 Stéphanie Bessoles†
Stéphanie Bessoles† Camille Grandclément
Camille Grandclément Elisenda Alari-PahissaJasmine GehrigBeena Jeevan-Raj
Elisenda Alari-PahissaJasmine GehrigBeena Jeevan-Raj Werner Held*
Werner Held*- Department of Oncology, Ludwig Center for Cancer Research, University of Lausanne, Lausanne, Switzerland
Natural Killer (NK) cells use germ line encoded receptors to detect diseased host cells. Despite the invariant recognition structures, NK cells have a significant ability to adapt to their surroundings, such as the presence or absence of MHC class I molecules. It has been assumed that this adaptation occurs during NK cell development, but recent findings show that mature NK cells can also adapt to the presence or absence of MHC class I molecules. Here, we summarize how NK cells adjust to changes in the expression of MHC class I molecules. We propose an extension of existing models, in which MHC class I recognition during NK cell development sequentially instructs and maintains NK cell function. The elucidation of the molecular basis of the two effects may identify ways to improve the fitness of NK cells and to prevent the loss of NK cell function due to persistent alterations in their environment.
Development and Function of Conventional NK Cells
Natural killer (NK) cells, which mediate innate immunity to certain pathogens, are able to eliminate malignant cells and regulate innate and adaptive immune responses. These functions depend on a significant number of distinct germ-line encoded receptors. While a few of these receptors are specific for pathogens (1) or for pathogen-derived components expressed on infected host cells, the majority of NK cell receptors are specific for endogenous self-ligands. This type of receptor allows NK cells to respond to host cells whose expression of specific self-ligands deviates from normal.
Conventional NK cells arise from committed progenitors present in the bone marrow. The understanding of the transcriptional network guiding NK cell differentiation has significantly improved recently [for a review, see Ref. (2)]. Expression of the transcription factor Nfil3 (E4BP4) commits common lymphoid progenitors to the NK cell lineage (3). E4BP4 controls the expression of Eomes as well as Id2 (3) and Eomes and T-bet are essential for the development of conventional NK cells downstream of NK committed progenitors (4, 5). Immature NK cells undergo sequential differentiation, expansion, and maturation processes. They acquire a significant number of distinct activation and co-activation receptors, some of which depend on T-bet and Eomes (5). In addition, NK cells show variegated expression of inhibitory receptors, many of which are specific for MHC class I molecules. Furthermore, during their maturation in the bone marrow, NK cells directly acquire an effector program. Bone marrow NK cells constitutively express high levels of Perforin, Granzyme B, and IFNγ mRNAs that are, however, not translated into protein (6, 7). The trigger for the acquisition of this effector program is currently not known, but IL-17 signaling may play a role (8).
The execution of effector functions by peripheral NK cells is controlled at two additional levels. The translation of preformed Perforin and Granzyme B mRNA is dependent on NK cell priming by cytokines such as IL-15 (9). Of note, NK cells in human or wild mice display a primed phenotype, which likely reflects a more frequent exposure to pathogens. Finally, the exocytosis of lytic granules and the production and release of cytokines are tightly controlled by inhibitory and activating NK cell surface receptors. NK cells are stimulated by “non-self,” “stress induced self,” and/or “constitutive self” ligands on host cells. They are inhibited by ubiquitously expressed MHC class I molecules, which are recognized by inhibitory Ly49 (mouse), CD94/NKG2A (mouse human), or KIR family NK cell receptors (human). In general, an excess of activating over inhibitory signals triggers the production and release of effector molecules, which can lead to the death of the infected or transformed host cell (10). “Missing-self” recognition refers to NK cell-mediated responses to cells, which lack inhibitory MHC class I molecules.
Adaptations of NK Cell to Their Environment
Despite the use of invariant germ-line encoded recognition receptors to detect diseased host cells, phenotypes and responses can significantly vary depending on the milieu that surrounds NK cells. Much of the diversity of human NK cells, which was recently estimated at 6,000–30,000 phenotypically distinct NK cell subsets, arises based on environmental variables, rather than genetic cues (11). The persistent exposure of NK cells to cell-bound stress ligands (12, 13) or viral ligands (14, 15) induced the downregulation of the cognate activation receptor and its non-responsiveness to re-stimulation. Dysfunction of activation receptors could be induced in mature NK cells and extended in some cases to activation receptors that had not been triggered [termed cross-tolerance induction (12)]. Removal of the activating ligand resulted in the restoration of receptor function. Similar effects were observed when NK cells were exposed to tumors in vitro and in vivo (12, 16). In addition to reduced function, persistent exposure to activating ligands was also associated with partial or almost complete loss of NK cells (15, 17). Chronic activation may thus affect, in a hierarchical manner, the function of cognate, followed by non-cognate activation receptors and eventually NK cell viability, similar to the exhaustion of CD8 T cells (18). The functional changes may also reflect an adaption of NK cells to their surroundings.
NK Cell Education
Optimal NK cell responses depend on an adaptive process, which adjusts NK cells functionally and phenotypically to the presence of MHC class I molecules. This process is globally referred to as NK cell education. Activation receptors on NK cells, which can recognize MHC class I, respond efficiently to stimulation, while NK cells, which cannot recognize MHC class I, respond poorly (19–22) (Figure 1A). NK cell licensing is used to describe the MHC class I-dependent change in the responsiveness of activation receptors (22). Licensing enables NK cells to respond to occasional aberrant cells that have lost MHC class I molecules (missing-self recognition). NK cell education is also associated with phenotypic changes, which include modifications of the repertoire of inhibitory receptors specific for MHC class I molecules (23, 24). However, these repertoire changes are subtle and their significance is uncertain, especially since they do not ensure that each NK cell can recognize MHC class I. Progress in the mechanistic understanding of NK cell education is hampered by a lack of cell surface markers to identify educated NK cells. Even though it is generally assumed that education is a developmental process, it is not known at which stage of development or maturation NK cells are educated.
KEY CONCEPT 1. NK cell education
Education refers to all phenotypic and functional changes in NK cells imposed by the expression of MHC class I molecules.
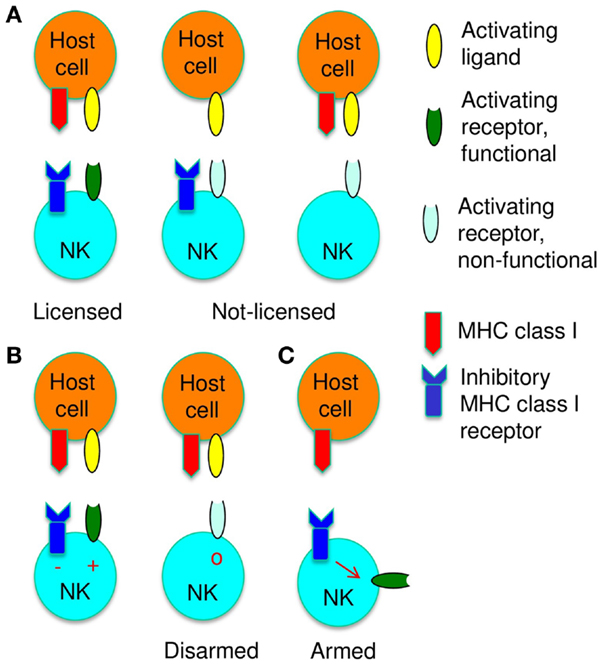
Figure 1. Functional adaptation of NK cells to self-MHC class I is shown. (A) NK cells are licensed, i.e., their activation receptors respond efficiently to stimulation (dark green) when NK cells can bind MHC class I. When the host lacks MHC class I molecules (middle) or when NK cells lack MHC class I receptors (right), activation receptors respond poorly to stimulation (light green), i.e., NK cells are not licensed. (B) Activation pathways are disarmed when NK cells are chronically stimulated in the absence of inhibitory signals from MHC class I receptors. (C) Signals from MHC class I receptors arm NK cells, i.e., instruct NK cell activation pathways to become responsive.
KEY CONCEPT 2. NK cell licensing
Licensing refers to changes in the function of activating NK cell receptors that depend on the recognition of MHC class I molecules by inhibitory NK cell receptors.
Disarming NK Cells
One possible mechanism to explain NK cell education is that activation receptors become by default responsive to stimulation at some point during NK cell development. Acquisition of an inhibitory receptor specific for self-MHC class I would neutralize activation signaling and this would keep the activation pathway competent to respond. If NK cells cannot bind MHC class I, persistent stimulation would eventually disarm all activation pathways (NK cell disarming) (25) (Figure 1B), perhaps similar to the chronic activation described above. However, the relevant receptors, which activate NK cells in response to constitutively expressed self-ligands, are incompletely defined, even though SLAM family receptors explain in part the preferential reactivity of NK cells to normal hematopoietic cells (26).
KEY CONCEPT 3. NK cell disarming
Disarming refers to a possible mechanism to explain licensing: Chronic stimulation of NK cells in the absence of MHC class I-dependent inhibition results in a reduced responsiveness of activating NK cell receptors.
This model is supported by the analysis of mixed bone marrow chimeras (27), MHC class I mosaic mice (28), and the adoptive transfer of NK cells from MHC class I-sufficient into MHC-deficient recipient mice (29). In all cases, NK cell function is dominantly impaired, when a significant fraction of the host cells lacks inhibitory MHC class I ligands. The transfer experiments show that mature NK cells are disarmed when transferred into an MHC class I-deficient host, indicating that disarming is not necessarily a developmentally regulated process.
The defining feature of this model is that the factors needed to inhibit NK cells are at the same time essential for NK cell licensing. MHC class I receptors cannot transduce inhibitory signals when their cytoplasmic ITIM (immune receptor tyrosine-based inhibition motifs) is mutated. Consistent with the above notion, such receptors do not license NK cells (22, 30, 31). Inhibitory signals are further transduced via the recruitment and activation of the Src homology 2 domain-containing phosphatase 1 (SHP-1). While NK cell licensing was dependent on SHP-1 function in some studies (27, 32) it was not necessary in others (22, 33). The conditional deletion of SHP-1 in NK cells will be essential to clarify this issue. Disarming as the unique explanation for NK cell licensing was called into question based on mice expressing a variant of the murine Ly49A receptor, in which the flexible stalk region was replaced with a rigid stalk derived from the CD72 receptor (see below) (30, 31). The variant receptor readily inhibited NK cell-mediated lysis of target cells. However, these NK cells did not mediate missing-self recognition and their NK1.1 activation receptor was not licensed (30, 31). These results suggest that the inhibitory function of an MHC class I receptor is not necessarily sufficient for NK cell licensing.
Arming NK Cells
An alternative mechanism for licensing is that MHC class I recognition instructs NK cell function. Accordingly, NK cell activation receptors would by default respond poorly to stimulation. The engagement of MHC class I receptors would be needed to render activation receptors responsive (NK cell arming) (25)(Figure 1C). Indirect support for this model came from the study of the aforementioned Ly49A variant, which suggested that the inhibitory function of the MHC class I receptor did not suffice to license NK cells (30, 31). The notion that ITIM-bearing Ly49 family member can perform functions in addition to inhibition is supported by the analysis of Ly49Q. This receptor enhanced, rather than inhibited, TLR9-mediated signaling events in plasmacytoid dendritic cells. The positive effect depended on the Ly49Q ITIM, but did not require SHP-1, SHP-2, or SHIP phosphatase activity (34). Thus ITIM-bearing Ly49 receptors can improve cellular activation. Finally, leukemia patients receiving an allogeneic stem cell transplant may have donor-derived NK cells that exert reactivity against residual leukemic cells. This depends on KIR on donor-derived NK cells that can recognize HLA class I of the donor but not of the host (KIR ligand mismatch) (35). These NK cells thus appear to be licensed by the presence of donor HLA class I (36). Similarly, an improved NK cell control-mediated of murine cytomegalovirus infection after allogeneic stem cell transplantation was dependent on the MHC class I of the donor rather than that of the host (37). Based on the disarming model, the absence of MHC class I ligand from a fraction of cells is expected to dominantly induce NK cell dysfunction. The above transplantation settings provided evidence that the presence of donor-type MHC class I molecules instructed NK cell function, consistent with arming.
KEY CONCEPT 4. NK cell arming
Arming provides an alternative mechanistic explanation for licensing. MHC class I recognition instructs NK cells to render their activating receptors responsive to stimulation.
NK Cell Education and the Role of MHC Class I Recognition in trans and cis
Murine Ly49 family receptors have the capacity to interact with MHC class I expressed on other cells (in trans) as well as on the NK cell itself (in cis) (38). Even though a significant number of additional cell surface receptors can bind ligand expressed in the plane of the same membrane (39), there is currently no evidence that KIR or CD94/NKG2A receptors are influenced by MHC class I expression in cis. For Ly49 receptors, combined structural and functional analyses showed that trans and cis interactions are mediated by two distinct conformations of Ly49 receptors (40). The existence of distinct Ly49A conformations on the surface of live cells was confirmed using a FRET-based approach (41). A question arising from these studies was whether cis interaction contributed to NK cell education. The aforementioned Ly49A variant was actually designed to allow trans interaction while preventing cis interaction. This was achieved by replacing the flexible Ly49A stalk with a rigid stalk from the CD72 molecules, thus forcing the ligand binding domains away from the NK cell membrane. As mentioned above, this receptor inhibited the function of cytokine stimulated NK cells but failed to license NK cells. Thus trans interaction was not sufficient to license NK cells, indicating that cis interaction played a role (31). This issue was addressed further using a shortened Ly49A receptor, which could no longer inhibit NK cells (absence of functional trans binding) but which retained the ability to bind in cis. This receptor also failed to license NK cells (30). Thus cis interaction alone was also not sufficient to license NK cells, raising the possibility that both cis and trans binding were required. The same conclusion was drawn based on an independent approach, in which the H-2Dd ligand of the Ly49A receptor was switched off in a cell type specific fashion. The deletion of H-2Dd selectively from NK cells (in cis) prevented NK cell-mediated missing-self recognition. Missing-self recognition was also impaired when H-2Dd was deleted selectively from T cells, i.e., in a situation where trans interactions with a specific cell type was abrogated (30). These data were consistent with the analysis of the modified Ly49A receptors and suggested that both cis and trans recognition of MHC class I molecules played a role for licensing when NK cells developed at steady state.
On the other hand, a role of cis interaction for NK cell licensing was called into question based on adoptive transfer experiments (29, 42) and the induction of MHC class I expression in vivo (43). Following adoptive transfer of MHC class I-deficient NK cells into MHC class I-sufficient mice, NK cells readily acquired improved function. Thus, the expression of endogenous MHC class I genes by NK cells was not needed to license NK cells. However, MHC class I-deficient NK cells are known to acquire significant quantities of MHC class I molecules from surrounding cells via their Ly49 receptors (44, 45). It was thus possible that licensing occurred via cis interactions with the acquired MHC class I molecules (42). When H-2Dd was selectively deleted from NK cells, Ly49A+ NK cells acquired H-2Dd from surrounding cells, however, licensing was not observed (30). Thus trans recognition seems sufficient to improve function following an acute exposure of peripheral NK cells to MHC class I. One possibility to explain the discrepancy to the findings described above may be that the developmental stage determines how NK cells respond to MHC class I encounter. Consistent with this possibility, modifications of the MHC class I receptor repertoire are observed when NK cells develop at steady state (23, 24) but are not evident following the adoptive transfer of MHC class I-deficient NK cells into MHC class I-sufficient hosts (42) or the acute induction of MHC class I expression in vivo using a tetracycline regulated expression system (43).
A Sequential Arming/Disarming Model for NK Cell Education
To account for a role of both cis and trans recognition for NK cell licensing of immature NK cells, we propose a sequential arming/disarming model for NK cell education. This model suggests that cis interactions instruct NK cell activation pathways and that trans interactions provide inhibitory signals, which prevent disarming (Figure 2). If so, the lack of cis and trans recognition should have distinct effects on NK cells. When MHC class I was absent form NK cells (cis), NK cells were not licensed. In contrast, NK cells appeared licensed, i.e., the NK1.1 activation receptor was functional, when MHC class I was absent from T cells (trans). A similar discrepancy between licensing and missing-self reactivity was reported in a CD1 knock out mouse strain. However, the reason for the defect is currently unknown, since it is independent of the targeted mutation (46). While the basis for this novel type of NK cell tolerance needs to be explored, impaired missing-self recognition induced by the absence of MHC class I in cis or from some cells in trans seems to be based on different mechanisms.
KEY CONCEPT 5. Sequential NK cell arming and disarming
We propose that MHC class I recognition during NK cell development first instructs NK cells to render their activating receptors responsive to stimulation (arming) and then prevents the chronic activation of NK cells, which would reduce the responsiveness of activating NK cell receptors (disarming).
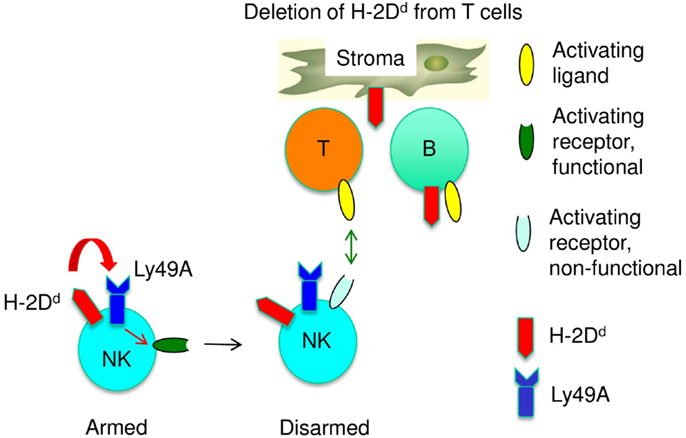
Figure 2. A sequential arming/disarming model for the adaptation of immature NK cells to self-MHC class I is shown. The co-expression of Ly49A and H-2Dd by NK cells, and thus their interaction in cis, renders NK cell activation receptors functional (arming). The absence of H-2Dd from other cells in trans (e.g., T cells) results in chronic NK cell stimulation by T cells, which will eventually disarm activation pathways.
An additional aspect of NK cell education is the modification of the repertoire of MHC class I receptors (23, 24, 47). The expression of a wild type Ly49A transgene in H-2Dd mice is known to induce significant changes in the endogenous MHC class I receptor repertoire. In contrast, repertoire changes were limited when H-2Dd was deleted from NK cells (cis), when Ly49A could only bind in trans and when the Ly49A ITIM was mutated. Conversely, repertoire changes were substantial when MHC class I was deleted from T cells, i.e., when Ly49A could bind in cis but not to certain cells in trans. Thus, the repertoire was modified whenever MHC class I was recognized in cis and when the ITIM was intact. However, when Ly49A could not also productively or continuously interact with MHC class I in trans, NK cell function was not improved. Thus repertoire changes chiefly depended on cis interaction while improved NK cell function required both cis and trans interactions. Accordingly, we propose that cis interaction would instruct NK cell activation receptors to become responsive. As a consequence, NK cells would modify the MHC class I receptor repertoire and trans interactions would be needed to maintain the functionality of NK cell activation pathways. If MHC class I receptors interact only in cis, but cannot not productively (shortened Ly49A variant) or continuously (T cell specific H-2Dd deletion) interact in trans, NK cells are not protected from the adverse effects of chronic stimulation and activation pathways would be disarmed (Figure 2). If MHC class I receptors interact only in trans, NK cell activation receptors are not instructed and the MHC class I receptor repertoire does not adapt.
Concluding Remarks
Natural killer cells functionally adapt to the presence of MHC class I molecules in order to detect cells that lack this key identifier of “normal self.” We propose a model in which MHC class I specific receptors have two roles, i.e., to first establish and then to maintain NK cell function. During NK cell development, Ly49 receptors mediate these two functions via cis and trans interactions, respectively. MHC class I receptors, for which there is no evidence for cis binding, which includes CD94/NKG2A or KIR, may also educate NK cells in a sequential fashion but perhaps via distinct types of signals based on trans interactions. In addition to directly address this issue, it will be important to determine whether the developmental or maturational stage differentially impacts the functional adaptation of NK cells to the presence and absence of MHC class I. Elucidating the molecular mechanisms of how NK cells adapt, may allow the identification of therapeutic targets that prevent NK cells from adapting to their environment, including growing tumors.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported in part by grants from the Swiss National Science Foundation and the Swiss Cancer League (Oncosuisse) (to Werner Held).
Author Biography
Werner Held obtained a Ph.D. in Immunology from the University of Bern. He did postdoctoral training with H. Acha-Orbea and H.R. MacDonald (Lausanne Branch of the Ludwig Institute) and with D.H. Raulet (University of California at Berkeley). After receiving a career development award (START) from the Swiss National Science Foundation he became group head at the Lausanne Branch of the Ludwig Institute. Since 2011, he is a Full Professor at the Ludwig Center for Cancer Research of the University of Lausanne.
References
1. Li SS, Kyei SK, Timm-McCann M, Ogbomo H, Jones GJ, Shi M, et al. The NK receptor NKp30 mediates direct fungal recognition and killing and is diminished in NK cells from HIV-infected patients. Cell Host Microbe (2013) 14:387–97. doi: 10.1016/j.chom.2013.09.007
2. Huntington ND, Nutt SL, Carotta S. Regulation of murine natural killer cell commitment. Front Immunol (2013) 4:14. doi:10.3389/fimmu.2013.00014
3. Male V, Nisoli I, Kostrzewski T, Allan DS, Carlyle JR, Lord GM, et al. The transcription factor E4bp4/Nfil3 controls commitment to the NK lineage and directly regulates Eomes and Id2 expression. J Exp Med (2014) 211:635–42. doi:10.1084/jem.20132398
4. Gascoyne DM, Long E, Veiga-Fernandes H, de Boer J, Williams O, Seddon B, et al. The basic leucine zipper transcription factor E4BP4 is essential for natural killer cell development. Nat Immunol (2009) 10:1118–24. doi:10.1038/ni.1787
5. Gordon SM, Chaix J, Rupp LJ, Wu J, Madera S, Sun JC, et al. The transcription factors T-bet and Eomes control key checkpoints of natural killer cell maturation. Immunity (2012) 36:55–67. doi:10.1016/j.immuni.2011.11.016
6. Fehniger TA, Cai SF, Cao X, Bredemeyer AJ, Presti RM, French AR, et al. Acquisition of murine NK cell cytotoxicity requires the translation of a pre-existing pool of granzyme B and perforin mRNAs. Immunity (2007) 26:798–811. doi:10.1016/j.immuni.2007.04.010
7. Stetson DB, Mohrs M, Reinhardt RL, Baron JL, Wang Z-E, Gapin L, et al. Constitutive cytokine mRNA mark Natural Killer (NK) and NK T cells poised for rapid effector function. J Exp Med (2003) 198:1069–76. doi:10.1084/jem.20030630
8. Bar E, Whitney PG, Moor K, Reis ESC, Leibundgut-Landmann S. IL-17 regulates systemic fungal immunity by controlling the functional competence of NK cells. Immunity (2014) 40:117–27. doi:10.1016/j.immuni.2013.12.002
9. Lucas M, Schachterle W, Oberle K, Aichele P, Diefenbach A. Dendritic cells prime natural killer cells by trans-presenting interleukin 15. Immunity (2007) 26:503–17. doi:10.1016/j.immuni.2007.03.006
10. Long EO, Sik Kim H, Liu D, Peterson ME, Rajagopalan S. Controlling natural killer cell responses: integration of signals for activation and inhibition. Annu Rev Immunol (2013) 31:227–58. doi:10.1146/annurev-immunol-020711-075005
11. Horowitz A, Strauss-Albee DM, Leipold M, Kubo J, Nemat-Gorgani N, Dogan OC, et al. Genetic and environmental determinants of human NK cell diversity revealed by mass cytometry. Sci Transl Med (2013) 5:208ra145. doi:10.1126/scitranslmed.3006702
12. Coudert JD, Scarpellino L, Gros F, Vivier E, Held W. Sustained NKG2D engagement induces cross-tolerance of multiple distinct NK cell activation pathways. Blood (2008) 111:3571–8. doi:10.1182/blood-2007-07-100057
13. Oppenheim DE, Roberts SJ, Clarke SL, Filler R, Lewis JM, Tigelaar RE, et al. Sustained localized expression of ligand for the activating NKG2D receptor impairs natural cytotoxicity in vivo and reduces tumor immunosurveillance. Nat Immunol (2005) 6:928–37. doi:10.1038/ni1239
14. Sun JC, Lanier LL. Tolerance of NK cells encountering their viral ligand during development. J Exp Med (2008) 205:1819–28. doi:10.1084/jem.20072448
15. Tripathy SK, Keyel PA, Yang L, Pingel JT, Cheng TP, Schneeberger A, et al. Continuous engagement of a self-specific activation receptor induces NK cell tolerance. J Exp Med (2008) 205:1829–41. doi:10.1084/jem.20072446
16. Gill S, Vasey AE, De Souza A, Baker J, Smith AT, Kohrt HE, et al. Rapid development of exhaustion and down-regulation of eomesodermin limit the antitumor activity of adoptively transferred murine natural killer cells. Blood (2012) 119:5758–68. doi:10.1182/blood-2012-03-415364
17. De Colvenaer V, Taveirne S, Hamann J, de Bruin AM, De Smedt M, Taghon T, et al. Continuous CD27 triggering in vivo strongly reduces NK cell numbers. Eur J Immunol (2010) 40:1107–17. doi:10.1002/eji.200939251
19. Anfossi N, Andre P, Guia S, Falk CS, Roetynck S, Stewart CA, et al. Human NK cell education by inhibitory receptors for MHC class I. Immunity (2006) 25:331–42. doi:10.1016/j.immuni.2006.06.013
20. Belanger S, Tu MM, Rahim MM, Mahmoud AB, Patel R, Tai LH, et al. Impaired natural killer cell self-education and “missing-self” responses in Ly49-deficient mice. Blood (2012) 120:592–602. doi:10.1182/blood-2012-02-408732
21. Fernandez NC, Treiner E, Vance RE, Jamieson AM, Lemieux S, Raulet DH. A subset of natural killer cells achieve self-tolerance without expressing inhibitory receptors specific for self MHC molecules. Blood (2005) 105:4416–23. doi:10.1182/blood-2004-08-3156
22. Kim S, Poursine-Laurent J, Truscott SM, Lybarger L, Song YJ, Yang L, et al. Licensing of natural killer cells by host major histocompatibility complex class I molecules. Nature (2005) 436:709–13. doi:10.1038/nature03847
23. Sternberg-Simon M, Brodin P, Pickman Y, Onfelt B, Karre K, Malmberg KJ, et al. Natural killer cell inhibitory receptor expression in humans and mice: a closer look. Front Immunol (2013) 4:65. doi:10.3389/fimmu.2013.00065
24. van Bergen J, Thompson A, van Pel M, Retiere C, Salvatori D, Raulet DH, et al. HLA reduces killer cell Ig-like receptor expression level and frequency in a humanized mouse model. J Immunol (2013) 190:2880–5. doi:10.4049/jimmunol.1200650
25. Raulet DH. Missing self recognition self tolerance of natural killer (NK) cells. Semin Immunol (2006) 18:145–50. doi:10.1016/j.smim.2006.03.003
26. Dong Z, Cruz-Munoz ME, Zhong MC, Chen R, Latour S, Veillette A. Essential function for SAP family adaptors in the surveillance of hematopoietic cells by natural killer cells. Nat Immunol (2009) 10:973–80. doi:10.1038/ni.1763
27. Joncker NT, Fernandez NC, Treiner E, Vivier E, Raulet DH. NK cell responsiveness is tuned commensurate with the number of inhibitory receptors for self-MHC class I: the rheostat model. J Immunol (2009) 182:4572–80. doi:10.4049/jimmunol.0803900
28. Johansson MH, Bieberich C, Jay G, Kärre K, Höglund P. Natural killer cell tolerance in mice with mosaic expression of major histocompatibility complex class I molecules. J Exp Med (1997) 186:353–64. doi:10.1084/jem.186.3.353
29. Joncker NT, Shifrin N, Delebecque F, Raulet DH. Mature natural killer cells reset their responsiveness when exposed to an altered MHC environment. J Exp Med (2010) 207:2065–82. doi:10.1084/jem.20100570
30. Bessoles S, Angelov GS, Back J, Leclercq G, Vivier E, Held W. Education of murine NK cells requires both cis and trans recognition of MHC class I molecules. J Immunol (2013) 191:5044–51. doi:10.4049/jimmunol.1301971
31. Chalifour A, Scarpellino L, Back J, Brodin P, Devèvre E, Gros F, et al. A role for cis interaction between the inhibitory Ly49A receptor and MHC class I for NK cell education. Immunity (2009) 30:337–47. doi:10.1016/j.immuni.2008.12.019
32. Lowin-Kropf B, Kunz B, Beermann F, Held W. Impaired natural killing of MHC class I-deficient targets by NK cells expressing a catalytically inactive form of SHP-1. J Immunol (2000) 165:1314–21. doi:10.4049/jimmunol.165.3.1314
33. Orr MT, Murphy WJ, Lanier LL. ‘Unlicensed’ natural killer cells dominate the response to cytomegalovirus infection. Nat Immunol (2010) 11:321–7. doi:10.1038/ni.1849
34. Rahim MM, Tai LH, Troke AD, Mahmoud AB, Abou-Samra E, Roy JG, et al. Ly49Q positively regulates type I IFN production by plasmacytoid dendritic cells in an immunoreceptor tyrosine-based inhibitory motif-dependent manner. J Immunol (2013) 190:3994–4004. doi:10.4049/jimmunol.1200873
35. Ruggeri L, Capanni M, Urbani E, Perruccio K, Shlomchik WD, Tosti A, et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science (2002) 295:2097–100. doi:10.1126/science.1068440
36. Haas P, Loiseau P, Tamouza R, Cayuela JM, Moins-Teisserenc H, Busson M, et al. NK-cell education is shaped by donor HLA genotype after unrelated allogeneic hematopoietic stem cell transplantation. Blood (2011) 117:1021–9. doi:10.1182/blood-2010-02-269381
37. Sungur CM, Tang-Feldman YJ, Zamora AE, Alvarez M, Pomeroy C, Murphy WJ. Murine NK-cell licensing is reflective of donor MHC-I following allogeneic hematopoietic stem cell transplantation in murine cytomegalovirus responses. Blood (2013) 122:1518–21. doi:10.1182/blood-2013-02-483503
38. Held W, Mariuzza RA. Cis interactions of immunoreceptors with MHC and non-MHC ligands. Nat Rev Immunol (2008) 8:269–78. doi:10.1038/nri2278
39. Held W, Mariuzza RA. Cis–trans interactions of cell surface receptors: biological roles and structural basis. Cell Mol Life Sci (2011) 68:3469–78. doi:10.1007/s00018-011-0798-z
40. Back J, Malchiodi EL, Cho S, Scarpellino L, Schneider P, Kerzic MC, et al. Distinct conformations of Ly49 natural killer cell receptors mediate MHC class I recognition in trans and cis. Immunity (2009) 31:598–608. doi:10.1016/j.immuni.2009.07.007
41. Back J, Angelov GS, Mariuzza RA, Held W. The interaction with H-2D(d) in cis is associated with a conformational change in the Ly49A NK cell receptor. Front Immunol (2011) 2:55. doi:10.3389/fimmu.2011.00055
42. Elliott JM, Wahle JA, Yokoyama WM. MHC class I-deficient natural killer cells acquire a licensed phenotype after transfer into an MHC class I-sufficient environment. J Exp Med (2010) 207:2073–9. doi:10.1084/jem.20100986
43. Ebihara T, Jonsson AH, Yokoyama WM. Natural killer cell licensing in mice with inducible expression of MHC class I. Proc Natl Acad Sci U S A (2013) 110:E4232–7. doi:10.1073/pnas.1318255110
44. Sjöström A, Eriksson M, Carboni C, Johansson MH, Sentman CL, Kärre K, et al. Acquisition of external major histocompatibility complex class I molecules by natural killer cells expressing inhibitory Ly49 receptors. J Exp Med (2001) 194:1519–30.
45. Zimmer J, Ioannidis V, Held W. H-2D ligand expression by Ly49A+ Natural Killer (NK) cells precludes ligand uptake from environmental cells: implications for NK cell function. J Exp Med (2001) 194:1531–9. doi:10.1084/jem.194.10.1531
46. Wickstrom SL, Oberg L, Karre K, Johansson MH. A genetic defect in mice that impairs missing self recognition despite evidence for normal maturation and MHC class I-dependent education of NK cells. J Immunol (2014) 192:1577–86. doi:10.4049/jimmunol.1300960
Keywords: NK cells, MHC class I, education, licensing, Ly49 receptors
Citation: Bessoles S, Grandclément C, Alari-Pahissa E, Gehrig J, Jeevan-Raj B and Held W (2014) Adaptations of natural killer cells to self-MHC class I. Front. Immunol. 5:349. doi: 10.3389/fimmu.2014.00349
Received: 07 March 2014; Paper pending published: 04 July 2014;
Accepted: 08 July 2014; Published online: 22 July 2014.
Edited by:
Carsten Watzl, Leibniz Research Centre for Working Environment and Human Factors – IfADo, GermanyReviewed by:
Michael G. Brown, University of Virginia School of Medicine, USADoris Urlaub, Leibniz Research Centre for Working Environment and Human Factors – IfADo, Germany
Copyright: © 2014 Bessoles, Grandclément, Alari-Pahissa, Gehrig, Jeevan-Raj and Held. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence:d2VybmVyLmhlbGRAdW5pbC5jaA==
†Present address: Stéphanie Bessoles, Institut Curie Centre de Recherche INSERM U932, Paris, France