 Greisa Vila
Greisa Vila Jens Otto L. Jørgensen2
Jens Otto L. Jørgensen2 Anton Luger
Anton Luger Günter K. Stalla
Günter K. Stalla- 1Division of Endocrinology and Metabolism, Department of Internal Medicine III, Medical University of Vienna, Vienna, Austria
- 2Department of Endocrinology and Internal Medicine, Aarhus University Hospital, Aarhus, Denmark
- 3Max Planck Institute of Psychiatry, Munich, Germany
Acromegaly is characterized by chronic overproduction of growth hormone (GH) that leads to insulin resistance, glucose intolerance and, ultimately, diabetes. The GH-induced sustained stimulation of lipolysis plays a major role not only in the development of insulin resistance and prediabetes/diabetes, but also in the reduction of lipid accumulation, making acromegaly a unique case of severe insulin resistance in the presence of reduced body fat. In the present review, we elucidate the effects of GH hypersecretion on metabolic organs, describing the pathophysiology of impaired glucose tolerance in acromegaly, as well as the impact of acromegaly-specific therapies on glucose metabolism. In addition, we highlight the role of insulin resistance in the development of acromegaly-associated complications such as hypertension, cardiac disease, sleep apnea, polycystic ovaries, bone disease, and cancer. Taken together, insulin resistance is an important metabolic hallmark of acromegaly, which is strongly related to disease activity, the development of comorbidities, and might even impact the response to drugs used in the treatment of acromegaly.
Introduction
The main physiological roles of growth hormone (GH) are the regulation of postnatal growth and lipolysis. These actions are highly dependent on the nutritional state, which partitions the metabolic actions of GH (1). GH is released from pituitary somatotroph cells in a pulsatile fashion that is tightly controlled by hormones and nutrients (2–5). GH secretion is enhanced by growth hormone-releasing hormone, fasting, stress, exercise and hypoglycemia, and suppressed by somatostatin, insulin, insuline-like growth factor I (IGF-I), glucose and free fatty acids (1, 3–10).
GH excess in acromegaly results, with few exceptions, from a benign tumor of pituitary somatotroph cells and leads to chronically increased GH concentrations, which do not respond to the classical physiological feed-back inhibition (11). Therefore, acromegaly is characterized by a concomitant increase in both GH and IGF-I production and activity. The main metabolic consequence of acromegaly is insulin resistance, which may progress to diabetes. The underlying pathophysiological mechanisms are increased lipolysis, reduced peripheral glucose utilization and enhanced gluconeogenesis (12). Acromegaly is a unique condition of concomitant increases in GH, IGF-I, and insulin concentrations, where the increase in insulin resistance, paradoxically, is associated with reduced total body fat and even reduced fat accumulation in metabolic organs such as the liver (13, 14).
In the present review, we discuss the mechanisms leading to insulin resistance in patients with acromegaly, the pathophysiological implications of insulin resistance in the comorbidities of acromegaly, as well as the relationship between glucose homeostasis and disease activity in acromegaly.
Whole-Body Glucose Homeostasis in Acromegaly
GH and Glucose Homeostasis
The insulin-antagonistic effects of GH were initially described about 85 years ago following the observation that hypophysectomy performed in dogs improved hyperglycemia and experimental diabetes (15). In the second half of the twentieth century, studies performed using pituitary human GH extracts demonstrated direct effects of GH on lipolysis and hyperglycemia (16, 17). It was hypothesized that the GH-induced insulin antagonistic effect is strongly related to the lipolytic effects of GH, as free fatty acids released from fat stores inhibit glucose disposal, resulting in insulin resistance (16, 18). This constitutes a favorable metabolic adaptation to fasting and exercise (where insulin levels and activity are low) by providing lipid utilization at the expense of glucose. By contrast, GH is suppressed postprandially where insulin activity is maximal (16, 17). In addition, GH and insulin pathways have been shown to cross-talk at the postreceptor level in rodent models and in vitro (19), but this has not been replicated in human in vivo models (20). Since the physiological reciprocal temporal pattern of GH and insulin is abolished in active acromegaly, where the continous GH elevation chronically activates intracellular GH signaling, it remains possible that this could impair insulin signaling, hence causing insulin resistance.
GH signaling in muscle and fat tissues is confirmed already 30 min after a GH surge (21). Intravenous administration of GH in healthy human volunteers leads within 2 h to an increase in free fatty acids together with reduced glucose uptake and oxidation in the muscle in concomitance with increased non-oxidative glucose diposal and increased endogenous glucose production (17, 22). The GH-induced impairment of glucose uptake is causally linked to the concomitant activation of lipolysis, as the administration of the antilipolytic agent acipimox abrogates GH actions on insulin sensitivity (23). It is likely that also the GH-induced stimulation of gluconeogenesis is positively influenced by the increased free fatty acid levels (24).
Taken together, GH-induced insulin resistance seems to be mainly the consequence of the increased lipolysis, impaired insulin action in peripheral tissues leading to reduced glucose uptake, and also increased gluconeogenesis in the liver [(17); Figure 1].
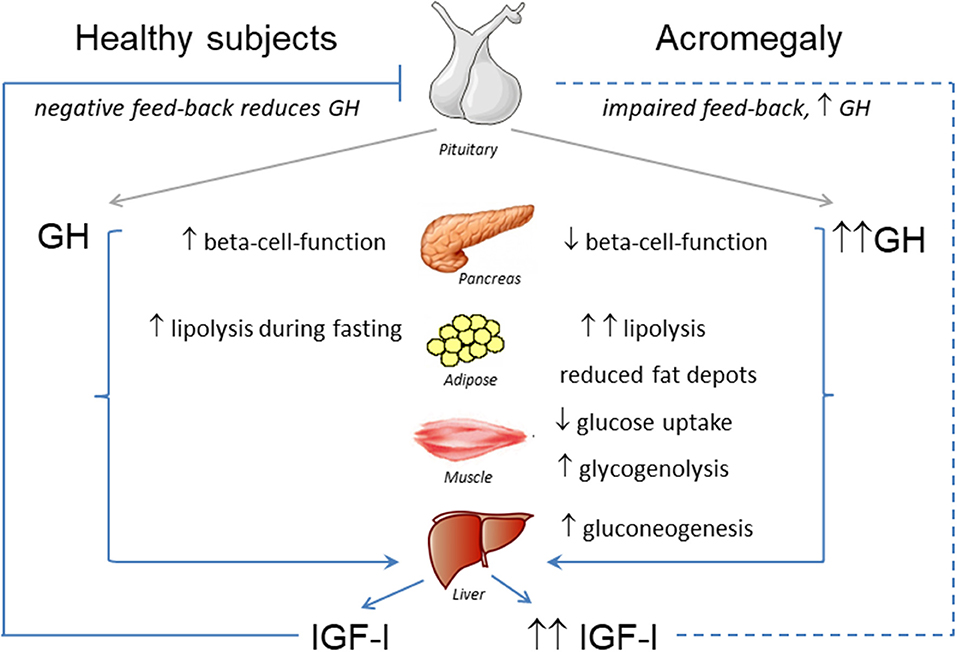
Figure 1. Effects of GH-IGF-I hypersecretion in metabolic organs.
IGF-I and Glucose Homeostasis
Prolonged administration of GH in the context of a positive energy balance leads to production of IGF-I by the liver. IGF-I is a single-chain polypeptide with 50% amino acid sequence homology with insulin, and its synthesis is stimulated not only by GH, but also by insulin (25). Acute increases in IGF-I concentrations exert insulin-like effects on glucose transport and circulating glucose concentrations, also in the absence of the insulin receptor (26). Nevertheless, circulating IGF-I does not cause hypoglycemia, as >90% is bound to specific binding proteins (27). Exogenous administration of IGF-I in high doses, on the other hand, mimicks administration of insulin and therefore acutely lowers plasma glucose levels (28). The same group showed that prolonged administration of IGF-I during several days does not impact circulating glucose concentrations, maybe because the increased IGF-I activity was balanced by a feedback-induced suppression of GH as well as insulin (29). Taken together, IGF-I has insulin-agonistic actions, thereby potentially counteracting the insulin-antagonistic effects of GH. Cross-sectional population studies have shown that both low and high IGF-I are associated with increased insulin resistance, highlighting the complexity of the IGF-I system, which probably reflects that IGF-I is a sex- and age-dependent biomarker of not only GH activity but also nutritional status (30). Circulating IGF-I per se, however, probably plays a very minor role in the regulation of glucose homeostasis in acromegaly.
Glucose Metabolism in Acromegaly
The overall effect of acromegaly on glucose metabolism is mainly determined by the insulin-antagonistic effects of chronically increased GH, which induces hepatic and peripheral insulin resistance as previously mentioned (12, 17, 31, 32). This is followed by a compensatory increase in beta-cell-function, which aims at maintaining euglycemia (33, 34) (Figure 1). Over time, chronic insulin resistance and fatty acid-induced lipotocixity deteriorate beta-cell-function eventually leading to diabetes (33, 34).
An impairment of glucose metabolism is observed in over 50% of patients with newly diagnosed acromegaly (35). Glucose homeostasis is related to disease activity in acromegaly, as higher IGF-I concentrations were found associated to lower insulin sensitivity (36). Nutrient intake and glucose physiologically suppress GH secretion in healthy subjects, but not in patients with acromegaly (37). Indeed, the latter is utilized in the biochemical evaluation of acromegaly disease activity (38).
Surgical cure of acromegaly improves insulin sensitivity and lowers circulating glucose and insulin concentrations (32, 34, 39).
Body Composition in Acromegaly
GH transgenic mice are resistant to high fat diet induced obesity, exhibiting an accumulation of lean tissue, and no increase in adipose tissue mass (40, 41). Based on the lipolytic effects of growth hormone one would predict a remarkable reduction of total body fat in acromegaly. Indeed, all studies evaluating body composition consistently describe reduced total fat content and also reduced organ-specific fat deposition in patients with acromegaly (13, 14, 42, 43). Nevertheless, one study demonstrated higher intermuscular adipose tissue depots in the presence of lower visceral and subcutaneous fat in patients with acromegaly (44). This is compatible with the observation of increased intramyocellular triglyceride (IMTG) accumulation in healthy subjects after 8 days high dose GH administration (45). Whether IMTG per se contributes to insulin resistance as indicated by other studies not involving GH (46) is uncertain, and it is noteworthy that IMTG also increases following exercise in fit individuals without compromising insulin sensitivity (47).
Ectopic lipids, however, play an important role in the pathophysiology of insulin resistance accompanying obesity in the general population (48). The presence of insulin resistance in the absence of hepatic lipid accumulation in patients with acromegaly is unique, and it is likely that the small increase in intramuscular fat in active acromegaly mainly reflects increased lipid oxidation in muscle. In all instances, biochemical control of acromegaly reverses this picture, increasing total fat mass and reducing lean body mass, while improving insulin sensitivity (13, 42, 43).
Pathophysiology of Insulin Resistance in Acromegaly
Fat Metabolism
Lipids constitute the main energy reserves in human physiology, being primarily stored in the adipose tissue as lipid droplets containing triacylglycerides surrounded by a phospholipid monolayer. An acute reduction of circulating free fatty acid levels stimulates GH secretion (10, 49). GH, in turn, stimulates lipolysis in humans leading to increased concentrations of free fatty acids and glycerol (50). Endogenous GH is essential for the increased lipolytic rate found during prolonged fasting, with fasting-induced peaks in endogenous GH secretion being crucial for the increased rate of lipolysis during starvation (51, 52). During the fed state, GH secretion is suppressed and insulin becomes the main regulator of substrate metabolism (10, 49). This feeding-induced shift between insulin and GH in the control of substrate metabolism was already suggested in 1963 by Rabinowitz and Zierler, with GH being responsible for the utilization of endogenous lipids during fasting and stress, thereby sparing glucose and proteins (16).
Acromegaly is associated with increased circulating levels of lipid intermediates, as well as with increased lipid uptake by the muscle, suggesting that fatty acids are a major fuel substrate in these patients (32). This sustained stimulation of lipolysis has three main consequences: (1) further deterioration of insulin sensitivity, (2) impairment of beta-cell function, and (3) reduction of whole-body fat. Surgical cure of acromegaly is followed by a normalization of lipolysis and glucose metabolism (32).
Beta-Cells
GH stimulation exerts insulinotrophic effects on β-cells in vitro (53) and in vivo, augmenting glucose-induced insulin secretion without playing a major role in basal insulin secretion (54). The pathophysiology underlying beta-cell-failure in acromegaly seems similar to that observed during the development of type 2 diabetes where insulin resistance leads to a compensatory hyperfunction of beta-cells (33, 34). Insulin resistance and lipotoxicity eventually lead to beta-cell dysfunction with failure to fully counterbalance the increased needs for insulin secretion (33, 34). Several studies have shown an improvement in beta-cell function after normalization of GH concentrations in patients with acromegaly (39, 55). Despite direct effects of GH on glucose metabolism, other hormonal alterations accompanying acromegaly may influence beta-cell function: 1) Acromegaly is associated with increased postprandial glucose-dependent insulinotropic polypeptide (GIP) concentrations, which in turn stimulate insulin secretion and influence postprandial (but not fasting) hyperinsulinemia (56); 2) A cross-sectional study found that beta-cell function strongly and directly correlates with the bone marker osteocalcin, revealing that the bone-beta-cell cross-talk initially described in other populations is also present in patients with acromegaly (57).
Liver
GH signaling in the liver is essential for the production of IGF-I and for the maintenance of a normal hepatic lipid metabolism (58). GH-induced hepatic IGF-I production depends on the local availability of insulin, with increased hepatic sensitivity to GH in the presence of high portal insulin levels (59). Higher portal insulin concentrations are associated with increased IGF-I concentrations, partially also due to the insulin-induced inhibition of hepatic IGFBP1 production (60). Patients with acromegaly have an increased glucose turnover, as GH increases hepatic glucose production by increasing glycogenolysis (17). Chronic GH administration impairs insulin sensitivity in the liver, thereby reducing the ability of insulin to suppress gluconeogenesis and glucose output (12, 32, 61).
Muscle
Local GH perfusion in the brachial artery leads to a rapid decrease in muscle glucose uptake and oxidation (17, 62, 63). These rapid GH effects in muscle cells could be either direct or secondary to the increased lipid utilization (17). GH signaling in muscle induces signal transducer and activator of transcription 5 (STAT5) phosphorylation and increased expression of canonical GH-dependent genes including IGF-I and cytokine-inducible SH2-containing protein (17, 21). The molecular mechanisms subserving GH-induced insulin resistance in human subjects remain uncertain. Studies in rodent models show impairment of insulin signaling at the level of phosphoinositide 3-kinase (PI3K) activity (64), but this is not observed in human studies in vivo (20, 21, 65, 66).
On the other hand, it has been shown that GH infusion increases lipolysis and suppresses pyruvate dehydrogenase activity, which indicates substrate competition between glucose and lipid intermediates (67) in accordance with the Randle hypothesis (18). However, it is noteworthy that human in vivo studies rely mainly on mRNA and protein expression in crude biopsies at specific time points, which may not be able to detect real time changes in complex signaling pathways.
Adipose Tissue
In general, circulating GH concentrations are inversely correlated to adipose tissue mass in both mice and humans (68). GH directly impairs glucose utilization in 3T3 adipocytes and also impairs insulin signaling in adipose tissue by influencing p85α expression that suppresses PI3K activity (69–72). GH receptor signaling phosphorylates the tyrosine residues on STAT5, leading to STAT5 activation (73). STAT5 mediates the GH-effects on lipolysis by increasing the transcription of several metabolic genes such as peroxisome proliferator-activated receptor (PPAR)-gamma and fatty acid synthase (73, 74). In addition, both STAT5- and mitogen-activated protein kinase (MEK)/extracellular signal-regulated kinase (ERK)-dependent intracellular signaling mediate the effects on GH in suppressing mRNA and protein levels of fat-specific protein 27 (FSP27), a negative regulator of lipolysis (75). The importance of FSP27 supression for GH signaling in adipocytes is highlighted by the fact that FSP27 overexpression fully abrogates the effects of GH on lipolysis and insulin resistance in adipose tissue, mainly by inhibiting PPAR-gamma phosphorylation (75, 76).
Acromegaly is also associated with decreased expression of the insulin-sensitizing adipokine adiponectin, but also with increased circulating concentrations of the proinflammatory adipokine visfatin, which is linked to enhanced inflammation and insulin resistance in many tissues (77–79). In addition, GH excess in acromegaly increases the expression of proinflammatory cytokines within the adipose tissue, which in turn might also contribute to the increased insulin resistance (80). This finding confirms that adipose tissue inflammation can be found also in the absence of increased adipose tissue mass. The functionality rather than the size of adipose tissue mass seems to determine the phenotype, as successful treatment of acromegaly resulted in a reduction of lean body mass and increase of total body fat mass together with improvement of insulin sensitivity and a reduction in proinflammatory cytokines (80).
Role of Insulin-Resistance in the Development of Acromegaly-Associated Comorbidities
Diabetes
Diabetes is a late consequence of impaired glucose metabolism in acromegaly, and occurs when the increased beta-cell function fails to compensate for the chronically increased insulin resistance (33, 34) (Figure 2). The prevalence of diabetes in patients with acromegaly is 20–35% at initial disease diagnosis (81, 82). The frequency of diabetes is related to disease control in acromegaly, and IGF-I concentrations are higher in patients with diabetes, when compared to patients with impaired glucose tolerance or normal glucose metabolism (81, 83). Age and positive family history for diabetes were found to be independently associated with impairment of glucose metabolism in acromegaly (35).
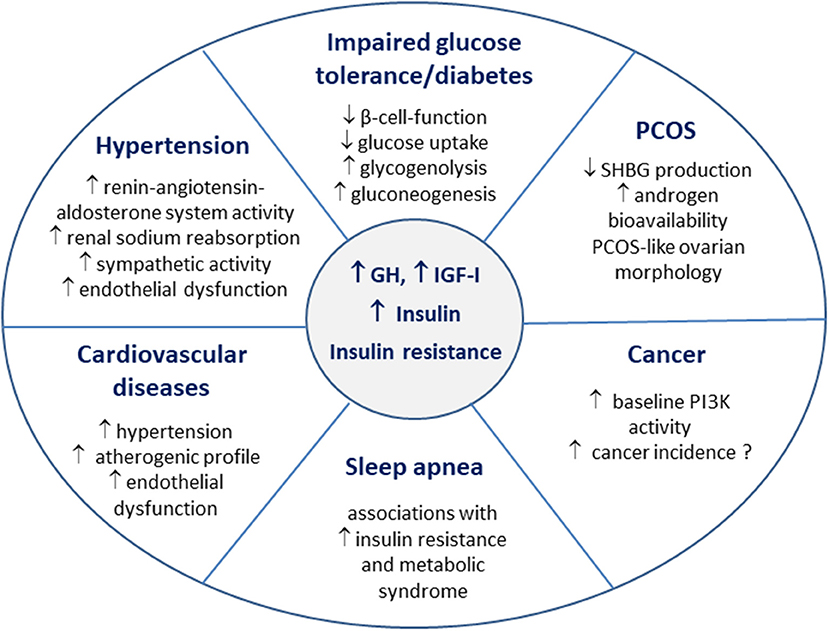
Figure 2. Insulin resistance in the pathophysiology of acromegaly comorbidities.
Hypertension
Insulin resistance is thought to be one of the main factors contributing to hypertension in the general population (84, 85). In acromegaly, hypertension is the most prevalent cardiovascular comorbidity, described in up to 60% of published acromegaly cohorts (86–90). The contribution of insulin resistance in the development of hypertension was studied in GH transgenic mice, in which increased systolic blood pressure appears at the age of 3–6 months and the prevalence of hypertension increases with age (91). In this model hyperinsulinemia develops very early, and unlike hypertension, even improves with age. The authors conclude that hyperinsulinemia might play a role in the pathophysiology of hypertension, but is not mandatory for the maintenance of hypertension (91).
Hyperinsulinemia may lead to increased sodium absorption in the kidney by activating the renin-angiotensin-aldosterone system, thereby also increasing the circulating plasma volume (92). In addition, both insulin and growth hormone increase sympathetic nervous activity (93). In accordance with this, patients with acromegaly lack the nocturnal fall in norepinephrine and blood pressure levels (94).
Insulin resistance and hyperinsulinemia impair also endothelium-dependent vasodilatation and increase oxidative stress in endothelial cells (95, 96). Indeed, flow-mediated vasodilatation, a functional test used for detecting endothelial dysfunction was lower in patients with acromegaly when compared to age- and gender-matched controls (97). Transsphenoidal surgery led to an increase/normalization of flow-mediated vasodilatation only in subjects that showed significant improvements in glucose, insulin concentrations and insulin resistance (97). The authors conclude that endothelial function in patients with acromegaly strongly relates to insulin resistance and does not always change following rapid improvement in GH and IGF-I concentrations (97). Interestingly intima media thickness of common carotid arteries, but not the prevalence of atheroclerotic plaques, is increased in patients with acromegaly (98). Taken together, insulin resistance is one of the main pathophysiological mechanisms contributing to the development of hypertension in acromegaly (99). In routine clinical practice, patients with acromegaly and impaired glucose tolerance/diabetes have higher blood pressure values than patients with acromegaly and normal glucose tolerance (100). In larger populations, the presence of diabetes in patients with acromegaly is associated with a significantly increased prevalence of hypertension (101).
Cardiovascular Disease
Insulin resistance is an independent risk factor for cardiovascular disease in the general population mediating the association between hyperglycemia and cardiovascular risk (102–104). Impaired glucose tolerance directly correlates to the severity of acromegalic cardiomyopathy (100). Patients with acromegaly and diabetes show an increased prevalence of cardiovascular diseases (101). GH and IGF-I excess lead to morphological heart changes (86, 105). Acromegalic cardiomyopathy displays similar features as diabetic cardiomyopathy, and the main morphological difference between the two is the lack of intracardiac lipid accumulation in acromegaly (14, 99). The coexistence of hypertension further aggravates cardiomyopathy in acromegalic patients (105, 106). Although there is no evidence for a direct effect of insulin resistance on heart morphology and dysfunction in acromegaly, the atherogenic properties of insulin resistance are thought to contribute to the pathophysiology of cardiovascular diseases [(99); Figure 2].
Obstructive Sleep Apnea
Obstructive sleep apnea is the most common sleep disorder in patients with acromegaly, and also specifically associated with increased mortality (89, 90, 92, 99). The prevalence of sleep apnea in acromegaly ranges from 45 to 80% and does not reliably relate to disease activity, with conflicting studies on its reversibility following successful treatment of acromegaly (92, 107).
The pathophysiology of sleep apnea in acromegaly is mainly based on soft tissue thickening of bronchial, pharyngeal, and laryngeal mucosa, as well as craniofacial morphological changes (92). Nevertheless, several studies support a bidirectional link between sleep apnea and insulin resistance/metabolic syndrome in the general population (108, 109).
Polycystic Ovary Syndrome (PCOS)
Both GH and IGF-I are known to affect ovarian function and morphology (110). Hypogonadism and menstrual irregularities are often reported in women with acromegaly, as volume effects of the pituitary adenoma and GH excess may lead to hypogonadotropic hypogonadism (111). Insulin directly modulates steroidogenesis in ovarian cells, and insulin resistance plays an important role in the pathogenesis of PCOS (112). GH modulates ovarian function both directly and via IGF-I (110). Hence, not only GH/IGF-I, but also hyperinsulinemia and insulin resistance accompanying acromegaly seem to be involved in the pathogenesis of a PCOS-like ovarian morphology, which was described in 50% of women with acromegaly (113). Although androgen levels are not generally increased in women with acromegaly, the circulating levels of sex hormone binding globulin (SHBG) were found to be low in a retrospective analysis (114). Insulin resistance is associated with decreased SHBG production in the liver, which in turn leads to increased androgen bioavailability, and this along with a direct stimulatory effect of GH on hair growth seems to be one of the reasons underlying the increased prevalence of hirsutism in women with acromegaly (113, 115). Therefore, the clinical phenotype of PCOS is determined not only by the severity of hyperandrogenism, but also by the degree of insulin resistance/hyperinsulinemia (Figure 2).
Cancer
High IGF-I levels are associated with an increased rate of malignancies in the general population (116, 117). Elevated GH and IGF-I concentrations might promote the development and progression of malignancies in patients with acromegaly but this remains a controversial topic (90). A recent meta-analysis revealed a moderate increased cancer risk, but this is mainly observed in single center studies (118). While older retrospective studies showed increased mortality due to cancer in acromegaly, the more recently published reports on acromegaly cohorts with normalized GH and IGF-I levels seem to indicate that the cancer mortality is comparable to the one observed in the general population (117, 119).
Insulin resistance and metabolic syndrome are also associated with an increased incidence of cancer (120). Indeed, the PI3K pathway, a common target of GH and insulin signaling, remains an important treatment target in malignancies (121). Other mitogenic pathways induced by both GH and insulin are MAPK/ERK and Ras-like GTPases (117). To date, there is no direct evidence on an additive effect of insulin resistance in the development of cancer in patients with acromegaly.
Bone Disease
Patients with acromegaly display a significant impairment of bone microarchitecture, increased bone formation and resorption, as well as an increased incidence of fractures (122–124). The frequency of fractures in patients with acromegaly relates to disease activity, male gender and concomitant hypogonadotropic hypogonadism (123). Importantly, the prevalence of diabetes is higher in acromegalic patients experiencing fractures (125). Diabetes is associated with an increased fracture risk also after biochemical control of acromegaly (125). To date the pathophysiological evidence linking insulin resistance with bone diseases is scarce, but cohort studies show impaired bone turnover, increased incidence of osteoporosis and fracture risk in non-acromegalic patients with diabetes (126, 127). GH and IGF-1 increase bone turnover and acromegaly is associated with distinct alterations in bone compartments showing lower trabecular bone quantitative parameters, while cortical bone density seems better preserved and found decreased only in patients with vertebral fractures (128). In contrast, diabetes is associated with increased cortical porosity and thinning by trabecularization of the endosteal part of cortical bone (129). So it appears that diabetes and acromegaly affect bone morphology differently. To date it is not clear whether insulin resistance plays a role in the association between fracture risk and diabetes in patients with acromegaly; or whether this is a simple marker of disease severity and thereby associated with other acromegaly complications.
Insulin Resistance in Relationship to Disease Activity and Therapy of Acromegaly
Biomarkers of glucose metabolism strongly relate to disease activity in patients with acromegaly, where IGF-I serves as a biomarker of overall disease control in acromegaly (35, 36). The impaired glucose metabolism often improves following successful pituitary surgery, and patients in remission have less prevalent diabetes than patients with persistent active disease necessitating medical therapy (32, 34, 39, 81). One of the main factors determining the normalization of glucose metabolism after surgical cure of acromegaly is the beta-cell state. Patients with preserved beta-cell function achieve a normalization of glucose tolerance after surgery, while impaired beta-cell function leads to persistently abnormal glucose metabolism also after successful surgery (55).
The relationship between parameters of glucose homeostasis and drugs used for treating acromegaly has been subject to extensive studies, as recently reviewed (130). A recent report compared the effect of the three main treatment modalities on glucose metabolism in patients with biochemically controlled acromegaly, finding out that plasma glucose decreases after successful surgery and after pegvisomant therapy, but increases in patients using first-generation somatostatin analogs (SSA) (131). First-generation somatostatin analogs (SSA) control GH secretion and IGF-I production, thereby lowering disease activity and improving insulin sensitivity in acromegaly. In parallel, however, they suppress secretion of insulin as well as of gastric and gut peptides, so their overall effect on glucose hoemostasis is not straightforward, but marked deterioration of glucose metabolism is rarely encountered (130, 132). Colao et al. described that the effects of SSAs on glucose metabolism depend on the status of glucose impairment before starting the therapy: SSA may increase plasma glucose levels in patients with normal or impaired glucose tolerance, and this effect was abolished after adding metformin (132). In patients with diabetes, both impairment and improvement of glucose tolerance were observed, and some cases needed optimization of diabetes therapy, but all patients had HbA1c < 6.5% at the end of the study (132). A recent meta-analysis including 47 studies on this topic found a high heterogeneity in fasting glucose and HbA1c outcomes, revealing a significant HbA1c increase over time (133). They describe a marginal and non-significant increase in fasting glucose, which became significant only in the subgroup of patients receiving SSA as second-line therapy, while glucose 2 h after OGTT significantly increased (133). In addition, they observed an improvement in insulin resistance and beta-cell function (133).
The multireceptor-ligand pasireotide more strongly suppresses insulin secretion and gut hormones and therefore hyperglycemia is observed in more than half of the patients (130, 134, 135). In healthy volunteers, pasireotide decreases insulin secretion and the incretin effect, but does not impact insulin sensitivity (135). In patients with acromegaly, improvement of disease control under pasireotide increases insulin sensitivity, but the concomitant impairment of beta-cell function is the main player determining the deterioration of glucose metabolism (130). In 13.2% of patients receiving pasireotide, treatment was withdrawn due to severe hyperglycemia (136).
The growth hormone antagonist pegvisomant was the first acromegaly medication to show a significant improvement in glucose metabolism with overnight reductions in endogenous glucose production and free fatty acid concentrations (137–139). Pegvisomant ameliorates all aspects of glucose metabolism and reduces the need for antidiabetic medications (101, 130). Therefore, pegvisomant is an attractive option in acromegalic patients with poorly controlled diabetes, and its dose requirements also depend on the severity of diabetes (140). The positive effects of pegvisomant on glucose metabolism are preserved when it is combined with an SSA (141–143).
Table 1 summarizes the effects of acromegaly-specific therapies on glucose metabolism. The relationship between dopamine agonist therapy and glucose metabolism has been subject to only a few reports, showing a reduction in basal and stimulated insulin levels (130, 144). Nevertheless, the impact of dopamine agonists on glucose metabolism was extensively studied in patients with prolactinomas, confirming their positive effect in reducing insulin resistance and ameliorating beta-cell function (145).

Table 1. Glucose homeostasis in relation to acromegaly treatment options.
Summary
Insulin resistance is an important metabolic hallmark of acromegaly caused mainly by the insulin-antagonizing effects of GH in general and the lipolytic effects of GH in particular. The degree of impairment of glucose metabolism is positively related to disease activity in acromegaly and is usually reversed after acromegaly treatment. Insulin resistance plays an important role in the development of acromegaly-specific comorbidities and further studies are needed for elucidating the role of drugs that improve insulin resistance on long-term patient outcomes in acromegaly.
Author Contributions
GV wrote the initial version of the manuscript. JJ, AL, and GS reviewed the literature and critically revised the manuscript.
Conflict of Interest Statement
GV has received lecture fees from IPSEN, Novartis, and Pfizer and serves on advisory boards for Pfizer and Novartis. JJ has received lecture fees and reserach grants from IPSEN, Novartis, and Pfizer and serves on advisory boards for Pfizer and IPSEN. AL has received honoraria for lectures from Ipsen, Novartis and Pfizer, participation in advisory boards from Ionis, Ipsen, Novartis, Pfizer. GS has received consultancy fees and/or reimbursements of delegate fees for conferences/educational events and/or travel expenses and/or funding for research projects from Pfizer, Ipsen, Lilly, Shire, Novartis, Sandoz, NovoNordisk, and HRA.
Abbreviations
ERK, extracellular signal-regulated kinase; FSP27, fat-specific protein 27; GH, growth hormone; IGF-I, insuline-like growth factor I; IMTG, intramyocellular triglyceride; MEK, mitogen-activated protein kinase; PCOS, polycystic ovary syndrome; PI3K, phosphoinositide 3-kinase; PPAR, peroxisome proliferator-activated receptor; SHBG, sex hormone binding globulin; SSA, somatostatin analog; STAT5, Signal transducer and activator of transcription 5.
References
1. Hawkes CP, Grimberg A. Insulin-like growth-factor-I is a marker for the nutritional state. Pediatr Endocrinol Rev. (2015) 13:499–511.
2. Albertsson-Wikland K, Rosberg S, Karlberg J, Groth T. Analysis of 24-hour growth hormone profiles in healthy boys and girls of normal stature: relation to puberty. J Clin Endocrinol Metab. (1994) 78:1195–201. doi: 10.1210/jc.78.5.1195
3. Melmed S. Insulin suppresses growth hormone secretion by rat pituitary cells. J Clin Invest. (1984) 73:1425–33. doi: 10.1172/JCI111347
4. Yamashita S, Melmed S. Insulin-like growth factor I action on rat anterior pituitary cells: suppression of growth hormone secretion and messenger ribonucleic acid levels. Endocrinology. (1986) 118:176–82. doi: 10.1210/endo-118-1-176
5. Roth J, Glick SM, Yalow RS, Berson SA. Hypoglycemia: a potent stimulus to secretion of growth hormone. Science. (1963) 140:987–8. doi: 10.1126/science.140.3570.987
6. Luger A, Watschinger B, Deuster P, Svoboda T, Clodi M, Chrousos GP. Plasma growth hormone and prolactin responses to graded levels of acute exercise and to a lactate infusion. Neuroendocrinology. (1992) 56:112–7. doi: 10.1159/000126912
7. Vila G, Maier C, Riedl M, Noworny P, Ludvik B, Luger A, et al. Bacterial endotoxin induces biphasic changes in plasma ghrelin in healthy humans. J Clin Endocrinol Metab. (2007) 92:3930–4. doi: 10.1210/jc.2007-1194
8. Hartman ML, Clayton PE, Johnson ML, Celniker A, Perlman AJ, Alberti KG, et al. A low dose euglycemic infusion of recombinant human insulin-like growth factor I rapidly suppresses fasting-enhanced pulsatile growth hormone secretion in humans. J Clin Invest. (1993) 91:2453–62. doi: 10.1172/JCI116480
9. Nakagawa K, Obara T. Suppression of plasma growth hormone levels with glucose infusion in patients with acromegaly. Clin Endocrinol (Oxf). (1976) 5:713–6. doi: 10.1111/j.1365-2265.1976.tb03874.x
10. Casanueva F, Villanueva L, Penalva A, Vila T, Cabezas-Cerrato J. Free fatty acid inhibition of exercise-induced growth hormone secretion. Horm Metab Res. (1981) 13:348–50. doi: 10.1055/s-2007-1019263
11. Ho PJ, DeMott-Friberg R, Barkan AL. Regulation of pulsatile growth hormone secretion by fasting in normal subjects and patients with acromegaly. J Clin Endocrinol Metab. (1992) 75:812–9. doi: 10.1210/jc.75.3.812
12. Hansen I, Tsalikian E, Beaufrere B, Gerich J, Haymond M, Rizza R. Insulin resistance in acromegaly: defects in both hepatic andextrahepatic insulin action. Am J Physiol. (1986) 250:E269–73. doi: 10.1152/ajpendo.1986.250.3.E269
13. Reyes-Vidal CM, Mojahed H, Shen W, Jin Z, Arias-Mendoza F, Fernandez JC, et al. Adipose tissue redistribution and ectopic lipid deposition in active acromegaly and effects of surgical treatment. J Clin Endocrinol Metab. (2015) 100:2946–55. doi: 10.1210/jc.2015-1917
14. Winhofer Y, Wolf P, Krššák M, Wolfsberger S, Tura A, Pacini G, et al. No evidence of ectopic lipid accumulation in the pathophysiology of the acromegalic cardiomyopathy. J Clin Endocrinol Metab. (2014) 99:4299–306. doi: 10.1210/jc.2014-2242
15. Houssay BA. The hypophysis and metabolism. N Engl J Med. (1936) 214:961–86. doi: 10.1056/NEJM193605142142001
16. Rabinowitz D, Zierler KL. A metabolic regulating device based on the actions of human growth hormone and of insulin singly and together on the human forearm. Nature. (1963) 199:913–5. doi: 10.1038/199913a0
17. Møller N, Jørgensen JO. Effects of growth hormone on glucose, lipid, and protein metabolism in human subjects. Endocr Rev. (2009) 30:152–77. doi: 10.1210/er.2008-0027
18. Randle PJ, Garland PB, Hales CN, Newsholme EA. The glucose fatty acid cycle: its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet. (1963) 1:785–9. doi: 10.1016/S0140-6736(63)91500-9
19. Dominici FP, Argentino DP, Muñoz MC, Miquet JG, Sotelo AI, Turyn D. Influence of the crosstalk between growth hormone and insulin signalling on the modulation of insulin sensitivity. Growth Horm IGF Res. (2005) 15:324–36. doi: 10.1016/j.ghir.2005.07.001
20. Jessen N, Djurhuus CB, Jørgensen JO, Jensen LS, Møller N, Lund S, et al. Evidence against a role for insulin-signaling proteins PI 3-kinase and Akt in insulin resistance in human skeletal muscle induced by short-term GH infusion. Am J Physiol Endocrinol Metab. (2005) 288:E194–9. doi: 10.1152/ajpendo.00149.2004
21. Jørgensen JO, Jessen N, Pedersen SB, Vestergaard E, Gormsen L, Lund SA, et al. GH receptor signaling in skeletal muscle and adipose tissue in human subjects following exposure to an intravenous GH bolus. Am J Physiol Endocrinol Metab. (2006) 291:E899–905. doi: 10.1152/ajpendo.00024.2006
22. Møller N, Jørgensen JO, Schmitz O, Møller J, Christiansen J, Alberti KG, et al. Effects of a growth hormone pulse on total and forearm substrate fluxes in humans. Am J Physiol. (1990) 258:E86–91. doi: 10.1152/ajpendo.1990.258.1.E86
23. Nielsen S, Møller N, Christiansen JS, Jørgensen JO. Pharmacological antilipolysis restores insulin sensitivity during growth hormone exposure. Diabetes. (2001) 50:2301–8. doi: 10.2337/diabetes.50.10.2301
24. Schwarz JM, Mulligan K, Lee J, Lo JC, Wen M, Noor MA, et al. Effects of recombinant human growth hormone on hepatic lipid and carbohydrate metabolism in HIV-infected patients with fat accumulation. J Clin Endocrinol Metab. (2002) 87:942. doi: 10.1210/jcem.87.2.8391
25. Frara S, Maffezzoni F, Mazziotti G, Giustina A. Current and emerging aspects of diabetes mellitus in acromegaly. Trends Endocrinol Metab. (2016) 27:470–83. doi: 10.1016/j.tem.2016.04.014
26. Di Cola G, Cool MH, Accili D. Hypoglycemic effect of insulin-like growth factor-1 in mice lacking insulin receptors. J Clin Invest. (1997) 99:2538–44. doi: 10.1172/JCI119438
27. Froesch ER, Schmid C, Schwander J, Zapf J. Actions of insulin-like growth factors. Annu Rev Physiol. (1985) 47:443–67. doi: 10.1146/annurev.ph.47.030185.002303
28. Guler HP, Zapf J, Froesch ER. Short-term metabolic effects of recombinant human insulin-like growth factor I in healthy adults. N Engl J Med. (1987) 317:137–40. doi: 10.1056/NEJM198707163170303
29. Guler HP, Schmid C, Zapf J, Froesch ER. Effects of recombinant insulin-like growth factor I on insulin secretion and renal function in normal human subjects. Proc Natl Acad Sci USA. (1989) 86:2868–72. doi: 10.1073/pnas.86.8.2868
30. Friedrich N, Thuesen B, Jørgensen T, Juul A, Spielhagen C, Wallaschofksi H, et al. The association between IGF-I and insulin resistance: a general population study in Danish adults. Diabetes Care. (2012) 35:768–73. doi: 10.2337/dc11-1833
31. Karlander S, Vranić M, Efendić S. Increased glucose turnover and glucose cycling in acromegalic patients with normal glucose tolerance. Diabetologia. (1986) 29:778–83. doi: 10.1007/BF00873216
32. Møller N, Schmitz O, Jøorgensen JO, Astrup J, Bak JF, Christensen SE, et al. Basal- and insulin-stimulated substrate metabolism in patients with active acromegaly before and after adenomectomy. J Clin Endocrinol Metab. (1992) 74:1012–9. doi: 10.1210/jcem.74.5.1569148
33. Defronzo RA. Banting lecture. From the triumvirate to the ominous octet: a new paradigm for the treatment of type 2 diabetes mellitus. Diabetes. (2009) 58:773–95. doi: 10.2337/db09-9028
34. Kasayama S, Otsuki M, Takagi M, Saito H, Sumitani S, Kouhara H, et al. Impaired β-cell function in the presence of reduced insulin sensitivity determines glucose tolerance status in acromegalic patients. Clin Endocrinol (Oxf). (2000) 52:549–55. doi: 10.1046/j.1365-2265.2000.00986.x
35. Alexopoulou O, Bex M, Kamenicky P, Mvoula AB, Chanson P, Maiter D. Prevalence and risk factors of impaired glucose tolerance and diabetes mellitus at diagnosis of acromegaly: a study in 148 patients. Pituitary. (2014) 17:81–9. doi: 10.1007/s11102-013-0471-7
36. Puder JJ, Nilavar S, Post KD, Freda PU. Relationship between disease-related morbidity and biochemical markers of activity in patients with acromegaly. J Clin Endocrinol Metab. (2005) 90:1972–8. doi: 10.1210/jc.2004-2009
37. Earll JM, Sparks LL, Forsham PH. Glucose suppression of serum growth hormone in the diagnosis of acromegaly. JAMA. (1967) 201:628–30. doi: 10.1001/jama.1967.03130080070021
38. Katznelson L, Laws ER Jr, Melmed S, Molitch ME, Murad MH, Utz A, et al. Acromegaly: an endocrine society clinical practice guideline. J Clin Endocrinol Metab. (2014) 99:3933–51. doi: 10.1210/jc.2014-2700
39. Roelfsema F, Frolich M. Glucose tolerance and plasma immunoreactive insulin levels in acromegalics before and after selective transsphenoidal surgery. Clin Endocrinol (Oxf). (1985) 22:531–7. doi: 10.1111/j.1365-2265.1985.tb00153.x
40. Berryman DE, List EO, Kohn DT, Coschigano KT, Seeley RJ, Kopchick JJ. Effect of growth hormone on susceptibility to diet-induced obesity. Endocrinology. (2006) 147:2801–8. doi: 10.1210/en.2006-0086
41. Olsson B, Bohlooly YM, Fitzgerald SM, Frick F, Ljungberg A, Ahrén B, et al. Bovine growth hormone transgenic mice are resistant to diet-induced obesity but develop hyperphagia, dyslipidemia, and diabetes on a high-fat diet. Endocrinology. (2005) 146:920–30. doi: 10.1210/en.2004-1232
42. Bredella MA, Schorr M, Dichtel LE, Gerweck AV, Young BJ, Woodmansee WW, et al. Body composition and ectopic lipid changes with biochemical control of acromegaly. J Clin Endocrinol Metab. (2017) 102:4218–25. doi: 10.1210/jc.2017-01210
43. Madsen M, Krusenstjerna-Hafstrøm T, Møller L, Christensen B, Vendelbo MH, Pedersen SB, et al. Fat content in liver and skeletal muscle changes in a reciprocal manner in patients with acromegaly during combination therapy with a somatostatin analog and a GH receptor antagonist: a randomized clinical trial. J Clin Endocrinol Metab. (2012) 97:1227–35. doi: 10.1210/jc.2011-2681
44. Freda PU, Shen W, Heymsfield SB, Reyes-Vidal CM, Geer EB, Bruce JN, et al. Lower visceral and subcutaneous but higher intermuscular adipose tissue depots in patients with growth hormone and insulin-like growth factor I excess due to acromegaly. J Clin Endocrinol Metab. (2008) 93:2334–43. doi: 10.1210/jc.2007-2780
45. Krag MB, Gormsen LC, Guo Z, Christiansen JS, Jensen MD, Nielsen S, et al. Growth hormone-induced insulin resistance is associated with increased intramyocellular triglyceride content but unaltered VLDL-triglyceride kinetics. Am J Physiol Endocrinol Metab. (2007) 292:E920–7. doi: 10.1152/ajpendo.00374.2006
46. Muoio DM. Revisiting the connection between intramyocellular lipids and insulin resistance: a long and winding road. Diabetologia. (2012) 55:2551–4. doi: 10.1007/s00125-012-2597-y
47. Goodpaster BH, He J, Watkins S, Kelley DE. Skeletal muscle lipid content and insulin resistance: evidence for a paradox in endurance-trained athletes. J Clin Endocrinol Metab. (2001) 86:5755–61. doi: 10.1210/jcem.86.12.8075
48. Petersen MC, Shulman GI. Mechanisms of insulin action and insulin resistance. Physiol Rev. (2018) 98:2133–223. doi: 10.1152/physrev.00063.2017
49. Imaki T, Shibasaki T, Shizume K, Masuda A, Hotta M, Kiyosawa Y, et al. The effect of free fatty acids on growth hormone (GH)-releasing hormone-mediated GH secretion in man. J Clin Endocrinol Metab. (1985) 60:290–3. doi: 10.1210/jcem-60-2-290
50. Moller L, Dalman L, Norrelund H, Billestrup N, Frystyk J, Moller N, et al. Impact of fasting on growth hormone signaling and action in muscle and fat. J Clin Endocrinol Metab. (2009) 94:965–72. doi: 10.1210/jc.2008-1385
51. Norrelund H, Djurhuus C, Jorgensen JO, Nielsen S. Effects of growth hormone on urea, glucose and lipid metabolism and insulin sensitivity during fasting in GH-deficient patients. Am J Physiol. (2003) 88:E3292–8. doi: 10.1152/ajpendo.00092.2003
52. Sakharova AA, Horowitz JF, Surya S, Goldenberg N, Harber MP, Symons K, et al. Role of growth hormone in regulating lipolysis, proteolysis, and hepatic glucose production during fasting. J Clin Endocrinol Metab. (2008) 93:2755–9. doi: 10.1210/jc.2008-0079
53. Davidson MB. Effect of growth hormone on carbohydrate and lipid metabolism. Endocr Rev. (1987) 8:115–31. doi: 10.1210/edrv-8-2-115
54. Pierluissi J, Pierluissi R, Ashcroft SJ. Effects of growth hormone on insulin release in the rat. Diabetologia. (1980) 19:391–6. doi: 10.1007/BF00280526
55. Kinoshita Y, Fujii H, Takeshita A, Taguchi M, Miyakawa M, Oyama K, et al. Impaired glucose metabolism in Japanese patients with acromegaly is restored after successful pituitary surgery if pancreatic β-cell function is preserved. Eur J Endocrinol. (2011) 164:467–73. doi: 10.1530/EJE-10-1096
56. Peracchi M, Porretti S, Gebbia C, Pagliari C, Bucciarelli P, Epaminonda P, et al. Increased glucose-dependent insulinotropic polypeptide (GIP) secretion in acromegaly. Eur J Endocrinol. (2001) 145:R1–4. doi: 10.1530/eje.0.145r001
57. Ueland T, Fougner SL, Godang K, Lekva T, Schurgers LJ, Scholz H, et al. Associations between body composition, circulating interleukin-1 receptor antagonist, osteocalcin, and insulin metabolism in active acromegaly. J Clin Endocrinol Metab. (2010) 95:361–8. doi: 10.1210/jc.2009-0422
58. Fan Y, Menon RK, Cohen P, Hwang D, Clemens T, DiGirolamo DJ, et al. Liver-specific deletion of the growth hormone receptor reveals essential role of growth hormone signaling in hepatic lipid metabolism. J Biol Chem. (2009) 284:19937–44. doi: 10.1074/jbc.M109.014308
59. Leung KC, Doyle N, Ballesteros M, Waters MJ, Ho KK. Insulin regulation of human hepatic growth hormone receptors: divergent effects on biosynthesis and surface translocation. J Clin Endocrinol Metab. (2000) 85:4712–20. doi: 10.1210/jc.85.12.4712
60. Frystyk J, Delhanty PJ, Skjaerbaek C, Baxter RC. Changes in the circulating IGF system during short-term fasting and refeeding in rats. Am J Physiol. (1999) 277:E245–52. doi: 10.1152/ajpendo.1999.277.2.E245
61. Forrest L, Sedmak C, Sikder S, Grewal S, Harman SM, Blackman MR, et al. Effects of growth hormone on hepatic insulin sensitivity and glucose effectiveness in healthy older adults. Endocrine. (2019) 63:497–506. doi: 10.1007/s12020-018-01834-4
62. Rabinowitz D, Klassen GA, Zierler KL. Effect of human growth hormone on muscle and adipose tissue metabolism in the forearm of man. J Clin Invest. (1965) 44:51–61. doi: 10.1172/JCI105126
63. Fineberg SE, Merimee TJ. Acute metabolic effects of human growth hormone. Diabetes. (1974) 23:499–504. doi: 10.2337/diab.23.6.499
64. Barbour LA, Mizanoor Rahman S, Gurevich I, Leitner JW, Fischer SJ, et al. Increased P85alpha is a potent negative regulator of skeletal muscle insulin signaling and induces in vivo insulin resistance associated with growth hormone excess. J Biol Chem. (2005) 280:37489–94. doi: 10.1074/jbc.M506967200
65. Krusenstjerna-Hafstrøm T, Clasen BF, Møller N, Jessen N, Pedersen SB, Christiansen JS, et al. Growth hormone (GH)-induced insulin resistance is rapidly reversible: an experimental study in GH-deficient adults. J Clin Endocrinol Metab. (2011) 96:2548–57. doi: 10.1210/jc.2011-0273
66. Krusenstjerna-Hafstrøm T, Madsen M, Vendelbo MH, Pedersen SB, Christiansen JS, Møller N, et al. Insulin and GH signaling in human skeletal muscle in vivo following exogenous GH exposure: impact of an oral glucose load. PLoS ONE. (2011) 6:e19392. doi: 10.1371/journal.pone.0019392
67. Nellemann B, Vendelbo MH, Nielsen TS, Bak AM, Høgild M, Pedersen SB, et al. Growth hormone-induced insulin resistance in human subjects involves reduced pyruvate dehydrogenase activity. Acta Physiol (Oxf). (2014) 210:392–402. doi: 10.1111/apha.12183
68. Berryman DE, List EO. Growth hormone's effect on adipose tissue: quality versus quantity. Int J Mol Sci. (2017) 18:1621. doi: 10.3390/ijms18081621
69. Schwartz J. Growth hormone directly alters glucose utilization in 3T3 adipocytes. Biochem Biophys Res Commun. (1984) 125:237–43. doi: 10.1016/S0006-291X(84)80359-9
70. del Rincon JP, Iida K, Gaylinn BD, McCurdy CE, Leitner JW, Barbour LA, et al. Growth hormone regulation of p85a expression and phosphoinositide 3-kinase activity in adipose tissue: mechanism for growth hormone-mediated insulin resistance. Diabetes. (2007) 56:1638–46. doi: 10.2337/db06-0299
71. Bergan HE, Kittilson JD, Sheridan MA. PKC and ERK mediate GH-stimulated lipolysis. J Mol Endocrinol. (2013) 51:213–24. doi: 10.1530/JME-13-0039
72. Gorin E, Tai LR, Honeyman TW, Goodman HM. Evidence for a role of protein kinase C in the stimulation of lipolysis by growth hormone and isoproterenol. Endocrinology. (1990) 126:2973–82. doi: 10.1210/endo-126-6-2973
73. Baik M, Yu JH, Hennighausen L. Growth hormone-STAT5 regulation of growth, hepatocellular carcinoma, and liver metabolism. Ann N Y Acad Sci. (2011) 1229:29–37. doi: 10.1111/j.1749-6632.2011.06100.x
74. Hogan JC, Stephens JM. The regulation of fatty acid synthase by STAT5A. Diabetes. (2005) 54:1968–75. doi: 10.2337/diabetes.54.7.1968
75. Sharma R, Luong Q, Sharma VM, Harberson M, Harper B, Colborn A, et al. Growth hormone controls lipolysis by regulation of FSP27 expression. J Endocrinol. (2018) 239:289–301. doi: 10.1530/JOE-18-0282
76. Sharma VM, Vestergaard ET, Jessen N, Kolind-Thomsen P, Nellemann B, Nielsen TS, et al. Growth hormone acts along the PPARγ-FSP27 axis to stimulate lipolysis in human adipocytes. Am J Physiol Endocrinol Metab. (2019) 316:E34–42. doi: 10.1152/ajpendo.00129.2018
77. Lam KS, Xu A, Tan KC, Wong LC, Tiu SC, Tam S. Serum adiponectin is reduced in acromegaly and normalized after correction of growth hormone excess. J Clin Endocrinol Metab. (2004) 89:5448–53. doi: 10.1210/jc.2003-032023
78. Sucunza N, Barahona MJ, Resmini E, Fernández-Real JM, Ricart W, Farrerons J, et al. A link between bone mineral density and serum adiponectin and visfatin levels in acromegaly. J Clin Endocrinol Metab. (2009) 94:3889–96. doi: 10.1210/jc.2009-0474
79. Kumari B, Yadav UCS. Adipokine Visfatin's role in pathogenesis of diabesity and related metabolic derangements. Curr Mol Med. (2018) 18:116–25. doi: 10.2174/1566524018666180705114131
80. Olarescu NC, Bollerslev J. The impact of adipose tissue on insulin resistance in acromegaly. Trends Endocrinol Metab. (2016) 27:226–37. doi: 10.1016/j.tem.2016.02.005
81. González B, Vargas G, de Los Monteros ALE, Mendoza V, Mercado M. Persistence of diabetes and hypertension after multimodal treatment of acromegal. J Clin Endocrinol Metab. (2018) 103:2369–75. doi: 10.1210/jc.2018-00325
82. Petrossians P, Daly AF, Natchev E, Maione L, Blijdorp K, Sahnoun-Fathallah M, et al. Acromegaly at diagnosis in 3173 patients from the liège acromegaly survey (LAS) database. Endocr Relat Cancer. (2017) 24:505–18. doi: 10.1530/ERC-17-0253
83. Espinosa-de-los-Monteros AL, González B, Vargas G, Sosa E, Mercado M. Clinical and biochemical characteristics of acromegalic patients with different abnormalities in glucose metabolism. Pituitary. (2011) 14:231–5. doi: 10.1007/s11102-010-0284-x
84. Ferrannini E, Haffner SM, Stern MP. Essential hypertension: an insulin-resistant state. J Cardiovasc Pharmacol. (1990) 15(Suppl. 5):S18–25. doi: 10.1097/00005344-199000005-00004
85. Saad MF, Rewers M, Selby J, Howard G, Jinagouda S, Fahmi S, et al. Insulin resistance and hypertension: the insulin resistance atherosclerosis study. Hypertension. (2004) 43:1324–31. doi: 10.1161/01.HYP.0000128019.19363.f9
86. Berg C, Petersenn S, Lahner H, Herrmann BL, Buchfelder M, Droste M, et al. Cardiovascular risk factors in patients with uncontrolled and long-term acromegaly: comparison with matched data from the general population and the effect of disease control. J Clin Endocrinol Metab. (2010) 95:3648–56. doi: 10.1210/jc.2009-2570
87. Arosio M, Reimondo G, Malchiodi E, Berchialla P, Borraccino A, De Marinis L, et al. Predictors of morbidity and mortality in acromegaly: an Italian survey. Eur J Endocrinol. (2012) 167:189–98. doi: 10.1530/EJE-12-0084
88. Portocarrero-Ortiz LA, Vergara-Lopez A, Vidrio-Velazquez M, Uribe-Diaz AM, García-Dominguez A, Reza-Albarrán AA, et al. The mexican acromegaly registry: clinical and biochemical characteristics at diagnosis and therapeutic outcomes. J Clin Endocrinol Metab. (2016) 101:3997–4004. doi: 10.1210/jc.2016-1937
89. Dal J, Feldt-Rasmussen U, Andersen M, Kristensen LØ, Laurberg P, Pedersen L, et al. Acromegaly incidence, prevalence, complications and long-term prognosis: a nationwide cohort study. Eur J Endocrinol. (2016) 175:181–90. doi: 10.1530/EJE-16-0117
90. Gadelha MR, Kasuki L, Lim DS, Fleseriu M. Systemic complications of acromegaly and the impact of the current treatment landscape: an update. Endocr Rev. (2019) 40:268–332. doi: 10.1210/er.2018-00115
91. Jara A, Benner CM, Sim D, Liu X, List EO, Householder LA, et al. Elevated systolic blood pressure in male GH transgenic mice is age dependent. Endocrinology. (2014) 155:975–86. doi: 10.1210/en.2013-1899
92. Powlson AS, Gurnell M. Cardiovascular disease and sleep-disordered breathing in acromegaly. Neuroendocrinology. (2016) 103:75–85. doi: 10.1159/000438903
93. Bondanelli M, Ambrosio MR, degli Uberti EC. Pathogenesis and prevalence of hypertension in acromegaly. Pituitary. (2001) 4:239–49. doi: 10.1023/A:1020798430884
94. Bondanelli M, Ambrosio MR, Franceschetti P, Margutti A, Trasforini G, degli Uberti EC. Diurnal rhythm of plasma catecholamines in acromegaly. J Clin Endocrinol Metab. (1999) 84:2458–67. doi: 10.1210/jc.84.7.2458
95. Mather KJ, Steinberg HO, Baron AD. Insulin resistance in the vasculature. J Clin Invest. (2013) 123:1003–4. doi: 10.1172/JCI67166
96. Wu HY, Jeng YY, Yue CJ, Chyu KY, Hsueh WA, Chan TM. Endothelial-dependent vascular effects of insulin and insulin-like growth factor I in the perfused rat mesenteric artery and aortic ring. Diabetes. (1994) 43:1027–32. doi: 10.2337/diab.43.8.1027
97. Sakai H, Tsuchiya K, Nakayama C, Iwashima F, Izumiyama H, Doi M, et al. Improvement of endothelial dysfunction in acromegaly after transsphenoidal surgery. Endocr J. (2008) 55:853–9. doi: 10.1507/endocrj.K07E-125
98. Colao A, Spiezia S, Cerbone G, Pivonello R, Marzullo P, Ferone D, et al. Increased arterial intima-media thickness by B-M mode echodoppler ultrasonography in acromegaly. Clin Endocrinol (Oxf). (2001) 54:515–24. doi: 10.1046/j.1365-2265.2001.01256.x
99. Colao A, Ferone D, Marzullo P, Lombardi G. Systemic complications of acromegaly: epidemiology, pathogenesis, and management. Endocr Rev. (2004) 25:102–52. doi: 10.1210/er.2002-0022
100. Colao A, Baldelli R, Marzullo P, Ferretti E, Ferone D, Gargiulo P, et al. Systemic hypertension and impaired glucose tolerance are independently correlated to the severity of the acromegalic cardiomyopathy. J Clin Endocrinol Metab. (2000) 85:193–9. doi: 10.1210/jc.85.1.193
101. Brue T, Lindberg A, Jan van der Lely A, Akerblad AC, Koltowska-Häggström M, Gomez R, et al. Diabetes in patients with acromegaly treated with pegvisomant: observations from acrostudy. Endocrine. (2019) 63:563–72. doi: 10.1007/s12020-018-1792-0
102. Hills SA, Balkau B, Coppack SW, Dekker JM, Mari A, Natali A, et al. The EGIR-RISC STUDY (The European group for the study of insulin resistance: relationship between insulin sensitivity and cardiovascular disease risk): I. methodology and objectives. Diabetologia. (2004) 47:566–70. doi: 10.1007/s00125-004-1335-5
103. Ning F, Tuomilehto J, Pyörälä K, Onat A, Söderberg S, Qiao Q, et al. Cardiovascular disease mortality in Europeans in relation to fasting and 2-h plasma glucose levels within a normoglycemic range. Diabetes Care. (2010) 33:2211–6. doi: 10.2337/dc09-2328
104. Færch K, Bergman B, Perreault L. Does insulin resistance drive the association between hyperglycemia and cardiovascular risk? PLoS ONE. (2012) 7:e39260. doi: 10.1371/journal.pone.0039260
105. López-Velasco R, Escobar-Morreale HF, Vega B, Villa E, Sancho JM, Moya-Mur JL, et al. Cardiac involvement in acromegaly: specific myocardiopathy or consequence of systemic hypertension? J Clin Endocrinol Metab. (1997) 82:1047–53. doi: 10.1210/jcem.82.4.3876
106. Holdaway IM, Rajasoorya RC, Gamble GD. Factors influencing mortality in acromegaly. J Clin Endocrinol Metab. (2004) 89:667–74. doi: 10.1210/jc.2003-031199
107. Ramos-Levi AM, Marazuela M. Sleep apnea syndrome in acromegalic patients: can morphological evaluation guide us to optimize treatment? Endocrine. (2016) 51:203–4. doi: 10.1007/s12020-015-0787-3
108. Ford ES. Risks for all-cause mortality, cardiovascular disease, and diabetes associated with the metabolic syndrome: a summary of the evidence. Diabetes Care. (2005) 28:1769–78. doi: 10.2337/diacare.28.7.1769
109. Gaines J, Vgontzas AN, Fernandez-Mendoza J, Bixler EO. Obstructive sleep apnea and the metabolic syndrome: the road to clinically-meaningful phenotyping, improved prognosis, and personalized treatment. Sleep Med Rev. (2018) 42:211–9. doi: 10.1016/j.smrv.2018.08.009
110. Vila G, Luger A. Growth hormone deficiency and pregnancy - any role for substitution? Minerva Endocrinol. (2018) 43:451–7. doi: 10.23736/S0391-1977.18.02834-1
111. Galdiero M, Pivonello R, Grasso LF, Cozzolino A, Colao A. Growth hormone, prolactin, and sexuality. J Endocrinol Invest. (2012) 35:782–94. doi: 10.1007/BF03345805
112. Diamanti-Kandarakis E, Dunaif A. Insulin resistance and the polycystic ovary syndrome revisited: an update on mechanisms and implications. Endocr Rev. (2012) 33:981–1030. doi: 10.1210/er.2011-1034
113. Kaltsas GA, Androulakis II, Tziveriotis K, Papadogias D, Tsikini A, Makras P, et al. Polycystic ovaries and the polycystic ovary syndrome phenotype in women with active acromegaly. Clin Endocrinol (Oxf). (2007) 67:917–22. doi: 10.1111/j.1365-2265.2007.02987.x
114. Kaltsas GA, Mukherjee JJ, Jenkins PJ, Satta MA, Islam N, Monson JP, et al. Menstrual irregularity in women with acromegaly. J Clin Endocrinol Metab. (1999) 84:2731–5. doi: 10.1210/jcem.84.8.5858
115. Winters SJ, Gogineni J, Karegar M, Scoggins C, Wunderlich CA, Baumgartner R, et al. Sex hormone-binding globulin gene expression and insulin resistance. J Clin Endocrinol Metab. (2014) 99:E2780–8. doi: 10.1210/jc.2014-2640
116. Pollak M. The question of a link between insulin-like growth factor physiology and neoplasia. Growth Horm IGF Res. (2000) 10(Suppl. B):S21–4. doi: 10.1016/S1096-6374(00)80005-2
117. Boguszewski CL, Boguszewski MCDS. Growth hormone's links to cancer. Endocr Rev. (2019) 40:558–74. doi: 10.1210/er.2018-00166
118. Dal J, Leisner MZ, Hermansen K, Farkas DK, Bengtsen M, Kistorp C, et al. Cancer incidence in patients with acromegaly: a cohort study and meta-analysis of the literature. J Clin Endocrinol Metab. (2018) 103:2182–8. doi: 10.1210/jc.2017-02457
119. Bolfi F, Neves AF, Boguszewski CL, Nunes-Nogueira VS. Mortality in acromegaly decreased in the last decade: a systematic review and meta-analysis. Eur J Endocrinol. (2018) 179:59–71. doi: 10.1530/EJE-18-0255
120. Esposito K, Chiodini P, Colao A, Lenzi A, Giugliano D. Metabolic syndrome and risk of cancer: a systematic review and meta-analysis. Diabetes Care. (2012) 35:2402–11. doi: 10.2337/dc12-0336
121. Esposito A, Viale G, Curigliano G. Safety, tolerability, and management of toxic effects of phosphatidylinositol 3-kinase inhibitor treatment in patients with cancer: a review. JAMA Oncol. (2019). doi: 10.1001/jamaoncol.2019.0034. [Epub ahead of print].
122. Kotzmann H, Bernecker P, Hübsch P, Pietschmann P, Woloszczuk W, Svoboda T, et al. Bone mineral density and parameters of bone metabolism in patients with acromegaly. J Bone Miner Res. (1993) 8:459–65. doi: 10.1002/jbmr.5650080410
123. Mazziotti G, Bianchi A, Bonadonna S, Cimino V, Patelli I, Fusco A, et al. Prevalence of vertebral fractures in men with acromegaly. J Clin Endocrinol Metab. (2008) 93:4649–55. doi: 10.1210/jc.2008-0791
124. Silva PPB, Amlashi FG, Yu EW, Pulaski-Liebert KJ, Gerweck AV, Fazeli PK, et al. Bone microarchitecture and estimated bone strength in men with active acromegaly. Eur J Endocrinol. (2017) 177:409–20. doi: 10.1530/EJE-17-0468
125. Mazziotti G, Gola M, Bianchi A, Porcelli T, Giampietro A, Cimino V, et al. Influence of diabetes mellitus in vertebral fractures in men with acromegaly. Endocrine. (2011) 40:102–8. doi: 10.1007/s12020-011-9486-x
126. Heilmeier U, Patsch JM. Diabetes and bone. Semin Musculoskelet Radiol. (2016) 20:300–4. doi: 10.1055/s-0036-1592366
127. Gilbert MP, Pratley RE. The impact of diabetes and diabetes medications on bone health. Endocr Rev. (2015) 36:194–213. doi: 10.1210/er.2012-1042
128. KuŽma M, Vaňuga P, Ságová I, Pávai D, Jackuliak P, Killinger Z, et al. Non-invasive DXA-derived bone structure assessment of acromegaly patients: a cross-sectional study. Eur J Endocrinol. (2019) 180:201–11. doi: 10.1530/EJE-18-0881
129. Ferrari SL, Abrahamsen B, Napoli N, Akesson K, Chandran M, Eastell R, et al. Diagnosis and management of bone fragility in diabetes: an emerging challenge. Osteoporos Int. (2018) 29:2585–96. doi: 10.1007/s00198-018-4650-2
130. Ferraù F, Albani A, Ciresi A, Giordano C, Cannavò S. Diabetes secondary to acromegaly: physiopathology, clinical features and effects of treatment. Front Endocrinol (Lausanne). (2018) 9:358. doi: 10.3389/fendo.2018.00358
131. Briet C, Ilie MD, Kuhn E, Maione L, Brailly-Tabard S, Salenave S, et al. Changes in metabolic parameters and cardiovascular risk factors after therapeutic control of acromegaly vary with the treatment modality. Data from the Bicêtre cohort, and review of the literature. Endocrine. (2019) 63:348–60. doi: 10.1007/s12020-018-1797-8
132. Colao A, Auriemma RS, Savastano S, Galdiero M, Grasso LF, Lombardi G, et al. Glucose tolerance and somatostatin analog treatment in acromegaly: a 12-month study. J Clin Endocrinol Metab. (2009) 94:2907–14. doi: 10.1210/jc.2008-2627
133. Cozzolino A, Feola T, Simonelli I, Puliani G, Pozza C, Giannetta E, et al. Somatostatin analogs and glucose metabolism in acromegaly: a meta-analysis of prospective interventional studies. J Clin Endocrinol Metab. (2018) 103:2089–99. doi: 10.1210/jc.2017-02566
134. Colao A, Bronstein MD, Freda P, Gu F, Shen CC, Gadelha M, et al. Pasireotide versus octreotide in acromegaly: a head-to-head superiority study. J Clin Endocrinol Metab. (2014) 99:791–9. doi: 10.1210/jc.2013-2480
135. Henry RR, Ciaraldi TP, Armstrong D, Burke P, Ligueros-Saylan M, Mudaliar S. Hyperglycemia associated with pasireotide: results from a mechanistic study in healthy volunteers. J Clin Endocrinol Metab. (2013) 98:3446–53. doi: 10.1210/jc.2013-1771
136. Bronstein MD, Fleseriu M, Neggers S, Colao A, Sheppard M, Gu F, et al. Switching patients with acromegaly from octreotide to pasireotide improves biochemical control: crossover extension to a randomized, double-blind, Phase III study. BMC Endocr Disord. (2016) 16:16. doi: 10.1186/s12902-016-0096-8
137. Higham CE, Rowles S, Russell-Jones D, Umpleby AM, Trainer PJ. Pegvisomant improves insulin sensitivity and reduces overnight free fatty acid concentrations in patients with acromegaly. J Clin Endocrinol Metab. (2009) 94:2459–63. doi: 10.1210/jc.2008-2086
138. Barkan AL, Burman P, Clemmons DR, Drake WM, Gagel RF, Harris PE, et al. Glucose homeostasis and safety in patients with acromegaly converted from long-acting octreotide to pegvisomant. J Clin Endocrinol Metab. (2005) 90:5684–91. doi: 10.1210/jc.2005-0331
139. Lindberg-Larsen R, Møller N, Schmitz O, Nielsen S, Andersen M, Orskov H, et al. The impact of pegvisomant treatment on substrate metabolism and insulin sensitivity in patients with acromegaly. J Clin Endocrinol Metab. (2007) 92:1724–8. doi: 10.1210/jc.2006-2276
140. Droste M, Domberg J, Buchfelder M, Mann K, Schwanke A, Stalla G, et al. Therapy of acromegalic patients exacerbated by concomitant type 2 diabetes requires higher pegvisomant doses to normalise IGF1 levels. Eur J Endocrinol. (2014) 171:59–68. doi: 10.1530/EJE-13-0438
141. Urbani C, Sardella C, Calevro A, Rossi G, Scattina I, Lombardi M, et al. Effects of medical therapies for acromegaly on glucose metabolism. Eur J Endocrinol. (2013) 169:99–108. doi: 10.1530/EJE-13-0032
142. Muhammad A, Coopmans EC, Delhanty PJD, Dallenga AHG, Haitsma IK, Janssen JAMJL, et al. Efficacy and safety of switching to pasireotide in acromegaly patients controlled with pegvisomant and somatostatin analogues: PAPE extension study. Eur J Endocrinol. (2018) 179:269–77. doi: 10.1530/EJE-18-0353
143. Jørgensen JO, Feldt-Rasmussen U, Frystyk J, Chen JW, Kristensen LØ, Hagen C, et al. Cotreatment of acromegaly with a somatostatin analog and a growth hormone receptor antagonist. J Clin Endocrinol Metab. (2005) 90:5627–31. doi: 10.1210/jc.2005-0531
144. Rau H, Althoff PH, Schmidt K, Badenhoop K, Usadel KH. Bromocriptine treatment over 12 years in acromegaly: effect on glucose tolerance and insulin secretion. Clin Investig. (1993) 71:372–8. doi: 10.1007/BF00186626
145. Auriemma RS, Granieri L, Galdiero M, Simeoli C, Perone Y, Vitale P, et al. Effect of cabergoline on metabolism in prolactinomas. Neuroendocrinology. (2013) 98:299–310. doi: 10.1159/000357810
146. Christiansen Arlien- Søborg M, Trolle C, Alvarson E, Bæk A, Dal J, Jørgensen JOL. Biochemical assessment of disease control in acromegaly: reappraisal of the glucose suppression test in somatostatin analogue (SA) treated patients. Endocrine. (2017) 56:589–94. doi: 10.1007/s12020-017-1258-9
147. Shen M, Wang M, He W, He M, Qiao N, Ma Z, et al. Impact of long-acting somatostatin analogues on glucose metabolism in acromegaly: a hospital-based study. Int J Endocrinol. (2018) 2018:3015854. doi: 10.1155/2018/3015854
148. Drake WM, Rowles SV, Roberts ME, Fode FK, Besser GM, Monson JP, et al. Insulin sensitivity and glucose tolerance improve in patients with acromegaly converted from depot octreotide to pegvisomant. Eur J Endocrinol. (2003) 149:521–7. doi: 10.1530/eje.0.1490521
149. Feola T, Cozzolino A, Simonelli I, Sbardella E, Pozza C, Giannetta E, et al. Pegvisomant improves glucose metabolism in acromegaly: a meta-analysis of prospective interventional studies. J Clin Endocrinol Metab. (2019) 104:2892–902. doi: 10.1210/jc.2018-02281
150. Shimon I, Adnan Z, Gorshtein A, Baraf L, Saba Khazen N, Gershinsky M, et al. Efficacy and safety of long-acting pasireotide in patients with somatostatin-resistant acromegaly: a multicenter study. Endocrine. (2018) 62:448–55. doi: 10.1007/s12020-018-1690-5
151. De Marinis L, Bianchi A, Fusco A, Cimino V, Mormando M, Tilaro L, et al. Long-term effects of the combination of pegvisomant with somatostatin analogs (SSA) on glucose homeostasis in non-diabetic patients with active acromegaly partially resistant to SSA. Pituitary. (2007) 10:227–32. doi: 10.1007/s11102-007-0037-7
Keywords: glucose, insulin, growth hormone, IGF-I, acromegaly complications, diabetes, comorbidities, pathophysiology
Citation: Vila G, Jørgensen JOL, Luger A and Stalla GK (2019) Insulin Resistance in Patients With Acromegaly. Front. Endocrinol. 10:509. doi: 10.3389/fendo.2019.00509
Received: 05 May 2019; Accepted: 12 July 2019;
Published: 30 July 2019.
Edited by:
Marek Bolanowski, Wroclaw Medical University, PolandReviewed by:
Marija Pfeifer, University of Ljubljana, SloveniaOdelia Cooper, Cedars-Sinai Medical Center, United States
Copyright © 2019 Vila, Jørgensen, Luger and Stalla. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Günter K. Stalla, c3RhbGxhQHBzeWNoLm1wZy5kZQ==; Z3VlbnRlci5zdGFsbGFAbWVkaWNvdmVyLmRl