 Corentin Jouault1,2,3*†
Corentin Jouault1,2,3*† Michael S. Engel4,5†
Michael S. Engel4,5† Diying Huang6†Juliette Berger1Philippe Grandcolas1†
Diying Huang6†Juliette Berger1Philippe Grandcolas1† Evgeny E. Perkovsky7,8†Frédéric Legendre1†‡
Evgeny E. Perkovsky7,8†Frédéric Legendre1†‡ André Nel1†‡
André Nel1†‡- 1Institut de Systématique, Évolution, Biodiversité (ISYEB) Muséum National d’Histoire Naturelle, CNRS, Sorbonne Université, EPHE, Université des Antilles, Paris, France
- 2Univ. Rennes, CNRS, Géosciences Rennes, UMR 6118, Rennes, France
- 3CNRS, UMR 5554 Institut des Sciences de l’Évolution de Montpellier, Montpellier, France
- 4Division of Invertebrate Zoology, American Museum of Natural History, New York, NY, United States
- 5Division of Entomology, Department of Ecology and Evolutionary Biology, Natural History Museum, University of Kansas, Lawrence, KS, United States
- 6State Key Laboratory of Palaeobiology and Stratigraphy, Center for Excellence in Life and Paleoenvironment, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China
- 7Schmalhausen Institute of Zoology, National Academy of Sciences of Ukraine, Kiev, Ukraine
- 8Borissiak Paleontological Institute of the Russian Academy of Sciences, Moscow, Russia
In several insect eusocial lineages, e.g., some aphids, thrips, ants, some stingless bees, and termites, task specialization is brought to its climax with a sterile soldier caste solely devoted to colony defense. In Isoptera, while the reproductives are defenseless, the soldiers have unique morpho-physiological specializations whose origin and evolution remain unresolved. Here we report on two instances of Cretaceous fossil termite reproductives belonging to different families († Valkyritermes inopinatus gen. et sp. nov. and an unpublished specimen from the Crato Formation), with intriguing phragmotic soldier-like heads and functional wings. These individuals, herein called Valkyries, are the first termite reproductives known with defensive features and suggest that phragmosis arose at least in the Early Cretaceous. Valkyries resemble modern neotenic soldiers except for their complete wings. Their discovery supports the hypothesis that the division between reproductive (indicated by the winged condition of Valkyries) and defensive tasks (indicated by the phragmotic head) has not always been complete in termite history. We explore two alternative scenarios regarding the origin of Valkyries (i.e., relatively recent and convergent origins vs. plesiomorphic condition) and discuss how they might relate to the development of soldiers. We argue that, in both cases, Valkyries likely evolved to face external threats, a selective pressure that could also have favored the origin of soldiers from helpers. Valkyries highlight the developmental flexibility of termites and illustrate the tortuous paths that evolution may follow.
Introduction
Task specialization is a defining aspect of eusocial societies, appearing independently in lineages as diverse as naked mole rats, snapping shrimp, and insects, the latter of whom made famous such societal organizations (Wilson, 1971, 2000). In these lineages, there is a caste system including reproductives that are the parents of all the other members of the colony (in the most typical cases), and workers devoted to the care of the young, the search for food, and to the protection of the colony (Wilson, 1971, 2000). If these divisions are essentially fixed in the majority of the non-arthropods lineages, some insect clades show a further specialization, related to the defense of the colony (Wilson, 1971), which is paramount given that reproductives are generally poorly equipped for defense, relying on colony members to provide for their protection. Some eusocial insects are thus renowned for having developed a third caste, the soldier caste, purely devoted to colony defense (Wilson, 1971; Crespi, 1992; Yamamura, 1993; Stern and Foster, 1996; Queller and Strassmann, 1998; Šobotník and Dahlsjö, 2017).
In all extant termites, alates or reproductives are highly vulnerable, particularly during the foundation of new colonies and until the development of the first generation of offspring (a period of several weeks or months). In fact, they lack any specialization that would ensure their defense or that of the first generation of workers. This absence is particularly surprising because another line of eusocial insects (ants) comprises species whose reproductives and soldiers share defensive morphological adaptations (e.g., Blepharidatta: Brandão et al., 2001). To ensure the defense of the colony, termite soldiers possess numerous specializations described below. Soldiers are present in nearly all termite genera (secondary lost in a few Termitidae) so that the soldier caste is reconstructed to be synapomorphic for all extant termites (e.g., Hare, 1937; Noirot and Pasteels, 1987; Roisin, 2000). Finding fossil reproducers with soldier-like heads bring new data that can enlighten what we know about the evolution of defensive strategies in termites.
Termite soldiers are surely the most striking example of hyperspecialization since they possess an arsenal of impressive adaptations for defense, including phragmotic heads, massive mandibles, chemical projections, etc. (Šobotník et al., 2010a,b, 2012; Šobotník and Dahlsjö, 2017). At its simplest, soldiers are sterile and develop from helpers (via a presoldier stage), the helpers being the developmental origin of numerous castes or intercastes (Noirot, 1956; Myles, 1980, 1986; Myles and Chang, 1984; Roisin and Pasteels, 1985; Thorne et al., 2003). Interestingly, an alternative to the classic sterile soldiers, called “brachypterous neotenics” or fertile “soldier neotenics,” is not uncommonly found in termite colonies of early diverging families (e.g., Archotermopsidae) (Thorne, 1997; Myles, 1999). Such individuals have been thought to be involved in the origin of the sterile defensive caste, with fertile soldier neotenics originating in the context of agonistic interactions among intercolonial reproductives (Thorne et al., 2003). Reproductive soldiers are often observed early in the life cycle of a colony, perhaps reflecting the greater vulnerability to intercolonial reproductive warfare during this stage (Thorne et al., 2003). Although studies investigating the origin of the termite soldier caste are multifarious, none has yet identified morphological specializations of alates for defense (other than the fontanelle present in the analogous soldiers). Here we report on two instances of Cretaceous termites with phragmotic soldier-like heads and functional wings, belonging to different families. Fully winged, these fossils are undoubtedly imagoes but they share morphological features with some neotenic soldiers, which is unique for termite alates. We argue that these findings shed light on the origin and evolution of defense in termites, and support the hypothesis that the soldier caste most likely originated from helpers, and not neotenics, following inter-colonial aggressions and/or predation pressures.
Materials and Methods
Locality and Repository
Valkyritermes inopinatus gen. et sp. nov.: Noije Bum Hill, Hukawng Valley (26°29′N, 96°35′E), Kachin State, Myanmar (see Grimaldi and Ross, 2017 for detailed map); upper Albian to lower Cenomanian, mid-Cretaceous. Repository: Geological Department and Museum (IGR) of the University of Rennes (France). The specimen is embedded in a piece of yellow Kachin amber. Radiometric data and taphonomic analysis of pholadids established an Early Cenomanian age (98.79 ± 0.62 Ma), based on zircons from volcanic clasts found within the amber-bearing sediments (Shi et al., 2012; Smith and Ross, 2017). Some ammonites found in the amber-bearing bed and within amber corroborate a late Albian/early Cenomanian age (Cruickshank and Ko, 2003; Yu et al., 2019). The specimen was legally obtained and donated to the Geological Department and Museum (IGR) of the University of Rennes (France).
Preparation, Examination, and Study of Specimen
The amber piece containing the type specimen of †Valkyritermes inopinatus gen. et sp. nov. was polished with a polisher (Buehler EcoMet 30) by using a thin silicon carbide sanding paper (grit size = 7,000). Then, the fragment was embedded with cedar oil between microscopic slides to minimize light scattering during focus stacking. The specimen of †Valkyritermes inopinatus gen. et sp. nov. was examined with a Leica MZ APO stereomicroscope, and a Nikon SMZ25 stereomicroscope. Photographs were taken using a Canon EOS 5D mark II mounted on a Leica MZ APO stereomicroscope. All images are digitally stacked photomicrographic composites of several individual focal planes, obtained using HeliconFocus 6.7 software. Figures were composed with Adobe Illustrator CC 2019 and Photoshop CC 2019 software.
The systematic classification and the morphological terminology follow Krishna et al. (2013) except for wing venation that follows Schubnel et al. (2019), and we retain the ranked name Isoptera (Lo et al., 2007), although as subordinate to Blattodea.
Nomenclatural acts established herein are registered in ZooBank, the proposed online registration system for the International Code of Zoological Nomenclature (ICZN). The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through and standard web browser by appending the LSID to the prefix “http://zoobank.org/”. The LSID for this publication is urn:lsid:zoobank.org:pub:2831A9DD-63AD-43DE-AC7F-3446035520FD.
Results
Systematic Paleontology
Blattodea Brunner von Wattenwyl, 1882.
Isoptera Brullé, 1832.
Family Kalotermitidae Froggatt, 1896.
Valkyritermes Jouault, Engel, & Nel gen. nov.
urn:lsid:zoobank.org:act:DEF01FFE-0CCC-47B3-B550-B0218DC33E17.
Type Species
Valkyritermes inopinatus Jouault, Engel, & Nel sp. nov.
Etymology
The generic name is a combination of valkyrie from Old Norse valkyrja (meaning, “chooser of the slain”), and the Latin noun termes (meaning, “termite”), commonly used for names among Isoptera; gender masculine.
Diagnosis
Alates (Figure 1) with “phragmotic” head, frons concave, frontal flange slightly elevated, with wide median notch (Figures 2A,B); in dorsal view, flange follows a semi-circular arch (Figures 2A,B); frontal horn slightly sclerotized; head with anterior corners orthogonal, posterior corners rounded; antenna moniliform with 15 articles (Figure 2A); postclypeus triangular shaped (narrower anteriorly) (Figure 2A); mandibles form and dentition imago-like, covered by labrum; left mandible apical tooth elongate, sharp; two marginal teeth, widely separated (with broad v-shaped incision in LM1–LM2 interdental space, similar to left imago mandible of Kalotermes flavicollis) (Figures 2B,C); right mandible with long, sharp apical tooth, two marginal teeth, RM1–RM2 interdental space narrower than interdental space of left mandible (Figures 2B,C); pronotum slightly trapezoidal, slightly wider than head, anterior margin concave, posterior margin medially concave, with dorsal surface flatly arched in profile, sides convex (Figure 1); tarsi tetramerous; tibial spur formula 3-3-3(?); with one inner additional spine on mesotibia (mb); arolium present; forewing with large scale, larger than hind wing scale, with humeral margin well-defined, arched, claval suture nearly straight, suture convex (Figures 1B, 2D); Sc, RA, and RP heavily sclerotized, Sc short terminating within scale, RA simple, RP running parallel to costal margin, with seven branches joining wing margin, radial field narrow along entire length, terminating anterior to wing apex (Figures 1B, 2D); M weak, running midway between RP and CuA, with one bifurcation just beyond wing midlength, at tangent of RP3, terminating in three or four branches (depending on wing), not contacting RP apically, anteriormost termination near wing apex; CuA weak and unsclerotized, with at least 10 branches joining posterior margin, apicalmost termination near apical fifth of wing length (Figures 1B, 2D).

Figure 1. Termite valkyrie from early Cenomanian amber. Valkyritermes inopinatus gen. et sp. nov., Kachin amber (ca. 98 Ma, mid-Cretaceous).
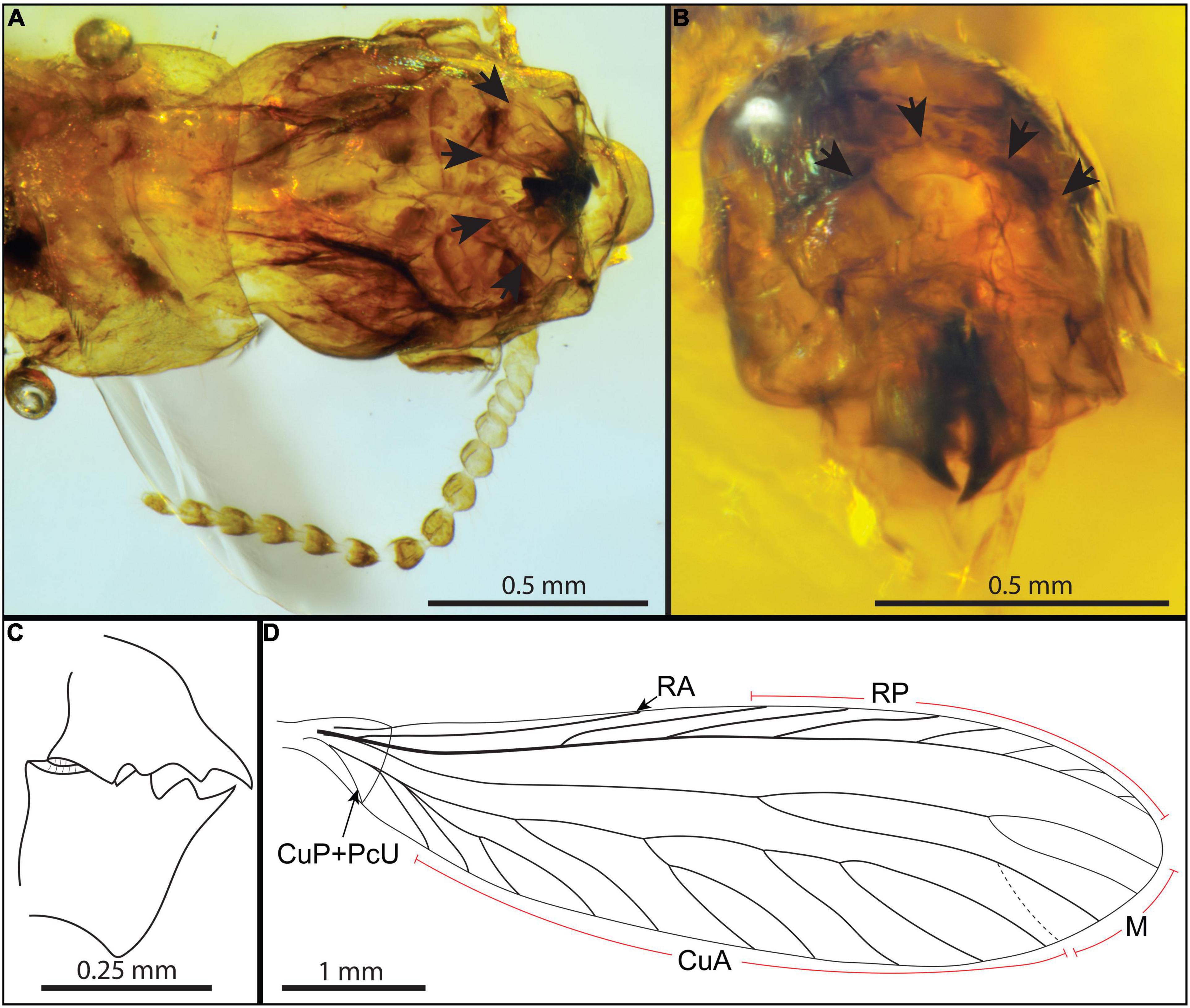
Figure 2. Detailed views and interpretative drawings of morphological features of the valkyrie Valkyritermes inopinatus gen. et sp. nov. (A) Microphotography of head in dorsal view, arrows: flange. (B) Microphotography of head in face view, arrows: flange. (C) Line drawing of mandibles. (D) Interpretative line drawing of forewing.
Valkyritermes inopinatus Jouault, Engel, & Nel sp. nov.
urn:lsid:zoobank.org:act:48FD1A7D-7456-4678-B704-2A5C58FFE7C5.
Etymology
The specific epithet is from the Latin adjective inoptus, meaning “unexpected”, and refers to the rather unexpected nature of the discovery in Kachin amber.
Holotype
IGR.BU-027 (Figures 1, 2), housed in the Geological Department and Museum (IGR) of the University of Rennes (France).
Type Locality and Horizon
Noije Bum Hill, Hukawng Valley, Kachin State, Myanmar; upper Albian to lower Cenomanian, mid-Cretaceous.
Description
Alate specimen (Figure 1), body 4.03 mm long (from tip of labrum to abdomen apex). Head robust, ca. 0.8 mm long and ca. 0.63 mm wide excluding compound eyes (slightly flattened dorso-ventrally because of a fracture), square-shaped in dorsal view, prognathous, sides parallel to slightly convex, anterolateral corner orthogonal, postero-lateral corners rounded (Figures 2A,B); three (?) maxillary palpomeres, combined length as preserved ca. 0.25 mm; at least three labial palpomeres present (not well preserved proximally); compound eyes, ca. 0.2 mm long, circular, situated laterally near head mid-length, and separated from posterior head margin by more than their length (Figure 2A); ocelli not visible; fontanelle absent (Figures 2A,B); antenna moniliform with 15 articles (Figure 2A). Pronotum ca. 0.50 mm long and 0.67 mm wide (Figure 1).
Legs slender (Figure 1); profemur ca. 0.75 mm long, protibia ca. 0.62 mm long, protarsus at least 0.25 mm long; protibia with three spurs f1, f2, f3; mesofemur medially swollen ca. 0.37 mm long, mesotibia ca. 0.56 mm long, mesotarsus ca. 0.30 mm long; mesotibia with three observable spurs m1, m2, m3, with one additional inner spine on mesotibia (mb); metafemur at least 0.70 mm long, metatibia ca. 0.71 mm long, combined length of metatarsi 0.32 mm; metatibia with three observable spurs h1, h2, h3(?) (h3 seems to be extremely short and not fully preserved); all tarsi tetramerous; pretarsus with arolium. Forewing 5.4 mm long, ca. 1.60 mm wide (measured at mid-length); scale ca. 0.7 mm long (Figure 1). Hind wing slightly shorter than forewing, with Sc, RA, and RP similar to forewing.
Abdomen (Figure 1) ca. 2.3 mm long with at least nine observable segments, conspicuously dorso-ventrally flattened and not well defined due to preservation; abdominal segments widest at mid-length. Cerci and styli not preserved. Color. Not preserved.
Remarks
The body of the specimen is generally compressed due to preservation. It is also extensively desiccated with the cuticle distorted, cleared, and collapsed in places rendering the observation and interpretation of certain features challenging. Valkyritermes shares characters with Kalotermitidae (Krishna, 1961; Krishna et al., 2013), a family previously recorded from the same deposit (Engel et al., 2007). In particular, the combination of lacking an anal lobe on the hind wing; tetramerous tarsi; antenna with 15 articles; left mandible with two marginal teeth, marginal teeth fused basally with angled v-shaped separation; right mandible without subsidiary tooth; fontanelle absent; pronotum flatly arched; tibial spur formula 3-3-3(?); forewing scale overlapping hind wing scale; forewing Sc short; Rs sclerotized, running parallel to costal margin, and with number of anterior branches meeting costal margin; M not sclerotized (Krishna, 1961; Krishna et al., 2013). The plesiomorphic retention of M and CuA arising within the forewing scale is similar to that observed in the genera Postelectrotermes, Electrotermes, and Proelectrotermes (likely also true for Kachinitermopsis), as is the combination of one additional spine (mb) on the mesotibia and the presence of pretarsal arolia (Krishna et al., 2013). Additionally, the forewing R terminates before wing midlength, M does not contact RP, CuP terminates at the suture on the posterior margin, and the wing membrane is not nodulose, all features that when taken in combination with the aforementioned traits serve to distinguish the species and genus from all other living and fossil Kalotermitidae.
General Comment on Systematics
In addition to this new Valkyrie, an alate with similar soldier like features is known from the Early Cretaceous Crato Formation. This specimen possesses a stout rectangular head (phragmotic) virtually similar to that of soldier and is fully winged. The morphological features and the wing venation of this additional specimen (pentamerous tarsi, developed wing venation, large pronotum) suggest strong affinities with early diverged termite lineages but its preservation precludes a confident placement in one family. Therefore, we consider that it belongs to the “Meiatermes-grade” and shares affinities with the family Mastotermitidae. This specimen increases the temporal range of the Valkyries from the late Aptian to the early Cenomanian. Because the description of this specimen would neither satisfy the guidelines proposed by Cisneros et al. (2022) nor the requirements of the Brazilian law, we only mention its presence without describing it formally.
Additionally, an alate with putatively soldier-like features was previously reported from Kachin amber (i.e., Tanytermes, although this example may merely be an alate with a more elongate head rather than being truly a Valkyrie, meaning further study is needed) (Engel et al., 2007), but was a less striking case than Valkyritermes. Naturally, we cannot ascertain that these Valkyries do not belong to fossil species already defined on the basis of other castes (e.g., species described from “normal” alate imagoes or soldiers), but features of wing venation and other body structures not typically linked to caste differences tend to indicate that the two specimens mentioned or described herein correspond to new species. For instance, the wing venation of the Kachin kalotermitid described herein differ notably from the coeval kalotermitids previously documented from typical alates or shed wings (Engel et al., 2007; Engel and Delclòs, 2010).
Discussion
With wings and phragmotic heads, Valkyries are unique termites featuring both alates and soldier morphological characters. They likely had reproductive capacities because, in extant termites, all winged individuals are able to reproduce. In addition, their soldier-like features are not as pronounced as those of sterile soldiers (i.e., with elongate or heavily toothed mandibles), which further strengthens the hypothesis that Valkyries were able to reproduce and took care of their offspring. The soldiers’ elongate mandibles do not allow them to attend to the brood, as founders do in their incipient colonies (e.g., Nalepa, 2015; Chouvenc and Su, 2017). Therefore, Valkyries were undoubtedly winged reproductives but we argue that they might nonetheless shed light on soldier evolution and task partitioning in termites.
Elaborating on two alternative scenarios about the origin of Valkyries, we more specifically address the following questions (Figure 3): (1) Although the oldest known Valkyrie, reported herein (Figure 4B), is older than the first occurrences of true apterous soldiers, do Valkyries necessarily predate sterile soldiers? (2) Were “weapons” shared between different castes in early diverging lineages like those exhibiting Valkyries? If yes, what can it tell us about the evolution of task partitioning in termites? (3) Because developmental flexibility is mainly understood as decreasing along termite evolution, do Valkyries reflect an early stage prior to the canalization of more fixed developmental pathways? (4) Because developmental flexibility seems linked to feeding groups (e.g., high developmental flexibility in wood-feeders), are Valkyries more likely in some feeding-groups than in others?
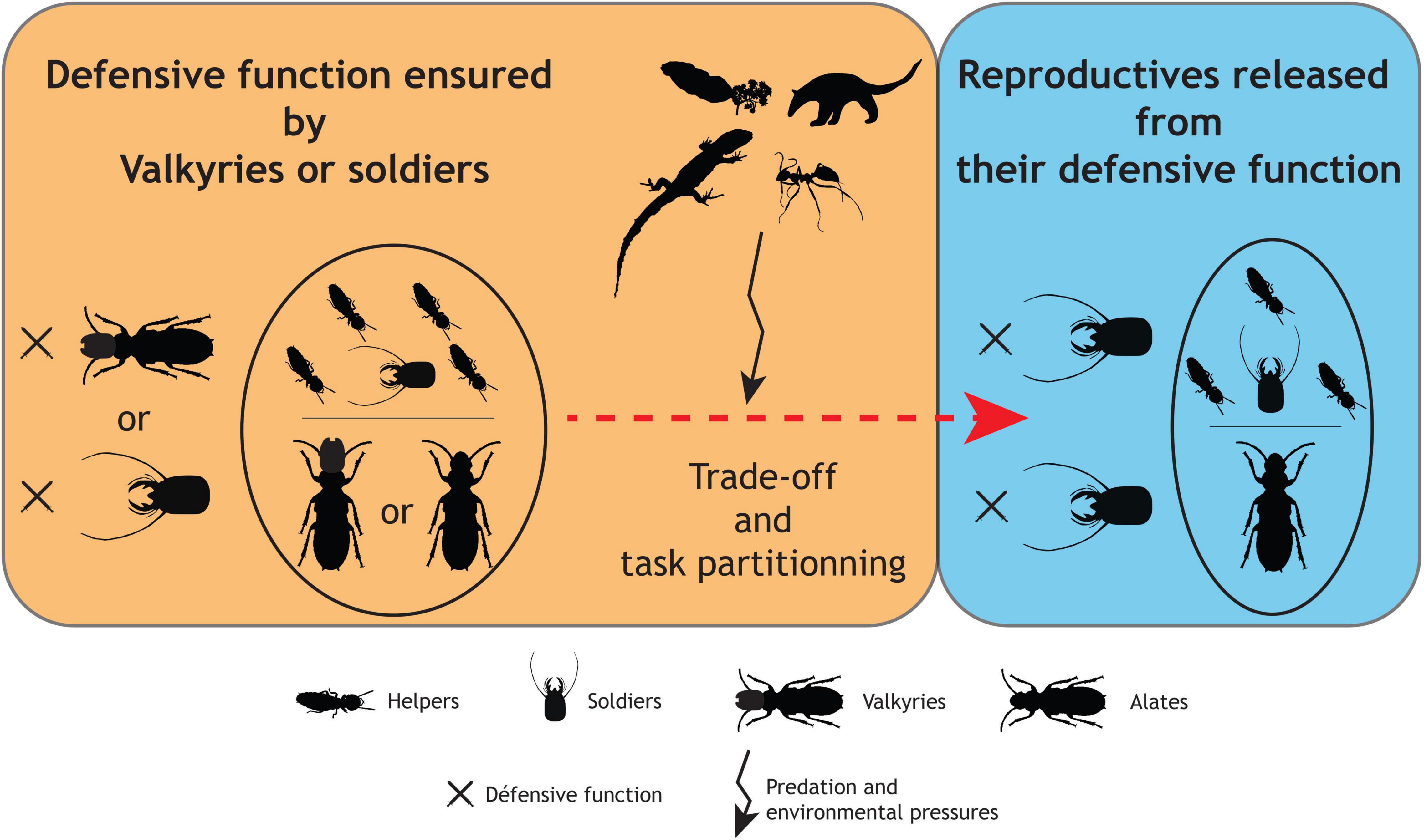
Figure 3. Hypothesis on the evolution of defensive and reproductive functions in termite societies: insight from the Valkyries (silhouettes from or modified from http://phylopic.org/). Reproductive functions are assumed to be initially performed by Valkyries or “classic” alates when soldiers may have been present or not in the colony or not (orange panel). Later, as a result of predation and environmental pressures, and due to their putative high cost, Valkyries would have been counter-selected, resulting in a better task partitioning (blue panel).
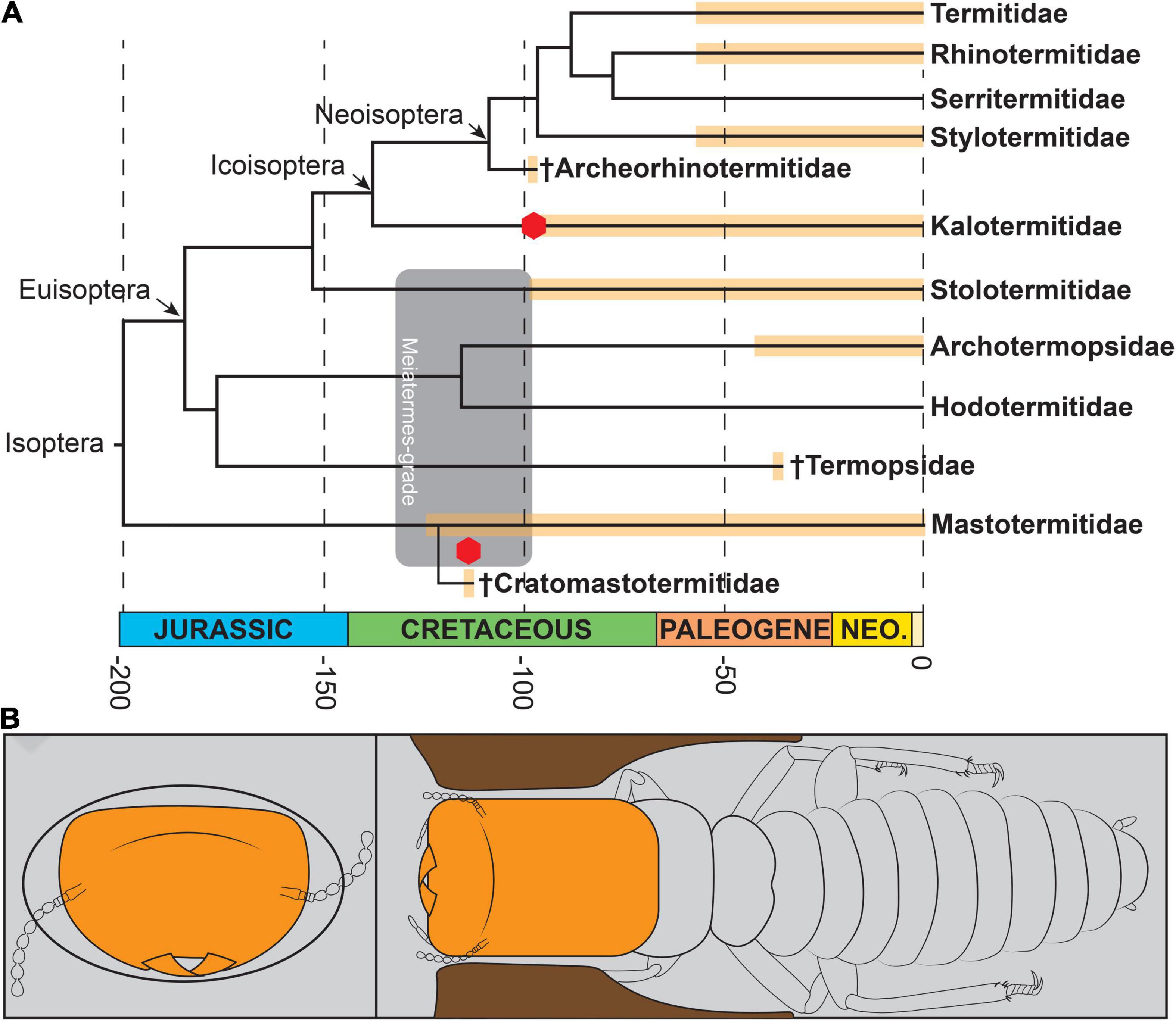
Figure 4. Dated phylogeny of termites, and phragmotic behavior of a Valkyrie. (A) Dated phylogeny of living and fossil taxa of Isoptera with occurrences of Valkyries indicated by red hexagon (datings from Jouault et al., 2021). (B) Schematic reconstruction of phragmotic behavior of a Valkyrie.
Evolution of Defensive Function in Early Termite Societies: Insights From Valkyries
Current Hypotheses About the Origin of the Soldier Caste
While the primary caste division in eusocial societies is becoming better understood, with helpers originating from offspring of the primary reproductives that remain within the nest and aid in raising their siblings and other close relatives, the eventual rise of the soldier caste is more enigmatic. Indeed, soldiers in different lineages of eusocial societies have different evolutionary origins, ranging from polyethism among helpers (e.g., stingless bees) to fixed developmental pathways (Lin and Michener, 1972; Stern and Foster, 1996; Legendre et al., 2008, 2013; Grüter et al., 2012; Korb, 2016; Miura and Maekawa, 2020). In extant termites, soldiers are an irreversible, specialized caste in terms of morphology, behavior, physiology, and development, arising from helpers or late-instar larvae (Šobotník et al., 2010a,b; Šobotník and Dahlsjö, 2017).
There have been two non-mutually exclusive hypotheses regarding the evolutionary origin of a soldier caste in termites (Šobotník et al., 2010a). Under one hypothesis, intrinsic benefits of aiding a flourishing colony drove helpers performing tasks within the colony to assume defensive roles rather than proceeding on to develop as neotenics or alates (Šobotník et al., 2010a). The other hypothesis posits that soldier behaviors and morphologies were selected by intracolonial or intercolonial aggressions among neotenics, themselves derived from helpers (Thorne et al., 2003). These two scenarios are not mutually exclusive as in both cases helpers may have originally assumed defensive tasks either before or after developing into neotenics or reproductives, with defensive specializations in morphology and physiology appearing subsequently.
Convergent and Relatively Recent Valkyries?
Soldiers are present in almost all termite species and are considered to be an apomorphy of termites (e.g., Hare, 1937; Noirot and Pasteels, 1987; Roisin, 2000). Then, it is possible that Valkyries do not predate sterile soldiers, although the oldest known Valkyrie, reported herein, is older than the first occurrences of true apterous soldiers (e.g., Engel et al., 2016; Zhao et al., 2020; Jouault et al., 2021). But given the nature of the fossil record, particularly for Isoptera, such absences of occurrence cannot be equated with evidence of actual absence. In other words, Valkyries would be relatively recent and convergent, and would express genetic pathways of soldiers already present at the time. This would imply that defensive weapons would be concomitantly expressed in two different castes, a situation only observed today with the fontanelle of neoisopterans. But why would Valkyries have evolved in taxa that already had a soldier caste? One important driver could be the supposedly strong selective pressure endured during the foundation of the colony, when no soldier is yet present. In several extant species, the first soldiers emerge months after the colony foundation (e.g., Mensa-Bonsu, 1976; Armua, 1998; Hanus et al., 2006), although they might develop earlier in some more derived species (e.g., Rhinotermitidae: Hanus et al., 2006). In Valkyries, founders would have been able to defend their incipient colonies until the development of the first soldiers. Both Valkyries are known from a period called the Cretaceous Terrestrial Revolution (KTR) (between 125 and 80 Ma) with major paleoenvironmental changes, including intense diversification of angiosperms and ants (Lloyd et al., 2008) that may have favored the development of this new morph (Figure 3).
But later, why did Valkyries disappear given the persistent threats faced by defenseless incipient colonies? The loss of Valkyries may have resulted from a trade-off. Producing alates with defensive weapons, particularly a large head filled with muscles or cuticular projections as in Valkyritermes, is costly and may extend their time of development (Figure 3). If sterile soldiers became increasingly effective—which remain to be studied because soldiers were already quite diversified 100 Ma—and sometimes developed precociously (Hanus et al., 2006), the advantages provided by defensively capable alates would have been less important, allowing for alates to be freed from defensive purposes. This better task partitioning would have eliminated much of the danger to the reproductives, preventing them from risking their life—and that of their colonies—if threatened (Figure 3).
Valkyries as Early as the Proto-Termite Colonies?
Alternatively, the discovery of Valkyries in two early diverging lineages from >90 Ma in distant deposits could indicate that Valkyries may have been present in other termite families at the same period or even before. Also, because the fossilization process of an organism is extraordinary and occurs rarely, assuming that Valkyries were more abundant than what we observe is plausible. Thus, Valkyries could have been present since the beginning of termite evolutionary history, which would imply that Valkyries from other early diverging lineages remain to be unearthed. But, more interestingly, it would suggest that Valkyries would result from a relic developmental pathway, a pathway that is no longer observed in extant termites. This would be in line with the high developmental flexibility of early lineages with totipotent pseudergates like Kalotermitidae, in which manipulations using juvenile hormone or transplants of corpora allata produced soldiers with reproductive features (Lebrun, 1970; Myles, 1980; Myles and Chang, 1984), although admittedly kalotermitids are more derived than the earliest termites and may not be reflective of groundplan conditions.
In this scenario, and following current hypotheses about proto-termites (e.g., Nalepa, 1988, 1994; Thorne, 1997), defense would have been ensured by the founders (i.e., Valkyries) of comparatively small colonies, like those of most Archotermopsidae, Hodotermitidae, and many other non-termitids today (Figure 3). This scenario would further indicate that a subsequent task partitioning would have been selected, with a defense-dedicated soldier caste, which could evolve all the more easily given that the genetic material for defensive morphologies was already present in Valkyries (Figure 3). In this scenario, however, Valkyries would not necessarily predate soldiers (i.e., Valkyries and soldiers could have evolved contemporaneously) and some of them, at least, would have persisted in some lineages for several million years (Figure 4A).
Interestingly, Valkyries are present in lineages that today are known to have either a “wood-nesting” type and a late separation between the “normal” alate and soldier paths (e.g., Kalotermitidae, “linear” pathway; Figure 4A) or a “foraging” type and an early separation between “normal” alate and soldier paths (Revely et al., 2021; “bifurcated” pathway). One could hypothesize that the presence of Valkyries would be more likely in highly flexible lineages (i.e., “wood-nesting” type) than in “foraging” lineages because individuals engage irreversibly in a given path much later (Revely et al., 2021). If this were the case, then the presence of Valkyries in both types could reflect that in most lineages, including some “foraging” ones, developmental pathways were supposedly ancestrally “linear” (Noirot, 1985; Legendre et al., 2013; but see Watson and Sewell, 1985).
Striking Morphology for Novel Ecological Implications
Both scenarios—convergent or plesiomorphic Valkyries—have their own advantages and limitations in terms of explanatory power. A convergent origin is a parsimonious hypothesis as to the current records of Valkyries but it raises the question of why they originated and then disappeared without obvious changes in terms of selective pressures (although we admit that critical selective pressures are not always easily observed, particularly in distant palaeoecological settings). On the other hand, finding Valkyries from other lineages could contribute to tip the scale in favor of the plesiomorphic Valkyries hypothesis (Figure 4A). In both cases, however, the prime mover for Valkyries would be external threats, whether from other termite colonies, ants, or other predators. Those selective pressures could have also played a role in the origin of the soldier caste, which then would have evolved, not from neotenics and intra-colonial aggressions, but from helpers (Šobotník et al., 2010a contra Thorne et al., 2003).
Termite soldiers have acquired many morphological, chemical, and physiological innovations contributing to an effective defense of their nests. Among them, arguably the most spectacular example of extensive morphological modification acquired for defense is the “phragmotic” head, used to plug nest entrances and prevent the entry of enemies. The head of extant alate termites does not possess defensive modifications, other than the fontanelle that is also present in corresponding soldiers and is associated with a defensive secretion in most lineages. The phragmotic head of Valkyritermes is remarkable (Figures 1, 2B,C) because it is the first record of such unique soldier-like specialization in a winged termite, and the earliest morphological evidence of defense in winged termites.
Arguably, Valkyries would have an increased fitness compared to that of founders devoid of defensive “weapons”. Recording their particular morphology during the KTR is noteworthy as these events have presumably affected Isoptera like many other lineages. Therefore, it is unsurprising that striking morphologies and an early ability to defend incipient colonies were developed during this period (Figure 4B), perhaps to cope with the diversification of termite predators (i.e., ants: Barden and Grimaldi, 2016; small insectivorous reptiles: Daza et al., 2016). The Cretaceous is a transitional period for eusocial insects during which they begin to diversify (e.g., Branstetter et al., 2017; Jouault et al., 2021: Figure 5), maybe because of the rapid angiosperm diversification (Silvestro et al., 2021). Stem groups or transitional lineages of eusocial insects are found at this time (e.g., Barden et al., 2020), sometimes from complete assemblages with unusual morphologies and that disappeared after the K-Pg crisis (e.g., Jiang et al., 2021). The drivers of these extinctions remain to be established.
Conclusion
The Valkyries reported here show that, in termites, the division of reproductive and defensive tasks was not always complete. This raises the question of how task partitioning evolved in these insects, a topic intertwined with the evolution of their developmental plasticity. Valkyries show that the full extent of this plasticity is today lost (perhaps through canalization of certain gene expressions), better controlled, and only found when undifferentiated helpers (i.e., pseudergates) are subject to hormonal modifications (Myles and Chang, 1984). As a result, Valkyries represent a missing alternative or a relic evolutionary “experiment” from ancient times (Figure 4A).
The discovery of Valkyries suggests that phragmosis arose at least in the Early Cretaceous (Figure 4), a period of major environmental changes, and possibly developed under selective pressures from other gallery insect invaders against which Valkyries would likely have played a critical defensive role for their incipient colonies. We argue that the simultaneous presence of Valkyries and soldiers in termite colonies is plausible so that Valkyries do not necessarily pre-date soldiers (Figure 4). Valkyries would have disappeared in favor of a caste strictly devoted to the defense of the nest, probably when developmental pathways may have become more canalized and better defined. It remains to be discovered what proto-termite colonies might have been like and how they were defended before the appearance of soldier morphologies (either as true soldiers or Valkyries).
Data Availability Statement
All data needed to evaluate the conclusions are present in the manuscript. All data related to this manuscript are available from the corresponding authors upon reasonable request. Nomenclatural acts established herein are registered in ZooBank: urn:lsid:zoobank.org:pub:2831A9DD-63AD-43DE-AC7F-3446035520FD.
Author Contributions
CJ and AN designed the research. CJ, ME, and AN participated in morphological studies and drafted the manuscript, to which FL, JB, PG, DH, and EP, contributed. CJ, FL, ME, and AN revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by French National Research Agency grant no. ANR-19-CE02-0023 (project SOCIOGENOMICS), the Second Tibetan Plateau Scientific Expedition and Research (2019QZKK0706), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB26000000), and the National Natural Science Foundation of China (41925008 and 41688103).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank AJ (editor) for managing the article during the editing process and for its responsiveness to our various requests. This manuscript greatly benefited from the comments and insightful corrections of two reviewers. This manuscript was contributed by CJ and JB during their Ph.D. projects.
References
Armua, A. C. (1998). Study of the postembryonic development of Nasutitermes corniger (Isoptera: Termitidae) under laboratory conditions. Revista de la Soc. Entomol. Argent. 57, 29–32.
Barden, P., and Grimaldi, D. (2016). Adaptive radiation in socially advanced stem-group ants from the Cretaceous. Curr. Biol. 26, 515–521. doi: 10.1016/j.cub.2015.12.060
Barden, P., Perrichot, V., and Wang, B. (2020). Specialized predation drives aberrant morphological integration and diversity in the earliest ants. Curr. Biol. 30, 3818–3824. doi: 10.1016/j.cub.2020.06.106
Brandão, C. R. F., Diniz, J. L. M., Silva, P. R., Albuquerque, N. L., and Silvestre, R. (2001). The first case of intranidal phragmosis in ants. The ergatoid queen of Blepharidatta conops (Formicidae, Myrmicinae) blocks the entrance of the brood chamber. Insect. Soc. 48, 251–258. doi: 10.1007/PL00001774
Branstetter, M. G., Danforth, B. N., Pitts, J. P., Faircloth, B. C., Ward, P. S., Buffington, M. L., et al. (2017). Phylogenomic insights into the evolution of stinging wasps and the origins of ants and bees. Curr. Biol. 27, 1019–1025. doi: 10.1016/j.cub.2017.03.027
Chouvenc, T., and Su, N. -Y. (2017). Irreversible transfer of brood care duties and insights into the burden of caregiving in incipient subterranean termite colonies. Ecol. Entomol. 42, 777–784. doi: 10.1111/een.12443
Cisneros, J.C., Raja, N.B., Ghilardi, A.L., Dunne, E.M., Pinheiro, F.L., Regalado Fernández, O.M., et al. (2022). Digging deeper into colonial palaeontological practices in modern day Mexico and Brazil. Royal Soc. Open Sci. 9:210898. doi: 10.1098/rsos.210898
Crespi, B. J. (1992). Eusociality in Australian gall thrips. Nature 359, 724–726. doi: 10.1038/359724a0
Cruickshank, R. D., and Ko, K. (2003). Geology of an amber locality in the Hukawng Valley, Northern Myanmar. J. Asia. Earth Sci. 21, 441–455. doi: 10.1016/S1367-9120(02)00044-5
Daza, J. D., Stanley, E.L., Wagner, P., Bauer, A.M., and Grimaldi, D.A. (2016). Mid-Cretaceous amber fossils illuminate the past diversity of tropical lizards. Sci. Adv. 2:e1501080. doi: 10.1126/sciadv.1501080
Engel, M. S., Barden, P., Riccio, M. L., and Grimaldi, D. A. (2016). Morphologically specialized termite castes and advanced sociality in the Early Cretaceous. Curr. Biol. 26, 522–530. doi: 10.1016/j.cub.2015.12.061
Engel, M. S., Grimaldi, D.A., and Krishna, K. (2007). Primitive termites from the Early Cretaceous of Asia (Isoptera). Stuttgarter Beiträge zur Naturkunde 371, 1–32.
Engel, M.S., and Delclòs, X. (2010). Primitive termites in Cretaceous amber from Spain and Canada (Isoptera). J. Kansas Entomol. Soc. 83, 111–128. doi: 10.2317/JKES0908.06.1
Grimaldi, D. A., and Ross, A. J. (2017). “Extraordinary Lagerstatten in amber, with particular reference to the Cretaceous of Burma”. in Terrestrial Conservation Lagerstatten: Windows into the Evolution of Life on Land, eds N Fraser and H. D. Sues (Edinburgh: Dunedin Academic Press), ?287–329.
Grüter, C., Menezes, C., Imperatriz-Fonseca, V. L., and Ratnieks, F. L. W. (2012). A morphologically specialized soldier caste improves colony defense in a Neotropical eusocial bee. Proc. Natl. Acad. Sci. U.S.A. 109, 1182–1186. doi: 10.1073/pnas.1113398109
Hanus, R., Šobotník, J., Valterová, I., and Lukáš, J. (2006). The ontogeny of soldiers in Prorhinotermes simplex (Isoptera, Rhinotermitidae). Insect. Soc. 53, 249–257. doi: 10.1007/s00040-006-0865-x
Hare, L. (1937). Termite phylogeny as evidenced by soldier mandible development. Ann. Entomol. Soc. Am. 30, 459–486. doi: 10.1093/aesa/30.3.459
Jiang, R. X., Zhang, H. R., Eldredge, K. T., Song, X. B., Li, Y. Y., Tihelka, E., et al. (2021). Further evidence of Cretaceous termitophily: description of new termite hosts of the trichopseniine Cretotrichopsenius (Coleoptera: Staphylinidae), with emendations to the classification of lower termites (Isoptera). Palaeoentomology 4, 374–389. doi: 10.11646/palaeoentomology.4.4.13
Jouault, C., Legendre, F., Grandcolas, P., and Nel, A. (2021). Revising dating estimates and the antiquity of eusociality in termites using the fossilized birth-death process. Syst. Entomol. 46, 592–610. doi: 10.1111/syen.12477
Korb, J. (2016). Genes underlying reproductive division of labor in termites, with comparisons to social Hymenoptera. Front. Ecol. Evol. 4:45. doi: 10.3389/fevo.2016.00045
Krishna, K. A. (1961). Generic revision and phylogenetic study of the family Kalotermitidae. Bull. Am. Museum Nat. Hist. 122, 303–408.
Krishna, K., Grimaldi, D. A., Krishna, V., and Engel, M. S. (2013). Treatise on the Isoptera of the world. Bull. Am. Museum Nat. Hist. 377, 1–2704. doi: 10.1206/377.1
Lebrun, D. (1970). Intercastes expérimentaux de Calotermes flavicollis Fabr. Insect. Soc. 17, 159–176. doi: 10.1007/bf02226190
Legendre, F., Whiting, M. F., and Grandcolas, P. (2013). Phylogenetic analyses of termite post-embryonic sequences illuminate caste and developmental pathway evolution. Evol. Dev. 15, 146–157. doi: 10.1111/ede.12023
Legendre, F., Whiting, M. F., Bordereau, C., Cancello, E. M., Evans, T. A., and Grandcolas, P. (2008). The phylogeny of termites (Dictyoptera: Isoptera) based on mitochondrial and nuclear markers: implications for the evolution of the worker and pseudergates castes, and foraging behaviors. Mol. Phylogen. Evol. 48, 615–627. doi: 10.1016/j.ympev.2008.04.017
Lloyd, G. T., Davis, K. E., Pisani, D., Tarver, J. E., Ruta, M., Sakamoto, M., et al. (2008). Dinosaurs and the Cretaceous Terrestrial Revolution. Proc. Royal Soc B 275, 2483–2490. doi: 10.1098/rspb.2008.0715
Lo, N., Engel, M. S., Stephen, C., Christine, A., Gaku, T., Grimaldi, D. A., et al. (2007). Save Isoptera: A comment on Inward, et al. Biol. Lett. 3, 562–563. doi: 10.1098/rsbl.2007.0264
Mensa-Bonsu, A. (1976). The biology and development of Prorotermes adamsoni (Froggatt) (Isoptera, Hodotermitidae). Insect. Soc. 23, 155–166. doi: 10.1007/bf02223848
Miura, T., and Maekawa, K. (2020). The making of the defensive caste: physiology, development, and evolution of the soldier differentiation in termites. Evol. Dev. 22, 425–437. doi: 10.1111/ede.12335
Myles, T. G. (1980). The production of soldier alate double castes with juvenile hormone analog in Neotermes connexus Kalotermitidae Isoptera. Am. Zool. 20:937.
Myles, T. G. (1986). Reproductive soldiers in the Termopsidae (lsoptera). Pan Pacific Entomol. 62, 293–299.
Myles, T. G. (1999). Review of secondary reproduction in termites (Insecta: Isoptera) with comments on its role in termite ecology and social evolution. Sociobiology 33, 1–91.
Myles, T. G., and Chang, F. (1984). The caste systems and caste mechanisms of Neotermes connexus (Isoptera: Kalotermitidae). Sociobiology 9, 163–321.
Nalepa, C. A. (1988). Cost of parental care in the woodroach Cryptocercus punctulatus Scudder (Dictyoptera: Cryptocercidae). Behav. Ecol. Sociobiol. 23, 135–140. doi: 10.1007/bf00300348
Nalepa, C. A. (1994). “Nourishment and the origin of termite eusociality”. In Nourishment and Evolution in Insect Societies, eds J. H. Hunt and C. A. Nalepa (Boulder, Colorado: Westview Press), 57–104.
Nalepa, C. A. (2015). Origin of termite eusociality: trophallaxis integrates the social, nutritional, and microbial environments. Ecol. Entomol. 40, 323–335. doi: 10.1111/een.12197
Noirot, C. (1956). Les sexués de remplacement chez les Termites supérieurs (Termitidae). Insect. Soc. 3, 145–158. doi: 10.1007/BF02230675
Noirot, C. (1985). “Pathways of caste development in the lower termites”. In Caste Differentiation in Social Insects, eds J. A. L. Watson, B. M. Okot-Kotber, and C. Noirot. (Oxford: Pergamon Press), 75–86. doi: 10.1016/b978-0-08-030783-1.50011-2
Noirot, C., and Pasteels, J. M. (1987). Ontogenetic development and the evolution of the worker caste in termites. Experientia 43, 851–952. doi: 10.1007/bf01951642
Queller, D. C., and Strassmann, J. E. (1998). Kin selection and social insects: social insects provide the most surprising predictions and satisfying tests of kin selection. BioScience 48, 165–175. doi: 10.2307/1313262
Revely, L., Sumner, S., and Eggleton, P. (2021). The plasticity and developmental potential of termites. Front. Ecol. Evol. 9:552624. doi: 10.3389/fevo.2021.552624
Roisin, Y. (2000). Diversity and evolution of caste patterns. In Termites: Evolution, Sociality, Symbioses, Ecology, eds T. Abe, D. E. Bignell, and M. Higashi (Dordrecht: Kluwer). 95–119 doi: 10.1007/978-94-017-3223-9_5
Roisin, Y. and Pasteels, J. M. (1985). Imaginal polymorphism and polygyny in the Neo-Guinean termite Nasutitermes princeps (Desneux). Insect. Soc. 32, 140–157. doi: 10.1007/BF02224229
Schubnel, T., Desutter-Grandcolas, L., Legendre, F., Prokop, J., Mazurier, A., Garrouste, R., Grandcolas, P., and Nel, A. (2019). To be or not to be: postcubital vein in insects revealed by microtomography. Syst. Entomol. 45, 327–336. doi: 10.1111/syen.12399
Shi, G. -H., Grimaldi, D. A., Harlow, G. E., Wang, J., Wang, J., Yang, M. C., et al. (2012). Age constraint on Burmese amber based on U-Pb dating of zircons. Cretaceous Res. 37, 155–163. doi: 10.1016/j.cretres.2012.03.014
Silvestro, D., Bacon, C. D., Ding, W., Zhang, Q., Donoghue, P. C. J., Antonelli, A., and Xing, Y. (2021). Fossil data support a pre-Cretaceous origin of flowering plants. Nat. Ecol. Evol. 5, 449–457. doi: 10.1038/s41559-020-01387-8
Smith, R. D. A., and Ross, A. J. (2017). Amberground pholadid bivalve borings and inclusions in Burmese amber: implications for proximity of resin producing forests to brackish waters, and the age of the amber. Earth Environ. Sci. Transact. Royal Soc. Edinburgh 107, 239–247. doi: 10.1017/S1755691017000287
Šobotník, J., and Dahlsjö, C. A. L. (2017). Isoptera. In Reference Module in Life Sciences. Amsterdam: Elsevier.
Šobotník, J., Bourguignon, T., Hanus, R., Demianová, Z., Pytelková, J., Mareš, M., et al. (2012). Explosive backpacks in old termite workers. Science 337:436. doi: 10.1126/science.1219129
Šobotník, J., Hanus, R., Piskorski, R., Urbanova, K., Wimmer, Z., Weyda, F., et al. (2010a). Impact of a juvenile hormone analogue on the anatomy and the frontal gland secretion of Prorhinotermes simplex (Isoptera: Rhinotermitidae). J. Insect Physiol. 56, 65–72. doi: 10.1016/j.jinsphys.2009.09.004
Šobotník, J., Jirošová, A., and Hanus, R. (2010b). Chemical warfare in termites. J. Insect Physiol. 59, 1012–1021. doi: 10.1016/j.jinsphys.2010.02.012
Stern, D. L., and Foster, W. A. (1996). The evolution of soldiers in aphids. Biol. Rev. 71, 27–79. doi: 10.1111/j.1469-185X.1996.tb00741.x
Thorne, B. L. (1997). Evolution of eusociality in termites. Ann. Rev. Ecol. Syst. 28, 27–54. doi: 10.1146/annurev.ecolsys.28.1.27
Thorne, B. L., Breisch, N. L., and Muscedere, M. L. (2003). Evolution of eusociality and the soldier caste in termites: influence of intraspecific competition and accelerated inheritance. Proc. Natl. Acad. Sci. U.S.A. 100, 12808–12813. doi: 10.1073/pnas.2133530100
Watson, J. A. L., and Sewell, J. J. (1985). Caste development in Mastotermes and Kalotermes: which is primitive? In Caste Differentiation in Social Insects, eds: J. A. L. Watson, B. M. Okot-Kotber, and C. Noirot. (Oxford: Pergamon Press), 27–40 doi: 10.1016/b978-0-08-030783-1.50008-2
Yamamura, N. (1993). Different evolutionary conditions for worker and soldier castes: genetic systems explaining caste distribution among eusocial insects. J. Theore. Biol. 161, 111–117. doi: 10.1006/jtbi.1993.1043
Yu, T., Thomson, U., Mu, L., Ross, A., Kennedy, J., Broly, P., et al. (2019). An ammonite trapped in Burmese amber. Proc. Natl. Acad. Sci. U.S.A. 116, 11345–11350. doi: 10.1073/pnas.1821292116
Keywords: insecta, Blattodea, evolution, eusociality, soldiers, phylogeny
Citation: Jouault C, Engel MS, Huang D, Berger J, Grandcolas P, Perkovsky EE, Legendre F and Nel A (2022) Termite Valkyries: Soldier-Like Alate Termites From the Cretaceous and Task Specialization in the Early Evolution of Isoptera. Front. Ecol. Evol. 10:737367. doi: 10.3389/fevo.2022.737367
Received: 06 July 2021; Accepted: 11 April 2022;
Published: 26 May 2022.
Edited by:
André Jasper, Universidade do Vale do Taquari - Univates, BrazilReviewed by:
Subir Bera, University of Calcutta, IndiaThomas Chouvenc, University of Florida, United States
Copyright © 2022 Jouault, Engel, Huang, Berger, Grandcolas, Perkovsky, Legendre and Nel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Corentin Jouault, jouaultc0@gmail.com
†ORCID: Corentin Jouault, orcid.org/0000-0002-3680-5172; Michael S. Engel, orcid.org/0000-0003-3067-077X; Diying Huang, orcid.org/0000-0002-5637-4867; Philippe Grandcolas, orcid.org/0000-0003-1374-1222; Evgeny E. Perkovsky, orcid.org/0000-0002-7959-4379; Frédéric Legendre, orcid.org/0000-0001-5900-8048; André Nel, orcid.org/0000-0002-4241-7651
‡These authors share senior authorship