 Susana L. Silva1,2
Susana L. Silva1,2 Adriana S. Albuquerque1
Adriana S. Albuquerque1 Paula Matoso1Bénédicte Charmeteau-de-Muylder3,4,5
Paula Matoso1Bénédicte Charmeteau-de-Muylder3,4,5 Rémi Cheynier3,4,5
Rémi Cheynier3,4,5 Dário Ligeiro6Miguel Abecasis7Rui Anjos7João T. Barata1Rui M. M. Victorino1,2
Dário Ligeiro6Miguel Abecasis7Rui Anjos7João T. Barata1Rui M. M. Victorino1,2 Ana E. Sousa1*
Ana E. Sousa1*
- 1Faculdade de Medicina, Instituto de Medicina Molecular, Universidade de Lisboa, Lisboa, Portugal
- 2Centro Hospitalar de Lisboa Norte, Hospital de Santa Maria, Lisboa, Portugal
- 3Cytokines and Viral Infections, Immunology Infection and Inflammation Department, Institut Cochin, INSERM, U1016, Paris, France
- 4CNRS, UMR8104, Paris, France
- 5Université Paris Descartes, Paris, France
- 6Centro de Sangue e Tranplantação de Lisboa, Instituto Português de Sangue e Transplantação, IP, Lisboa, Portugal
- 7Departamento do Coração, Hospital de Santa Cruz, Centro Hospitalar de Lisboa Ocidental, Carnaxide, Portugal
Naive CD4 T-cell maintenance is critical for immune competence. We investigated here the fine-tuning of homeostatic mechanisms of the naive compartment to counteract the loss of de novo CD4 T-cell generation. Adults thymectomized in early childhood during corrective cardiac surgery were grouped based on presence or absence of thymopoiesis and compared with age-matched controls. We found that the preservation of the CD31− subset was independent of the thymus and that its size is tightly controlled by peripheral mechanisms, including prolonged cell survival as attested by Bcl-2 levels. Conversely, a significant contraction of the CD31+ naive subset was observed in the absence of thymic activity. This was associated with impaired responses of purified naive CD4 T-cells to IL-7, namely, in vitro proliferation and upregulation of CD31 expression, which likely potentiated the decline in recent thymic emigrants. Additionally, we found no apparent constraint in the differentiation of naive cells into the memory compartment in individuals completely lacking thymic activity despite upregulation of DUSP6, a phosphatase associated with increased TCR threshold. Of note, thymectomized individuals featuring some degree of thymopoiesis were able to preserve the size and diversity of the naive CD4 compartment, further arguing against complete thymectomy in infancy. Overall, our data suggest that robust peripheral mechanisms ensure the homeostasis of CD31− naive CD4 pool and point to the requirement of continuous thymic activity to the maintenance of IL-7-driven homeostatic proliferation of CD31+ naive CD4 T-cells, which is essential to secure T-cell diversity throughout life.
Introduction
Long-term preservation of the naive CD4 T-cell pool is vital to ensure immunity to foreign antigens and to maintain peripheral tolerance (1, 2). Naive CD4 T-cells are preserved throughout life due to a dynamic balance between thymic generation, and peripheral proliferation, survival, death, or differentiation into memory/effector cells (1, 3).
The thymus is known to be functional up to the sixth decade of life, even though an age-dependent decline in thymic activity occurs (4). Thymic output can be estimated through the quantification of TCR rearrangement circles (TRECs), which are excision by-products generated during T-cell development in the thymus (4). These episomal DNA fragments are progressively diluted with cell division in the periphery (5), being thus enriched in recent thymic emigrant cells (RTEs). Aging is associated with a progressive reduction in TREC levels (4). This decline is much more striking than the one observed in naive CD4 T-cell counts, indicating that the loss of cell replenishment due to thymic involution is complemented by peripheral dilution due to cell proliferation (4, 6–8). In fact, in contrast to mice, the establishment and maintenance of the human naive CD4 T-cell compartment are currently thought to significantly rely on post-thymic T-cell proliferation (3, 9, 10). Several in silico studies suggest that thymic output per se is insufficient to guarantee the size of the peripheral naive T-cell compartment without a major contribution of cell proliferation in the periphery (3, 9, 10). This homeostatic proliferation is driven by self-peptide/MHC interaction and/or cytokines, namely, IL-7 (9, 11, 12).
IL-7 is essential for thymopoiesis and plays a key role in peripheral naive T-cell survival through the induction of Bcl-2 (11, 13). In addition, IL-7 induces low-level naive T-cell proliferation (11, 13), which is particularly important in lymphopenic clinical settings (11, 13). In steady-state conditions, this homeostatic proliferation within the naive CD4 T-cell compartment is mainly restricted to the subset expressing CD31 [platelet endothelial cell adhesion molecule (PECAM-1)] (14), a population that includes the RTEs and is thought to have a broadly diverse TCR repertoire (12). We have also shown that IL-7 increases the levels of expression of CD31 in this subset (14). The biological significance of CD31 expression is still debatable, though it has been suggested that it may limit TCR-mediated naive CD4 T-cell responses through inhibitory signaling ascribed to its cytoplasmic immune-receptor tyrosine-base inhibitory motifs (15). In agreement, the homeostatic proliferation of CD31− naive CD4 T-cells is thought to be mainly mediated by low-affinity self-peptide/MHC interactions (16). Of note, CD31 expression is lost after TCR stimulation of naive CD4 T-cells (2, 17–19).
There are few studies on human naive CD4 T-cell homeostasis, and the interplay between peripheral mechanisms and the age-associated decline in thymic output remains unclear (3, 9, 10). Adults thymectomized early in infancy due to corrective cardiac surgery provide a unique setting to address this issue (20–27). Using this clinical model, we show here that thymic activity is required to ensure IL-7-mediated peripheral homeostatic proliferation, whereas the homeostasis of the CD31− compartment is preserved in the absence of thymic activity.
Materials and Methods
Study Design
Blood was collected from 22 adult patients submitted to thymectomy during corrective cardiac surgery in early childhood and 20 age-matched healthy controls. All the subjects gave written informed consent for blood sampling and processing. The study was approved by the Ethical Boards of Faculdade de Medicina da Universidade de Lisboa, Centro Hospitalar Lisboa Norte, and Hospital de Santa Cruz, Portugal.
Cell Isolation and Cell Culture
Peripheral blood mononuclear cells (PBMCs) were isolated from freshly collected heparinized blood via Ficoll-Paque PLUS (GE Healthcare, Uppsala, Sweden). Naive CD4 T-cells were subsequently purified by negative selection (purity > 96%, StemCell Technologies, Grenoble, France). Purified naive CD4 T-cells were cultured at 1 × 106 cells/ml with either IL-7 (10 ng/ml; R&D Systems, Minneapolis, MN, USA) or IL-2 (20 IU/ml; NIH/AIDS Research and Reference Program, Division of AIDS, NIAID, Hoffman-La Roche), for up to 13 days (d), with media replacement at d3 and d7, as we have previously described (14, 28).
Flow Cytometry
Ex vivo phenotypic analysis was performed in freshly collected whole blood using an eight-color staining protocol and a panel of monoclonal antibodies previously described (28). Purified naive CD4 T-cells were surface stained ex vivo and upon culture, followed by intracellular staining using eBioscience FoxP3 kit (eBioscience, San Diego, CA, USA), as described (28). At least 150,000 events were acquired for each sample on a BD LSRFortessa (BD Biosciences, San Jose, CA, USA). Data were analyzed using FlowJo software (TreeStar, Ashland, OR, USA) after doublet exclusion. Results are presented as proportion of a cell population or as mean fluorescence intensity (MFI) of a given marker within the specified population.
TCR Activation
Purified untouched naive CD4 T-cells were cultured at 1 × 106 cells/ml (25,000 cells/well) and stimulated with increasing concentrations of beads coated with anti-CD3 and anti-CD28 monoclonal antibodies (Dynabeads, Thermo Fischer Scientific), in order to generate a dose–response curve of TCR stimulation at the single-cell level. In agreement with a previously reported protocol (29), antibody against CD40L (clone 89-76, BD Bioscience) was added to the culture media to improve the staining. After 6 h, cells were collected and surface stained, as described (28), for the marker of cell differentiation (CD45RO) and the additional activation marker (CD69).
TREC Quantification
Signal joint (sj) and DβJβTREC analyses were conducted as described (5, 30). Briefly, multiplex PCR amplification for sjTREC, DJβ1TRECs (Dβ1-Jβ1.1 to 1.6), or DJβ2TRECs (Dβ2-Jβ2.1 to 2.7), together with the CD3γ chain was performed in triplicate on lysed PBMC. TREC and CD3γ quantifications were then performed using a LightCycler™ in independent experiments, with the same first-round serial dilution standard curve. This highly sensitive nested quantitative PCR assay allowed detection of 1 copy in 105 cells for any excision circle. The sj/βTREC ratio [sjTREC/105 cells/(DJβ1TRECs/105 cells + DJβ2TRECs/105 cells)] was calculated as described (30).
mRNA Quantification
Total RNA was extracted from purified naive CD4 T-cells using Quick-RNA MicroPrep (Zymo Research Corporation, Irvine, CA, USA). cDNA was synthesized from 50 ng of RNA (SuperScript III Reverse Transcriptase, Thermo Fischer Scientific) and used to quantify the expression levels of KLF2, FOXP1, P21, BIM, DUSP4, and DUSP6 in duplicates, using TaqMan Gene Expression Assays on a ViiA7 Sequence Detection system (both from Thermo Fischer Scientific). Results are expressed as ΔCT normalized to the medium CT levels of GAPDH and HPRT.
TCR Spectratyping Analysis
Total RNA was extracted from 105 to 106 cells with RNeasy kit (Qiagen, MD, USA), and first-strand cDNA synthesized from 1 to 2 µg of RNA (SuperScript III) using an equivolume mixture of random hexamers and oligo (dT). Amplification of the TCRVβ CDR3 was performed using primers specific for each TRBV family and a common TRCB reverse primer (31), followed by a run-off reaction that extends each different PCR product with a second TRCB FAM-labeled primer; and the third step, in which each different fluorescent TRBV-TRBC PCR fragment was separated using a capillary electrophoresis-based DNA automated sequencer. Data were collected and analyzed with GeneMapper v4.0 (Thermo Fischer Scientific) for size and fluorescence intensity determination.
Statistical Analysis
Statistical analysis was performed with Graph Prism Version 5.01 (GraphPad Software, San Diego, CA, USA). The following tests were used for analyzing epidemiological data and results from ex vivo studies as appropriate: Wilcoxon-Signed Rank/paired T-test for pairwise comparisons and unpaired T-test/Mann–Whitney for unpaired comparisons, for Gaussian and non-Gaussian distribution respectively. Cultures were analyzed using one-way ANOVA. Results were expressed as median (interquartile range or range when n < 4). P values <0.05 were considered significant.
Results
Evidence of Thymus Activity in Adults Thymectomized during the First Year of Life
We studied a cohort of 22 adults submitted to thymectomy in early childhood during corrective cardiac surgery and 20 age-matched healthy controls (Table 1; Table S1 in Supplementary Material). Of note, the thymic function is relatively stable in healthy individuals during the age-period spanned (4). The thymectomized patients were stratified into two groups according to evidence of residual thymic activity (Table 1; Table S1 in Supplementary Material). No thymic activity (∅Thy) strictly refers to cases with surgical reports of complete thymus removal and levels of sjTRECs clearly below the lower level found in controls (P < 0.0001, Figure 1A), as we previously reported (28). Individuals with some degree of thymic activity (Thy) featured sjTREC levels within the range of age-matched controls, though significantly lower (P = 0.0061, Figure 1A).
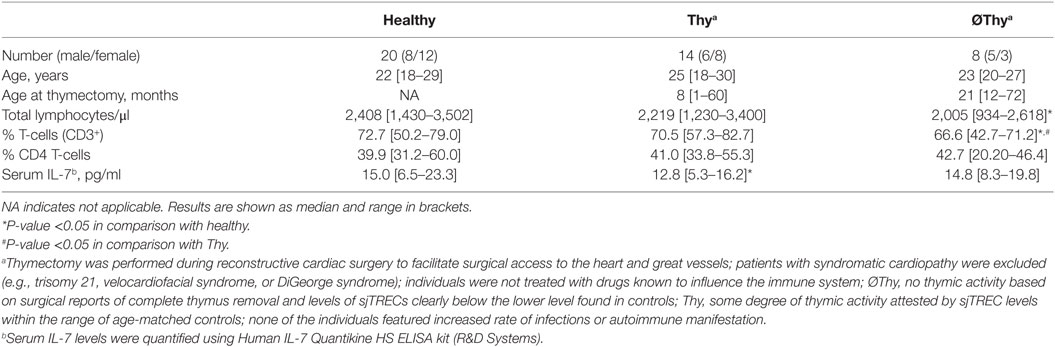
Table 1. Clinical–epidemiological characteristics of cohorts.
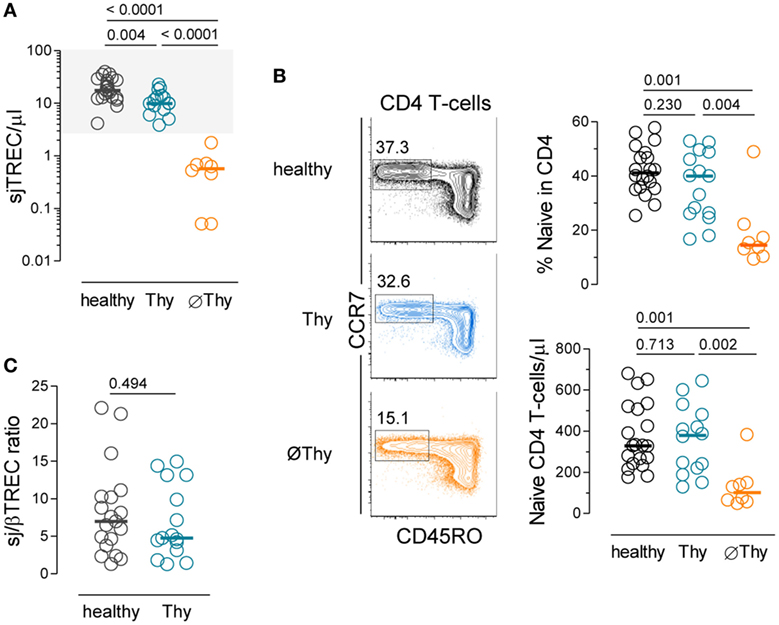
Figure 1. Naive CD4 T-cell compartment in adults thymectomized early in life. (A) Quantification of sjTREC levels in peripheral blood mononuclear cells (PBMCs) from thymectomized patients without (∅Thy), or with evidence of thymic activity (Thy), and in age-matched healthy individuals. (B) Contour plots illustrating CD45RO and CCR7 analysis within circulating CD4 T-cells of representative individuals from the three cohorts; graphs show naive CD4 T-cell frequency (top) and counts (bottom). (C) sj/βTREC ratio quantified in PBMCs from Thy and healthy individuals. Each dot represents one individual, bars represent median, and P values are shown.
Interestingly, these two groups showed almost no overlap between age at thymectomy, which was performed during the first year of life in all Thy cases except two and later on in all ∅Thy patients (Table S1 in Supplementary Material). A previous study also found an association between preservation of thymic activity and younger age at thymectomy, which was attributed to loss of thymus regenerative capacity in children older than 18 months (24, 32). It is also noteworthy that complete thymic tissue removal is more likely to occur after the first year of life due to the surgical procedures required for the type of cardiac defects (Table S1 in Supplementary Material), as well as due to age-related anatomic specificities (21) Of note, individuals with syndromatic cardiac defects were not included (Table S1 in Supplementary Material).
In agreement with complete lack of thymic activity, the ∅Thy group featured decreases in naive CD4 T-cell frequency and absolute numbers, which were statistically significant not only in comparison with healthy individuals (P = 0.0012 and P = 0.0006, respectively, Figure 1B) but also with Thy (P = 0.0041 and P = 0.0019, respectively, Figure 1B). These cells featured a truly naive phenotype based on an extensive panel of naive markers and lack of expression of molecules associated with a memory phenotype, as we have previously reported (28).
On the other hand, Thy patients showed no reduction in lymphocyte counts (Table 1) and maintained the naive CD4 T-cell compartment (Figure 1B). In order to estimate their effective thymic output, we quantified the sj/βTREC ratio, which reflects the number of proliferation cycles undergone by precursor T-cells during their intra-thymic differentiation and directly correlates with thymic activity (30). We observed similar levels of sj/βTREC ratio in Thy and age-matched healthy individuals (Figure 1C). Together with close to normal sjTREC contents, this observation supports a major contribution of thymic recovery to the maintenance of the size of naive CD4 T-cell compartment upon partial thymectomy.
We further assessed the impact of the degree of thymectomy on the structural diversity of naive CD4 T-cells by spectratyping analysis of their TCR repertoire. The distribution of the CDR3 lengths within each different Vβ family is considered to reflect the overall sequence diversity (33). A diverse polyclonal TCR repertoire is associated with a Gaussian distribution of CDR3 lengths, whereas skewed TCR repertoires feature a reduced number of peaks. A relatively preserved TCR diversity was observed in Thy patients as compared to age-matched controls (P = 0.2620), supporting that their degree of thymic activity was sufficient to ensure the preservation of the quality of the naive CD4 T-cell compartment (Figures 2A,B). Patients with no thymic activity exhibited higher numbers of non-polyclonal Gaussian families than both Thy (P < 0.0001) and healthy individuals (P = 0.0001), as shown in Figure 2.
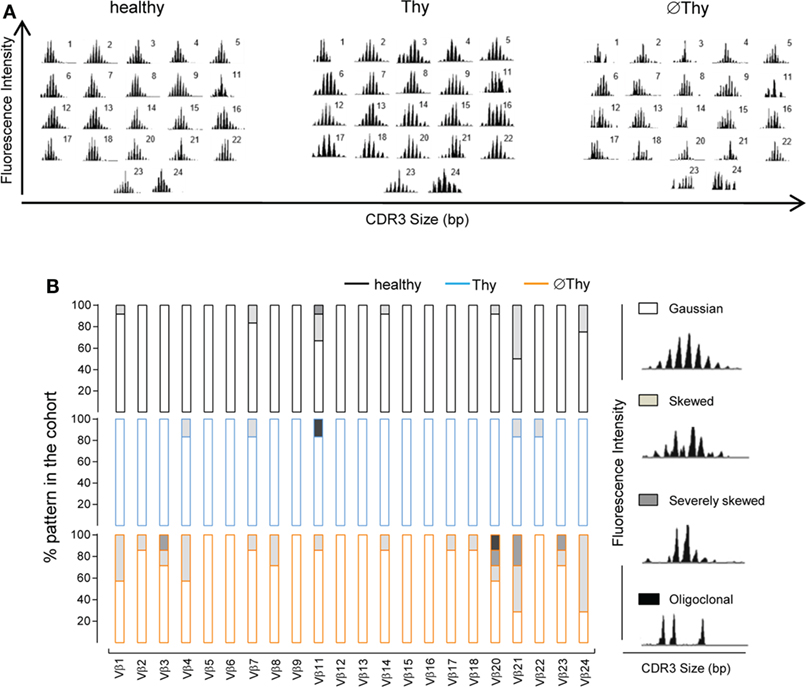
Figure 2. Spectratyping analysis of naive CD4 T-cell diversity in adults thymectomized early in life. CD3 length distribution within each of the 22 Vβ families of purified naive CD4 T-cells analyzed by spectratyping: (A) illustrative examples from thymectomized patients without (right, ∅Thy1 in Table S1 in Supplementary Material) and with evidence of thymic activity (middle, Thy6 in Table S1 in Supplementary Material), as well as age-matched healthy (left) individuals; (B) proportion of the illustrated patterns within each Vβ family in the three cohorts (∅Thy, bottom, n = 7; Thy, middle, n = 6; healthy, top, n = 12).
Of note, these differences between the two thymectomized cohorts could not be attributed to a distinct prevalence of CMV infection, since a similar proportion of individuals with IgG seropositivity against CMV was observed in ∅Thy and Thy cohorts (Table S1 in Supplementary Material). Moreover, the contraction of the naive CD4 T-cell compartment in ∅Thy individuals was not restricted to those seropositive for CMV, as previously reported (21).
Overall, peripheral homeostatic mechanisms were unable to prevent the contraction of the naive CD4 T-cell compartment upon complete thymus removal in infancy, whereas the maintenance of some degree of thymic activity allowed preservation of naive CD4 T-cells with a diverse TCR repertoire into adulthood.
Lack of Thymic Activity Does Not Associate with Increased Threshold for TCR Activation of Naive CD4 T-Cells
The maintenance of naive T-cells is also determined by the rate of their differentiation into memory–effector cells. Of note, both central and effector memory CD4 T-cell counts in individuals lacking thymic activity were found to be similar to those in age-matched healthy controls (Figure 3A). Therefore, we assessed the expression of a panel of genes known to be involved in the regulation of cell quiescence and/or of the threshold for TCR-mediated cell activation in purified naive CD4 T-cells from thymectomized and healthy individuals (Figure 3B). No alterations were found in the expression levels of the following genes: Krüppel-like factor 2 (KLF-2) (34), the transcription factor FOXP1 (35), CDKN1A (encoding the cyclin-dependent kinase inhibitor p21cip1/waf) (36), the proapoptotic Bcl-2 family member BIM (37), and the dual-specificity protein phosphatase DUSP4 (38). However, individuals with no thymic activity featured significantly higher DUSP6 transcript levels than controls, an increase not observed in those with some preservation of thymopoiesis (Figure 3B).
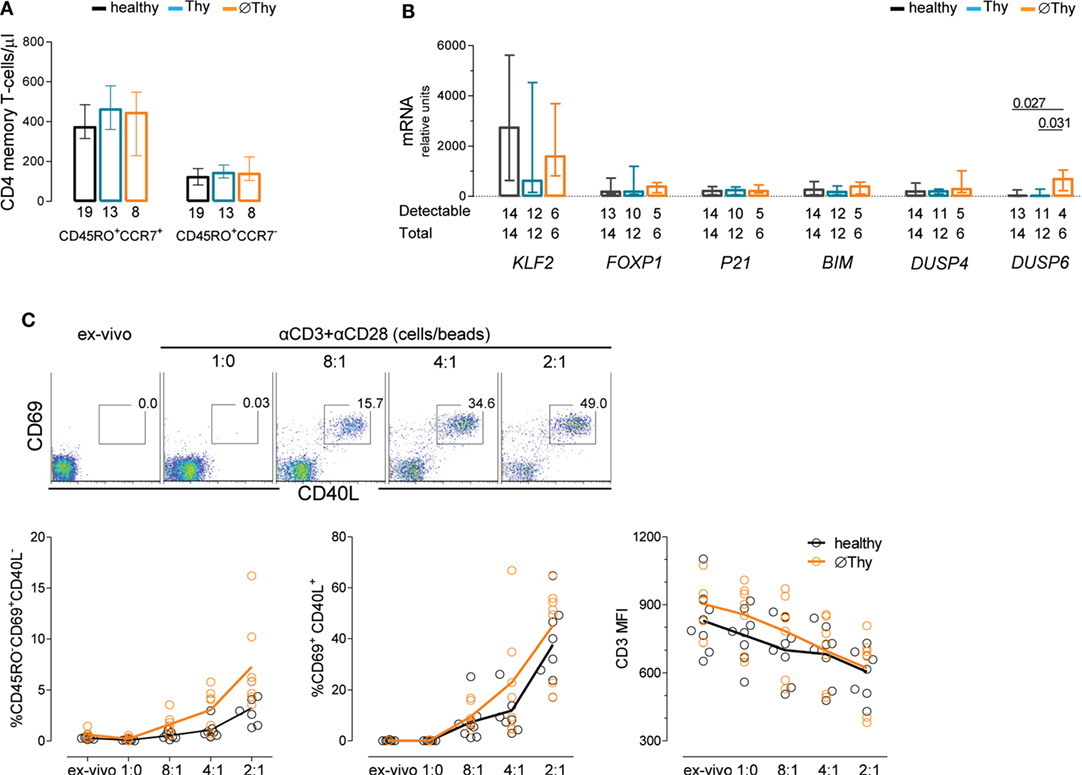
Figure 3. Maintenance of naive CD4 T-cell quiescence upon thymectomy. (A) Absolute numbers of circulating central memory (CD45RO+CCR7+) and effector memory (CD45RO+CCR7−) cells in thymectomized patients without (∅Thy) and with evidence of thymic activity (Thy), as well as age-matched healthy individuals; numbers below the graph indicate the number of individuals. (B) mRNA expression levels of genes involved in cell cycle or maintenance of naive phenotype quantified in purified naive CD4 T-cells from the three cohorts; ΔCT normalized to the medium CT levels of GAPDH and HPRT are shown; numbers below the graph indicate the total number of samples tested and those with levels above the detection threshold of the respective gene. (C) Purified naive CD4 T-cells from ∅Thy and healthy individuals were stimulated (6 h) with increasing concentrations of beads coated with anti-CD3 and anti-CD28 monoclonal antibodies with dot plots illustrating the upregulation of the activation markers CD40L and CD69 in one ∅Thy subject, and graphs showing frequencies of CD69+CD40L− cells (left), CD69+CD40L+ cells (middle), and CD3 mean fluorescence intensity (right); each dot represents one individual; lines connect means; and the two cohorts were compared with two-way ANOVA. Bars represent median and interquartile range. P values <0.05 are shown.
DUSP6 is highly specific for ERKs, leading to reduction of ERK activity that is critical for efficient TCR signaling (38, 39). Therefore, high DUSP6 levels might be associated with an increase in the threshold for TCR-induced activation in ∅Thy. To test this possibility, we performed a dose–response TCR stimulation of purified naive CD4 T-cells and quantified the upregulation of the early activation markers CD40L and CD69, in parallel with the downregulation of CD3 expression and induction of the memory marker CD45RO. Contrarily to our expectation, individuals completely lacking thymic activity responded to TCR stimulation as efficiently as healthy subjects (Figure 3C).
In conclusion, we found no apparent constraint in the differentiation of naive CD4 T-cells into the memory compartment in individuals completely lacking thymic activity.
Preservation of the CD31− Compartment of CD4 Naive T-Cells in the Absence of Thymic Activity
The CD31 molecule has been shown to be expressed in all RTEs and to be lost upon TCR stimulation of naive CD4 T-cells (12, 18). Next, we investigated the contribution of peripheral cell survival and cell cycling to the homeostasis of the CD31+ and CD31− naive CD4 T-cell subsets according to the degree of thymic activity.
The CD31+ compartment was preserved in Thy patients (Figures 4A,B). Conversely, it was significantly contracted in the ∅Thy cohort, both in frequency (Figure 4A) and absolute counts (Figure 4B), as expected in the absence of thymic activity (3, 20). Of note, the median level of CD31 expression within CD31+ naive CD4 T-cells was not significantly different in thymectomized individuals (CD31 MFI: healthy 4,494 [3,490–4,872]; Thy 3,573 [3,255–4,453]; ØThy 3,936 [2,658–4,170]; P > 0.05).
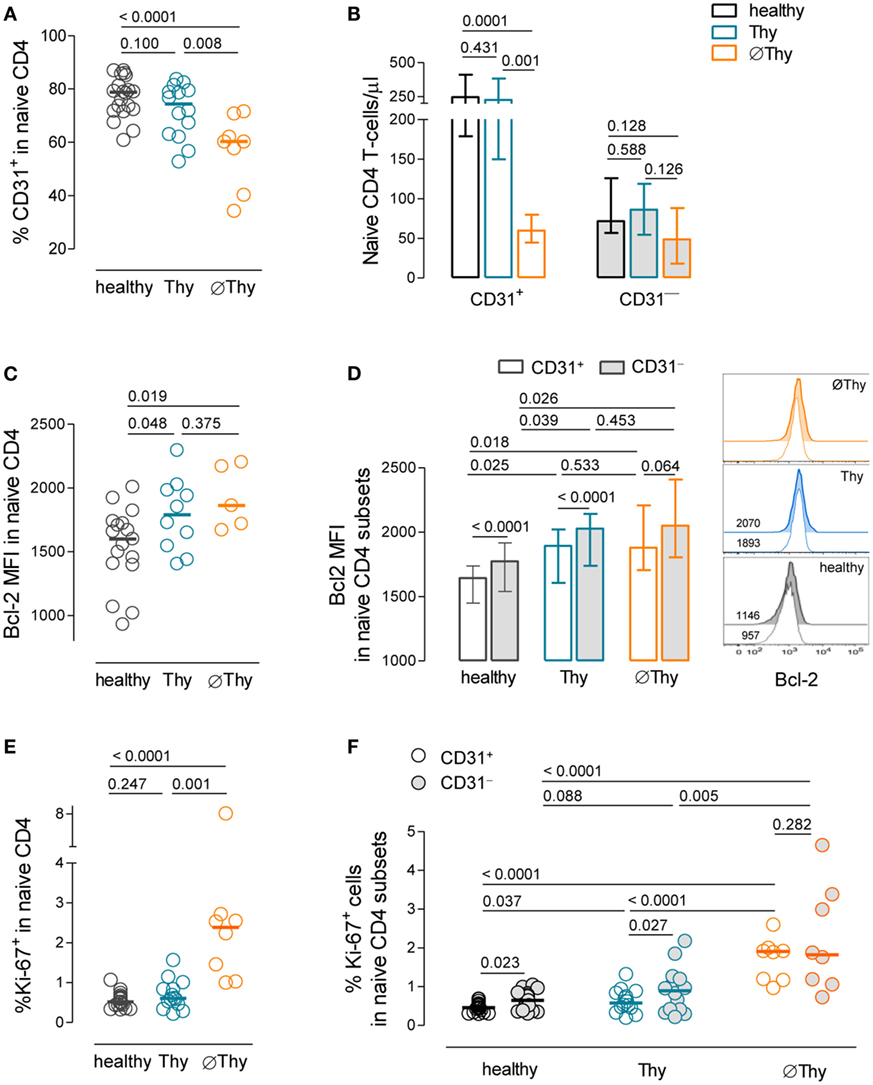
Figure 4. Homeostasis of the CD31+ and CD31− subsets of naive CD4 T-cells upon thymectomy. (A) Frequency of CD31+ cells within naive CD4 T-cells in thymectomized patients without (∅Thy) and with evidence of thymic activity (Thy), as well as age-matched healthy individuals. (B) Absolute numbers of CD31+ and CD31− naive CD4 T-cells in ∅Thy (n = 8), Thy (n = 14), and healthy (n = 19) individuals. (C) Bcl-2 mean fluorescence intensity (MFI) within naive CD4 T-cells. (D) Bcl-2 MFI within the CD31+ and CD31− subsets in ∅Thy (n = 5), Thy (n = 9), and healthy (n = 17) individuals, as illustrated in the representative overlay histograms (numbers indicate Bcl-2 MFI). (E,F) Frequency of cycling cells (Ki-67+) within naive CD4 T-cells (E) and within the CD31+ and CD31− subsets (F) in the three cohorts. In scatter graphs, each dot represents one individual, and bars represent median; bar graphs show median and interquartile range. P values are shown.
Importantly, despite the marked naive CD4 T-cell lymphopenia, ∅Thy featured preserved CD31− naive CD4 T-cell counts (Figure 4B). This finding adds to previous data on aged individuals reporting preservation of the CD31− compartment in parallel with the progressive decline of CD31+ cell counts (40) and argues in favor of the robustness of the homeostasis of CD31− naive CD4 T-cells in individuals lacking thymic activity.
The expression levels of the survival marker Bcl-2 were upregulated in both thymectomized cohorts as compared to healthy controls (∅Thy: P = 0.0187; Thy: P = 0.0487; Figure 4C), suggesting increased naive CD4 T-cell survival irrespectively of the presence of thymic activity, which persisted for more than 20 years post-thymectomy (Table 1). This increase was observed in both CD31+ and CD31− naive T-cells (Figure 4D). Of note, we found that the CD31− compartment featured significantly higher Bcl-2 MFI than CD31+ cells both in healthy and Thy individuals, a difference that was attenuated in ∅Thy (Figure 4D).
Regarding proliferation, a significant increase in the frequency of cycling cells within total naive CD4 T-cells was found only in ∅Thy (P < 0.0001 to healthy, P = 0.0007 to Thy; Figure 4E). The proportion of Ki-67+ cells was significantly higher in the CD31− than in the CD31+ compartment in both healthy individuals and in Thy patients (Figure 4F). Nevertheless, its relative contribution to the pool of proliferating naive CD4 T-cells is minor (<1%), given the large overrepresentation of CD31+ cells in these individuals (Figure 4A). On the other hand, ∅Thy individuals featured an increase in the relative representation of the CD31− subset, associated with the loss of CD31+ cells (Figure 4A), in parallel with a significant increase in the frequency of cycling cells irrespective of CD31 expression (CD31− subset: P < 0.0001 to healthy; P = 0.0048 to Thy; CD31+ subset: P < 0.0001 to healthy; P < 0.0001 to Thy; Figure 4F).
Thus, we showed that the CD31− naive CD4 T-cell compartment was maintained in the absence of thymic output, in association with both expanded cell survival and increased proliferation.
Naive CD4 T-Cells Feature Reduced Proliferative Response to IL-7 In Vitro in the Absence of Thymic Activity In Vivo
Cytokine-driven homeostatic mechanisms are crucial for naive CD4 T-cell maintenance, and IL-7 is considered the key cytokine in these processes (13). Therefore, we hypothesized that naive CD4 T-cells adjust their intrinsic ability to respond to IL-7 in order to counteract the decline in thymic output. Thus, we investigated the impact of IL-7 on purified naive CD4 T-cells, using a 13d culture system previously optimized in our laboratory (14, 28). We were able to purify the required amount of naive CD4 T-cells from 5 ØThy and 11 Thy individuals, which were compared with samples from 14 healthy subjects. We found comparable ex vivo levels of IL-7Rα-chain (CD127) expression within naive CD4 T-cells in healthy and ∅Thy, and significantly higher levels in Thy individuals (Figure 5A). Others have shown that the proximal signaling through the IL-7 receptor is preserved in adults thymectomized in infancy, as assessed by STAT5 phosphorylation upon short-term stimulation with IL-7 (20).
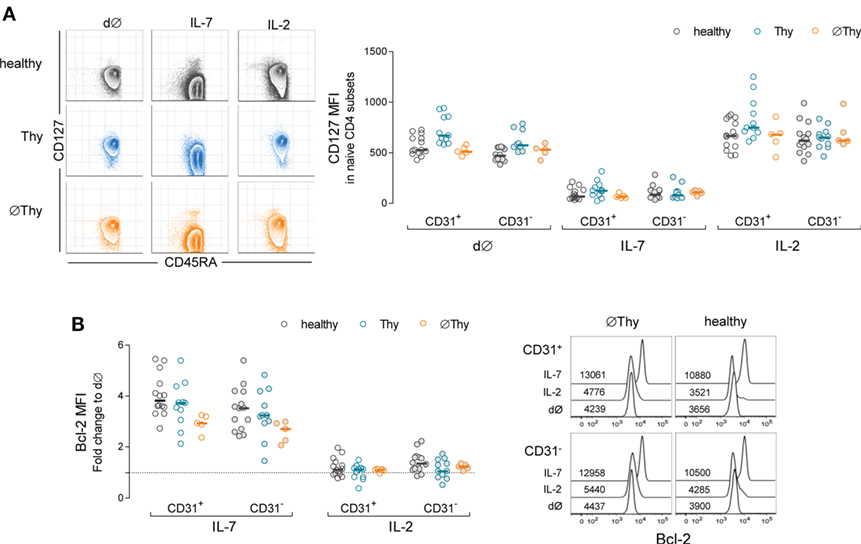
Figure 5. Impact of thymectomy on naive CD4 T-cell ability to respond to IL-7. Purified naive CD4 T-cells from thymectomized individuals without (∅Thy) or with evidence of thymic activity (Thy), and age-matched healthy controls were cultured for 13d with IL-7 or IL-2. (A) Illustrative contour plots from one individual of each cohort showing maintenance of naive phenotype (CD45RA+) and downregulation of IL-7Rα (CD127) upon culture with IL-7 but not with IL-2; graph shows CD127 mean fluorescence intensity (MFI) within the CD31+ and CD31− compartments ex vivo (d∅) and upon culture with IL-2 or IL-7 in the three cohorts: CD127 downregulation in response to IL-7 was statistically significant as compared to both d∅ and IL-2 in all cohorts/subsets (P < 0.001); no significant differences were found between d∅ and IL-2, except for CD31+ cells in healthy (P < 0.05); the comparison between cohorts revealed no significant differences, except for the levels of CD127 MFI at d∅ in Thy in comparison to both healthy (CD31+: P < 0.01; CD31−: P < 0.001) and ∅Thy (CD31+: P < 0.01) individuals. (B) Fold change of Bcl-2 MFI within CD31+ and CD31− naive CD4 T-cells upon culture with IL-7 or IL-2 as compared to d∅ in the three cohorts: Bcl-2 upregulation with IL-7 was significant in all cohorts/subsets (P < 0.001) without inter-cohort differences, except for CD31+ naive CD4 T-cells in ∅Thy in comparison with healthy (P < 0.05); overlay histograms illustrate Bcl-2 expression within gated CD31+ and CD31− naive CD4 T-cells in d∅ and upon culture with IL-7 or IL-2 in a ∅Thy and an healthy individual. Each dot represents one individual, and bars represent median.
As illustrated in Figure 5A, the cells preserved their naive phenotype upon culture with either IL-7 or IL-2, including those from ∅Thy individuals. The expected IL-7-mediated downregulation of CD127 expression (14, 41) was comparable in all individuals and, therefore, independent of the degree of thymic activity (Figure 5A). Of note, no changes occurred in the control culture condition with IL-2 (Figure 5A). Additionally, the upregulation of CD25 and CD95 by IL-7 (14, 42) was also similar in all cohorts, both in the CD31+ and CD31− compartments (data not shown).
An important physiological role of IL-7 relies on Bcl-2 induction (43). We found a clear upregulation of Bcl-2 expression, in both CD31+ and CD31− subsets in cultures with IL-7 in both thymectomized cohorts, which was not observed with IL-2 (Figure 5B).
We then investigated the proliferative response of naive CD4 T-cells using the cell-cycling marker Ki-67, which we have shown to be the best approach to reveal low-level IL-7-driven proliferation (14). Unexpectedly, we found that in contrast to healthy and Thy cohorts, ∅Thy featured no significant increase in the frequency of cycling cells in response to IL-7, as compared to both ex vivo and cultures with IL-2 (Figure 6A). As shown in Figure 6B, naive CD4 T-cell recovery per well was significantly higher upon culture with IL-7 than IL-2 in healthy (fold change 1.07 [0.86–1.54] versus 0.79 [0.51–1.63]; P = 0.0137) and Thy (fold change 1.15 [0.16–1.55] versus 0.65 [0.06–1.32]; P = 0.0259), but not in ∅Thy (fold change 0.87 [0.65–1.77] versus 0.54 [0.34–1.66]; P = 0.2234). The cell recovery upon culture with IL-7 was significantly lower in ∅Thy as compared to healthy individuals (P = 0.0236).
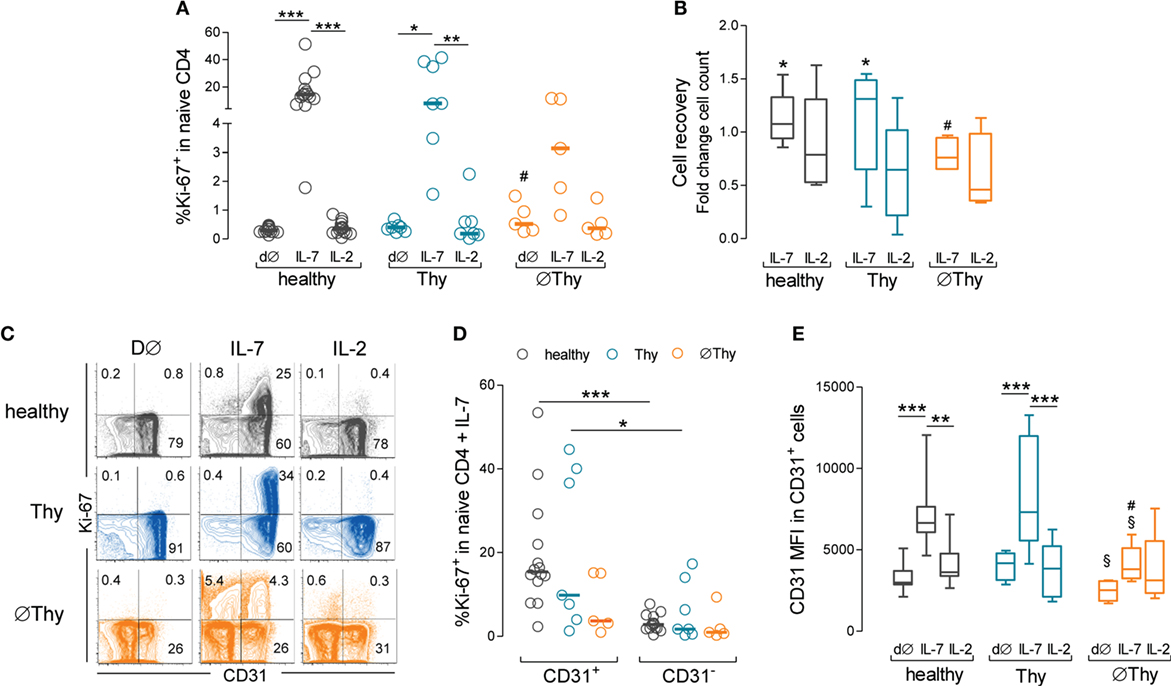
Figure 6. Impact of thymectomy on naive CD4 T-cell ability to proliferate in vitro in response to IL-7. Purified naive CD4 T-cells from thymectomized patients without (∅Thy) or with evidence of thymic activity (Thy), and from age-matched healthy controls were cultured for 13d with IL-7 or IL-2. (A) Frequency of cycling cells (Ki-67+) within total naive CD4 T-cells ex vivo (d∅) and upon culture with IL-7 or IL-2. (B) Cell recovery counts upon culture with IL-7 or IL-2. (C) Contour plots illustrate the analysis of Ki-67 versus CD31 in representative ∅Thy, Thy, and healthy individuals. (D) Frequency of Ki-67+ cells within CD31+ and CD31− naive CD4 T cells. (E) CD31 expression (mean fluorescence intensity) within the CD31+ compartment at d∅ and upon culture with IL-7 or IL-2 in ∅Thy (n = 5), Thy (n = 7), and healthy (n = 14) individuals. Each dot represents one individual, and bars represent median and interquartile range. Significant P values are shown: *P < 0.05, **P < 0.01, and ***P < 0.001 for comparisons between conditions within each cohort; #P < 0.05 for comparisons between healthy and ∅Thy or Thy; §P < 0.05 for comparisons between ∅Thy and Thy.
We have previously shown that the IL-7-induced proliferation was restricted to the CD31+ subset in healthy subjects (14), which was confirmed here (Figures 6C,D). Notably, a similar profile was observed in Thy individuals (Figures 6C,D). By contrast, patients lacking thymic activity featured reduced proliferation upon IL-7 stimulation (Figures 6C,D) and showed no significant increase in the frequency of cycling cells (Ki-67+) within the CD31+, as compared to the CD31− subset (Figures 6C,D). As a consequence, the proportion of CD31+ within cycling cells was significantly lower in ∅Thy, as compared to both healthy and Thy individuals (P < 0.0001 and P = 0.007, respectively).
We have also previously shown that IL-7 upregulates the levels of expression of CD31 within the CD31+ naive CD4 T-cell subset in a PI3K-dependent manner (14, 28). Of note, the ∅Thy cohort reached significantly lower levels of upregulation of CD31 MFI within the CD31+ subset in response to IL-7, as compared to healthy (P = 0.0035), despite featuring comparable ex vivo levels (P = 0.1052) (Figure 6E). Moreover, the upregulation of CD31 MFI was also significantly lower in ∅Thy than in Thy (P = 0.0177) individuals, although the latter featured significantly higher ex vivo levels of CD31 MFI within the CD31+ subset (P = 0.0177) (Figure 6E). These findings suggest that naive CD4 T-cells from ∅Thy patients lost the ability to respond to IL-7, possibly through the PI3K pathway, in agreement with their impaired proliferation. Conversely, they preserved the ability to upregulate Bcl-2 in response to IL-7, which we have shown previously that is not PI3K-dependent (14, 28).
Altogether, these data showed that the ability of naive CD4 T-cells to proliferate and upregulate CD31 in response to IL-7 was impaired in individuals completely lacking thymic activity.
Discussion
We investigated here mechanisms of peripheral naive CD4 T-cell homeostasis in adults with different degrees of thymus impairment since early infancy. We found that the size of the CD31− compartment was similar in healthy and thymectomized subjects, supporting the existence of thymus-independent homeostasis, possibly driven by self-peptide/MHC. On the other hand, proliferation mediated by IL-7, the main homeostatic cytokine, was severely impaired in the absence of thymopoiesis.
Thymectomy performed during corrective cardiac surgery in infancy is widely recognized as a powerful model to investigate the thymus contribution to naive T-cell maintenance beyond the establishment of the T-cell compartment. Nevertheless, a wide heterogeneity of findings has been reported (22–26, 32, 44–47). Our study focused on adults thymectomized during infancy/early childhood within a relatively narrow age range, which were grouped according to absence (∅Thy) or presence (Thy) of thymopoiesis based on circulating sjTRECs/μl (5). Our molecular strategy to stringently rule out the existence of thymic output in thymectomized patients overcomes the limitations of other approaches based solely on surgical reports (21, 23, 32, 44–46) and/or thoracic imaging (21, 32, 45, 48), which may have neglected thymic regeneration or ectopic thymus (49).
Of note, after the exclusion of the thymectomized patients lacking thymic activity, we found that both size and diversity of the naive CD4 T-cell compartment were preserved to a median of 21 years post-thymectomy. This likely occurred through both peripheral mechanisms and thymus regeneration, as supported by our finding of sj/βTREC ratios in Thy patients within the range of healthy age-matched controls. These data strengthen the recommendation to avoid complete thymectomy during cardiac surgery (44, 50), which is particularly relevant after the first year of life given the observed association between younger age at thymectomy and thymic recovery (24, 32).
The thymus provides a unique environment to generate a diverse TCR repertoire (51). This process that involves genomic recombination and gene editing at the individual cell level (52) imposes major challenges to the quantification of TCR diversity, particularly when sample availability is limited (52–57), leading us to opt for a standard approach using spectratyping. To our knowledge, there is only one study assessing the diversity of purified naive CD4 T-cells from three thymectomized children/adolescents that reported conservation of the spectratyping profiles (25). We showed here that the diversity of the TCR repertoire within the naive CD4 T-cell compartment was preserved in thymectomized individuals with some degree of remaining thymic activity and significantly contracted in patients completely lacking thymopoiesis. Although ØThy featured no major infections or autoimmunity, this profile of premature immune senescence (21, 22, 46, 58) is likely to have clinical implications not yet evaluated, since successful corrective cardiac surgery in young children only became a routine practice three decades ago, precluding extended follow-up studies (45, 59).
Of note, patients lacking thymic activity featured no major change of the transcript levels of genes involved in cell quiescence and survival of naive CD4 T-cells, except for the significant increase in DUSP6. This phosphatase enhances the TCR activation threshold by decreasing ERK phosphorylation (39). However, no significant change was observed in the activation of purified naive CD4 T-cells, suggesting that this pathway does not limit their differentiation into the memory–effector pool in completely thymectomized patients. Accordingly, they featured an increase in cycling cells within the CD31− subset that is thought to mainly proliferate in response to TCR stimulation by low-affinity self-peptide/MHC (2, 18).
We showed here that the maintenance of the CD31− subset is independent of thymic output, and that robust peripheral mechanisms ensure the homeostasis of this population. This is in agreement with the CD31− preservation that others have reported during age-associated thymic involution (12, 40). Our study revealed that, even in healthy young adults, the levels of the prosurvival molecule Bcl-2 were significantly higher in CD31− than in CD31+ naive CD4 T-cells, emphasizing the contribution of anti-apoptotic pathways for the homeostasis of the CD31− subset (60).
IL-7 is known to play a crucial role in naive CD4 T-cell homeostasis, not only by enhancing thymopoiesis (61, 62) but also through the peripheral induction of survival and proliferation (14, 28, 56, 63). We show here that there is no reduction of IL-7Rα expression within naive CD4 T-cells from thymectomized individuals, and others have shown that IL-7Rα proximal signaling, as assessed by STAT5 phosphorylation, were preserved (20). Our data support the notion that in the absence of thymopoiesis, there is mainly an impairment in the peripheral responses to IL-7 that are PI3K-dependent, namely, proliferation and CD31 upregulation, whereas Bcl-2 induction, which does not rely on this pathway, is relatively preserved. We have previously reported preservation of naive regulatory T-cells in the same ØThy cohort, despite the marked contraction of conventional naive CD4 T-cells (28). In the current study, we further analyzed the expression of the regulatory marker FoxP3 within cycling naive CD4 T-cells, and found that in contrast to the conventional cells, the FoxP3+ featured significant proliferation rates upon culture with IL-7 (median fold change of Ki-67+ cells as compared to ex vivo levels: 3.41 for FoxP3+ versus 1.32 for Foxp3−, P = 0.0248, n = 5). These findings point to a defect of conventional naive CD4 T-cells. The comparison of the two naive CD4 T-cell subsets is therefore a promising strategy to clarify the mechanisms underlying the defective IL-7 response in complete thymectomized individuals. These data will foster our understanding of IL-7 signaling in human naive CD4 T-cells and possibly identify druggable targets.
Functional heterogeneity within naive CD4 T-cells may result from the maturation process that RTEs undergo in the periphery, which may vary throughout life (64–66). It is expectable that cells with privileged response to IL-7 are more abundant in the first years of life, when accelerated growth and constant exposure to new antigens demand for peripheral expansion to ensure continuous replenishment of the naive compartment. IL-7-induced proliferation is known to be higher in mature single-positive thymocytes than peripheral T-cells (28, 66) and in cord blood than adult naive CD4 T-cells (14). Moreover, it is plausible that in elderly, an impaired ability of circulating naive CD4 T-cells to proliferate in response to IL-7 contributes to their decline, in parallel with thymic involution (40). In this context, the low-level homeostatic proliferation of naive CD4 T-cells will progressively rely on self-peptide MHC interactions, which in addition to constrain the repertoire may promote aging-associated autoreactivity.
Our data suggest a scenario where proliferative responses to IL-7 would be favored in a narrow window of time upon thymic egress, which has important implications to the therapeutic use of IL-7 in clinical settings known to be associated with thymic injury, namely, HIV/AIDS (67, 68) and chemotherapy (69, 70). Therefore, the requirement for ongoing thymopoiesis questions the suggested benefit of IL-7 therapy in the recovery of lymphopenia in thymectomized individuals (21).
In complete thymectomized individuals, the homeostatic proliferation of naive CD4 T-cells is likely to be mostly related to TCR stimulation by low-affinity self-peptide/MHC in both CD31− and CD31+ subsets, which likely contributes to further constrain their TCR repertoire.
In conclusion, our investigation of the interplay of thymic output and peripheral mechanisms to the maintenance of the naive CD4 T-cell compartment uncovered the need for continued thymic activity to the IL-7-driven peripheral proliferation of naive CD4 T-cells. These findings are of particular relevance for lymphopenic clinical settings and aging, demanding the appraisal of thymus targeting strategies in order to maximize the peripheral effect of IL-7.
Ethics Statement
The study was approved by the Ethical Boards of Faculdade de Medicina da Universidade de Lisboa, Centro Hospitalar Lisboa Norte, and Hospital de Santa Cruz, Portugal. All the subjects gave written informed consent for blood sampling and processing. Vulnerable populations, namely, minors, pregnant women, or persons with disabilities were not included.
Author Contributions
SS, AA, JB, RV, and AS designed the study; SS, AA, PM, BC-M, DL, and RC performed research; SS, MA, and RA collected clinical data; AS supervised the study; SS and AS wrote the paper.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank the sample donors for their collaboration, as well as Rita Tendeiro, Helena Nunes-Cabaço, Maria Conceição Pereira-Santos, Ana Serra-Caetano, Russell B. Foxall, Ana R. Pires, and Ana I. Pinheiro for technical assistance and discussion of the results.
Funding
This work was supported by Fundação para a Ciência e Tecnologia (FCT; POCI2010/IC/83068/2007 to RV; PTDC/SAU-MIC/109786/2009 to AS), and Calouste Gulbenkian Foundation (P132532/2013 to AS). SS, AA, and PM received FCT scholarships.
Supplementary Material
The Supplementary Material for this article can be found online at http://journal.frontiersin.org/article/10.3389/fimmu.2017.00020/full#supplementary-material.
References
1. Appay V, Sauce D. Naive T cells: the crux of cellular immune aging? Exp Gerontol (2014) 54:90–3. doi:10.1016/j.exger.2014.01.003
2. Kohler S, Thiel A. Life after the thymus: CD31+ and CD31- human naive CD4+ T-cell subsets. Blood (2009) 113(4):769–74. doi:10.1182/blood-2008-02-139154
3. den Braber I, Mugwagwa T, Vrisekoop N, Westera L, Mogling R, de Boer AB, et al. Maintenance of peripheral naive T cells is sustained by thymus output in mice but not humans. Immunity (2012) 36(2):288–97. doi:10.1016/j.immuni.2012.02.006
4. Douek DC, McFarland RD, Keiser PH, Gage EA, Massey JM, Haynes BF, et al. Changes in thymic function with age and during the treatment of HIV infection. Nature (1998) 396(6712):690–5. doi:10.1038/25374
5. Dion ML, Sekaly RP, Cheynier R. Estimating thymic function through quantification of T-cell receptor excision circles. Methods Mol Biol (2007) 380:197–213. doi:10.1007/978-1-59745-395-0_12
6. Zhang L, Lewin SR, Markowitz M, Lin HH, Skulsky E, Karanicolas R, et al. Measuring recent thymic emigrants in blood of normal and HIV-1-infected individuals before and after effective therapy. J Exp Med (1999) 190(5):725–32. doi:10.1084/jem.190.5.725
7. Utsuyama M, Hirokawa K, Kurashima C, Fukayama M, Inamatsu T, Suzuki K, et al. Differential age-change in the numbers of CD4+CD45RA+ and CD4+CD29+ T cell subsets in human peripheral blood. Mech Ageing Dev (1992) 63(1):57–68. doi:10.1016/0047-6374(92)90016-7
8. Stulnig T, Maczek C, Bock G, Majdic O, Wick G. Reference intervals for human peripheral blood lymphocyte subpopulations from ’healthy’ young and aged subjects. Int Arch Allergy Immunol (1995) 108(3):205–10. doi:10.1159/000237155
9. Pekalski ML, Ferreira RC, Coulson RM, Cutler AJ, Guo H, Smyth DJ, et al. Postthymic expansion in human CD4 naive T cells defined by expression of functional high-affinity IL-2 receptors. J Immunol (2013) 190(6):2554–66. doi:10.4049/jimmunol.1202914
10. Westera L, van Hoeven V, Drylewicz J, Spierenburg G, van Velzen JF, de Boer RJ, et al. Lymphocyte maintenance during healthy aging requires no substantial alterations in cellular turnover. Aging Cell (2015) 14(2):219–27. doi:10.1111/acel.12311
11. Tan JT, Dudl E, LeRoy E, Murray R, Sprent J, Weinberg KI, et al. IL-7 is critical for homeostatic proliferation and survival of naive T cells. Proc Natl Acad Sci U S A (2001) 98(15):8732–7. doi:10.1073/pnas.161126098
12. Kohler S, Wagner U, Pierer M, Kimmig S, Oppmann B, Mowes B, et al. Post-thymic in vivo proliferation of naive CD4+ T cells constrains the TCR repertoire in healthy human adults. Eur J Immunol (2005) 35(6):1987–94. doi:10.1002/eji.200526181
13. Fry TJ, Mackall CL. The many faces of IL-7: from lymphopoiesis to peripheral T cell maintenance. J Immunol (2005) 174(11):6571–6. doi:10.4049/jimmunol.174.11.6571
14. Azevedo RI, Soares MV, Barata JT, Tendeiro R, Serra-Caetano A, Victorino RM, et al. IL-7 sustains CD31 expression in human naive CD4+ T cells and preferentially expands the CD31+ subset in a PI3K-dependent manner. Blood (2009) 113(13):2999–3007. doi:10.1182/blood-2008-07-166223
15. Newton-Nash DK, Newman PJ. A new role for platelet-endothelial cell adhesion molecule-1 (CD31): inhibition of TCR-mediated signal transduction. J Immunol (1999) 163(2):682–8.
16. Ernst B, Lee DS, Chang JM, Sprent J, Surh CD. The peptide ligands mediating positive selection in the thymus control T cell survival and homeostatic proliferation in the periphery. Immunity (1999) 11(2):173–81. doi:10.1016/S1074-7613(00)80092-8
17. Torimoto Y, Rothstein DM, Dang NH, Schlossman SF, Morimoto C. CD31, a novel cell surface marker for CD4 cells of suppressor lineage, unaltered by state of activation. J Immunol (1992) 148(2):388–96.
18. Kimmig S, Przybylski GK, Schmidt CA, Laurisch K, Mowes B, Radbruch A, et al. Two subsets of naive T helper cells with distinct T cell receptor excision circle content in human adult peripheral blood. J Exp Med (2002) 195(6):789–94. doi:10.1084/jem.20011756
19. Demeure CE, Byun DG, Yang LP, Vezzio N, Delespesse G. CD31 (PECAM-1) is a differentiation antigen lost during human CD4 T-cell maturation into Th1 or Th2 effector cells. Immunology (1996) 88(1):110–5. doi:10.1046/j.1365-2567.1996.d01-652.x
20. Sauce D, Larsen M, Fastenackels S, Roux A, Gorochov G, Katlama C, et al. Lymphopenia-driven homeostatic regulation of naive T cells in elderly and thymectomized young adults. J Immunol (2012) 189(12):5541–8. doi:10.4049/jimmunol.1201235
21. Sauce D, Larsen M, Fastenackels S, Duperrier A, Keller M, Grubeck-Loebenstein B, et al. Evidence of premature immune aging in patients thymectomized during early childhood. J Clin Invest (2009) 119(10):3070–8. doi:10.1172/JCI39269
22. Prelog M, Keller M, Geiger R, Brandstatter A, Wurzner R, Schweigmann U, et al. Thymectomy in early childhood: significant alterations of the CD4(+)CD45RA(+)CD62L(+) T cell compartment in later life. Clin Immunol (2009) 130(2):123–32. doi:10.1016/j.clim.2008.08.023
23. Eysteinsdottir JH, Freysdottir J, Skaftadottir I, Helgason H, Haraldsson A, Ogmundsdottir HM. Vbeta usage and T regulatory cells in children following partial or total thymectomy after open heart surgery in infancy. Scand J Immunol (2009) 69(2):162–8. doi:10.1111/j.1365-3083.2008.02203.x
24. van Gent R, Schadenberg AW, Otto SA, Nievelstein RA, Sieswerda GT, Haas F, et al. Long-term restoration of the human T-cell compartment after thymectomy during infancy: a role for thymic regeneration? Blood (2011) 118(3):627–34. doi:10.1182/blood-2011-03-341396
25. Halnon NJ, Cooper P, Chen DY, Boechat MI, Uittenbogaart CH. Immune dysregulation after cardiothoracic surgery and incidental thymectomy: maintenance of regulatory T cells despite impaired thymopoiesis. Clin Dev Immunol (2011) 2011:915864. doi:10.1155/2011/915864
26. Halnon NJ, Jamieson B, Plunkett M, Kitchen CM, Pham T, Krogstad P. Thymic function and impaired maintenance of peripheral T cell populations in children with congenital heart disease and surgical thymectomy. Pediatr Res (2005) 57(1):42–8. doi:10.1203/01.PDR.0000147735.19342.DE
27. Gudmundsdottir J, Oskarsdottir S, Skogberg G, Lindgren S, Lundberg V, Berglund M, et al. Early thymectomy leads to premature immunologic ageing: an 18-year follow-up. J Allergy Clin Immunol (2016) 138(5):1439–43.e10. doi:10.1016/j.jaci.2016.05.014
28. Silva SL, Albuquerque AS, Serra-Caetano A, Foxall RB, Pires AR, Matoso P, et al. Human naive regulatory T-cells feature high steady-state turnover and are maintained by IL-7. Oncotarget (2016) 7(11):12163–75. doi:10.18632/oncotarget.7512
29. Chattopadhyay PK, Yu J, Roederer M. A live-cell assay to detect antigen-specific CD4+ T cells with diverse cytokine profiles. Nat Med (2005) 11(10):1113–7. doi:10.1038/nm1293
30. Dion ML, Poulin JF, Bordi R, Sylvestre M, Corsini R, Kettaf N, et al. HIV infection rapidly induces and maintains a substantial suppression of thymocyte proliferation. Immunity (2004) 21(6):757–68. doi:10.1016/j.immuni.2004.10.013
31. Albuquerque AS, Marques JG, Silva SL, Ligeiro D, Devlin BH, Dutrieux J, et al. Human FOXN1-deficiency is associated with alphabeta double-negative and FoxP3+ T-cell expansions that are distinctly modulated upon thymic transplantation. PLoS One (2012) 7(5):e37042. doi:10.1371/journal.pone.0037042
32. van den Broek T, Delemarre EM, Janssen WJ, Nievelstein RA, Broen JC, Tesselaar K, et al. Neonatal thymectomy reveals differentiation and plasticity within human naive T cells. J Clin Invest (2016) 126(3):1126–36. doi:10.1172/JCI84997
33. Pannetier C, Even J, Kourilsky P. T-cell repertoire diversity and clonal expansions in normal and clinical samples. Immunol Today (1995) 16(4):176–81. doi:10.1016/0167-5699(95)80117-0
34. Kuo CT, Veselits ML, Leiden JM. LKLF: a transcriptional regulator of single-positive T cell quiescence and survival. Science (1997) 277(5334):1986–90. doi:10.1126/science.277.5334.1986
35. Feng X, Wang H, Takata H, Day TJ, Willen J, Hu H. Transcription factor Foxp1 exerts essential cell-intrinsic regulation of the quiescence of naive T cells. Nat Immunol (2011) 12(6):544–50. doi:10.1038/ni.2034
36. Khanna AK, Plummer M, Nilakantan V, Pieper GM. Recombinant p21 protein inhibits lymphocyte proliferation and transcription factors. J Immunol (2005) 174(12):7610–7. doi:10.4049/jimmunol.174.12.7610
37. Tsukamoto H, Huston GE, Dibble J, Duso DK, Swain SL. Bim dictates naive CD4 T cell lifespan and the development of age-associated functional defects. J Immunol (2010) 185(8):4535–44. doi:10.4049/jimmunol.1001668
38. Huang CY, Tan TH. DUSPs, to MAP kinases and beyond. Cell Biosci (2012) 2(1):24. doi:10.1186/2045-3701-2-24
39. Li G, Yu M, Lee WW, Tsang M, Krishnan E, Weyand CM, et al. Decline in miR-181a expression with age impairs T cell receptor sensitivity by increasing DUSP6 activity. Nat Med (2012) 18(10):1518–24. doi:10.1038/nm.2963
40. Kilpatrick RD, Rickabaugh T, Hultin LE, Hultin P, Hausner MA, Detels R, et al. Homeostasis of the naive CD4+ T cell compartment during aging. J Immunol (2008) 180(3):1499–507. doi:10.4049/jimmunol.180.3.1499
41. Park JH, Yu Q, Erman B, Appelbaum JS, Montoya-Durango D, Grimes HL, et al. Suppression of IL7Ralpha transcription by IL-7 and other prosurvival cytokines: a novel mechanism for maximizing IL-7-dependent T cell survival. Immunity (2004) 21(2):289–302. doi:10.1016/j.immuni.2004.07.016
42. Kinter AL, Godbout EJ, McNally JP, Sereti I, Roby GA, O’Shea MA, et al. The common gamma-chain cytokines IL-2, IL-7, IL-15, and IL-21 Induce the expression of programmed death-1 and its ligands. J Immunol (2008) 181(10):6738–46. doi:10.4049/jimmunol.181.10.6738
43. Hassan J, Reen DJ. IL-7 promotes the survival and maturation but not differentiation of human post-thymic CD4(+) T cells. Eur J Immunol (1998) 28(10):3057–65. doi:10.1002/(SICI)1521-4141(199810)28:10<3057::AID-IMMU3057>3.3.CO;2-Q
44. Kurobe H, Tominaga T, Sugano M, Hayabuchi Y, Egawa Y, Takahama Y, et al. Complete but not partial thymectomy in early infancy reduces T-cell-mediated immune response: three-year tracing study after pediatric cardiac surgery. J Thorac Cardiovasc Surg (2013) 145(3):e1–2. doi:10.1016/j.jtcvs.2012.12.015
45. Prelog M, Wilk C, Keller M, Karall T, Orth D, Geiger R, et al. Diminished response to tick-borne encephalitis vaccination in thymectomized children. Vaccine (2008) 26(5):595–600. doi:10.1016/j.vaccine.2007.11.074
46. Mancebo E, Clemente J, Sanchez J, Ruiz-Contreras J, De Pablos P, Cortezon S, et al. Longitudinal analysis of immune function in the first 3 years of life in thymectomized neonates during cardiac surgery. Clin Exp Immunol (2008) 154(3):375–83. doi:10.1111/j.1365-2249.2008.03771.x
47. Eysteinsdottir JH, Freysdottir J, Haraldsson A, Stefansdottir J, Skaftadottir I, Helgason H, et al. The influence of partial or total thymectomy during open heart surgery in infants on the immune function later in life. Clin Exp Immunol (2004) 136(2):349–55. doi:10.1111/j.1365-2249.2004.02437.x
48. Kamali K, Ghahartars M, Amirghofran AA. Evaluation of thymic changes after median sternotomy in children. Iran J Med Sci (2014) 39(3):289–92.
49. Sanei B, Tabatabie SA, Bigdelian H, Hashemi SM, Davarpanah AH, Jafari HR, et al. Distribution of mediastinal ectopic thymic tissue in patients without thymic disease. Adv Biomed Res (2015) 4:18. doi:10.4103/2277-9175.149849
50. Sauce D, Appay V. Altered thymic activity in early life: how does it affect the immune system in young adults? Curr Opin Immunol (2011) 23(4):543–8. doi:10.1016/j.coi.2011.05.001
51. Carpenter AC, Bosselut R. Decision checkpoints in the thymus. Nat Immunol (2010) 11(8):666–73. doi:10.1038/ni.1887
52. Attaf M, Huseby E, Sewell AK. alphabeta T cell receptors as predictors of health and disease. Cell Mol Immunol (2015) 12(4):391–9. doi:10.1038/cmi.2014.134
53. Naylor K, Li G, Vallejo AN, Lee WW, Koetz K, Bryl E, et al. The influence of age on T cell generation and TCR diversity. J Immunol (2005) 174(11):7446–52. doi:10.4049/jimmunol.174.11.7446
54. Goronzy JJ, Lee WW, Weyand CM. Aging and T-cell diversity. Exp Gerontol (2007) 42(5):400–6. doi:10.1016/j.exger.2006.11.016
55. Qi Q, Liu Y, Cheng Y, Glanville J, Zhang D, Lee JY, et al. Diversity and clonal selection in the human T-cell repertoire. Proc Natl Acad Sci U S A (2014) 111(36):13139–44. doi:10.1073/pnas.1409155111
56. Parker R, Dutrieux J, Beq S, Lemercier B, Rozlan S, Fabre-Mersseman V, et al. Interleukin-7 treatment counteracts IFN-alpha therapy-induced lymphopenia and stimulates SIV-specific cytotoxic T lymphocyte responses in SIV-infected rhesus macaques. Blood (2010) 116(25):5589–99. doi:10.1182/blood-2010-03-276261
57. Arstila TP, Casrouge A, Baron V, Even J, Kanellopoulos J, Kourilsky P. A direct estimate of the human alphabeta T cell receptor diversity. Science (1999) 286(5441):958–61. doi:10.1126/science.286.5441.958
58. Zlamy M, Prelog M. Thymectomy in early childhood: a model for premature T cell immunosenescence? Rejuvenation Res (2009) 12(4):249–58. doi:10.1089/rej.2009.0864
59. Brearley S, Gentle TA, Baynham MI, Roberts KD, Abrams LD, Thompson RA. Immunodeficiency following neonatal thymectomy in man. Clin Exp Immunol (1987) 70(2):322–7.
60. Duszczyszyn DA, Williams JL, Mason H, Lapierre Y, Antel J, Haegert DG. Thymic involution and proliferative T-cell responses in multiple sclerosis. J Neuroimmunol (2010) 221(1–2):73–80. doi:10.1016/j.jneuroim.2010.02.005
61. van Lent AU, Dontje W, Nagasawa M, Siamari R, Bakker AQ, Pouw SM, et al. IL-7 enhances thymic human T cell development in “human immune system” Rag2-/-IL-2Rgammac-/- mice without affecting peripheral T cell homeostasis. J Immunol (2009) 183(12):7645–55. doi:10.4049/jimmunol.0902019
62. Okamoto Y, Douek DC, McFarland RD, Koup RA. Effects of exogenous interleukin-7 on human thymus function. Blood (2002) 99(8):2851–8. doi:10.1182/blood.V99.8.2851
63. Barata JT, Silva A, Brandao JG, Nadler LM, Cardoso AA, Boussiotis VA. Activation of PI3K is indispensable for interleukin 7-mediated viability, proliferation, glucose use, and growth of T cell acute lymphoblastic leukemia cells. J Exp Med (2004) 200(5):659–69. doi:10.1084/jem.20040789
64. Gibbons D, Fleming P, Virasami A, Michel ML, Sebire NJ, Costeloe K, et al. Interleukin-8 (CXCL8) production is a signatory T cell effector function of human newborn infants. Nat Med (2014) 20(10):1206–10. doi:10.1038/nm.3670
65. Lewis DB, Haines C, Ross D. Protein tyrosine kinase 7: a novel surface marker for human recent thymic emigrants with potential clinical utility. J Perinatol (2011) 31(Suppl 1):S72–81. doi:10.1038/jp.2010.187
66. Okazaki H, Ito M, Sudo T, Hattori M, Kano S, Katsura Y, et al. IL-7 promotes thymocyte proliferation and maintains immunocompetent thymocytes bearing alpha beta or gamma delta T-cell receptors in vitro: synergism with IL-2. J Immunol (1989) 143(9):2917–22.
67. Levy Y, Lacabaratz C, Weiss L, Viard JP, Goujard C, Lelievre JD, et al. Enhanced T cell recovery in HIV-1-infected adults through IL-7 treatment. J Clin Invest (2009) 119(4):997–1007. doi:10.1172/JCI38052
68. Sereti I, Dunham RM, Spritzler J, Aga E, Proschan MA, Medvik K, et al. IL-7 administration drives T cell-cycle entry and expansion in HIV-1 infection. Blood (2009) 113(25):6304–14. doi:10.1182/blood-2008-10-186601
69. Rosenberg SA, Sportes C, Ahmadzadeh M, Fry TJ, Ngo LT, Schwarz SL, et al. IL-7 administration to humans leads to expansion of CD8+ and CD4+ cells but a relative decrease of CD4+ T-regulatory cells. J Immunother (2006) 29(3):313–9. doi:10.1097/01.cji.0000210386.55951.c2
Keywords: naive CD4 T-cells, T-cell homeostasis, IL-7, thymus, thymectomy
Citation: Silva SL, Albuquerque AS, Matoso P, Charmeteau-de-Muylder B, Cheynier R, Ligeiro D, Abecasis M, Anjos R, Barata JT, Victorino RMM and Sousa AE (2017) IL-7-Induced Proliferation of Human Naive CD4 T-Cells Relies on Continued Thymic Activity. Front. Immunol. 8:20. doi: 10.3389/fimmu.2017.00020
Received: 04 September 2016; Accepted: 05 January 2017;
Published: 19 January 2017
Edited by:
Rene De Waal Malefyt, Merck Research Laboratories Palo Alto, USAReviewed by:
Vasileios Bekiaris, Technical University of Denmark, DenmarkAvery August, Cornell University, USA
Copyright: © 2017 Silva, Albuquerque, Matoso, Charmeteau-de-Muylder, Cheynier, Ligeiro, Abecasis, Anjos, Barata, Victorino and Sousa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana E. Sousa, YXNvdXNhQG1lZGljaW5hLnVsaXNib2EucHQ=