 Amit Kishore
Amit Kishore Veronika Žižková
Veronika Žižková Lenka Kocourková
Lenka Kocourková Martin Petřek
Martin Petřek- 1Department of Pathological Physiology, Laboratory of Immunogenomics, Faculty of Medicine and Dentistry, Palacký University, Olomouc, Czech Republic
- 2Institute of Molecular and Translational Medicine, Faculty of Medicine and Dentistry, Palacký University, Olomouc, Czech Republic
Introduction
Idiopathic pulmonary fibrosis (IPF) is a specific form of chronic, progressive fibrosing interstitial pneumonia with poor diagnosis and a median survival of 2–3 years from initial diagnosis (1, 2). The cellular inflammation drives the fibrotic response in lung and plays a major role in IPF pathogenesis (3). Inflammatory cells (majorly, type 2 alveolar epithelial cells) release TGF-β, the key mediator of pulmonary fibrosis, that regulates several profibrotic cytokines/chemokines, their receptors, receptor subunits, and growth factors inducing process of epithelial–mesenchymal transition (EMT) (3, 4). Among the pro-inflammatory and profibrotic cytokines involved in IPF pathogenesis, interleukin (IL)-1 (4), IL-1β (5), IL-4, IL-5 (6), IL-6Rα (4), angiogenic IL-8/CXCL-8 (7), IL-13, its receptor IL-13 Rα2 (8), and IL-33 (9) have been implicated in accelerated inflammation and irreversible damage to lung architecture with loss of alveolar-capillary barrier basal membrane leading to persistent fibrosis. Genes encoding these factors exhibit nucleotide variation that could affect the severity of immune/inflammatory reactions and extent of any subsequent dysregulated fibroproliferative activity in disease development. Furthermore, variants in mucin-encoding genes (10–13) and in genes for pathogen-associated molecular patterns (PAMPs) receptors of innate immunity known as toll-like receptors (TLRs) (14, 15) have been also implicated in IPF immunopathogenesis and related to rapid progression of the disease. Investigations of these “candidate” gene variant(s) e.g., in case-control association studies may, therefore, provide novel insight into underlying mechanism of IPF susceptibility/disease outcome and, further, may aid to develop novel diagnostic approaches and eventually therapeutic interventions based on genetic information (16).
A candidate gene study typically involves genotyping 5–50 single nucleotide polymorphisms (SNPs) within gene(s) for its coding and non-coding/regulatory regions (17). Irrespective of the number of tested gene variants; for a standard conductance, data collection, and transparent reporting of a genetic association study, the recommendations of STrengthening the REporting of Genetic Association studies (STREGA) and STrengthening the Reporting of OBservational studies in Epidemiology (STROBE) should be considered (16). In case-control studies, knowledge of frequency distribution of candidate gene loci/variants among normal (healthy control) population(s) is necessary and could be useful also for genetically related population(s) to determine the gene variants associated with disease and/or its clinical course.
The role of inflammatory and profibrotic mechanisms involving gene variation has been investigated in IPF and spectrum of susceptible polymorphic gene variants, including those in genes of immune reactions and signaling processes, have been recently reported from both genome-wide association studies (GWAS) and population-based case-control studies (14, 18, 19) performed mostly in US Caucasians, and also in some other ethnicities. The nominated gene variants, summarized in Table 1, are of different functions and implicate some yet-unanticipated pathways in IPF pathogenesis, including endoplasmic reticulum stress and unfolded protein response, cellular senescence, DNA-damage response, and already known Wnt–β-catenin signaling (20). The distribution of SNPs may greatly differ with populations (ethnicities), for example, a high frequency of MUC5B rs35705950*T allele in IPF cases is observed among European-Americans (14–34%) (21, 22), while its low frequency is characteristic for Asians, such as Chinese (3.3%) (23), Japanese (3.4%) (24), and Korean (1.0%) cohorts (11). Similarly, MUC2 rs7934606*A allele exhibit frequency of 41% in Europeans and 1% in Asians (1000 Genomes Project Phase 3 allele frequencies). Furthermore, in context of participation of more than one gene in IPF pathogenesis, it will be important to analyze multiple susceptible gene variants. The approach of analyzing common and rare genetic factors in IPF susceptibility may provide novel insights into IPF and it could also be helpful in identifying population-specific rare variants, predominant panel of candidate gene variants for IPF risk and in understanding the basis of variable disease severity or progression among different populations.
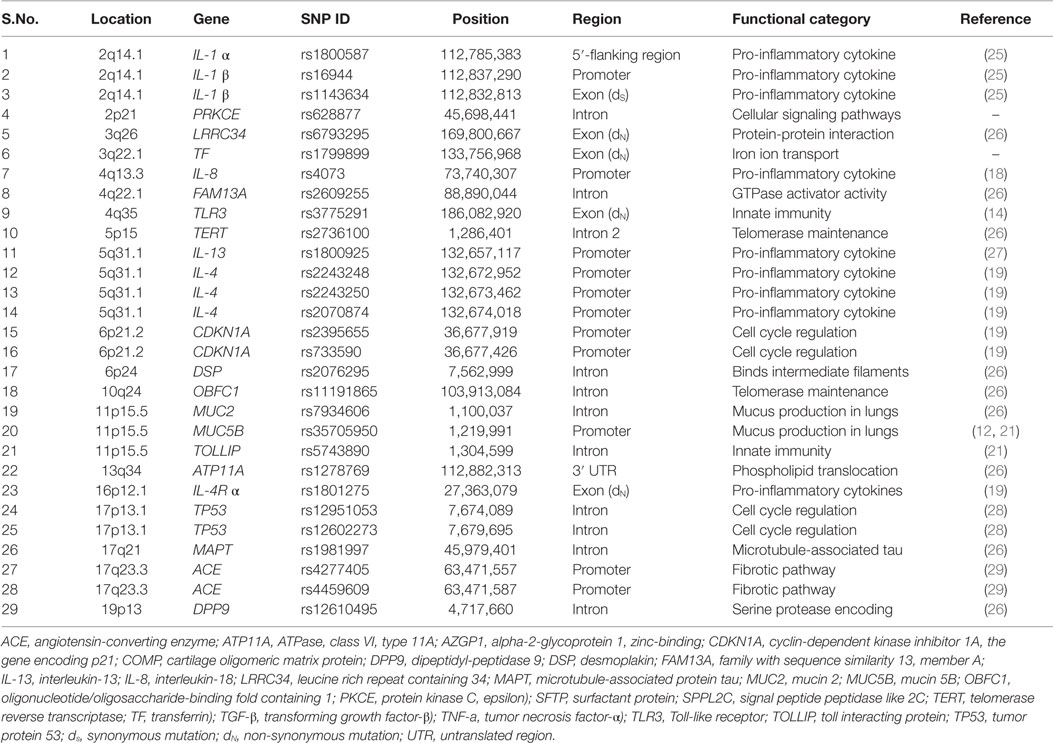
Table 1. List of candidate SNPs investigated in the study.
No complex data have been yet reported on IPF-related variants in Slavonic populations, including Czechs. Starting our investigations of plausible multiple IPF susceptibility polymorphisms primarily in Czech and also related populations, we adopted allele-specific MALDI-TOF mass spectrometry-based SNPs genotyping assay for determination of gene variation in the relevant targets. Several IPF susceptible SNPs in genes of various functional categories were multiplexed, and in the first phase genotyped in probands from normal (healthy) Czech population using Sequenom MassARRAY platform. In the current dataset manuscript, we, besides genotyping methodology, report the genotype, allele, and phenotype (carriage rate) frequencies for plausible IPF susceptibility variants among normal population of Czech Republic of Western Slavonic (Caucasian) ancestry.
Materials and Methods
Characteristics of the Study Group
Ninety-six unrelated healthy subjects (45 males, 51 females), free of any disease as assessed by physician’s enquiry about their personal and family history, were enrolled. The mean age ±standard deviation was 34.5 ± 8.9 years and ranged from 18–57 years. All were Caucasians, and as assessed by surname and tracking personal history of Czech (Western Slavonic) ancestry living in Moravian region of the Czech Republic. All probands were informed about the purpose of the study and provided informed consent; the study was realized with the approval of the institutional ethics committee. Genomic DNA was isolated from peripheral blood leukocytes by standard salting out method (30).
A list of reference single nucleotide polymorphism database ID (rsSNP) was prepared for IPF-associated gene variants identified from available literature (GWAS and case-control studies) (Table 1). SNP genotyping using Sequenom iPLEX and MALDI-TOF-based MassARRAY platform allows analysis of upto 40 SNPs in a single well reaction. Primers were designed using Assay Design Suite v2.0 (ADS v 2.0) available under online tools of Agena Bioscience (https://www.mysequenom.com/Tools).
Assay Design, PCR Amplification, and Genotyping
For online genotyping assay design of multiplexed SNPs using ADS v2.0, an input list of SNP IDs (rsSNP of each target) was provided and following steps were followed: (1) the sequence for each rsSNP was retrieved from database and formatted accordingly, (2) the proximal SNPs for each rsSNP were identified from database, and (3) optimal primer areas were identified that result in a unique amplicons containing a target for the extension primer. To avoid extension primer rejection due to insufficient known bases, the proximal base was replaced with inosine. (4) PCR and extension primers were designed and checked for false priming, hairpin/dimer formation. The primer multiplexes with mass separation of analytes (alleles) were created. The PCR primer plex was prepared for PCR amplification, and extension primer mix was prepared for single base extension (SBE)-based iPLEX reaction. The SBE pool plex consists of multiplexed primers that anneal adjacent to polymorphic site for each reaction present together in the multiplexed assay pool. Thus, several individual DNA polymorphisms with their corresponding SNP sites could be analyzed in a single reaction. Due to the inverse relationship between peak intensity and extend primer mass, the extension primers in iPLEX assays were adjusted for concentration to ensure the possible equal intensity of extension primers.
For multiplexed PCR amplification and genotyping assay, protocol described in literature was followed (31). In brief, multiplexed PCR amplification was performed using 10 ng DNA template. A cleanup reaction for amplicons plex was performed with shrimp alkaline phosphatase (SAP) mix. These SAP cleaned amplicons plex were further subjected to SBE reaction with iPLEX mix containing extend primer plex. The iPLEX reaction products were treated with a cationic resin (SpectroCLEAN, Sequenom) to remove salts, such as Na+, K+, and Mg2+ ions. The desalted iPLEX extend amplicons were dispensed on SpectroCHIPs using MassARRAY® Nanodispenser RS 1000 station and analyzed on matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS)-based MassARRAY analyzer for target allele(s). As assay control of SNP genotyping, a duplicate, positive, and negative control samples were included in each assay plate. In each assay, the specific ddNTP incorporated at target site could be identified with peak representing increase in mass of extend primer. The assay peak spectrum and call cluster plot resulting from MALDI-TOF MS analysis were analyzed with MassARRAY Typer 4.0.20 software that traces primer masses to the assayed alleles.
Statistical Analysis
Each SNP was tested for Hardy–Weinberg Equilibrium (HWE) by Pearson’s goodness-of-fit, Chi-square (χ2) test. SNPs within HWE (p > 0.05) and sufficiently common (Minor Allele Frequency, MAF > 5%) in the general population were included. Phenotype frequency (carriage rate) was calculated as proportion of individuals carrying one or two copies of a particular allele on one or both (maternal or paternal) chromosomes.
Results
Using online Assay Design Suite v2.0 for primer designing, two plexes were generated. Plexes I and II consisted of 22 and 7 SNPs, respectively. For a successful assay, inosine was used instead of proximal SNP in the extend primer sequence for rs2736100, rs2243250, and rs4277405 of plex I.
In assay control of multiplexed SNP genotyping, SNP rs2395655 in CDKN1A showed low assay-success rate (< 95%) and two SNPs DSP rs2076295 and TOLLIP rs5743890 were found as positive in no template control. These SNPs failed the quality control assessments and were removed from further analysis (Table 2). The genotyping assays success rates for all other analyzed SNPs were 98–100%. In our Czech healthy control population, all analyzed SNPs were in HWE, except for IL-4Rα rs1801275 that exhibited minor deviation (p = 0.04) reflecting a small anomaly, so the locus was not excluded from analysis (Table 2).
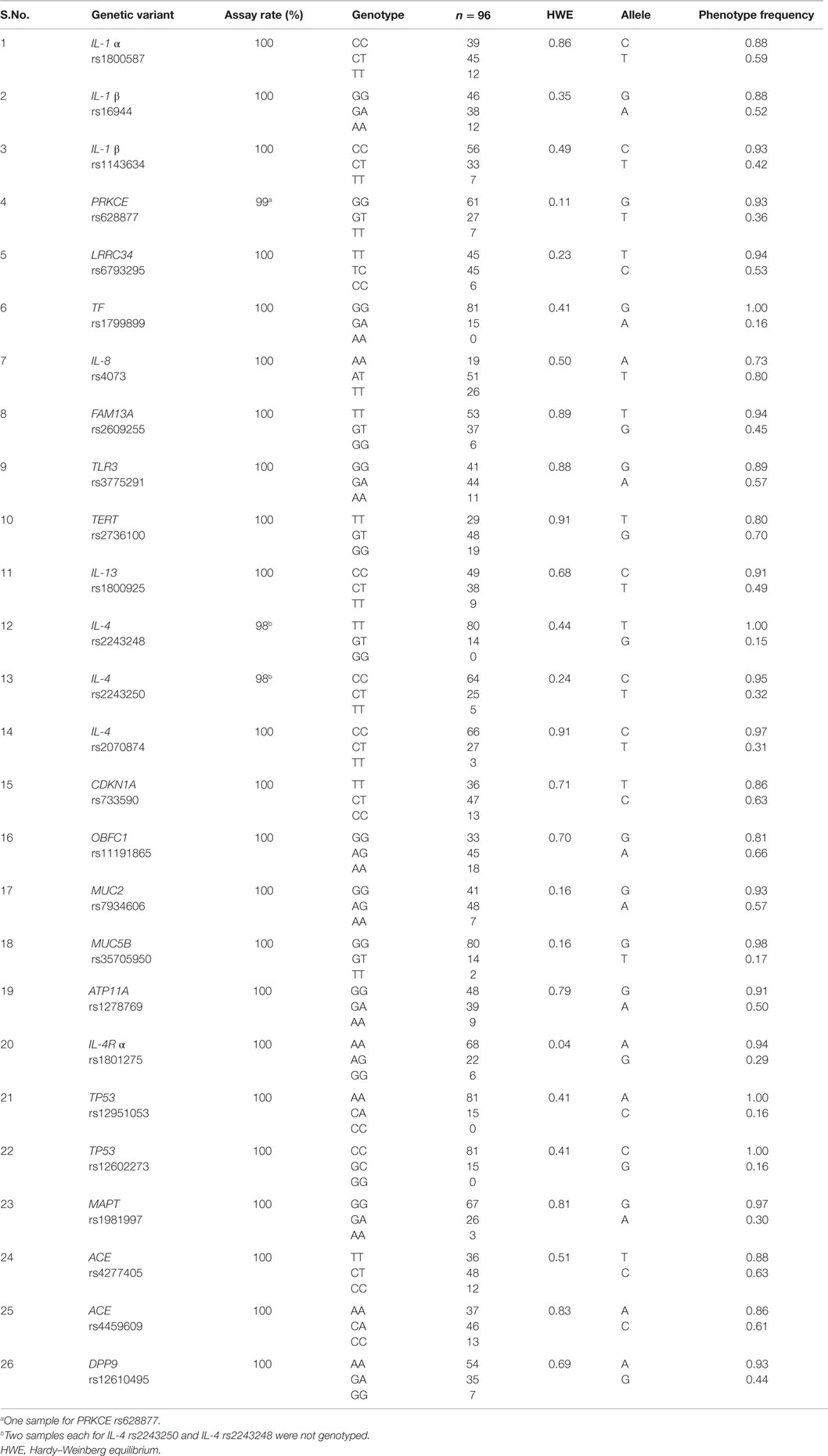
Table 2. Genotype distribution and phenotype frequency of candidate SNPs in normal Czech population.
Among analyzed SNPs in cytokines, their receptor and sub-units, IL-4 rs2243248 exhibited highest genotype (TT = 0.85), allele frequency (T = 0.93) and carriage rate (T = 1.00). Besides cytokines, we also report allelic frequency of rs3775291 in TLR3, an innate immune system receptor. In present report, four out of 26 SNPs, viz., TP53 rs12951053, TP53 rs12602273, TF rs1799899, and IL-4 rs2243248 showed complete absence of their respective homozygous genotype CC, GG, AA, and GG, and exhibited high phenotype frequency (1.00) for allele A, C, G, and T, respectively (Table 2). For MUC5B rs35705950*T risk-allele, allelic and phenotype frequencies were found as 9% and 17%, respectively.
The genotype frequency and allele frequency for the 26 analyzed SNPs are available online at ALlele FREquency Database with Sample UID: SA004336Q (http://alfred.med.yale.edu/alfred/pophetgraph.asp?sampleuid=SA004336Q&cutoff=0.25) and will be publicly available at dbSNP database with the release of dbSNP Build (B144) (http://www.ncbi.nlm.nih.gov/SNP/snp_viewTable.cgi?handle=LIGP).
Discussion
The present dataset reports the genotype distribution, genotype, allele, and phenotype frequency of 26 gene variants involved in immune-related pathomechanisms of IPF in normal Czech population using Sequenom MassARRAY based genotyping platform. Besides the relevance to the delineation of immunogenetic component of IPF, the knowledge of frequency distribution of gene variants in normal populations is of considerable importance for their evaluation as genetic markers in susceptibility, manifestation, prognosis, and potentially treatment of diseases in different populations (32).
A SNP rs35705950 in the putative promoter of MUC5B has been shown to exhibit strong association with both familial interstitial pneumonia and IPF (33). The observed rs35705950*T risk-allele frequency of 9% in normal Czech population was in concordance with other reports in normal Caucasians of European-American descents, as 9–11% in American (33), 10% in UK Caucasians (34), 11% in French (22), and 4.3% in Germans (24) populations among Europeans. Interestingly, the MUC5B promoter polymorphism is observed less frequently in normal Asian populations, such as 0.8% in Japanese (24), 0.7% in Chinese (23), and <1% in Koreans (11). Overall, mucin glycoprotein encoding MUC5B has role in normal lung function by regulating immune function, microbial population, airway infection, and mucociliary clearance in lungs (35, 36).
Among analyzed cytokines, IL-4 has significant role in IPF pathogenesis by regulating fibroblast functions, such as chemotaxis, proliferation, collagen synthesis, myofibroblast differentiation, and Th1/Th2 equilibrium (19). The angiogenic IL-8 was shown as predictive for early stage of IPF (37) and as poor IPF survival (38). Additionally, IL-13 and IL-13 pathway markers (39) and the innate immune signaling receptor TLR3 have been suggested as potential markers of rapidly progressive form of IPF. Several recent studies have suggested that defective TLRs are linked to dysregulated fibrogenesis and have key role in myofibroblast activation, increased profibrotic cytokines, collagen deposition, fibrosis, and tissue destruction and, thus, promoting the progression of disease during the later phase of IPF (14, 15, 40, 41).
Of the four variants that exhibited absence of homozygous genotypes in this data report: (1) the frequency of TP53 rs12951053 CC genotype has been reported as 6% in Caucasian HC (28), 1.2% in European and Africans and relatively higher in Asian (11.9% in Han Chinese and 11.6% in Japanese) populations (http://snp-nexus.org/temp/snpnexus_10220/results.html); (2) For TP53 rs12602273, CC genotype frequency has been reported as 3% in Caucasian healthy controls (28); (3) For TF rs1799899, AA genotype frequency has been reported as 0.6% in European, and 0.0% among African, Han Chinese, and Japanese populations (http://snp-nexus.org/temp/snpnexus_10168/results.html); and (4) For IL-4 rs2243248, low GG genotype frequency (IL-4, -1098 G/T) has been reported in another independent study for characterization of IL-4 gene polymorphism in a relatively small cohort of IPF patients of same ethnicity (19).
The present findings are widely applicable in IPF genetics research in other related populations as well. In a current research initiatives in immunogenetics by HLA-NET network, a working group for population definitions and sampling strategies in population genetics analyses strongly recommend the usage of geographical and/or cultural criteria (with anthropological considerations) to describe human populations instead of a priori misclassifications of racial and ethnic groups (42). In this context, Central Europe populations have been demonstrated as similar and genetically homogeneous (32, 43, 44). Therefore, the present findings are relevant for IPF gene case-control studies not only in Czech but also in neighboring populations, namely Slovak and Polish, and also in Germans and Austrians, as we could recently exemplify in preliminary investigations of immune-related IPF susceptible variants in Czech and German population cohorts (10, 13).
Conclusion
The present data on a spectrum of 26 gene variants including 10 variants of immune and inflammatory response (cytokines/chemokines and TLR) and their frequency distribution in normal Czech (Western Slavonic, Caucasian) population has wider application as standard control along with cases in association studies for IPF. It is also relevant in other fibrotic lung diseases among Czech and genetically related/neighboring population(s) and in the wider context for further delineation of the role of immune and inflammatory reactions in this debilitating disease.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Grant support: CZ.1.07/2.3.00/30.0041, LO1304, and IGA PU LF_2015_020.
References
1. Raghu G, Collard HR, Egan JJ, Martinez FJ, Behr J, Brown KK, et al. An official ATS/ERS/JRS/ALAT statement: idiopathic pulmonary fibrosis: evidence-based guidelines for diagnosis and management. Am J Respir Crit Care Med (2011) 183:788–824. doi: 10.1164/rccm.2009-040GL
2. Spagnolo P, Sverzellati N, Rossi G, Cavazza A, Tzouvelekis A, Crestani B, et al. Idiopathic pulmonary fibrosis: an update. Ann Med (2015) 47:15–27. doi:10.3109/07853890.2014.982165
3. Agostini C, Gurrieri C. Chemokine/cytokine cocktail in idiopathic pulmonary fibrosis. Proc Am Thorac Soc (2006) 3:357–63. doi:10.1513/pats.200601-010TK
4. Zhang J, Tian XJ, Zhang H, Teng Y, Li R, Bai F, et al. TGF-beta-induced epithelial-to-mesenchymal transition proceeds through stepwise activation of multiple feedback loops. Sci Signal (2014) 7:ra91. doi:10.1126/scisignal.2005304
5. Strieter RM, Mehrad B. New mechanisms of pulmonary fibrosis. Chest (2009) 136:1364–70. doi:10.1378/chest.09-0510
6. Furuie H, Yamasaki H, Suga M, Ando M. Altered accessory cell function of alveolar macrophages: a possible mechanism for induction of Th2 secretory profile in idiopathic pulmonary fibrosis. Eur Respir J (1997) 10:787–94.
7. Antoniou KM, Tzouvelekis A, Alexandrakis MG, Sfiridaki K, Tsiligianni I, Rachiotis G, et al. Different angiogenic activity in pulmonary sarcoidosis and idiopathic pulmonary fibrosis. Chest (2006) 130:982–8. doi:10.1378/chest.130.4.982
8. Barnes JC, Lumsden RV, Worrell J, Counihan IP, O’Beirne SL, Belperio JA, et al. CXCR3 is required for the IL-13 mediated upregulation of IL-13Ralpha2 in pulmonary fibroblasts. Am J Respir Cell Mol Biol (2014) 53:217–25. doi:10.1165/rcmb.2013-0433OC
9. Luzina IG, Pickering EM, Kopach P, Kang PH, Lockatell V, Todd NW, et al. Full-length IL-33 promotes inflammation but not Th2 response in vivo in an ST2-independent fashion. J Immunol (2012) 189:403–10. doi:10.4049/jimmunol.1200259
10. Kishore A, Zissel G, Zizkova V, Mueller-Quernheim J, Petrek M. Association of mucin (MUC2, MUC5B) gene variants with idiopathic pulmonary fibrosis (IPF) in a German population: a pilot study using MassARRAY technology. Tissue Antigens (2015) 85:340. doi:10.1111/tan.12557
11. Peljto AL, Selman M, Kim DS, Murphy E, Tucker L, Pardo A, et al. The MUC5B promoter polymorphism is associated with IPF in a Mexican cohort but is rare among Asian ancestries. Chest (2015) 147:460–4. doi:10.1378/chest.14-0867
12. Peljto AL, Zhang Y, Fingerlin TE, Ma SF, Garcia JG, Richards TJ, et al. Association between the MUC5B promoter polymorphism and survival in patients with idiopathic pulmonary fibrosis. JAMA (2013) 309:2232–9. doi:10.1164/ajrccm-conference.2015.191.1_MeetingAbstracts.A4382
13. Petrek M, Kishore A, Zizkova V. Genetic association study for idiopathic pulmonary fibrosis in a Czech population: results from a pilot study using MassARRAY technology. Am J Respir Crit Care Med (2015) 191:A4382. doi:10.1164/ajrccm-conference.2015.191.1_MeetingAbstracts.A4382
14. O’Dwyer DN, Armstrong ME, Trujillo G, Cooke G, Keane MP, Fallon PG, et al. The toll-like receptor 3 L412F polymorphism and disease progression in idiopathic pulmonary fibrosis. Am J Respir Crit Care Med (2013) 188:1442–50. doi:10.1164/rccm.201304-0760OC
15. O’Dwyer DN, Armstrong ME, Kooblall M, Donnelly SC. Targeting defective toll-like receptor-3 function and idiopathic pulmonary fibrosis. Expert Opin Ther Targets (2015) 19:507–14. doi:10.1517/14728222.2014.988706
16. Little J, Higgins JP, Ioannidis JP, Moher D, Gagnon F, von Elm E, et al. Strengthening the reporting of genetic association studies (STREGA): an extension of the STROBE statement. Eur J Epidemiol (2009) 24:37–55. doi:10.1007/s10654-008-9302-y
17. Balding DJ. A tutorial on statistical methods for population association studies. Nat Rev Genet (2006) 7:781–91. doi:10.1038/nrg1916
18. Ahn MH, Park BL, Lee SH, Park SW, Park JS, Kim DJ, et al. A promoter SNP rs4073T>A in the common allele of the interleukin 8 gene is associated with the development of idiopathic pulmonary fibrosis via the IL-8 protein enhancing mode. Respir Res (2011) 12:73. doi:10.1186/1465-9921-12-73
19. Vasakova M, Sterclova M, Matej R, Olejar T, Kolesar L, Skibova J, et al. IL-4 polymorphisms, HRCT score and lung tissue markers in idiopathic pulmonary fibrosis. Hum Immunol (2013) 74:1346–51. doi:10.1016/j.humimm.2013.07.011
20. Kropski JA, Lawson WE, Young LR, Blackwell TS. Genetic studies provide clues on the pathogenesis of idiopathic pulmonary fibrosis. Dis Model Mech (2013) 6:9–17. doi:10.1242/dmm.010736
21. Noth I, Zhang Y, Ma SF, Flores C, Barber M, Huang Y, et al. Genetic variants associated with idiopathic pulmonary fibrosis susceptibility and mortality: a genome-wide association study. Lancet Respir Med (2013) 1:309–17. doi:10.1016/S2213-2600(13)70045-6
22. Borie R, Crestani B, Dieude P, Nunes H, Allanore Y, Kannengiesser C, et al. The MUC5B variant is associated with idiopathic pulmonary fibrosis but not with systemic sclerosis interstitial lung disease in the European Caucasian population. PLoS One (2013) 8:e70621. doi:10.1371/journal.pone.0070621
23. Wang C, Zhuang Y, Guo W, Cao L, Zhang H, Xu L, et al. Mucin 5B promoter polymorphism is associated with susceptibility to interstitial lung diseases in Chinese males. PLoS One (2014) 9:e104919. doi:10.1371/journal.pone.0104919
24. Horimasu Y, Ohshimo S, Bonella F, Tanaka S, Ishikawa N, Hattori N, et al. MUC5B promoter polymorphism in Japanese patients with idiopathic pulmonary fibrosis. Respirology (2015) 20:439–44. doi:10.1111/resp.12466
25. Hutyrova B, Pantelidis P, Drabek J, Zurkova M, Kolek V, Lenhart K, et al. Interleukin-1 gene cluster polymorphisms in sarcoidosis and idiopathic pulmonary fibrosis. Am J Respir Crit Care Med (2002) 165:148–51. doi:10.1164/ajrccm.165.2.2106004
26. Fingerlin TE, Murphy E, Zhang W, Peljto AL, Brown KK, Steele MP, et al. Genome-wide association study identifies multiple susceptibility loci for pulmonary fibrosis. Nat Genet (2013) 45:613–20. doi:10.1038/ng.2609
27. Ding M, Sheng H, Shen W, Zhen J, Xi L, Zeng J, et al. Association of the SNP rs1800925(C/T) in the interleukin-13 gene promoter with pulmonary function in Chinese Han patients with idiopathic pulmonary fibrosis. Cell Biochem Biophys (2013) 67:905–9. doi:10.1007/s12013-013-9580-1
28. Korthagen NM, van Moorsel CH, Barlo NP, Kazemier KM, Ruven HJ, Grutters JC. Association between variations in cell cycle genes and idiopathic pulmonary fibrosis. PLoS One (2012) 7:e30442. doi:10.1371/journal.pone.0030442
29. Uh ST, Kim TH, Shim EY, Jang AS, Park SW, Park JS, et al. Angiotensin-converting enzyme (ACE) gene polymorphisms are associated with idiopathic pulmonary fibrosis. Lung (2013) 191:345–51. doi:10.1007/s00408-013-9469-1
30. Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res (1988) 16:1215. doi:10.1093/nar/16.3.1215
31. Bradic M, Costa J, Chelo IM. Genotyping with Sequenom. Methods Mol Biol (2011) 772:193–210. doi:10.1007/978-1-61779-228-1_11
32. Kubistova Z, Mrazek F, Tudos Z, Kriegova E, Ambruzova Z, Mytilineos J, et al. Distribution of 22 cytokine gene polymorphisms in the healthy Czech population. Int J Immunogenet (2006) 33:261–7. doi:10.1111/j.1744-313X.2006.00609.x
33. Seibold MA, Wise AL, Speer MC, Steele MP, Brown KK, Loyd JE, et al. A common MUC5B promoter polymorphism and pulmonary fibrosis. N Engl J Med (2011) 364:1503–12. doi:10.1056/NEJMoa1013660
34. Stock CJ, Sato H, Fonseca C, Banya WA, Molyneaux PL, Adamali H, et al. Mucin 5B promoter polymorphism is associated with idiopathic pulmonary fibrosis but not with development of lung fibrosis in systemic sclerosis or sarcoidosis. Thorax (2013) 68:436–41. doi:10.1136/thoraxjnl-2012-201786
35. Roy MG, Livraghi-Butrico A, Fletcher AA, McElwee MM, Evans SE, Boerner RM, et al. Muc5b is required for airway defence. Nature (2014) 505:412–6. doi:10.1038/nature12807
36. Fletcher AA, Evans CM. Regulation of microbial populations and immune functions by Muc5b. Ann Am Thorac Soc (2014) 11:S80–S. doi:10.1513/AnnalsATS.201308-263MG
37. Sterclova M, Vasakova M, Metlicka M, Pavlicek J, Kolesar L, Striz I. Angiostatic versus angiogenic chemokines in IPF and EAA. Respir Med (2009) 103:1651–6. doi:10.1016/j.rmed.2009.05.012
38. Richards TJ, Kaminski N, Baribaud F, Flavin S, Brodmerkel C, Horowitz D, et al. Peripheral blood proteins predict mortality in idiopathic pulmonary fibrosis. Am J Respir Crit Care Med (2012) 185:67–76. doi:10.1164/rccm.201101-0058OC
39. Murray LA, Zhang H, Oak SR, Coelho AL, Herath A, Flaherty KR, et al. Targeting interleukin-13 with tralokinumab attenuates lung fibrosis and epithelial damage in a humanized SCID idiopathic pulmonary fibrosis model. Am J Respir Cell Mol Biol (2014) 50:985–94. doi:10.1165/rcmb.2013-0342OC
40. Hogaboam CM, Trujillo G, Martinez FJ. Aberrant innate immune sensing leads to the rapid progression of idiopathic pulmonary fibrosis. Fibrogenesis Tissue Repair (2012) 5:S3. doi:10.1186/1755-1536-5-S1-S3
41. Trujillo G, Meneghin A, Flaherty KR, Sholl LM, Myers JL, Kazerooni EA, et al. TLR9 differentiates rapidly from slowly progressing forms of idiopathic pulmonary fibrosis. Sci Transl Med (2010) 2:57ra82. doi:10.1126/scitranslmed.3001510
42. Sanchez-Mazas A, Vidan-Jeras B, Nunes JM, Fischer G, Little AM, Bekmane U, et al. Strategies to work with HLA data in human populations for histocompatibility, clinical transplantation, epidemiology and population genetics: HLA-NET methodological recommendations. Int J Immunogenet (2012) 39:459–72. doi:10.1111/j.1744-313X.2012.01113.x
43. Lundmark PE, Liljedahl U, Boomsma DI, Mannila H, Martin NG, Palotie A, et al. Evaluation of HapMap data in six populations of European descent. Eur J Hum Genet (2008) 16:1142–50. doi:10.1038/ejhg.2008.77
Keywords: MUC5B, cytokines, single nucleotide polymorphism, susceptibility, sequenom MassARRAY, idiopathic pulmonary fibrosis, Czech normal population, association study
Citation: Kishore A, Žižková V, Kocourková L and Petřek M (2015) A dataset of 26 candidate gene and pro-inflammatory cytokine variants for association studies in idiopathic pulmonary fibrosis: frequency distribution in normal Czech population. Front. Immunol. 6:476. doi: 10.3389/fimmu.2015.00476
Received: 14 August 2015; Accepted: 02 September 2015;
Published: 22 September 2015
Edited by:
Masaaki Murakami, Hokkaido University, JapanReviewed by:
Takayuki Yoshimoto, Tokyo Medical University, JapanDaisuke Kamimura, Hokkaido University, Japan
Copyright: © 2015 Kishore, Žižková, Kocourková and Petřek. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martin Petřek, Department of Pathological Physiology, Faculty of Medicine and Dentistry, Palacky University, Hněvotínská Street 3, Olomouc 77515, Czech Republic,bWFydGluLnBldHJla0Bmbm9sLmN6