 Sana Ullah1†
Sana Ullah1† Shahid Ahmad
Shahid Ahmad Saleem Ullah
Saleem Ullah Sana Ullah
Sana Ullah Ghulam Nabi
Ghulam Nabi- 1Centre of Biotechnology and Microbiology, University of Peshawar, Peshawar, Pakistan
- 2School of Ecology and Environment, Hainan University, Haikou, Hainan, China
- 3Academy of Plateau Science and Sustainability, College of Life Sciences, Qinghai Normal University, Xining, China
- 4Department of Zoology, Division of Science and Technology, University of Education, Lahore, Pakistan
- 5Institute of Nature Conservation, Polish Academy of Sciences, Krakow, Poland
- 6Key Laboratory of Adaptation and Evolution of Plateau Biota, Laboratory of Plateau Fish Evolutionary and Functional Genomics, Qinghai Key Laboratory of Animal Ecological Genomics, Northwest Institute of Plateau Biology, Chinese Academy of Science, Xining, China
Over the years, the vaste expansion of plastic manufacturing has dramatically increased the environmental impact of microplastics [MPs] and nanoplastics [NPs], making them a threat to marine and terrestrial biota because they contain endocrine disrupting chemicals [EDCs] and other harmful compounds. MPs and NPs have deleteriouse impacts on mammalian endocrine components such as hypothalamus, pituitary, thyroid, adrenal, testes, and ovaries. MPs and NPs absorb and act as a transport medium for harmful chemicals such as bisphenols, phthalates, polybrominated diphenyl ether, polychlorinated biphenyl ether, organotin, perfluorinated compounds, dioxins, polycyclic aromatic hydrocarbons, organic contaminants, and heavy metals, which are commonly used as additives in plastic production. As the EDCs are not covalently bonded to plastics, they can easily leach into milk, water, and other liquids affecting the endocrine system of mammals upon exposure. The toxicity induced by MPs and NPs is size-dependent, as smaller particles have better absorption capacity and larger surface area, releasing more EDC and toxic chemicals. Various EDCs contained or carried by MPs and NPs share structural similarities with specific hormone receptors; hence they interfere with normal hormone receptors, altering the hormonal action of the endocrine glands. This review demonstrates size-dependent MPs’ bioaccumulation, distribution, and translocation with potential hazards to the endocrine gland. We reviewed that MPs and NPs disrupt hypothalamic-pituitary axes, including the hypothalamic-pituitary-thyroid/adrenal/testicular/ovarian axis leading to oxidative stress, reproductive toxicity, neurotoxicity, cytotoxicity, developmental abnormalities, decreased sperm quality, and immunotoxicity. The direct consequences of MPs and NPs on the thyroid, testis, and ovaries are documented. Still, studies need to be carried out to identify the direct effects of MPs and NPs on the hypothalamus, pituitary, and adrenal glands.
1 Introduction
xIn recent decades, plastic pollution has become one of the most widespread and enduring anthropogenic alterations in all environmental compartments of our planet’s surface (1) and therefore, considered as a stratigraphic marker for the Anthropocene (2). A culture of single-use plastic, rapid and inexpensive plastic production and non-circular economic models led to the creation of over 368 million metric tons (Mt) of single-use plastic in 2019 (3). It is expected that if the current production and waste management trends continue, approximately 12,000 Mt of plastic waste will end up in the natural environment by 2050 (4). To date, plastic debris has affected 3876 species only in the aquatic environment (https://litterbase.awi.de/interaction_detail; date assessed July 28, 2022), and by the year 2050, plastic will be found in the digestive tract of 99% of all sea bird species (5). Globally mammals are already at-risk due to several reasons, including climate change (6), however, bioaccumulation of the microplastic (MP) and associated toxic chemical additives are accelerating the risk of extinction (7, 8). Despite its ubiquitous distribution, current knowledge about the health effects of MP and associated chemicals in mammals is limited. Therefore, we aim to highlight how MP affects mammalian endocrine glands, which could contribute to their conservation and management, especially in vulnerable populations.
1.1 Rising global plastic and MPs pollution in the terrestrial and aquatic environment
Plastic waste is a contemporary societal and ecological issue due to its indispensable nature and ubiquitous use in daily life, associated with long-term detrimental effects on organisms (9). Plastic consumption will continue to rise in response to global population growth, and as plastic degrades, microplastics (<5 mm) and nanoplastics (< 1 nm) enter the terrestrial ecosystem in several ways (10). Depending on their manufactured and fragmented origin, MPs can be classified as primary or secondary (11). Primary MPs are primarily designed into small sizes for commercial practices like personal care products, whereas secondary MPs are fragmented from larger plastics by various physical and biological methods (10), such as UV radiation, temperature changes, and wave action (12). Due to sewage sludge applications, each year, 63–430 and 44–300 thousand tons of MPs are added to agro-ecosystems in Europe and North America (13). In marine ecosystems, approximately 5-13 million tons of plastic debris enter the ocean each year (8). Consequently, the world’s upper ocean currently comprises 24.4 trillion pieces, (8.2 × 104 ~ 57.8 × 104 tons) of micro-plastic (14), and its concentration might exceed 250 million metric tons by 2025 (8, 15). In 2015, Oceanographers estimated 15-51 trillion MP particles floating on water surfaces worldwide (16). In Europe, approximately, 63,000–430,000 tonnes of MPs entered the farmlands annually (17), while in most part of the world, data regarding MP loading in agriculture farmlands are unavailable.
1.2 Chemical composition of MPs and their endocrine-disrupting effects
MPs generated from plastic degradation persist for hundreds and thousands of years in the environment (18). Plastic bottles, disposable diapers, and polystyrene foam have a life span of 450, 500, and >5000 years [https://www.goecopure.com/lifespan-of-plastic.aspx]. Plastic additives used during plastic processing contribute up to 70% of plastics (18). More than 10,000 chemicals are identified as plastic additives, and 2400 chemicals have been classified as detrimental to marine and terrestrial biota (19). These additives include plasticizers, antioxidants, UV stabilizers, dyes, and flame retardants. Some of them are of serious concern, such as alkylphenol (20), polybrominated diphenyl ethers [PBDEs] (21), phthalates, organotins, perfluorinated compounds, dioxin (22), bisphenol A [BPA] and heavy metals like chromium, lead, and cadmium (8, 23). Approximately, 1000 chemicals classified as endocrine disruptor chemicals (EDCs) alter the expression of various hormone receptors and interfere with the synthesis, secretion, transport, and action of hormones, leading to endocrine and developmental abnormalities (23, 24). Nearly nine different forms of MPs are reported in human feces from multiple countries, clearly validating the presence of MPs in the human food chain and warning us about their harmful effects on human health (25). MPs and their composite toxic additives can cross various biological membranes, blood-brain barriers and both can interfere with various hormone receptors, thereby disrupting different hypothalamic axes such as the hypothalamic-pituitary-thyroid axis [HPT], the hypothalamic-pituitary-adrenal axis [HPA], and hypothalamic-pituitary-gonadal axis (HPG; Figure 1) (23, 26–29). Various metabolic disorders, gut dysbiosis, and intestinal barrier dysfunction induced by MPs have been explored using rats and mice as model organisms. Similarly, neurobehavioral changes, disrupted thyroid status, and biochemical stress are the direct consequences of MPs exposure in rats (30).
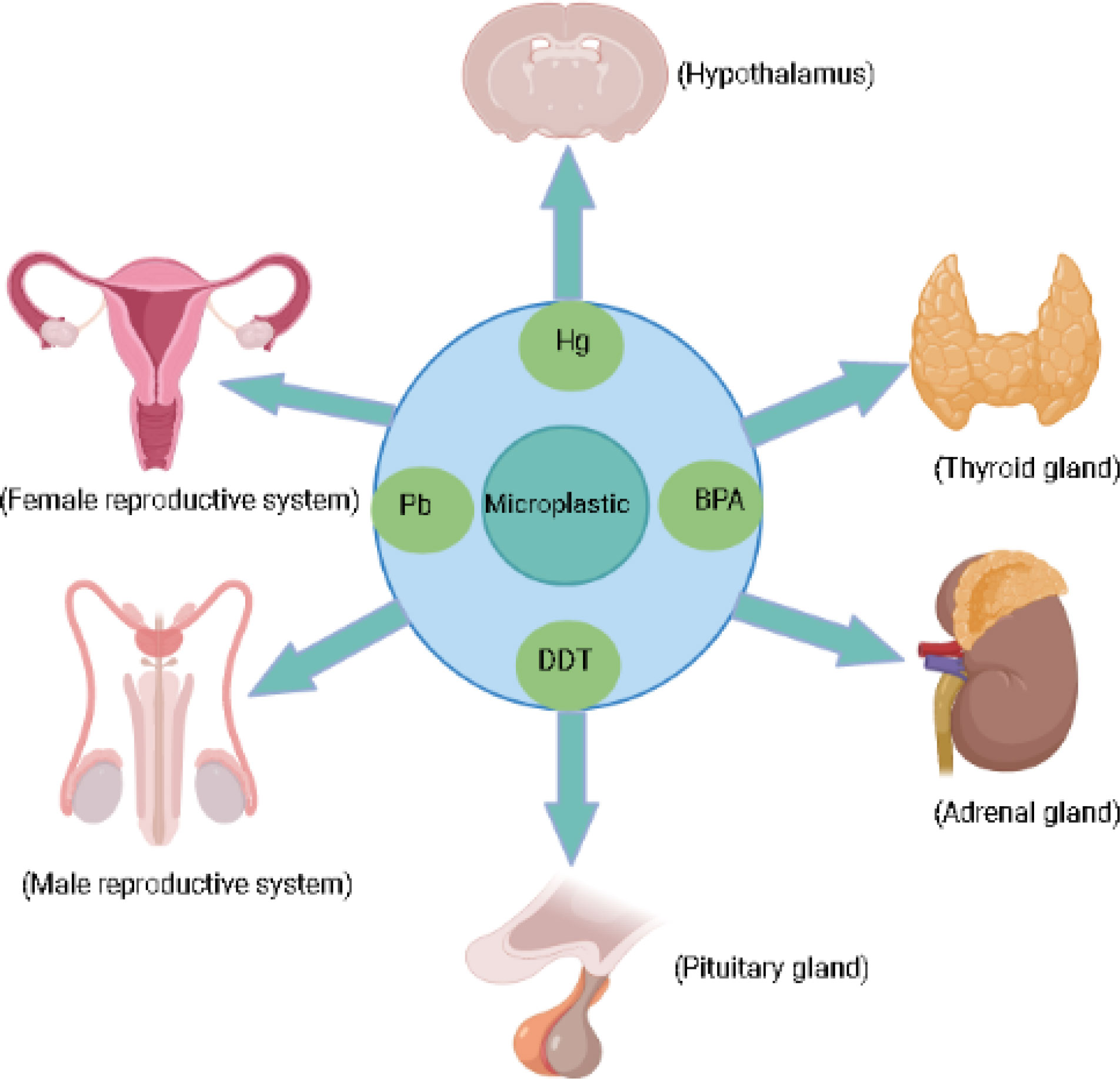
Figure 1 Micoplastic and their associated chemicals exposure can effect endocrine glands.
1.3 MPs bioaccumulation and biomagnification due to their “Trojan Horse” effects in mammals
The large surface area and hydrophobic surface of MPs make them a suitable medium for carrying many pollutants such as EDCs, heavy metals, and other toxic organic chemicals, making them harmful to mammals through bioaccumulation and biomagnification processes (31, 32). These are called “Trojan Horse Effects” of MPs (33), and induce several synergistic, behavioral, histological, and biomolecular alternations (32). Many EDCs and other pollutants are added as additives or absorbed by MPs; after being consumed directly or indirectly through the food web, MPs increase their bioaccumulation in mammals (8, 21, 34). MPs accumulate in various body parts and are involved in biochemical pathways, affecting cell functioning by crossing biological membranes in a size-dependent manner (35). Studies have shown that MPs of size 0.1-10 µm can cross biological membranes, blood-brain barrier, and even placenta, enhancing the possibilities of their bio-accumulation in secondary tissues such as the liver and brain (23). While MPs <150µm can cross the gastrointestinal tract, those <5µm can accumulate in macrophages and be carried to the blood circulation and the spleen (22). Similarly, MPs <10µm trans-locate from the gut to the circulatory system and can accumulate in the liver, kidney, and brain (36).
This review warns about the accumulation of MPs in marine and terrestrial ecosystems. Furthermore, it implies that studies need to be carried out to deeply understand the action mechanism of MPs, nanoplastics [NPs], and associated chemicals in aquatic and terrestrial biota and their long-term detrimental consequences on the mammalian endocrine system.
2 Effects of MPs, NPs, and associated chemicals on the mammalian thyroid gland
The thyroid gland is an essential endocrine gland responsible for normal brain function, growth, and neurological development of all animals (37). The thyroid functions under the HPT axis and affects almost every organ in the body (38), therefore disruption in thyroid homeostasis can be detrimental and will affect the body’s overall health status. Long-term exposure to plastic particles and associated chemicals has been shown to exhaust thyroid endocrine function by weakening its driving forces in regulating growth, development, metabolism, and reproduction (39). MPs additives and pollutants, such as PBDEs, BPA, phthalates, and organotin act as thyroid-disrupting chemicals [TDCs] (22) (Table 1). Similarly, MPs cause thyroid dysfunction and developmental abnormalities once ingested with associated POPs and EDCs (43). Phthalate causes a reduction in thyroid weight during childhood exposure and associated with developmental abnormalities and hyperactivity of the thyroid gland (44, 45). These TDCs associated with plastic enter the body through the gastrointestinal tract and interfere with T4 and T3 biochemical pathways, while their circulation adversely affects thyroid hormone production and metabolism, affecting other organs like the brain in primary developmental stages (46). Several TDCs circulating in the blood form complexes with protein of thyroid hormones and eventually reach the brain and bind with thyroid hormone receptors, disrupting thyroid health (46). TDCs are also responsible for the prevalence of subclinical thyroid conditions known as “subclinical thyroid disease” [SCTD]. In SCTD, the body observes abnormal low or high levels of thyroid-stimulating hormones [TSH] (47). Similarly, BPA can interfere with thyroid hormone action and impairs thyroid functions by inhibiting T3 binding to its receptor and suppressing transcriptional activity mediated by thyroid hormone receptors (48). Phthalates also disturb the normal thyroid system by interfering with gene expression in the HPT axis and metabolic activity (27). It impairs thyroid function through various mechanisms, including inhibition of T3 protein binding, antagonistic interactions, and disruption of throid receptor’s transcriptional activity (49).

Table 1 The effects of MPs on the mammalian thyroid gland.
PBDEs and polybrominated biphenyl [PBBs] as flame retardants in plastic (50) can decrease the circulating level of thyroid hormones and are associated with impaired thyroid function (51). It has been reported that five weeks of rats’ exposure to 1, 3, 6, and 10mg/kg/day of polystyrene nanoplastics [PS-NPs] suppress the serum level of T3, FT3, and FT4 synthesis and circulating level of thyroid hormones (41) (Table 2). Few researchers have demonstrated phthalates association with altered FT4 and total T3 in pregnant women (53). It has been shown that BPA and phthalates cause endocrine toxicity at all levels in animals (45).
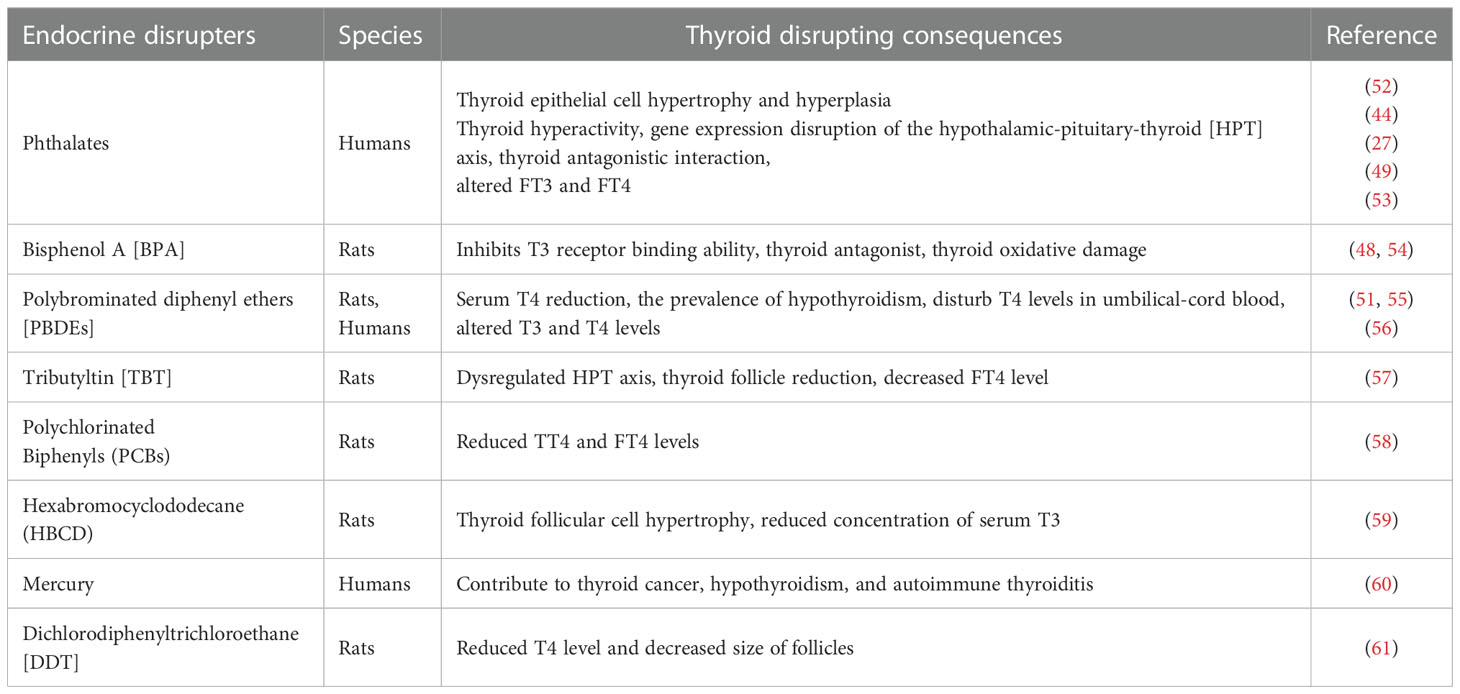
Table 2 The MPs additives effects on the mammalian thyroid gland.
Thyroid hormone levels in pregnant rats and their progeny are altered by brominated flame retardant chemicals, resulting in obesity, heart illness, early puberty, and insulin resistance in their children (62). The PBDEs and PBBs can dissolve in lipids and fats, so their accumulation is easy in wildlife and marine animals once exposed to these chemicals (21). PBDEs reduce the circulating levels of thyroid hormones through changes in T4 binding, and reduction in serum T4 (55). PBDEs disrupt the HPT axis (63) by altering the gene transcriptions of multiple genes like thyroid stimulating hormone subunit [tsh], deiodinase type 2 [deio2], and NK2 homeobox 1 [nkx2.1] (64). These genes, regulating thyroid development and TSH synthesis, are extremely sensitive to PBDEs (64).
3 Effect of MPs, NPs, and associated chemicals on the male reproductive system
Due to the small size of MPs, they can easily enter the organism’s reproductive cells, tissues, and organs altering normal morphology, histology, and physiological functions of the male reproductive system (65). In recent years MPs’ toxicity to the reproductive system got much more attention because they have caused widespread male reproductive abnormalities in mice making them a potential hazard (66) (Table 3). Currently, this environmental issue with respect to mammalian reproductive health is poorly understood (65). However, the harmful effects of MPs on mice’s reproductive health might provide new insight into the deleterious effects of MPs on the mammalian reproductive system.

Table 3 MPs effects on the mammalian male reproductive system.
The limited preliminary studies have suggested MPs bioaccumulation in mammalian testes with subsequent adverse reproductive outcomes (65). MPs ≤10 μm have been observed to accumulate in mice testes reducing testosterone [T] concentration, sperm quality and causes testicular inflammation (66). It also penetrates the testicular tissues such as Leydig cells, germ cells, and Sertoli cells (66). The accumulation of MPs in these cells may profoundly contribute to our understanding of MPs’ effects on mammalian reproductive health.
MPs contaminated with phthalate esters [PAEs] accumulated in the testes and altered testicular weight and sperm physiology by reducing sperm number and vitality (68). MPs cause morphological alternation in sperm like loss of sperm acrosome, cephalic having a small head, acephalia having no head, and tailless sperm (66). Exposure to these plastic particles at developmental stages resulted in shrank germ cells and decreased sperm density in the seminiferous tubules (73). MPs also induces irregular rearrangement of spermatid in testicular seminiferous tubules reducing spermatids’ number (67). A recent study (67) suggested PS-MPs reduce sperm production in mice through testicular injury. Deng et al. (68) observed spermatogenic disruption through altered acid phosphatase [ACP], superoxide dismutase [SOD], and malonaldehyde [MDA] levels in testes. ACP present in Sertoli cells (74) providing structural and nutritional support to spermatogenesis, were significantly increased while SOD and MDA levels were also increased inducing oxidative stress in testes (68). Oxidative stress is the key factor responsible for male infertility due to the increased cell division rate and mitochondrial oxygen consumption in testicular tissues (75). PS-MPs not only cause spermatogenic defects and testicular abnormalities but also penetrate the blood-testis-barrier [BTB] (66) in Sertoli cells and accumulate in the organism testes (30). According to (67) mice exposed to 5 µm of MPs shows a reduction in sperm activity and testicular tissue damage by activating the p38 mitogen-activated protein kinases [MAPK] pathway.
Spermatogenesis is an essential differentiation mechanism that requires T secretion, nutritional support provided by the sertoli cell, and a suitable environment for the production of sperm protected by BTB (66, 76). Following exposure to MPs, the T secretion declined and causes BTB disruption (66) with dysregulated testicular spermatogenesis (67). MPs enter testicular tissues by disrupting BTB, which serves as a physical barrier preventing the penetration of toxic chemicals to the testis, thus providing a healthy environment for spermatogenesis (66, 77) (Table 3). Due to BTB disruption, some proteins expression linked with BTB like Claudin 11, N-Cadherin, Connexin and Occludin were significantly reduced (71). Excess bioaccumulation of MPs induces increased germ cell apoptosis and disrupted spermatogenesis by causing abscission and irregular arrangement of spermatogenic cells (72) and sperm DNA fragmentation; a primary factor responsible for reproductive impairment (30). In mice, MPs exposure resulted in the shedding of spermatogenic cells and the structural disruption of seminiferous tubules (71).
The development and regulation of the reproductive system depend on the HPG axis, which alludes to the connection between the hypothalamus, pituitary, and gonads (78). Reproductive regulation initiates at the hypothalamus level due to gonadotropin-releasing hormone (GnRH) secretion by neurosecretory cells to the hypothalamic-hypophysial system (79). In response to GnRH secretion, pituitary releases FSH and LH that control gonadal functions (79). In males, the HPG axis is responsible for T secretion and regulation of spermatogenesis (80). MPs disrupt the HPG axis (26) as their exposure in male mice reduces the serum concentration of FSH, LH, and T while estradiol level significantly increases (81). Therefore the reproductive abnormalities caused by MPs due to HPG axis disruption include delayed gonadal maturation and the altered ratio of sex hormones that hindered reproductive development (26).
Some studies have shown PS-MPs increase reactive oxygen species [ROS] in male zebrafish liver and gonads. Their exposure to MPs increases apoptosis in testes, affecting gamete production (82) and interact with plasma membrane permeability of gametes, preventing gamete binding and offspring growth (26). In zebrafish testis, silver nanoparticles induce increased cell apoptosis due to overexpression of apoptotic genes like BAX, caspase-3 and caspase-9 (83). MPs also cause the thickness of the basement membrane of the zebrafish testis, due to which the production of spermatozoa is attenuated and undergoes the atrophy of seminiferous tubules (82).
BPA as essential plasticizers causes abnormal spermatogenesis, disruption of BTB, production of poor semen quality, and oxidative stress (84) (Table 4). BPA and BPS consumption alters T secretions and causes cell proliferation by interfering with many receptors (91). PBDEs as persistent flame retardants alter sperm DNA methylation by disrupting the hypothalamic-pituitary-testicular axis, affecting the functional ability of Leydig cells and spermatogenesis (86). MPs contaminated with phthalates can also cause oxidative stress in testes, change the sperm physiology (68), and their esters like dibutyl phthalate [DBP], diethylhexyl phthalate [DEHP], and diisopentyl phthalate [DiPeP] are recognized as anti-androgenic endocrine disrupters (85). Nonylphenol as a persistent pollutant inhibits steroidogenesis and alters enzyme localization like P450 aromatase, 3β-hydroxysteroid dehydrogenase and 17β-hydroxysteroid dehydrogenase in Podarcis Siculus (92). Similarly, DDT causes testicular injury and possible transgenerational effects on epigenomes and transcriptomes of future generations (93). Some anthropogenic EDCs such as cadmium [Cd] and lead [Pb] beyond a certain levels have a close association with infertility in mice and humans (94) (Table 4). Both Cd and Pb adversely alters the HPG axis that harms testicular tissues directly (40) and disrupt spermatogenesis, spermiogenesis, and steroidogenesis (95). Mice administration to varying doses of PSMPs for six weeks induced a significant increase in sperm deformities rate and a decrease in sperm motility (72). PSMPs also cause a reduction in mice’s serum T levels and decrease the functional activity of various enzymes involved in sperm metabolisms such as succinate dehydrogenase and lactase dehydrogenase (72).

Table 4 MPs additives effects on the male mammalian reproductive system.
4 Effects of MPs, NPs, and their associated chemicals on the female reproductive system
Little is known about the harmful effects of MPs and NPs on the ovaries of mammals but studies conducted on rats and aquatic organisms may provide new insights into deadly consequences caused by plastic particles in ovaries. The harmful effects of MPs and their compound substances are linked to a dysfunctional female reproductive system (26) (Table 5).

Table 5 Microplastics effects on the mammalian female reproductive system.
Research has shown MPs accumulation in rat’s ovaries and granulosa cells, reducing the growth of the follicles, decreasing the level of anti-Mullerian hormone [AMH] (99), estradiol, and causing an irregular estrous cycle and abnormal folliculogenesis (101). Granulosa cells are the essential somatic cells of the ovary responsible for normal ovarian development and maturation and play a significant role in folliculogenesis (102). In addition, PS-MPs also induce fibrosis of the ovary through activation of the Wnt/β-Catenin signaling pathway and apoptosis of granulosa cells by oxidative stress, reducing normal ovarian reserve capacity in rats (99, 103). The wnt/β-Catenin signaling pathway is essential for maintaining tissue homeostasis and regulating embryonic maturation, cell proliferation, and apoptosis (104).
MPs as a transport medium for their composite EDCs (105) induce various endocrine disorders like infertility, precocious puberty, hormone-based tumors, several metabolic problems, disruption of granulosa cell steroidogenesis, and polycystic ovary syndrome [PCOS] (106, 107) (Table 6). Endocrine disrupting plastic additives like PBDEs, BPA, phthalates, organotins (20), nonylphenols, octylphenols (113), and biocides like TBT, mercury, arsenic, copper, cadmium, and lead (114) can transfer from pregnant women to the fetal bloodstream through a placental barrier causing neurodevelopmental abnormalities in infants (115). In mice and monkeys, BPA disrupts oocyte development (116) and induces impairment and disruption of steroidogenesis in humans, ovine, swine (107), and murine granulosa cells (117). Several concerns have been raised demonstrating phthalates effect on granulosa cell steroidogenesis in humans (118), mice (108), and rats (119), and increase cell proliferation in swine when exposed to NPs with their composite EDCs (120). Similarly, cadmium disrupts gonadal steroidogenesis and inhibits the binding of FSH to its specific receptor, and alters steroid production of ovarian granulosa cells (121).

Table 6 The effects of MPs additives on the female mammalian reproductive system.
Some aquatic studies have also shown the bioaccumulation of PS-MPs in female fish embryo yolk sacs and female eggs, affecting the normal physiology of offspring in female fish (122, 123). The ability of polystyrene MPs to interfere with plasma proteins connected to oocytes facilitates MPs’ cross-generational transfer (123). In addition, PSMPs delay ovarian development, decline the reproductive ability of marine medaka (26) and reduce superoxide dismutase [SOD], catalase [CAT], glutathione S-transferase [GST], and glutathione peroxidase [GSH-PX] in Oryzias melastigma ovary (99, 123). Similarly, MPs disrupt the HPG axis by down-regulating the transcription of genes like GnRH, vitellogenin [Vtg], and choriogenin [Chg] in the steroidogenesis pathway (26) while its combined exposure with phenanthrene [Phe] increases the accumulation of Phe in the ovaries of marine medaka, disrupting ovarian development and the HPG axis (124).
5 Effects of MPs, NPs, and their additives on the hypothalamus
Hypothalamus is an essential part of the endocrine system that connects the nervous system to the endocrine system and secretes both inhibiting and releasing hormones that signal the pituitary gland to release various important hormones to the whole endocrine system (125). The detrimental consequences of MPs on the mammalian hypothalamus are poorly documented. However, there is clear evidence of mammalian hypothalamic-pituitary axes disruption caused by MPs and their composite EDCs that alter hormonal balance through feedback mechanisms (126, 127) (Table 7).
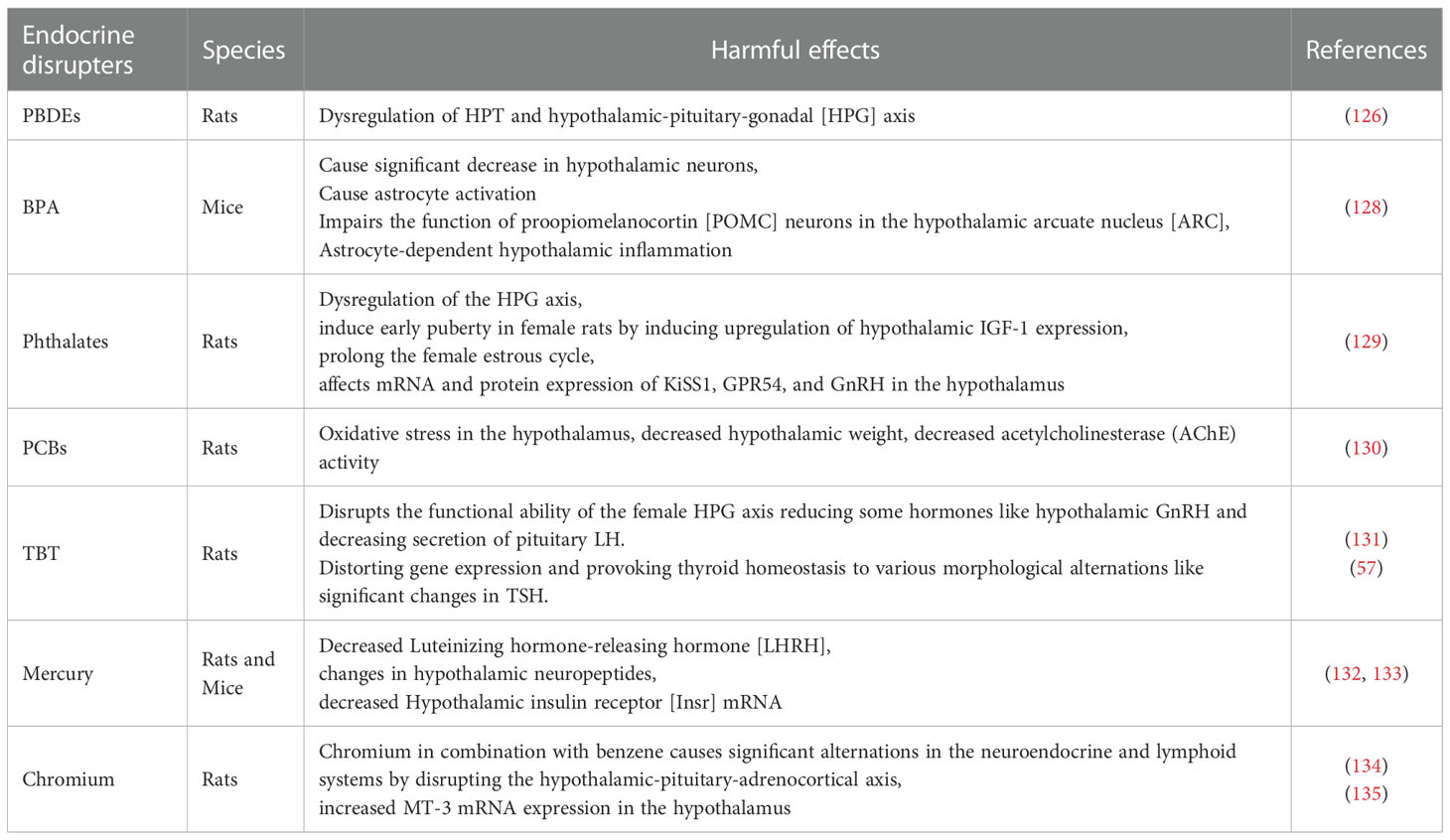
Table 7 MPs additives effects on the mammalian hypothalamus.
MPs and NPs accumulation have been observed in fish brain tissues, crossing the blood-brain barrier and causing neurotoxic effects (136). MPs also decrease hypothalamic kisspeptin level in zebrafish, responsible for their reproduction (137), and interferes with the HPT axis that distorts their gene expression (138).
Plastic additives like BPA and BPS cause decreased hypothalamic neurons and neuroendocrine disruption (139) while polybisphenols disturb the expression of HPT-axis genes (140). Mice exposed to BPA have shown astrocyte activation and various inflammatory actions in the hypothalamus through activation of the toll-like receptor [TLR4], a receptor that plays a vital role in inflammatory responses in the central nervous system (128). The hypothalamic inflammation induced by BPA impairs the function of proopiomelanocortin [POMC] neurons in the hypothalamic arcuate nucleus [ARC] (128). BPA also alters regulatory and inhibitory responses in the HPG axis and its chronic exposure increases expression levels of GnRH1, Kiss1, and FSH in exposed male and female rodents (141). Similarly, BPA induces increased anteroventral periventricular nucleus [AVPV] Kiss1 neurons in male offspring and enhanced Kiss1 cell number in the rostral periventricular area of the third ventricle of female offspring (126).
Phthalates cause hormonal imbalance by interacting with nuclear receptors, hormonal receptors, signaling pathways, and modulate gene expression linked with reproduction thereby, disrupting the HPG axis that affects fertility (142, 143). Due to HPG axis disruption, phthalates alter the levels of GnRH by interacting with genes of G protein-coupled receptors [GPCRs] on pituitary cells and disrupt FSH and LH ratio by interfering with their receptors on Leydig cells, which consequently disrupt the normal activity of steroidogenic enzymes and steroid hormones (142). DEHP exposure induces early puberty in female rats by inducing upregulation of hypothalamic insulin-like Growth Factor-1 [IGF-1] expression which alters normal hormonal levels of growth hormone [GH] and IGF-1 in the hypothalamus (129). DEHP also adversely affects mRNA and protein expression of KiSS1, GPR54, and GnRH in the hypothalamus thereby interfering with the hypothalamic regulatory mechanism affecting normal gonadal development and hypothalamic hormonal balance in pubertal rats (129).
Similarly, nonylphenol disrupts the negative feedback mechanism of the hypothalamic-pituitary-adrenal [HPA] axis by inhibiting estrogen binding with its receptor (28). TBT disrupts the functional ability of the female HPG axis reducing hormones like hypothalamic GnRH and decreasing the secretion of pituitary LH (125). These abnormalities induced by TBT are associated with dysregulated ovarian steroidogenesis, irregular folliculogenesis, oxidative stress, fibrosis, and abnormal alternations in estrogen and testosterone levels (144). Furthermore, TBT is capable of reducing thyroid follicles, distorting gene expression in the HPT axis and provoking thyroid homeostasis to various morphological alternations like changes in TSH and FT4 levels (57).
Mercury exposure causes adverse alternation in the circulating level of some hormones like FSH, LH, inhibin, and androgen, causing reproductive disruption (121, 145) through its pathogenic changes in the HPA axis and HPG axis (146). Studies found a higher concentration of HPA axis hormones in people exposed to heavy metals (147). Some heavy metal present in electronic waste of electronic devices alters the HPA axis and increases the secretion of corticotropin-releasing hormone [CRH], and adrenocorticotropic hormone [ACTH] (147). Lead (Pb) distorts the mechanism of neurotransmission (148) in the brain and causes variations in the regulation of hypothalamic neurotransmitters that affect the functional ability of gonadotropic hormones by influencing the control of the HPG axis (121, 149). Chromium is a neurotoxicant that increases GR activity and metallothionein isoform 3 [MT3] in the hypothalamus (150). Chromium in combination with benzene causes alternations in the neuroendocrine and lymphoid systems by disrupting the hypothalamic-pituitary-adrenocortical axis (134).
6 Effects of MPs, NPs, and their associated chemicals on the pituitary gland
The pituitary gland is a neuroendocrine organ having an essential role in major physiological functions such as growth, sexual development, metabolism, and stress responses (151). Through various regulatory axes, the hypothalamus and pituitary gland regulate neuroendocrine actions including the HPT axis, HPG axis, HPA axis, and HP-somatotrophic axis (126). The hypothalamus-pituitary [HP] axis plays a major integrative role in controlling the mammalian endocrine system. It maintains a balanced homeostatic condition and is responsible for essential hormone secretion that regulates the thyroid, adrenal gland, gonads, somatic growth, and many other functions (126). The hormones secreted by the hypothalamus play a major role in controlling pituitary functions such as metabolism, lactation, growth, and milk secretion (126). Pituitary glands consist of two lobes, the anterior pituitary, and posterior pituitary, linked by the intermediate lobe. The hormones secreted by the anterior pituitary gland into the bloodstream are adrenocorticotrophic hormone, follicle-stimulating hormones, luteinizing hormones, thyroid-stimulating hormone, growth hormone, and prolactin while the posterior pituitary releases oxytocin and antidiuretic hormone (152).
There is a dearth of studies regarding the harmful effects of MPs on the pituitary and we did not find out any research data that have explored the MPs toxicity on the mammalian pituitary gland. However, there is some evidence on the dysregulation of hypothalamic-pituitary axes caused by MPs and their composite EDCs (126, 127) like the HPT axis (138) and HPG axis (127) (Table 8).

Table 8 The effects of [MPs] additives on the mammalian pituitary gland.
HP axis is vulnerable to a variety of MPs composite EDCs (105) such as BPA (162), PCBs, PBDEs, PBBs, dichlorodiphenyltrichloroethane [DDT] (163), and TBT (164) (Table 8). Extensive use of these EDCs in synthetic products, as well as their incorrect disposal, results in a range of environmental contamination, leading to endocrine disruption (126). The consequences of EDCs carried by MPs and NPs (105) on the pituitary gland are the induction of a non-cancerous pituitary tumor known as prolactinoma and stimulation of pituitary hormones like prolactin and TSH (165). The minimal amount of estrogen required to induce tumor is far higher than the normal level so, it is questionable and doubtful whether weak estrogenic disrupters might act as carcinogenic in the pituitary gland (165).
BPA as an essential additive (126) disrupts the regulatory mechanism of the HPT axis through altered TSH levels and affects the pituitary directly by altering its response to Thyrotropin-releasing hormone [TRH] released by the hypothalamus (153).
Mice studies have shown a significant decrease in the expression of pituitary Esr1 with reduced hypothalamic Esr1 expression in rats exposed to 10mg/kg/day diisopentyl phthalate (85). PBDEs as flame retardants alter TH balance at multiple stages of the HPT-axis (154, 155) and exert carcinogenic effects in the thyroid and pituitary of male rats and the uterus of the female rats (154).
Mercury bio-accumulates in the pituitary and thyroid glands and causes endocrine toxicity by altering HP thyroid/gonadal axis (158). Cadmium and arsenic, among the most harmful EDCs, adversely affect the endocrine by altering the secretion of hormones (166).
Mercury inhibits pituitary gland LH and FSH secretion, causing spermatogenesis and sperm count disruption in males and ovarian dysfunction and dysregulated menstruation in females (146). Both Cadmium and arsenic exert xenoestrogenic effects on the interior part of the pituitary gland and reduce LH secretion (167). Arsenic causes neurological abnormalities, and increases mRNA expressions of genes responsible for oxidative responses thus, inducing oxidative stress and apoptosis (168). Similarly, the combined exposure of Pb and cadmium affects the LH and FSH levels in proestrus rats while Pb exposure alone causes a reduction in the fluidity of the pituitary membrane (94).
7 Effects of MPs, NPs, and their associated chemicals on the adrenal gland
The adrenal gland is an essential endocrine gland located on the top of the kidneys and composed of the adrenal cortex and adrenal medulla that release hormones like cortisol, aldosterone, epinephrine, and norepinephrine (169). Like the hypothalamus and pituitary, we did not find any research-based analysis about the direct consequences of MPs on the mammalian adrenal gland, except the findings of Stojanović et al. (170) that have identified an increase in the relative weight of rat’s adrenal gland. However, in zebrafish, PS-NPs affect glucose homeostasis by increasing cortisol secretion and such alternation in cortisol levels causes behavioral changes by interfering with brain cells’ electrical activity that alters important molecules like neurotransmitters, enzymes, and receptors (171).
Toxicological findings have recognized the adrenal gland as the most sensitive organ to EDCs because of the critical role of glucocorticoids secreted by the adrenal cortex in maintaining homeostasis (172) (Table 9). EDCs may disrupt HPA (186), which induces stress responses causing altered behavioral, neuronal, and immune functions while other abnormalities associated with disrupted HPA axis include anxiety, metabolic disorders, and post-traumatic stress disorder [PTSD] (187).
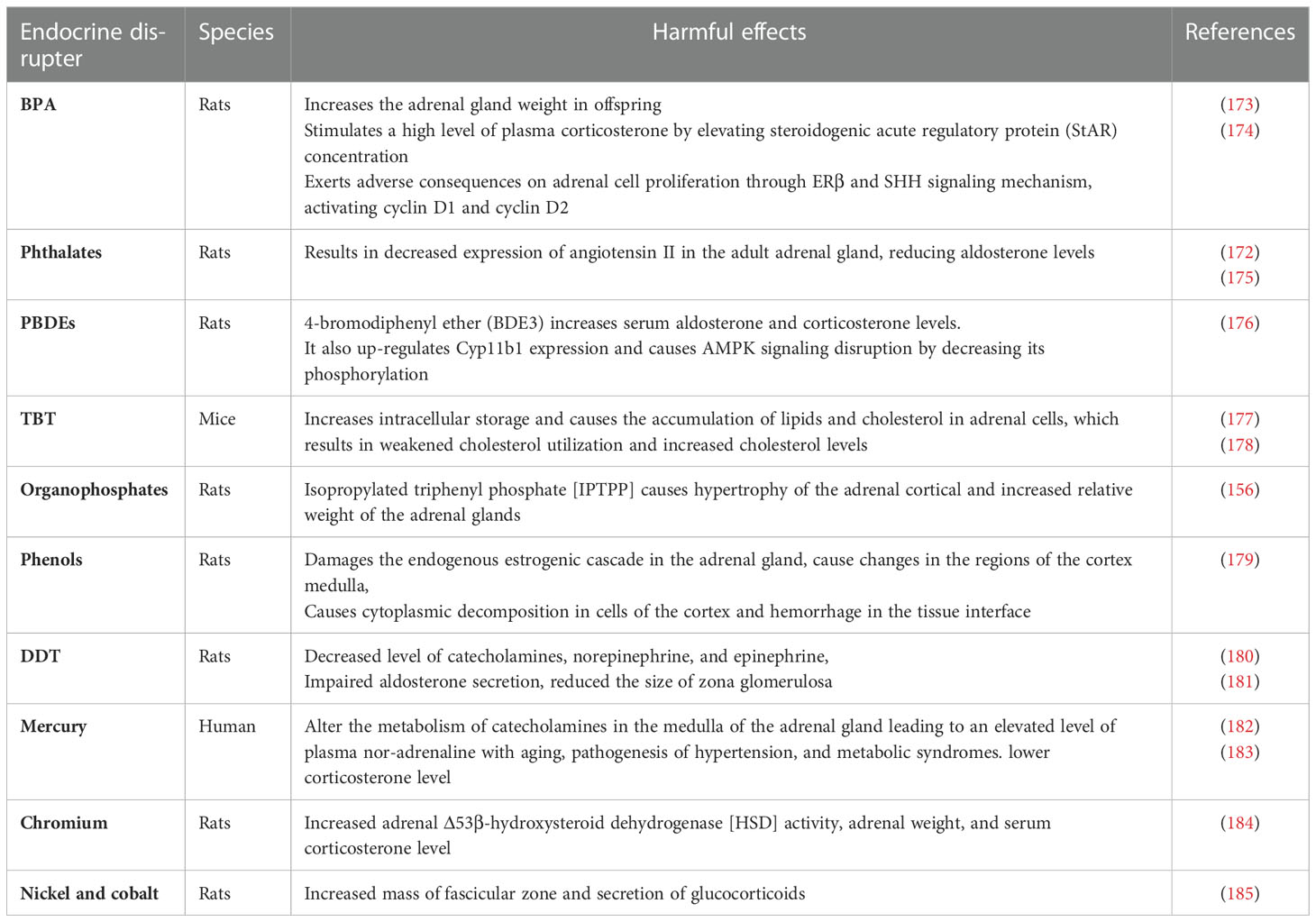
Table 9 The effects of [MPs] additives on the mammalian pituitary gland.
BPA as EDC plays an essential role in the development of non-functional adrenal incidentaloma [NFAI] (174) and causes increased adrenal gland weight in offspring of both male and female rats when exposed to food containing BPA of 25mg/kg (173). BPA causes high level of plasma corticosterone by elevating steroidogenic acute regulatory protein [StAR] concentration (174) and altering adrenal cell proliferation through ERβ and Sonic Hedgehog Signaling [SHH], activating cyclin D1 and cyclin D2 (188). BPA also reduces the immunoreactivity of smooth muscle actin [SMA] in smooth muscles of the adrenal capsule and alters the immunoreactivity of adrenal contractile proteins in rats (189). It has been observed that BPA induces a considerable increase in the adrenal index, vascular congestion, cellular destruction, reduced antioxidant enzymes, and decreased expression of vimentin proteins as well as alpha-smooth muscle actin (189). DEHP as an essential plasticizer is associated with decreased expression of angiotensin II in the adult adrenal gland, reducing aldosterone levels (172). Postpartum exposure to 300 mg/kg of DEHP significantly reduced corticosterone levels while 500 mg/kg of DEHP increases corticosterone and ACTH levels and 10 mg/kg of DEHP triggers glucocorticoid receptor [GR] in the HPA axis, resulting in anxiety-like behavior in premature rats (175). PBDEs, such as 4-bromodiphenyl ether [BDE3] increased the level of serum aldosterone and corticosterone (176). BDE3 up-regulates Cyp11b1 expression and causes AMPK signaling disruption by decreasing its phosphorylation in rats exposed to 200mg/kg of BDE3 (176). TBT is an oxidative endocrine disrupter, which increases intracellular storage (177) and causes the accumulation of lipids and cholesterol in adrenal cells, which results in weakened cholesterol utilization and increased cholesterol levels (178). Organophosphates like isopropylated triphenyl phosphate [IPTPP] cause hypertrophy of the adrenal cortical in zona fasciculate and increase the relative weight of the adrenal gland (156).
Nonyl phenols and octylphenol derived from ethoxylates (190) act as an ED, damaging the endogenous estrogenic cascade in the adrenal gland. Nonylphenol causes adrenal disruption by decreasing the noradrenaline cells leading to lethargy and altering stress response in the body (28). While octyl phenol causes changes in the regions of the cortex medulla, cytoplasmic decomposition in cortex cells, and hemorrhage in the tissue interface of pregnant rats (179). DDT, a widespread ED causes cell atrophy and degenerative effects in the adrenal cortex mainly in the zona fasciculate and zona reticularis (191). DDT can bio-accumulate in the thymus, brain, and even in adipose tissue, and induces impairments of both cortex and medulla of the adrenal gland, disrupting hormonal secretion in cortical and chromaffin cells as well as suppressing the thyroxine hydroxylase production in chromaffin cells (191).
The adrenal gland is also vulnerable to heavy metals like mercury, cadmium, cobalt, and copper that affect the zona glomerulosa of the rat adrenal gland (192) and dysregulate HPA axis, altering the hormonal secretion of corticosterone in response to various stressors and interfering with steroid hormones metabolism (183). Among these toxic metals, mercury alters the metabolism of catecholamines in the adrenal medulla, leading to an increased level of plasma nor-adrenaline with aging. Its chronic exposure is associated with the pathogenesis of hypertension and metabolic syndromes (183).
8 Conclusion
The abundance and distribution of MPs derived from plastic degradation across the globe are so extensive that we can claim of living in a plastic world. Endocrine toxicity induced by MPs is an emerging issue, despite the subject being rarely documented, there is growing evidence for ingested MPs bioaccumulation in mammalian tissues and organs with deleterious outcomes including endocrine abnormalities, reproductive toxicity, gut microbiota dysbiosis, and defective immunological responses in rodents, rats and mice. Various EDCs or toxic chemicals present in plastic as an additive or adsorbed by MPs enter the body easily, acting as agonists or antagonists for a wide range of hormonal receptors, and induce endocrine toxicity. The identification of adverse consequences of MPs on the mammalian endocrine system is a great challenge due to their rising levels in both terrestrial and aquatic ecosystems. However, there are still no conclusive research reports that have determined the direct consequences of MPs and NPs on the hypothalamus, pituitary, and adrenal gland. So further research studies are essential to be performed to determine the potential hazards of MPs and NPs to regulate laws that reduce exposure to these small plastic particles.
Author contributions
Conceptualization, SU (1st author) and GN. methodology, SU (1st author) and GN. writing—original draft preparation, SU (1st author), SaU, and GN. writing—review and editing, SU (1st author), SA, XG, SaU, SU (5th author), GN. funding acquisition, XG, KW. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by the Natural Science Foundation of Qinghai Province (2022-ZJ-936Q) and Qinghai Kunlun Ying Cai Action Project No. Qing Ren Zi (2020) 18.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Deonie A, Allen S, Abbasi S, Baker A, Bergmann M, Brahney J, et al. Microplastics and nanoplastics in the marine-atmosphere environment. Nat Rev Earth Environ (2022) 3:1–13. doi: 10.1038/s43017-022-00292-x
2. Reinhold L, Wagreich M, Williams M, Wolfe AP. The geological cycle of plastics and their use as a stratigraphic indicator of the Anthropocene. Anthropocene (2016) 13:4–17. doi: 10.1016/j.ancene.2016.01.002
4. Roland G, Jambeck JR, Law KL. Production, use, and fate of all plastics ever made. Sci Adv (2017) 3(7):e1700782. doi: 10.1126/sciadv.1700782
5. Chris W, Sebille EV, Hardesty BD. Threat of plastic pollution to seabirds is global, pervasive, and increasing. Proc Natl Acad Sci (2015) 112(38):11899–904. doi: 10.1073/pnas.1502108112
6. Davidson AD, Shoemaker KT, Weinstein B, Costa GC, Brooks TM, Ceballos G, et al. Geography of current and future global mammal extinction risk. PloS One (2017) 12(11):e0186934. doi: 10.1371/journal.pone.0186934
7. Santos RG, Machovsky-Capuska GE, Andrades R. Plastic ingestion as an evolutionary trap: Toward a holistic understanding. Science (2021) 373(6550):56–60. doi: 10.1126/science.abh0945
8. Ghulam N, Ahmad S, Ullah S, Zada S, Sarfraz M, Guo X, et al. The adverse health effects of increasing microplastic pollution on aquatic mammals. J King Saud University-Science (2022) 34(4):102006. doi: 10.1016/j.jksus.2022.102006
9. Galloway TS, Cole M, Lewis C. Interactions of microplastic debris throughout the marine ecosystem. Nat Ecol Evol (2017) 1(5):1–8. doi: 10.1038/s41559-017-0116
10. Benson NU, Agboola OD, Fred-Ahmadu OH, De-la-Torre GE, Oluwalana A, Williams A. Micro (Nano) plastics prevalence, food web interactions and toxicity assessment in aquatic organisms: A review. Front Mar Sci (2022) 9:291. doi: 10.3389/fmars.2022.851281
11. Kershaw PJ, Rochman CM. Sources, fate and effects of microplastics in the marine environment: part 2 of a global assessment. Reports and Studies-IMO/FAO/Unesco-IOC/WMO/IAEA/UN/UNEP Joint Group of Experts on the Scientific Aspects of Marine Environmental Protection (GESAMP) (2015) Eng No. 93
12. Jaikumar G, Brun NR, Vijver MG, Bosker T. Reproductive toxicity of primary and secondary microplastics to three cladocerans during chronic exposure. Environ pollut (2019) 249:638–46. doi: 10.1016/j.envpol.2019.03.085
13. Baho DL, Bundschuh M, Futter MN. Microplastics in terrestrial ecosystems: Moving beyond the state of the art to minimize the risk of ecological surprise. Global Change Biol (2021) 27(17):3969–86. doi: 10.1111/gcb.15724
14. Atsuhiko I, Azuma T, Cordova MR, Cózar A, Galgani F, Hagita R, et al. A multilevel dataset of microplastic abundance in the world’s upper ocean and the laurentian great lakes. Microplastics Nanoplastics (2021) 1(1):1–14. doi: 10.1186/s43591-021-00013-z
15. Jambeck JR, Geyer R, Wilcox C, Siegler TR, Perryman M, Andrady A, et al. Plastic waste inputs from land into the ocean. Science (2015) 347(6223):768–71. doi: 10.1126/science.1260352
17. Luca N, Futter M, Langaas S. Environmental Science & Technology (2016) 50(20):10777–9. doi: 10.1021/acs.est.6b04140
18. Laura R, Marcos R, Hernández A. Potential adverse health effects of ingested micro-and nanoplastics on humans. lessons learned from in vivo and in vitro mammalian models. J Toxicol Environ Health Part B (2020) 23(2):51–68. doi: 10.1080/10937404.2019.1700598
19. Helene W, Wang Z, Hellweg S. Deep dive into plastic monomers, additives, and processing aids. Environ Sci Technol (2021) 55(13):9339–51. doi: 10.1021/acs.est.1c00976
20. Chen Q, Allgeier A, Yin D, Hollert H. Leaching of endocrine disrupting chemicals from marine microplastics and mesoplastics under common life stress conditions. Environ Int (2019) 130:104938. doi: 10.1016/j.envint.2019.104938
21. Zoeller TR. Environmental chemicals targeting thyroid. Hormones (2010) 9(1):28–40. doi: 10.14310/horm.2002.1250
22. Kurunthachalam K, Vimalkumar K. A review of human exposure to microplastics and insights into microplastics as obesogens. Front Endocrinol (2021) 12:978. doi: 10.3389/fendo.2021.724989
23. Campanale C, Massarelli C, Savino I, Locaputo V, Uricchio VF. A detailed review study on potential effects of microplastics and additives of concern on human health. Int J Environ Res Public Health (2020) 17(4):1212. doi: 10.3390/ijerph17041212
24. Yilmaz B, Terekeci H, Sandal S, Kelestimur F. Endocrine disrupting chemicals: Exposure, effects on human health, mechanism of action, models for testing and strategies for prevention. Rev endocr Metab Disord (2020) 21(1):127–47. doi: 10.1007/s11154-019-09521-z
25. Schwabl P, Köppel S, Königshofer P, Bucsics T, Trauner M, Reiberger T, et al. Detection of various microplastics in human stool: A prospective case series. Ann Internal Med (2019) 171(7):453–57. doi: 10.7326/M19-0618
26. Wang J, Li Y, Lu L, Zheng M, Zhang X, Tian H, et al. Polystyrene microplastics cause tissue damages, sex-specific reproductive disruption and transgenerational effects in marine medaka (Oryzias melastigma). Environ pollut (2019) 254:113024. doi: 10.1016/j.envpol.2019.113024
27. Derakhshan A, Shu H, Broeren MAC, Lindh CH, Peeters RP, Kortenkamp A, et al. Association of phthalate exposure with thyroid function during pregnancy. Environ Int (2021) 157:106795. doi: 10.1016/j.envint.2021.106795
28. De Falco M, Sellitti A, Sciarrillo R, Capaldo A, Valiante S, Iachetta G, et al. Nonylphenol effects on the hpa axis of the bioindicator vertebrate, Podarcis sicula lizard. Chemosphere (2014) 104:190–96. doi: 10.1016/j.chemosphere.2013.11.014
29. Van Sebille E, Wilcox C, Lebreton L, Maximenko N, Hardesty BD, Van Franeker JA, et al. A global inventory of small floating plastic debris. Environ Res Lett (2015) 10(12):124006. doi: 10.1088/1748-9326/10/12/124006
30. Amereh F, Babaei M, Eslami A, Fazelipour S, Rafiee M. The emerging risk of exposure to nano (Micro) plastics on endocrine disturbance and reproductive toxicity: From a hypothetical scenario to a global public health challenge. Environ pollut (2020) 261:114158. doi: 10.1016/j.envpol.2020.114158
31. De Sá LC, Oliveira M, Ribeiro F, Lopes Rocha T, Futter MN. Studies of the effects of microplastics on aquatic organisms: What do we know and where should we focus our efforts in the future? Sci total Environ (2018) 645:1029–39. doi: 10.1016/j.scitotenv.2018.07.207
32. Hu L, Zhao Y, Xu H. Trojan Horse in the intestine: A review on the biotoxicity of microplastics combined environmental contaminants. J Hazard Mater (2022) 439:129652. doi: 10.1016/j.jhazmat.2022.129652
33. Schell T, Rico A, Cherta L, Nozal L, Dafouz R, Giacchini R, et al. Influence of microplastics on the bioconcentration of organic contaminants in fish: Is the “Trojan horse” effect a matter of concern? Environ pollut (2022) 306:119473. doi: 10.1016/j.envpol.2022.119473
34. Alava JJ. Modeling the bioaccumulation and biomagnification potential of microplastics in a cetacean foodweb of the northeastern pacific: A prospective tool to assess the risk exposure to plastic particles. Front Mar Sci (2020) 7:793. doi: 10.3389/fmars.2020.566101
35. De Felice B, Sabatini V, Antenucci S, Gattoni G, Santo N, Bacchetta R, et al. Polystyrene microplastics ingestion induced behavioral effects to the Cladoceran daphnia magna. Chemosphere (2019) 231:423–31. doi: 10.1016/j.chemosphere.2019.05.115
36. Banerjee A, Shelver WL. Micro-and nanoplastic-mediated pathophysiological changes in rodents, rabbits, and chickens: A review. J Food Prot (2021) 84(9):1480–95. doi: 10.4315/JFP-21-117
37. Bernal J. Thyroid hormone receptors in brain development and function. Nat Clin Pract Endocrinol Metab (2007) 3(3):249–59. doi: 10.1038/ncpendmet0424
38. Shahid MA, Ashraf MA, Sharma S. Physiology, Thyroid Hormone. In: StatPearls. StatPearls Publishing, Treasure Island (FL); (2022). Available at: https://www.ncbi.nlm.nih.gov/books/NBK500006/.
39. Kloas W, Urbatzka R, Opitz R, Würtz S, Behrends T, Hermelink B, et al. Endocrine disruption in aquatic vertebrates. Ann New York Acad Sci (2009) 1163(1):187–200. doi: 10.1111/j.1749-6632.2009.04453.x
40. Saradha B, Mathur PP. Effect of environmental contaminants on Male reproduction. Environ Toxicol Pharmacol (2006) 21(1):34–41. doi: 10.1016/j.etap.2005.06.004
41. Amereh F, Eslami A, Fazelipour S, Rafiee M, Zibaii MI, Babaei M. Thyroid endocrine status and biochemical stress responses in adult Male wistar rats chronically exposed to pristine polystyrene nanoplastics. Toxicol Res (2019) 8(6):953–63. doi: 10.1039/c9tx00147f
42. Kim J, Maruthupandy M, An KS, Lee KH, Jeon S, Kim J-S, et al. Acute and subacute repeated oral toxicity study of fragmented microplastics in sprague-dawley rats. Ecotoxicol Environ Saf (2021) 228:112964. doi: 10.1016/j.ecoenv.2021.112964
43. Gallo F, Fossi C, Weber R, Santillo D, Sousa J, Ingram I, et al. Marine litter plastics and microplastics and their toxic chemicals components: The need for urgent preventive measures. Environ Sci Europe (2018) 30(1):1–14. doi: 10.1186/s12302-018-0139-z
44. Boas M, Frederiksen H, Feldt-Rasmussen U, Skakkebæk NE, Hegedüs L, Hilsted L, et al. Childhood exposure to phthalates: Associations with thyroid function, insulin-like growth factor I, and growth. Environ Health Perspect (2010) 118(10):1458–64. doi: 10.1289/ehp.0901331
45. Mathieu-Denoncourt J, Wallace SJ, de Solla SR, Langlois VS. Plasticizer endocrine disruption: Highlighting developmental and reproductive effects in mammals and non-mammalian aquatic species. Gen Comp Endocrinol (2015) 219:74–88. doi: 10.1016/j.ygcen.2014.11.003
46. Andra SS, Makris KC. Thyroid disrupting chemicals in plastic additives and thyroid health. J Environ Sci Health Part C (2012) 30(2):107–51. doi: 10.1080/10590501.2012.681487
47. Ayala AR, Danese MD, Ladenson PW. When to treat mild hypothyroidism. Endocrinol Metab Clinics (2000) 29(2):399–415. doi: 10.1016/S0889-8529(05)70139-0
48. Moriyama K, Tagami T, Akamizu T, Usui T, Saijo M, Kanamoto N, et al. Thyroid hormone action is disrupted by bisphenol a as an antagonist. J Clin Endocrinol Metab (2002) 87(11):5185–90. doi: 10.1210/jc.2002-020209
49. Ishihara A, Sawatsubashi S, Yamauchi K. Endocrine disrupting chemicals: Interference of thyroid hormone binding to transthyretins and to thyroid hormone receptors. Mol Cell Endocrinol (2003) 199(1-2):105–17. doi: 10.1016/S0303-7207(02)00302-7
50. Shao M, Jiang J, Li M, Wu L, Hu M. Recent developments in the analysis of polybrominated diphenyl ethers and polybrominated biphenyls in plastic. Rev Anal Chem (2016) 35(3):133–43. doi: 10.1515/revac-2016-0012
51. Oulhote Y, Chevrier J, Bouchard MF. Exposure to polybrominated diphenyl ethers (Pbdes) and hypothyroidism in Canadian women. J Clin Endocrinol Metab (2016) 101(2):590–98. doi: 10.1210/jc.2015-2659
52. Liu C, Zhao L, Wei L, Li L. Dehp reduces thyroid hormones Via interacting with hormone synthesis-related proteins, deiodinases, transthyretin, receptors, and hepatic enzymes in rats. Environ Sci pollut Res (2015) 22(16):12711–19. doi: 10.1007/s11356-015-4567-7
53. Huang P-C, Tsai C-H, Liang W-Y, Li S-S, Huang H-B, Kuo P-L. Early phthalates exposure in pregnant women is associated with alteration of thyroid hormones. PloS One (2016) 11(7):e0159398. doi: 10.1371/journal.pone.0159398
54. Mohammed ET, Khalid S H, Ahmed AE, Tarek Aly M, Aleya L, Abdel-Daim MM. Ginger extract ameliorates bisphenol a (Bpa)-induced disruption in thyroid hormones synthesis and metabolism: Involvement of nrf-2/Ho-1 pathway. Sci Total Environ (2020) 703:134664. doi: 10.1016/j.scitotenv.2019.134664
55. Stoker TE, Laws SC, Crofton KM, Hedge JM, Ferrell JM, Cooper RL. Assessment of de-71, a commercial polybrominated diphenyl ether (Pbde) mixture, in the edsp Male and female pubertal protocols. Toxicol Sci (2004) 78(1):144–55. doi: 10.1093/toxsci/kfh029
56. Abdelouahab N, Langlois M-F, Lavoie L, Corbin F, Pasquier J-C, Takser L. Maternal and cord-blood thyroid hormone levels and exposure to polybrominated diphenyl ethers and polychlorinated biphenyls during early pregnancy. Am J Epidemiol (2013) 178(5):701–13. doi: 10.1093/aje/kwt141
57. Andrade MN, Santos-Silva AP, Rodrigues-Pereira P, Paiva-Melo FD, Correa de Lima NJ, Pires Teixeira M, et al. The environmental contaminant tributyltin leads to abnormalities in different levels of the hypothalamus-Pituitary-Thyroid axis in female rats. Environ pollut (2018) 241:636–45. doi: 10.1016/j.envpol.2018.06.006
58. Hallgren S, Sinjari T, Håkansson H, Darnerud P. Effects of polybrominated diphenyl ethers (Pbdes) and polychlorinated biphenyls (Pcbs) on thyroid hormone and vitamin a levels in rats and mice. Arch Toxicol (2001) 75(4):200–08. doi: 10.1007/s002040000208
59. Saegusa Y, Fujimoto H, Woo G-H, Inoue K, Takahashi M, Mitsumori TK, Hirose M, Nishikawa A, Shibutani M, et al. Developmental toxicity of brominated flame retardants, tetrabromobisphenol a and 1, 2, 5, 6, 9, 10-hexabromocyclododecane, in rat offspring after maternal exposure from mid-gestation through lactation. Reprod Toxicol (2009) 28(4):456–67. doi: 10.1016/j.reprotox.2009.06.011
60. Pamphlett R, Doble PA, Bishop DP. Mercury in the human thyroid gland: Potential implications for thyroid cancer, autoimmune thyroiditis, and hypothyroidism. PloS One (2021) 16(2):e0246748. doi: 10.1371/journal.pone.0246748
61. Yaglova NV, Yaglov VV. Cytophysiological changes in the follicular epithelium of the thyroid gland after long-term exposure to low doses of dichlorodiphenyltrichloroethane (Ddt). Bull Exp Biol Med (2017) 162(5):699–702. doi: 10.1007/s10517-017-3691-4
62. Gore AC, Crews D, Doan LL, La Merrill M, Patisaul H, Zota A. Introduction to endocrine disrupting chemicals (Edcs). A guide Public interest organ policy-makers (2014), 21–2.
63. Yu L, Han Z, Liu C. A review on the effects of PBDEs on thyroid and reproduction systems in fish. Gen Comp Endocrinol (2015) 219:64–73. doi: 10.1016/j.ygcen.2014.12.010
64. Yu L, Deng J, Shi X, Liu C, Yu K, Zhou B. Exposure to DE-71 alters thyroid hormone levels and gene transcription in the hypothalamic–pituitary–thyroid axis of zebrafish larvae. Aquat Toxicol (2010) 97(3):226–33. doi: 10.1016/j.aquatox.2009.10.022
65. D’Angelo S, Meccariello R. Microplastics: A threat for Male fertility. Int J Environ Res Public Health (2021) 18(5):2392. doi: 10.3390/ijerph18052392
66. Jin H, Ma T, Sha X, Liu Z, Zhou Y, Meng X, et al. Polystyrene microplastics induced Male reproductive toxicity in mice. J hazard mater (2021) 401:123430. doi: 10.1016/j.jhazmat.2020.123430
67. Hou B, Wang F, Liu T, Wang Z. Reproductive toxicity of polystyrene microplastics: In vivo experimental study on testicular toxicity in mice. J hazard mater (2021) 405:124028. doi: 10.1016/j.jhazmat.2020.124028
68. Deng Y, Yan Z, Shen R, Huang Y, Ren H, Zhang Y. Enhanced reproductive toxicities induced by phthalates contaminated microplastics in Male mice (Mus musculus). J hazard mater (2021) 406:124644. doi: 10.1016/j.jhazmat.2020.124644
69. Wang X, Zhang X, Sun K, Wang S, Gong D. Polystyrene microplastics induce apoptosis and necroptosis in swine testis cells Via Ros/Mapk/Hif1α pathway. Environ Toxicol (2022) 37(10):2483–92. doi: 10.1002/tox.23611
70. Deng Y, Chen H, Huang Y, Wang Q, Chen W, Chen D. Polystyrene microplastics affect the reproductive performance of Male mice and lipid homeostasis in their offspring. Environ Sci Technol Lett (2022) 9(9):752–57. doi: 10.1021/acs.estlett.2c00262
71. Li S, Wang Q, Yu H, Yang L, Sun Y, Xu N, et al. Polystyrene microplastics induce blood–testis barrier disruption regulated by the mapk-Nrf2 signaling pathway in rats. Environ Sci pollut Res (2021) 28(35):47921–31. doi: 10.1007/s11356-021-13911-9
72. Xie X, Ting D, Duan J, Xie J, Yuan J, Chen M. Exposure to polystyrene microplastics causes reproductive toxicity through oxidative stress and activation of the P38 mapk signaling pathway. Ecotoxicol Environ Saf (2020) 190:110133. doi: 10.1016/j.ecoenv.2019.110133
73. Huang T, Zhang W, Lin T, Liu S, Sun Z, Liu F, et al. Maternal exposure to polystyrene nanoplastics during gestation and lactation induces hepatic and testicular toxicity in Male mouse offspring. Food Chem Toxicol (2022) 160:112803. doi: 10.1016/j.fct.2021.112803
74. Peruquetti RL, Taboga SR, Vilela de Azeredo Oliveira MT. Expression of acid phosphatase in the seminiferous epithelium of zebrrates. Genet Mol Res (2010) 9:620–28. doi: 10.4238/vol9-2gmr730
75. Asadi N, Bahmani M, Kheradmand A, Rafieian-Kopaei M. The impact of oxidative stress on testicular function and the role of antioxidants in improving it: A review. J Clin Diagn res: JCDR (2017) 11(5):IE01. doi: 10.7860/JCDR/2017/23927.9886
76. C Yan C, Mruk DD. The blood-testis barrier and its implications for Male contraception. Pharmacol Rev (2012) 64(1):16–64. doi: 10.1124/pr.110.002790
77. Mruk DD, Cheng CY. The mammalian blood-testis barrier: Its biology and regulation. Endocr Rev36(5) (2015) 36:564–91. doi: 10.1210/er.2014-1101
78. Peper JS, Brouwer RM, van Leeuwen M, Schnack HG, Boomsma DI, Kahn RS, et al. Hpg-axis hormones during puberty: A study on the association with hypothalamic and pituitary volumes. Psychoneuroendocrinology (2010) 35(1):133–40. doi: 10.1016/j.psyneuen.2009.05.025
79. Kurylowicz A. Endocrine disorders accompanying obesity-effect or cause? Role Obes Hum Health Dis (2021) 13. doi: 10.5772/intechopen.98793
80. Dwyer AA, Quinton R. Anatomy and physiology of the hypothalamic-Pituitary-Gonadal (Hpg) axis. In Advanced Pract Endocrinol Nurs (2019), 839–52. doi: 10.1007/978-3-319-99817-6_43
81. Wei Z, Wang Y, Wang S, Xie J, Han Q, Chen M. Comparing the effects of polystyrene microplastics exposure on reproduction and fertility in Male and female mice. Toxicology (2022) 465:153059. doi: 10.1016/j.tox.2021.153059
82. Qiang L, Cheng J. Exposure to polystyrene microplastics impairs gonads of zebrafish (Danio rerio). Chemosphere (2021) 263:128161. doi: 10.1016/j.chemosphere.2020.128161
83. Ma Y-B, Lu C-J, Junaid M, Jia P-P, Yang L, Zhang J-H, et al. Potential adverse outcome pathway (Aop) of silver nanoparticles mediated reproductive toxicity in zebrafish. Chemosphere (2018) 207:320–28. doi: 10.1016/j.chemosphere.2018.05.019
84. Chioccarelli T, Manfrevola F, Migliaccio M, Altucci L, Porreca V, Fasano S, et al. Fetal-perinatal exposure to bisphenol-a affects quality of spermatozoa in adulthood mouse. Int J Endocrinol (2020) 2020. doi: 10.1155/2020/2750501
85. da Silva N, Tatiana G, Curi Z, Lima Tolouei SE, Tapias Passoni M, Sari Hey GB, et al. Effects of diisopentyl phthalate exposure during gestation and lactation on hormone-dependent behaviours and hormone receptor expression in rats. J Neuroendocrinol (2019) 31(12):e12816. doi: 10.1111/jne.12816
86. Yu Y-j, Lin B-g, Chen X-c, Qiao J, Li L-z, Liang Y, et al. Polybrominated diphenyl ethers in human serum, semen and indoor dust: Effects on hormones balance and semen quality. Sci Total Environ (2019) 671:1017–25. doi: 10.1016/j.scitotenv.2019.03.319
87. Kanimozhi V, Palanivel K, Akbarsha MA, Kadalmani B. Molecular mechanisms of tributyltin-induced alterations in cholesterol homeostasis and steroidogenesis in hamster testis: In vivo and in vitro studies. J Cell Biochem (2018) 119(5):4021–37. doi: 10.1002/jcb.26564
88. Williams RS, Curnick DJ, Brownlow A, Barber JL, Barnett J, Davison NJ, et al. Polychlorinated biphenyls are associated with reduced testes weights in harbour porpoises (Phocoena phocoena). Environ Int (2021) 150:106303. doi: 10.1016/j.envint.2020.106303
89. Kuriyama SN, Chahoud I. In utero exposure to low-dose 2, 3′, 4, 4′, 5-pentachlorobiphenyl (Pcb 118) impairs Male fertility and alters neurobehavior in rat offspring. Toxicology (2004) 202(3):185–97. doi: 10.1016/j.tox.2004.05.006
90. Massányi P, Massányi M, Madeddu R, Stawarz R, Lukáč N. Effects of cadmium, lead, and mercury on the structure and function of reproductive organs. Toxics (2020) 8(4):94. doi: 10.3390/toxics8040094
91. Ullah A, Pirzada M, Jahan S, Ullah H, Shaheen G, Rehman H, et al. Bisphenol a and its analogs bisphenol b, bisphenol f, and bisphenol s: Comparative in vitro and in vivo studies on the sperms and testicular tissues of rats. Chemosphere (2018) 209:508–165. doi: 10.1016/j.chemosphere.2018.06.089
92. Di Lorenzo M, Mileo A, Laforgia V, De Falco M, Rosati L. Alkyphenol exposure alters steroidogenesis in male lizard Podarcis siculus. Animals (2021) 11(4):1003. doi: 10.3390/ani11041003
93. Burgos-Aceves MA, Migliaccio V, Di Gregorio I, Paolella G, Lepretti M, Faggio C, et al. 1, 1, 1-trichloro-2, 2-bis (p-chlorophenyl)-ethane (DDT) and 1, 1-Dichloro-2, 2-bis (p, p’-chlorophenyl) ethylene (DDE) as endocrine disruptors in human and wildlife: A possible implication of mitochondria. Environ Toxicol Pharmacol (2021) 87:103684. doi: 10.1016/j.etap.2021.103684
94. Plunk EC, Richards SM. Endocrine-disrupting air pollutants and their effects on the hypothalamus-Pituitary-Gonadal axis. Int J Mol Sci (2020) 21(23):9191. doi: 10.3390/ijms21239191
95. Pandya C, Pillai P, Nampoothiri LP, Bhatt N, Gupta S, Gupta S. Effect of lead and cadmium Co-exposure on testicular steroid metabolism and antioxidant system of adult Male rats. Andrologia (2012) 44:813–225. doi: 10.1111/j.1439-0272.2010.01137.x
96. Liu Z, Zhuan Q, Zhang L, Meng L, Fu X, Hou Y. Polystyrene microplastics induced female reproductive toxicity in mice. J hazard mater (2022) 424:127629. doi: 10.1016/j.jhazmat.2021.127629
97. Park E-J, Han J-S, Park E-J, Seong E, Lee G-H, Kim D-W, et al. Repeated-oral dose toxicity of polyethylene microplastics and the possible implications on reproduction and development of the next generation. Toxicol Lett (2020) 324:75–85. doi: 10.1016/j.toxlet.2020.01.008
98. Hu J, Qin X, Zhang J, Zhu Y, Zeng W, Lin Y, et al. Polystyrene microplastics disturb maternal-fetal immune balance and cause reproductive toxicity in pregnant mice. Reprod Toxicol (2021) 106:42–50. doi: 10.1016/j.reprotox.2021.10.002
99. An R, Xifeng W, Yang L, Zhang J, Wang N, Xu F, et al. Polystyrene microplastics cause granulosa cells apoptosis and fibrosis in ovary through oxidative stress in rats. Toxicology (2021) 449:152665. doi: 10.1016/j.tox.2020.152665
100. Hou J, Lei Z, Cui L, Hou Y, Yang L, An R, et al. Polystyrene microplastics lead to pyroptosis and apoptosis of ovarian granulosa cells Via Nlrp3/Caspase-1 signaling pathway in rats. Ecotoxicol Environ Saf (2021) 212:112012. doi: 10.1016/j.ecoenv.2021.112012
101. Haddadi A, Kessabi K, Boughammoura S, Ben Rhouma M, Mlouka R, Banni M, et al. Exposure to microplastics leads to a defective ovarian function and change in cytoskeleton protein expression in rat. Environ Sci pollut Res (2022) 29:1–13. doi: 10.1007/s11356-021-18218-3
102. Tu J, Hoi-Hung Cheung A, Leung-Kwok Chan C, Chan W-Y. The role of micrornas in ovarian granulosa cells in health and disease. Front Endocrinol (2019) 10:174. doi: 10.3389/fendo.2019.00174
103. Li X, Zhang T, Lv W, Wang H, Chen H, Xu Q, et al. Intratracheal administration of polystyrene microplastics induces pulmonary fibrosis by activating oxidative stress and Wnt/B-catenin signaling pathway in mice. Ecotoxicol Environ Saf (2022) 232:113238. doi: 10.1016/j.ecoenv.2022.113238
104. Silva-García O, Valdez-Alarcón JJ, Baizabal-Aguirre VM. Wnt/B-catenin signaling as a molecular target by pathogenic bacteria. Front Immunol (2135) 2019). doi: 10.3389/fimmu.2019.02135
105. Laskar N, Upendra K. Plastics and microplastics: A threat to environment. Environ Technol Innovation (2019) 14:100352. doi: 10.1016/j.eti.2019.100352
106. Konieczna A, Rutkowska A, Rachon D. Health risk of exposure to bisphenol a (Bpa). Roczniki Państwowego Zakładu Higieny (2015) 66(1):5–11.
107. Amar S, Binet A, Téteau O, Desmarchais A, Papillier P, Lacroix M, et al. Bisphenol s impaired human granulosa cell steroidogenesis in vitro. Int J Mol Sci (2020) 21(5):1821. doi: 10.3390/ijms21051821
108. Lai F-N, Liu J-C, Li L, Ma J-Y, Liu X-L, Liu Y-P, et al. Di (2-ethylhexyl) phthalate impairs steroidogenesis in ovarian follicular cells of prepuberal mice. Arch Toxicol (2017) 91(3):1279–92. doi: 10.1007/s00204-016-1790-z
109. Shi X, Wang X, Peng L, Chen Y, Liu C, Yang Q, et al. Associations between polybrominated diphenyl ethers (Pbdes) levels in adipose tissues and female menstrual cycle and menstrual bleeding duration in shantou, China. Environ pollut (2022) 301:119025. doi: 10.1016/j.envpol.2022.119025
110. de Araújo JFP, Podratz PL, Sena GC, Merlo E, Freitas-Lima LC, Mori Ayub JG, et al. The obesogen tributyltin induces abnormal ovarian adipogenesis in adult female rats. Toxicol Lett (2018) 295:99–114. doi: 10.1016/j.toxlet.2018.06.1068
111. Pocar P, Fiandanese N, Secchi C, Berrini A, Fischer B, Schmidt J-S, et al. Effects of polychlorinated biphenyls in cd-1 mice: Reproductive toxicity and intergenerational transmission. Toxicol Sci (2012) 126(1):213–26. doi: 10.1093/toxsci/kfr327
112. Steinberg RM, Walker DM, Juenger TE, Woller MJ, Gore AC. Effects of perinatal polychlorinated biphenyls on adult female rat reproduction: Development, reproductive physiology, and second generational effects. Biol Reprod (2008) 78(6):1091–101. doi: 10.1095/biolreprod.107.067249
113. Loyo-Rosales JE, Rosales-Rivera GC, Lynch AM, Rice CP, Torrents A. Migration of nonylphenol from plastic containers to water and a milk surrogate. J Agric Food Chem (2004) 52:2016–20. doi: 10.1021/jf0345696
114. Marta S, Pereiro P, Figueras A, Novoa B. An integrative toxicogenomic analysis of plastic additives. J hazard mater (2021) 409:124975. doi: 10.1016/j.jhazmat.2020.124975
115. Ong H-T, Samsudin H, Soto-Valdez H. Migration of endocrine-disrupting chemicals into food from plastic packaging materials: An overview of chemical risk assessment, techniques to monitor migration, and international regulations. Crit Rev Food Sci Nutr (2022) 62(4):957–79. doi: 10.1080/10408398.2020.1830747
116. Hunt PA, Koehler KE, Susiarjo M, Hodges CA, Ilagan A, Voigt RC, et al. Bisphenol a exposure causes meiotic aneuploidy in the female mouse. Curr Biol 13(7) (2003) 13:546–53. doi: 10.1016/S0960-9822(03)00189-1
117. Yong C, Ying Q, Valiyaveettil S, Luen Tang B. Toxicity of microplastics and nanoplastics in mammalian systems. Int J Environ Res Public Health (2020) 17(5):1509. doi: 10.3390/ijerph17051509
118. Ma Y, Zhang J, Zeng R, Qiao X, Cheng R, Nie Y, et al. Effects of the dibutyl phthalate (Dbp) on the expression and activity of aromatase in human granulosa cell line kgn. Ann Clin Lab Sci (2019) 49(2):175–82.
119. Yu G. Effect of mono-(2-ethylhexyl) phthalate (MEHP) on proliferation of and steroid hormone synthesis in rat ovarian granulosa cells in vitro J Cell Physiol (2018) 4:3629–37. doi: 10.1002/jcp.26224
120. Basini G, Bussolati S, Andriani L, Grolli S, Ramoni R, Bertini S, et al. Nanoplastics impair in vitro swine granulosa cell functions. Domest Anim Endocrinol (2021) 76:1066115. doi: 10.1016/j.domaniend.2021.106611
121. Chakraborty SB. Non-essential heavy metals as endocrine disruptors: Evaluating impact on reproduction in teleosts. Pap presented at Proc Zool Soc (2021) 74:417–431. doi: 10.1007/s12595-021-00399-x
122. Pitt JA, Kozal JS, Jayasundara N, Massarsky A, Trevisan R, Geitner N, et al. Uptake, tissue distribution, and toxicity of polystyrene nanoparticles in developing zebrafish (Danio rerio). Aquat Toxicol (2018) 194:185–94. doi: 10.1016/j.aquatox.2017.11.017
123. Pitt JA, Trevisan R, Massarsky A, Kozal JS, Levin ED, Di Giulio RT. Maternal transfer of nanoplastics to offspring in zebrafish (Danio rerio): A case study with nanopolystyrene. Sci Total Environ (2018) 643:324–34. doi: 10.1016/j.scitotenv.2018.06.186
124. Li Y, Yang G, Wang J, Lu L, Li X, Zheng Y, et al. Microplastics increase the accumulation of phenanthrene in the ovaries of marine medaka (Oryzias melastigma) and its transgenerational toxicity. J hazard mater (2022) 424:127754. doi: 10.1016/j.jhazmat.2021.127754
125. Hiller-Sturmhöfel S, Bartke A. The endocrine system: An overview. Alcohol Health Res World (1998) 22(3):153.
126. Graceli, Jones B, Dettogni RS, Merlo E, Niño O, da Costa CS, Zanol JF, et al. The impact of endocrine-disrupting chemical exposure in the mammalian hypothalamic-pituitary axis. Mol Cell Endocrinol (2020) 518:110997. doi: 10.1016/j.mce.2020.110997
127. Haibo J, Yan M, Pan C, Liu Z, Sha X, Jiang C, et al. Chronic exposure to polystyrene microplastics induced Male reproductive toxicity and decreased testosterone levels Via the lh-mediated Lhr/Camp/Pka/Star pathway. Part fibre Toxicol (2022) 19(1):1–17. doi: 10.1186/s12989-022-00453-2
128. Ma Q, Deng P, Lin M, Yang L, Li L, Guo L, et al. Long-term bisphenol a exposure exacerbates diet-induced prediabetes Via Tlr4-dependent hypothalamic inflammation. J hazard mater (2021) 402:123926. doi: 10.1016/j.jhazmat.2020.123926
129. Liu T, Wang Y, Yang M, Shao P, Duan L, Li M, et al. Di-(2-Ethylhexyl) phthalate induces precocious puberty in adolescent female rats. Iranian J Basic Med Sci (2018) 21(8):848. doi: 10.22038/IJBMS.2018.28489.6905
130. Muthuvel R, Venkataraman P, Krishnamoorthy G, Gunadharini DN, Kanagaraj P, Jone Stanley A, et al. Antioxidant effect of ascorbic acid on pcb (Aroclor 1254) induced oxidative stress in hypothalamus of albino rats. Clin Chimica Acta (2006) 365(1-2):297–303. doi: 10.1016/j.cca.2005.09.006
131. Sena GC, Freitas-Lima LC, Merlo E, Podratz PL, de Araújo JFP, Brandão PAA, et al. Environmental obesogen tributyltin chloride leads to abnormal hypothalamic-Pituitary-Gonadal axis function by disruption in Kisspeptin/Leptin signaling in female rats. Toxicol Appl Pharmacol (2017) 319:22–38. doi: 10.1016/j.taap.2017.01.021
132. Oliveira FRT, Ferreira JR, Corrêa dos Santos CM, Miranda Macêdo LE, de Oliveira RB, Rodrigues JA, et al. Estradiol reduces cumulative mercury and associated disturbances in the hypothalamus–pituitary axis of ovariectomized rats. Ecotoxicol Environ Saf (2006) 63(3):488–93. doi: 10.1016/j.ecoenv.2004.12.024
133. Ferrer B, Peres TV, Dos Santos AA, Bornhorst J, Morcillo P, Ludvig Gonçalves C, et al. Methylmercury affects the expression of hypothalamic neuropeptides that control body weight in C57bl/6j mice. Toxicol Sci (2018) 163(2):557–68. doi: 10.1093/toxsci/kfy052
134. Karaulov AV, Smolyagin AI, Mikhailova IV, Stadnikov AA, Ermolina EV, Filippova YV, Kuzmicheva NA, Vlata Z, Djordjevic AB, Tsitsimpikou C, et al. Assessment of the combined effects of chromium and benzene on the rat neuroendocrine and immune systems. Environ Res (2022) 207:112096. doi: 10.1016/j.envres.2021.112096
135. Nudler SI, Quinteros FA, Miler EA, Cabilla JP, Ronchetti SA, Duvilanski BH. Chromium vi administration induces oxidative stress in hypothalamus and anterior pituitary gland from Male rats. Toxicol Lett (2009) 185(3):187–92. doi: 10.1016/j.toxlet.2009.01.003
136. Prüst M, Meijer J, Westerink RHS. The plastic brain: Neurotoxicity of micro-and nanoplastics. Part fibre Toxicol (2020) 17(1):1–16. doi: 10.1186/s12989-020-00358-y
137. Sarasamma S, Audira G, Siregar P, Malhotra N, Lai Y-H, Liang S-T, et al. Nanoplastics cause neurobehavioral impairments, reproductive and oxidative damages, and biomarker responses in zebrafish: Throwing up alarms of wide spread health risk of exposure. Int J Mol Sci (2020) 21(4):1410. doi: 10.3390/ijms21041410
138. Zhao X, Liu Z, Ren X, Duan X. Parental transfer of nanopolystyrene-enhanced tris (1, 3-Dichloro-2-Propyl) phosphate induces transgenerational thyroid disruption in zebrafish. Aquat Toxicol (2021) 236:105871. doi: 10.1016/j.aquatox.2021.105871
139. Rosenfeld CS. Neuroendocrine disruption in animal models due to exposure to bisphenol a analogues. Front Neuroendocrinol (2017) 47:123–335. doi: 10.1016/j.yfrne.2017.08.001
140. Wang J, Li X, Li P, Li L, Zhao L, Ru S, et al. Porous microplastics enhance polychlorinated biphenyls-induced thyroid disruption in juvenile Japanese flounder (Paralichthys olivaceus). Mar pollut Bull (2022) 174:113289. doi: 10.1016/j.marpolbul.2021.113289
141. Xi W, Lee CKF, Yeung WSB, Giesy JP, Wong MH, Zhang X, et al. Effect of perinatal and postnatal bisphenol a exposure to the regulatory circuits at the hypothalamus–Pituitary–Gonadal axis of cd-1 mice. Reprod Toxicol (2011) 31(4):409–17. doi: 10.1016/j.reprotox.2010.12.002
142. Hlisníková H, Petrovičová I, Kolena B, Šidlovská M, Sirotkin A. Effects and mechanisms of phthalates’ action on reproductive processes and reproductive health: A literature review. Int J Environ Res Public Health (2020) 17(18):6811. doi: 10.3390/ijerph17186811
143. Li X-N, Li H-X, Yang T-N, Li X-W, Huang Y-Q, Zhu S-Y, et al. Di-(2-Ethylhexyl) phthalate induced developmental abnormalities of the ovary in quail (Coturnix japonica) Via disruption of the hypothalamic-Pituitary-Ovarian axis. Sci Total Environ (2020) 741:140293. doi: 10.1016/j.scitotenv.2020.140293
144. Barbosa KL, Dettogni RS, da Costa CS, Gastal EL, Raetzman LT, Flaws JA, et al. Tributyltin and the female hypothalamic-Pituitary-Gonadal disruption. Toxicol Sci (2021) 186:179–89. doi: 10.1093/toxsci/kfab141
145. Kumari S, Jamwal R, Mishra N, Singh DK. Recent developments in environmental mercury bioremediation and its toxicity: A review. Environ Nanotechnol Monit Manage (2020) 13:100283. doi: 10.1016/j.enmm.2020.100283
146. Rice KM, Walker EM Jr, Wu M, Gillette C, Blough ER. Environmental mercury and its toxic effects. J Prev Med Public Health (2014) 47(2):74. doi: 10.3961/jpmph.2014.47.2.74
147. Li Z, Li X, Qian Y, Guo C, Wang Z, Wei Y. The sustaining effects of e-Waste-Related metal exposure on hypothalamus-Pituitary-Adrenal axis reactivity and oxidative stress. Sci Total Environ (2020) 739:139964. doi: 10.1016/j.scitotenv.2020.139964
148. Kim J-H, Kang J-C. Effects of Sub-chronic exposure to lead (Pb) and ascorbic acid in juvenile rockfish: Antioxidant responses, Mt gene expression, and neurotransmitters. Chemosphere (2017) 171:520–27. doi: 10.1016/j.chemosphere.2016.12.094
149. Qu J, Niu H, Wang J, Wang Q, Li Y. Potential mechanism of lead poisoning to the growth and development of ovarian follicle. Toxicology (2021) 457:152810. doi: 10.1016/j.tox.2021.152810
150. Wise JJP, Young JL, Cai J, Cai L. Current understanding of hexavalent chromium [Cr (Vi)] neurotoxicity and new perspectives. Environ Int (2022) 158:106877. doi: 10.1016/j.envint.2021.106877
153. Fernandez MO, Bourguignon NS, Arocena P, Rosa M, Libertun C, Lux-Lantos V. Neonatal exposure to bisphenol a alters the hypothalamic-Pituitary-Thyroid axis in female rats. Toxicol Lett (2018) 285:81–6. doi: 10.1016/j.toxlet.2017.12.029
154. Dunnick JK, Pandiri AR, Merrick BA, Kissling GE, Cunny H, Mutlu E, et al. Carcinogenic activity of pentabrominated diphenyl ether mixture (De-71) in rats and mice. Toxicol Rep (2018) 5:615–245. doi: 10.1016/j.toxrep.2018.05.010
155. Butt CM, Miranda ML, Stapleton HM. Development of an analytical method to quantify pbdes, oh-bdes, hbcds, 2, 4, 6-tbp, eh-tbb, and beh-tebp in human serum. Anal bioanal Chem (2016) 408(10):2449–59. doi: 10.1007/s00216-016-9340-3
156. Wade MG, Kawata A, Rigden M, Caldwell D, Holloway AC. Toxicity of flame retardant isopropylated triphenyl phosphate: Liver, adrenal, and metabolic effects. Int J Toxicol (2019) 38(4):279–90. doi: 10.1177/1091581819851502
157. Sarkar R, Mohanakumar KP, Chowdhury M. Effects of an organophosphate pesticide, quinalphos, on the hypothalamo-Pituitary-Gonadal axis in adult Male rats. J Reprod Fertil (2000) 118(1):29–38. doi: 10.1530/reprod/118.1.29
158. Bjørklund G, Chirumbolo S, Dadar M, Pivina L, Lindh U, Butnariu M, et al. Mercury exposure and its effects on fertility and pregnancy outcome. Basic Clin Pharmacol Toxicol (2019) 125(4):317–27. doi: 10.1111/bcpt.13264
159. McGregor AJ, Mason HJ. Occupational mercury vapour exposure and testicular, pituitary and thyroid endocrine function. Hum Exp Toxicol (1991) 10(3):199–203. doi: 10.1177/096032719101000309
160. Petrusz P, Weaver CM, Grant LD, Mushak P, Krigman MR. Lead poisoning and reproduction: Effects on pituitary and serum gonadotropins in neonatal rats. Environ Res (1979) 19(2):383–91. doi: 10.1016/0013-9351(79)90063-X
161. Lafuente A, Márquez N, Perez-Lorenzo M, Pazo D, Esquifino AI. Cadmium effects on hypothalamic-Pituitary-Testicular axis in Male rats. Exp Biol Med (2001) 226(6):605–11. doi: 10.1177/153537020122600615
162. Fernández M, Bianchi M, Lux-Lantos V, Libertun C. Neonatal exposure to bisphenol a alters reproductive parameters and gonadotropin releasing hormone signaling in female rats. Environ Health Perspect (2009) 117(5):757–62. doi: 10.1289/ehp.0800267
163. Yaglova NV, Tsomartova DA, Obernikhin SS, Nazimova SV. Alteration of rat adrenal cortex after low-dose exposure to endocrine disrupting chemical dichlorodiphenyltrichloroethane since the first day of postnatal development. Voprosy Pitaniia (2017) 86(4):70–6. doi: 10.24411/0042-8833-2017-00061
164. Merlo E, Podratz PL, Sena GC, De Araújo JFP, Lima LCF, Alves ISS, et al. The environmental pollutant tributyltin chloride disrupts the hypothalamic-Pituitary-Adrenal axis at different levels in female rats. Endocrinology (2016) 157(8):2978–95. doi: 10.1210/en.2015-1896
165. Fujimoto N. Effects of endocrine disruptors on the pituitary gland. J Toxicol Pathol (2001) 14(1):65–5. doi: 10.1293/tox.14.65
166. Sabir S, Hamid Akash MS, Fiayyaz F, Saleem U, Mehmood MH, Rehman K. Role of cadmium and arsenic as endocrine disruptors in the metabolism of carbohydrates: Inserting the association into perspectives. Biomed Pharmacother (2019) 114:108802. doi: 10.1016/j.biopha.2019.108802
167. Ronchetti SA, Novack GV, Bianchi MS, Crocco MC, Duvilanski BH, Cabilla JP. In vivo xenoestrogenic actions of cadmium and arsenic in anterior pituitary and uterus. Reproduction (2016) 152(1):1–10. doi: 10.1530/REP-16-0115
168. Ronchetti SA, Bianchi MS, Duvilanski BH, Cabilla JP. In vivo and in vitro arsenic exposition induces oxidative stress in anterior pituitary gland. Int J Toxicol (2016) 35(4):463–75. doi: 10.1177/1091581816645797
169. Rosol TJ, Yarrington JT, Latendresse J, Capen CC. Adrenal gland: Structure, function, and mechanisms of toxicity. Toxicol Pathol (2001) 29(1):41–8. doi: 10.1080/019262301301418847
170. Stojanović Z, Todorović A, Filipović N, Veljković F, Guševac Stojanović I. A single dose of microplastic particles induces changed in organ weight of Male wistar rats. Int Conf Fundam Appl Aspects Phys Chemistry-Phys Chem (2021) 2021:119–22
171. Brun NR, Van Hage P, Hunting ER, Haramis A-PG, Vink SC, Vijver MG, et al. Polystyrene nanoplastics disrupt glucose metabolism and cortisol levels with a possible link to behavioural changes in larval zebrafish. Commun Biol (2019) 2(1):1–9. doi: 10.1038/s42003-019-0629-6
172. Campioli E, Martinez-Arguelles DB, Papadopoulos V. In utero exposure to the endocrine disruptor di-(2-Ethylhexyl) phthalate promotes local adipose and systemic inflammation in adult Male offspring. Nutr Diabetes (2014) 4(5):e115–e15. doi: 10.1038/nutd.2014.13
173. Medwid S, Guan H, Yang K. Prenatal exposure to bisphenol a disrupts adrenal steroidogenesis in adult mouse offspring. Environ Toxicol Pharmacol (2016) 43:203–08. doi: 10.1016/j.etap.2016.03.014
174. Eker F, Gungunes A, Durmaz S, Kisa U, Celik ZR. Nonfunctional adrenal incidentalomas may be related to bisphenol-a. Endocrine (2021) 71(2):459–66. doi: 10.1007/s12020-020-02502-2
175. Hlisníková H, Petrovičová I, Kolena B, Šidlovská M, Sirotkin A. Effects and mechanisms of phthalates’ action on neurological processes and neural health: A literature review. Pharmacol Rep (2021) 73(2):386–404. doi: 10.1007/s43440-021-00215-5
176. Chen X, Mo J, Zhang S, Li X, Huang T, Zhu Q, et al. 4-bromodiphenyl ether causes adrenal gland dysfunction in rats during puberty. Chem Res Toxicol (2019) 32(9):1772–79. doi: 10.1021/acs.chemrestox.9b00123
177. Yan ,H, Hao G, Cheng D, Kou R, Zhang C, Si J. Tributyltin reduces the levels of serum adiponectin and activity of akt and induces metabolic syndrome in Male mice. Environ Toxicol (2018) 33(7):752–58. doi: 10.1002/tox.22562
178. Solaiman A, Seddik S. Histological study of the effect of tributyltin on the adrenal cortical cells of adult Male albino rats. Egypt J Histol (2020) 43(1):104–21. doi: 10.21608/EJH.2019.13126.1127
179. Di Lorenzo M, Sciarrillo R, Rosati L, Sellitti A, Barra T, De Luca A, et al. Effects of alkylphenols mixture on the adrenal gland of the lizard Podarcis sicula. Chemosphere (2020) 258:127239. doi: 10.1016/j.chemosphere.2020.127239
180. Yaglova NV, Tsomartova DA, Yaglov VV. Effect of prenatal and postnatal exposure to low doses of ddt on catecholamine secretion in rats in different period of ontogeny. Bull Exp Biol Med (2017) 163(4):422–24. doi: 10.1007/s10517-017-3819-6
181. Yaglova NV, Obernikhin SS, Yaglov VV, Tsomartova DA, Nazimova SV, Timokhina EP. Ultrastructural mechanisms of impaired aldosterone synthesis in rats exposed to ddt during prenatal and postnatal development. Bull Exp Biol Med (2020) 170(1):101–05. doi: 10.1007/s10517-020-05013-2
182. Pamphlett R, Kum Jew S, Doble PA, Bishop DP. Mercury in the human adrenal medulla could contribute to increased plasma noradrenaline in aging. Sci Rep (2021) 11(1):1–14. doi: 10.1038/s41598-021-82483-y
183. Soto M, Lewis R, Curtis JT. Chronic exposure to inorganic mercury alters stress responses in Male prairie voles (Microtus ochrogaster). Horm Behav (2019) 109:53–5. doi: 10.1016/j.yhbeh.2019.02.008
184. Chandra AK, Chatterjee A, Ghosh R, Sarkar M, Chaube SK. Chromium induced testicular impairment in relation to adrenocortical activities in adult albino rats. Reprod Toxicol (2007) 24(3-4):388–96. doi: 10.1016/j.reprotox.2007.07.009
185. Mikheeva EV, Zhigal’skiĭ OA, Mamina VP, Baĭtimirova EA. Adaptation of the bank vole (Clethrionomys glareolus schreber) to conditions of biogeochemical province with abnormally high content of nickel, cobalt and chromium. Zhurnal Obshchei Biol (2006) 67(3):212–21. doi: 10.1016/j.ecoenv.2021.112034
186. Lauretta R, Sansone A, Sansone M, Romanelli F, Appetecchia M. Endocrine disrupting chemicals: Effects on endocrine glands. Front Endocrinol (2019) 10:178. doi: 10.3389/fendo.2019.00178
187. Kinlein SA, Wilson CD, Karatsoreos IN. Dysregulated hypothalamic–Pituitary–Adrenal axis function contributes to altered endocrine and neurobehavioral responses to acute stress. Front Psychiatry (2015) 6:31. doi: 10.3389/fpsyt.2015.00031
188. Medwid S, Guan H, Yang K. Bisphenol a stimulates adrenal cortical cell proliferation via erβ-mediated activation of the sonic hedgehog signalling pathway. J Steroid Biochem Mol Biol (2018) 178:254–62. doi: 10.1016/j.jsbmb.2018.01.004
189. Olukole SG, Olaniyi Lanipekun D, Ola-Davies EO, Bankole OO. Melatonin attenuates bisphenol a-induced toxicity of the adrenal gland of wistar rats. Environ Sci pollut Res (2019) 26(6):5971–82. doi: 10.1007/s11356-018-4024-5
190. Asimakopoulos AG, Thomaidis NS, Koupparis MA. Recent trends in biomonitoring of bisphenol a, 4-T-Octylphenol, and 4-nonylphenol. Toxicol Lett (2012) 210(2):141–54. doi: 10.1016/j.toxlet.2011.07.032
191. Timokhina EP, Yaglov VV, Nazimova SV. Dichlorodiphenyltrichloroethane and the adrenal gland: From toxicity to endocrine disruption. Toxics (2021) 9(10):243. doi: 10.3390/toxics9100243
Keywords: microplastics, nanoplastics, mammalian endocrine system, endocrine abnormalities, endocrine disrupting chemicals, plastic additives, environmental pollution
Citation: Ullah S, Ahmad S, Guo X, Ullah S, Ullah S, Nabi G and Wanghe K (2023) A review of the endocrine disrupting effects of micro and nano plastic and their associated chemicals in mammals. Front. Endocrinol. 13:1084236. doi: 10.3389/fendo.2022.1084236
Received: 30 October 2022; Accepted: 05 December 2022;
Published: 16 January 2023.
Edited by:
Bodil Holst, Swedish University of Agricultural Sciences, SwedenReviewed by:
Luigi Rosati, University of Naples Federico II, ItalySergio Minucci, Università della Campania Luigi Vanvitelli, Italy
Copyright © 2023 Ullah, Ahmad, Guo, Ullah, Ullah, Nabi and Wanghe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ghulam Nabi, Z2h1bGFtbmFiaXFhdUBnbWFpbC5jb20=; Kunyuan Wanghe, d2FuZ2hla3VueXVhbkBnbWFpbC5jb20=
†These authors have contributed equally to this work and share first authorship