 María Luisa Pita-López
María Luisa Pita-López Alejandra Pera
Alejandra Pera Rafael Solana
Rafael Solana- 1Research Center in Molecular Biology of Chronic Diseases (CIBIMEC), CUSUR University of Guadalajara, Guzmán, Mexico
- 2Clinical Division, Brighton and Sussex Medical School, University of Sussex, Brighton, UK
- 3Maimonides Biomedicine Institute of Cordoba (IMIBIC), Reina Sofia Hospital, University of Córdoba, Córdoba, Spain
Human natural killer (NK)-like CD8+ T-cells are singular T-cells that express both T and NK cell markers such as CD56; their frequencies depend on their differentiation and activation during their lifetime. There is evidence of the presence of these innate CD8+ T-cells in the human umbilical cord, highlighting the necessity of investigating whether the NK-like CD8+ T-cells arise in the early stages of life (gestation). Based on the presence of cell surface markers, these cells have also been referred to as CD8+KIR+ T-cells, innate CD8+ T-cells, CD8+CD28−KIR+ T-cells or NKT-like CD8+CD56+ cells. However, the functional and co-signaling significance of these NK cell receptors on NK-like CD8+ T-cells is less clear. Also, the diverse array of costimulatory and co-inhibitory receptors are spatially and temporally regulated and may have distinct overlapping functions on NK-like CD8+ T-cell priming, activation, differentiation, and memory responses associated with different cell phenotypes. Currently, there is no consensus regarding the functional properties and phenotypic characterization of human NK-like CD8+ T-cells. Environmental factors, such as aging, autoimmunity, inflammation, viral antigen re-exposure, or the presence of persistent tumor antigens have been shown to allow differentiation (“adaptation”) of the NK-like CD8+ T-cells; the elucidation of this differentiation process and a greater understanding of the characteristics of these cells could be important for their eventual in potential therapeutic applications aimed at improving protective immunity. This review will attempt to elucidate an understanding of the characteristics of these cells with the goal toward their eventual use in potential therapeutic applications aimed at improving protective immunity.
Introduction
T lymphocytes derive their name from their site of maturation in the thymus. In particular, T cytotoxic (Tc) cells that express CD8 are activated upon interaction with an MHC-class I complex on the surface of an altered-self cell (e.g., virus-infected cell or tumor cell) in the presence of appropriate cytokines. T-cell co-signaling is largely context dependent and relies on a diverse array of costimulatory and co-inhibitory receptors spatiotemporally regulated, which may have distinct or overlapping functions in T-cell priming, activation, differentiation, and memory responses (1). The total cytotoxic CD8+ T-cell pool is exposed to different microenvironmental stimuli (both TcR dependent and independent) and the resulting phenotype and cytokine secretion will determine an individual T-cell or T-cell clone’s effector or regulating functional capacities, including tissue residence/homing and organ homeostasis (2).
In addition to CD8+ T lymphocytes, natural killer (NK) cells have a crucial role in the recognition and killing of virus-infected/tumor cells, but unlike CD8+ T-cells, they use a repertoire of germ-line encoded inhibitory/activating receptors that recognize “missing self”/“altered-self” antigens on the target cells leading to cytotoxicity and cytokine production (3). These NK cell receptors (NKRs) are also expressed on certain subsets of T-cells. One example is NKR-CD56, which has been found to be elevated in both peripheral blood cells and in tumor-infiltrating lymphocytes in patients with colorectal cancer (4). In many clinical circumstances, the expression of different NKRs on T-cells is associated with prolonged antigen stimulation, suggesting that these receptors play a crucial role in the homeostasis of antigen-experienced T-cells.
Cumulative evidence supports the existence of T-cell subsets, with characteristics that bridge innate and adaptive immunity, which are relevant in inflammation and viral and tumor surveillance, and which could have a role in the pathogenesis of autoimmune diseases. These NKR-expressing cytotoxic T lymphocytes (CTL) have been termed NK T (NKT) cells. Thus, NKT cells are naturally occurring, although rare, T-cells that express both T and NK cell receptors (5). However, there is some confusion with the use of the term “NKT-cell.” On one hand, CD1d-restricted cells, which have a semi-invariant TcR, are frequently called NKT-cells or invariant NKT (iNKT) cells; on the other hand, highly specialized effector memory CD8+ T-cells expressing NKRs are also referred as NKT-like cells. Therefore, to avoid confusion, we will call the later, NK-like CD8+ T-cells.
Natural killer-like CD8+ T-cell differentiation occurs after the induction of transduction signals that activate/inhibit the expression of certain CD8+ T-cells genes, determining the activation state, proliferation, and differentiation (6). Indeed, prolonged antigen stimulation may induce changes in the CD8+ T-cell receptor repertoire leading to the expression of NKRs; and chronic antigen stimulation of T cells also leads to other phenotypic changes such as the loss of costimulatory molecules (e.g., CD28) (5). Usually, CD8+ T-cell memory subsets display specific responses based on the expression of killer cell immunoglobulin-like receptors (KIRs) used to distinguish unhealthy cellular targets from the healthy host cells (7, 8). However, high antigen concentrations can bypass the KIR-mediated inhibition of T-cell activation. Dynamic KIR expression may mediate T-cell tolerance to self-antigens by down regulating self-reactive T-cells (9). Nevertheless, the functional significance of the inhibitory or activating NKRs on NK-like CD8+ T-cells is still unclear.
Natural killer-like CD8+ T-cells have been described using different names, for example: CD56+CD8+ NKT-like cells (10), CD28−KIR+ CD8+ T-cells (5), KIR+CD8+ T-cells—particularly those expressing NKG2A—(11), or the general term innate CD8+ T-cells—since NKR+ αβT-cells likely represent immune effector cells that are capable of combining innate and adaptive functions (12). Table 1 summarizes the most relevant information regarding the characterization of NK-like CD8+ T-cells.
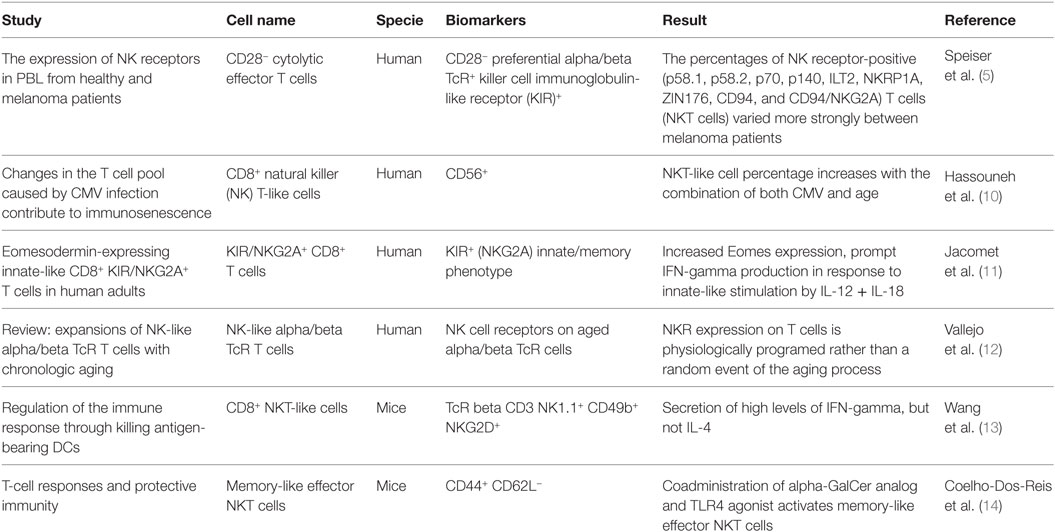
Table 1. Characterization of NK-like CD8+ T-cells.
In both the human umbilical cord and in healthy adults, an “innate/memory-like” CD8+ T-cell subset that expresses KIR and NKG2A has been described. These cells were EOMEShi, exhibited potent antigen-independent cytotoxic activity, and produced IFN-gamma in response to IL-12 + IL-18 (11). The differentiation of these cells, similar to that of the overall pool of CD8+ T-cells, can be influenced by aging. Specifically, age has been associated with increased susceptibility to infections and inflammatory diseases (15), cancer, and autoimmunity (16). The differentiation of human NK-like CD8+ T-cells is initiated after viral/tumor antigen priming and may be influenced by other factors such as aging, autoimmunity, or inflammation. These cytotoxic CD8+ T-cells, when activated, implement a differential expression of NKRs, leading to memory and migration responses. According to the expression of the CD8 marker on their surface, these cells are classified as bright or dim and display different functional properties (Figure 1A). In this context, we reviewed the literature regarding NK-like CD8+ T-cells and their phenotypic characterization associated with viral infection, immunosenescence, and diseases.
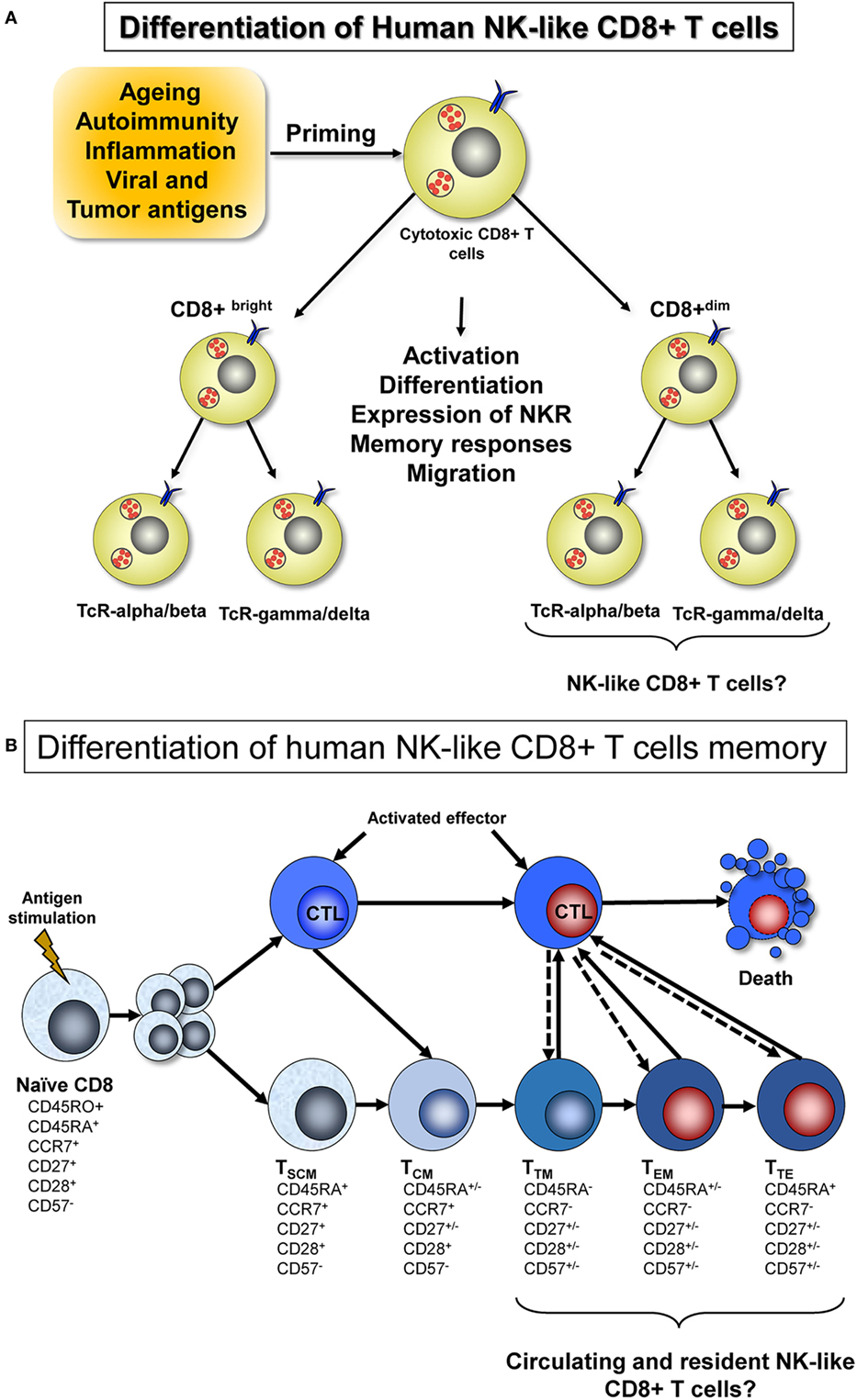
Figure 1. (A) Differentiation of human NK-like CD8+ T-cells and expression of TcR. The total pool of cytotoxic CD8+ T-cells is exposed to different TcR-alpha/beta and TcR-gamma/delta dependent or independent microenvironmental stimuli. From this pool originate the NK-like CD8+ T cells, which can be induced by the transduction of signals that activate or inhibit gene expression that, in turn, determines cytokine secretion, effector/regulating functions, migration/tissue retention, activation state, proliferation, and differentiation. Other factors that influence this process are aging, autoimmunity, inflammation, and the presence of viral and tumor antigens. (B) Differentiation of human NK-like CD8+ T-cell memory. Differentiation of CD8+ T-cell memory starts after naïve T-cell activation. According to the recently proposed model of progressive differentiation, the fate of T-cells depend on the duration of signaling and the presence or absence of cytokines (17). Thus, a single naïve cytotoxic T lymphocyte will differentiate gradually to different memory subsets: stem cell memory (TSCM), central memory (TCM), transitional memory (TTM) cells, effector memory (TEM), and terminally differentiated effector memory (TTE). In consequence, brief antigen stimulation will generate TCM cells and TTM cells, the later being more differentiated than TCM cells but not as fully differentiated as TEM cells, in terms of phenotype. On the other hand, sustained stimulation together with presence of cytokines will generate TEM and TTE cells, which most probably include NK-like CD8+ T cells both circulating or tissue resident.
Phenotypic Characterization of NK-like CD8+ T-Cells
Immunological memory is the ability of the immune system to respond more rapidly and effectively to previously encountered pathogens. This is a classical feature of adaptive immunity, which is derived from unique patterns of gene expression. A faster and stronger transcription of previously activated genes occurs in memory T-cells compared with naïve cells. This ability to remember past transcriptional responses is termed “adaptive transcriptional memory” (18). After acute infections, CD8+ T-cell memory differentiation leads to the generation of functionally distinct populations, with either proliferative potential or cytotoxic effector functions, that recirculate into lymphoid tissues or remain tissue-resident (6). This phenomenon depends on the expression of several receptors, including the C-C chemokine receptor 7 (CCR7) and CD45RA, which have been used to discriminate naïve (N; CD45RA+CCR7+), central memory (TCM; CD45RA−CCR7+), transitional memory (TTM; CD45RA−CCR7−), and terminally differentiated T-cells (TEMRA or TTE: CD45RA+CCR7−) (15). A further classification divides the CCR7−CD8+ T-cell subpopulation into three distinct memory subsets according to the expression of CD45RA: CD45RAnull, CD45RAdim, and CD45RAbright (19).
There are several models of memory CD8+ T-cell differentiation (20) and two new subsets have been recently described: the “stem cell-like memory T-cells” (TSCM) (21) and the “transitional memory (TTM) T-cells” (17). Differentiation of circulating memory CD8+ T-cells starts after antigen challenge and subsequent naïve T-cell activation. The recently described progressive differentiation model proposes that the fate of the T-cells depends on the duration of signaling and the presence or absence of cytokines. Thus, a single naïve cytotoxic T lymphocyte will differentiate gradually to different memory subsets (Figure 1B). In consequence, brief antigen stimulation will generate TCM and TTM cells, while sustained stimulation together with the presence of cytokines will lead to TEM and TTE (TEMRA) cells that re-express CD45RA (17, 22–24). Additionally, another type of memory cell has been described, the “resident memory T-cells” (TRM), which are non-recirculating memory T-cells with long-term persistence in epithelial barrier tissues. As shown in Figure 1B, it is probable that NK-like CD8+ T-cells emerge from TTM, TEM, or TTE CD8+ T-cell phenotypes; these cells could be circulating or tissue resident.
In addition, some authors suggest that the CD45RO+CD45RA− T-cells comprise diverse memory subsets, including TCM, TSCM, TEM, and TRM subsets, which are heterogeneous in their generation, distribution, and function (24). Thus, TRM cells may persist in the absence of antigens and display several effector functions. Moreover, TRM cells could have evolved to provide rapid immune protection against pathogens. However, autoreactive, aberrantly activated, and malignant TRM cells can contribute to numerous human inflammatory diseases (25).
CD8+ T-cells in lymphoid tissues are naïve, while in mucosal sites, these cells are IFN-gamma producing TEM cells. The T-cell activation marker, CD69, is constitutively expressed by memory T-cells in all tissues, distinguishing them from circulating subsets. TRM cells expressing CD69 are also present in human mucosal and peripheral tissue sites (24). However, the mucosal memory T-cells exhibit additional distinct phenotypic and functional properties (26). In particular, human intrahepatic lymphocytes are rich in CD1d-unrestricted T-cells that co-express NKRs (NK-like CD8+ T-cells), and it is possible that the hepatic epithelial cells and the cytokine milieu play a role in the shaping of these cells. For example, IL-15 is capable of inducing Ag-independent upregulation of NKRs in the CD8+CD56− T-cells. This increased percentage of intrahepatic NK-like CD8+ T-cells could be in part due to a local CD8+ T-cell differentiation (27) and could explain how NK-like CD8+ T-cells differentiate in the human liver.
The distribution and tissue residence of naïve, central and effector memory, and terminal effector subsets is contingent on both their differentiation state and tissue localization. Moreover, T-cells homeostasis, driven by cytokine or TcR-mediated signals, is different in CD4+ or CD8+ T-cell lineages and varies with their differentiation stage and tissue localization (28). In this sense, it is important to investigate NK-like CD8+ T-cells with respect to memory phenotype, functional properties, and long-term differential fates following acute infection or chronic diseases.
NK-like CD8+ T-Cells in Virus Infection and Immunosenescence
Throughout an individual’s lifetime, the memory T-cell percentage undergoes dynamic changes that can be classified into three phases: memory generation during infancy and early childhood, memory homeostasis, which occurs after age 20–25, and immunosenescence (24). This last term refers to the deterioration of the immune system associated with aging, and it is characterized by substantial alterations of the T-lymphocyte subsets (29). An increased expression of NK cell markers on T-cells has been reported to be associated with aging and chronic activation of the immune system, as reflected in the accumulation of effector/senescent T-cells (30). This memory subpopulation is interesting, because the senescence of human TTE CD8+ T-cells is stringently controlled by distinct and reversible cell signaling events (31). Also, there is evidence of a differential regulation of NKR expression between T-cells and NK cells suggesting that NKR expression on T-cells is physiologically programed rather than a random event of the aging process (12). This may suggest that the NK-like CD8+ T-cells have a functional plasticity with respect to their “adaptation” that allows them to respond to different stimuli.
CMV and HIV infection strongly affect CD8+ T-cell differentiation and maturation, enhancing immunosenescence due to the accumulation of highly differentiated TEM and TTE cells (32, 33). The CD57 antigen has been traditionally used to characterize terminally differentiated “senescent” cells, as CD57+ T-cells exhibit a reduced proliferative capacity and altered functional properties (34). Of note, the expansion of CD57+CD8+ T-cells is a hallmark of latent CMV infection (35). The CD57+CD8+ T-cell subset is functionally heterogeneous, and includes highly cytotoxic TTE cells that express intermediate levels of EOMES, as well as non-cytotoxic EOMEShi TTE cells with high proliferative capacity (36). These above-referenced studies highlight the existence of functional heterogeneity among the CD8+ T-cell memory subsets. Regarding NK-like CD8+ T cells, high percentages of these cells also express the CD57 marker and likewise arise after CMV infection (10). It has been well established that NK-like CD8+ T-cells expand with age (30, 37–39) and some studies suggest that their frequency is increased in CMV-seropositive individuals (40–42). However, recent work performed in healthy young individuals indicates that NK-like CD8+ T-cell frequency is not affected by CMV latent infection. Thus, the authors propose that these cells accumulate with age in the CMV-seropositive individuals, rather than with CMV infection per se (10).
Moreover, NK-like CD8+ T-cells from Epstein–Barr virus (EBV)-associated tumor patients are quantitatively and functionally impaired and in a human-thymus-SCID chimera model, the EBV-induced human NK-like CD8+ T-cells synergize with NK-like CD4+ T-cells suppressing EBV-associated tumors upon induction of a Th1-bias (43). Additionally, in women with human papillomavirus (HPV)-associated cervical neoplasia, there are increased levels of CD28−, TEM, and CD16+CD56+ CD8+ T-cells in peripheral blood, probably associated with the chronic infection with HPV (44). As we mentioned above, NK-like CD8+ T-cells possess a diverse TcR repertoire and there is evidence that these cells can function as antigen-specific suppressive cells that regulate the immune response through killing antigen-bearing dendritic cells (13). The class-I MHC-restricted T-cell-associated molecule (CRTAM) has been shown to be expressed only on activated class-I MHC-restricted T-cells, including NK-like CD8+ and conventional CD8+ T-cells. Of note, this molecule is a surface marker of activation associated with human viral infections and autoimmune diseases (45). These studies show that the NK-like CD8+ T-cells interact with other cells and that chronic stimulation determines their phenotype.
NK-like CD8+ T-Cells and Disease
There is evidence in the literature of an immune suppressor role for the CD8+CD28− T-cells (Ts) and the CD3+CD56+ T-cells. Patients with B-cell non-Hodgkin’s lymphoma had significantly higher percentages of Ts cells and NKT-like cells than healthy people, suggesting that, in this type of lymphoma, these cell subsets may possibly have an immunosuppressive role (46). It has been suggested that tumor-induced dysfunction of CTL in patients with multiple myeloma may contribute to immune escape and causes clonal T-cell immunosenescence, but not exhaustion, as a predominant feature. These cells exhibited a senescent secretory effector phenotype: KLRG-1+/CD57+/CD160+/CD28− (47) and may possibly be NK-like CD8+ T-cells with TEM or TTE phenotype. Furthermore, the use of ex vivo-expanded NK and NK-like T-cells has been reported seems to be safe and it could be an approach for further clinical evaluation in cancer patients (47).
Patients with Behcet’s uveitis also showed an increased number of CD8+ T-cells and CD8+CD56+ (NKT-like) cells in the aqueous humor, indicating a possible role for these subsets in the immunopathogenesis of the disease (48). CD56+CD8+ NKT-cells express more IFN-gamma and KIR in patients with leishmaniasis compared with healthy subjects (49). Similarly, loss of CD28 was associated with an increased percentage of T and NK-like T-cells producing IFN-gamma or TNF-alpha in patients with chronic obstructive pulmonary diseases (44). Furthermore, targeting peripheral blood pro-inflammatory CD28− T-cells and NK-like CD8+ T-cells by inhibiting CD137 expression may possibly be of relevance to the treatment of bronchiolitis obliterans syndrome (50). In this regard, the percentage of CD57+CD8+ T-cells is the strongest immunologic predictor of future cutaneous squamous cell carcinoma and was correlated with increasing CD8+ T-cell differentiation (36). As mentioned above, a high percentage of CD57+CD8+ T cells are NK-like.
The human activating receptor NKG2D recognizes a diverse family of ligands (MICA, MICB, and ULBPs 1–6), leading to the activation of effector cells and triggering the lysis of target T-cells. Differential expression of NKG2D is regulated in the different T-cell subsets by epigenetic mechanisms (51). The NKG2D receptor–ligand system plays an important role in the immune response to infections, tumors, transplanted grafts, and autoantigens. In lung cancer patients, NK-like CD8+ T-cells exhibit low expression of NKG2D, which correlates with the pathological stage (52). Thus, understanding the regulation of human NK-like CD8+ T-cells activation could be a strategy to manipulate T-cell-mediated responses including tumoral responses and infections.
Patients with Behcet’s uveitis also showed an increased number of CD8+ T-cells and CD8+CD56+ (NKT-like) cells in the aqueous humor, indicating a possible role for these subsets in the immunopathogenesis of the disease (48). A skewed distribution and lower frequencies of circulating activated CD161+ NK-like CD8+ T-cells was observed in patients with common variable immunodeficiency disorders, suggesting a probable regulatory function of these cells (53). CD161 is expressed by several T-cell subsets, including CD8+, NK-like CD8+, CD4+, and TcR-gamma/delta cells and all CD161+ lymphocytes display a shared innate response to IL-12 + IL-18 in which CD161 can act as a costimulatory receptor (54). Additionally, IL-23 responsiveness is restricted to the CD161+ subset in CD45RO+CD8+ memory T-cells (55). Moreover, both the frequency and the absolute number of CD161+CD8+ T-cells are decreased in the peripheral blood of patients suffering from systemic lupus erythematosus (56). A skewed distribution and lower frequencies of circulating activated CD161+ NK-like CD8+ T-cells was observed in patients with common variable immunodeficiency disorders, suggesting a probable regulatory function of these cells (53).
Finally, evidence from murine research has shown that harnessing the immune adjuvant properties of NK-like CD8+ T-cells can be an effective strategy to generate immunological memory and anticancer immunity. This effect was associated with the IFN-gamma-dependent expansion of KLRG1+CD8+ effector T-cells (57). Another study in mice assessed the vaccine induction of CD8+ T-cell responses and protective immunity after coadministration of alpha-GalCer analog and TLR4 agonist. The results showed a robust CD8+ T-cell response to PyCS protein and WT-1 antigen and activation of memory-like effector NK-like CD8+ T-cells, with a CD44+CD62L− phenotype (14).
Future Research Considerations
This review supports the concept that NK-like CD8+ T-cells are part of a cell subset associated with the acquisition of differential marker profiles, although there is little information regarding the functional properties of these cells in humans. Thus, there are several questions to clarify. First, are NK-like CD8+ T-cells CD3+/CD8+/CD56+ bright or dim? Second, are they TcR-alpha/beta+, TcR-gamma/delta+, or TcR-gamma/delta−? Third, do these cells contain variant or semi-invariant chains? Fourth, do they have CD8+/alpha-beta or CD8+/alpha-alpha? It would also be interesting to evaluate whether these subpopulations differ in their abilities to stimulate other immune cells and if they have diverse immunoregulatory functions including activation/suppression of diverse cells. Additionally, more information is needed with regards to how different environmental factors, such as autoimmunity, inflammation, viral antigen re-exposure, or persistent tumor antigens might allow the differentiation (“adaptation”) of the memory NK-like CD8+ T-cells. Finally, it is important to be cognizant of the different NK/T-cell-like cell populations and exclude the NK-like CD8+ T-cells when analyzing the immune responses mediated by conventional CD8+ T-cells and vice versa.
Nutrient/metabolic regulators can influence NK-like CD8+ T-cell differentiation, which could be analyzed through nutrigenomics contributing to the knowledge regarding the differentiation of this subset. As the balance between activating/inhibitory receptors controls NK-like CD8+ T-cells immune responses, it should also be considered that the expression of these receptors could depend on the cell differentiation state, the age, and/or diseases of the individual.
In conclusion, a thorough functional and phenotypic characterization of human NK-like CD8+ T-cells will be fundamental in order to provide mechanistic insight into the functional adaptation of these cells to aging, autoimmunity, inflammation, viral, and tumor antigens and toward their exploitation in potential therapeutic applications.
Author Contributions
M-PL conceived and participated in the design and coordination of the manuscript. AP and RS provided helpful discussions and edited the manuscript. All authors wrote, read, and approved the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
To Jesse Haramati from CUCBA University of Guadalajara México, for the style correction of the manuscript.
Funding
This work was supported by Coordination of Research and Graduate Studies with the number SAC/CI/027/16, CUSUR, University of Guadalajara México.
References
1. Legat A, Speiser DE, Pircher H, Zehn D, Fuertes Marraco SA. Inhibitory receptor expression depends more dominantly on differentiation and activation than “exhaustion” of human CD8 T cells. Front Immunol (2013) 4:455. doi: 10.3389/fimmu.2013.00455
2. Alter G, Rihn S, Streeck H, Teigen N, Piechocka-Trocha A, Moss K, et al. Ligand-independent exhaustion of killer immunoglobulin-like receptor-positive CD8+ T cells in human immunodeficiency virus type 1 infection. J Virol (2008) 82(19):9668–77. doi:10.1128/JVI.00341-08
3. Carrillo-Bustamante P, Kesmir C, de Boer RJ. The evolution of natural killer cell receptors. Immunogenetics (2016) 68(1):3–18. doi:10.1007/s00251-015-0869-7
4. Takii Y, Hashimoto S, Iiai T, Watanabe H, Hatakeyama K, Abo T. Increase in the proportion of granulated CD56+ T cells in patients with malignancy. Clin Exp Immunol (1994) 97(3):522–7. doi:10.1111/j.1365-2249.1994.tb06120.x
5. Speiser DE, Valmori D, Rimoldi D, Pittet MJ, Lienard D, Cerundolo V, et al. CD28-negative cytolytic effector T cells frequently express NK receptors and are present at variable proportions in circulating lymphocytes from healthy donors and melanoma patients. Eur J Immunol (1999) 29(6):1990–9. doi:10.1002/(SICI)1521-4141(199906)29:06<1990::AID-IMMU1990>3.0.CO;2-9
6. Bottcher J, Knolle PA. Global transcriptional characterization of CD8+ T cell memory. Semin Immunol (2015) 27(1):4–9. doi:10.1016/j.smim.2015.03.001
7. Rajalingam R. Human diversity of killer cell immunoglobulin-like receptors and disease. Korean J Hematol (2011) 46(4):216–28. doi:10.5045/kjh.2011.46.4.216
8. Hombach AA, Abken H. Young T cells age during a redirected anti-tumor attack: chimeric antigen receptor-provided dual costimulation is half the battle. Front Immunol (2013) 4:135. doi:10.3389/fimmu.2013.00135
9. Huard B, Karlsson L. KIR expression on self-reactive CD8+ T cells is controlled by T-cell receptor engagement. Nature (2000) 403(6767):325–8. doi:10.1038/35002105
10. Hassouneh F, Campos C, Lopez-Sejas N, Alonso C, Tarazona R, Solana R, et al. Effect of age and latent CMV infection on CD8+ CD56+ T cells (NKT-like) frequency and functionality. Mech Ageing Dev (2016) 158:38–45. doi:10.1016/j.mad.2015.12.003
11. Jacomet F, Cayssials E, Basbous S, Levescot A, Piccirilli N, Desmier D, et al. Evidence for eomesodermin-expressing innate-like CD8(+) KIR/NKG2A(+) T cells in human adults and cord blood samples. Eur J Immunol (2015) 45(7):1926–33. doi:10.1002/eji.201545539
12. Vallejo AN, Mueller RG, Hamel DL Jr, Way A, Dvergsten JA, Griffin P, et al. Expansions of NK-like alphabetaT cells with chronologic aging: novel lymphocyte effectors that compensate for functional deficits of conventional NK cells and T cells. Ageing Res Rev (2011) 10(3):354–61. doi:10.1016/j.arr.2010.09.006
13. Wang C, Liu X, Li Z, Chai Y, Jiang Y, Wang Q, et al. CD8(+)NKT-like cells regulate the immune response by killing antigen-bearing DCs. Sci Rep (2015) 5:14124. doi:10.1038/srep14124
14. Coelho-Dos-Reis JG, Huang J, Tsao T, Pereira FV, Funakoshi R, Nakajima H, et al. Co-administration of alpha-GalCer analog and TLR4 agonist induces robust CD8(+) T-cell responses to PyCS protein and WT-1 antigen and activates memory-like effector NKT cells. Clin Immunol (2016) 168:6–15. doi:10.1016/j.clim.2016.04.014
15. Vasudev A, Ying CT, Ayyadhury S, Puan KJ, Andiappan AK, Nyunt MS, et al. Gamma/delta T cell subsets in human aging using the classical alpha/beta T cell model. J Leukoc Biol (2014) 96(4):647–55. doi:10.1189/jlb.5A1213-650RR
16. Fulop T Jr, Larbi A, Dupuis G, Pawelec G. Ageing, autoimmunity and arthritis: perturbations of TCR signal transduction pathways with ageing – a biochemical paradigm for the ageing immune system. Arthritis Res Ther (2003) 5(6):290–302. doi:10.1186/ar1019
17. Mahnke YD, Brodie TM, Sallusto F, Roederer M, Lugli E. The who’s who of T-cell differentiation: human memory T-cell subsets. Eur J Immunol (2013) 43(11):2797–809. doi:10.1002/eji.201343751
18. Dunn J, McCuaig R, Tu WJ, Hardy K, Rao S. Multi-layered epigenetic mechanisms contribute to transcriptional memory in T lymphocytes. BMC Immunol (2015) 16:27. doi:10.1186/s12865-015-0089-9
19. Pita-Lopez ML, Gayoso I, DelaRosa O, Casado JG, Alonso C, Munoz-Gomariz E, et al. Effect of ageing on CMV-specific CD8 T cells from CMV seropositive healthy donors. Immun Ageing (2009) 6:11. doi:10.1186/1742-4933-6-11
20. Kaech SM, Cui W. Transcriptional control of effector and memory CD8+ T cell differentiation. Nat Rev Immunol (2012) 12(11):749–61. doi:10.1038/nri3307
21. Scholz G, Jandus C, Zhang L, Grandclement C, Lopez-Mejia IC, Soneson C, et al. Modulation of mTOR signalling triggers the formation of stem cell-like memory T cells. EBioMedicine (2016) 4:50–61. doi:10.1016/j.ebiom.2016.01.019
22. Kaech SM, Hemby S, Kersh E, Ahmed R. Molecular and functional profiling of memory CD8 T cell differentiation. Cell (2002) 111(6):837–51. doi:10.1016/S0092-8674(02)01139-X
23. Flynn JK, Gorry PR. Stem memory T cells (TSCM)-their role in cancer and HIV immunotherapies. Clin Transl Immunology (2014) 3(7):e20. doi:10.1038/cti.2014.16
24. Farber DL, Yudanin NA, Restifo NP. Human memory T cells: generation, compartmentalization and homeostasis. Nat Rev Immunol (2014) 14(1):24–35. doi:10.1038/nri3567
25. Clark RA. Resident memory T cells in human health and disease. Sci Transl Med (2015) 7(269):269rv1. doi:10.1126/scitranslmed.3010641
26. Sathaliyawala T, Kubota M, Yudanin N, Turner D, Camp P, Thome JJ, et al. Distribution and compartmentalization of human circulating and tissue-resident memory T cell subsets. Immunity (2013) 38(1):187–97. doi:10.1016/j.immuni.2012.09.020
27. Correia MP, Cardoso EM, Pereira CF, Neves R, Uhrberg M, Arosa FA. Hepatocytes and IL-15: a favorable microenvironment for T cell survival and CD8+ T cell differentiation. J Immunol (2009) 182(10):6149–59. doi:10.4049/jimmunol.0802470
28. Thome JJ, Yudanin N, Ohmura Y, Kubota M, Grinshpun B, Sathaliyawala T, et al. Spatial map of human T cell compartmentalization and maintenance over decades of life. Cell (2014) 159(4):814–28. doi:10.1016/j.cell.2014.10.026
29. Pawelec G, Solana R. Immunosenescence. Immunol Today (1997) 18(11):514–6. doi:10.1016/S0167-5699(97)01145-6
30. Tarazona R, DelaRosa O, Alonso C, Ostos B, Espejo J, Pena J, et al. Increased expression of NK cell markers on T lymphocytes in aging and chronic activation of the immune system reflects the accumulation of effector/senescent T cells. Mech Ageing Dev (2000) 121(1–3):77–88. doi:10.1016/S0047-6374(00)00199-8
31. Henson SM, Macaulay R, Riddell NE, Nunn CJ, Akbar AN. Blockade of PD-1 or p38 MAP kinase signaling enhances senescent human CD8(+) T-cell proliferation by distinct pathways. Eur J Immunol (2015) 45(5):1441–51. doi:10.1002/eji.201445312
32. Weltevrede M, Eilers R, de Melker HE, van BD. Cytomegalovirus persistence and T-cell immunosenescence in people aged fifty and older: a systematic review. Exp Gerontol (2016) 77:87–95. doi:10.1016/j.exger.2016.02.005
33. Tavenier J, Langkilde A, Haupt TH, Henriksen JH, Jensen FK, Petersen J, et al. Immunosenescence of the CD8(+) T cell compartment is associated with HIV-infection, but only weakly reflects age-related processes of adipose tissue, metabolism, and muscle in antiretroviral therapy-treated HIV-infected patients and controls. BMC Immunol (2015) 16:72. doi:10.1186/s12865-015-0136-6
34. Kared H, Martelli S, Ng TP, Pender SL, Larbi A. CD57 in human natural killer cells and T-lymphocytes. Cancer Immunol Immunother (2016) 65(4):441–52. doi:10.1007/s00262-016-1803-z
35. Pera A, Campos C, Corona A, Sanchez-Correa B, Tarazona R, Larbi A, et al. CMV latent infection improves CD8+ T response to SEB due to expansion of polyfunctional CD57+ cells in young individuals. PLoS One (2014) 9(2):e88538. doi:10.1371/journal.pone.0088538
36. Simonetta F, Hua S, Lecuroux C, Gerard S, Boufassa F, Saez-Cirion A, et al. High eomesodermin expression among CD57+ CD8+ T cells identifies a CD8+ T cell subset associated with viral control during chronic human immunodeficiency virus infection. J Virol (2014) 88(20):11861–71. doi:10.1128/JVI.02013-14
37. Looney RJ, Falsey A, Campbell D, Torres A, Kolassa J, Brower C, et al. Role of cytomegalovirus in the T cell changes seen in elderly individuals. Clin Immunol (1999) 90(2):213–9. doi:10.1006/clim.1998.4638
38. Peralbo E, Alonso C, Solana R. Invariant NKT and NKT-like lymphocytes: two different T cell subsets that are differentially affected by ageing. Exp Gerontol (2007) 42(8):703–8. doi:10.1016/j.exger.2007.05.002
39. Almehmadi M, Flanagan BF, Khan N, Alomar S, Christmas SE. Increased numbers and functional activity of CD56(+) T cells in healthy cytomegalovirus positive subjects. Immunology (2014) 142(2):258–68. doi:10.1111/imm.12250
40. Bergstrom I, Backteman K, Lundberg A, Ernerudh J, Jonasson L. Persistent accumulation of interferon-gamma-producing CD8+CD56+ T cells in blood from patients with coronary artery disease. Atherosclerosis (2012) 224(2):515–20. doi:10.1016/j.atherosclerosis.2012.07.033
41. Romo N, Fito M, Guma M, Sala J, Garcia C, Ramos R, et al. Association of atherosclerosis with expression of the LILRB1 receptor by human NK and T-cells supports the infectious burden hypothesis. Arterioscler Thromb Vasc Biol (2011) 31(10):2314–21. doi:10.1161/ATVBAHA.111.233288
42. Shmeleva EV, Boag SE, Murali S, Bennaceur K, Das R, Egred M, et al. Differences in immune responses between CMV-seronegative and – seropositive patients with myocardial ischemia and reperfusion. Immun Inflamm Dis (2015) 3(2):56–70. doi:10.1002/iid3.49
43. Xiao W, Li L, Zhou R, Xiao R, Wang Y, Ji X, et al. EBV-induced human CD8(+) NKT cells synergise CD4(+) NKT cells suppressing EBV-associated tumours upon induction of Th1-bias. Cell Mol Immunol (2009) 6(5):367–79. doi:10.1038/cmi.2009.48
44. Pita-Lopez ML, Ortiz-Lazareno PC, Navarro-Meza M, Santoyo-Telles F, Peralta-Zaragoza O. CD28-, CD45RA(null/dim) and natural killer-like CD8+ T cells are increased in peripheral blood of women with low-grade cervical lesions. Cancer Cell Int (2014) 14(1):97. doi:10.1186/s12935-014-0097-5
45. Patino-Lopez G, Hevezi P, Lee J, Willhite D, Verge GM, Lechner SM, et al. Human class-I restricted T cell associated molecule is highly expressed in the cerebellum and is a marker for activated NKT and CD8+ T lymphocytes. J Neuroimmunol (2006) 171(1–2):145–55. doi:10.1016/j.jneuroim.2005.09.017
46. Shi YX, Zhang XS, Liu DG, Guan ZZ, Jiang WQ. [Distribution of CD8+CD28- T cells and CD3+CD56+ NKT cells in peripheral blood of patients with B-cell non-Hodgkin’s lymphoma]. Ai Zheng (2004) 23(11 Suppl):1437–42.
47. Barkholt L, Alici E, Conrad R, Sutlu T, Gilljam M, Stellan B, et al. Safety analysis of ex vivo-expanded NK and NK-like T cells administered to cancer patients: a phase I clinical study. Immunotherapy (2009) 1(5):753–64. doi:10.2217/imt.09.47
48. Yu HG, Lee DS, Seo JM, Ahn JK, Yu YS, Lee WJ, et al. The number of CD8+ T cells and NKT cells increases in the aqueous humor of patients with Behcet’s uveitis. Clin Exp Immunol (2004) 137(2):437–43. doi:10.1111/j.1365-2249.2004.02536.x
49. Kumari S, Jamal F, Shivam P, Thakur A, Kumar M, Bimal S, et al. Leishmania donovani skews the CD56(+) Natural Killer T cell response during human visceral leishmaniasis. Cytokine (2015) 73(1):53–60. doi:10.1016/j.cyto.2015.01.011
50. Hodge G, Hodge S, Reynolds PN, Holmes M. Targeting peripheral blood pro-inflammatory CD28null T cells and natural killer T-like cells by inhibiting CD137 expression: possible relevance to treatment of bronchiolitis obliterans syndrome. J Heart Lung Transplant (2013) 32(11):1081–9. doi:10.1016/j.healun.2013.07.017
51. Fernandez-Sanchez A, Baragano RA, Carvajal PR, Sanz AB, Ortiz A, Ortega F, et al. DNA demethylation and histone H3K9 acetylation determine the active transcription of the NKG2D gene in human CD8+ T and NK cells. Epigenetics (2013) 8(1):66–78. doi:10.4161/epi.23115
52. Cheng N, Han F, Wang Y, Mai X, Su W. [Expression and clinical significance of CD8+natural killer T cell stimulatory receptor NKG2D in peripheral blood of lung cancer patients]. Zhongguo Fei Ai Za Zhi (2010) 13(10):962–7. doi:10.3779/j.issn.1009-3419.2010.10.06
53. Carvalho KI, Melo KM, Bruno FR, Snyder-Cappione JE, Nixon DF, Costa-Carvalho BT, et al. Skewed distribution of circulating activated natural killer T (NKT) cells in patients with common variable immunodeficiency disorders (CVID). PLoS One (2010) 5(9):e12652. doi:10.1371/journal.pone.0012652
54. Fergusson JR, Smith KE, Fleming VM, Rajoriya N, Newell EW, Simmons R, et al. CD161 defines a transcriptional and functional phenotype across distinct human T cell lineages. Cell Rep (2014) 9(3):1075–88. doi:10.1016/j.celrep.2014.09.045
55. Shen H, Zhang W, Abraham C, Cho JH. Age and CD161 expression contribute to inter-individual variation in interleukin-23 response in CD8+ memory human T cells. PLoS One (2013) 8(3):e57746. doi:10.1371/journal.pone.0057746
56. Mitsuo A, Morimoto S, Nakiri Y, Suzuki J, Kaneko H, Tokano Y, et al. Decreased CD161+CD8+ T cells in the peripheral blood of patients suffering from rheumatic diseases. Rheumatology (Oxford) (2006) 45(12):1477–84. doi:10.1093/rheumatology/kel119
Keywords: NK-like CD8+ T-cells, memory, T-cell differentiation, immunosenescence, aging, CMV, natural killer receptors, CD56
Citation: Pita-López ML, Pera A and Solana R (2016) Adaptive Memory of Human NK-like CD8+ T-Cells to Aging, and Viral and Tumor Antigens. Front. Immunol. 7:616. doi: 10.3389/fimmu.2016.00616
Received: 25 October 2016; Accepted: 06 December 2016;
Published: 19 December 2016
Edited by:
Fernando A. Arosa, University of Beira Interior, PortugalReviewed by:
Koji Yasutomo, University of Tokushima, JapanSian M. Henson, Queen Mary University of London, UK
Copyright: © 2016 Pita-López, Pera and Solana. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María Luisa Pita-López, bWFyaWEucGl0YUBjdXN1ci51ZGcubXg=