 Elisabeth R. Krow-Lucal
Elisabeth R. Krow-Lucal Joseph M. McCune
Joseph M. McCune- Division of Experimental Medicine, Department of Medicine, University of California San Francisco, San Francisco, CA, USA
Introduction
Pregnancy poses a challenge to normal mechanisms of immune recognition and rejection: both the mother and her fetus are exposed to allogeneic cells from one to the other. In the case of the mother, these cells are fetal cells carrying paternal antigens; in the case of the fetus, they are maternal cells expressing non-inherited maternal alloantigens (1, 2). Since adaptive immune recognition of these alloantigens could result in mutual rejection and an end to the pregnancy, there are extensive mechanisms in place to inhibit such responses, including poor antigen presentation (3), non-canonical MHC expression, and unique placental and decidual immunomodulatory cell populations (4). The reader is referred to several excellent reviews on this subject (4–7).
Given the inherent difficulties attending experiments in humans, studies of the fetal–maternal interface have focused primarily on inbred strains of laboratory mice. There are, however, major differences between the biology of immune system development of such mice and that found in humans, making it challenging to relate findings in one species to the other. In mice, by example, mature αβ T cells colonize peripheral lymphoid organs during very late gestation and do not fully populate the periphery until after birth (8). By contrast, mature αβ T cells can be found in the periphery of the human fetus as early as 10–12 gestational weeks (5, 9). Thus, early hypotheses posited that in utero tolerance was maintained by a passive or inert fetal immune system (similar to that found in the mouse) (Figure 1A). However, current research suggests that there exist distinct fetal programs both in the T and myeloid compartments that contribute to the unique environment in utero, both in mice and in humans (Figures 1B,C).
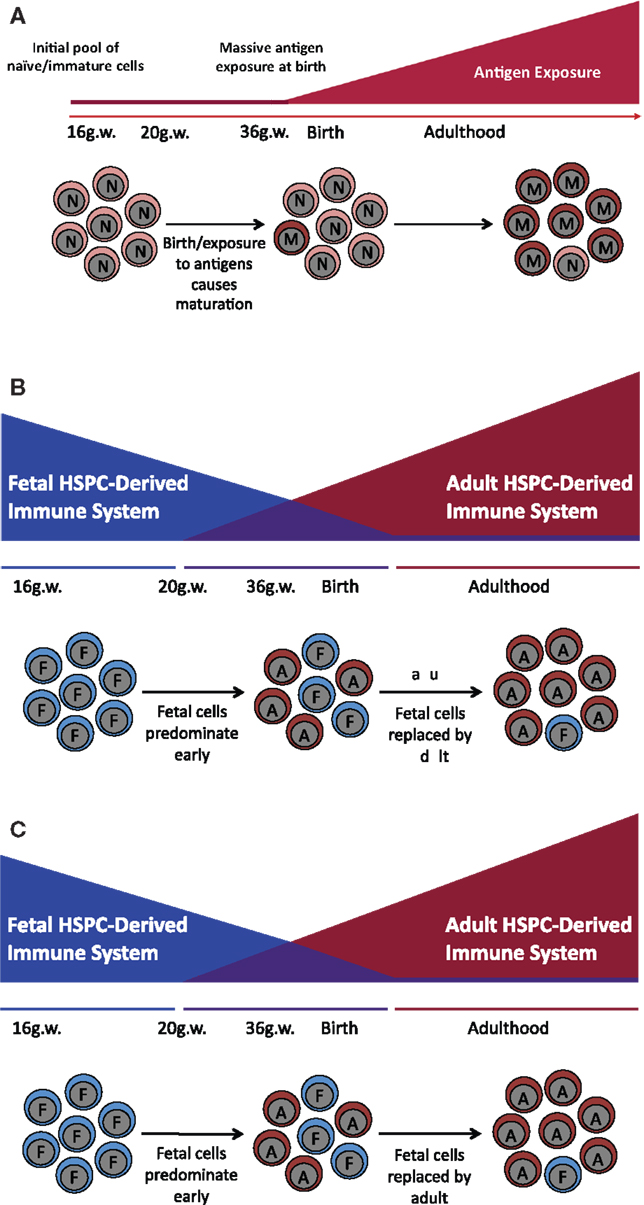
Figure 1. Models of human immune development. Throughout different stages of development, fetal T and myeloid cells, as compared to their adult counterparts, specifically populate a subset of tissues including the epithelium (DETC, non-DETC cells, and Langerhans cells), the brain (microglia), and the heart (cardiac macrophages). This differential population of tissues depending on developmental stage suggests that immune maturation does not proceed in a linear fashion and is rather a specifically timed layering of cells with distinct functions, giving rise to specialized tissue-resident populations. (A) Linear model of immune development. Previous work suggested that the fetal immune system is completely naïve. After birth, it is exposed to massive antigenic doses and matures into the adult immune system. (B) The layered model of immune development. Several types of HSPC appear sequentially and function at specific times during development in a cell autonomous manner, creating unique layers of HSPC-derived cells with different functional outcomes. During gestation, cells are derived from a fetal HSPC and have a specific functional outcome; while in the adult, the vast majority of the cells come from an adult HSPC that has an adult functional outcome. This model suggests that there is a period between 20 gestational weeks and 6 months to a year after birth where there is an admixture of fetal and adult-derived cells, and that it is this admixture of functionally different cell populations that gives rise to inter-individual differences observed in the neonatal immune response. (C) The maturation model of immune development. Fetal cells mature into adult cells such that at birth there are cells with a mixture of both fetal and adult characteristics.
Fetal T Cell Development and Function
Early work in quail chick embryos demonstrated that thymic T cell development occurs in sequential waves, each of which can be identified by differential stem cell colonization of thymic tissue and by unique TCRs (10). These waves appear to be developmentally regulated as they wax and wane according to embryonic gestational age (10), and further work in mice has identified discrete TCR (γδ) utilization during fetal and neonatal development as compared to the adult TCR (αβ) (11–15). Fetal-derived γδ T cells have limited TCR diversity, suggesting a distinct and limited antigen recognition repertoire (12). Furthermore, these cells appear to localize to specific tissues, including the epithelium (16) and the intestine (17). This localization and restricted TCR repertoire suggest that these fetal-derived cells may play a unique role in barrier sites and, as they are developmentally restricted, may be important for promoting tolerance to skin and gut microbiota in early life.
Because of their distinct TCR repertoire and anatomical location, multiple fetal-derived functional populations have been characterized in mice, including dendritic epidermal T cells (DETCs) and non-DETC γδ T cell populations found in the dermis (18, 19). DETCs are the first T cells and seed the epidermis early in development (20). These cells have been implicated in the inhibition of inflammatory skin conditions (21), protection against cutaneous malignancies (22, 23), and wound repair (24, 25). Non-DETC γδ T cell populations have been shown to be the primary producers of IL-17 (18, 19) in the skin and may play a role in response to infection.
These functions may be indicative of a fetal-specific program, ontologically geared toward appropriate development and maintenance of in utero tolerance. Work in humans has demonstrated that while fetal T cells are capable of recognizing and responding to alloantigen in utero (1), these cells preferentially differentiate into T regulatory (Treg) cells, capable of suppressing immune responses (1, 26). Furthermore, these studies show that the fetal T cells are derived from a fetal hematopoietic stem/progenitor cell (HSPC) in the fetal liver and fetal bone marrow, which gives rise to downstream progeny that are distinct from those generated by adult bone marrow-resident HSPC.
Taken together, these data suggest that there are developmentally restricted windows of T cell development in which fetal T cells, functionally distinct from their adult counterparts, arise from discrete HSPC, and seed specific anatomical locations (Figure 1B,C).
Fetal Myeloid Development and Function
The discovery that distinct HSPC give rise to either fetal or adult T cell progeny raises the possibility that there may be differences in other lineages of blood cells as well. Thus, there has been extensive work in human and mouse models elucidating the pathways of regulation of fetal as compared to adult hemoglobin in red blood cells (27–30). Work in mice has meanwhile demonstrated that the presence of fetal-derived B1 cells, with distinct innate-like functions as compared to the adult B2 cells (31–36). Given the multilineage potential of HSPCs, it was also evident that there should be a distinct lineage of fetal- or adult-derived myeloid cells.
In mice, the first hematopoietic progenitors emanate from the extra-embryonic yolk sac and are engaged in primitive hematopoiesis (E7.0–E9.0) (37, 38). “Definitive hematopoiesis” occurs independently in the aorta, gonads, and mesonephros (AGM) region (37, 38). At E10.5, progenitors colonize the fetal liver, the major site of hematopoiesis early in development (38). These waves of hematopoiesis promote egress of various monocyte and macrophage populations, which then give rise to various tissue-resident myeloid populations, including microglia (39), Langerhans cells (LC) (40), and cardiac macrophages (41).
Microglia are the resident macrophage population in the brain and are associated with brain inflammatory diseases. Studies have shown that microglia arise from primitive myeloid progenitors (before E8.0) and are not replaced by circulating monocytes in the adult. Whether or not these fetal-derived cells have distinct functions that differ from an equivalent adult counterpart remains unknown.
Langerhans cells are found in the epidermis of both human and mouse skin. Recent work has demonstrated that LCs are derived from a yolk sac myeloid population during early embryogenesis and then replaced by fetal liver monocytes late in embryogenesis (40). LCs were originally described as pro-inflammatory antigen presenting cells (42); in recent years, however, it has become evident that they can also be involved in – and, indeed, are essential for – the induction of Tregs after infection (43), UV irradiation (44), and glucocorticosteroid stimulation (45). In humans, LCs are able to induce IL-22, but not IL-17, producing T cells (46), potentially suggesting a role in barrier maintenance as opposed to inflammatory processes. LCs also have a limited Toll-like receptor repertoire, including low TLR2, TLR4, and TLR5 expression, leading to attenuated responses to both Gram-positive and Gram-negative bacteria (while leaving viral responses completely intact) (47). Similar to the limited TCR repertoire, this suggests that LCs may be playing a role in tolerization to the skin microbiome.
Recent work highlights the fact that other fetal-derived populations exist in various organs, including lung, liver, spleen, and kidney (41). Furthermore, it has been shown that these populations persist and regenerate in situ, rather than being replaced by the circulating adult monocyte pool (40, 41, 48–50). Thus, these functionally distinct fetal-derived myeloid populations persist into adulthood and can affect immunological outcomes throughout the life of the organism.
Models of Immune Development
Fetal-derived lymphoid and myeloid cells colonize specific anatomical locations and have distinct functions from their adult counterparts. Many of these functions seem tied to barrier integrity and induction of tolerogenic mechanisms. Ontologically, this could be a developmental program designed to allow in utero tolerance to non-inherited maternal alloantigens as well as to promote tolerance to commensal bacteria. These distinct functions, as well as the identification of a fetal HSPC (26), suggest that immune maturation in humans may proceed in a layered fashion (51), with a fetal system that pre-dominates in utero and an adult system that pre-dominates later in life (Figure 1).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Mold JE, Michaëlsson J, Burt TD, Muench MO, Beckerman KP, Busch MP, et al. Maternal alloantigens promote the development of tolerogenic fetal regulatory T cells in utero. Science (2008) 322:1562–5. doi:10.1126/science.1164511
2. Stevens A, Nelson JL. Maternal and fetal microchimerism: implications for human diseases. NeoReviews (2002) 3:e11–9. doi:10.1542/neo.3-1-e11
3. Erlebacher A, Vencato D, Price KA, Zhang D, Glimcher LH. Constraints in antigen presentation severely restrict T cell recognition of the allogeneic fetus. J Clin Invest (2007) 117:1399–411. doi:10.1172/JCI28214
4. Erlebacher A. Immunology of the maternal-fetal interface. Annu Rev Immunol (2013) 31:387–411. doi:10.1146/annurev-immunol-032712-100003
5. Mold JE, McCune JM. Immunological tolerance during fetal development: from mouse to man. Adv Immunol (2012) 115:73–111. doi:10.1016/B978-0-12-394299-9.00003-5
6. Munoz-Suano A, Hamilton AB, Betz AG. Gimme shelter: the immune system during pregnancy. Immunol Rev (2011) 241:20–38. doi:10.1111/j.1600-065X.2011.01002.x
7. Erlebacher A. Mechanisms of T cell tolerance towards the allogeneic fetus. Nat Rev Immunol (2013) 13:23–33. doi:10.1038/nri3361
8. Friedberg SH, Weissman IL. Lymphoid tissue architecture II. Ontogeny of peripheral T and B cells in mice: evidence against Peyer’s patches as the site of generation of B cells. J Immunol (1974) 113:1477–92.
9. Haynes BF, Martin ME, Kay HH, Kurtzberg J. Early events in human T cell ontogeny. Phenotypic characterization and immunohistologic localization of T cell precursors in early human fetal tissues. J Exp Med (1988) 168:1061–80. doi:10.1084/jem.168.3.1061
10. Coltey M, Bucy RP, Chen CH, Cihak J, Lösch U, Char D, et al. Analysis of the first two waves of thymus homing stem cells and their T cell progeny in chick-quail chimeras. J Exp Med (1989) 170:543–57. doi:10.1084/jem.170.2.543
11. Bonyhadi M, Weiss A, Tucker PW, Tigelaar RE, Allison JP. Delta is the Cx-gene product in the γ/δ antigen receptor of dendritic epidermal cells. Nature (1987) 330:574–6. doi:10.1038/330574a0
12. Ito K, Bonneville M, Takagaki Y, Nakanishi N, Kanagawa O, Krecko EG, et al. Different gamma delta T-cell receptors are expressed on thymocytes at different stages of development. Proc Natl Acad Sci U S A (1989) 86:631–5. doi:10.1073/pnas.86.2.631
13. Houlden BA, Cron RQ, Coligan JE, Bluestone JA. Systematic development of distinct T cell receptor-gamma delta T cell subsets during fetal ontogeny. J Immunol (1988) 141:3753–9.
14. Francis Elliott J, Rock EP, Patten PA, Davis MM, Chien Y-H. The adult T-cell receptor 5-chain is diverse and distinct from that of fetal thymocytes. Nature (1988) 331:627–31. doi:10.1038/331627a0
15. Heilig JS, Tonegawa S. Diversity of murine gamma genes and expression in fetal and adult T lymphocytes. Nature (1986) 322:836–40. doi:10.1038/322836a0
16. Leandersson K, Jaensson E, Ivars F. T cells developing in fetal thymus of T-cell receptor alpha-chain transgenic mice colonize gammadelta T-cell-specific epithelial niches but lack long-term reconstituting potential. Immunology (2006) 119:134–42. doi:10.1111/j.1365-2567.2006.02415.x
17. Bonneville M, Janeway CA Jr, Ito K, Haser W, Ishida I, Nakanishi N, et al. Intestinal intraepithelial lymphocytes are a distinct set of γδ T cells. Nature (1988) 336:479–81. doi:10.1038/336479a0
18. Gray EE, Suzuki K, Cyster JG. Cutting edge: identification of a motile IL-17-producing gammadelta T cell population in the dermis. J Immunol (2011) 186:6091–5. doi:10.4049/jimmunol.1100427
19. Sumaria N, Roediger B, Ng LG, Qin J, Pinto R, Cavanagh LL, et al. Cutaneous immunosurveillance by self-renewing dermal gammadelta T cells. J Exp Med (2011) 208:505–18. doi:10.1084/jem.20101824
20. Havran WL, Allison JP. Developmentally ordered appearance of thymocytes expressing different T-cell antigen receptors. Nature (1988) 335:443–5. doi:10.1038/335443a0
21. Girardi M, Lewis J, Glusac E, Filler RB, Geng L, Hayday AC, et al. Resident skin-specific γδ T cells provide local, nonredundant regulation of cutaneous inflammation. J Exp Med (2002) 195:855–67. doi:10.1084/jem.20012000
22. Girardi M, Glusac E, Filler RB, Roberts SJ, Propperova I, Lewis J, et al. The distinct contributions of murine T cell receptor (TCR)gammadelta+ and TCRalphabeta+ T cells to different stages of chemically induced skin cancer. J Exp Med (2003) 198:747–55. doi:10.1084/jem.20021282
23. Girardi M, Oppenheim DE, Steele CR, Lewis JM, Glusac E, Filler R, et al. Regulation of cutaneous malignancy by gammadelta T cells. Science (2001) 294:605–9. doi:10.1126/science.1063916
24. Sharp LL, Jameson JM, Cauvi G, Havran WL. Dendritic epidermal T cells regulate skin homeostasis through local production of insulin-like growth factor 1. Nat Immunol (2005) 6:73–9. doi:10.1038/ni1152
25. Jameson J, Ugarte K, Chen N, Yachi P, Fuchs E, Boismenu R, et al. A role for skin gammadelta T cells in wound repair. Science (2002) 296:747–9. doi:10.1126/science.1069639
26. Mold JE, Venkatasubrahmanyam S, Burt TD, Michaëlsson J, Rivera JM, Galkina SA, et al. Fetal and adult hematopoietic stem cells give rise to distinct T cell lineages in humans. Science (2010) 330:1695–9. doi:10.1126/science.1196509
27. Wilber A, Nienhuis AW, Persons DA. Transcriptional regulation of fetal to adult hemoglobin switching: new therapeutic opportunities. Blood (2011) 117:3945–53. doi:10.1182/blood-2010-11-316893
28. Wilber A, Tschulena U, Hargrove PW, Kim YS, Persons DA, Barbas CF III, et al. A zinc-finger transcriptional activator designed to interact with the gamma-globin gene promoters enhances fetal hemoglobin production in primary human adult erythroblasts. Blood (2010) 115:3033–41. doi:10.1182/blood-2009-08-240556
29. Rochette J, Dodé C, Leturcq F, Krishnamoorthy R. Level and composition of fetal hemoglobin expression in normal newborn babies are not dependent on β cluster DNA haplotype. Am J Hematol (1990) 34:223–4. doi:10.1002/ajh.2830340313
30. Sankaran VG, Xu J, Byron R, Greisman HA, Fisher C, Weatherall DJ, et al. A Functional Element Necessary for Fetal Hemoglobin Silencing. N Engl J Med (2011) 365:807–14. doi:10.1056/NEJMoa1103070
31. Kantor AB. The development and repertoire of B-1 cells (CD5 B cells). Immunol Today (1991) 12:389–91. doi:10.1016/0167-5699(91)90136-H
32. Griffin DO, Holodick NE, Rothstein TL. Human B1 cells in umbilical cord and adult peripheral blood express the novel phenotype CD20+ CD27+ CD43+ CD70-. J Exp Med (2011) 208:67–80. doi:10.1084/jem.20101499
33. Montecino-Rodriguez E, Dorshkind K. B-1 B cell development in the fetus and adult. Immunity (2012) 36:13–21. doi:10.1016/j.immuni.2011.11.017
34. Montecino-Rodriguez E, Dorshkind K. Formation of B-1 B Cells from Neonatal B-1 Transitional Cells Exhibits NF-κB Redundancy. J Immunol (2011) 187:5712–9. doi:10.4049/jimmunol.1102416
35. Hayakawa K, Hardy RR, Herzenberg LA, Herzenberg LA. Progenitors for Ly-1 B cells are distinct from progenitors for other B cells. J Exp Med (1985) 161:1554–68. doi:10.1084/jem.161.6.1554
36. Herzenberg LA, Stall AM. Conventional and Ly-1 B-cell lineages in normal and mu transgenic mice. Cold Spring Harb Symp Quant Biol (1989) 54(Pt 1):219–25. doi:10.1101/SQB.1989.054.01.027
37. Lichanska AM, Hume DA. Origins and functions of phagocytes in the embryo. Exp Hematol (2000) 28:601–11. doi:10.1016/S0301-472X(00)00157-0
38. Orkin SH, Zon LI. Hematopoiesis: an evolving paradigm for stem cell biology. Cell (2008) 132:631–44. doi:10.1016/j.cell.2008.01.025
39. Ginhoux F, Greter M, Leboeuf M, Nandi S, See P, Gokhan S, et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science (2010) 330:841–5. doi:10.1126/science.1194637
40. Hoeffel G, Wang Y, Greter M, See P, Teo P, Malleret B, et al. Adult Langerhans cells derive predominantly from embryonic fetal liver monocytes with a minor contribution of yolk sac-derived macrophages. J Exp Med (2012) 209:1167–81. doi:10.1084/jem.20120340
41. Epelman S, Lavine KJ, Beaudin AE, Sojka DK, Carrero JA, Calderon B, et al. Embryonic and adult-derived resident cardiac macrophages are maintained through distinct mechanisms at steady state and during inflammation. Immunity (2014) 40:91–104. doi:10.1016/j.immuni.2013.11.019
42. Romani N, Brunner PM, Stingl G. Changing views of the role of Langerhans cells. J Invest Dermatol (2012) 132:872–81. doi:10.1038/jid.2011.437
43. Kautz-Neu K, Noordegraaf M, Dinges S, Bennett CL, John D, Clausen BE, et al. Langerhans cells are negative regulators of the anti-Leishmania response. J Exp Med (2011) 208:885–91. doi:10.1084/jem.20102318
44. Obhrai JS, Oberbarnscheidt M, Zhang N, Mueller DL, Shlomchik WD, Lakkis FG, et al. Langerhans cells are not required for efficient skin graft rejection. J Invest Dermatol (2008) 128:1950–5. doi:10.1038/jid.2008.52
45. Stary G, Klein I, Bauer W, Koszik F, Reininger B, Kohlhofer S, et al. Glucocorticosteroids modify Langerhans cells to produce TGF-β and expand regulatory T cells. J Immunol (2011) 186:103–12. doi:10.4049/jimmunol.1002485
46. Fujita H, Nograles KE, Kikuchi T, Gonzalez J, Carucci JA, Krueger JG. Human Langerhans cells induce distinct IL-22-producing CD4+ T cells lacking IL-17 production. Proc Natl Acad Sci U S A (2009) 106:21795–800. doi:10.1073/pnas.0911472106
47. van der Aar AM, Sylva-Steenland RM, Bos JD, Kapsenberg ML, de Jong EC, Teunissen MB. Cutting edge: loss of TLR2, TLR4, and TLR5 on Langerhans cells abolishes bacterial recognition. J Immunol (2007) 178:1986–90. doi:10.4049/jimmunol.178.4.1986
48. Schulz C, Gomez Perdiguero E, Chorro L, Szabo-Rogers H, Cagnard N, Kierdorf K, et al. A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science (2012) 336:86–90. doi:10.1126/science.1219179
49. Yona S, Kim KW, Wolf Y, Mildner A, Varol D, Breker M, et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity (2013) 38:79–91. doi:10.1016/j.immuni.2012.12.001
50. Hashimoto D, Chow A, Noizat C, Teo P, Beasley MB, Leboeuf M, et al. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity (2013) 38:792–804. doi:10.1016/j.immuni.2013.04.004
Keywords: T cell, myeloid cells, human, immune development, immune function
Citation: Krow-Lucal ER and McCune JM (2014) Distinct functional programs in fetal T and myeloid lineages. Front. Immunol. 5:314. doi: 10.3389/fimmu.2014.00314
Received: 05 June 2014; Accepted: 23 June 2014;
Published online: 07 July 2014.
Edited by:
Arnaud Marchant, Université Libre de Bruxelles, BelgiumReviewed by:
Deena Gibbons, Kings College London, UKCopyright: © 2014 Krow-Lucal and McCune. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence:bWlrZS5tY2N1bmVAdWNzZi5lZHU=