tang, Ashur-Fabian, Incerpi and Mousa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paul J. Davis, Pharmaceutical Research Institute, Albany College of Pharmacy and Health Sciences, One Discovery Drive, Rensselaer, NY 12144, USA e-mail:cGRhdmlzLm9yZHdheXN0QGdtYWlsLmNvbQ==