 Jordi Altirriba
Jordi Altirriba Anne-Laure Poher
Anne-Laure Poher Françoise Rohner-Jeanrenaud
Françoise Rohner-Jeanrenaud- Laboratory of Metabolism, Department of Internal Medicine Specialties, Faculty of Medicine, University of Geneva, Geneva, Switzerland
This review summarizes the existing literature on the effects of oxytocin administration in the treatment of obesity in different animal models and in humans, focusing on the central control of food intake, the oxytocin effects on adipose tissue, and the relationships between oxytocin and leptin. Oxytocin is a hypothalamic nonapeptide synthesized mainly in the paraventricular and supraoptic nuclei projecting to the pituitary, where it reaches the peripheral circulation, as well as to other brain regions. Moreover, leptin modulates oxytocin levels and activates oxytocin neurons in the hypothalamic paraventricular nucleus, which innervates the nucleus of the solitary tract, partly responsible for the brain-elicited oxytocin effects. Taking into account that oxytocin is located downstream leptin, it was hypothesized that oxytocin treatment would be effective in decreasing body weight in leptin-resistant DIO animals, as well as in those with leptin or with leptin receptor deficiency. Several groups have demonstrated that in such animal models (rats, mice, and rhesus monkeys), central or peripheral oxytocin administration decreases body weight, mainly due to a decrease in fat mass, demonstrating that an oxytocin treatment is able to partly overcome leptin deficiency or resistance. Moreover, a pilot clinical study demonstrated the efficiency of oxytocin in the treatment of obesity in human subjects, confirming the results obtained in the different animal models. Larger multicenter studies are now needed to determine whether the beneficial effects of oxytocin treatment can apply not only to obese but also to type 2 diabetic patients. These studies should also shed some light on the molecular mechanisms of oxytocin action in humans.
1. Introduction
According to the World Health Organization, 2.8 million people die worldwide due to overweight or obesity, which represents one of the five leading global risks for mortality (1). Aging is another situation that contributes to the development of overweight or obesity. Indeed, it is well known that the body mass index tends to increase throughout adulthood, to level off from the age of 50 onward (2). Moreover, the presence of overweight or obesity in young and middle age was reported to result in adverse consequences for health care costs in older age (3).
Overall, the key component of the obesity epidemic is long-term dysregulation of energy balance comprising increased energy intake and/or reduced energy expenditure. Despite active research and impressive improvements in the understanding of the regulation of energy balance, there are only a very limited number of drugs that can be used for the efficient treatment of obesity and its comorbidities. Given the well-known state of leptin resistance, present in the vast majority of overweight/obese subjects, together with the fact that leptin exerts a whole array of beneficial effects on body weight homeostasis, one of the common approaches is to identify leptin targets that are able to mimic leptin’s effects, thereby bypassing leptin resistance. Along this line, oxytocin was recently considered as a potential interesting candidate, although it was historically recognized for its role in parturition and lactation (4). The aim of this review is to summarize the existing literature demonstrating the effects of oxytocin on the feeding behavior and peripheral metabolism, as well as to describe the interactions between oxytocin and the leptin signaling pathway.
Virtually, all vertebrate species were found to have an oxytocin-like nonapeptide that supports reproductive functions (5, 6). This impressive evolutionary conservation illustrates the significance of oxytocin in the survival of species. In mammals, oxytocin is synthesized mainly in the central nervous system, within both the magnocellular and parvocellular neurons of the paraventricular nucleus (PVN), as well as by the magnocellular neurons of the supraoptic nucleus (SON) in the hypothalamus. Unlike magnocellular oxytocin neurons, which project to the neurohypophysis (posterior pituitary), where they secrete oxytocin into the circulation, parvocellular oxytocin neurons of the PVN project centrally to various brain areas, including the arcuate nucleus of the hypothalamus (ARC) (7), the ventral tegmental area, the nucleus of the solitary tract (NTS), and the spinal cord (5, 6). Oxytocin release also occurs locally in the SON and PVN via somatodendritics. Several peripheral organs, such as the ovary, uterus, placenta, testis, thymus, kidney, heart, blood vessels, skin, and the gastrointestinal (GI) tract also synthesize oxytocin, although to a lesser extent than hypothalamic neurons [for reviews, see Ref. (5, 6, 8)]. To date, only one oxytocin receptor (OXTR) type has been identified. Its wide distribution within the brain (e.g., basal ganglia, hypothalamus, nuclei of the hindbrain), as well as in various peripheral tissues (e.g., heart, thymus, pancreas, adipocytes, and GI tract) (5) is in keeping with its large variety of physiological actions [for review, see Ref. (5)].
In 1989, Arletti et al. described that oxytocin administration (intraperitoneally or intracerebroventricularly) reduced food intake in rats, due to both a reduced meal size and an increased latency to feeding (9). Oxytocin is now well recognized as an anorexigenic neuropeptide, which effects are mediated by reducing gastric emptying and GI transit, as well as by suppressing the feeding reward circuit (4, 10). Furthermore, oxytocin and OXTR knockout mice were shown to develop late-onset obesity (11, 12), without alterations in food intake, pointing to the fact that oxytocin is controlling metabolic homeostasis, not only via an effect on food intake but also by modulating energy expenditure (11, 13) [for review, see Ref. (4)]. In keeping with the roles of oxytocin described in rodents (see below), patients with Prader–Willi syndrome (whose main characteristics are cognitive disabilities, chronic food craving, and morbid obesity) seem to present a deficit in the oxytocin producing neurons of the PVN (14), probably due to a loss of function or deletion of SIM1, a transcription factor controlling the expression of the OXT gene (15, 16).
Summary
Oxytocin can be considered as an anorexigenic peptide. Lack of oxytocin leads to the development of obesity in rodents, as well as in humans. This is partly independent of its effect on food intake, suggesting an additional role of oxytocin in the regulation of metabolic homeostasis.
2. Central Oxytocin Circuits and Interaction with Leptin in the Control of Food Intake
Different oxytocin neuron populations may fulfill different functions. Their interactions with leptin also appear to be specific for some distinct neurons. Close examination of the literature in the field allows to unravel the importance of precise leptin–oxytocin circuits in the regulation of food intake.
2.1. PVN to NTS Oxytocin Circuit
Accumulating evidence suggests that oxytocin neurons in the PVN mediate the anorexigenic effect of leptin (Figure 1). Thus, the oxytocin synthesizing neurons in the PVN exhibit a strong expression of the leptin receptor (17). Central leptin administration was shown to activate parvocellular oxytocin neurons of the PVN, which project to the NTS (18–20), where they are known to innervate POMC/CART neurons (21, 22). The low Oxt gene expression observed in the PVN during fasting was recovered after peripheral leptin administration (20, 23). Additionally, the leptin effect in decreasing food intake was partially blunted in adult mice with ablation of oxytocin neurons (24) and it was prevented by the injection of an OXTR antagonist within the fourth ventricle (19) (close to the NTS). Finally, the involvement of oxytocin neurons in leptin action was substantiated in mice with deletion of Socs3 (an inhibitor of the Jak-Stat leptin signaling pathway) in the mediobasal hypothalamus (25), which includes the PVN. Indeed, as a result of improved leptin signaling, these mice exhibited decreased food intake and body weight. Interestingly, the oxytocin content of the dorsal vagal complex (including the NTS) was increased in response to leptin administration in these mice, and the leptin-induced decrease in food intake was abolished by injection of an OXTR antagonist into the NTS (25). Collectively, these data suggest that oxytocin neurons of the PVN could mediate the leptin-activated hindbrain (NTS)-containing satiety circuit.
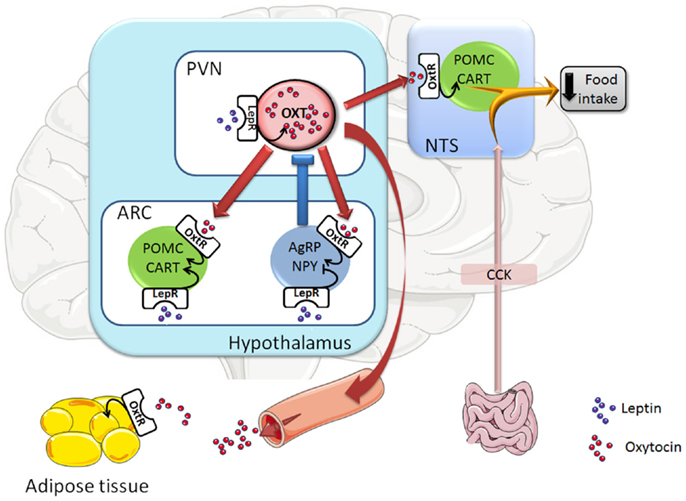
Figure 1. Representative scheme of the oxytocin neuronal circuits controlling food intake. Leptin, through the leptin receptor (LepR), activates oxytocin (Oxt) neurons in the hypothalamic paraventricular nucleus (PVN) and POMC/CART neurons in the hypothalamic arcuate nucleus (ARC), whereas it inhibits AgRP/NPY neurons in the ARC. Once oxytocin neurons are activated, they are innervating and activating POMC/CART neurons in the hypothalamic ARC nucleus and the hindbrain NTS nucleus (which is receiving also innervation from the gut), leading to a decrease in food intake. Moreover, activation of oxytocin neurons also triggers a negative feedback loop as PVN oxytocin neurons innervate AgRP/NPY neurons in the ARC nucleus, which inhibit oxytocin neurons in the PVN. At last, oxytocin neurons innervate the posterior pituitary and their activation leads to the secretion of oxytocin into the circulation, a way by which oxytocin can reach its target tissues, such as white adipose tissue. Arrow-headed lines and bar-headed lines indicate activation and inhibition, respectively. Oxytocin neurons and innervations are drawn in red, POMC/CART neurons in green, AgRP/NPY neurons in blue, leptin protein in purple circles, and oxytocin protein in red circles. Abbreviations: Oxt, oxytocin; POMC, pro-opiomelanocortin; CART, cocaine- and amphetamine-regulated transcript; AgRP, agouti-related protein; NPY, neuropeptide Y; CCK, cholecystokinin; LepR, leptin receptor; OxtR, oxytocin receptor; ARC, arcuate nucleus; PVN, paraventricular nucleus; NTS, nucleus of the solitary tract.
One of the ways by which leptin was reported to decrease food intake is by enhancing the efficiency of gut-derived anorexigenic hormones, such as cholecystokinin (CCK) (26). Thus, in line with a role of oxytocin as a mediator of leptin’s effect on food intake, it was shown that the satiety effect of CCK is partly blunted by the central injection of an OXTR antagonist or by destruction of the OXTR expressing neurons in the NTS (27–29). In contrast, other data obtained in oxytocin-deficient mice showed that oxytocin is dispensable for feeding inhibition by CCK (24, 30). A possible explanation that could reconcile both types of results (OXTR antagonist and OXTR cell ablation vs. oxytocin knockout) is that the oxytocin antagonist used (d(CH2)5[Tyr(Me)2,Thr4,Tyr-NH2(9)] OVT) (27–29) is an inverse agonist, which would not only block the oxytocin signal but also inhibit the neurons, which express the OXTR. It is also possible that, in oxytocin knockout mice, compensatory mechanisms may offset the absence of oxytocin. Further studies are, however, needed to clarify the role of oxytocin on the anorexigenic signal from gut-derived hormones.
2.2. PVN to ARC Oxytocin Circuit
In addition to PVN oxytocin neurons projecting to the NTS, other oxytocin neurons in the PVN also innervate POMC and AgRP neurons in the ARC (7). Moreover, it was reported that AgRP neurons target and inhibit oxytocin neurons (31), constituting a short feedback loop system in the hypothalamic control of food intake. It should be mentioned, however, that intra-ARC oxytocin administration decreased food intake to a lesser extent than lateral ventricle injection, suggesting that ARC activation only partially mediates the anorexigenic effect of oxytocin (7).
2.3. SON Oxytocin Neurons
With regard to the oxytocin neurons in the SON, they also display a high expression of the leptin receptor (17). However, the leptin’s effects on oxytocin in this nucleus are not clear. Thus, icv leptin administration did not modify the Oxt gene expression in SON neurons (32), neither did it activate OXT neurons in this nucleus, as assessed by the measurement of c-fos expression (19). In addition, leptin was shown to exert either inhibitory or stimulatory effects on the electrical activity of SON oxytocin neurons as studied in brain slices, or in vivo (i.v. administration in rats) (33, 34) respectively. Further studies are required to understand the role of oxytocin in the SON, although SON oxytocin neurons do not appear to be involved in the control of food intake.
Summary
It appears that among the hypothalamic oxytocin neurons involved in the control of food intake, those projecting from the PVN to the NTS are the most important ones in mediating the leptin’s effect.
3. Oxytocin Treatment in Animal Models of Obesity and Diabetes: A Way to Overcome Leptin Resistance?
Leptin administration to lean mice and rats leads to decreased food intake and body weight, effects that are blunted in obese rodents (35, 36). Moreover, diet-induced obese rodents are well known to display high-circulating leptin levels, reflecting their increased fat mass. Together, these data imply that obesity is associated with a state of leptin resistance (37). Taking into account that oxytocin is located downstream of leptin, mediating part of its effects, it would be expected that oxytocin treatment would be effective in decreasing body weight in leptin-resistant DIO animals, as well as in those with leptin or with leptin receptor deficiency.
3.1. Diet-Induced Obese Rodents
Recent data demonstrate that oxytocin treatment (subcutaneous or intracerebroventricular) of DIO mice, rats and rhesus monkeys was able to decrease body weight, mainly due to a decrease in fat mass, with a concomitant improvement in glucose tolerance and insulin sensitivity (38–42). Additional results obtained using a pair-feeding paradigm or the use of low oxytocin doses (which did not alter food intake) allowed concluding that at least part of the body weight-lowering effect of oxytocin was independent from changes in food intake (38, 39). It is noteworthy at that point that the OXTR is highly expressed in adipose tissue [at a similar level as in most of the classical oxytocin target tissues (43, 44)] and is upregulated in some mice models of obesity (44, 45). This could explain the occurrence of oxytocin effects on adipose tissue, as well as the difference between the effects of the oxytocin treatment in lean and obese animals. Indeed, it was observed that chronic oxytocin administration to lean mice was almost without any effect after the first day of treatment (during which a decrease in food intake and body weight was noted) (44).
3.2. Rodents with Leptin Receptor Deficiency or with Impaired Leptin Receptor Signaling
As alluded to above, oxytocin administration was also successful in treating obesity in animal models of leptin deficiency or reduced leptin receptor signaling [Zucker rats (21), Koletsky rats (41), ob/ob mice (44), and db/db mice (22, 46)]. Thus, 2 weeks of subcutaneous oxytocin infusion to ob/ob mice decreased body weight gain, mainly due to an effect on the fat mass. This was accompanied by a decrease in food intake, which was mainly observed during the first week of the treatment. Surprisingly, glucose tolerance of oxytocin-treated ob/ob mice was worsened, probably due to an increased activity of the hypothalamic–pituitary–adrenal axis, which led to increased corticosterone levels and enhanced hepatic gluconeogenesis (44). Interestingly, longer oxytocin treatment (12 weeks) in db/db mice also induced a decrease in body weight gain with a decrease in the fat mass, without any change in food intake. In contrast to ob/ob animals, oxytocin-treated db/db mice exhibited improved glucose tolerance and insulin sensitivity (46).
3.3. Oxytocin Treatment in Other Rodent Animal Models
Oxytocin treatment was also demonstrated to exert beneficial effects in other animal models, improving glucose metabolism in a diabetic model of pancreatic beta cell ablation by streptozotocin (47) and decreasing fat mass gain in ovarectomized mice (48).
Summary
Oxytocin treatment is able to overcome leptin resistance or leptin deficiency as it decreases body weight and fat mass in DIO animals, as well as in rodents with leptin deficiency or reduced leptin receptor signaling. In most experimental conditions, glucose metabolism is improved by the oxytocin treatment, except during short-term experiments in leptin-deficient mice.
4. Adipose Tissue as the Main Target of Oxytocin Action in Obesity
Adipose tissue seems to be one of the main targets of oxytocin in obesity, as the decrease in fat mass, in response to an in vivo oxytocin treatment, was observed in all the different models of rodent obesity studied so far (38, 39, 44, 46). However, considering data obtained in vitro, ex vivo, or in vivo during acute or chronic experiments, the reported results are contradictory.
4.1. In Vitro
In ex vivo experiments with adipose tissue or in cell cultures, oxytocin was without any effect on lipid metabolism in rabbit adipose tissue (49), whereas it reportedly increased lipogenesis (50, 51) or lipolysis (38) in rat adipocytes. It was also demonstrated that oxytocin inhibits adipocyte differentiation (52), increases glucose oxidation, and stimulates pyruvate dehydrogenase activity (53).
4.2. In Vivo
In acute in vivo experiments, oxytocin administration resulted in highly variable changes in non-esterified fatty acid (NEFA) and triglyceride (TG) levels, according to the experimental conditions used (e.g., sex, timing, feeding conditions, etc.) (54). Another acute study indicated that oxytocin increased circulating NEFA levels in dehydrated, but not in normally hydrated rats (55).
In chronic in vivo studies, it was shown that 15 days of treatment with oxytocin enhanced lipolysis in DIO rats (38) and promoted futile cycling (increase in both lipolysis and glycerogenesis) in ob/ob mice (44). In lean animals, such a chronic treatment increased adipogenesis in rats (56) and was without any effect in mice (44). Interestingly, a much longer treatment (12 weeks) was reported to decrease the size of adipocytes, the mass of fat pads, as well as to induce browning of white adipose tissue (i.e., presence of brown or beige/brite adipocytes in white adipose depots) in lean and obese db/db mice (46).
In humans, while acute oxytocin injection produced an increase in NEFA levels a few days after delivery (57), it resulted in decreased NEFA levels in non-pregnant healthy subjects (58). Furthermore, chronic treatment of obese humans with nasal oxytocin delivery did not significantly modify the TG levels, although there was some trend toward a decrease in this parameter (47).
4.3. Possible Explanations for the Contrary Results Obtained in Different Experimental Setups
As detailed above, although in vivo oxytocin treatment appears to consistently reduce fat mass in several models of obesity, discrepant results pertaining to oxytocin effects on glucose metabolism were observed. Concerning oxytocin effects on adipose tissue, conflicting results were reported depending on the experimental approach used (in vitro, ex vivo, in vivo).
One of the main reasons underlying the description of discrepant results is the use of different models, doses, and durations of the oxytocin treatments. Alternatively, as suggested by Muchmore and colleagues (59), the contradictory data could arise from the existence of low- and high-affinity OXTRs with opposite effects in response to their activation. Thus, low-oxytocin levels would exert insulin-like activities through interaction with high-affinity OXTRs, while at high concentrations, oxytocin would inhibit glycogen deposition (likely through an activation of glycogenolysis) and induce lipolysis, through its action on the low-affinity receptors (59). It should be mentioned at that point that, although the hypothesis of dose-dependent varying affinity of oxytocin for its receptor is interesting, it was postulated more than 30 years ago using an in vitro approach and was not validated further since then.
Another important variable, which could influence the in vivo oxytocin effects, is the type of diet consumed. Indeed, by modulating the membrane composition (60), it is conceivable that the diet may modulate the affinity of oxytocin for its receptor. This was actually demonstrated in the presence of divalent cations (such as magnesium) or cholesterol (5) and could occur when animals are fed a high-fat diet to induce obesity, glucose intolerance, and insulin resistance.
Differences in the expression of the OXTR (44, 45), as well as in oxytocinase activity in adipose tissue (61) could also modulate the type of oxytocin effects on glucose or lipid metabolism.
At last, it has been demonstrated that oxytocin neurons innervate white adipose tissue through the sympathetic nervous system (62) and oxytocin knockout mice display a decreased sympathetic tone (12). Altogether, oxytocin may therefore influence adipose tissue metabolism both by direct and indirect action on this tissue via the sympathetic nervous system. Therefore, the possible different sympathetic tone in the various models used could potentially also explain some of the divergent results obtained.
Summary
Although the data in non-obese animals and in in vitro experiments bring about some contradictory results, it is clear that oxytocin treatment in obese animals decreases the fat mass. In most instances, this is accompanied by improvements in glucose metabolism. The mechanisms underlying these beneficial effects remain to be determined.
5. Oxytocin Treatment in Obese Patients
Conflicting literature exists regarding the fact that circulating oxytocin levels are increased (63), unchanged (64), or decreased in obese (65) compared to lean human subjects. Moreover, highly variable results were reported using different types of measurements (66–68). This is an important issue as it leads to different conclusions about the activity of the oxytocin system in human obesity. This should prompt us to urgently define a standard and reliable method allowing drawing relevant conclusions on that topic.
Independently from this issue and taking into consideration that part of the leptin actions seems to be mediated by oxytocin (19), that chronic oxytocin treatment exerts promising effects on obesity and diabetes in various rodent models, as described above, it was hypothesized that oxytocin treatment could bypass the well-described leptin resistance, present not only in animals but also in obese human subjects (69).
As of now, two studies have been performed to determine the effects of oxytocin on food intake, body weight, and metabolic function in lean (70) and obese (47) volunteers. In lean healthy men (70), a single dose of 24 IU of intranasal oxytocin administration was shown to inhibit the reward – but not the hunger-driven eating, attenuating basal and postprandial levels of adrenocorticotropic hormone and of cortisol, and decreasing the postprandrial rise in plasma glucose, without modifying energy expenditure. Most interestingly, a clinical trial was performed by Zhang et al. (47), whereby intranasal oxytocin was administered to obese patients (BMI 30–36) at a dose of 24 IU, four times a day during 8 weeks. This treatment led to a significant and constant decrease in body weight (almost 9 kg) with a concomitant decrease in total- and LDL-cholesterol. It is noteworthy that this clinical trial was performed in male and female volunteers (pregnant and lactating females excluded) and that no adverse effect was described, reinforcing the fact that this treatment seems to be safe and effective in both sexes.
Summary
The beneficial effects of oxytocin treatment on body weight homeostasis obtained in animal models of obesity seem to apply to human obesity as well.
6. Oxytocin and Aging
Few data are available as yet on the oxytocin system during aging. Studies reported no change in hypothalamic oxytocin content in old rats (71), total number of oxytocin neurons in old human subjects (72), or plasma oxytocin levels in aged rats or humans (73–75). This is in contrast with very recent data demonstrating a decrease in oxytocin circulating levels in aged mice (76), pointing to possible species differences. Moreover, it has been proposed that PVN oxytocin secretion is increased in aged rats (73), which would fit with the decreased food intake that accompanies aging (77).
7. Future Perspectives
In view of the small cohort of the clinical trial mentioned above, in which oxytocin was administered to obese patients (9 subjects in the oxytocin-treated group and 11 subjects in the placebo-treated one) (47), larger multicenter clinical trials should be performed, in order to confirm the results and extent the conclusions. Moreover, these clinical trials should include not only obese but also type 2 diabetic patients, as the study by Zhang and collaborators (47) pointed to a tendency toward improvements in postprandial glucose and insulin levels in the oxytocin-treated group of obese subjects. This would also fit the observations that rodent models of type 2 diabetes (DIO rats and mice) exhibited improvements in glucose intolerance and insulin resistance in response to oxytocin treatment. Future studies should also shed some light on the molecular mechanisms by which oxytocin treatment can exert its beneficial effects.
Conflict of Interest Statement
Françoise Rohner-Jeanrenaud has a patent application (PCT/IB2011/052156) covering novel therapeutic uses of oxytocin. Jordi Altirriba and Anne-Laure Poher have no conflict of interest to declare.
References
1. World Health Organization. Global Health Risks: Mortality and Burden of Disease Attributable to Selected Major Risks. Geneva: World Health Organization (2009). Available from: http://www.who.int/healthinfo/global_burden_disease/GlobalHealthRisks_report_full.pdf
2. Wang YC, Colditz GA, Kuntz KM. Forecasting the obesity epidemic in the aging U.S. population. Obesity (Silver Spring) (2007) 15:2855–65. doi:10.1038/oby.2007.339
3. Daviglus ML, Liu K, Yan LL, Pirzada A, Manheim L, Manning W, et al. Relation of body mass index in young adulthood and middle age to Medicare expenditures in older age. JAMA (2004) 292:2743–9. doi:10.1001/jama.292.22.2743
4. Blevins JE, Ho JM. Role of oxytocin signaling in the regulation of body weight. Rev Endocr Metab Disord (2013) 14:311–29. doi:10.1007/s11154-013-9260-x
5. Gimpl G, Fahrenholz F. The oxytocin receptor system: structure, function, and regulation. Physiol Rev (2001) 81:629–83.
6. McEwen BB. General introduction to vasopressin and oxytocin: structure/metabolism, evolutionary aspects, neural pathway/receptor distribution, and functional aspects relevant to memory processing. Adv Pharmacol (2004) 50(1–50):655–708. doi:10.1016/S1054-3589(04)50001-7
7. Maejima Y, Sakuma K, Santoso P, Gantulga D, Katsurada K, Ueta Y, et al. Oxytocinergic circuit from paraventricular and supraoptic nuclei to arcuate POMC neurons in hypothalamus. FEBS Lett (2014) 588:4404–12. doi:10.1016/j.febslet.2014.10.010
8. Kiss A, Mikkelsen JD. Oxytocin – anatomy and functional assignments: a minireview. Endocr Regul (2005) 39:97–105.
9. Arletti R, Benelli A, Bertolini A. Influence of oxytocin on feeding behavior in the rat. Peptides (1989) 10:89–93. doi:10.1016/0196-9781(89)90082-X
10. Klockars A, Levine AS, Olszewski PK. Central oxytocin and food intake: focus on macronutrient-driven reward. Front Endocrinol (2015) 6:65. doi:10.3389/fendo.2015.00065
11. Takayanagi Y, Kasahara Y, Onaka T, Takahashi N, Kawada T, Nishimori K. Oxytocin receptor-deficient mice developed late-onset obesity. Neuroreport (2008) 19:951–5. doi:10.1097/WNR.0b013e3283021ca9
12. Camerino C. Low sympathetic tone and obese phenotype in oxytocin-deficient mice. Obesity (Silver Spring) (2009) 17:980–4. doi:10.1038/oby.2009.12
13. Kasahara Y, Takayanagi Y, Kawada T, Itoi K, Nishimori K. Impaired thermoregulatory ability of oxytocin-deficient mice during cold-exposure. Biosci Biotechnol Biochem (2007) 71:3122–6. doi:10.1271/bbb.70498
14. Swaab DF, Purba JS, Hofman MA. Alterations in the hypothalamic paraventricular nucleus and its oxytocin neurons (putative satiety cells) in Prader-Willi syndrome: a study of five cases. J Clin Endocrinol Metab (1995) 80:573–9. doi:10.1210/jcem.80.2.7852523
15. Kublaoui BM, Gemelli T, Tolson KP, Wang Y, Zinn AR. Oxytocin deficiency mediates hyperphagic obesity of Sim1 haploinsufficient mice. Mol Endocrinol (2008) 22:1723–34. doi:10.1210/me.2008-0067
16. Bonnefond A, Raimondo A, Stutzmann F, Ghoussaini M, Ramachandrappa S, Bersten DC, et al. Loss-of-function mutations in SIM1 contribute to obesity and Prader-Willi-like features. J Clin Invest (2013) 123:3037–41. doi:10.1172/JCI68035
17. Hakansson ML, Brown H, Ghilardi N, Skoda RC, Meister B. Leptin receptor immunoreactivity in chemically defined target neurons of the hypothalamus. J Neurosci (1998) 18:559–72.
18. Elmquist JK, Ahima RS, Maratos-Flier E, Flier JS, Saper CB. Leptin activates neurons in ventrobasal hypothalamus and brainstem. Endocrinology (1997) 138:839–42. doi:10.1210/endo.138.2.5033
19. Blevins JE, Schwartz MW, Baskin DG. Evidence that paraventricular nucleus oxytocin neurons link hypothalamic leptin action to caudal brain stem nuclei controlling meal size. Am J Physiol Regul Integr Comp Physiol (2004) 287:R87–96. doi:10.1152/ajpregu.00604.2003
20. Perello M, Raingo J. Leptin activates oxytocin neurons of the hypothalamic paraventricular nucleus in both control and diet-induced obese rodents. PLoS One (2013) 8:e59625. doi:10.1371/journal.pone.0059625
21. Maejima Y, Sedbazar U, Suyama S, Kohno D, Onaka T, Takano E, et al. Nesfatin-1-regulated oxytocinergic signaling in the paraventricular nucleus causes anorexia through a leptin-independent melanocortin pathway. Cell Metab (2009) 10:355–65. doi:10.1016/j.cmet.2009.09.002
22. Iwasaki Y, Maejima Y, Suyama S, Yoshida M, Arai T, Katsurada K, et al. Peripheral oxytocin activates vagal afferent neurons to suppress feeding in normal and leptin-resistant mice: a route for ameliorating hyperphagia and obesity. Am J Physiol (2015) 308:R360–9. doi:10.1152/ajpregu.00344.2014
23. Tung YC, Ma M, Piper S, Coll A, O’Rahilly S, Yeo GS. Novel leptin-regulated genes revealed by transcriptional profiling of the hypothalamic paraventricular nucleus. J Neurosci (2008) 28:12419–26. doi:10.1523/JNEUROSCI.3412-08.2008
24. Wu Z, Xu Y, Zhu Y, Sutton AK, Zhao R, Lowell BB, et al. An obligate role of oxytocin neurons in diet induced energy expenditure. PLoS One (2012) 7:e45167. doi:10.1371/journal.pone.0045167
25. Matarazzo V, Schaller F, Nedelec E, Benani A, Penicaud L, Muscatelli F, et al. Inactivation of Socs3 in the hypothalamus enhances the hindbrain response to endogenous satiety signals via oxytocin signaling. J Neurosci (2012) 32:17097–107. doi:10.1523/JNEUROSCI.1669-12.2012
26. Wang L, Barachina MD, Martinez V, Wei JY, Tache Y. Synergistic interaction between CCK and leptin to regulate food intake. Regul Pept (2000) 92:79–85. doi:10.1016/S0167-0115(00)00153-1
27. Olson BR, Drutarosky MD, Stricker EM, Verbalis JG. Brain oxytocin receptor antagonism blunts the effects of anorexigenic treatments in rats: evidence for central oxytocin inhibition of food intake. Endocrinology (1991) 129:785–91. doi:10.1210/endo-129-2-785
28. Blevins JE, Eakin TJ, Murphy JA, Schwartz MW, Baskin DG. Oxytocin innervation of caudal brainstem nuclei activated by cholecystokinin. Brain Res (2003) 993:30–41. doi:10.1016/j.brainres.2003.08.036
29. Baskin DG, Kim F, Gelling RW, Russell BJ, Schwartz MW, Morton GJ, et al. A new oxytocin-saporin cytotoxin for lesioning oxytocin-receptive neurons in the rat hindbrain. Endocrinology (2010) 151:4207–13. doi:10.1210/en.2010-0295
30. Mantella RC, Rinaman L, Vollmer RR, Amico JA. Cholecystokinin and D-fenfluramine inhibit food intake in oxytocin-deficient mice. Am J Physiol Regul Integr Comp Physiol (2003) 285:R1037–45. doi:10.1152/ajpregu.00383.2002
31. Atasoy D, Betley JN, Su HH, Sternson SM. Deconstruction of a neural circuit for hunger. Nature (2012) 488:172–7. doi:10.1038/nature11270
32. Yamamoto S, Morimoto I, Kai K, Arao T, Fujihira T, Morita E, et al. Centrally administered murine leptin stimulates plasma arginine-vasopressin secretion and increases the level of mRNA expression in the supraoptic nucleus of conscious rats. Neuroendocrinology (1999) 70:207–12. doi:10.1159/000054478
33. Honda K, Narita K, Murata T, Higuchi T. Leptin affects the electrical activity of neurones in the hypothalamic supraoptic nucleus. Brain Res Bull (2002) 57:721–5. doi:10.1016/S0361-9230(01)00788-2
34. Velmurugan S, Russell JA, Leng G. Systemic leptin increases the electrical activity of supraoptic nucleus oxytocin neurones in virgin and late pregnant rats. J Neuroendocrinol (2013) 25:383–90. doi:10.1111/jne.12016
35. Seeley RJ, Van Dijk G, Campfield LA, Smith FJ, Burn P, Nelligan JA, et al. Intraventricular leptin reduces food intake and body weight of lean rats but not obese Zucker rats. Horm Metab Res (1996) 28:664–8. doi:10.1055/s-2007-979874
36. Halaas JL, Boozer C, Blair-West J, Fidahusein N, Denton DA, Friedman JM. Physiological response to long-term peripheral and central leptin infusion in lean and obese mice. Proc Natl Acad Sci U S A (1997) 94:8878–83. doi:10.1073/pnas.94.16.8878
37. Frederich RC, Hamann A, Anderson S, Lollmann B, Lowell BB, Flier JS. Leptin levels reflect body lipid content in mice: evidence for diet-induced resistance to leptin action. Nat Med (1995) 1:1311–4. doi:10.1038/nm1295-1311
38. Deblon N, Veyrat-Durebex C, Bourgoin L, Caillon A, Bussier AL, Petrosino S, et al. Mechanisms of the anti-obesity effects of oxytocin in diet-induced obese rats. PLoS One (2011) 6:e25565. doi:10.1371/journal.pone.0025565
39. Maejima Y, Iwasaki Y, Yamahara Y, Kodaira M, Sedbazar U, Yada T. Peripheral oxytocin treatment ameliorates obesity by reducing food intake and visceral fat mass. Aging (Albany NY) (2011) 3:1169–77.
40. Zhang G, Bai H, Zhang H, Dean C, Wu Q, Li J, et al. Neuropeptide exocytosis involving synaptotagmin-4 and oxytocin in hypothalamic programming of body weight and energy balance. Neuron (2011) 69:523–35. doi:10.1016/j.neuron.2010.12.036
41. Morton GJ, Thatcher BS, Reidelberger RD, Ogimoto K, Wolden-Hanson T, Baskin DG, et al. Peripheral oxytocin suppresses food intake and causes weight loss in diet-induced obese rats. Am J Physiol Endocrinol Metab (2012) 302:E134–44. doi:10.1152/ajpendo.00296.2011
42. Blevins JE, Graham JL, Morton GJ, Bales KL, Schwartz MW, Baskin DG, et al. Chronic oxytocin administration inhibits food intake, increases energy expenditure, and produces weight loss in fructose-fed obese rhesus monkeys. Am J Physiol (2015) 308:R431–8. doi:10.1152/ajpregu.00441.2014
43. Boland D, Goren HJ. Binding and structural properties of oxytocin receptors in isolated rat epididymal adipocytes. Regul Pept (1987) 18:7–18. doi:10.1016/0167-0115(87)90045-0
44. Altirriba J, Poher AL, Caillon A, Arsenijevic D, Veyrat-Durebex C, Lyautey J, et al. Divergent effects of oxytocin treatment of obese diabetic mice on adiposity and diabetes. Endocrinology (2014) 155:4189–201. doi:10.1210/en.2014-1466
45. Gajdosechova L, Krskova K, Olszanecki R, Zorad S. Differential regulation of oxytocin receptor in various adipose tissue depots and skeletal muscle types in obese Zucker rats. Horm Metab Res (2015) 47(8):600–4. doi:10.1055/s-0034-1395677
46. Plante E, Menaouar A, Danalache BA, Yip D, Broderick TL, Chiasson JL, et al. Oxytocin treatment prevents the cardiomyopathy observed in obese diabetic male db/db mice. Endocrinology (2015) 156(4):1416–28. doi:10.1210/en.2014-1718
47. Zhang H, Wu C, Chen Q, Chen X, Xu Z, Wu J, et al. Treatment of obesity and diabetes using oxytocin or analogs in patients and mouse models. PLoS One (2013) 8:e61477. doi:10.1371/journal.pone.0061477
48. Beranger GE, Pisani DF, Castel J, Djedaini M, Battaglia S, Amiaud J, et al. Oxytocin reverses ovariectomy-induced osteopenia and body fat gain. Endocrinology (2014) 155:1340–52. doi:10.1210/en.2013-1688
49. Lewis GP, Matthews J. The mobilization of free fatty acids from rabbit adipose tissue in situ. Br J Pharmacol (1968) 34:564–78. doi:10.1111/j.1476-5381.1968.tb08485.x
50. Goren HJ, Hanif K, Dudley R, Hollenberg MD, Lederis K. Adenosine modulation of fat cell responsiveness to insulin and oxytocin. Regul Pept (1986) 16:125–34. doi:10.1016/0167-0115(86)90056-X
51. Eriksson H, Bjorgell P, Akerlund M, Hauksson A, Melin P. Effects of a tocolytic oxytocin analogue on lipid and carbohydrate metabolism. Gynecol Obstet Invest (1990) 29:97–100. doi:10.1159/000293310
52. Wilson EJ, Hollenberg MD. Effects of oxytocin and vasopressin on the preadipocyte 3T3-F442A cell line. Biochem Cell Biol (1987) 65:211–8. doi:10.1139/o87-027
53. Mukherjee SP, Mukherjee C. Stimulation of pyruvate dehydrogenase activity in adipocytes by oxytocin: evidence for mediation of the insulin-like effect by endogenous hydrogen peroxide independent of glucose transport. Arch Biochem Biophys (1982) 214:211–22. doi:10.1016/0003-9861(82)90024-8
54. Suva J, Caisova D, Stajner A. Modification of fat and carbohydrate metabolism by neurohypophyseal hormones. III. Effect of oxytocin on non-esterified fatty acid, glucose, triglyceride and cholesterol levels in rat serum. Endokrinologie (1980) 76:333–9.
55. Yano S, Kotani S, Miyatake A, Yamamura Y. Effect of neurohypophyseal hormones on FFA mobilization from the adipose tissue in the dehydrated rat. Endocrinol Jpn (1973) 20:345–7. doi:10.1507/endocrj1954.20.345
56. Eckertova M, Ondrejcakova M, Krskova K, Zorad S, Jezova D. Subchronic treatment of rats with oxytocin results in improved adipocyte differentiation and increased gene expression of factors involved in adipogenesis. Br J Pharmacol (2011) 162:452–63. doi:10.1111/j.1476-5381.2010.01037.x
57. Burt RL, Leake NH, Dannenburg WN. Metabolic activity of oxytocin in the puerperium. Nature (1963) 198:293. doi:10.1038/198293a0
58. Fabian E, Stork A, Kobilkova J, Fabianova J. Inhibition of lipolysis by oxytocin and vasopressin. Acta Univ Carol Med Monogr (1977) 77:141–5.
59. Muchmore DB, Little SA, De Haen C. A dual mechanism of action of ocytocin in rat epididymal fat cells. J Biol Chem (1981) 256:365–72.
60. Field CJ, Ryan EA, Thomson AB, Clandinin MT. Dietary fat and the diabetic state alter insulin binding and the fatty acyl composition of the adipocyte plasma membrane. Biochem J (1988) 253:417–24.
61. Gajdosechova L, Krskova K, Segarra AB, Spolcova A, Suski M, Olszanecki R, et al. Hypooxytocinaemia in obese Zucker rats relates to oxytocin degradation in liver and adipose tissue. J Endocrinol (2014) 220:333–43. doi:10.1530/JOE-13-0417
62. Shi H, Bartness TJ. Neurochemical phenotype of sympathetic nervous system outflow from brain to white fat. Brain Res Bull (2001) 54:375–85. doi:10.1016/S0361-9230(00)00455-X
63. Stock S, Granstrom L, Backman L, Matthiesen AS, Uvnas-Moberg K. Elevated plasma levels of oxytocin in obese subjects before and after gastric banding. Int J Obes (1989) 13:213–22.
64. Coiro V, Passeri M, Davoli C, D’Amato L, Gelmini G, Fagnoni F, et al. Oxytocin response to insulin-induced hypoglycemia in obese subjects before and after weight loss. J Endocrinol Invest (1988) 11:125–8. doi:10.1007/BF03350119
65. Qian W, Zhu T, Tang B, Yu S, Hu H, Sun W, et al. Decreased circulating levels of oxytocin in obesity and newly diagnosed type 2 diabetic patients. J Clin Endocrinol Metab (2014) 99(12):4683–9. doi:10.1210/jc.2014-2206
66. Szeto A, Mccabe PM, Nation DA, Tabak BA, Rossetti MA, Mccullough ME, et al. Evaluation of enzyme immunoassay and radioimmunoassay methods for the measurement of plasma oxytocin. Psychosom Med (2011) 73:393–400. doi:10.1097/PSY.0b013e31821df0c2
67. Zhang G, Zhang Y, Fast DM, Lin Z, Steenwyk R. Ultra sensitive quantitation of endogenous oxytocin in rat and human plasma using a two-dimensional liquid chromatography-tandem mass spectrometry assay. Anal Biochem (2011) 416:45–52. doi:10.1016/j.ab.2011.04.041
68. McCullough ME, Churchland PS, Mendez AJ. Problems with measuring peripheral oxytocin: can the data on oxytocin and human behavior be trusted? Neurosci Biobehav Rev (2013) 37:1485–92. doi:10.1016/j.neubiorev.2013.04.018
69. Proietto J, Thorburn AW. The therapeutic potential of leptin. Expert Opin Investig Drugs (2003) 12:373–8. doi:10.1517/13543784.12.3.373
70. Ott V, Finlayson G, Lehnert H, Heitmann B, Heinrichs M, Born J, et al. Oxytocin reduces reward-driven food intake in humans. Diabetes (2013) 62:3418–25. doi:10.2337/db13-0663
71. Melis MR, Stancampiano R, Fratta W, Argiolas A. Oxytocin concentration changes in different rat brain areas but not in plasma during aging. Neurobiol Aging (1992) 13:783–6. doi:10.1016/0197-4580(92)90102-4
72. Wierda M, Goudsmit E, Van Der Woude PF, Purba JS, Hofman MA, Bogte H, et al. Oxytocin cell number in the human paraventricular nucleus remains constant with aging and in Alzheimer’s disease. Neurobiol Aging (1991) 12:511–6. doi:10.1016/0197-4580(91)90081-T
73. Fliers E, Swaab DF. Activation of vasopressinergic and oxytocinergic neurons during aging in the Wistar rat. Peptides (1983) 4:165–70. doi:10.1016/0196-9781(83)90108-0
74. Zbuzek V, Fuchs AR, Zbuzek VK, Wu WH. Neurohypophyseal aging: differential changes in oxytocin and vasopressin release, studied in Fischer 344 and Sprague-Dawley rats. Neuroendocrinology (1988) 48:619–26. doi:10.1159/000125072
75. Chiodera P, Volpi R, Capretti L, Caiazza A, Marchesi M, Caffari G, et al. Oxytocin response to challenging stimuli in elderly men. Regul Pept (1994) 51:169–76. doi:10.1016/0167-0115(94)90206-2
76. Elabd C, Cousin W, Upadhyayula P, Chen RY, Chooljian MS, Li J, et al. Oxytocin is an age-specific circulating hormone that is necessary for muscle maintenance and regeneration. Nat Commun (2014) 5:4082. doi:10.1038/ncomms5082
Keywords: oxytocin, leptin, adipocyte, obesity, diabetes
Citation: Altirriba J, Poher A-L and Rohner-Jeanrenaud F (2015) Chronic oxytocin administration as a treatment against impaired leptin signaling or leptin resistance in obesity. Front. Endocrinol. 6:119. doi: 10.3389/fendo.2015.00119
Received: 18 February 2015; Accepted: 22 July 2015;
Published: 05 August 2015
Edited by:
Ez-Zoubir Amri, CNRS University of Nice-Sophia Antipolis, FranceReviewed by:
Marian Beekman, Leiden University Medical Center, NetherlandsMarie-Christine Pardon, University of Nottingham, UK
Copyright: © 2015 Altirriba, Poher and Rohner-Jeanrenaud. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jordi Altirriba, Laboratory of Metabolism, Department of Internal Medicine Specialties, Faculty of Medicine, University of Geneva, 1, rue Michel-Servet, Geneva CH-1211, Switzerland,am9yZ2UuYWx0aXJyaWJhQHVuaWdlLmNo