
- Division of Transplantation Pathology, Thomas E. Starzl Transplantation Institute, University of Pittsburgh Medical Center, Pittsburgh, PA, USA
HLA mismatching is an important risk factor for antibody-mediated rejection and transplant failure. With the realization HLA antibodies recognize epitopes rather than antigens, it has become apparent that donor-recipient compatibility should be assessed at the epitope level. Recent developments have increased our understanding of the structural basis of HLA antigenicity, i.e., the reactivity with specific antibody and, immunogenicity, i.e., the ability to induce an antibody response. HLAMatchmaker is a computer algorithm that considers each HLA antigen as a series of small configurations of polymorphic residues referred to as eplets as essential components of HLA epitopes. This article addresses the relevance of determining epitope-specificities of HLA antibodies in the identification of acceptable mismatches for sensitized patients considered for transplantation. Permissible mismatching for non-sensitized patients aimed to prevent or reduce HLA antibody responses could consider epitope loads of mismatched antigens and the recently developed non-self–self paradigm of epitope immunogenicity.
Introduction
HLA antibodies play an important role in transplant rejection and failure and they result after exposure to mismatched HLA antigens which can occur after transplantation as well as following blood transfusions or during pregnancy. Traditionally, antibodies have been described as specific for HLA antigens such as anti-A1, anti-B7, and anti-DR1, or for serologically cross-reacting HLA antigens such as the A2-CREG and the B7-CREG. It has now become apparent that HLA antigens carry multiple epitopes which can be defined by molecular structural modeling and amino acid sequence differences between alleles. HLAMatchmaker represents an epitope-based approach to assess HLA compatibility and to select suitable donors for patients in need of an organ transplant (Duquesnoy, 2002, 2006). Three recent reviews describe the concept of HLAMatchmaker and its usefulness in HLA epitope matching for organ transplantation (Duquesnoy, 2008a, 2011a; Duquesnoy and Marrari, 2009).
Briefly, HLAMatchmaker considers each HLA antigen as a string of amino acid configurations as key elements of epitopes that can elicit specific alloantibodies. The original version used triplets, i.e., linear sequences of three residues (Duquesnoy, 2002), but the so-called eplet version is based on stereochemical modeling of protein antigen–antibody complexes and the contributions of critical amino acid residues that dominate in antigen–antibody binding (Duquesnoy, 2006). The residues of such patches are within a three Ångstrom radius of a non-self residue. Each eplet is assigned a position number in the amino acid sequence and the notation system lists only polymorphic residues marked with the standard letter code. HLAMatchmaker programs consider Class I (Duquesnoy, 2006), Class II (Duquesnoy and Askar, 2007), and MICA compatibility and antibody analysis (Duquesnoy et al., 2008a). The www.HLAMatchmaker.net website is an information resource and has Excel based analysis programs that can be downloaded free of charge.
HLA Epitope Antigenicity
Recent developments have increased our understanding of the structural basis of HLA antigenicity, i.e., reactivity with specific antibody. HLA antibodies are specific for epitopes that can be defined by single eplets or pairs consisting of a non-self eplet presented by the immunizing antigen and a self eplet shared by the antibody producer and the immunizer (Duquesnoy et al., 2005; Marrari et al., 2010).
The application of HLAMatchmaker to the analysis of antibody reactivity may increase our understanding of otherwise unexplained sensitization patterns induced by a given mismatch. For instance, sensitization to certain HLA-C mismatches can lead to antibodies reacting with epitopes shared with HLA-B antigens (Lomago et al., 2010; Duquesnoy and Marrari, 2011). These findings demonstrate that sensitization induced by an epitope on a HLA-C mismatch may cause other class I antigens to become unacceptable mismatches because they share that epitope although the patient may have never been exposed to such antigens. HLAMatchmaker can also explain unexpected reactivity patterns of class II antibodies. For instance, patients sensitized by a DR2 mismatch have often antibodies reacting with DR1 (Marrari and Duquesnoy, 2009). Such antibodies are induced by DR51 which is in strong linkage disequilibrium with DR2. They are specific for the 96EV eplet shared between DR51 and DR1. Conversely, sensitization by a DR1 mismatch can lead to antibodies that react also with DR51 but not with DR2. These findings demonstrate the importance of DRB3/4/5 eplets in DRB-specific antibody responses of kidney transplant recipients (Duquesnoy et al., 2008b).
HLA-DQ and HLA-DP heterodimers have distinct eplet repertoires and certain DQ and DP eplets react often with class II antibodies (Duquesnoy, 2008b; Duquesnoy et al., 2008b). DQ antibodies can also recognize pairs of eplets shared between DQA and DQB chains (Tambur et al., 2010).
HLA Mismatch Acceptability for Sensitized Patients
In the clinical setting of transplantation, it has become apparent that HLA epitopes rather than antigens are important for analyzing antibody specificity. The highly sensitized patient represents an enigma for kidney transplantation: not only is it difficult to find a suitably matched donor but subsequent kidney transplants are often less successful. The analysis of serum reactivity for HLA antibodies has two goals. Most commonly used is the identification of antibody-defined HLA antigens that should be considered unacceptable. This system is designed to identify donors who must be excluded but it does not necessarily mean that all other HLA antigens would be compatible for a patient. The other goal is to determine HLA antigens that are acceptable mismatches. This strategy represents a direct approach of finding a compatible donor for a sensitized patient (Claas et al., 1989, 2004).
HLAMatchmaker is a useful tool in the analysis of serum antibody reactivity of sensitized patients and the identification of potential donors with acceptable mismatches (Claas et al., 2004, 2005; Duquesnoy et al., 2004; Doxiadis et al., 2005; Goodman et al., 2006). Eurotransplant has incorporated HLAMatchmaker in the Acceptable Mismatch program to identify donors for highly sensitized patients (Claas et al., 2004, 2005; Doxiadis et al., 2005). This approach shortens the waiting time for a suitable kidney donor and leads to excellent graft survivals comparable to those seen with non-sensitized recipients (Claas et al., 2004).
HLA Immunogenicity and Epitope Load
HLAMatchmaker can be used as a quantitative tool to determine the degree of a mismatch, i.e., the number of mismatched eplets or triplets. A given HLA antigen mismatch has an epitope load that is primarily determined by the recipient’s HLA type representing a repertoire of self epitopes to which no alloantibodies can be made. For some patients, a mismatched antigen might be structurally compatible whereas for other patients it has multiple mismatched epitopes (Duquesnoy, 2008a). Table 1 shows examples of eplet mismatches of two class I alleles for eight HLA phenotypes. For each allele, one can readily identify cases with low or high eplet loads.
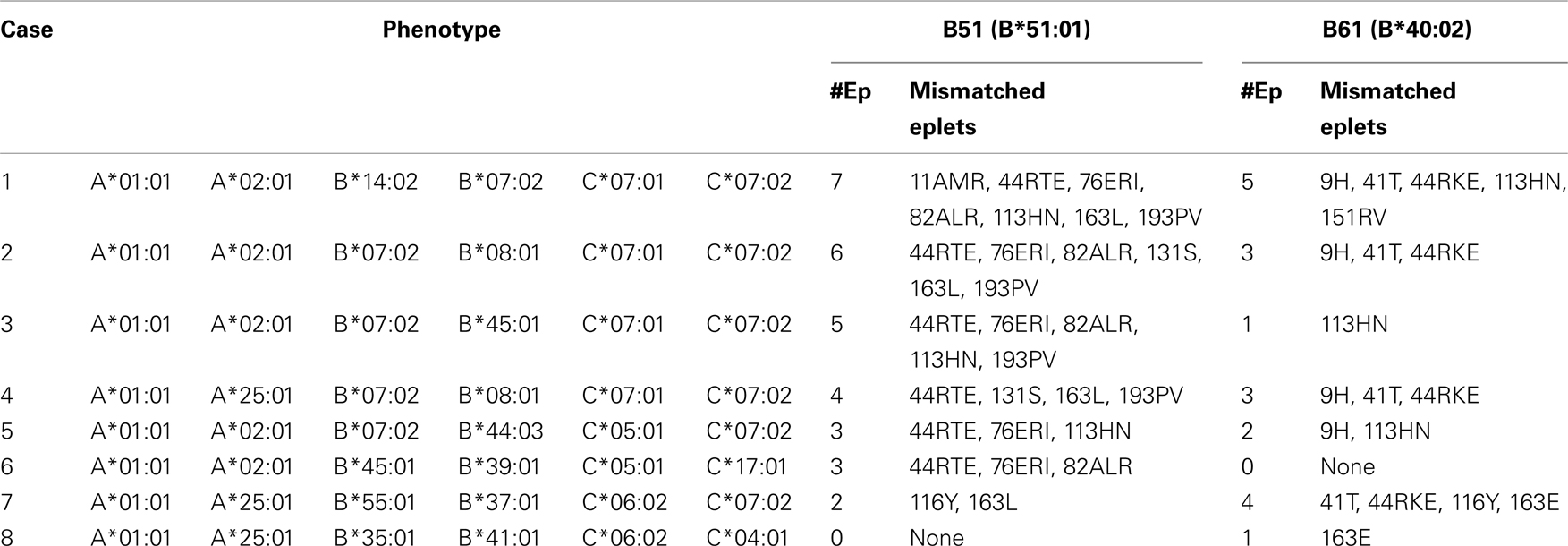
Table 1. Two examples showing how the HLA phenotype of the recipient affects the eplet load of a class I allele mismatch (Adapted from Duquesnoy, 2008b).
The incidence of the anti-class I antibody response induced by a transplant or during pregnancy correlates with the number of non-self triplets or eplets on mismatched antigens (Dankers et al., 2004; Goodman et al., 2006; Mihaylova et al., 2006; Kosmoliaptsis et al., 2008). Anti-HLA-C antibody responses by patients with rejected kidney transplants also correlate with eplet loads of HLA-C mismatches (Duquesnoy and Marrari, 2011).
Conventional class II matching criteria consider only HLA-DR antigen but this approach is an insufficient reflection of class II compatibility because antibodies against other class II mismatches including DP and DQ have been shown to diminish transplant success. Each DR antigen should be viewed as a package of DR + DQ + DP antigens and the overall class II epitope load depends on the patient’s DR, DQ, DP type. Table 2 illustrates how DR haplotypes have different epitope loads if DRB3/4/5, DQB, and DQA are included. Let as assume that DR11, 16 corresponds to DRB1*11:01, DRB3*02:01, DQB1*03:01, DQA1*05:01/DRB1*16:01, DRB5*02:02, DQB1*05:02, DQA1*01:02 genotype, and that the serological DR antigens correspond to the common DR–DQ haplotypes shown in Table 2. The DR12 and DR15 mismatches have the lowest eplet loads namely 6 and 9, whereas DR7 and DR9 have 28 and 27 mismatched eplets, respectively. The donor’s DRB1*16:02 has two eplet differences with the recipient’s DRB1*16:01. This DRB1*15:01 haplotype has no DRB1 eplet mismatches but its DRB5*01:01 and DQB1*06:02 have 2 and 7 mismatched eplets, respectively. These examples are merely intended to illustrate that high-resolution DR, DQ typing can provide detailed class II matching information at the eplet level.
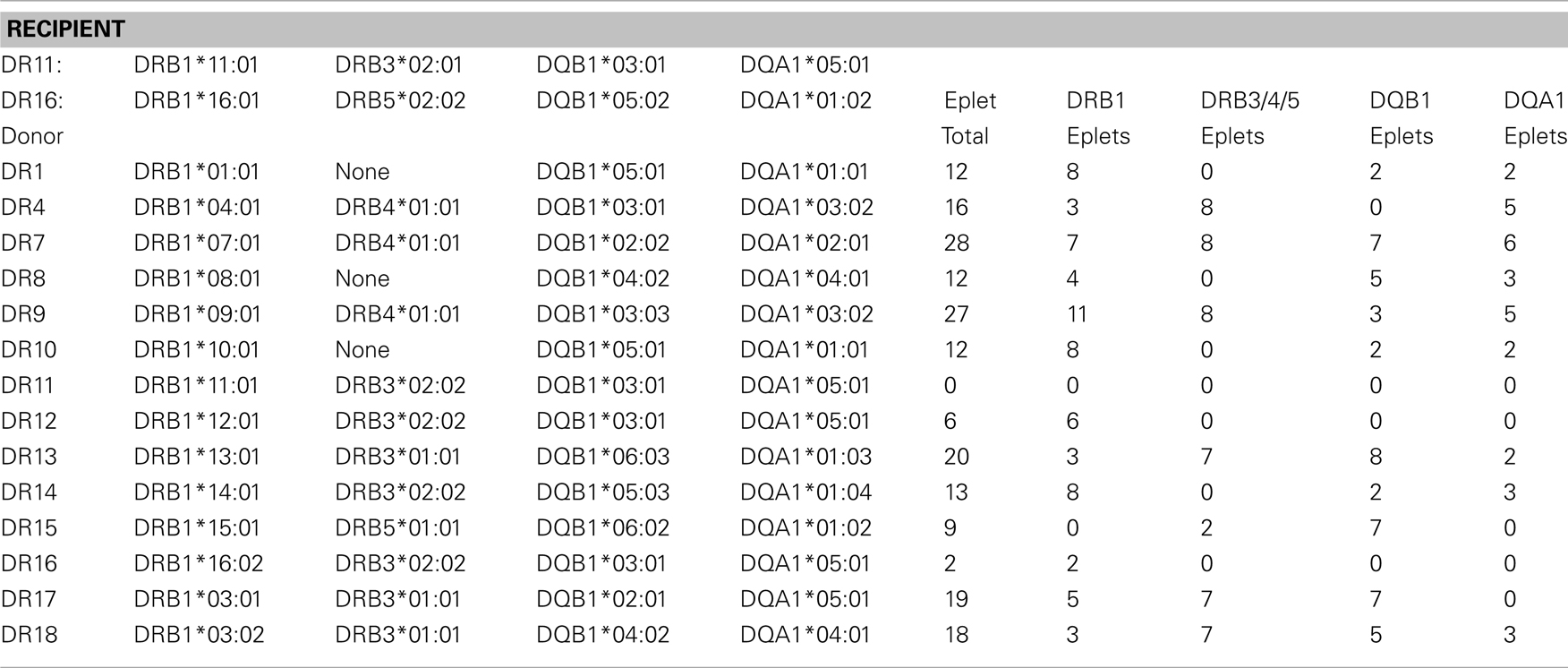
Table 2. Example showing how a high-resolution DR–DQ haplotype shows differences in class II eplet loads among serologically defined DR antigen mismatches (Adapted from Duquesnoy, 2008a).
Epitope loads of class II mismatches affect specific antibody responses. Donor-specific, DRB1-reactive antibodies are less often detectable than antibodies against other class II epitopes (Duquesnoy et al., 2008b). Antibody absence correlates with low numbers of mismatched DRB1 eplets. In contrast, donor-specific DRB3/4/5 mismatches induce more antibody responses and they have higher numbers of incompatible eplets. Anti-DQ antibodies are rather common and this correlates with more mismatched eplets on DQB and DQA than on DRB1 (Duquesnoy et al., 2008b). About one-third of class II sensitized patients have anti-DP antibodies reactive with a few DPB eplets and an allelic pair of DPA eplets.
Information about epitope loads of HLA mismatches seems clinically useful in the management of transplant patients. Epitope loads can be interpreted as risk factors for antibody-mediated rejection in the clinical management of transplant recipients and may eventually lead to new strategies for HLA mismatch permissibility to reduce alloimmunization and increase transplant survival.
The Non-Self–Self Paradigm of HLA Eplet Immunogenicity
Our studies on eplet pairs have always shown that the immunizing allele has one eplet that is non-self whereas as the other is a self eplet shared with the antibody producer (Duquesnoy et al., 2005; Marrari et al., 2010). This suggests an autoreactive component of the alloantibody response to an HLA mismatch and a recent report has expanded this view to the non-self–self paradigm of eplet immunogenicity (Duquesnoy, 2011b). This paradigm is based on current concepts about epitope–antibody interactions and the development of B-cells specific for autologous proteins.
The variable domains of immunoglobulin heavy and light chains determine antibody reactivity and each chain has three complementarity determining regions (CDRs) with hypervariable loops that interact with different parts of an epitope (Poljak et al., 1973; Chothia and Lesk, 1987). The third CDR of the heavy chain (CDR-H3) lies generally in the center of the antigen–binding site and plays a major role in epitope recognition. CDR-H3 has considerable variability in its length (Kabat et al., 1977; Wu et al., 1993; MacCallum et al., 1996; Shirai et al., 1999) whereas the other CDRs form limited numbers of so-called canonical structures that stabilize the binding with CDR-H3 (Chothia and Lesk, 1987; Kuroda et al., 2009). Stereochemical analyses of crystallized antigen–antibody complexes have defined a structural epitope as that part of the antigen that makes contact with all CDRs of antibody (Davies et al., 1990; Padlan, 1994; MacCallum et al., 1996). A structural epitope has about 15–25 surface residues and within it lies the functional epitope consisting of energetic residues that dominate the binding with antibody (Getzoff et al., 1988; Novotny, 1991; Laune et al., 1997; Van Regenmortel, 2002). Functional epitopes comprise one patch or a pair of patches of energetic residues separated far enough to be contacted by different CDRs of antibody. In order to be immunogenic, a functional epitope must have at least one non-self residue, i.e., the antibody producer’s homologous proteins must have a different residue in the corresponding sequence position. Such residues must be on the molecular surface so they can make contact with the specificity-determining CDRs of antibody. The surface of a structural epitope is about 700–900 Å2 (Davies et al., 1990; Padlan, 1994; MacCallum et al., 1996).
There is no information about the structure of an HLA antigen complexed with alloantibody but Ziegler’s group has obtained detailed information about a crystallized antigen–antibody complex involving a melanoma-associated peptide bound to HLA-A1 (Hulsmeyer et al., 2005). The structural epitope involves four contact residues of the peptide and 10 contact residues in the α helices of HLA-A1. These contact residues are on a molecular surface of about 900 Å2. What are the possible dimensions of structural HLA alloepitopes which have mismatched eplets as functional epitopes? Considering the 700–900-Å2 surface of a structural epitope one can calculate that surface residues within a 15-Å radius of a centrally located eplet could make contact with antibody. These residues can be identified with the “select by distance” command of the Cn3D structure software program (Hogue, 1997) using informative HLA molecular models downloaded from http://www.ncbi.nlm.nih.gov/Structure.
The non-self–self paradigm of eplet immunogenicity considers the hypothesis that B-cells carry low-affinity immunoglobulin receptors for self-HLA epitopes (Duquesnoy, 2011b). Their interactions with self-HLA will not lead to B-cell activation or antibody production. In contrast, exposure to HLA mismatches induces often strong alloantibody responses. It seems that the activation of a self-HLA specific B-cell by a non-self eplet requires that the remainder of the structural epitope on the immunizing antigen has considerable structural similarity with the corresponding self-HLA epitope of the antibody producer. To assess the validity of the non-self–self paradigm, we have determined polymorphic residue differences within a 15-Å radius of an eplet on the immunizer and the alleles of the antibody producer. The goal is to identify the antibody producer’s structural epitope that has an identical or very similar residue composition as the immunizing epitope with the mismatched eplet.
Table 3 summarizes data in three recent publications as experimental support of this hypothesis (Duquesnoy, 2011b; Marrari et al., 2011; Duquesnoy et al., submitted). For each antibody response, at least one allele of the antibody producer has no or few differences with the immunizing allele in antibody-accessible positions defined by a 15-Å radius of the mismatched eplet, the presumed dimension of a structural epitope. Most of the data were obtained with human monoclonal antibodies (produced by Arend Mulder, Leiden University Medical Center) that were specific for eplets or eplet pairs. In each case, the antibody producer had at least one allele which had no or very residue differences with the immunizing allele; they are highlighted in Table 3. Two patients had antibodies against the 145R + 82LR pair presented by the immunizing B*44:02; this allele has no residue differences with the self B*13:02 allele except for 145L rather than 145R. It should be noted that both eplets are present on one or more alleles of the antibody producer and, according to HLAMatchmaker, they are considered intralocus and interlocus matches which should not induce antibodies. None of the alleles of these antibody producers had however, the combination of these eplets and the non-self–self paradigm for HLA epitope immunogenicity has offered a ready explanation why the pair of self-145R and self-82LR eplets presented by B*44:02 had induced specific antibodies. The B*15:01 induced monoclonal 12 is also specific for a pair of self eplets, namely s69TNT + s80NRG (Table 3). Within the 15-Å radius of this pair, the B*07:02 allele of the antibody producer had no residue differences with B*15:01.
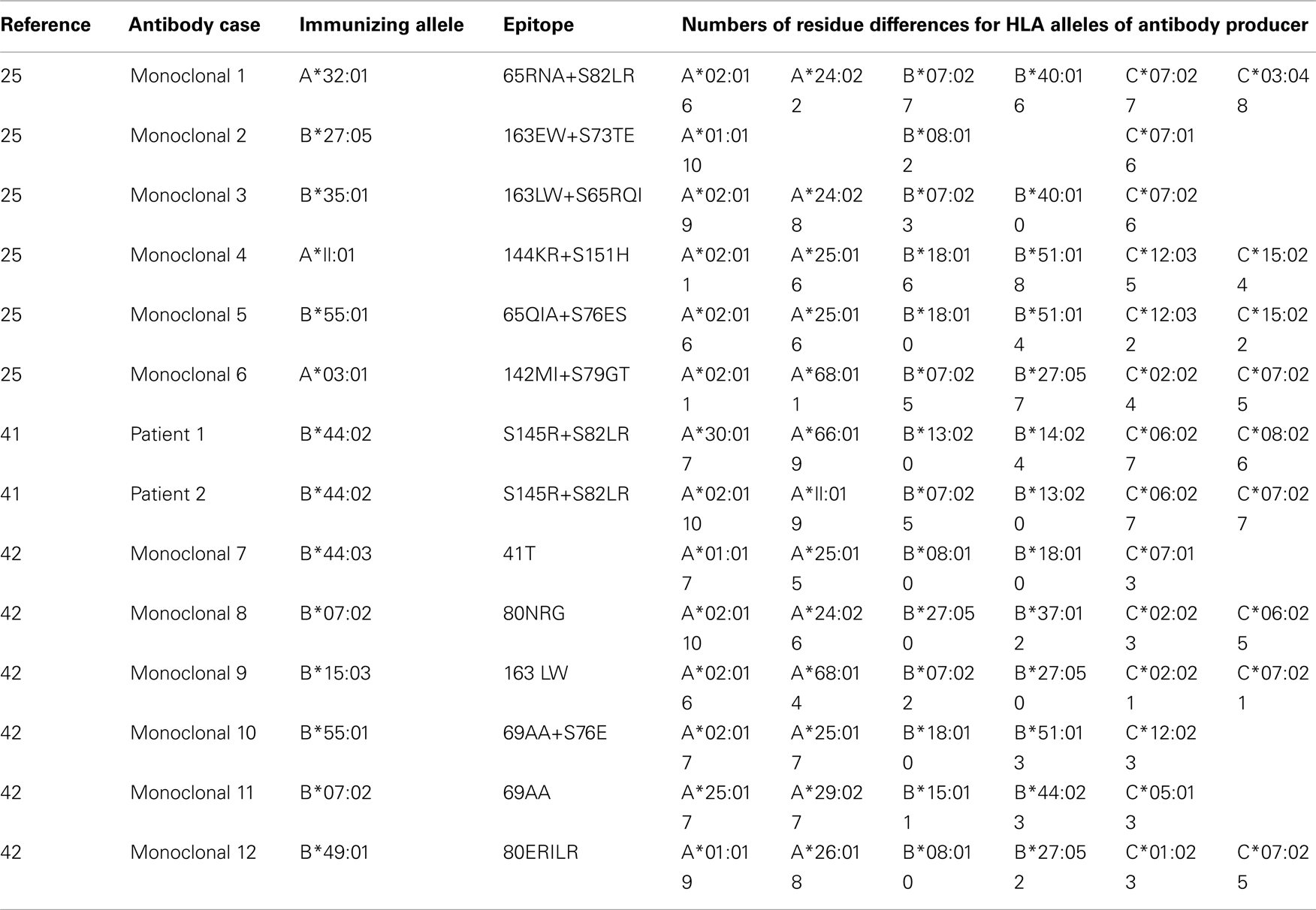
Table 3. Examples of polymorphic residue differences within 15 Å of mismatched eplets on antibody-defined epitopes on immunizing alleles and the alleles of the antibody producer.
These findings support the concept that HLA antibodies originate from B-cells with self-HLA Immunoglobulin receptors that recognize mismatched eplets as non-self entities on immunizing antigens. The humoral alloresponse to an HLA mismatch is not well understood. It is well known that sensitized patients develop specific antibodies to a restricted number of mismatched epitopes (Duquesnoy et al., 1990; Rodey et al., 1994). The non-self–self paradigm of HLA epitope immunogenicity may explain this phenomenon. It is possible that the antibody response to an eplet requires that its structural epitope on the immunizing HLA antigen must be structurally very similar to the corresponding self epitope of the antibody producer. Any antigen with significant structural epitope differences with patient’s self epitopes might prevent B-cell activation and subsequent HLA antibody production. The non-self–self concept of HLA immunogenicity may become clinically useful regarding predicting antibody responses to HLA mismatches but it needs of course, experimental verification in the clinical setting.
Conclusion
Prevention of HLA sensitization represents a significant challenge for the non-sensitized transplant candidate. Two causes, namely blood transfusions and the transplant itself should be approached as being potentially preventable. Perfect HLA matching is somewhat impractical because it can be done for small numbers of patients. On the other hand, information about epitope loads and the application of the non-self–self paradigm of epitope immunogenicity may useful in assessing risks for antibody-mediated rejection after transplantation and the clinical management of transplant recipients. Such permissible mismatching may lead to new strategies to identify suitable donors with minimal risks for allosensitization.
Conflict of Interest Statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Chothia, C., and Lesk, A. (1987). Canonical structures for the hypervariable regions of immunoglobulins. J. Mol. Biol. 196, 901–917.
Claas, F. H., Gijbels, Y., von Veen, A., de Waal, L. P., D’Amaro, J., Persijn, G. G., and van Rood, J. J. (1989). Selection of cross-match negative HLA-A and/or -B mismatched donors for highly sensitized patients. Transplant. Proc. 21, 665–666.
Claas, F. H. J., Dankers, M. K., Oudshoorn, M., van Rood, J. J., Mulder, A., Roelen, D. L., Duquesnoy, R. J., and Doxiadis, I. I. N. (2005). Differential immunogenicity of HLA mismatches in clinical transplantation. Transpl. Immunol. 14, 187–191.
Claas, F. H. J., Witvliet, M., Duquesnoy, R. J., Persijn, G., and Doxiadis, I. I. N. (2004). The acceptable mismatch program as a fast tool to transplant highly sensitized patients awaiting a post-mortal kidney: short waiting time and excellent graft outcome. Transplantation 78, 190–193.
Dankers, M. K. A., Witvliet, M. D., Roelen, D. L., De Lange, P., Korfage, N., Persijn, G. G., Duquesnoy, R. J., Doxiadis, I. I. N., and Claas, F. H. J. (2004). The number of amino acid triplet differences between patient and donor is predictive for the antibody reactivity against mismatched HLA antigens. Transplantation 128, 1236–1239.
Davies, D., Padlan, E., and Sheriff, S. (1990). Antibody-antigen complexes. Annu. Rev. Biochem. 59, 439–473.
Doxiadis, I. I. N., Duquesnoy, R. J., and Claas, F. H. J. (2005). Extending options for highly sensitized patients to receive a suitable kidney graft. Curr. Opin. Immunol. 17, 536–540.
Duquesnoy, R. (2008a). Clinical usefulness of HLAMatchmaker in HLA epitope matching for organ transplantation. Curr. Opin. Immunol. 20, 594–601.
Duquesnoy, R. (2008b). HLA class II antibodies and transplant outcome (invited editorial). Transplantation 86, 638–640.
Duquesnoy, R. (2011a). Antibody-reactive epitope determination with HLAMatchmaker and its application in transplantation. Tissue Antigens 77, 525–534.
Duquesnoy, R. (2011b). The antibody response to an HLA mismatch: a model for nonself-self discrimination in relation to HLA epitope immunogenicity. Int. J. Immunogenet. doi: 10.1111/j.1744-313X.2011.01042.x. [Epub ahead of print].
Duquesnoy, R., and Marrari, M. (2011). Detection of antibodies against HLA-C epitopes in patients with rejected kidney transplants. Transpl. Immunol. 24, 164–171.
Duquesnoy, R., Mostecki, J., Hariharan, J., and Balasz, I. (2008a). A structurally based epitope analysis of MICA antibody specificity patterns. Hum. Immunol. 69, 826–832.
Duquesnoy, R., Awadalla, Y., Lomago, J., Jelinek, L., Howe, J., Zern, D., Hunter, B., Martell, J., Girnita, A., and Zeevi, A. (2008b). Retransplant candidates have donor-specific antibodies that react with structurally defined HLA-DR, DQ, DP epitopes. Transpl. Immunol. 18, 352–360.
Duquesnoy, R. J. (2002). HLAMatchmaker: a molecularly based algorithm for histocompatibility determination. I. Description of the algorithm. Hum. Immunol. 63, 339–352.
Duquesnoy, R. J. (2006). A structurally based approach to determine HLA compatibility at the humoral immune level. Hum. Immunol. 67, 847–862.
Duquesnoy, R. J., and Askar, M. (2007). HLAMatchmaker: a molecularly based algorithm for histocompatibility determination V. Eplet matching for HLA-DR, HLA-DQ and HLA-DP. Hum. Immunol. 68, 12–25.
Duquesnoy, R. J., and Marrari, M. (2009). HLAMatchmaker-based definition of structural HLA epitopes detected by alloantibodies. Curr. Opin. Organ Transplant. 14, 403–409.
Duquesnoy, R. J., Mulder, A., Askar, M., Fernandez-Vina, M., and Claas, F. H. J. (2005). HLAMatchmaker-based analysis of human monoclonal antibody reactivity demonstrates the importance of an additional contact site for specific recognition of triplet-defined epitopes. Hum. Immunol. 66, 749–761.
Duquesnoy, R. J., White, L. T., Fierst, J. W., Vanek, M., Banner, B. F., Iwaki, Y., and Starzl, T. E. (1990). Multiscreen serum analysis of highly sensitized renal dialysis patients for antibodies toward public and private class I HLA determinants. Implications for computer-predicted acceptable and unacceptable donor mismatches in kidney transplantation. Transplantation 50, 427–437.
Duquesnoy, R. J., Witvliet, M. J., Doxiadis, I. I. N., de Fijter, H., and Claas, F. H. J. (2004). HLAMatchmaker-based strategy to identify acceptable HLA class I mismatches for highly sensitized kidney transplant candidates. Transpl. Int. 7, 31–38.
Getzoff, E. D., Tainer, J. A., Lerner, R. A., and Geysen, H. M. (1988). The chemistry and mechanisnm of antibody binding to protein antigens. Adv. Immunol. 43, 1–98.
Goodman, R., Taylor, C., O’Rourke, C., Lynch, A., Bradley, A., and Key, K. (2006). Utility of HLAMatchmaker and single-antigen HLA-antibody detection beads for identification of acceptable mismatches in highly sensitised patients awaiting kidney transplantation. Transplantation 81, 1331–1336.
Hogue, C. (1997). Cn3D: a new generation of three-dimensional molecular structure viewer. Trends Biochem. Sci. 22, 314–316.
Hulsmeyer, M., Chames, P., Hillig, R., Stanfield, R., Held, G., Coulie, P., Alings, C., Wille, G., Saenger, W., Uchanska-Ziegler, B., Hoogenboom, H., and Ziegler, A. (2005). A major histocompatibility complex-peptide-restricted antibody and T cell receptor molecules recognize their target by distinct binding modes. Crystal structure of human leukocyte antigen (HLA)-A1 -MAGE-A1 complex with Fab-Hyb3. J. Biol. Chem. 280, 2972–2980.
Kabat, E. A., Wu, T., and Bilofsky, H. (1977). Unusual distribution of amino acids in complementarity-determining (hypervarible) segments of heavy and light chains of immunoglobulins and their possible roles in specificity of antibody-combining sites. J. Biol. Chem. 252, 6609–6617.
Kosmoliaptsis, V., Bradley, J., Sharples, L., Chaudhry, A., Key, T., Goodman, R., and Taylor, C. (2008). Predicting the immunogenicity of human leukocyte antigen class I alloantigens using structural epitope analysis determined by HLAMatchmaker. Transplantation 85, 1817–1825.
Kuroda, D., Shirai, H., Kobori, M., and Nakamura, H. (2009). Systematic classification of CDR-L3 in antibodies: implications of the light chain subtypes and the VL-VH interface. Proteins 75, 139–146.
Laune, D., Molina, K., Ferrieres, G., Mani, J.-C., Cohen, P., Simon, D., Bernardi, T., Piechaczyk, M., Pau, B., and Granier, C. (1997). Systematic exploration of the antigen binding activity of synthetic peptides isolated from the variable regions of immunoglobulins. J. Biol. Chem. 272, 30937–30944.
Lomago, J., Jelenik, L., Zern, D., Howe, J., Martell, J., Zeevi, A., and Duquesnoy, R. (2010). How did a patient who types for HLA-B*4403 develop antibodies that react with HLA-B*4402? Hum. Immunol. 71, 176–178.
MacCallum, R. M., Martin, A. C. R., and Thornton, J. M. (1996). Antibody-antigen interactions: contact analysis and binding site topography. J. Mol. Biol. 262, 732–745.
Marrari, M., Conca, R., Praticò-Barbato, L., Amoroso, A., and Duquesnoy, R. (2011). Brief report: why did two patients who type for HLA-B13 have antibodies that react with All Bw4 antigens except HLA-B13? Transpl. Immunol. 25, 217–220.
Marrari, M., and Duquesnoy, R. (2009). Why can sensitization by a HLA-DR2 mismatch lead to antibodies that react also with HLA-DR1? Hum. Immunol. 70, 403–409.
Marrari, M., Mostecki, J., Mulder, A., Balazs, I., Claas, F., and Duquesnoy, R. (2010). Human monoclonal antibody reactivity with HLA class I epitopes defined by pairs of mismatched eplets and self eplets. Transplantation 90, 1468–1472.
Mihaylova, A., Baltadjieva, D., Boneva, P., Ivanova, M., Penkova, K., Marinova, D., Mihailova, S., Paskalev, E., Simeonov, P., and Naumova, E. (2006). Clinical relevance of anti-HLA antibodies detected by flow-cytometry bead-based assays – single-center experience. Hum. Immunol. 67, 787–794.
Poljak, R., Amzel, L., Avey, H., Chen, B., Phizackerley, R., and Saul, F. (1973). Three-dimensional structure of the Fab’ fragment of a human immunoglobulin at 2,8-A resolution. Proc. Natl. Acad. Sci. U.S.A. 70, 3305–3310.
Rodey, G. E., Neylan, J. F., Whelchel, J. D., Revels, K. W., and Bray, R. A. (1994). Epitope specificity of HLA class I alloantibodies. I. Frequency analysis of antibodies to private versus public specificities in potential transplant recipients. Hum. Immunol. 39, 272–280.
Shirai, H., Kidera, A., and Nakamura, H. (1999). H3-rules: identification of CDR-H3 structures in antibodies. FEBS Lett. 455, 188–197.
Tambur, A., Leventhal, J. R., Friedewald, J., and Ramon, D. (2010). The complexity of human leukocyte antigen (HLA)-DQ antibodies and its effect on virtual crossmatching. Transplantation 90, 1117–1124.
Van Regenmortel, M. (2002). Reductionism and the search for structure-function relationships in antibody molecules. J. Mol. Recognit. 15, 240–247.
Keywords: HLA antibody, HLA epitope, HLAMatchmaker, eplet, non-self–self paradigm of HLA epitope immunogenicity
Citation: Duquesnoy RJ (2011) Humoral alloimmunity in transplantation: relevance of HLA epitope antigenicity and immunogenicity. Front. Immun. 2:59. doi: 10.3389/fimmu.2011.00059
Received: 07 September 2011;
Paper pending published: 06 October 2011;
Accepted: 17 October 2011;
Published online: 08 November 2011.
Edited by:
Hildegard Theresia Greinix, Medical University of Vienna, AustriaReviewed by:
Bronwen Elizabeth Shaw, Royal Marsden NHS Foundation Trust, UKChristophe Legendre, Hôpital Necker, France
Gottfried Fischer, Medizinische Universität Wien, Austria
Georg Böhmig, Medical University Vienna, Austria
Copyright: © 2011 Duquesnoy. This is an open-access article subject to a non-exclusive license between the authors and Frontiers Media SA, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and other Frontiers conditions are complied with.
*Correspondence: René J. Duquesnoy, Thomas E. Starzl Biomedical Research Tower, Room W1552, Pittsburgh, PA 15213, USA. e-mail:ZHVxdWVzbm95ckB1cG1jLmVkdQ==