 Emilia Pachulec
Emilia Pachulec Vanessa Neitzke-Montinelli
Vanessa Neitzke-Montinelli João P. B. Viola
João P. B. Viola- Program of Cellular Biology, Brazilian National Cancer Institute (INCA), Rio de Janeiro, Brazil
Nuclear factor of activated T cells (NFAT) 2 null mutant mice die in utero of cardiac failure, precluding analysis of the role of NFAT2 in lymphocyte responses. Only the NFAT2−/−/Rag-1−/− chimeric mice model gave insight into the role of NFAT2 transcription factor in T lymphocyte development, activation, and differentiation. As reports are mainly focused on the role of NFAT2 in CD4+ T lymphocytes activation and differentiation, we decided to investigate NFAT2’s impact on CD8+ T lymphocyte responses. We report that NFAT2 is phosphorylated and inactive in the cytoplasm of naive CD8+ T cells, and upon TCR stimulation, it is dephosphorylated and translocated into the nucleus. To study the role of NFAT2 in CD8+ T responses, we employed NFAT2fl/flCD4-Cre mice with NFAT2 deletion specifically in T cells. Interestingly, the absence of NFAT2 in T cells resulted in increased percentage of non-conventional innate-like CD8+ T cells. These cells were CD122+, rapid producer of interferon gamma (IFN-γ) and had characteristics of conventional memory CD8+ T cells. We also observed an expansion of PLZF+ expressing CD3+ thymocyte population in the absence of NFAT2 and increased IL-4 production. Furthermore, we found that CD8+ T lymphocytes deficient in NFAT2 had reduced activation, proliferation, and IFN-γ and IL-2 production at suboptimal TCR strength. NFAT2 absence did not significantly influence differentiation of CD8+ T cells into cytotoxic effector cells but reduced their IFN-γ production. This work documents NFAT2 as a negative regulator of innate-like CD8+ T cells development. NFAT2 is required for complete CD8+ T cell responses at suboptimal TCR stimulation and regulates IFN-γ production by cytotoxic CD8+ T cells in vitro.
Introduction
CD8+ T cells are the key mediators in cell-mediated immunity against intracellular bacteria, viral infections, and tumor cells. Conventional CD8+ T cells develop in the thymus from a bone marrow-derived multipotent T cells progenitor through a series of defined and coordinated developmental stages of differentiation, positive and negative selection, and maturation. Mature single positive (SP) CD8+ T cells leave the thymus and migrate to peripheral lymphoid organs. In the peripheral lymphoid organs, naive CD8+ T cells, which are activated by specific antigens, differentiate into effector T cells that are able to kill target cells through the release of cytotoxic granules, secretion of cytokines, such as interferon gamma (IFN-γ) and Fas/FasL-dependent pathway. A small number of activated CD8+ T cells develop into memory T cells that provide long-term protection against reinfection (1). There are several subsets of memory CD8+ T cells, and among them, central and effector memory T cells (2). Central memory T cells express high level of CD62L and reside mainly in peripheral lymphoid organs, whereas effector memory T cells express low level of CD62L and are present mainly in non-lymphoid tissues. The development of effector or memory CD8+ T cells is dependent on the environment created by the innate immune system, for example, the presence of cytokines such as IL-12 promoting effector CD8+ T cells differentiation (3) or IL-7 and IL-15 required for the maintenance and survival of memory CD8+ T cells (4).
In addition to conventional CD8+ T cells, there are also several subsets of CD8+ T cells that acquire functions as a result of their maturation process rather than through antigen activation. These cells are believed to be selected by hematopoietic cells and are dependent on signals transduced through signaling lymphocyte activation molecule (SLAM) family members (5, 6). A subset of these cells, called memory-like or innate-like T cells, has been described in a few gene knockout animal models (5–14) and humans (15). These cells share characteristics with conventional memory CD8+ T cells, such as high expression of CD44 and CD122 (5, 6), and with innate cells as they can rapidly secrete IFN-γ upon stimulation (16). It has been reported that memory-like CD8+ T cells play important roles in the innate response against infections such as Listeria monocytogenes, chronic infections with viruses such as Herpes virus (17–19), and tumor cell lines in vitro (20).
Nuclear factor of activated T cells (NFAT) was originally described as a transcription factor inducing the expression of interleukin 2 (IL-2) (21). The NFAT family of transcription factors consists of five members, named NFAT1–5, and the main forms expressed in T cells are NFAT1 and NFAT2 (22). NFAT1 is constitutively expressed in T cells (23), whereas NFAT2 is induced upon T-cell receptor stimulation (24). NFAT proteins reside phosphorylated in the cytoplasm. In activated lymphocytes, NFAT is dephosphorylated by calcineurin (25–28), translocates from the cytoplasm into the nucleus (29–31), where coupled with other transcription factors (26, 32) binds to the promotor regions of multiple genes to induce their transcription. Previous studies showed that NFAT proteins play regulatory roles during T-cell differentiation and effector functions. NFAT1 deficiency in T cells diminished Th1 differentiation and induced IL-4 production (33). NFAT1 was also reported to contribute to IL-21 expression and to limit the immunosuppressive function of CD4+CD25+Foxp3+GITR+ T regulatory (Treg) cells (34).
The role of NFAT2 in T-cell differentiation is not fully understood, as the total inactivation of NFAT2 gene in mice led to an early death of mice embryos (35). Previous analysis on Th1- and Th2-skewed T cells isolated from NFAT2−/−/Rag-1−/− chimeric mice revealed an involvement of NFAT2 in the induction of the Th2-cytokines IL-4 and IL-6, whereas it had no effect on IFN-γ and IL-2 expression in Th1 cells (36–38). NFAT2 binding sites were found within the Il17a promoter (39) and the Il10 promoter (40). Recently, NFAT2 has been shown as a positive regulator of RORγT and Th17 cytokines during TGF-β-mediated Th17-cell differentiation (41). NFAT2-deficient TGF-β-induced iTreg cells showed a slight reduction of CD25 and Foxp3 expression as compared to WT cells (42), indicating no essential role for NFAT2 in iTreg cell development. Up to now, most of the available studies are focused on the role of NFAT2 in CD4+ T lymphocytes differentiation and little is known about its function in CD8+ T lymphocytes responses.
In this study, we analyzed the role of NFAT2 in CD8+ T cell development and differentiation with the help of conditional NFAT2-deficient mice that were generated by crossing NFAT2fl/fl mice to CD4-Cre mice. These mice show a functional NFAT2 deficiency beyond double positive (DP) thymocytes, consequently CD8+ mature T cells. Our results indicate that NFAT2 plays an important role in the development of innate-like CD8+ T cells in the thymus. We further demonstrate that conditional inactivation of NFAT2 in T cells alter the threshold of CD8+ T cell activation, proliferation, and cytokines production but not differentiation. NFAT2 is not essential for differentiation into effector CD8+ T lymphocytes in vitro; however, it is important for IFN-γ production by cytotoxic CD8+ T cells.
Materials and Methods
Animals
C57BL/6 and NFAT2fl/flCD4-Cre were bred and maintained at the Brazilian National Cancer Institute (INCA) animal facility (Rio de Janeiro, RJ, Brazil). Also, 6- to 12-week-old mice were used in all experiments. NFAT2fl/flCD4-Cre mice were generated in Dr. Anjana Rao’s laboratory (La Jolla Institute for Allergy and Immunology, San Diego, CA, USA). All animal experiments were performed in accordance with the Brazilian Government’s ethical and animal experimental regulations. The experiments were approved and conducted according to the animal welfare guidelines of the Ethics Committee of Animal Experimentation from INCA (CEUA process no. 004/13).
Cell Culture
Primary cells were cultured in DMEM supplemented with 10% FCS, l-glutamine, streptomycin–penicillin, essential and non-essential amino acids, vitamins, HEPES, and 2-ME (all from Gibco) in a humidified environment containing 5% CO2 at 37°C. P815 cell line was cultured in RPMI supplemented with 10% FCS, l-glutamine, streptomycin–penicillin, sodium pyruvate, and 2-ME (all from Gibco). CD8+ T cells were purified from total lymph nodes (inguinal, brachial, and axillary). Cells were isolated with Dynal Mouse CD8 Negative Isolation Kit (Invitrogen). For cytometric purity analysis, cells were stained with anti-CD4-PE and anti-CD8-FITC Abs (both from BD Pharmingen) and analyzed by flow cytometry on a FACScan (BD Biosciences). Cell populations were isolated to >95% purity. Total lymph nodes or purified CD8+ T lymphocytes were activated in vitro for indicated times with plate-bound anti-CD3 (1 μg/ml; BD Pharmingen; otherwise indicated) plus soluble anti-CD28 (1 μg/ml; BD Pharmingen). To differentiate CD8+ T lymphocytes into cytotoxic CD8+T lymphocytes in vitro, 1 × 106 cells were activated for 48 h with plate-bound anti-CD3 (1 μg/ml) plus soluble anti-CD28 (1 μg/ml). Cells were expanded daily for 4 days with 200 U/ml of murine recombinant IL-2 (43). At day 5, cells were analyzed by flow cytometry and in cytotoxicity assay in vitro.
Cytoplasmic and Nuclear Protein Extraction
Purified CD8+ T lymphocytes (3 × 106) were stimulated or left untreated for indicated times with plate-bound anti-CD3 plus soluble anti-CD28 (both at 1 μg/ml). To extract cytoplasmic fraction, cells were lysed in buffer containing 10 mM Tris-Cl pH 7.5, 10 mM NaCl, 3 mM MgCl2, 0.5 mM DTT, 0.1 mM EDTA, 0.5% NP-40, 1 mM PMSF, and protease inhibitor. Nuclear extracts were obtained by lysis of the nuclei with buffer containing 40 mM Tris-Cl pH 7.5, 10 mM EDTA, 60 mM sodium pyrophosphate, and 5% SDS followed by incubation at 100°C for 15 min.
Western Blot
Total protein extract from 3 × 106 T lymphocytes was obtained from cells lysis in buffer containing 40 mM Tris pH 7.5, 60 mM sodium pyrophosphate, 10 mM EDTA, and 5% SDS, followed by incubation at 100°C for 15 min. Total cell lysates were resolved by SDS-PAGE, and the separated proteins were transferred onto a nitrocellulose membrane. The antibodies used were GAPDH monoclonal antibody 6C5 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) and NFAT2 monoclonal antibody 7A6 (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The immunodetection was performed with the ECL Western Blotting Detection Kit (GE Healthcare).
Immunofluorescence
Intracellular localization of NFAT2 protein was analyzed in purified CD8+ T cells (5 × 105 cells) from C57BL/6 mice by immunofluorescence staining. Briefly, cells were stimulated or left unstimulated for indicated time with plate-bound anti-CD3 plus soluble anti-CD28 (both at 1 μg/ml). Then, cells were fixed in 4% paraformaldehyde, permeabilized with 0.5% Non-idet P-40, and stained with anti-NFAT2 Ab. The cells were photographed with Confocal Laser Scanning Microscope FV10i-O Olympus.
Flow Cytometry
To analyze cell surface expression levels of the various markers, the cell suspensions (1 × 106) were incubated with following mAbs: anti-CD3-APC, anti-B220-FITC, anti-CD4-PE, anti-CD4-FITC, anti-CD8-FITC, anti-NK1.1-PerCP.Cy5.5, anti-PLZF-PE, anti-CD44-FITC, anti-CD69-APC, anti-CD62L-PE, anti-CD62L-PerCP, anti-CD25-APC, anti-CD122-PE, and anti-CD127-PE (all BD Pharmingen) acquired in a FACScan (Becton Dickinson, Mountain View, CA, USA) and analyzed using FlowJo software.
Intracellular Staining
For intracellular cytokine staining, 1 × 106 cells were stimulated in vitro for 6 h with PMA (10 nM) plus ionomycine (1 μM, both from Calbiochem). Brefeldin A (1:1000; BD Pharmingen) was added to the culture for last 2 h. Cells were harvested and stained with anti-CD8-FITC Abs. Then, cells were fixed, permeabilized, and stained with anti-IFN-γ-FITC, anti-IL-2-PE, anti-IL-4-APC, and anti-Granzime B-FITC Abs. For PLZF intracellular staining, cells were harvested and stained with anti-CD3-APC, fixed, permeablized, and stained with anti-PLZF-PE. Samples were analyzed by flow cytometry on a FACScan (BD Biosciences) and FlowJo software.
Proliferation Assay
Purified CD8+ T lymphocytes (5 × 106) were stained with CFSE Cell Proliferation Assay (Invitrogen) according to manufacturer’s instructions, then stimulated or not with plate-bound anti-CD3 (0.25 μg/ml) plus soluble anti-CD28 (1 μg/ml) for indicated times in the absence or presence of 200 U/ml of IL-2. Carboxyfluorescein diacetate succinimidyl ester (CFSE) dilution was analyzed by flow cytometry on FACScan (BD Biosciences) and FlowJo Software.
RNA Extractions and Real-time RT-PCR
Total RNA from total or sorted into CD44low and CD44high naive CD8+ T lymphocytes, total thymocytes, CD4+ thymocytes, and total splenocytes was extracted using Trizol LS Reagent (Invitrogen), and first-strand cDNA was synthesized using ImProm-II Reverse Transcription System (Promega) or Superscript II kit (Life Technology) for sorted CD44low and CD44high CD8+ T lymphocytes and CD4+ thymocytes. Real-time polymerase chain reactions for NFAT2 and PLZF were performed using SYBR Green master mix (Applied Biosystems) and for Blimp-1, Tbx21, IL-4, Eomes using Taqman probes. Actin or HPRT were used as endogenous control. Sequences of primers used for SyberGreen real-time PCR are NFAT2-F 5′-CCAGAAAATAACATGCGAGCC-3′, NFAT2-R 5′-GTGGGATGTGAACTCGGAAG-3′, PLZF-F 5′-GAGCAGTGCAGCGTGTGT-3′, and PLZF-R 5′-AACCGTTTTCCGCAGAGTT-3′ (44), Actin-F 5′-ATGGTGGGAATGGGTCAGAAG-3′, Actin-R 5′-TTCTCCATGTCGTCCCAGTTG-3′. All procedures were performed according to the manufacturers’ instructions.
In Vitro Cytotoxicity Assay
Target cells (P815) at 106/ml were labeled with 10-μM Calcein-AM (Molecular Probes) in complete medium for 30 min at 37°C. The assay was performed in V bottom 96-well microtiter plates with E:T ratios ranging from 2:1 to 0.5:1 with wells for spontaneous (only target cells in complete medium) and maximum release (only target cells in medium plus 2% Triton X-100) in the presence of 1 μg/ml of soluble anti-CD3, in triplicate. Each well contained from 1 × 105 to 2.5 × 104 lymphocytes in 100 μl of complete medium and 5 × 104 target cells/50 μl of complete medium. After incubation at 37°C in 5% CO2 for 4 h, 75 μl of each supernatant was harvested and transferred into new plates. Samples were measured using a Spectramax Gemini dual-scanning microplate spectrofluorimeter (Molecular Devices) (excitation: 485 nm; emission: 530). Data were expressed as arbitrary fluorescent units (AFU). Percent lysis was calculated with the following formula: [(A sample − A spontaneous)/(A max − A spontaneous)] × 100.
Statistical Analysis
For single comparisons, unpaired Student’s t-test was used. Flow cytometry and mean fluorescence intensity analysis was performed using FlowJo 10.1r5 software and analyzed for statistical significance by unpaired two-tailed Student’s t-test using Prism (GraphPad Software). Differences with p < 0.05 were considered statistically significant.
Results
NFAT2 Is Present and Functional in CD8+ T Lymphocytes
TCR stimulation results in calcium mobilization and consequent activation of NFAT transcription factors. NFAT is activated by the phosphatase calcineurin, which dephosphorylates multiple phosphoserines in the regulatory domain of NFAT, leading to NFAT nuclear translocation (45).
To evaluate the NFAT2 expression and activation, CD8+ T lymphocytes from WT mice were purified, left unstimulated, or stimulated in vitro with anti-CD3 plus anti-CD28 for different times. As expected, TCR stimulation increased both NFAT2 mRNA and protein level (Figures 1A,B). In unstimulated cells, NFAT2 was present in the cytoplasm in an inactive phosphorylated form (Figures 1C,D). At 24 h post-activation, we observed NFAT2 dephosphorylation (Figures 1B,C), and at 48 h post-activation, we detected NFAT2 in the nucleus (Figures 1C,D). These data show that NFAT2 protein is present and functional in CD8+ T lymphocytes.
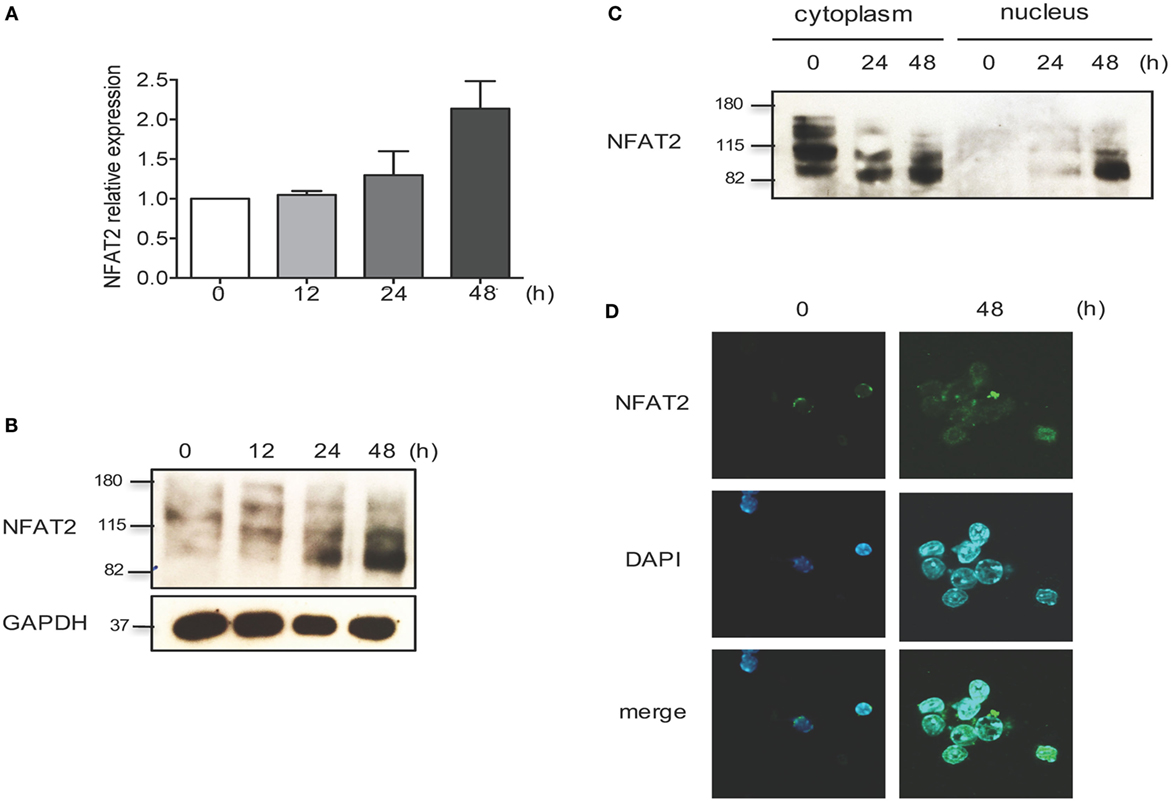
Figure 1. NFAT2 is present and functional in CD8+ T lymphocytes. CD8+ T lymphocytes were purified from naive C57BL/6 mice, as described, and then left unstimulated (0) or stimulated in vitro with anti-CD3 plus anti-CD28 (both at 1 μg/ml) for indicated times. (A) Total RNA was isolated, and NFAT2 mRNA levels were analyzed by real-time RT-PCR assay using SYBR green master mix. The data are normalized to the β-actin levels. Data are shown as mean ± SD of three independent experiments. Detection of NFAT2 transcription factor in CD8+ T cells in total lysates (B) and in cytoplasmic and nuclear fractions (C) by Western blot. (D) Cellular localization of NFAT2 protein in CD8+ T cells by immunofluorescence staining. All data are representative of at least two independent experiments.
Development of Innate-Like CD8+ T Cells in the Absence of NFAT2
To study how NFAT2 regulates CD8+ T cell development and responses, we used mice with T cell-specific deficiency in NFAT2. We used CD4-cre mice crossed to mice with NFAT2 flanked by loxP sites (NFAT2fl/fl). In this model, the NFAT2 gene is excised at the DP stage of thymocyte development, affecting both SP CD4 and CD8 mature T cells. The absence of NFAT2 protein expression in CD8+ T cells from NFAT2fl/flCD4-Cre mice was confirmed by Western blot (Figure 2A).
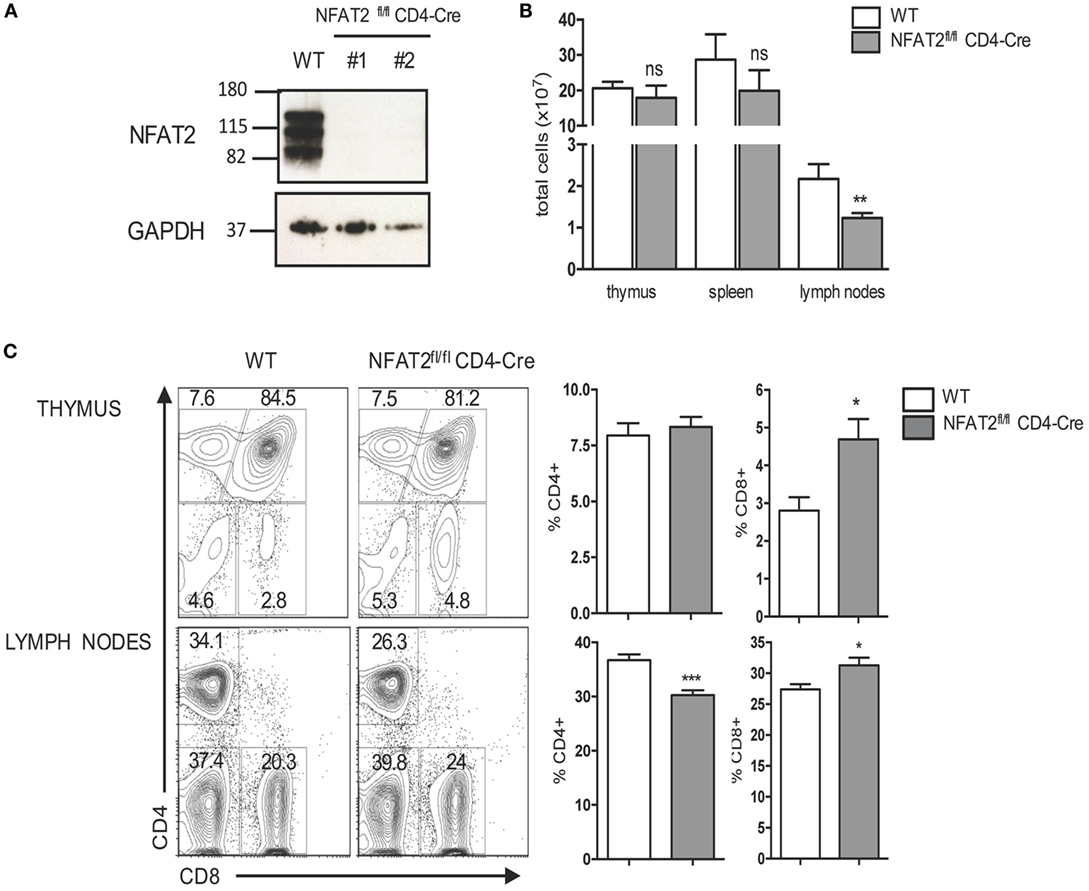
Figure 2. NFAT2 deficiency increases the frequency of CD8+ T cells. (A) Detection of NFAT2 transcription factor in CD8+ T cells’ total protein lysates from WT and NFAT2fl/flCD4-Cre mice by Western blot. (B) Thymus, spleens, and lymph nodes were harvested from WT or NFAT2fl/flCD4-Cre mice, and the total number of cells were counted using Trypan blue exclusion. (C) Flow cytometric analysis of thymocytes and total lymph nodes cells from WT or NFAT2fl/fl CD4-Cre mice. Shown is the frequency of CD4- and CD8-expressing cells. Right, the percentage of CD4+ and CD8+ cells for all the mice. All results are representative of at least two independent experiments. All data are shown as mean ± SD. The ns indicates not significant, *indicates p < 0.05, and **indicates p < 0.01 compared to WT mice.
NFAT2fl/flCD4-Cre mice showed only little changes in total cell numbers in the thymus and spleen, but a significant decrease of total cell number in lymph nodes (Figure 2B), and consistently NFAT2fl/flCD4-Cre mice had reduced size of lymph nodes (data not shown). Thymus of NFAT2fl/flCD4-Cre mice had similar frequency of DP as well as CD4+ single positive (SP) thymocytes as WT (Figure 2C). Interestingly, there was an increase in the frequency of CD8+ SP thymocytes (Figure 2C). To examine whether observed difference was thymus-specific or a general characteristic, we then analyzed spleen and lymph nodes of NFAT2fl/flCD4-Cre mice. The spleen of NFAT2fl/flCD4-Cre mice had significant reduction of the frequency of total T lymphocytes, in particular CD4+, and a slight increase of B lymphocytes, while no change was observed for CD8+ T cells (data not shown). Lymph nodes of NFAT2fl/flCD4-Cre mice showed significantly lower number of CD8+ T lymphocytes in NFAT2fl/flCD4-Cre mice compared to WT (data not shown), however, with only little changes in the frequency of total T and B lymphocytes. Similarly, to the spleen, we observed a decrease in the frequency of CD4+ T cells (Figure 2C), and as seen in the thymus, an increase in the frequency of CD8+ T lymphocytes (Figure 2C).
A higher proportion of CD8+ T cells in the thymus and lymph nodes of NFAT2fl/flCD4-Cre mice compared to WT was expressing CD44, an activated T cell marker. An increase in the frequency of CD8+CD44high cells was also observed in the spleen of NFAT2fl/flCD4-Cre mice even without a significant increase in the frequency of CD8+ T cells (Figure 3A). As the percentage of CD8+CD44high cells was increased in the thymus, lymph nodes, and spleen, it suggested that this is not dependent on preferential enrichment of these cells in specific tissues.
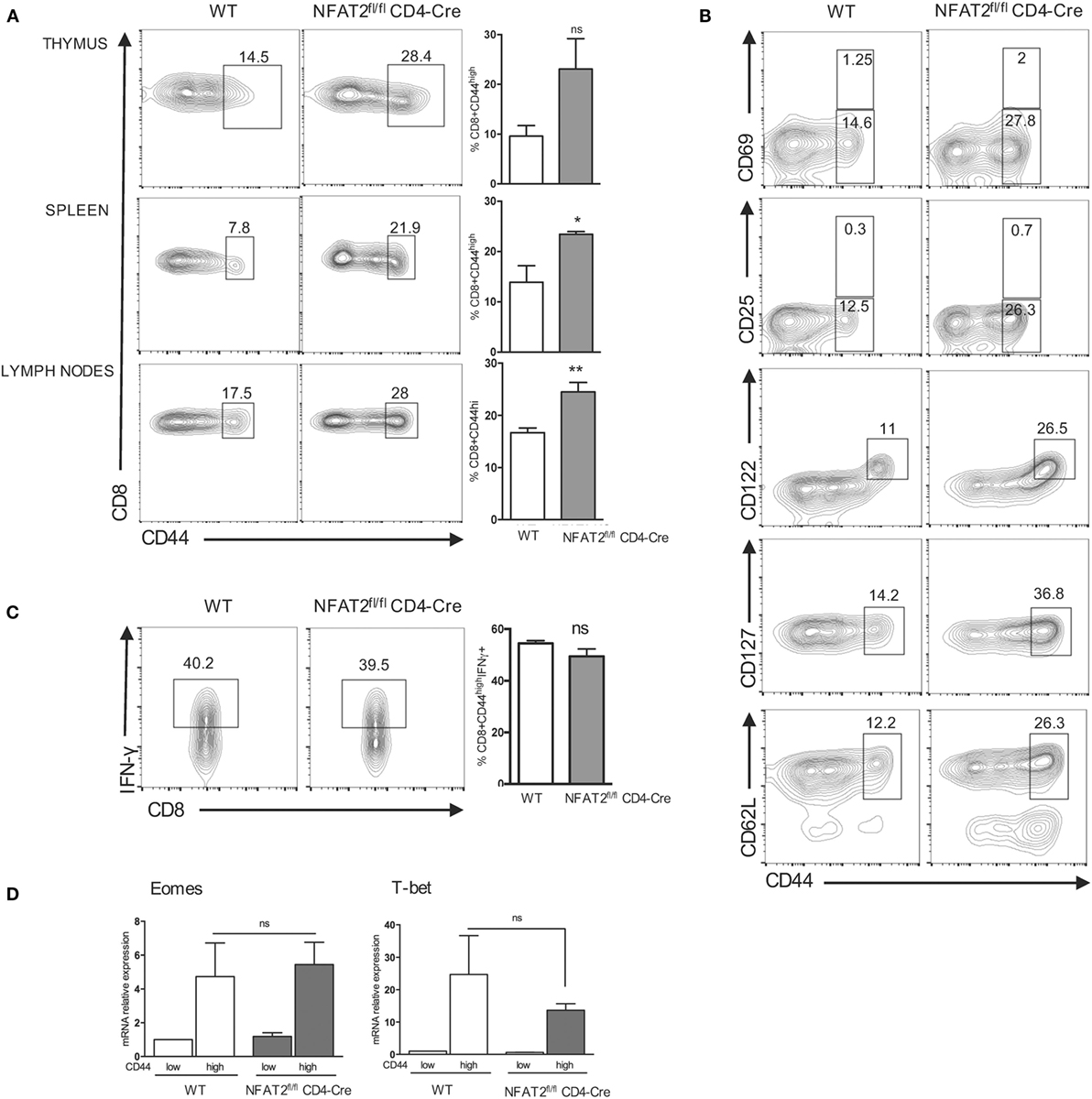
Figure 3. NFAT2 deficiency leads to the development of innate-like CD8+ T cells. (A) Flow cytometric analysis CD8+ T cells for CD44 expression. Right, the percentage of CD8+CD44high cells for all the mice. (B) Total CD8+ T cells were analyzed for the expression of CD69, CD25, CD122, CD127, and CD62L. Shown are the percentages of T CD8+CD44high populations expressing or not specific surface markers. (C) Total CD8+ T cells were stimulated with PMA plus ionomycin for 6 h, and cells in CD8+CD44high gate were analyzed for IFN-γ production. Right, the percentage of CD8+CD44highIFN-γ+ cells for all the mice. (D) Total RNA was isolated from CD8+CD44high and CD8+CD44low populations from lymph nodes of WT and NFAT2fl/flCD4-Cre mice, and Tbx21 and Eomes mRNA levels were analyzed by real-time RT-PCR assay using Taqman probe. The data are normalized to the HPRT levels. All results are representative of at least two independent experiments. All data are shown as mean ± SD. The ns indicates not significant, *indicates p < 0.05, and **indicates p < 0.01 compared to WT mice.
To exclude the fact that CD8+CD44high T cells are present due to an activation, we examined the expression of two activation markers: CD69 (early activation marker) and CD25 (IL-2 receptor α-chain). CD8+CD44high population from both WT and NFAT2fl/flCD4-Cre mice expressed low and similar percentage of CD69+ and CD25+ cells showing that CD8+CD44high population is not pre-activated in vivo (Figure 3B).
Expression of CD122, the IL-2Rβ chain, has been reported to define the population of innate-like CD8+ T cells (46), and we found that the CD8+CD44high population in both WT and NFAT2fl/fl CD4-Cre mice expressed CD122, although the NFAT2fl/flCD4-Cre mice carried larger percentage of these cells (Figure 3B).
Moreover, NFAT2fl/flCD4-Cre mice had larger percentage of CD8+CD44high CD127+ and CD8+CD44high CD62L+ compared to WT (Figure 3B), further showing that CD8+CD44high population display conventional memory T cell characteristics without previous antigen exposure.
Another characteristic of innate-like CD8+ T cells is the rapid production of large amount of IFN-γ upon stimulation (16). To determine if these innate-like CD8+ cells exhibit similar functional behavior, we analyzed their ability to rapidly secrete IFN-γ upon stimulation. Comparison of IFN-γ production by PMA plus ionomycin stimulated CD8+CD44high T cells in vitro indicated that NFAT2-deficient cells behave similarly to those from WT mice in rapid (within 6 h) secretion of this cytokine. A similar percentage of the CD8+CD44high T cells responded from both WT and NFAT2fl/flCD4-Cre mice (Figure 3C). By contrast, CD8+CD44low T cells did not secrete any appreciable IFN-γ in this time period (data not shown).
Further analysis of CD8+CD44high population revealed that cells from both NFAT2fl/flCD4-Cre and WT mice expressed elevated Tbx21 and Eomes mRNA level compared to respective CD8+CD44low counterparts. Eomes expression remained similar (Figure 3D), while Tbx21 mRNA was reduced without a significant difference in CD8+CD44high cells from NFAT2fl/flCD4-Cre animals compared to WT.
Taken together, these results demonstrate that NFAT2 deficiency leads to the development of innate-like CD8+ T cells without affecting theirs characteristics, such as surface markers and transcription factors expression and IFN-γ production.
NFAT2 Deficiency in T Cells Results in the Expansion of PLZF+ T Cells
Interleukin-4 is a cytokine classically associated with CD4+ T helper type 2 differentiation (47) but has been recently shown in different mice models to be also required for the development of innate-like CD8+ T lymphocytes through Eomes upregulation (9–11, 44, 48–50). To compare the IL-4 production between NFAT2fl/flCD4-Cre and WT mice, we stimulated WT and NFAT2-deficient thymocytes, splenocytes, and total lymph nodes cells ex vivo with PMA plus ionomycine for 6 h. We analyzed the production of IL-4 by CD3+ cells. We observed that NFAT2fl/flCD4-Cre mice had higher percentage of CD3+IL-4+ cells in the thymus and lymph nodes and only small increase in the spleen (Figure 4A). Consistently, IL-4 mRNA was clearly more abundant in the thymus, lymph nodes, and even the spleen of NFAT2fl/flCD4-Cre mice (Figure 4B).
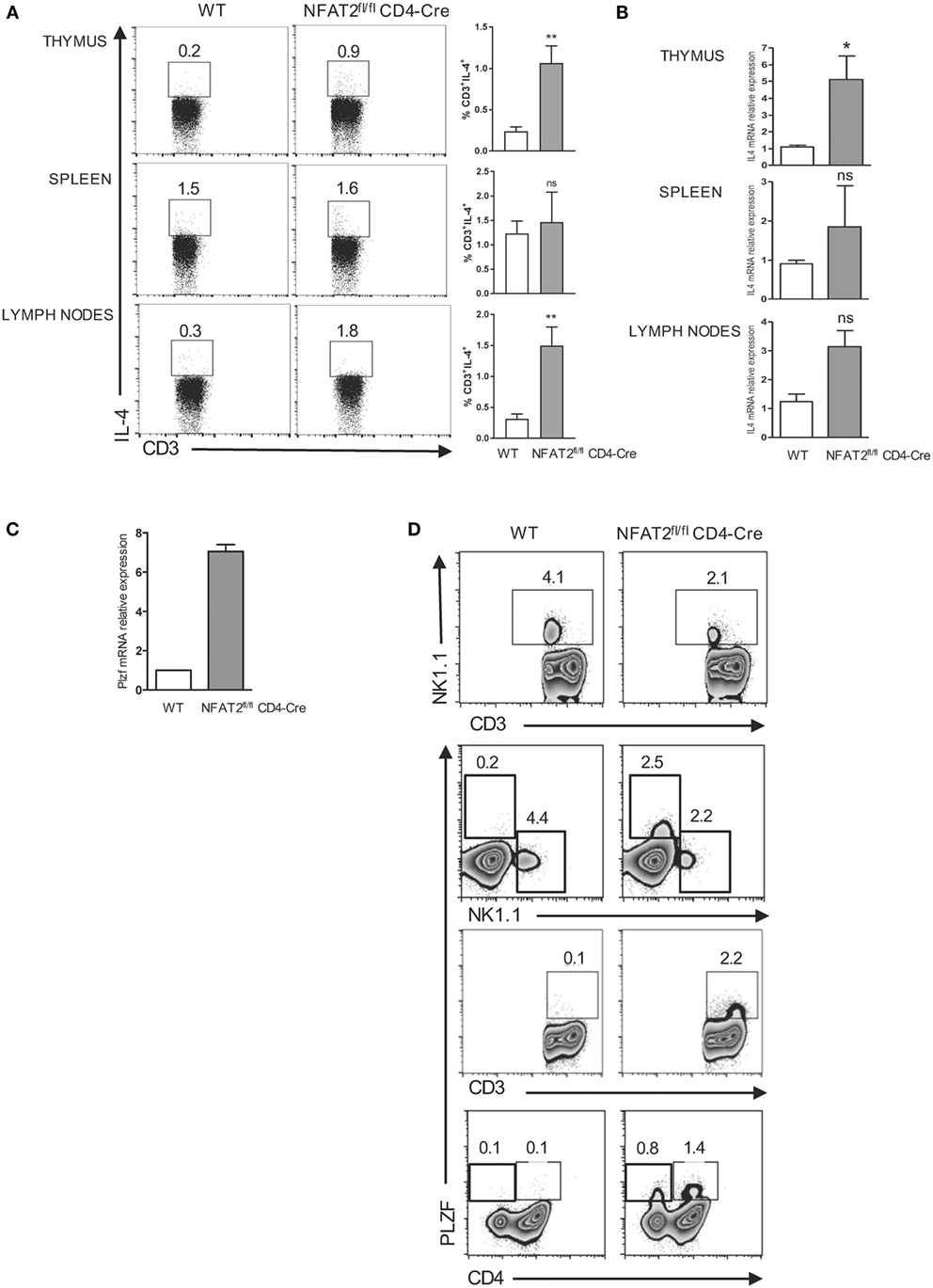
Figure 4. Expansion of PLZF expressing cells in the absence of NFAT2. Total thymocytes, splenocytes, and lymph nodes cells were stimulated with PMA plus ionomycin for 6 h. IL-4-producing cells were analyzed in CD3+ gate. Right, the percentage of CD3+IL-4+ cells for all the mice (A). Total RNA was isolated from thymus, spleen, and lymph nodes from WT and NFAT2fl/flCD4-Cre mice, and IL-4 mRNA levels were analyzed by real-time RT-PCR assay using Taqman probe. The data are normalized to the HPRT levels (B). Total RNA was isolated from CD4+ thymocytes and analyzed for PLZF mRNA level by real-time RT-PCR using SyberGreen. The data are normalized to the actin levels (C). Flow cytometric analysis of CD3+, CD4+, and NK1.1+ thymocytes for PLZF expression (D). All results are representative of at least two independent experiments. All data are shown as mean ± SD. The ns indicates not significant, *indicates p < 0.05, and **indicates p < 0.01 compared to WT mice.
In several gene-deficiency models, IL-4 is produced by an expanded population of T cells expressing the transcription factor PLZF. First, we tested whether NFAT2 absence affects the PLZF expression, by measuring the PLZF mRNA expression in the thymus by real-time PCR. Indeed, we found much higher PLZF mRNA level in NFAT2-deficient CD4+ thymocytes compared to WT (Figure 4C). Interestingly, in NFAT2fl/flCD4-Cre mice, we observed the reduction of CD3+ NK1.1+ as compared to WT. Further analysis showed that PLZF+ T cell population in NFAT2fl/flCD4-Cre mice was NK1.1−, CD4+, or CD4− (Figure 4D). Thus, the absence of NFAT2 results in the expansion of PLZF+, NK1.1−, CD4+, or CD4− expressing T cells and increases the IL-4 production in the thymus and the periphery, which, in consequence, may lead to the higher frequency of innate-like CD8+ T cells.
The Absence of NFAT2 Increases the Threshold of T Cell Responses
Nuclear factor of activated T cell transcription factors are ubiquitous regulators of gene expression during cellular activation; however, most reports are focused mainly on CD4+ T lymphocytes [for review, see Ref. (23)]. To evaluate the specific role of NFAT2 in CD8+ T cell activation, we purified CD8+ T lymphocytes from lymph nodes of WT and NFAT2fl/flCD4-Cre mice, and we left them either unstimulated or stimulated for 24 and 48 h with different doses of anti-CD3 plus 1 μg/ml of anti-CD28. As expected, more than 85% of unstimulated cells were positive for CD62L, a protein highly expressed on naive cells, and only few expressed CD69 and CD25 activation markers (data not shown). TCR stimulation with 1, 0.5, and 0.25 μg/ml of anti-CD3 for 24 h led to a reduction of both CD69 and CD25 expression in NFAT2-deficient cells compared to WT (Figure 5, upper panel). The dose of 0.1 μg/ml of anti-CD3 was not sufficient to activate CD8+ T cells (Figure 5). Interestingly, TCR stimulation with 1 and 0.5 μg/ml of anti-CD3 for 48 h resulted in a similar cell activation as shown by CD69 and CD25 expression between WT and NFAT2-deficient cells (Figure 5, lower panel), indicating that NFAT2 absence was compensated by another transcription factor, likely NFAT1. Only at lower anti-CD3 concentration (0.25 μg/ml), the cell activation of NFAT2-deficient cells remained significantly reduced in comparison to WT (Figure 5, lower panel), indicating that the absence of NFAT2 increases the threshold of CD8+ T cell activation at suboptimal TCR stimulation.
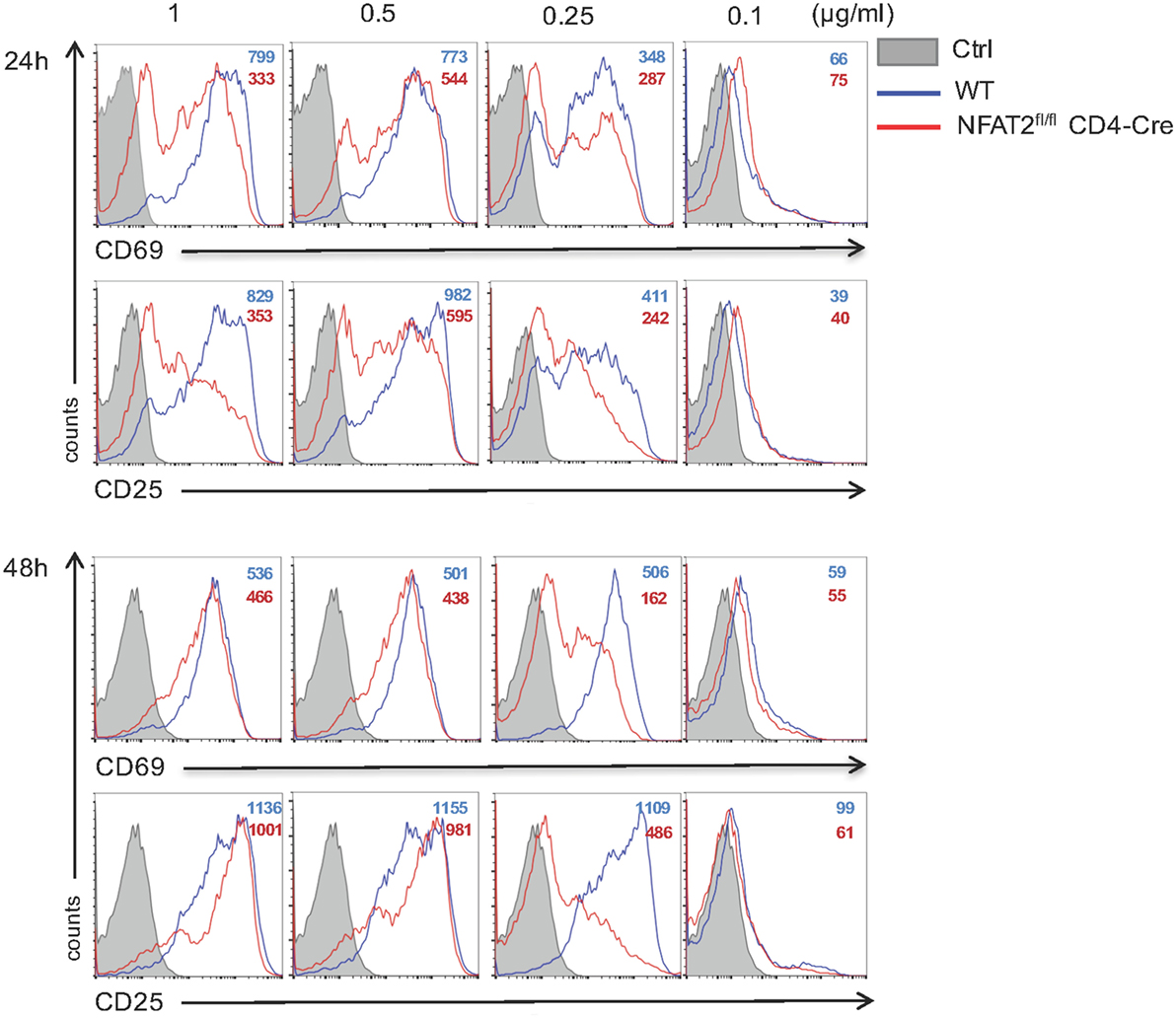
Figure 5. NFAT2 absence increases the threshold of CD8+ T cells activation. Total lymph nodes cells from WT and NFAT2fl/flCD4-Cre mice were activated for 24 and 48 h with different doses of anti-CD3 and 1 μg/ml of anti-CD28. Cells in CD8+ gate were analyzed for the expression of CD69 and CD25. All data are representative of at least three independent experiments.
As NFAT2 deficiency in CD8+ T cells increased the threshold of T cell activation, we next tested the effect of NFAT2 absence on CD8+ T cell proliferation. Purified CD8+ T cells were stained with CFSE and then activated for different times with 0.25 μg/ml anti-CD3 plus 1 μg/ml of anti-CD28. At indicated times, cell proliferation was monitored by flow cytometric measurement of CFSE dye dilution. Low concentration of anti-CD3 led to a reduced proliferation of NFAT2-deficient CD8+ T cells with division indexes of 3.19 for WT and 2.64 for NFAT2-deficient cells after 72 h, indicating that NFAT2 absence increases the threshold of triggering CD8+ T cell into division (Figure 6A).
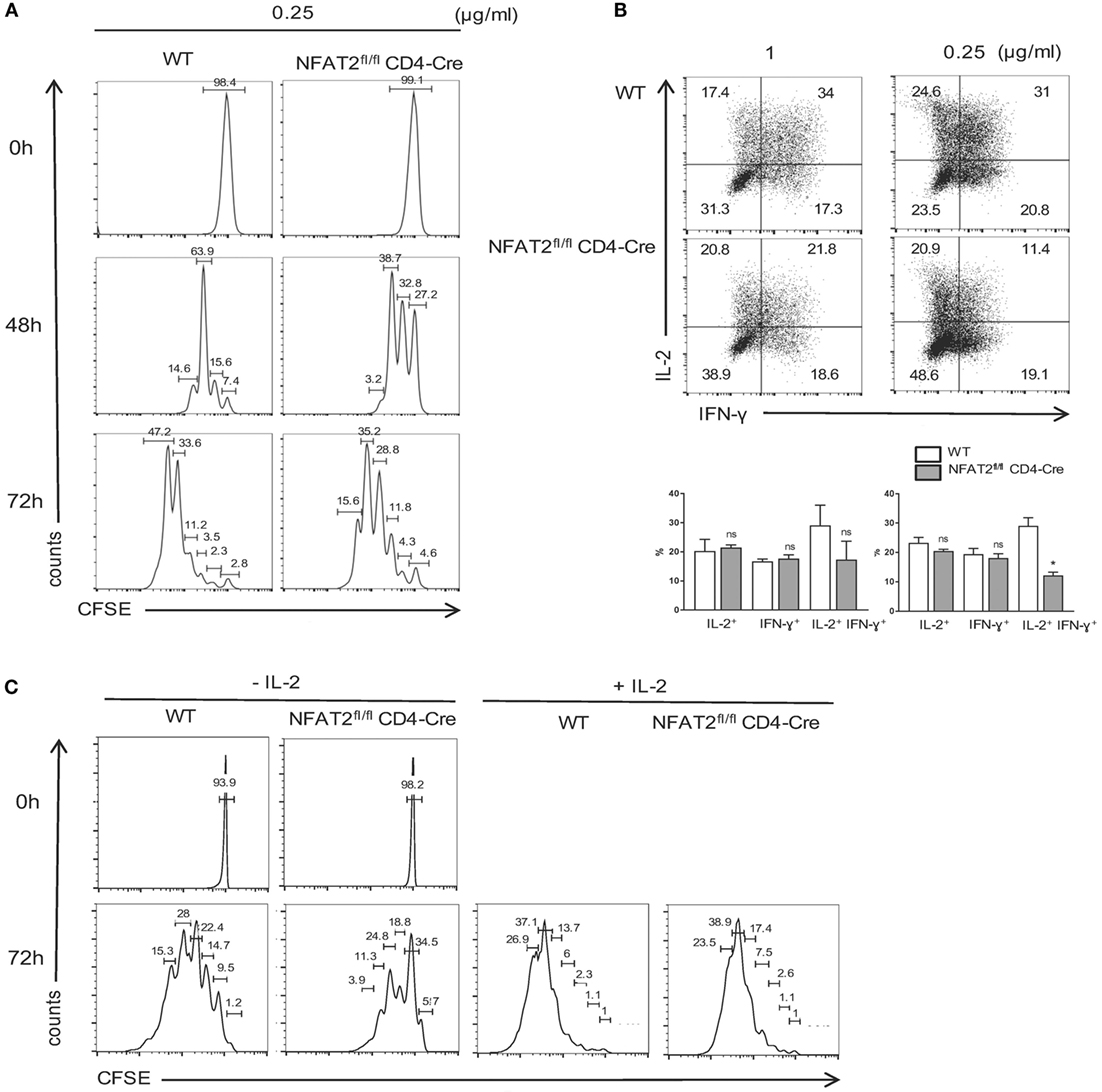
Figure 6. NFAT2 deficiency increases the threshold of CD8+ T cells proliferation and cytokines production. (A) CD8+ T cells from lymph nodes of WT and NFAT2fl/flCD4-Cre mice were purified, stained with CFSE as described, and activated with 0.25 μg/ml of anti-CD3 plus 1 μg/ml of anti-CD28 for indicated times. At each time point, CFSE dye dilution was analyzed by flow cytometry. (B) CD8+ T cells from lymph nodes of WT and NFAT2fl/flCD4-Cre mice were purified and activated for 48 h with either 1 or 0.25 μg/ml of anti-CD3 plus 1 μg/ml of anti-CD28. Then, cells were restimulated with PMA plus ionomycin for 6 h, and intracellular IL-2 and IFN-γ production was analyzed by flow cytometry. Shown is the percentage of IL-2- and IFN-γ-expressing cells. (C) CD8+ T cells from lymph nodes of WT and NFAT2fl/flCD4-Cre mice were purified, stained with CFSE as described, and activated with 0.25 μg/ml of anti-CD3 plus 1 μg/ml of anti-CD28 in the absence or presence of 200 U/ml IL-2 for indicated times. At each time point, CFSE dye dilution was analyzed by flow cytometry. All data are representative of at least two independent experiments.
Consistent with defects in cell activation and proliferation, NFAT2-deficient CD8+ T cells were also weak producers of both IFN-γ and IL-2. The slight difference was observed even at strong TCR stimulation, but clear decrease of the percentage of IFN-γ- and IL-2-producing cells was observed at suboptimal dose of anti-CD3 (Figure 6B). Next, we decided to determine whether defect of IL-2 production in NFAT2-deficient CD8+ T cells results in delayed cell proliferation. Purified CD8+ T cells were stained with CFSE and then activated for different times with 0.25 μg/ml anti-CD3 plus 1 μg/ml of anti-CD28 in the absence or presence of 200 U/ml of IL-2. At indicated times, cell proliferation was monitored by flow cytometric measurement of CFSE dye dilution. Addition of IL-2 restored the proliferation of NFAT2-deficient CD8+ T cells at 48 h (data not shown) and 72 h post-activation (Figure 6C) demonstrating that reduced proliferation in the absence of NFAT2 is due to decreased IL-2 secretion.
NFAT2 Regulates IFN-γ Production by Cytotoxic CD8+ T Cells
Antigen-induced CD8+ T cells differentiate into cytotoxic CD8+ T cells (CTLs) that kill infected cells and release cytokines, such as IFN-γ (1). To analyze if NFAT2-deficient CD8+ T cells have any other intrinsic defects besides increased threshold of activation, proliferation, and cytokine secretion, we determined if differentiation and function of CTLs in vitro were intact. To differentiate CTLs in vitro, we followed as reported (43). Briefly, purified CD8+ T cells from WT and NFAT2fl/flCD4-Cre mice were stimulated for 48 h with 1 μg/ml of anti-CD3 plus anti-CD28 and expanded daily with murine recombinant IL-2 (200 U/ml) (Figure 7A). At day 5, cells were collected and analyzed. The WT cytotoxic CD8+ T cells differentiated in vitro showed a downregulation of NFAT2 mRNA expression compared to naive CD8+ T cells (Figure 7B). No significant difference in cell viability and proliferation between WT and NFAT2-deficient CTLs was observed using the Trypan blue exclusion method (Figure 7C). The expression of activation markers, CD44 and CD25, was also comparable between WT and NFAT2-deficient CTLs (Figure 7D), indicating that NFAT2 does not influence either proliferation or activation of cytotoxic CD8+ T cells.
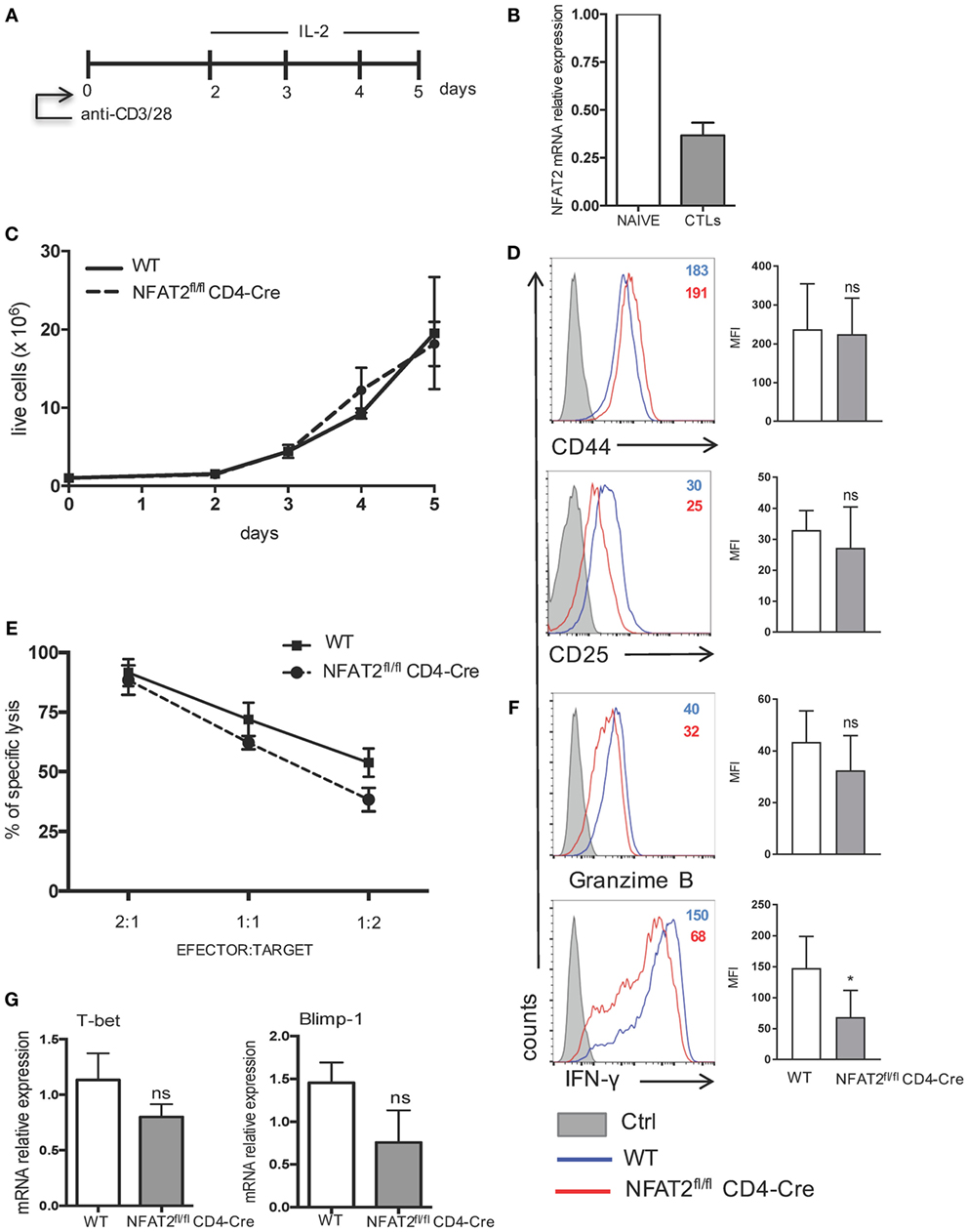
Figure 7. IFN-γ production by cytotoxic CD8+ T cells is regulated by NFAT2. (A) Schematic representation of cytotoxic CD8+ T cell differentiation in vitro. Purified CD8+ T cells from WT and NFAT2fl/flCD4-Cre mice were differentiated in vitro into cytotoxic CD8+ T lymphocytes as described. (B) Total RNA isolated from naive CD8+ T cells (NAIVE) or from in vitro differentiated cytotoxic CD8+ T lymphocytes (CTLs) were analyzed for NFAT2 mRNA levels by real-time RT-PCR assay using SYBR green master mix. The data are normalized to the β-actin levels. (C) Total number of live WT and NFAT2-deficient CD8+ T lymphocytes was recorded daily during in vitro differentiation using Trypan blue exclusion method. (D) Flow cytometric analysis of CD44 and CD25 expression on differentiated cytotoxic WT and NFAT2-deficient CD8+ T cells at day 5. The analysis of MFI from three independent experiments on the right. (E) At day 5, differentiated cytotoxic CD8+ T cells were tested in cytotoxicity assay against P815 cells as described at indicated efector:target ratios or restimulated with PMA plus ionomycin for 6 h, and intracellular granzime B and IFN-γ production was analyzed by flow cytometry. The analysis of MFI from three independent experiments on the right (F). (G) Total RNA was isolated from differentiated in vitro WT and NFAT2-deficient cytotoxic CD8+ T lymphocytes, and T-bet and Blimp-1 mRNA levels were analyzed by real-time RT-PCR assay using Taqman probes. The data are normalized to the HPRT RNA levels. All data are shown as mean ± SD of three independent experiments. The ns indicates not significant and *indicates p < 0.05 compared to WT mice.
CD8+ T cytotoxic activity was measured as the ability to effectively lyse P815 cells. NFAT2-deficient CTLs had slightly reduced potential to lyse P815 cells at 1:1 and 1:2 effector:target ratios (Figure 7E). Consistently, granzyme B expression was slightly decreased without statistical significance in NFAT2-deficient CTLs (Figure 7F). Remarkably, NFAT2-deficient CTLs produced significantly less IFN-γ compared to their proficient counterparts (Figure 7F).
Transcription factors T-bet and Blimp-1 are reported to play a crucial role in promoting cytotoxic CD8+ T cell differentiation (51, 52); therefore, we tested what is the effect of NFAT2 absence in CTLs on the expression of T-bet and Blimp-1. NFAT2-deficient CTLs had lower expression of both T-bet and Blimp-1, although in both cases, the difference remained insignificant (Figure 7G).
Taken together, these data demonstrate that NFAT2 does not significantly affect the differentiation of CD8+ T cells into cytotoxic T cells but positively controls the IFN-γ production.
Discussion
In this study, we characterized the role of NFAT2 in CD8+ T cell development and responses. During analysis of mice lacking NFAT2 in T cells, we noted that these mice in comparison to WT mice had an increased percentage of CD8+CD44highCD122+ cells in the thymus, spleen, and lymph nodes. Upon stimulation with PMA plus ionomycine, CD8+CD44high cells rapidly produced IFN-γ; however, in the same manner as WT CD8+CD44high indicating that NFAT2 is not essential for IFN-γ production. Rapid IFN-γ production by stimulated naive CD8+ T cells was reported to be dependent on NFAT1 (53), thus NFAT2 seems only to regulate the development of CD8+CD44highCD122+ cells.
CD8+CD44highCD122+ population is known as innate-like CD8+ T cells, has the phenotype of memory CD8+ T cells, and function as innate cells. In contrast to the so-called “true” or “conventional” memory cells that are induced by TCR stimulation with foreign antigen, this population is present in a steady state without contact with the foreign antigen. There have been numerous studies that have investigated innate-like cells in different experimental contexts. Mice deficient in or expressing mutant forms of diverse T-cell signaling molecules or transcription factors, including the scaffold protein SLP-76 (54), Itk (7, 55), the transcription factor KLF2 (Kruppel-like factor 2) (13, 44), the histone acetyltransferase CBP (CREB-binding protein), and the transcriptional regulator Id3 (inhibitor of DNA-binding 3) (12), also display an increased frequency and numbers of innate-like CD8+ T cells [reviewed in Ref. (56)]. Similar population of CD8+CD44high cells have been reported in mice lacking classical class I molecules, and this phenotype appears to be class 1b restricted (7, 57). Together, the similar phenotypes of these different mouse strains define a network of diverse effector proteins involved in a TCR signaling pathway to gene expression, which together specify increased generation of innate-like CD8+ T cells.
Based on the origin, CD8+CD44high cells are grouped in two populations. The first is called homeostatic memory or virtual memory that arises both in the lymphopenia and physiological environment in the periphery, in an IL-15-dependent manner (58). CD8+CD44high cells appear upon the transfer of naive T cells into lymphopenic mice (44). Consistently, patients recovering from bone marrow transplant, older people, and mice also showed elevated percentages of T cells with a similar phenotype (59–61). In addition, viral and other infections can lead to expanded populations of T cells bearing this phenotype (19).
The second group is IL-4-induced memory cells or innate-like CD8+ T cells identified in gene-deficient or mutant mice and also in normal mice (62). A common feature of these mice is the expansion of one or more subsets of PLZF+ thymocytes secreting IL-4, which in turn drive the expansion of the innate-like CD8+ T cells in a cell-extrinsic manner (10, 11, 44, 48). PLZF+ cells are rare or not activated to produce IL-4 in WT mice; however, distinct genetic alterations influence their generation and activation, and consequently increase innate-like CD8+ T cells presence. We show that NFAT2 deficiency in T cells results in the expansion of PLZF+ NK1.1−CD4+ and CD4− T cells. Consistently, we also observed increased IL-4 mRNA and IL-4 production by CD3+ cells in the thymus, spleen, and lymph nodes of NFAT2fl/flCD4-Cre mice. Therefore, innate-like CD8+ T cells development in NFAT2fl/flCD4-Cre mice seem to also be IL-4 dependent. To explain how NFAT2 exactly regulates the development of PLZF+ thymocytes and IL-4 production by those cells, further investigation is required.
Both virtual memory and innate CD8+ T cells are phenotypically and functionally very similar. They differ in cytokine dependence, IL-15 for virtual memory, and IL-4 for innate-like CD8+; origin, arising in the periphery vs. thymus, respectively, and differential expression profile of certain genes, such as Tnfarsf1a, Nfkb2, Eomes, and Bcl2l11 (63). Interestingly, virtual memory CD8+ T cells were also reported to develop in the presence of IL-4 overproduction in the periphery (64). It is still unclear whether these cell populations represent different subsets of alternative memory CD8+ T cells or if they are the same cell population that acquired its phenotype with different pathways. If we claim two distinct populations of alternative memory CD8+ T cells, it is possible that CD8+CD44high population in NFAT2fl/flCD4-Cre mice is heterogeneous and is comprised of both innate-like CD8+ T cells developed in the thymus and virtual memory CD8+ T cells developed in the periphery, both in IL-4-dependent manner.
In the absence of NFAT2, CD8+ T cells showed increased threshold for activation, proliferation, and cytokines production under suboptimal TCR stimulation. The fact that CD69 and CD25 activation markers were strongly reduced at 24 h post-activation and then at 48 h post-activation were similar to WT level indicates that NFAT2 deficiency has been compensated by other transcription factor, likely NFAT1. Only at suboptimal TCR stimulation, NFAT2 was crucial for complete CD8+ T cell responses.
Nuclear factor of activated T cells 2 deficiency resulted as well in the reduced capacity of CD8+ T cells to proliferate under suboptimal anti-CD3 concentration. Previous studies using NFAT2−/−/Rag-1−/− chimeric mice likewise reported impaired proliferation of NFAT2-deficient T cells (38) but independently of autocrine IL-2 secretion. In the EAE model, NFAT2 deficiency reduced the proliferation capacity of MOG-specific T cells in IL-2-dependent and -independent manners (41). In our model, CD8+ T cells produced less IL-2 under suboptimal TCR stimulation, and NFAT2 deficiency reduced proliferation in IL-2-dependent manner.
Nuclear factor of activated T cells 2 is not required for the cytotoxic activity of CD8+ T cells in vitro but appears to be important for their IFN-γ production. IFN-γ production by cytotoxic CD8+ T cells is regulated by T-bet and Blimp-1 transcription factors. There was only a slight decrease in T-bet expression but clear reduction of Blimp-1 expression in the absence of NFAT2. Whether NFAT2 influences the IFN-γ expression by directly binding to the IFN-γ promoter or the reduction of IFN-γ protein expression is secondary to the downregulation of Blimp-1, and/or T-bet transcription factors have to be further investigated. Similar defect in IFN-γ production was also found in CD8+ T cytotoxic cells in EAE model in vivo (41). NFAT2−/−/Rag-1−/− chimeric mice developed comparable cytotoxic T lymphocyte responses to the virus indicating that NFAT2 is not essential for the helper function in vivo (38).
In conclusion, we demonstrate in this study that NFAT2 is involved in several aspects of CD8+ T cell biology. In regard to T cell development, NFAT2 is a negative regulator of innate-like CD8+ T cells and PLZF+ expressing T cells development. What concerns CD8+ T cell responses upon TCR stimulation, we show that the absence of NFAT2 increases the threshold of CD8+ T cell activation and decreases IFN-γ and IL-2 production and thus delays cell proliferation in an IL-2-dependent manner. Furthermore, we show that although NFAT2 does not impact cytotoxic CD8+ T cells differentiation in vitro, it is clearly required for IFN-γ production. Finally, we show that NFAT2 regulates both innate immune response via innate-like CD8+ T cells and adaptive immune response by controlling IFN-γ production by effector CD8+ T lymphocytes.
Author Contributions
EP performed most of the experiments, analyzed the data, contributed to experimental design, and wrote the manuscript; VN-M performed some of the experiments, participated in the discussions, and helped with the CD8+ T lymphocytes differentiation and in vitro cytotoxicity assay; JV designed and was responsible for the study, assisted with data analysis, and wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are especially grateful to Dr. Anjana Rao from the La Jolla Institute for Allergy and Immunology (San Diego, CA, USA) for kindly providing the NFAT2fl/flCD4-Cre mice.
Funding
This work was supported by grants to JV from the CNPq (307296/2011-3 and 476314/2012-7), FAPERJ (112.056/2012, 102.308/2013, 110.794/2013, and 101.147/2013), and INCT-Cancer (573806/2008-0 and 170.026/2008). EP was supported by a CNPq and Brazilian Ministry of Health fellowships, and VN-M was supported by a Brazilian Ministry of Health fellowship.
References
1. Zhang N, Bevan MJ. CD8(+) T cells: foot soldiers of the immune system. Immunity (2011) 35:161–8. doi:10.1016/j.immuni.2011.07.010
2. Sallusto F, Geginat J, Lanzavecchia A. Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu Rev Immunol (2004) 22:745–63. doi:10.1146/annurev.immunol.22.012703.104702
3. Takemoto N, Intlekofer AM, Northrup JT, Wherry EJ, Reiner SL. Cutting edge: IL-12 inversely regulates T-bet and eomesodermin expression during pathogen-induced CD8+ T cell differentiation. J Immunol (2006) 177:7515–9. doi:10.4049/jimmunol.177.11.7515
4. Prlic M, Williams MA, Bevan MJ. Requirements for CD8 T-cell priming, memory generation and maintenance. Curr Opin Immunol (2007) 19:315–9. doi:10.1016/j.coi.2007.04.010
5. Prince AL, Yin CC, Enos ME, Felices M, Berg LJ. The Tec kinases Itk and Rlk regulate conventional versus innate T-cell development. Immunol Rev (2009) 228:115–31. doi:10.1111/j.1600-065X.2008.00746.x
6. Readinger JA, Mueller KL, Venegas AM, Horai R, Schwartzberg PL. Tec kinases regulate T-lymphocyte development and function: new insights into the roles of Itk and Rlk/Txk. Immunol Rev (2009) 228:93–114. doi:10.1111/j.1600-065X.2008.00757.x
7. Broussard C, Fleischacker C, Horai R, Chetana M, Venegas AM, Sharp LL, et al. Altered development of CD8+ T cell lineages in mice deficient for the Tec kinases Itk and Rlk. Immunity (2006) 25:93–104. doi:10.1016/j.immuni.2006.05.011
8. Fukuyama T, Kasper LH, Boussouar F, Jeevan T, van Deursen J, Brindle PK. Histone acetyltransferase CBP is vital to demarcate conventional and innate CD8+ T-cell development. Mol Cell Biol (2009) 29:3894–904. doi:10.1128/MCB.01598-08
9. Lai D, Zhu J, Wang T, Hu-Li J, Terabe M, Berzofsky JA, et al. KLF13 sustains thymic memory-like CD8(+) T cells in BALB/c mice by regulating IL-4-generating invariant natural killer T cells. J Exp Med (2011) 208:1093–103. doi:10.1084/jem.20101527
10. Sharma A, Chen Q, Nguyen T, Yu Q, Sen JM. T cell factor-1 and beta-catenin control the development of memory-like CD8 thymocytes. J Immunol (2012) 188:3859–68. doi:10.4049/jimmunol.1103729
11. Verykokakis M, Boos MD, Bendelac A, Adams EJ, Pereira P, Kee BL. Inhibitor of DNA binding 3 limits development of murine slam-associated adaptor protein-dependent “innate” gammadelta T cells. PLoS One (2010) 5:e9303. doi:10.1371/journal.pone.0009303
12. Verykokakis M, Boos MD, Bendelac A, Kee BL. SAP protein-dependent natural killer T-like cells regulate the development of CD8(+) T cells with innate lymphocyte characteristics. Immunity (2010) 33:203–15. doi:10.1016/j.immuni.2010.07.013
13. Weinreich MA, Takada K, Skon C, Reiner SL, Jameson SC, Hogquist KA. KLF2 transcription-factor deficiency in T cells results in unrestrained cytokine production and upregulation of bystander chemokine receptors. Immunity (2009) 31:122–30. doi:10.1016/j.immuni.2009.05.011
14. Hu J, Sahu N, Walsh E, August A. Memory phenotype CD8+ T cells with innate function selectively develop in the absence of active Itk. Eur J Immunol (2007) 37:2892–9. doi:10.1002/eji.200737311
15. Jacomet F, Cayssials E, Basbous S, Levescot A, Piccirilli N, Desmier D, et al. Evidence for eomesodermin-expressing innate-like CD8(+) KIR/NKG2A(+) T cells in human adults and cord blood samples. Eur J Immunol (2015) 45:1926–33. doi:10.1002/eji.201545539
16. Kambayashi T, Assarsson E, Lukacher AE, Ljunggren HG, Jensen PE. Memory CD8+ T cells provide an early source of IFN-gamma. J Immunol (2003) 170:2399–408. doi:10.4049/jimmunol.170.5.2399
17. Berg RE, Cordes CJ, Forman J. Contribution of CD8+ T cells to innate immunity: IFN-gamma secretion induced by IL-12 and IL-18. Eur J Immunol (2002) 32:2807–16. doi:10.1002/1521-4141(2002010)32:10<2807::AID-IMMU2807>3.0.CO;2-0
18. Berg RE, Forman J. The role of CD8 T cells in innate immunity and in antigen non-specific protection. Curr Opin Immunol (2006) 18:338–43. doi:10.1016/j.coi.2006.03.010
19. Braaten DC, McClellan JS, Messaoudi I, Tibbetts SA, McClellan KB, Nikolich-Zugich J, et al. Effective control of chronic gamma-herpesvirus infection by unconventional MHC class Ia-independent CD8 T cells. PLoS Pathog (2006) 2:e37. doi:10.1371/journal.ppat.0020037
20. Oghumu S, Terrazas CA, Varikuti S, Kimble J, Vadia S, Yu L, et al. CXCR3 expression defines a novel subset of innate CD8+ T cells that enhance immunity against bacterial infection and cancer upon stimulation with IL-15. FASEB J (2015) 29:1019–28. doi:10.1096/fj.14-264507
21. Shaw JP, Utz PJ, Durand DB, Toole JJ, Emmel EA, Crabtree GR. Identification of a putative regulator of early T cell activation genes. Science (1988) 241:202–5. doi:10.1126/science.3260404
22. Rao A, Luo C, Hogan PG. Transcription factors of the NFAT family: regulation and function. Annu Rev Immunol (1997) 15:707–47. doi:10.1146/annurev.immunol.15.1.707
23. Macian F. NFAT proteins: key regulators of T-cell development and function. Nat Rev Immunol (2005) 5:472–84. doi:10.1038/nri1632
24. Northrop JP, Ho SN, Chen L, Thomas DJ, Timmerman LA, Nolan GP, et al. NF-AT components define a family of transcription factors targeted in T-cell activation. Nature (1994) 369:497–502. doi:10.1038/369497a0
25. Beals CR, Clipstone NA, Ho SN, Crabtree GR. Nuclear localization of NF-ATc by a calcineurin-dependent, cyclosporin-sensitive intramolecular interaction. Genes Dev (1997) 11:824–34. doi:10.1101/gad.11.7.824
26. Flanagan WM, Corthesy B, Bram RJ, Crabtree GR. Nuclear association of a T-cell transcription factor blocked by FK-506 and cyclosporin A. Nature (1991) 352:803–7. doi:10.1038/352803a0
27. Loh C, Shaw KT, Carew J, Viola JP, Luo C, Perrino BA, et al. Calcineurin binds the transcription factor NFAT1 and reversibly regulates its activity. J Biol Chem (1996) 271:10884–91. doi:10.1074/jbc.271.18.10884
28. McCaffrey PG, Perrino BA, Soderling TR, Rao A. NF-ATp, a T lymphocyte DNA-binding protein that is a target for calcineurin and immunosuppressive drugs. J Biol Chem (1993) 268:3747–52.
29. Liu J, Koyano-Nakagawa N, Amasaki Y, Saito-Ohara F, Ikeuchi T, Imai S, et al. Calcineurin-dependent nuclear translocation of a murine transcription factor NFATx: molecular cloning and functional characterization. Mol Biol Cell (1997) 8:157–70. doi:10.1091/mbc.8.1.157
30. Shibasaki F, Price ER, Milan D, McKeon F. Role of kinases and the phosphatase calcineurin in the nuclear shuttling of transcription factor NF-AT4. Nature (1996) 382:370–3. doi:10.1038/382370a0
31. Timmerman LA, Clipstone NA, Ho SN, Northrop JP, Crabtree GR. Rapid shuttling of NF-AT in discrimination of Ca2+ signals and immunosuppression. Nature (1996) 383:837–40. doi:10.1038/383837a0
32. Jain J, Valge-Archer VE, Sinskey AJ, Rao A. The AP-1 site at -150 bp, but not the NF-kappa B site, is likely to represent the major target of protein kinase C in the interleukin 2 promoter. J Exp Med (1992) 175:853–62. doi:10.1084/jem.175.3.853
33. Hodge MR, Ranger AM, Charles de la Brousse F, Hoey T, Grusby MJ, Glimcher LH. Hyperproliferation and dysregulation of IL-4 expression in NF-ATp-deficient mice. Immunity (1996) 4:397–405. doi:10.1016/S1074-7613(00)80253-8
34. Mehta DS, Wurster AL, Weinmann AS, Grusby MJ. NFATc2 and T-bet contribute to T-helper-cell-subset-specific regulation of IL-21 expression. Proc Natl Acad Sci U S A (2005) 102:2016–21. doi:10.1073/pnas.0409512102
35. de la Pompa JL, Timmerman LA, Takimoto H, Yoshida H, Elia AJ, Samper E, et al. Role of the NF-ATc transcription factor in morphogenesis of cardiac valves and septum. Nature (1998) 392:182–6. doi:10.1038/32419
36. Peng SL, Gerth AJ, Ranger AM, Glimcher LH. NFATc1 and NFATc2 together control both T and B cell activation and differentiation. Immunity (2001) 14:13–20. doi:10.1016/S1074-7613(01)00085-1
37. Ranger AM, Hodge MR, Gravallese EM, Oukka M, Davidson L, Alt FW, et al. Delayed lymphoid repopulation with defects in IL-4-driven responses produced by inactivation of NF-ATc. Immunity (1998) 8:125–34. doi:10.1016/S1074-7613(00)80465-3
38. Yoshida H, Nishina H, Takimoto H, Marengere LE, Wakeham AC, Bouchard D, et al. The transcription factor NF-ATc1 regulates lymphocyte proliferation and Th2 cytokine production. Immunity (1998) 8:115–24. doi:10.1016/S1074-7613(00)80464-1
39. Gomez-Rodriguez J, Sahu N, Handon R, Davidson TS, Anderson SM, Kirby MR, et al. Differential expression of interleukin-17A and -17F is coupled to T cell receptor signaling via inducible T cell kinase. Immunity (2009) 31:587–97. doi:10.1016/j.immuni.2009.07.009
40. Lee Y, Awasthi A, Yosef N, Quintana FJ, Xiao S, Peters A, et al. Induction and molecular signature of pathogenic TH17 cells. Nat Immunol (2012) 13:991–9. doi:10.1038/ni.2416
41. Reppert S, Zinser E, Holzinger C, Sandrock L, Koch S, Finotto S. NFATc1 deficiency in T cells protects mice from experimental autoimmune encephalomyelitis. Eur J Immunol (2015) 45:1426–40. doi:10.1002/eji.201445150
42. Vaeth M, Schliesser U, Muller G, Reissig S, Satoh K, Tuettenberg A, et al. Dependence on nuclear factor of activated T-cells (NFAT) levels discriminates conventional T cells from Foxp3+ regulatory T cells. Proc Natl Acad Sci U S A (2012) 109:16258–63. doi:10.1073/pnas.1203870109
43. Pipkin ME, Sacks JA, Cruz-Guilloty F, Lichtenheld MG, Bevan MJ, Rao A. Interleukin-2 and inflammation induce distinct transcriptional programs that promote the differentiation of effector cytolytic T cells. Immunity (2010) 32:79–90. doi:10.1016/j.immuni.2009.11.012
44. Weinreich MA, Odumade OA, Jameson SC, Hogquist KA. T cells expressing the transcription factor PLZF regulate the development of memory-like CD8+ T cells. Nat Immunol (2010) 11:709–16. doi:10.1038/ni.1898
45. Hogan PG, Lewis RS, Rao A. Molecular basis of calcium signaling in lymphocytes: STIM and ORAI. Annu Rev Immunol (2010) 28:491–533. doi:10.1146/annurev.immunol.021908.132550
46. Walzer T, Arpin C, Beloeil L, Marvel J. Differential in vivo persistence of two subsets of memory phenotype CD8 T cells defined by CD44 and CD122 expression levels. J Immunol (2002) 168:2704–11. doi:10.4049/jimmunol.168.6.2704
47. Paul WE. What determines Th2 differentiation, in vitro and in vivo? Immunol Cell Biol (2010) 88:236–9. doi:10.1038/icb.2010.2
48. Min HS, Lee YJ, Jeon YK, Kim EJ, Kang BH, Jung KC, et al. MHC class II-restricted interaction between thymocytes plays an essential role in the production of innate CD8+ T cells. J Immunol (2011) 186:5749–57. doi:10.4049/jimmunol.1002825
49. Gordon SM, Carty SA, Kim JS, Zou T, Smith-Garvin J, Alonzo ES, et al. Requirements for eomesodermin and promyelocytic leukemia zinc finger in the development of innate-like CD8+ T cells. J Immunol (2011) 186:4573–8. doi:10.4049/jimmunol.1100037
50. Huang W, Huang F, Kannan AK, Hu J, August A. ITK tunes IL-4-induced development of innate memory CD8+ T cells in a gammadelta T and invariant NKT cell-independent manner. J Leukoc Biol (2014) 96:55–63. doi:10.1189/jlb.1AB0913-484RR
51. Rutishauser RL, Martins GA, Kalachikov S, Chandele A, Parish IA, Meffre E, et al. Transcriptional repressor Blimp-1 promotes CD8(+) T cell terminal differentiation and represses the acquisition of central memory T cell properties. Immunity (2009) 31:296–308. doi:10.1016/j.immuni.2009.05.014
52. Joshi NS, Cui W, Chandele A, Lee HK, Urso DR, Hagman J, et al. Inflammation directs memory precursor and short-lived effector CD8(+) T cell fates via the graded expression of T-bet transcription factor. Immunity (2007) 27:281–95. doi:10.1016/j.immuni.2007.07.010
53. Teixeira LK, Fonseca BP, Vieira-de-Abreu A, Barboza BA, Robbs BK, Bozza PT, et al. IFN-gamma production by CD8+ T cells depends on NFAT1 transcription factor and regulates Th differentiation. J Immunol (2005) 175:5931–9. doi:10.4049/jimmunol.175.9.5931
54. Jordan MS, Smith JE, Burns JC, Austin JE, Nichols KE, Aschenbrenner AC, et al. Complementation in trans of altered thymocyte development in mice expressing mutant forms of the adaptor molecule SLP76. Immunity (2008) 28:359–69. doi:10.1016/j.immuni.2008.01.010
55. Horai R, Mueller KL, Handon RA, Cannons JL, Anderson SM, Kirby MR, et al. Requirements for selection of conventional and innate T lymphocyte lineages. Immunity (2007) 27:775–85. doi:10.1016/j.immuni.2007.09.012
56. Lee YJ, Jameson SC, Hogquist KA. Alternative memory in the CD8 T cell lineage. Trends Immunol (2011) 32:50–6. doi:10.1016/j.it.2010.12.004
57. Su J, Berg RE, Murray S, Forman J. Thymus-dependent memory phenotype CD8 T cells in naive B6.H-2Kb-/-Db-/- animals mediate an antigen-specific response against Listeria monocytogenes. J Immunol (2005) 175:6450–7. doi:10.4049/jimmunol.175.10.6450
58. White JT, Cross EW, Burchill MA, Danhorn T, McCarter MD, Rosen HR, et al. Virtual memory T cells develop and mediate bystander protective immunity in an IL-15-dependent manner. Nat Commun (2016) 7:11291. doi:10.1038/ncomms11291
59. Ernst DN, Weigle WO, Noonan DJ, McQuitty DN, Hobbs MV. The age-associated increase in IFN-gamma synthesis by mouse CD8+ T cells correlates with shifts in the frequencies of cell subsets defined by membrane CD44, CD45RB, 3G11, and MEL-14 expression. J Immunol (1993) 151:575–87.
60. Jackola DR, Ruger JK, Miller RA. Age-associated changes in human T cell phenotype and function. Aging (Milano) (1994) 6:25–34.
61. Stachel D, Futterer A, Haas RJ, Schmid I. Enhanced lymphocyte proliferation responses in pediatric patients early after myelosuppressive chemotherapy. Pediatr Blood Cancer (2004) 43:644–50. doi:10.1002/pbc.20140
62. Dobber R, Hertogh-Huijbregts A, Rozing J, Bottomly K, Nagelkerken L. The involvement of the intestinal microflora in the expansion of CD4+ T cells with a naive phenotype in the periphery. Dev Immunol (1992) 2:141–50. doi:10.1155/1992/57057
63. Huang W, Hu J, August A. Cutting edge: innate memory CD8+ T cells are distinct from homeostatic expanded CD8+ T cells and rapidly respond to primary antigenic stimuli. J Immunol (2013) 190:2490–4. doi:10.4049/jimmunol.1202988
Keywords: NFAT2, CD8+ T lymphocytes, innate-like CD8+ T cells, PLZF, IFN-γ
Citation: Pachulec E, Neitzke-Montinelli V and Viola JPB (2016) NFAT2 Regulates Generation of Innate-Like CD8+ T Lymphocytes and CD8+ T Lymphocytes Responses. Front. Immunol. 7:411. doi: 10.3389/fimmu.2016.00411
Received: 26 May 2016; Accepted: 22 September 2016;
Published: 06 October 2016
Edited by:
Loretta Tuosto, Sapienza University of Rome, ItalyReviewed by:
Francesca Granucci, University of Milano-Bicocca, ItalySho Yamasaki, Kyushu University, Japan
Avery August, Cornell University, USA
Copyright: © 2016 Pachulec, Neitzke-Montinelli and Viola. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: João P. B. Viola, anB2aW9sYUBpbmNhLmdvdi5icg==