



- 1Deutsches Rheuma-Forschungszentrum (DRFZ), Berlin, Germany
- 2Department of Rheumatology and Clinical Immunology, Charité – University Medicine Berlin, Berlin, Germany
In the absence of antigen, memory T helper (Th) cells are maintained in a resting state. Recently it has been shown that bone marrow (BM) is a major reservoir of resting memory Th cells. In a given immune response, less than 10% of the activated CD4 T cells are recruited to the pool of resting BM memory Th cells. Here we review recent evidence that CD69 and CD49b control homing of memory Th cell precursors to the BM. During the effector phase of an immune response, about 10% of activated CD4 T cells in the spleen express both CD69 and CD49b, and thus qualify as precursors of resting memory Th cells of BM. Loss or blockade of CD69 and CD49b expression on CD4 T cells impairs the generation of resting memory Th cells in the BM. Moreover, in the absence of BM memory Th cells in CD69-deficient mice, T-cell help for B cells is impaired, confirming the central role of BM memory Th cells in the maintenance of immunological memory.
Introduction
The immune system can memorize previously encountered specific antigens for many years, both by the secretion of specific antibodies (humoral memory) and by long-lived, pre-activated cells (reactive memory). It is still debated whether immunological memory is dependent on persistent antigen (1, 2) and whether it is dependent on long-lived effector cells as opposed to dedicated memory cells (3). Here we define immunological memory as “the maintenance of information in the absence of the original instruction”, i.e., memory cells are maintained in the absence of antigen (4). This definition distinguishes “resting” long-lived memory cells from the long-lived effector cells that develop during infection and persistent immune responses. Recent evidence indeed points to the existence of distinct professional memory lymphocytes. In particular, long-lived “memory” plasma cells have been identified, which mostly reside in the bone marrow (BM) and provide humoral memory (5 –7). Recently, also professional memory T helper (Th) cells have been located in the BM. They represent up to 10% of the Th cells generated in an immune reaction. It has been shown that they rest in terms of proliferation and gene expression (8). This finding provides a fascinating challenge for the understanding of how these memory Th cells are generated, maintained, and reactivated. It also raises questions about their relation to long-lived effector Th cells out of the BM. Here we review recent data about roles of CD49b and CD69 in generation of the precursors of memory Th cells and discuss new concepts on how BM memory Th cells are generated and on the particular role of the BM in this process.
Resting Memory
Memory Th cells, memory B cells and memory plasma cells are in a resting state, i.e., they rest in terms of proliferation (5, 7, 9, 10), while for memory CD8 T cells this is likely, but has not been formally demonstrated so far. Memory B cells are thought to be generated in the germinal centers (GCs), although some studies report that early memory B cells develop outside of GCs (11–13) and that affinity-matured IgM+ memory B cells develop in GCs (14, 15). Memory plasma cells are generated from activated B cells, in a way not fully understood, alternative to memory B cell differentiation (16, 17). While it has been described that memory B cells are maintained in the spleen (18, 19), memory plasma cells are maintained mostly in the BM (5, 6). Memory plasma cells of the BM are maintained in distinct survival niches, organized by CXCL12-expressing reticular stromal cells, but also containing other cells, like eosinophils, providing essential survival factors (10, 20, 21). A survival niche for memory B cells has not been identified so far. Memory plasma cells themselves are immobile. Although they express CXCR4, they do not migrate in response to its ligand CXCL12, while their immediate precursors, the plasmablasts do (22). Thus, it seems reasonable to assume that plasmablasts immigrate into the BM, dock onto CXCL12-expressing stromal cells and there differentiate into memory plasma cells [Figure 1; (7, 23)].
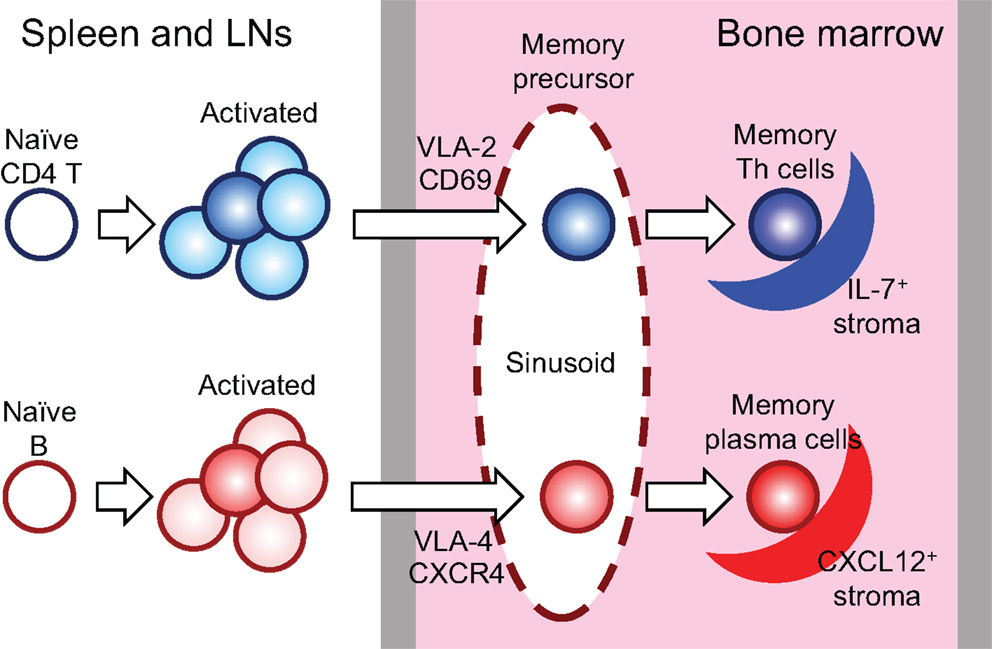
Figure 1. Generation, relocation, and maintenance of memory Th and plasma cells. In an immune response, 10–20% of activated lymphocytes in secondary lymphoid organs relocate into the BM and reside and rest there in the distinct stromal niches. LNs: lymph nodes.
Numerical Control of Resting Th Cell Memory in BM
More than 95% of antigen-specific memory Th cells in the BM individually colocalize with VCAM-1+IL-7+ stromal cells, which comprise about 2% of the total BM cell populations (8). These IL-7-expressing stromal cells probably form a survival niche for resting memory Th cells within the BM. Studies show the competition of newly generated memory Th cells with older ones (24, 25), suggesting the number of niches may be limited; this may determine the overall capacity of the BM for memory Th cells. We compared immune responses to a lymphocytic choriomeningitis virus (LCMV) peptide with responses to a natural LCMV infection and found that they differed substantially in terms of magnitude; these results provide insight as to how a limited number of BM survival niches regulate the size of the memory T-cell pool. In the memory phase of each immune response, the absolute numbers of specific memory Th cells in the BM during each of the responses were comparable (unpublished data). In addition, we detected similar absolute numbers of antigen-specific memory Th cells in the BM in responses to ovalbumin (OVA) plus lipopolysaccharide (LPS) or in response to OVA plus Alum; however, the numbers of effector cells specific for these antigens were different. These numbers games suggest that the BM efficiently restricts access by activated CD4 T cells derived from the pool of memory Th cells via a mechanism that probably involves IL-7-expressing stromal cell, which organizes a niche for exactly one memory Th cell [Figure 1; (23)].
The maintenance of Th cell memory is dependent on IL-7, because long-lived CD4 T cells isolated from the spleen and lymph nodes and then adoptively transferred into IL-7-deficient mice do not survive (26). Persistence of antigen at low levels, i.e., TCR signaling, has been suggested to be required to maintain Th cell memory (1, 2). However, studies using MHC class II-deficient mice (27) and targeted deletion of the TCR (28) show that the maintenance of Th cell memory is independent of TCR-mediated signaling. Professional memory Th cells of BM dock onto IL-7-expressing reticular stromal cells, and there are no MHC class II-expressing cells in their immediate neighborhood (8). Thus it seems likely that the IL-7-expressing stromal cells organize survival niches for memory Th cells of the BM and that TCR stimulation is not required for the maintenance of resting memory Th cells of BM (23). If it is so important for the memory Th cell to be in this dedicated BM niche, central questions for the understanding of immunological memory are: When and how do the T cells get there, and which T cells do get there?
To determine the dynamics of the establishment of professional memory Th cells in the BM, we tracked murine antigen-specific CD4 T cells, generated in intentional immune responses to a defined antigen, into the memory phase of immune responses (8). In immune responses of naive TCR-transgenic CD4 T cells, specific for LCMV or OVA, or wild-type CD4 T cells in response to 4-hydroxy-3-nitrophenylacetyl keyhole limpet hemocyanin (NP-KLH), antigen-specific CD44hi CD4 T cells were present in spleen and lymph nodes on day 4 and 7 (8). Then, the numbers of CD44hi T cells in the secondary lymphoid organs decreased; however, there was an increase in the number of antigen-specific CD44hi CD4 T cells in the BM. From day 30 to 60 post-immunization, more than 80% of the antigen-specific CD4 T cells were in the BM. In the BM they persisted in constant numbers for the rest of the observation period, 7–20 weeks. Beyond day 30 the antigen-specific Th cells are maintained in a resting state in the BM; these cells are not proliferating, having switched off most of their genes. When exactly the cells differentiate from activated cells into resting cells remains to be determined, as well as whether the T cells in BM also rest in terms of mobility.
CD49b/CD69-Dependent Generation of Resting Th Cell Memory
The phenotype of resting memory Th cells of BM is clearly distinct from that of long-lived Th cells in the secondary lymphoid organs and in the blood. They share high expression of CD44, and low expression of CD62L. But only the memory Th cells of BM highly express Ly-6C. They also express CD49b and CD69. While the role of Ly-6C is still enigmatic, it should be noted that humans do not have an orthologous gene. It could be speculated that activated, Ly-6Chi CD4 T cells of the secondary lymphoid organs are precursors of BM memory cells. But whether and if so, how Ly-6C is involved in their homing into the BM or their local maintenance remains unclear.
CD49b, or alpha2 integrin, together with beta1 integrin forms an adhesion molecule VLA-2, which binds mainly to collagens I, II, and XI (29, 30). BM-resident memory Th cells express both, VLA-2 and VLA-4, the alpha2- and the alpha4- beta1-integrin heterodimers. Notably, VLA-4 is expressed also by memory plasma cells, and antibodies to VLA-4 have been shown to eliminate plasma cells from BM (31). For memory Th cells this has not been tested so far. However, the homing of adoptively transferred BM memory Th cells to BM can be efficiently blocked by antibodies to alpha2 integrin (8). This observation alone points to a decisive role of CD49b in the trafficking of memory Th cell precursors to the BM niches for memory Th cells. It should be noted that among the putative ligands of VLA-2 are the collagens II and XI, and collagen XI is exclusively expressed in the BM (32). However, how collagen XI might guide memory Th cell precursors to their desired location remains to be determined.
CD69 is a member of the C-type lectin family and its extracellular ligand is unknown. It is expressed constitutively by more than 60% of the resting memory Th cells of BM (8, 33). This is surprising in the first place, since in general CD69 is considered as an activation marker, expressed by T cells activated in secondary lymphoid organs (34, 35). Jason G. Cyster and his colleagues then showed that CD69 in secondary lymphoid organs essentially acts as a retention signal (36). At first glance, retaining resting memory Th cells in the BM at times when their antigen is not around, would be a reasonable explanation for their expression of CD69. However, things seem to be more complicated. First of all, CD69-deficient mice do not develop a population of BM-resident memory Th cells, in the aftermath of an immune reaction (33). In the effector phase of an immune response, wild-type and CD69 −/− antigen-specific CD4 T cells can be detected readily, in comparable numbers in the secondary lymphoid organs and in the blood. Most notably, follicular helper T cells and GC B cells are present in numbers and phenotypes undistinguishable between wild-type and CD69−/− mice. However, transition of CD69−/− memory Th cell precursors into the BM is impaired. Very few CD69−/− CD4 T cells arrive in the BM and those that do, do not colocalize with IL-7-expressing stromal cells (33).
Differentiation into Resting Memory Cell Precursors
In mice, 10–20% of newly generated plasma cells persist in the BM during a secondary immune response (5, 7, 37, 38). In humans that have been vaccinated against tetanus, the proportion of specific antibodies that persists in the memory phase of the immune response is about 10% of that of the peak response, probably reflecting a corresponding elimination of 90% of the antibody-secreting cells had been generated (39). However, it is unclear how the 10% of memory plasma cell precursors, i.e., plasmablasts that finally make it are selected. In analogy to memory plasma cells, memory Th cells are generated from activated CD4 T cells, i.e., T-cell blasts. Upon immunization with protein antigens, about 10% of the splenic activated CD4 T cells generated at the peak of the response actually mature into BM memory Th cells. Does this mean that 10% of the T-cell blasts make it into the BM memory pool? And how are they qualified? We showed that both CD49b and CD69 are essential for the homing of memory Th cell precursors to the BM (8, 33). On day 6 post-immunization, about 10% of activated CD4 T cells expressed both CD49b and CD69 (unpublished data). It is an intriguing speculation, and hypothesis worthwhile testing, that coexpression of CD69 and CD49b by individual activated Th cells defines these cells as the precursors of resting memory Th cells. In addition, to improve vaccine efficacy, various vaccination strategies should be checked based on the efficiency in inducing CD69 and CD49b coexpressing memory Th cell precursors. Human BM CD4 T cells also preferentially express CD69 compared to peripheral blood CD4 T cells (40). The concept that memory Th cells reside and rest in the BM as observed in mice, may also extend to human Th cell memory and the particular role of the BM in memory maintenance, which should be investigated further.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the Leibniz Association (International Leibniz Research Cluster “ImmunoMemory”).
References
1. Gray D, Matzinger P. T cell memory is short-lived in the absence of antigen. J Exp Med (1991) 174:969–74. doi:10.1084/jem.174.5.969
2. Zinkernagel RM, Hengartner H. Protective ‘immunity’ by pre-existent neutralizing antibody titers and preactivated T cells but not by so-called ‘immunological memory’. Immunol Rev (2006) 211:310–9. doi:10.1111/j.0105-2896.2006.00402.x
3. Bell EB, Westermann J. CD4 memory T cells on trial: immunological memory without a memory T cell. Trends Immunol (2008) 29:405–11. doi:10.1016/j.it.2008.06.002
4. Radbruch A, Rajewsky K. The cellular basis of B cell memory. In: Frederick W, Alt TH, SN Michael editors. The Molecular Biology of B Cells. London: Elsevier Science (2004). p. 247–59.
5. Manz RA, Thiel A, Radbruch A. Lifetime of plasma cells in the bone marrow. Nature (1997) 388:133–4. doi:10.1038/40540
6. Slifka MK, Antia R, Whitmire JK, Ahmed R. Humoral immunity due to long-lived plasma cells. Immunity (1998) 8:363–72. doi:10.1016/S1074-7613(00)80541-5
7. Radbruch A, Muehlinghaus G, Luger EO, Inamine A, Smith KG, Dorner T, et al. Competence and competition: the challenge of becoming a long-lived plasma cell. Nat Rev Immunol (2006) 6:741–50. doi:10.1038/nri1886
8. Tokoyoda K, Zehentmeier S, Hegazy AN, Albrecht I, Grün JR, Löhning M, et al. Professional memory CD4+ T lymphocytes preferentially reside and rest in the bone marrow. Immunity (2009) 30:721–30. doi:10.1016/j.immuni.2009.03.015
9. Tomayko MM, Anderson SM, Brayton CE, Sadanand S, Steinel NC, Behrens TW, et al. Systematic comparison of gene expression between murine memory and naive B cells demonstrates that memory B cells have unique signaling capabilities. J Immunol (2008) 181:27–38.
10. Tokoyoda K, Radbruch A. Signals controlling rest and reactivation of T helper memory lymphocytes in bone marrow. Cell Mol Life Sci (2012) 69:1609–13. doi:10.1007/s00018-012-0969-6
11. Toyama H, Okada S, Hatano M, Takahashi Y, Takeda N, Ichii H, et al. Memory B cells without somatic hypermutation are generated from Bcl6-deficient B cells. Immunity (2002) 17:329–39. doi:10.1016/S1074-7613(02)00387-4
12. Linterman MA, Vinuesa CG. T follicular helper cells during immunity and tolerance. Prog Mol Biol Transl Sci (2010) 92:207–48. doi:10.1016/S1877-1173(10)92009-7
13. Kaji T, Ishige A, Hikida M, Taka J, Hijikata A, Kubo M, et al. Distinct cellular pathways select germline-encoded and somatically mutated antibodies into immunological memory. J Exp Med (2012) 209:2079–97. doi:10.1084/jem.20120127
14. Dogan I, Bertocci B, Vilmont V, Delbos F, Megret J, Storck S, et al. Multiple layers of B cell memory with different effector functions. Nat Immunol (2009) 10:1292–9. doi:10.1038/ni.1814
15. Pape KA, Taylor JJ, Maul RW, Gearhart PJ, Jenkins MK. Different B cell populations mediate early and late memory during an endogenous immune response. Science (2011) 331:1203–7. doi:10.1126/science.1201730
16. Zotos D, Tarlinton DM. Determining germinal centre B cell fate. Trends Immunol (2012) 33:281–8. doi:10.1016/j.it.2012.04.003
17. Shlomchik MJ, Weisel F. Germinal center selection and the development of memory B and plasma cells. Immunol Rev (2012) 247:52–63. doi:10.1111/j.1600-065X.2012.01124.x
18. Mamani-Matsuda MM, Cosma A, Weller S, Faili A, Staib C, Garçon L, et al. The human spleen is a major reservoir for long-lived vaccinia virus-specific memory B cells. Blood (2008) 111:4653–9. doi:10.1182/blood-2007-11-123844
19. Martinez-Gamboa L, Mei H, Loddenkemper C, Ballmer B, Hansen A, Lipsky PE, et al. Role of the spleen in peripheral memory B-cell homeostasis in patients with autoimmune thrombocytopenia purpura. Clin Immunol (2009) 130:199–212. doi:10.1016/j.clim.2008.09.009
20. Yoshida T, Mei H, Dörner T, Hiepe F, Radbruch A, Fillatreau S, et al. Memory B and memory plasma cells. Immunol Rev (2010) 237:117–39. doi:10.1111/j.1600-065X.2010.00938.x
21. Tangye SG. Staying alive: regulation of plasma cell survival. Trends Immunol (2011) 32:595–602. doi:10.1016/j.it.2011.09.001
22. Hauser AE, Debes GF, Arce S, Cassese G, Hamann A, Radbruch A, et al. Chemotactic responsiveness toward ligands for CXCR3 and CXCR4 is regulated on plasma blasts during the time course of a memory immune response. J Immunol (2002) 169:1277–82.
23. Tokoyoda K, Hauser AE, Nakayama T, Radbruch A. Organization of immunological memory by bone marrow stroma. Nat Rev Immunol (2010) 10:193–200. doi:10.1038/nri2727
24. Hataye J, Moon JJ, Khoruts A, Reilly C, Jenkins MK. Naive and memory CD4+ T cell survival controlled by clonal abundance. Science (2006) 312:114–6. doi:10.1126/science.1124228
25. Blair DA, Lefrançois L. Increased competition for antigen during priming negatively impacts the generation of memory CD4 T cells. Proc Natl Acad Sci U S A (2007) 104:15045–50. doi:10.1073/pnas.0703767104
26. Kondrack RM, Harbertson J, Tan JT, McBreen ME, Surh CD, Bradley LM. Interleukin 7 regulates the survival and generation of memory CD4 cells. J Exp Med (2003) 198:1797–806. doi:10.1084/jem.20030735
27. Swain SL, Hu H, Huston G. Class II-independent generation of CD4 memory T cells from effectors. Science (1999) 286:1381–3. doi:10.1126/science.286.5443.1381
28. Polic B, Kunkel D, Scheffold A, Rajewsky K. How alpha beta T cells deal with induced TCR alpha ablation. Proc Natl Acad Sci U S A (2001) 98:8744–9. doi:10.1073/pnas.141218898
29. Tuckwell D, Calderwood DA, Green LJ, Humphries MJ. Integrin alpha 2 I-domain is a binding site for collagens. J Cell Sci (1995) 108(Pt 4):1629–37.
30. Heino J. The collagen family members as cell adhesion proteins. Bioessays (2007) 29:1001–10. doi:10.1002/bies.20636
31. DiLillo DJ, Hamaguchi Y, Ueda Y, Yang K, Uchida J, Haas KM, et al. Maintenance of long-lived plasma cells and serological memory despite mature and memory B cell depletion during CD20 immunotherapy in mice. J Immunol (2008) 180:361–71.
32. Hanazawa A, Tokoyoda K, Hayashizaki K, Shinoda K, Yagita H, Okumura K, et al. CD49b-dependent establishment of T helper cell memory. Immunol Cell Biol (in press).
33. Shinoda K, Tokoyoda K, Hanazawa A, Hayashizaki K, Zehentmeier S, Hosokawa H, et al. Type II membrane protein CD69 regulates the formation of resting T-helper memory. Proc Natl Acad Sci U S A (2012) 109:7409–14. doi:10.1073/pnas.1118539109
34. Testi R, D’Ambrosio D, De Maria R, Santoni A. The CD69 receptor: a multipurpose cell-surface trigger for hematopoietic cells. Immunol Today (1994) 15:479–83. doi:10.1016/0167-5699(94)90193-7
35. Ziegler SF, Ramsdell F, Alderson MR. The activation antigen CD69. Stem Cells (1994) 12:456–65. doi:10.1002/stem.5530120502
36. Shiow LR, Rosen DB, Brdicková N, Xu Y, An J, Lanier LL, et al. CD69 acts downstream of interferon-alpha/beta to inhibit S1P1 and lymphocyte egress from lymphoid organs. Nature (2006) 440:540–4. doi:10.1038/nature04606
37. Manz RA, Löhning M, Cassese G, Thiel A, Radbruch A. Survival of long-lived plasma cells is independent of antigen. Int Immunol (1998) 10:1703–11. doi:10.1093/intimm/10.11.1703
38. Höfer T, Muehlinghaus G, Moser K, Yoshida T, Mei HE, Hebel K, et al. Adaptation of humoral memory. Immunol Rev (2006) 211:295–302. doi:10.1111/j.0105-2896.2006.00380.x
Keywords: T helper cells, immunological memory, bone marrow, CD49b, CD69, homing
Citation: Hanazawa A, Löhning M, Radbruch A and Tokoyoda K (2013) CD49b/CD69-dependent generation of resting T helper cell memory. Front. Immunol. 4:183. doi: 10.3389/fimmu.2013.00183
Received: 06 May 2013; Paper pending published: 29 May 2013;
Accepted: 24 June 2013; Published online: 10 July 2013.
Edited by:
Susan Swain, University of Massachusetts Medical School, USAReviewed by:
Paul Klenerman, University of Oxford, UKKai-Michael Toellner, University of Birmingham, UK
Copyright: © 2013 Hanazawa, Löhning, Radbruch and Tokoyoda. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Koji Tokoyoda, Deutsches Rheuma-Forschungszentrum (DRFZ), Charitéplatz 1, 10117 Berlin, Germany e-mail:dG9rb3lvZGFAZHJmei5kZQ==
†Andreas Radbruch and Koji Tokoyoda have contributed equally to this work.