 Christophe Moreau1,2,3†
Christophe Moreau1,2,3† Isabelle Bally1,2,3†
Isabelle Bally1,2,3† Anne Chouquet1,2,3
Anne Chouquet1,2,3 Barbara Bottazzi4
Barbara Bottazzi4 Berhane Ghebrehiwet5
Berhane Ghebrehiwet5 Christine Gaboriaud1,2,3*
Christine Gaboriaud1,2,3* Nicole Thielens1,2,3*
Nicole Thielens1,2,3*
- 1IBS, University of Grenoble Alpes, Grenoble, France
- 2CNRS, IBS, Grenoble, France
- 3IBS, CEA, Grenoble, France
- 4Immunopharmacology Laboratory, Humanitas Research Hospital, Rozzano, Italy
- 5Department of Medicine, Stony Brook University, Stony Brook, NY, USA
Complement C1q is a soluble pattern recognition molecule comprising six heterotrimeric subunits assembled from three polypeptide chains (A–C). Each heterotrimer forms a collagen-like stem prolonged by a globular recognition domain. These recognition domains sense a wide variety of ligands, including pathogens and altered-self components. Ligand recognition is either direct or mediated by immunoglobulins or pentraxins. Multivalent binding of C1q to its targets triggers immune effector mechanisms mediated via its collagen-like stems. The induced immune response includes activation of the classical complement pathway and enhancement of the phagocytosis of the recognized target. We report here, the first production of a single-chain recombinant form of human C1q globular region (C1q-scGR). The three monomers have been linked in tandem to generate a single continuous polypeptide, based on a strategy previously used for adiponectin, a protein structurally related to C1q. The resulting C1q-scGR protein was produced at high yield in stably transfected 293-F mammalian cells. Recombinant C1q-scGR was correctly folded, as demonstrated by its X-ray crystal structure solved at a resolution of 1.35 Å. Its interaction properties were assessed by surface plasmon resonance analysis using the following physiological C1q ligands: the receptor for C1q globular heads, the long pentraxin PTX3, calreticulin, and heparin. The 3D structure and the binding properties of C1q-scGR were similar to those of the three-chain fragment generated by collagenase digestion of serum-derived C1q. Comparison of the interaction properties of the fragments with those of native C1q provided insights into the avidity component associated with the hexameric assembly of C1q. The interest of this functional recombinant form of the recognition domains of C1q in basic research and its potential biomedical applications are discussed.
Introduction
The soluble defense collagens are oligomeric innate immune pattern recognition receptors (PRRs), which are composed of N-terminal collagen-like stems prolonged by C-terminal globular trimeric pattern recognition domains [reviewed in Ref. (1)]. According to the nature of their recognition domain, these PRRs can be divided into three families in human, namely, proteins with gC1q domains (C1q and adiponectin), C-type lectin carbohydrate recognition domains (lung surfactant proteins A and D, mannan-binding lectin (MBL), collectins kidney 1 and liver 1), and fibrinogen-like domains (ficolins). Following target recognition, C1q and collectins (except surfactant proteins) or ficolins have the capacity to trigger activation of the classical and lectin pathways of complement for microbial killing and phagocytosis, through proteases associated with their collagen stems.
The C1q molecule is a complex defense collagen, being assembled from six heterotrimeric subunits, each consisting of three homologous, yet distinct polypeptide chains (A–C) encoded by the C1QA, C1QB, and C1QC genes oriented in the A–C–B order on human chromosome 1p (2). C1q also features the most versatile recognition properties, being able to identify not only bacterial and viral pathogens, either directly or through other immune proteins such as antibodies and pentraxins, but also many altered self elements, including β-amyloid fibrils (3), the pathological form of the prion protein (4, 5), modified low-density lipoproteins (6), and apoptotic cells (7–9).
Production of the C1q globular region (C1q-GR) by limited proteolysis of the serum-derived protein with collagenase allowed resolution of its X-ray crystal structure. The resulting compact heterotrimeric structure revealed differences in the surface charges of the subunits, a key factor for the versatility of C1q binding properties (10, 11). A further step toward understanding C1q binding properties was accomplished with the production of recombinant forms of the individual gC1q domains fused to maltose-binding protein, which revealed that these domains are functionally autonomous modules with differential ligand-binding properties (12). Site-directed mutagenesis studies provided information about the residues involved in the interaction of C1q with some of its ligands (13–16). However, elucidation of the C1q recognition properties in the more physiological context of the heterotrimeric globular regions still awaits the availability of the corresponding recombinant fragment.
We report here, the production of a single-chain recombinant form of human C1q globular region (C1q-scGR). The three monomers have been linked in tandem to generate a single continuous polypeptide, based on a strategy previously used to generate a single-chain form of the homotrimeric globular domain of adiponectin, a protein structurally related to C1q (17). The C1q-scGR recombinant protein was produced at high yield in stably transfected mammalian cells. Its physicochemical, structural, and functional analysis shows that it is correctly folded and retains the ability to associate with physiological C1q ligands, including the long pentraxin PTX3, the receptor for the globular heads of C1q (gC1qR), calreticulin (CRT), and heparin. The interest of this fragment in basic research and its potential biomedical applications will be discussed.
Materials and Methods
Proteins and Reagents
C1q was purified from human serum and quantified, as described previously (18). The globular regions of C1q were prepared by collagenase digestion of C1q, as described previously (3), and their molar concentration estimated using a Mw value of 48,000 and an absorption coefficient (A1%, 1 cm) at 280 nm of 0.93. Recombinant human PTX3, gC1qR, and CRT were produced, as described previously (19–21). Streptavidin and heparin-biotin sodium salt (Mw 15 kDa) were procured from Sigma-Aldrich. Oligonucleotides were purchased from Eurogentec. Restriction and modification enzymes were from New England Biolabs.
Cloning of the Single-Chain Globular Domain of Human C1q
For recombinant protein expression in the baculovirus/insect cells system, a synthetic cDNA encoding residues 85–223 of mature C1qA, a Gly–Ser–Gly linker, residues 87–217 of mature C1qC (gC1qC), a Gly–Ser–Ala linker, and residues 90–226 of mature C1qB (gC1qB), cloned in frame with the melittin signal peptide of the pNT-Bac vector (22) (pNT-Bac–C1q-scGR), was purchased from GeneCust.
For expression in mammalian 293-F cells, an intermediate construct was generated from the pcDNA3.1–C1qA vector (23) by removing residues 1–87 of mature C1qA by site-directed mutagenesis, allowing in frame cloning of residues 88–223 of C1qA with the native signal peptide of C1qA (pcDNA3.1–gC1qA). A DNA fragment encoding the Gly–Ser–Gly linker, gC1qC, the Gly–Ser–Ala linker, and gC1qB was amplified using VentR polymerase and pNT-Bac–C1q-scGR as a template and inserted into pcDNA3.1–gC1qA by site-directed mutagenesis. The resulting construct (called pcDNA3.1–C1q-scGR) was characterized by restriction mapping and checked by double-stranded DNA sequencing (GATC Biotech).
Production of C1q-scGR in Eukaryotic Cells and Protein Purification
Generation of a recombinant baculovirus from the pNT-Bac–C1q-scGR plasmid using the Bac-to-Bac system (Invitrogen) and infection of Trichoplusia ni (High Five) insect cells was performed, as described previously (24). Stably transfected cells producing C1q-scGR were obtained by transfection of FreeStyle 293-F cells with the pcDNA3.1–C1q-scGR plasmid using 293-fectin and subsequent selection with 400 μg/ml G418 as recommended by the manufacturer (Invitrogen). The cells were expanded in the Freestyle expression medium (Invitrogen) and the culture supernatant harvested and replaced every 72 h up to three times.
The insect and mammalian cell culture supernatants containing C1q-scGR (500 ml) were dialyzed against 50 mM MES, 25 mM NaCl, pH 6.4, and loaded at 1.5 ml/min onto a SP Sepharose Fast Flow column (GE Healthcare) (50 ml) equilibrated in the same buffer. Elution was carried out by applying a 1-l linear gradient from 25 to 500 mM NaCl in the same buffer. The fractions containing the recombinant protein were identified by SDS-PAGE analysis, pooled, dialyzed against 50 mM Tris–HCl, 150 mM NaCl, pH 7.4, and concentrated to 1–5 mg/ml by ultrafiltration on a PM-10 membrane (Amicon). The molar concentration of C1q-scGR was estimated using an absorption coefficient (A1%, 1 cm) at 280 nm of 0.93 and a Mr value of 47,534, as determined by mass spectrometry.
SDS-PAGE, N-Terminal Sequence, and Liquid Chromatography–Electrospray Ionization–Time-of-Flight Mass Spectrometry Analyses
Recombinant C1q-scGR was analyzed by SDS-PAGE under non-reducing or reducing conditions using Tris–HCl gels containing 10% polyacrylamide. N-terminal sequence determination was performed using an Applied Biosystems gas-phase sequencer model 492 coupled online with an Applied Biosystems Model 140C HPLC system. Liquid chromatography–electrospray ionization–time-of-flight (LC–ESI–TOF) mass spectrometry analyses of purified C1q-scGR, before and after treatment with Clostridium perfringens type X neuraminidase (Sigma) (0.3 U/mg) for 5 h at 25°C, were performed using a 6210 LC–TOF mass spectrometer interfaced with LC pump system (Agilent Technologies). Samples were desalted on-line on a protein trap (Zorbax 300SB-C8, 5 μm, 5 mm × 0.3 mm, Agilent Technologies) before analysis.
Analytical Ultracentrifugation
Sedimentation velocity analysis was performed using a Beckman XL-I analytical ultracentrifuge and an AN-50 TI rotor (Beckman Coulter, Palo Alto, CA, USA). Three C1q-scGR samples at 0.9, 2.5, and 5 mg/ml were loaded into 12, 3, and 1.5 mm pathlength double-sector cells and centrifuged at 42,000 rpm at 6°C in 50 mM Tris–HCl, 150 mM NaCl, and pH 7.4. Data acquisition was done in absorbance (at 280 nm) and interference modes. The sedimentation coefficients were obtained by fitting the sedimentation velocity profiles to the non-interacting species model using the SEDFIT program,1 and the continuous distribution of sedimentation coefficients was obtained considering globular proteins. Solvent density was calculated at 1.00739 g/ml, and the partial specific volume was estimated at 0.724 ml/g, using the SEDNTERP program.2
Crystallization, Data Collection, and Structure Determination
Single-chain recombinant form of human C1q globular region was concentrated to 5 mg/ml, and standard crystallization kits were screened through the EMBL HTX Lab platform at 20°C. Several initial hits were reproduced manually, using the hanging drop method by mixing equal volumes (2 μl) of the protein and reservoir solutions and adding calcium in some reservoir solutions. The crystallization conditions used and the resulting crystal morphology were very similar to those obtained previously for plasma-derived C1q-GR (10). To obtain the crystal structures presented here, the following reservoir solutions were used: (1) 30% PEG 8000, 0.1M Hepes, pH 7.5; (2) 23% PEG 3350, 0.1M Tris, 0.2M NaCl, 50 mM CaCl2, pH 8.5. Diffraction data were recorded up to 1.35 or 1.55 Å resolution at the European Synchrotron Radiation facility (ESRF) beamline ID23-eh1 and auto-processed in the C2 space group (25). The data collection statistics are provided in Table 1.
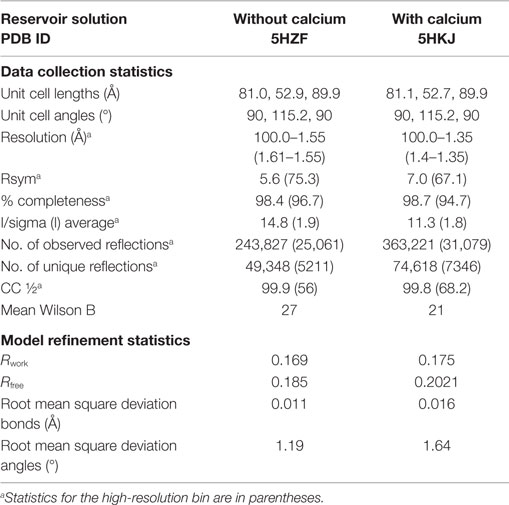
Table 1. Data collection and refinement statistics for C1q-scGR.
The position and orientation of the C1q-scGR trimeric globular domain in the asymmetric unit were determined with the molecular replacement software Phaser (26). Alternative cycles of refinement and graphics edition were performed using Refmac5 (27) and Coot (28), respectively. The final refinement cycles were performed using Phenix (29). Refinement statistics are provided in Table 1. Illustrations were prepared using Pymol (30).
Surface Plasmon Resonance Studies
Analyses were performed at 25°C using a Biacore 3000 instrument (GE Healthcare).
SPR Analyses on Immobilized C1q Protein Ligands
Calreticulin, gC1qR, and PTX3 were diluted to 20, 68, and 100 μg/ml in 10 mM sodium acetate pH 4.0, 4.0, and 3.5, respectively, and immobilized on a CM5 sensor chip (GE Healthcare) using the amine coupling chemistry in 10 mM Hepes, 150 mM NaCl, 3 mM EDTA, 0.005% surfactant P20, pH 7.4. The reference surface was submitted to the coupling steps without immobilized protein. Binding of C1q-scGR, C1q-GR, and C1q to immobilized CRT (3000–4700 RU), gC1qR (500–3400 RU), and PTX3 (5300–6200 RU) was measured at a flow rate of 20 μl/min in 50 mM Tris–HCl, 150 mM NaCl, 2 mM CaCl2, 0.005% surfactant P20, pH 7.4. The specific binding signal was obtained by subtracting the background signal over the reference surface. Regeneration of the surfaces was achieved by 10 μl injections of 10–20 mM NaOH.
SPR Analyses on Immobilized Heparin
Streptavidin (approximately 4000 RU) was immobilized on two flow cells of a CM5 sensor chip, as described previously (31). Biotinylated heparin was captured on the streptavidin surface in 10 mM Hepes, 150 mM NaCl, 0.005% surfactant P20, pH 7.4 (HBS-P) until a coupling level of 250–300 RU was obtained. Serum C1q-GR and recombinant C1q-scGR were injected over the heparin-bound surface at 20 μl/min in HBS-P. Surfaces were regenerated with 10 μl of 1M NaCl. The streptavidin surface without bound heparin was used as a reference.
SPR Data Evaluation
Data were analyzed by global fitting either to a 1:1 Langmuir binding model or to a two-state reaction binding model of both the association and dissociation phases for at least five concentrations simultaneously, using the BIAevaluation 3.2 software (GE Healthcare). Buffer blanks were subtracted from the data sets used for kinetic analysis (double referencing). Chi2 values were below 3.5 in all cases. For the two-state reaction (conformational change) model, the apparent dissociation constants were calculated from the rate constants: KD = 1/[(ka1/kd1) (1 + ka2/kd2)]. For the Langmuir binding model, the apparent equilibrium dissociation constants (KD) were calculated from the ratio of the dissociation and association rate constants (kd/ka).
Results and Discussion
Generation of a Single-Chain Recombinant Form of gC1q
A strategy derived from that used for expression of a single polypeptide protein containing three consecutive copies of the globular domain of adiponectin was chosen to produce C1q-scGR (17). As revealed by the X-ray crystal structures of the globular domains of adiponectin (32) and of C1q (10), the N- and C-termini of the three gC1q modules emerge at the base of the trimer. Short 3-amino acid linkers can thus connect the adjacent monomers A–C and C–B. The 5′-3′ A–C–B order chosen to generate C1q-scGR also corresponds to that of the three C1q genes on chromosome 1p (33) (Figure 1A).
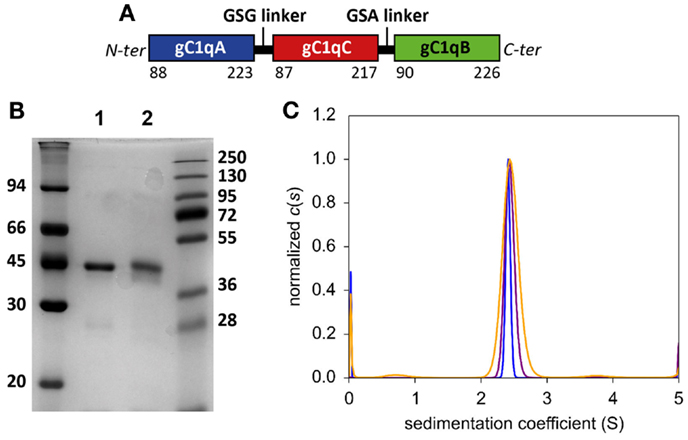
Figure 1. Biochemical characterization of purified C1q-scGR. (A) Schematic representation of mature recombinant C1q-scGR protein. (B) SDS-PAGE analysis of C1q-scGR, unreduced (lane 1) and reduced (lane 2). The molecular masses of unreduced and reduced markers (expressed in kilodaltons) are shown on the left and right sides of the gel, respectively. (C) Sedimentation velocity analysis of C1q-scGR. Analysis was performed as described in Section “Materials and Methods” and the continuous distribution of sedimentation coefficients is shown. Orange, purple and blue traces correspond to C1q-scGR samples at 0.9, 2.5, and 5 mg/ml, respectively (data acquisition in absorbance mode).
A first attempt to produce recombinant C1q-scGR was performed using a baculovirus/insect cells expression system, but the production yield was rather low since only 1 mg purified protein was recovered per liter of cell culture supernatant. In addition, the recombinant material was heterogeneous, consisting of a mixture of glycosylated and unglycosylated species (data not shown). Recombinant C1q-scGR was next produced in stably transfected 293-F mammalian cells and purified by cation-exchange chromatography. Up to 50 mg C1q-scGR could be purified from one liter of 293-F cells supernatant, which represents a 50-times higher yield compared to the baculovirus-infected insect cells. SDS-PAGE analysis showed a single band with an apparent mass of approximately 45 kDa under reducing and non-reducing conditions (Figure 1B).
N-terminal sequence analysis yielded the single sequence Lys–Asp–Gln–Pro–Arg, starting as expected at residue Lys 88 of C1qA chain. Mass spectrometry analysis yielded three peaks with masses of 47,749.84, 47,897.45, and 48,043.18 Da, accounting for a polypeptide chain with a predicted mass of 45,691.89 Da, and additional masses of 2059, 2205, and 2350 Da, corresponding to the three types of biantennary N-glycans (monosialylated, monosialylated fucosylated, and bisialylated) identified previously in serum-derived C1q (34, 35). The single N-glysosylation site at Asn 124 of C1qA is thus glycosylated in recombinant C1q-scGR. Sialidase treatment resulted in the appearance of two peaks with masses of 47,615.55 and 47,453.09 Da, compatible with asialylated biantennary N-glycans, fucosylated, or not (expected masses 1914 and 1768 Da).
Analysis of C1q-scGR by sedimentation velocity at three protein concentrations (0.9, 2.5, and 5 mg/ml) yielded a major peak accounting for 95 ± 3% of the signal with a sedimentation coefficient of 2.14 ± 0.4 S. Analysis in non-interacting species yielded a molecular mass of 41.4 ± 2 kDa, which is close to the mass measured by mass spectrometry, indicating that C1q-scGR is a monomer (Figure 1C).
X-ray Crystal Structure of C1q-scGR
Although sialidase treatment of serum-derived C1q-GR had been required to obtain crystals of this protein suitable for structure determination (10), the presence of sialic acids in C1q-scGR was not an obstacle to the determination of its crystal structure. Its X-ray structure, refined at 1.35 Å resolution (Table 1), allowed us to check that the linkers did not introduce any distortion. The recombinant and serum-derived C1q globular domains are indeed almost identical, as shown by their very small 0.1 Å RMS deviation on the 331 Cα common positions (Figure 2A). The main-chain trace of the segment encompassing the C–B linker and the first two residues of gC1qB (GSAKA) were modeled into the electron density (Figure 2B). This more rigid C–B linker only slightly alters the main-chain position of the two preceding and following residues, but the positions of their side-chains are conserved (Figure 2B). The A–C linker is more flexible, and only the first two residues of gC1qC were modeled into the electron density (Figure 2C). All the water molecules except one correspond to those observed in the structure of the plasma-derived protein (PDB code 2wnv) (36). In the absence of calcium in the reservoir solution, the electron density filling the calcium-binding site is best modeled as a magnesium ion (Figure 2D). With a RMS deviation of 0.08 Å on the 340 Cα common positions, the two refined structures (Table 1) are almost identical except for the nature of the bound ion, which is either magnesium or calcium. Thus, this ion substitution does not alter the calcium-binding environment and is reversible (Figure 2E). The presence of a single calcium (or magnesium) ion at the top of the heterotrimeric assembly has been proposed to contribute to the stability of the recognition domain of C1q (10).
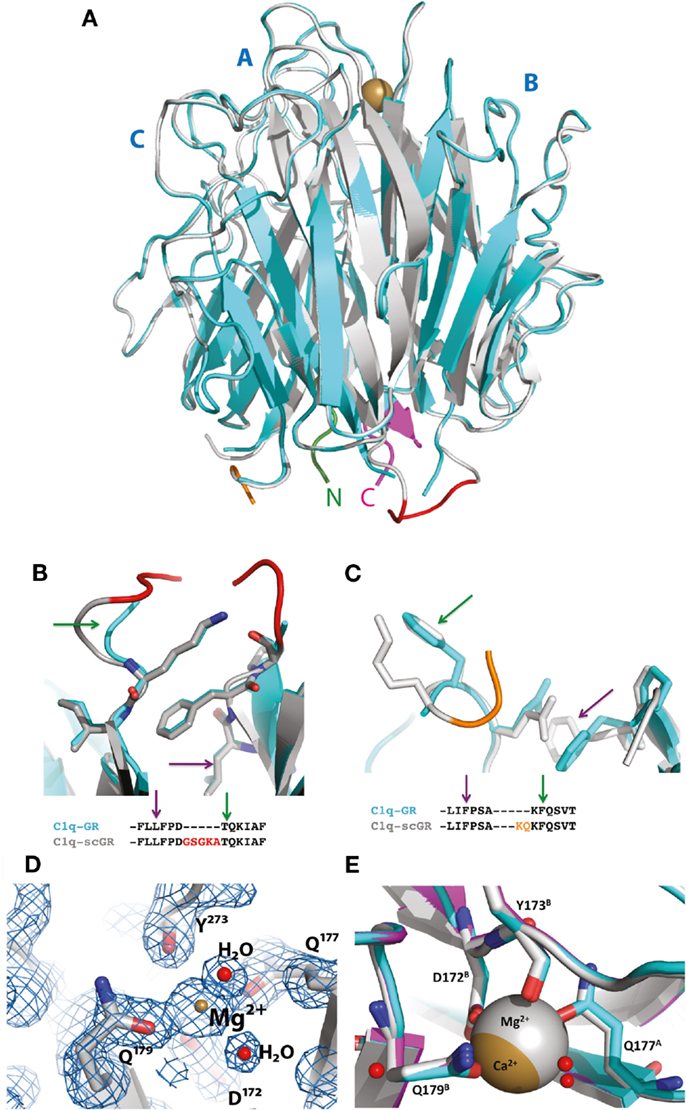
Figure 2. Crystal structure of C1q-scGR and its comparison with C1q-GR. (A) The global superposition of C1q-scGR (gray) and C1q-GR (cyan) structures illustrates their similarity, with only 0.1 Å RMS deviation on 331 common Cα positions. The A–C labels corresponding to the three C1q-GR subunits are shown in blue. The calcium ion is shown as a golden sphere. The N-terminal and C-terminal ends are shown in green and magenta, respectively. More details of the C–B (red) and A–C (orange) linkers are provided in (B) and (C), respectively. The green and magenta arrows link the details in these zooming sections with the positions in the sequences shown below. (D) In the absence of calcium in the crystallization reservoir, the calcium-binding site is occupied by a magnesium ion, as illustrated here in the 2mFo–DFc map contoured at 2 σ level. Introducing a calcium ion in the model at this position deteriorates the Rfree factor by 2% and introduces a negative peak at this position in the Fo–Fc electron density map (not shown). (E) The ion substitution does not alter the structure of the calcium-binding site environment, as illustrated by the superposition of the structures of C1q-GR (cyan), C1q-scGR with calcium (magenta), and C1q-scGR with magnesium (gray).
The X-ray crystal structure of a single-chain version of the homotrimeric globular head of adiponectin had validated the strategy used to generate the recognition domain of this C1q-related protein in a recombinant form (37). The fact that the 3D structure of C1q-scGR is virtually identical to that of the three-chain C1q-derived fragment shows that this strategy can be extended to other proteins containing trimeric gC1q domains.
Functional Properties of C1q-scGR
The interaction properties of C1q-scGR were analyzed by surface plasmon resonance (SPR) and compared to those of serum-derived C1q-GR, using known physiological ligands of C1q: the receptor for the globular heads of C1q (gC1qR), CRT, the long pentraxin PTX3, and heparin. All ligands were immobilized on the surface of a sensor chip and the serum-derived and recombinant forms of C1q-GR were used as soluble analytes. Both C1q-scGR and C1q-GR bound to the immobilized ligands with comparable association and dissociation rate constants, yielding comparable apparent KD values in the sub-micromolar range (Figure 3; Table 2). This indicated that the recombinant single-chain protein retained the binding capacities of its serum-derived counterpart and was therefore fully functional. The KD values obtained here for the interaction with CRT (494–510 nM) and gC1qR (304–344 nM) were in the same range as those reported previously for binding of C1q-GR to CRT (830 nM) (21) and gC1qR (370 nM) (38). The KD value obtained for binding to heparin (51 nM) is slightly lower than that obtained previously for the interaction of C1q-GR with 6 kDa heparin (154 nM) (36), a difference that might be explained by the higher molecular weight heparin (15 kDa) used in the present study. Similar results were obtained when the binding experiments were performed in the absence of added calcium in the running buffer (not shown). In light of our structural data, it is likely that the calcium-binding site was still occupied under these conditions, since no chelating agent such as EDTA was used to remove the bound ion.
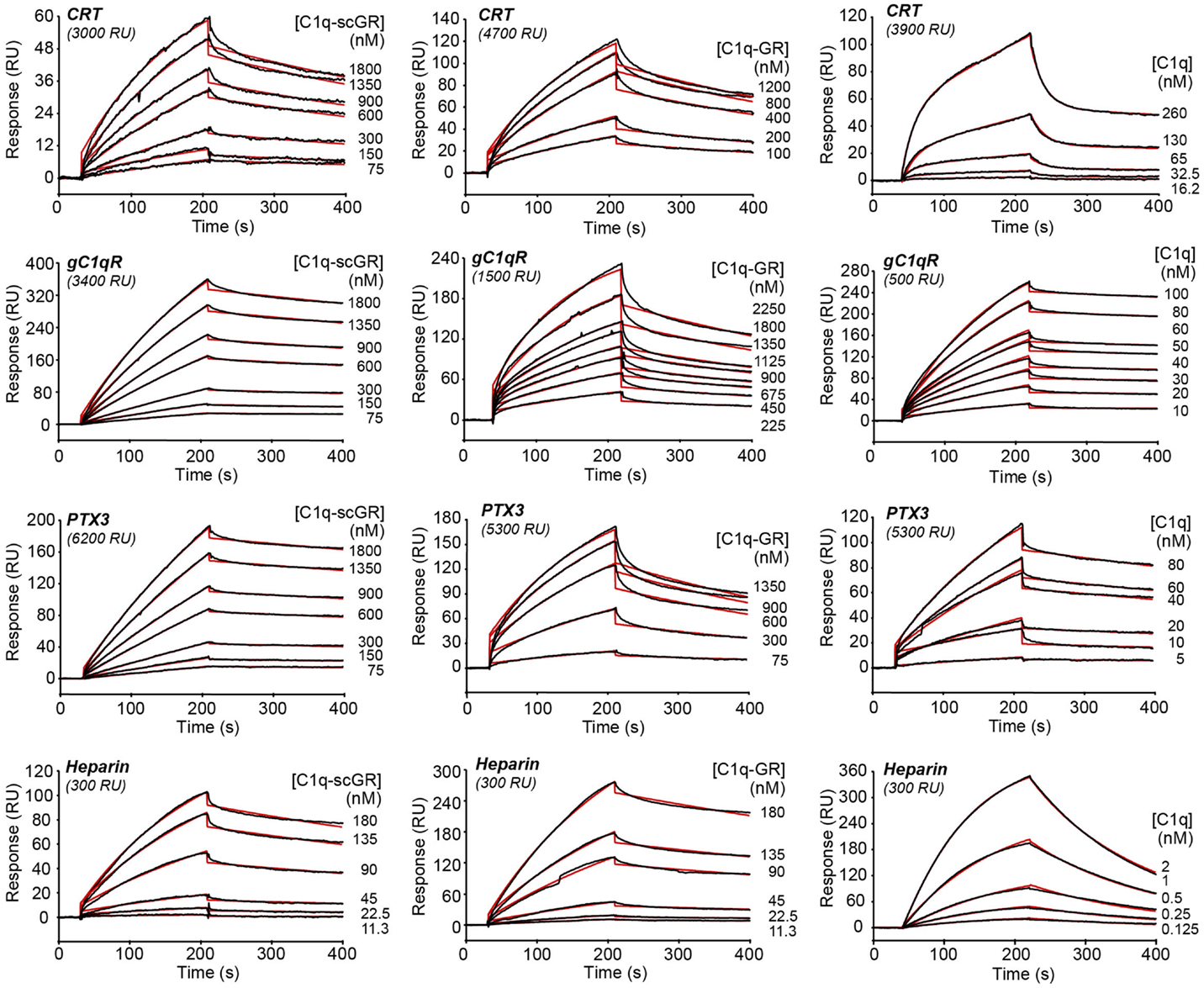
Figure 3. Kinetic analysis of the interaction of C1q-scGR, C1q-GR, and C1q with immobilized CRT, gC1qR, PTX3, and heparin. The C1q samples (60 μl) were injected at the indicated concentrations over immobilized CRT, gC1qR, PTX3 in 50 mM Tris–HCl, 150 mM NaCl, 2 mM CaCl2, 0.005% surfactant P20, pH 7.4, and over biotinylated heparin in 10 mM Hepes, 150 mM NaCl, 0.005% surfactant P20, pH 7.4. Fits are shown as red lines and were obtained by global fitting of the data using a 1:1 Langmuir binding model, except for the C1q–CRT interaction for which a two-state reaction (conformational change) model was used.
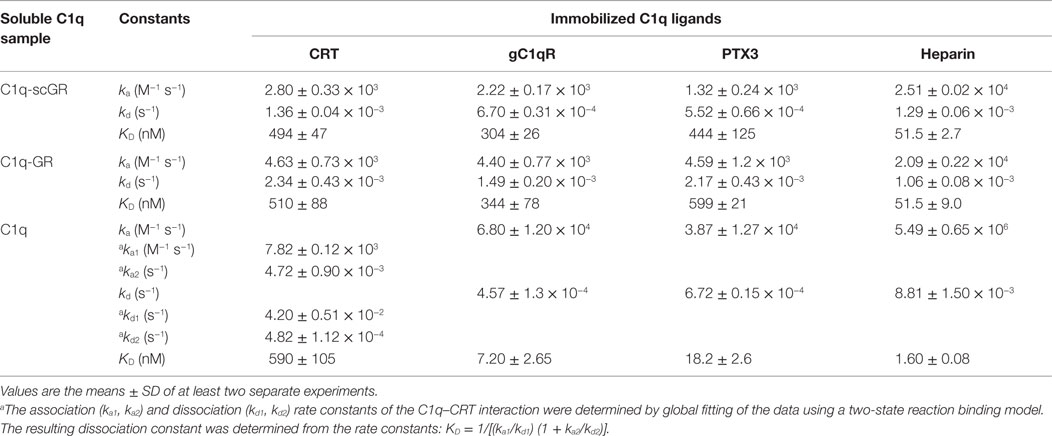
Table 2. Kinetic and dissociation constants for binding of C1q-scGR, C1q-GR, and C1q to immobilized C1q ligands.
When the interaction experiments were performed using full-length C1q as soluble analyte, the binding affinities for all ligands, except CRT, were in the nanomolar range (Figure 3; Table 2). The observation that the C1q globular domain binds to immobilized gC1qR, PTX3, and heparin with a lower affinity (41- to 47-, 24- to 32-, and 32-fold, respectively) than intact C1q is consistent with the fact that C1q-scGR and C1q-GR lack the binding avidity of the hexameric C1q molecule. The decrease in affinity resulted mainly from a decrease in the ka value, the kd remaining essentially in the same range. A similar 44-fold decreased affinity has been observed previously for the prion protein, another C1q ligand known to be recognized through C1q globular domain (39).
Proper kinetic analysis of the CRT–C1q interaction required the use of a two-state reaction binding model, taking into account conformational changes leading to an increasingly more stable complex formed in two steps, as reported previously for C1q binding to placenta-derived and recombinant CRT (21). The two groups of kinetic constants and the resulting apparent affinity constant (KD), are listed in Table 2. As proposed previously (21), our data also suggest that CRT recognition by intact C1q implies conformational changes that do not take place in the isolated GR domains. Interestingly, the apparent KD values obtained here for the interaction of CRT with C1q or its globular domain were all in the sub-micromolar range, suggesting an interaction mechanism differing from those involving other C1q ligands, such as gC1qR, PTX3, or the prion protein. It should be mentioned that a 2.4- to 7-fold increase in the C1q versus C1q-GR affinity to placenta-derived and recombinant CRT was observed previously, which corresponds anyway to a much lower avidity component compared to the other C1q ligands. Further investigation, including site-directed mutagenesis and/or structural analyses, will be needed to propose a relevant model for the CRT–C1q-GR interaction.
Of note, the same experimental SPR settings did not allow us to compare the binding affinity of C1q and its globular regions for IgG, a classical C1q ligand. Indeed, although full-length C1q readily bound to immobilized human IgG when injected at a concentration of 10 nM, only very weak binding of C1q-GR (either serum-derived or recombinant) injected at concentrations in the micromolar range was observed (Figure S1 in Supplementary Material). This precluded determination of kinetic constants for C1q-GR binding to IgG, but revealed, here too, a difference between full-length C1q and its recognition domains. It is well known that, under physiological conditions, efficient complement activation is triggered by multivalent binding of C1q to antigen–IgG complexes. These conditions can be reproduced artificially in ELISA tests using adsorbed heat-aggregated IgG. Therefore, it is not totally unexpected that the globular regions bind only weakly to immobilized single IgG molecules. The immobilized gC1qR and PTX3 molecules are naturally trimers and octamers, respectively, and it is expected that each of these ligands can bind to a single globular head of C1q, although with less affinity than to full-length C1q.
Conclusion
The availability of a recombinant functional form of the heterotrimeric globular regions of C1q opens the way for deciphering the molecular basis of the binding versatility of C1q by mapping the residues involved in the recognition of its numerous targets using site-directed mutagenesis. Although it has been shown previously that the three isolated subunits mediate different individual binding properties (12), it is now possible to assess the effects of single residue mutations in the heterotrimeric context of C1q-scGR, as it occurs in native C1q. In addition, given the compact structure of the domain, it appears likely that recognition of certain ligands will involve residues contributed by several subunits (10), a hypothesis that can now be tested experimentally. The availability of two recombinant forms of C1q, the full-length protein (23) and its recognition domain, will allow comparison of their binding and effector properties, taking into account the avidity provided by the hexameric full-length C1q molecule.
In addition to basic research, such artificial gC1q molecules should be of interest for biomedical applications. The contribution of complement in the pathogenesis of many important diseases, including neurodegenerative, infectious, and autoimmune disorders, is now well recognized and C1q is an attractive target for anti-complement therapy (40). C1q-scGR molecules, possibly engineered to form multimers, could thus be used in the fluid phase as natural competitors to inhibit the classical complement pathway at the initial recognition step without any risk of triggering the immune effector mechanisms mediated by the collagen-like regions.
Engineered C1q-scGR molecules might also be used to functionalize magnetic nanobeads or hemoadsorption filters for in vitro blood cleansing, as described recently for the carbohydrate recognition domain of human MBL, a recognition protein of the lectin complement pathway (41, 42). MBL-coated devices were shown to efficiently capture bacteria, fungi, and endotoxins from whole human blood and are promising tools in sepsis therapy. Using C1q-scGR would broader the field of application of such devices beyond pathogenic microorganisms because of the capacity of C1q to recognize a variety of potentially noxious altered self elements such as amyloid fibrils and the toxic form of the prion protein. In addition, the fact that C1q specifically reacts with circulating immune complexes and acute-phase proteins such as pentraxins might open the way to the use of C1q-scGR hemoadsorption for the treatment of patients with severe autoimmune diseases such as SLE. It should be mentioned indeed that plasma immunoadsorption on a C1q-column (prepared with immobilized serum-purified porcine C1q) has been successfully used to treat a few SLE patients (43, 44). Using recombinant C1q-scGR, it is now possible to address the potential contamination risks associated with animal serum-derived proteins.
Author Contributions
NT, CG, and IB designed the study; CM, IB, AC, and NT performed the research; CM, IB, CG, and NT analyzed the data; BB and BG contributed new reagents; NT, CG, and CM wrote the manuscript; all authors revised and approved the final version of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Gérard Arlaud for helpful suggestions and discussions at the early stages of this project. This work used the platforms of the Grenoble Instruct center (ISBG; UMS 3518 CNRS-CEA-UJF-EMBL) with support from FRISBI (ANR-10-INSB-05-02) and GRAL (ANR-10-LABX-49-01) within the Grenoble Partnership for Structural Biology (PSB). We thank Luca Signor, Jean-Pierre Andrieu, and Aline Le Roy for assistance and access to the mass spectrometry, protein N-terminal sequencing, and analytical ultracentrifugation facilities, respectively. Access to the European Synchrotron Radiation Facility beamline ID23-eh1 is acknowledged. This work was supported by a grant from the French National Research Agency (ANR-09-PIRI-0021). The contribution of the Italian Health Ministry (RF-2010-2310888) is gratefully acknowledged.
Supplementary Material
The Supplementary Material for this article can be found online at http://journal.frontiersin.org/article/10.3389/fimmu.2016.00079
Abbreviations
C1q-GR, globular region of C1q; C1q-scGR, single-chain globular region of C1q; CRT, calreticulin; gC1qR, receptor for the globular head of C1q; SPR, surface plasmon resonance.
Footnotes
References
1. Fraser DA, Tenner AJ. Directing an appropriate immune response: the role of defense collagens and other soluble pattern recognition molecules. Curr Drug Targets (2008) 9:113–22. doi: 10.2174/138945008783502476
3. Tacnet-Delorme P, Chevallier S, Arlaud GJ. Beta-amyloid fibrils activate the C1 complex of complement under physiological conditions: evidence for a binding site for A beta on the C1q globular regions. J Immunol (2001) 167:6374–81. doi:10.4049/jimmunol.167.11.6374
4. Klein MA, Kaeser PS, Schwarz P, Weyd H, Xenarios I, Zinkernagel RM, et al. Complement facilitates early prion pathogenesis. Nat Med (2001) 7:488–92. doi:10.1038/86567
5. Erlich P, Dumestre-Perard C, Ling WL, Lemaire-Vieille C, Schoehn G, Arlaud GJ, et al. Complement protein C1q forms a complex with cytotoxic prion protein oligomers. J Biol Chem (2010) 285:19267–76. doi:10.1074/jbc.M109.071860
6. Biro A, Thielens NM, Cervenak L, Prohaszka Z, Fust G, Arlaud GJ. Modified low density lipoproteins differentially bind and activate the C1 complex of complement. Mol Immunol (2007) 44:1169–77. doi:10.1016/j.molimm.2006.06.013
7. Korb LC, Ahearn JM. C1q binds directly and specifically to surface blebs of apoptotic human keratinocytes: complement deficiency and systemic lupus erythematosus revisited. J Immunol (1997) 158:4525–8.
8. Taylor PR, Carugati A, Fadok VA, Cook HT, Andrews M, Carroll MC, et al. A hierarchical role for classical pathway complement proteins in the clearance of apoptotic cells in vivo. J Exp Med (2000) 192:359–66. doi:10.1084/jem.192.3.359
9. Navratil JS, Watkins SC, Wisnieski JJ, Ahearn JM. The globular heads of C1q specifically recognize surface blebs of apoptotic vascular endothelial cells. J Immunol (2001) 166:3231–9. doi:10.4049/jimmunol.166.5.3231
10. Gaboriaud C, Juanhuix J, Gruez A, Lacroix M, Darnault C, Pignol D, et al. The crystal structure of the globular head of complement protein C1q provides a basis for its versatile recognition properties. J Biol Chem (2003) 278:46974–82. doi:10.1074/jbc.M307764200
11. Gaboriaud C, Frachet P, Thielens NM, Arlaud GJ. The human c1q globular domain: structure and recognition of non-immune self ligands. Front Immunol (2011) 2:92. doi:10.3389/fimmu.2011.00092
12. Kishore U, Gupta SK, Perdikoulis MV, Kojouharova MS, Urban BC, Reid KB. Modular organization of the carboxyl-terminal, globular head region of human C1q A, B, and C chains. J Immunol (2003) 171:812–20. doi:10.4049/jimmunol.171.2.812
13. Kojouharova MS, Tsacheva IG, Tchorbadjieva MI, Reid KB, Kishore U. Localization of ligand-binding sites on human C1q globular head region using recombinant globular head fragments and single-chain antibodies. Biochim Biophys Acta (2003) 1652:64–74. doi:10.1016/j.bbapap.2003.08.003
14. Kojouharova MS, Gadjeva MG, Tsacheva IG, Zlatarova A, Roumenina LT, Tchorbadjieva MI, et al. Mutational analyses of the recombinant globular regions of human C1q A, B, and C chains suggest an essential role for arginine and histidine residues in the C1q-IgG interaction. J Immunol (2004) 172:4351–8. doi:10.4049/jimmunol.172.7.4351
15. Roumenina LT, Ruseva MM, Zlatarova A, Ghai R, Kolev M, Olova N, et al. Interaction of C1q with IgG1, C-reactive protein and pentraxin 3: mutational studies using recombinant globular head modules of human C1q A, B, and C chains. Biochemistry (2006) 45:4093–104. doi:10.1021/bi052646f
16. Roumenina LT, Popov KT, Bureeva SV, Kojouharova M, Gadjeva M, Rabheru S, et al. Interaction of the globular domain of human C1q with Salmonella typhimurium lipopolysaccharide. Biochim Biophys Acta (2008) 1784:1271–6. doi:10.1016/j.bbapap.2008.04.029
17. Ge H, Xiong Y, Lemon B, Lee KJ, Tang J, Wang P, et al. Generation of novel long-acting globular adiponectin molecules. J Mol Biol (2010) 399:113–9. doi:10.1016/j.jmb.2010.03.062
18. Arlaud GJ, Sim RB, Duplaa AM, Colomb MG. Differential elution of Clq, Clr and Cls from human Cl bound to immune aggregates. Use in the rapid purification of Cl subcomponents. Mol Immunol (1979) 16:445–50. doi:10.1016/0161-5890(79)90069-5
19. Bottazzi B, Vouret-Craviari V, Bastone A, De Gioia L, Matteucci C, Peri G, et al. Multimer formation and ligand recognition by the long pentraxin PTX3. Similarities and differences with the short pentraxins C-reactive protein and serum amyloid P component. J Biol Chem (1997) 272:32817–23. doi:10.1074/jbc.272.52.32817
20. Ghebrehiwet B, Jesty J, Xu S, Vinayagasundaram R, Vinayagasundaram U, Ji Y, et al. Structure-function studies using deletion mutants identify domains of gC1qR/p33 as potential therapeutic targets for vascular permeability and inflammation. Front Immunol (2011) 2:58. doi:10.3389/fimmu.2011.00058
21. Paidassi H, Tacnet-Delorme P, Verneret M, Gaboriaud C, Houen G, Duus K, et al. Investigations on the C1q-calreticulin-phosphatidylserine interactions yield new insights into apoptotic cell recognition. J Mol Biol (2011) 408:277–90. doi:10.1016/j.jmb.2011.02.029
22. Rossi V, Bally I, Thielens NM, Esser AF, Arlaud GJ. Baculovirus-mediated expression of truncated modular fragments from the catalytic region of human complement serine protease C1s. Evidence for the involvement of both complement control protein modules in the recognition of the C4 protein substrate. J Biol Chem (1998) 273:1232–9. doi:10.1074/jbc.273.2.1232
23. Bally I, Ancelet S, Moriscot C, Gonnet F, Mantovani A, Daniel R, et al. Expression of recombinant human complement C1q allows identification of the C1r/C1s-binding sites. Proc Natl Acad Sci U S A (2013) 110:8650–5. doi:10.1073/pnas.1304894110
24. Jacquet M, Lacroix M, Ancelet S, Gout E, Gaboriaud C, Thielens NM, et al. Deciphering complement receptor type 1 interactions with recognition proteins of the lectin complement pathway. J Immunol (2013) 190:3721–31. doi:10.4049/jimmunol.1202451
25. Monaco S, Gordon E, Bowler MW, Delageniere S, Guijarro M, Spruce D, et al. Automatic processing of macromolecular crystallography X-ray diffraction data at the ESRF. J Appl Crystallogr (2013) 46:804–10. doi:10.1107/S0021889813006195
26. McCoy AJ, Grosse-Kunstleve RW, Adams PD, Winn MD, Storoni LC, Read RJ. Phaser crystallographic software. J Appl Crystallogr (2007) 40:658–74. doi:10.1107/S0021889807021206
27. Murshudov GN, Skubak P, Lebedev AA, Pannu NS, Steiner RA, Nicholls RA, et al. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr D Biol Crystallogr (2011) 67:355–67. doi:10.1107/S0907444911001314
28. Emsley P, Lohkamp B, Scott WG, Cowtan K. Features and development of Coot. Acta Crystallogr D Biol Crystallogr (2010) 66:486–501. doi:10.1107/S0907444910007493
29. Adams PD, Afonine PV, Bunkoczi G, Chen VB, Davis IW, Echols N, et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr D Biol Crystallogr (2010) 66:213–21. doi:10.1107/S0907444909052925
31. Rossi V, Bally I, Ancelet S, Xu Y, Fremeaux-Bacchi V, Vives RR, et al. Functional characterization of the recombinant human C1 inhibitor serpin domain: insights into heparin binding. J Immunol (2010) 184:4982–9. doi:10.4049/jimmunol.0902016
32. Shapiro L, Scherer PE. The crystal structure of a complement-1q family protein suggests an evolutionary link to tumor necrosis factor. Curr Biol (1998) 8:335–8. doi:10.1016/S0960-9822(98)70133-2
33. Sellar GC, Blake DJ, Reid KB. Characterization and organization of the genes encoding the A-, B- and C-chains of human complement subcomponent C1q. The complete derived amino acid sequence of human C1q. Biochem J (1991) 274(Pt 2):481–90. doi:10.1042/bj2740481
34. Mizuochi T, Yonemasu K, Yamashita K, Kobata A. The asparagine-linked sugar chains of subcomponent C1q of the first component of human complement. J Biol Chem (1978) 253:7404–9.
35. Pflieger D, Przybylski C, Gonnet F, Le Caer JP, Lunardi T, Arlaud GJ, et al. Analysis of human C1q by combined bottom-up and top-down mass spectrometry: detailed mapping of post-translational modifications and insights into the C1r/C1s binding sites. Mol Cell Proteomics (2010) 9:593–610. doi:10.1074/mcp.M900350-MCP200
36. Garlatti V, Chouquet A, Lunardi T, Vives R, Paidassi H, Lortat-Jacob H, et al. Cutting edge: C1q binds deoxyribose and heparan sulfate through neighboring sites of its recognition domain. J Immunol (2010) 185:808–12. doi:10.4049/jimmunol.1000184
37. Min X, Lemon B, Tang J, Liu Q, Zhang R, Walker N, et al. Crystal structure of a single-chain trimer of human adiponectin globular domain. FEBS Lett (2012) 586:912–7. doi:10.1016/j.febslet.2012.02.024
38. Tacnet P, Cheong EC, Goeltz P, Ghebrehiwet B, Arlaud GJ, Liu XY, et al. Trimeric reassembly of the globular domain of human C1q. Biochim Biophys Acta (2008) 1784:518–29. doi:10.1016/j.bbapap.2007.12.005
39. Blanquet-Grossard F, Thielens NM, Vendrely C, Jamin M, Arlaud GJ. Complement protein C1q recognizes a conformationally modified form of the prion protein. Biochemistry (2005) 44:4349–56. doi:10.1021/bi047370a
40. Roos A, Ramwadhdoebe TH, Nauta AJ, Hack CE, Daha MR. Therapeutic inhibition of the early phase of complement activation. Immunobiology (2002) 205:595–609. doi:10.1078/0171-2985-00157
41. Kang JH, Super M, Yung CW, Cooper RM, Domansky K, Graveline AR, et al. An extracorporeal blood-cleansing device for sepsis therapy. Nat Med (2014) 20:1211–6. doi:10.1038/nm.3640
42. Didar TF, Cartwright MJ, Rottman M, Graveline AR, Gamini N, Watters AL, et al. Improved treatment of systemic blood infections using antibiotics with extracorporeal opsonin hemoadsorption. Biomaterials (2015) 67:382–92. doi:10.1016/j.biomaterials.2015.07.046
43. Berner B, Scheel AK, Schettler V, Hummel KM, Reuss-Borst MA, Muller GA, et al. Rapid improvement of SLE-specific cutaneous lesions by C1q immunoadsorption. Ann Rheum Dis (2001) 60:898–9.
Keywords: innate immune recognition, complement, C1q, protein engineering, X-ray crystallography, surface plasmon resonance
Citation: Moreau C, Bally I, Chouquet A, Bottazzi B, Ghebrehiwet B, Gaboriaud C and Thielens N (2016) Structural and Functional Characterization of a Single-Chain Form of the Recognition Domain of Complement Protein C1q. Front. Immunol. 7:79. doi: 10.3389/fimmu.2016.00079
Received: 15 January 2016; Accepted: 18 February 2016;
Published: 02 March 2016
Edited by:
Uday Kishore, Brunel University London, UKCopyright: © 2016 Moreau, Bally, Chouquet, Bottazzi, Ghebrehiwet, Gaboriaud and Thielens. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christine Gaboriaud, Y2hyaXN0aW5lLmdhYm9yaWF1ZEBpYnMuZnI=;
Nicole Thielens, bmljb2xlLnRoaWVsZW5zQGlicy5mcg==
†Christophe Moreau and Isabelle Bally contributed equally to this work.