

 Zhaoyong Li3
Zhaoyong Li3- 1State Key Laboratory of Agrobiotechnology, College of Biological Sciences, China Agricultural University, Beijing, China
- 2Department of Biochemistry, Jiaozuo Teachers Colleague, Jiaozuo, Henan, China
- 3Beijing CRVAB Biotechnology Co., Ltd., Beijing, China
- 4National Engineering Laboratory for Animal Breeding, Key Laboratory of Animal Genetics and Breeding of the Ministry of Agriculture, College of Animal Science and Technology, China Agricultural University, Beijing, China
The VH replacement process is a RAG-mediated secondary recombination in which the variable region of a rearranged VHDJH is replaced by a different germline VH gene. In almost all human and mouse VH genes, two sequence features appear to be crucial for VH replacement. First, an embedded heptamer, which is located near the 3′ end of the rearranged VH gene, serves as a cryptic recombination signal sequence (3′cRSS) for the VH replacement process. Second, a short stretch of nucleotides located downstream of the 3′cRSS serve as a footprint of the original VH region, frequently encoding charged amino acids. In this review, we show that both of these two features are conserved in the VH genes of all jawed vertebrates, which suggests that the VH replacement process may be a conserved mechanism.
Introduction
In vertebrates, the largely diversified repertoire in the variable domain of immunoglobulins is initially generated through V(D)J recombination in developing B lymphocytes. V(D)J recombination is a somatic, cell type-specific process in which the separated germline variable (V), diversity (D) (only for immunoglobulin heavy chain, IgH), and joining (J) gene segments are assembled to form an exon that encodes the functional variable domain. The V(D)J recombination process is also a site-specific recombination that depends on the recognition, binding, and cleavage of recombination signal sequences (RSSs) by the enzymes encoded by the recombination-activating genes 1 and 2 (RAG-1 and RAG-2). RSSs flank all potentially functional V, D, and J gene segments. The consensus RSS consists of a conversed heptamer (CACAGTG) and an A-rich nonamer (ACAAAAACC), which are separated by approximately 12- or 23-bp spacers. V(D)J recombination occurs efficiently between two RSSs with different spacer lengths (Jung et al., 2006). Because the immunoglobulin gene loci of the IgH genes in tetrapods and teleosts are organized in a “translocon” fashion, a quasi-random selection and combination of the individual V, D, and J segments from the germline repertoire generates a large diversity of the antibody specificities (Flajnik, 2002; Nemazee, 2006). A further increase in this diversity is provided by the imprecise processing of the coding region junctions, including the deletion of nucleotides in the coding end and the addition of non-templated (N) nucleotides (Lieber et al., 2003).
Due to the stochastic nature of the V(D)J recombination process, B cell receptors (BCRs) that recognize autoantigens can trigger B cell central tolerance via anergy, clonal deletion, and receptor editing. Whereas anergy and clonal deletion inactivate or clear the self-reactive clones, receptor editing in immature B cells allows the cells to continue their immunoglobulin gene rearrangements to alter the specificity of their BCR (Nemazee, 2006). The secondary rearrangement occurs at the immunoglobulin kappa and lambda (Igκ and Igλ) loci, as previously demonstrated by many studies. However, the ongoing rearrangement of the IgH gene locus was once considered impossible because the primary rearrangement deletes the entire D locus and thus no D segments with the appropriate 5′ and 3′ 12-RSS sequences remain to join with new VH and JH segments (Gay et al., 1993; Tiegs et al., 1993; Prak and Weigert, 1995). Originally discovered in studies of murine pre-B cell lines, a type of secondary rearrangement called VH replacement, has now been documented as a receptor editing mechanism for the IgH gene in an increasing number of studies using knock-in mice and human normal and transformed B cells (Kleinfield et al., 1986; Reth et al., 1986; Chen et al., 1995; Cascalho et al., 1997; Zhang et al., 2003; Koralov et al., 2006). The VH replacement process closely resembles the mechanism of V(D)J recombination; this process uses a normal RSS of an upstream germline VH segment and a cryptic RSS (cRSS) that is embedded close to the 3′ end of the rearranged V exon to mediate a VH-to-VHDJH recombination. In addition, VH replacement usually produces junctional diversity or leads to frame-shifts in vivo (Covey et al., 1990; Chen et al., 1995; Koralov et al., 2006).
Conservation of the 3′cRSS and a Downstream Charged Amino Acid-Encoding Nucleotide Sequence in the VH Genes of Human and Mouse
In almost all known human germline VH genes (47/51), the cRSS is composed of a heptamer (TACTGTG) in the opposite orientation to the RSS of the germline VH segments. In addition, no conserved nonamer similar to the consensus nonamer is located upstream of the heptamer (Covey et al., 1990; Radic and Zouali, 1996). Similar conserved heptamers have been identified in more than 60% of the mouse VH nucleotide sequences that are available in GenBank (Chen et al., 1995). Some studies suggested that the VH replacement process is a RAG-mediated recombination process because of the detection of the double-stranded DNA breaks at the cRSS and the extrachromosomal DNA circles. Zhang et al. provided further evidence that the recombinant RAG-1/RAG-2 proteins can cleave the cRSS in vitro (Covey et al., 1990; Usuda et al., 1992; Zhang et al., 2003). Furthermore, many additional 3′ cryptic recombination signal sequence (3′cRSS)-like motifs that only contain the most conserved trinucleotide of the heptamer, 5′CAC (or 3′GTG), in both orientations of the coding region of the VH gene have been considered to play a role in VH gene revision, which is a second receptor replacement mechanism that occurs in germinal center B cells that may have undergone clonal expansion in response to antigen stimulation (Itoh et al., 2000; Wilson et al., 2000). Some predicted cRSSs that are initiated by the CAC motifs have been found to support detectable levels of recombination in extrachromosomal recombination assays (Davila et al., 2007). Therefore, any heptamer that contains a CAC motif at its 5′ end may have the potential to act as a cRSS for secondary rearrangement.
During each round of VH replacement, the recipient VH may leave a short stretch of nucleotides downstream of the 3′cRSS as a footprint. The analysis of the VH replacement footprints (the residual 3′ sequences of the replaced VH at the V-D junctions) in natural human IgH sequences by Zhang et al. indicated that the footprints frequently contribute charged amino acids to the IgH CDR3 region, regardless of the reading frame. In addition, 80% of the amino acids encoded by the 3′ end of human VH genes in all three reading frames are highly charged (Zhang et al., 2003). In the mouse, the arginine (Arg)-encoding AGA codon was also found at the 3′ end of most VH genes (Koralov et al., 2006). Previous studies have indicated that somatic mutations to Arg are common in the majority of high-affinity anti-dsDNA antibodies generated in autoimmune mice (Radic et al., 1993). Because the germline D genes and the normal VH-D and D-JH junctions of the IgH gene in the human and mouse rarely encode charged amino acids, the antibodies that contain VH replacement footprints may have a tendency to become autoreactive (Zhang et al., 2004). In addition, antibodies containing an Arg-rich CDR3 are negatively selected in a mouse strain in which the IgH repertoire is generated by VH replacement, although the level of anti-DNA antibodies in the sera of these mutant mice is still elevated (Koralov et al., 2006). A similar observation was recently made in humans. In systematic lupus erythematosus (SLE) patients, the frequency of VH replacement is significantly higher than in healthy individuals, and more than half of the autoreactive antibodies are encoded by VH replacement products with CDR3 regions that are rich in charged amino acids (Fan, 2009).
The cRSS near the 3′ end of VH genes and the charged amino acid-encoding nucleotide sequence following the 3′cRSS are conserved in both human and mouse. However, the conservation of these two features is not comprehensive to all six groups of jawed vertebrates (cartilaginous fishes, teleosts, amphibians, reptiles, birds, and mammals). Because the genomic organization of the VH genes in cartilaginous fishes and birds does not provide an advantageous condition for VH replacement (McCormack et al., 1991; Dooley and Flajnik, 2006), we will present a detailed analysis of the VH genes in the other four classes of jawed vertebrates, including six mammals (mouse, Norway rat, guinea pig, rabbit, African elephant, and gray short-tailed opossum), two reptiles (painted turtle and anole lizard), one amphibian (western clawed frog), and three teleosts (zebrafish, Atlantic salmon, and channel catfish), to determine whether these two features have been conserved throughout the evolution of jawed vertebrates.
Conservation of the 3′cRSS in the Functional VH Genes of Different Vertebrates
In our analysis, the functional germline VH sequences are available from the IMGT database (www.imgt.org) (for mouse and Norway rat), Ensembl genome database (www.ensembl.org) (for western clawed frog, painted turtle, and anole lizard) and other references (Ros et al., 2004; Danilova et al., 2005; Bengten et al., 2006; Wang et al., 2009; Yasuike et al., 2010; Guo et al., 2011, 2012).
Regarding only the canonical heptamer (TACTGTG) of the 3′cRSS, the percentage of VH genes with an embedded 3′cRSS varies widely among the listed species, from zero in the rabbit to 90.5% in the opossum (Table 1). If only those heptamers that contain the critical 3′ GTG (NNNNGTG) are considered to be functional 3′cRSSs, the 3′cRSS is present in more than 65% of the VH genes in all analyzed species except the channel catfish; in addition, the percentage of VH genes with this sequence is higher than 85% in most mammals (except the Norway rat), western clawed frog, zebrafish, and Atlantic salmon (Table 1). The first two nucleotides (GT) of the GTG motif of the 3′cRSS arise from the TGT codon for cysteine 104 (Cys104, IMGT numbering) and the third nucleotide (G) belonging to the following codon. More than 50% (67/122) of the heptamers that do not contain the GTG motif retain the GT nucleotides. In the majority of the germline VH genes from the most species analyzed below, the amino acid that follows Cys104 is alanine (Ala), which is encoded by a GCN codon (mouse 88/107, Norway rat 79/117, guinea pig 74/89, rabbit 12/12, African elephant 42/48, opossum 19/21, painted turtle 60/68, anole lizard 59/71, western clawed frog 31/38, zebrafish 32/33, and Atlantic salmon 43/50). Fanning et al. speculated that the 3′cRSS reflects the conservation of Cys104, which is critical for the structure of the H chain (Fanning et al., 1998). Our analysis supports this hypothesis, but the preference of the TGT codon for Cys104 and that of the following GCN codon for Ala is also important.
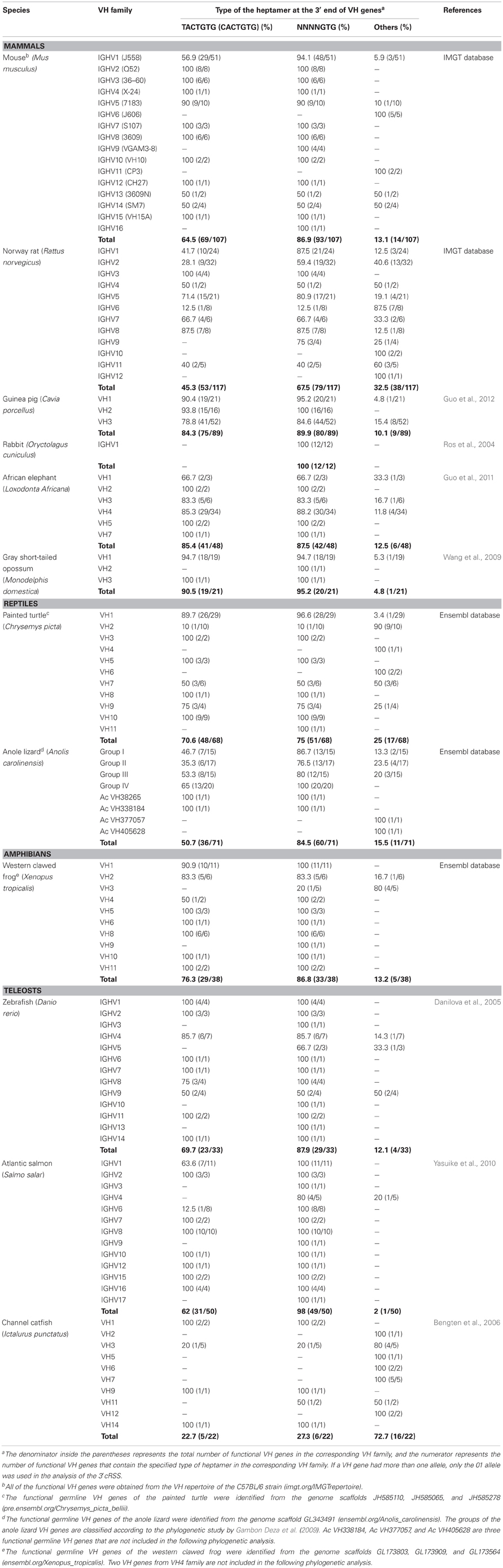
Table 1. Frequency of the 3′cRSS heptamer in the functional germline VH genes of 12 species.
To further determine whether the maintenance of the 3′cRSS is driven by an evolutionary force, we used a phylogenetic analysis to classify the VH sequences from all 12 species into eight groups. The A, B, and C groups include the mammalian clans I, II, and III, respectively, as well as a few of the VH genes from painted turtle, anole lizard, and western clawed frog. Group D only contains the reptile VH genes. Group E consists of the VH genes from painted turtle, anole lizard, western clawed frog, and teleosts; group F contains VH genes from western clawed frog and teleosts. All VH genes from group G and H belong to the teleosts (Figure 1). We then calculated the frequency of the 3′cRSS in each VH group. As shown in Table 2, group A to F possess a high proportion of VH genes that contain the canonical heptamer motif (TACTGTG > 55%). By contrast, most VH genes from two teleost-specific VH groups, G and H, do not contain the canonical heptamer motifs (TACTGTG < 40%). However, all eight groups, regardless of the divergence time, contain high the proportion of VH genes that contain the critical 3′ GTG in the heptamer motif (NNNNGTG > 69%). Because the VH genes are subjected to divergent evolution by the birth-and-death process (Ota and Nei, 1994) and six VH groups (A–F) contain genes that have persisted for a long time in living species from different classes, the high frequency of the 3′cRSS in VH groups that evolved relatively later suggests that the maintenance of the 3′cRSS was positively selected during the evolution of the VH genes.
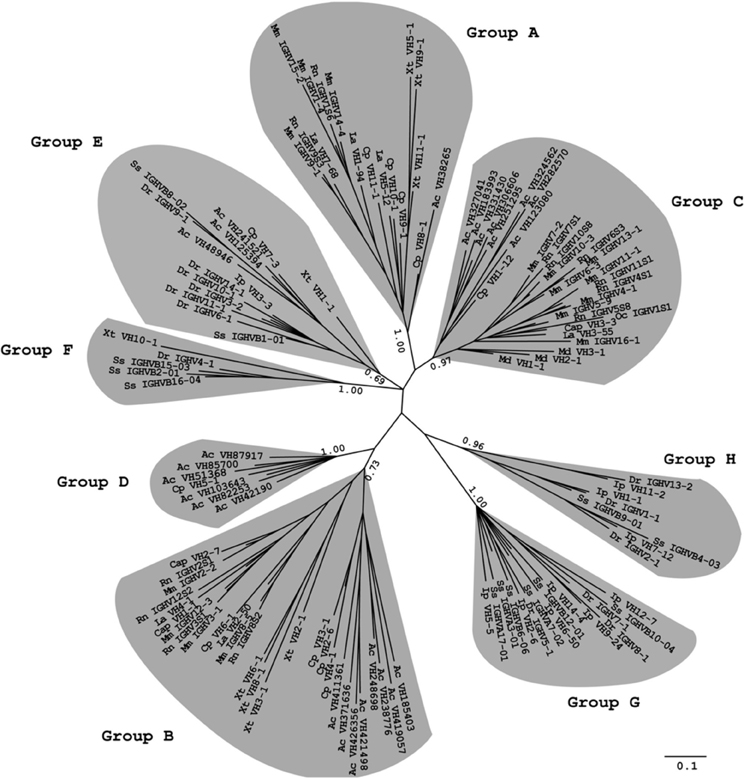
Figure 1. Phylogenetic analysis of the VH genes from 12 species. Phylogenetic trees were constructed using MrBayes3.1.2 and viewed in TreeView. Multiple DNA sequence alignments for the tree construction were performed using ClustalX2. Only the FR1-3 regions (as defined by the IMGT numbering system) of each sequence were utilized to construct the trees. Each VH family is represented with one sequence per species, which was chosen among the functional VH genes of the species. The optional VH genes of most species are named according to the nomenclature from the IMGT database (imgt.cines.fr) or other references. The only exceptions were the anole lizard, painted turtle, and western clawed frog, for which the VH genes were named using our own annotation. The species abbreviations used are as follows: Mm, mouse (Mus musculus); Rn, Norway rat (Rattus norvegicus); Cap, guinea pig (Cavia porcellus); Oc, rabbit (Oryctolagus cuniculus); La, African elephant (Loxodonta Africana); Md, gray short-tailed opossum (Monodelphis domestica); Cp, painted turtle (Chrysemys picta); Ac, anole lizard (Anolis carolinensis); Xt, western clawed frog (Xenopus tropicalis); Dr, zebrafish (Danio rerio); Ss, Atlantic salmon (Salmo salar); Ip, channel catfish (Ictalurus punctatus).
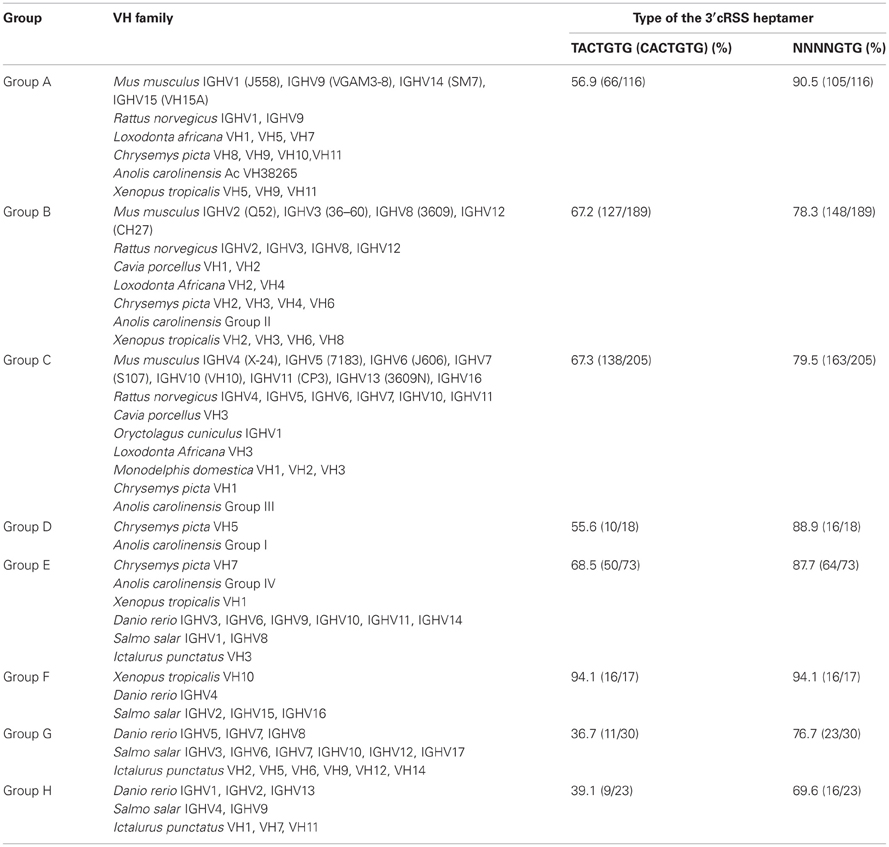
Table 2. Frequency of the 3′cRSS heptamer in the functional germline VH genes of the eight groups.
Conservation of the Downstream Charged Amino Acid-Encoding Nucleotide Sequence in the Functional VH Genes of Different Vertebrates
The germline VH genes from 11 species were analyzed to determine the frequency of charged amino acids that are encoded by the nucleotide sequence following the 3′cRSS in three reading frames (Figure 2). The length of the nucleotide sequence following the 3′cRSS is usually 7 nt in seven tetrapod species and 9 nt in teleosts. Therefore, the number of amino acids that are encoded by this sequence in three reading frames is usually one, two, and two in tetrapods and two, three, and two in teleosts (Figure 2A). Due to the high percentage of A and G nucleotides in this sequence in all 11 species, the frequency of charged amino acids that are encoded by all three reading frames is higher than the random frequency (14/64, ~22%) and the frequency of the charged amino acids encoded by functional germline DH genes (Figure 2B). In addition, it is noteworthy that the frequency of charged amino acids encoded by reading frame I is greater than 60% for all 11 species (Figure 2B). Moreover, most of the charged amino acids are positively charged (Norway rat 108/115, guinea pig 65/69, rabbit 12/12, African elephant 28/30, opossum 17/17, painted turtle 65/70, anole lizard 71/81, western clawed frog 28/34, zebrafish 35/45, Atlantic salmon 45/63, and channel catfish 27/38). The reading frame I is the only correct reading frame that can ensure the encoding of a natural H chain protein; reading frame II and III might also be found in VH replacement footprints if the primary rearrangement is non-functional; thus, the VH replacement process in all 11 species should be prone to generate CDR3s that are rich in positively charged amino acids if this mechanism is conserved in jawed vertebrates.
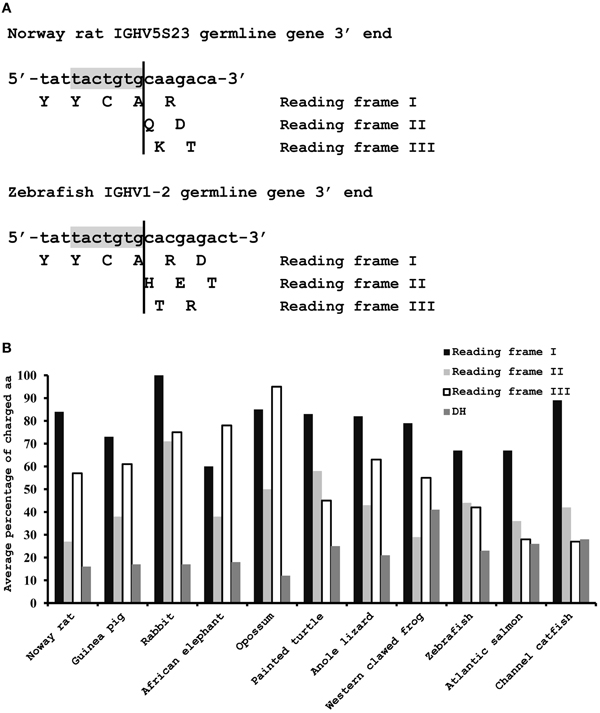
Figure 2. Charged amino acids encoded by the nucleotide sequence following the 3′cRSS of the functional germline VH genes. (A) Two examples of amino acids that are encoded by the nucleotide sequence following the 3′cRSS of the functional germline VH genes in three reading frames. The typical length of the nucleotide sequence following the 3′cRSS is 7 nt in tetrapods (above) and 9 nt in teleosts (below). The 3′cRSSs in the two sequences are shown in gray. (B) Average frequency of the charged amino acids encoded by the nucleotide sequence following the 3′cRSS of the functional germline VH genes in three reading frames and by all functional DH germline genes in 11 species. All functional germline DH sequences are available from the IMGT database (for Norway rat), Ensembl genome database (for painted turtle, genome scaffold JH584564), and other references (Ros et al., 2004; Danilova et al., 2005; Bengten et al., 2006; Zhao et al., 2006; Wang et al., 2009; Wei et al., 2009; Yasuike et al., 2010; Guo et al., 2011, 2012). For each DH gene, the reading frame which encodes the highest percentage of the charged amino acids are chosen to calculate the average percentage of the charged amino acids encoded by DH genes in certain species; thus, the actual average percentage of the charged amino acids encoded by the DH genes in certain species is much lower than the value showed in the figure.
Conclusion
The main conclusion from the present analysis is that both the 3′cRSS and the charged amino acid-encoding nucleotide sequence following the 3′cRSS are conserved among different classes of vertebrates, which suggests that the VH replacement may be a conserved mechanism in all jawed vertebrates. However, additional experimental evidence from species other than human and mouse are needed to support this hypothesis. The biological function of the VH replacement process in non-mammalian vertebrates is worth careful and thorough study.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the National Science Fund for Distinguished Young Scholars (30725029), the National Basic Research Program of China (973 Program-2010CB945300), and the National Natural Science foundation of China (31272433).
References
Bengten, E., Quiniou, S., Hikima, J., Waldbieser, G., Warr, G. W., Miller, N. W., et al. (2006). Structure of the catfish IGH locus: analysis of the region including the single functional IGHM gene. Immunogenetics 58, 831–844.
Cascalho, M., Wong, J., and Wabl, M. (1997). VH gene replacement in hyperselected B cells of the quasimonoclonal mouse. J. Immunol. 159, 5795–5801.
Chen, C., Nagy, Z., Prak, E. L., and Weigert, M. (1995). Immunoglobulin heavy chain gene replacement: a mechanism of receptor editing. Immunity 3, 747–755.
Covey, L. R., Ferrier, P., and Alt, F. W. (1990). VH to VHDJH rearrangement is mediated by the internal VH heptamer. Int. Immunol. 2, 579–583.
Danilova, N., Bussmann, J., Jekosch, K., and Steiner, L. A. (2005). The immunoglobulin heavy-chain locus in zebrafish: identification and expression of a previously unknown isotype, immunoglobulin Z. Nat. Immunol. 6, 295–302.
Davila, M., Liu, F., Cowell, L. G., Lieberman, A. E., Heikamp, E., Patel, A., et al. (2007). Multiple, conserved cryptic recombination signals in VH gene segments: detection of cleavage products only in pro B cells. J. Exp. Med. 204, 3195–3208.
Dooley, H., and Flajnik, M. F. (2006). Antibody repertoire development in cartilaginous fish. Dev. Comp. Immunol. 30, 43–56.
Fan, R. (2009). The Potential Role of VH Replacement in Editing and Generating Autoreactive Antibodies. Ph.D, The University of Alabama.
Fanning, L., Bertrand, F. E., Steinberg, C., and Wu, G. E. (1998). Molecular mechanisms involved in receptor editing at the Ig heavy chain locus. Int. Immunol. 10, 241–246.
Flajnik, M. F. (2002). Comparative analyses of immunoglobulin genes: surprises and portents. Nat. Rev. Immunol. 2, 688–698.
Gambon Deza, F., Sanchez Espinel, C., and Magadan Mompo, S. (2009). The immunoglobulin heavy chain locus in the reptile Anolis carolinensis. Mol. Immunol. 46, 1679–1687.
Gay, D., Saunders, T., Camper, S., and Weigert, M. (1993). Receptor editing: an approach by autoreactive B cells to escape tolerance. J. Exp. Med. 177, 999–1008.
Guo, Y., Bao, Y., Meng, Q., Hu, X., Ren, L., Li, N., et al. (2012). Immunoglobulin genomics in the guinea pig (Cavia porcellus). PLoS ONE 7:e39298. doi: 10.1371/journal.pone.0039298
Guo, Y., Bao, Y., Wang, H., Hu, X., Zhao, Z., Li, N., et al. (2011). A preliminary analysis of the immunoglobulin genes in the African elephant (Loxodonta africana). PLoS ONE 6:e16889. doi: 10.1371/journal.pone.0016889
Itoh, K., Meffre, E., Albesiano, E., Farber, A., Dines, D., Stein, P., et al. (2000). Immunoglobulin heavy chain variable region gene replacement as a mechanism for receptor revision in rheumatoid arthritis synovial tissue B lymphocytes. J. Exp. Med. 192, 1151–1164.
Jung, D., Giallourakis, C., Mostoslavsky, R., and Alt, F. W. (2006). Mechanism and control of V(D)J recombination at the immunoglobulin heavy chain locus. Annu. Rev. Immunol. 24, 541–570.
Kleinfield, R., Hardy, R. R., Tarlinton, D., Dangl, J., Herzenberg, L. A., and Weigert, M. (1986). Recombination between an expressed immunoglobulin heavy-chain gene and a germline variable gene segment in a Ly 1+ B-cell lymphoma. Nature 322, 843–846.
Koralov, S. B., Novobrantseva, T. I., Konigsmann, J., Ehlich, A., and Rajewsky, K. (2006). Antibody repertoires generated by VH replacement and direct VH to JH joining. Immunity 25, 43–53.
Lieber, M. R., Ma, Y., Pannicke, U., and Schwarz, K. (2003). Mechanism and regulation of human non-homologous DNA end-joining. Nat. Rev. Mol. Cell Biol. 4, 712–720.
McCormack, W. T., Tjoelker, L. W., and Thompson, C. B. (1991). Avian B-cell development: generation of an immunoglobulin repertoire by gene conversion. Annu. Rev. Immunol. 9, 219–241.
Nemazee, D. (2006). Receptor editing in lymphocyte development and central tolerance. Nat. Rev. Immunol. 6, 728–740.
Ota, T., and Nei, M. (1994). Divergent evolution and evolution by the birth-and-death process in the immunoglobulin VH gene family. Mol. Biol. Evol. 11, 469–482.
Prak, E. L., and Weigert, M. (1995). Light chain replacement: a new model for antibody gene rearrangement. J. Exp. Med. 182, 541–548.
Radic, M. Z., Mackle, J., Erikson, J., Mol, C., Anderson, W. F., and Weigert, M. (1993). Residues that mediate DNA binding of autoimmune antibodies. J. Immunol. 150, 4966–4977.
Radic, M. Z., and Zouali, M. (1996). Receptor editing, immune diversification, and self-tolerance. Immunity 5, 505–511.
Reth, M., Gehrmann, P., Petrac, E., and Wiese, P. (1986). A novel VH to VHDJH joining mechanism in heavy-chain-negative (null) pre-B cells results in heavy-chain production. Nature 322, 840–842.
Ros, F., Puels, J., Reichenberger, N., Van Schooten, W., Buelow, R., and Platzer, J. (2004). Sequence analysis of 0.5 Mb of the rabbit germline immunoglobulin heavy chain locus. Gene 330, 49–59.
Tiegs, S. L., Russell, D. M., and Nemazee, D. (1993). Receptor editing in self-reactive bone marrow B cells. J. Exp. Med. 177, 1009–1020.
Usuda, S., Takemori, T., Matsuoka, M., Shirasawa, T., Yoshida, K., Mori, A., et al. (1992). Immunoglobulin V gene replacement is caused by the intramolecular DNA deletion mechanism. EMBO J. 11, 611–618.
Wang, X., Olp, J. J., and Miller, R. D. (2009). On the genomics of immunoglobulins in the gray, short-tailed opossum Monodelphis domestica. Immunogenetics 61, 581–596.
Wei, Z., Wu, Q., Ren, L., Hu, X., Guo, Y., Warr, G. W., et al. (2009). Expression of IgM, IgD, and IgY in a reptile, Anolis carolinensis. J. Immunol. 183, 3858–3864.
Wilson, P. C., Wilson, K., Liu, Y. J., Banchereau, J., Pascual, V., and Capra, J. D. (2000). Receptor revision of immunoglobulin heavy chain variable region genes in normal human B lymphocytes. J. Exp. Med. 191, 1881–1894.
Yasuike, M., De Boer, J., Von Schalburg, K. R., Cooper, G. A., McKinnel, L., Messmer, A., et al. (2010). Evolution of duplicated IgH loci in Atlantic salmon, Salmo salar. BMC Genomics 11:486. doi: 10.1186/1471-2164-11-486
Zhang, Z., Burrows, P. D., and Cooper, M. D. (2004). The molecular basis and biological significance of VH replacement. Immunol. Rev. 197, 231–242.
Zhang, Z., Zemlin, M., Wang, Y. H., Munfus, D., Huye, L. E., Findley, H. W., et al. (2003). Contribution of Vh gene replacement to the primary B cell repertoire. Immunity 19, 21–31.
Keywords: immunoglobulin, VH replacement, 3′cRSS, VH replacement footprint, charged amino acid, vertebrate
Citation: Sun Y, Liu Z, Li Z, Lian Z and Zhao Y (2012) Phylogenetic conservation of the 3′ cryptic recombination signal sequence (3′cRSS) in the VH genes of jawed vertebrates. Front. Immun. 3:392. doi: 10.3389/fimmu.2012.00392
Received: 04 September 2012; Accepted: 05 December 2012;
Published online: 20 December 2012.
Edited by:
Zhixin (Jason) Zhang, University of Nebraska Medical Center, USAReviewed by:
Biao Zheng, Baylor College of Medicine and University of Texas Medical School, USAZhixin (Jason) Zhang, University of Nebraska Medical Center, USA
Copyright © 2012 Sun, Liu, Li, Lian and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Yi Sun and Yaofeng Zhao, State Key Laboratory of Agrobiotechnology, College of Biological Sciences, China Agricultural University, No.2 Yuanmingyan west Road, Haidian District, Beijing 100193, China. e-mail:c3VueWlAY2F1LmVkdS5jbg==;eWFvZmVuZ3poYW9AY2F1LmVkdS5jbg==