 Sabine Huenecke1*
Sabine Huenecke1* Claudia Cappel1
Claudia Cappel1 Ruth Esser2Verena Pfirrmann1Emilia Salzmann-Manrique1Sibille Betz1Eileen Keitl1Julia Banisharif-Dehkordi1
Ruth Esser2Verena Pfirrmann1Emilia Salzmann-Manrique1Sibille Betz1Eileen Keitl1Julia Banisharif-Dehkordi1 Shahrzad Bakhtiar1Christoph Königs1Andrea Jarisch1Jan Soerensen1
Shahrzad Bakhtiar1Christoph Königs1Andrea Jarisch1Jan Soerensen1 Evelyn Ullrich1,3Thomas Klingebiel1
Evelyn Ullrich1,3Thomas Klingebiel1 Peter Bader1
Peter Bader1 Melanie Bremm1
Melanie Bremm1
- 1Clinic for Pediatric and Adolescent Medicine, University Hospital, Frankfurt, Germany
- 2GMP Development Unit, Hannover Medical School, Institute of Cellular Therapeutics, Hannover, Germany
- 3LOEWE Center for Cell and Gene Therapy, Goethe University, Frankfurt, Germany
Natural killer (NK) cells play an important role following allogeneic hematopoietic stem cell transplantation (HSCT) exerting graft-versus-leukemia/tumor effect and mediating pathogen-specific immunity. Although NK cells are the first donor-derived lymphocytes reconstituting post-HSCT, their distribution of CD56++CD16− (CD56bright), CD56++CD16+ (CD56intermediate=int), and CD56+CD16++ (CD56dim) NK cells is explicitly divergent from healthy adults, but to some extent comparable to the NK cell development in early childhood. The proportion of CD56bright/CD56int/CD56dim changed from 15/8/78% in early childhood to 6/4/90% in adults, respectively. Within this study, we first compared the NK cell reconstitution post-HSCT to reference values of NK cell subpopulations of healthy children. Afterward, we investigated the reconstitution of NK cell subpopulations post-HSCT in correlation to acute graft versus host disease (aGvHD) and chronic graft versus host disease (cGvHD) as well as to viral infections. Interestingly, after a HSCT follow-up phase of 12 months, the distribution of NK cell subpopulations largely matched the 50th percentile of the reference range for healthy individuals. Patients suffering from aGvHD and cGvHD showed a delayed reconstitution of NK cells. Remarkably, within the first 2 months post-HSCT, patients suffering from aGvHD had significantly lower levels of CD56bright NK cells compared to patients without viral infection or without graft versus host disease (GvHD). Therefore, the amount of CD56bright NK cells might serve as an early prognostic factor for GvHD development. Furthermore, a prolonged and elevated peak in CD56int NK cells seemed to be characteristic for the chronification of GvHD. In context of viral infection, a slightly lower CD56 and CD16 receptor expression followed by a considerable reduction in the absolute CD56dim NK cell numbers combined with reoccurrence of CD56int NK cells was observed. Our results suggest that a precise analysis of the reconstitution of NK cell subpopulations post-HSCT might indicate the occurrence of undesired events post-HSCT such as severe aGvHD.
Introduction
The reconstitution of natural killer (NK) cells following allogeneic hematopoietic stem cell transplantation (HSCT) plays an important role in the response against residual malignant cells and the control of viral infections (1, 2). Independent of the graft source, NK cells typically regenerate within the first month following HSCT (3). However, there is an overrepresentation of CD56brightCD16neg NK cells in the early phase post-HSCT compared to healthy individuals (4, 5), where NK cells are composed of about 90% CD56dimCD16++ and 10% CD56brightCD16− cells. Whereas CD56dimCD16++ NK cells mediate cytotoxicity and antibody-dependent cellular cytotoxicity, the CD56brightCD16− subpopulation, which is mainly present in the early period post-HSCT, primarily secretes immunoregulatory cytokines. Presumably, the development from CD56bright to CD56dim NK cells corresponds to sequential steps of NK cell differentiation (6). In most patients, the ratio between CD56bright and CD56dim NK cells normalizes within the first 6 months post-HSCT influenced by the patient’s age and events following HSCT. A correlation between the reconstitution of NK cells and overall survival was described by few studies emphasizing their essential role in the defense of infections when T cell immunity is mainly absent (7, 8). Furthermore, NK cells were successfully applied as immunotherapy for patients with high-risk malignancies suffering from impending relapse following HSCT. In clinical studies, adoptively infused NK cells induced graft-versus-leukemia/tumor effect without concomitant severe graft versus host disease (GvHD) (9–11). In addition, recent studies ascribe GvHD reduction to NK cell function post-HSCT (12). In this work, we focused on the regeneration of CD56bright and CD56dim NK cells with a special regard on the population shifting between these subpopulations. We divided NK cells in three NK cell subpopulations. In the first step, we established reference values of CD56++CD16− (CD56bright), CD56++CD16+ (CD56int), and CD56+CD16++ (CD56dim) NK cells of healthy children and adolescents (n = 174). These reference values were matched and compared to the NK cell reconstitution of patients, who did not suffer from any viral infection or GvHD and are still alive after allogenic stem cell transplantation. In the next step, we investigated the associations between the reconstitution of the three different NK cell subpopulations in regard to the occurrence of events such as acute graft versus host disease (aGvHD) or chronic graft versus host disease (cGvHD) and severe viral infections in the first year post-HSCT in contrast to a cohort of patients without severe events assumed as control group.
Materials and Methods
Reference Cohort of Children and Adolescents
In this cross-sectional monocentric study (approval ethic committee Ref. No. 139/09), 174 donors (64 females and 110 males) were included. Residual peripheral blood samples of hematologically healthy children aged 1 month to 18 years were analyzed (patients were hospitalized, e.g., for cleft lip and palate correction). Inclusion criteria involved no incidence of immunodeficiency or infection (defined as >2 severe infections/year, >8 infections/year, persistent fungal infections, post-vaccinal complications, no evidence of acute bleeding, negative for CRP and normal leukocytes, lymphocytes, and neutrophil granulocytes).
Patients and Grafts
The reconstitution of NK cells and their subpopulation CD56bright, CD56int, and CD56dim was analyzed in n = 74 patients (n = 25 females and n = 49 males) transplanted from 2010 to 2016 (see Table 1). Indications for HSCT were high-risk acute lymphoblastic leukemia (n = 51), acute myeloid leukemia (n = 12), and myelodysplastic syndrome (n = 11). Median age at HSCT was 10.4 years (range: 1.3–24.4 years). Grafts were received from matched family donors (MSD; n = 20), matched unrelated donors (MUD) (n = 39), and haploidentical mismatched family donors with <8/10 HLA matches (MMFD; n = 15). No significant differences in the occurrence of GvHD were available in the different donor groups (MSD: 6/20, MUD: 10/39, and MMFD: 3/15). A second or third transplant was administered to n = 9 and n = 2 patients, respectively, because of relapse after first or second HSCT. Stem cell sources consisted of (bone marrow; n = 48), unmanipulated PBSC (n = 11), and T cell-depleted PBSC (n = 15). Post-transplant aGvHD occurred in n = 31 patients with grades I (n = 9), II (n = 3), III (n = 16), and IV (n = 3), cGvHD in 22 patients. Severe viral infections occurred in 18 patients including primary infection or reactivation with cytomegalovirus (CMV) (n = 8), adenovirus (ADV) (n = 5), and Epstein–Barr virus (EBV) (n = 5).
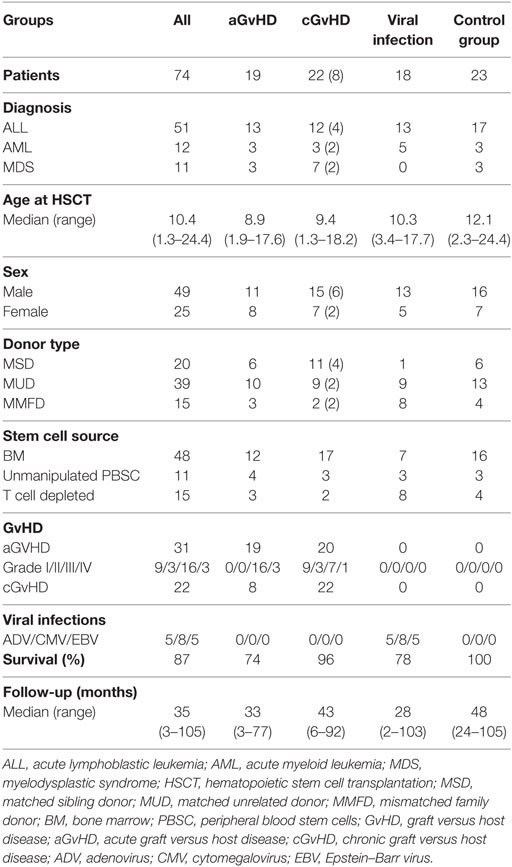
Table 1. Patient’s characteristics.
Study Design
This study was carried out in accordance with the Declaration of Helsinki and was approved by the Medical Ethics Committee of the Frankfurt University Hospital (Ref. No. #198/16). Peripheral blood was collected within the framework of a post-HSCT routine sampling for clinical follow-up from 2010 to 2016. In engrafted patients, samples were collected starting at day 15, within the first year monthly, within the second year three monthly, and afterward every 6 months until 36 months post-HSCT, respectively. In total, n = 925 measurements of n = 74 patients were included in this analysis.
Depending on the occurrence of unexpected events such as aGvHD (only grades III and IV, grades I and II excluded), cGvHD, and severe viral infections, the study group was partitioned. The patient group without aGvHD/cGvHD or any viral infections was chosen for comparison as control group.
Patients showing a distinct CD56int population in a sufficient absolute amount post-HSCT were elected for more detailed analysis applying a 10-color flow cytometry.
Assessment of NK Cells and Their Subpopulations
Natural killer cell subpopulations were analyzed on a FC500 flow-cytometer (Beckman Coulter, Krefeld, Germany) applying a five-color panel to estimate CD3+ T cells, CD19+ B cell, and CD3−CD56+ NK cells, including the differentiation into CD56++CD16−, CD56++CD16+, CD56+CD16++ NK cells, abbreviated as CD56bright, CD56int, and CD56dim NK cells, respectively. Absolute cell numbers were estimated from peripheral blood samples in a dual platform lyse-no-wash procedure as described previously (13). In brief, a tube of 100 µl EDTA-peripheral blood was labeled with CD45/CD56/CD19/CD3 tetraCHROME (clones B3821F4A/N901/J3-119/UCHT1) multi-color mAb conjugated with FITC, phycoerythrin (PE), phycoerythrin texas red (ECD), and phycoerythrin-cyanine 5 (PC5). CD16 phycoerythrin-cyanine 7 (PC7, clone: 6607118) was additionally added. For the measurement of T cells including T helper and cytotoxic T cells, we applied the tetraCHROME multi-color reagent CD45/CD4/CD8/CD3 (clones B3821F4A/SFCI12T4D11/SFCI21Thy2D3/UCHT1) conjugated with FITC, PE, ECD, and PC5. All reagents were acquired from Beckman Coulter Immutech (Marseille, France).
Differences in the expression profiles of CD56bright, CD56int, and CD56dim cells were analyzed applying two panels on a Navios™ 10-color flow cytometer (Beckman Coulter, Krefeld, Germany). Panel 1: CD226 = DNAM-1 (FITC; clone: KRA236), NKG2A (PE; clone: Z199), DUMP = CD3&CD14&CD19 (ECD; clones: UCHT1, RMO52, J3-119), CD117 (PC5.5; clone: 104D2D1), CD27 (PC7; clone: 1A4CD27), CD56 [allophycocyanin (APC); clone: N901], CD127 (APC-A700; clone: R34.34), CD16 (APC-A750; clone: 3G8), CD57 [Pacific Blue (PB); clone: NC1], CD45 [Krome Orange (KrO); clone: J.33]. Panel 2: CCR5 (FITC; clone: 2D7), killer cell immunoglobulin-like receptor (KIR) mix (CD158 + CD158b + CD158e1; clones: EB6B, GL183, Z27.3.7), DUMP = CD3 + CD14 + CD19 (ECD; clones: UCHT1, RMO52, J3-119), CD117 (PC5.5; clone: 104D2D1), CX3CR1 (PC7; clone: 2A9-1), CD56 (APC; clone: N901), CD62L (APC-A700; clone: DREG56), CD16 (APC-A750; clone: 3G8), CCR7 (PB; clone: G043H7), and CD45 (KrO; clone: J.33). All antibodies were purchased from Beckman Coulter Immutech except CCR5 (BD Biosciences, Heidelberg, Germany) and CX3CR1 (Biolegend, San Diego, CA, USA). Staining was performed, using 100 µl of peripheral blood for each tube followed by 15 min of incubation at room temperature and erythrocyte lysis applying NH4Cl reagent (Beckman Coulter, Marseille, France).
Stained Cyto-Comp™ cells were applied to compensate the fluorescence overlap. The flow cytometer fluidic stability and the optical alignment were daily tested using Flow-Check™ Fluorospheres (Beckman Coulter, Krefeld, Germany). For verification, Immunotrol cells (Beckman Coulter) were applied three times a day. Furthermore, we participate in an external quality assessment for the detection of T-, B-, and NK cells (INSTANT e. V.—provider for German round robin test, No 213).
Data evaluation was performed using Kaluza and CXP-software (Beckman Coulter).
Statistical Analysis
Statistical analysis was performed using GraphPad Prism 6 (GraphPad Software, San Diego, CA, USA). The NK subpopulation reference values of healthy children and adolescents were calculated with non-linear exponential regression analysis (equation: one-phase decay, least square fit; function: Y = (Y0 – Plateau) × exp(– K × X) + Plateau). Significant differences between groups were assessed by a non-paired two-tailed Mann–Whitney U test. p-Values <0.05 were regarded as significant and are indicated as *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001.
Results
Normalization of CD56bright to CD56dim Ratio Post-HSCT within the First Year Post-HSCT
The distribution of NK cell subpopulations post-HSCT is divergent from healthy individuals showing a high proportion of CD56bright cells shifting toward CD56dim NK cells. To estimate the reconstitution time to a normal CD56bright, CD56int, and CD56dim NK cell distribution and absolute cell counts post-HSCT, we first generated age-matched reference values of NK cell subpopulation frequencies and absolute NK cell counts for healthy children and adolescents (Figure S1 in Supplementary Material). Interestingly, the proportion of CD56bright, CD56int, and CD56dim in early childhood changed from 15, 8, and 78 to 6, 4, and 90% in adults, respectively. Subsequently, we matched the NK cell reconstitution of all patients of the control group without severe events (e.g., GvHD, infections) with the newly generated reference values (Figure 1). The frequency of CD56bright NK cells is elevated directly after HSCT but reaches the upper reference range following 2 months. However, the levels of CD56bright appear slightly increased remaining between the 50th and 90th percentile until 12 months post-HSCT (Figure 1A). Surprisingly, the percentage of CD56int cells matches the reference range within the first 2 months post-HSCT but increases starting from 3 months exceeding the 90th percentile of the reference range following 5 months post-HSCT. After this period, the CD56int fraction declines and converges to the 50th percentile of the reference values 12 months after HSCT (Figure 1B). By contrast, the CD56dim fraction remains below the 10th percentile until 8 months and reaches the 50th percentile of normal reference values after 12 months post-HSCT (Figure 1C). Comparable reconstitution profiles are also apparent for absolute values of CD56bright, CD56int, and CD56dim NK cells. However, absolute values after HSCT seem to be lower than NK cells of healthy children (Figures 1D–F).
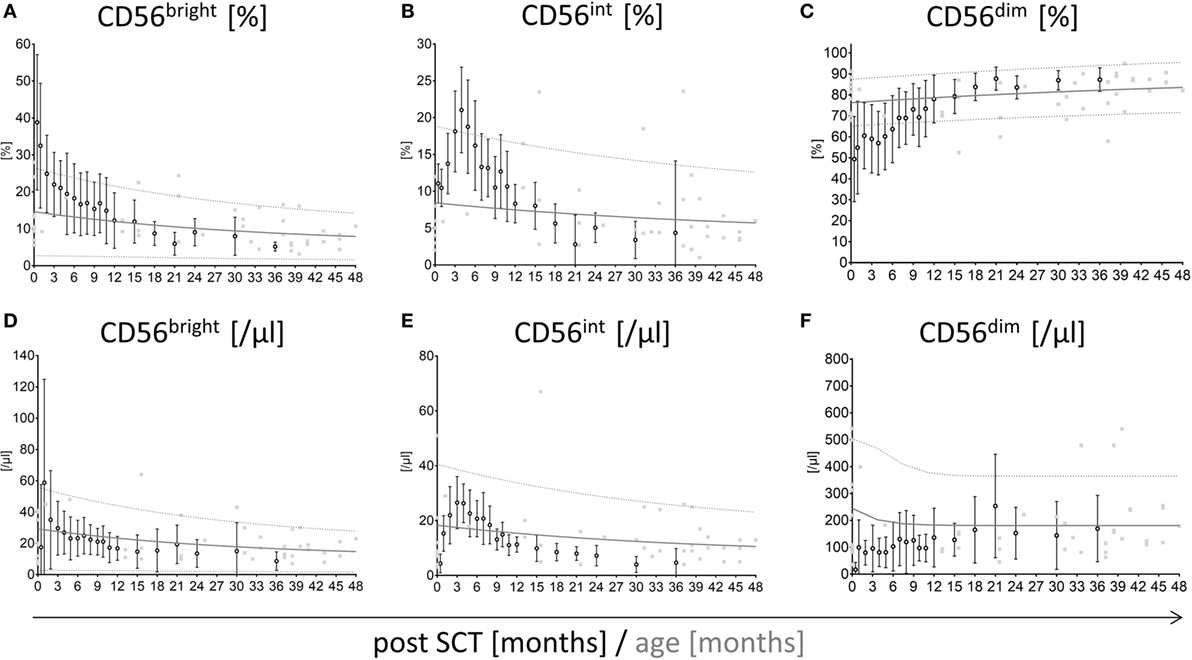
Figure 1. Reconstitution of CD56bright, CD56int, and CD56dim natural killer (NK) cells with respect to reference percentiles of healthy children. Reconstitution of CD56bright, CD56int, and CD56dim NK cells plotted into a graph showing the 10th, 90th (…., dotted line), and 50th ( , solid line) percentile of NK cell reference values of healthy children (n = 174) as well as their underlying measurements (
, solid line) percentile of NK cell reference values of healthy children (n = 174) as well as their underlying measurements ( , gray squares). Measurements of stem cell transplanted patients without graft-versus-host disease or viral infections (n = 23; mean with SD) were plotted into the reference model (○, black rimmed circles) at 19 time points until 36 months after hematopoietic stem cell transplantation (HSCT). Time points were 15 days, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 15, 18, 21, 14, 30, and 36 months after SCT. The CD56bright and CD56dim NK cells post-HSCT needed around 12 months to reach the distribution of healthy individuals NK cells. At that time point, the CD56bright, CD56int, and CD56dim NK cells met the 50th percentile of the reference range (A–C). CD56bright NK cells were elevated post-HSCT but already reached the reference range after 2 months (A). Intermediate NK cells matched the reference range directly after HSCT but showed a fast proliferation until 5 months followed by a decline until 12 months post-HSCT (B). CD56dim NK cells were underrepresented directly after HSCT, reaching the lower reference range after 8 months post-HSCT (C). The development of absolute cell counts of CD56bright, CD56int, and CD56dim NK cells is shown in subfigures (D–F).
, gray squares). Measurements of stem cell transplanted patients without graft-versus-host disease or viral infections (n = 23; mean with SD) were plotted into the reference model (○, black rimmed circles) at 19 time points until 36 months after hematopoietic stem cell transplantation (HSCT). Time points were 15 days, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 15, 18, 21, 14, 30, and 36 months after SCT. The CD56bright and CD56dim NK cells post-HSCT needed around 12 months to reach the distribution of healthy individuals NK cells. At that time point, the CD56bright, CD56int, and CD56dim NK cells met the 50th percentile of the reference range (A–C). CD56bright NK cells were elevated post-HSCT but already reached the reference range after 2 months (A). Intermediate NK cells matched the reference range directly after HSCT but showed a fast proliferation until 5 months followed by a decline until 12 months post-HSCT (B). CD56dim NK cells were underrepresented directly after HSCT, reaching the lower reference range after 8 months post-HSCT (C). The development of absolute cell counts of CD56bright, CD56int, and CD56dim NK cells is shown in subfigures (D–F).
Reconstitution of NK Cells with Regard to GvHD Development
Patients suffering from aGvHD with grade higher than III show significant differences in the reconstitution of NK cell subpopulations compared to patients without any severe events post-HSCT. The median time of first symptoms of aGvHD was 22 days post-HSCT (range: 13–84). Within the first, second, and third month 68, 89, and 100% of the affected patients showed first signs of aGvHD, respectively. Patients without events show conspicuously higher frequency of CD56bright NK cells within the first 3 months following HSCT compared to patients suffering from aGvHD with the most significant difference already 15 days following engraftment (p < 0.0001; Figure 2A). This could also be shown for the absolute CD56bright NK cell amount 1 month after HSCT (p < 0.01; Figure 2D). This tendency was less pronounced when analyzing NK cells in patients with lower grades aGvHD (data not shown). Furthermore, the reconstitution of NK cell subpopulations seems to be delayed in patients suffering from aGvHD. For the CD56int NK cell population, a displacement in time could be shown, which leads to a longer increase in CD56int frequency and absolute amount (Figures 2B,E). To reach the 50th percentile of normal reference values, patients with aGvHD need a prolonged reconstitution time taking at least two times longer compared to patients without events. Patients with lower GvHD grades were lying in between (data not shown). Even after 3 years of monitoring, a trend toward higher frequency of CD56bright and CD56int NK cells concomitant with lower CD56dim was seen in patients suffering from severe aGvHD (not significant, Figures 2A–C). These differences in the NK cell development post-HSCT were also found evaluating the absolute amounts of NK cell subpopulations (Figures 2D–F). Noteworthy, patients affected with aGvHD following HSCT also show a reduced absolute amount of CD56dim NK cells after 3 years post-HSCT (Figure 2F). Analyzing absolute NK cell count (including all three subgroups), we did not see a correlation between patients with and without aGvHD (Figure S2A in Supplementary Material). Analyzing cytotoxic T cells, we detected that patients reaching levels of cytotoxic T cells above 1,500/μl within the first year post-HSCT developed in almost all cases an aGvHD (Figure S2B in Supplementary Material). But this fact was only true for less than 25% of the total aGvHD patient cohort. Furthermore, we evaluated the NK cell regeneration of patients with primary aGvHD grade III or IV that became chronic. Thereby, we detected important differences in the development of NK cell subpopulations from CD56bright above CD56int to CD56dim NK cells. The development from CD56int to CD56dim NK cells is delayed for at least 2 years. Furthermore, patients with chronification of aGvHD with grade >III have a markedly elevated CD56int frequency (Figure 3A), which is also clearly visible in absolute cell count of CD56int NK cells (Figure 3B).
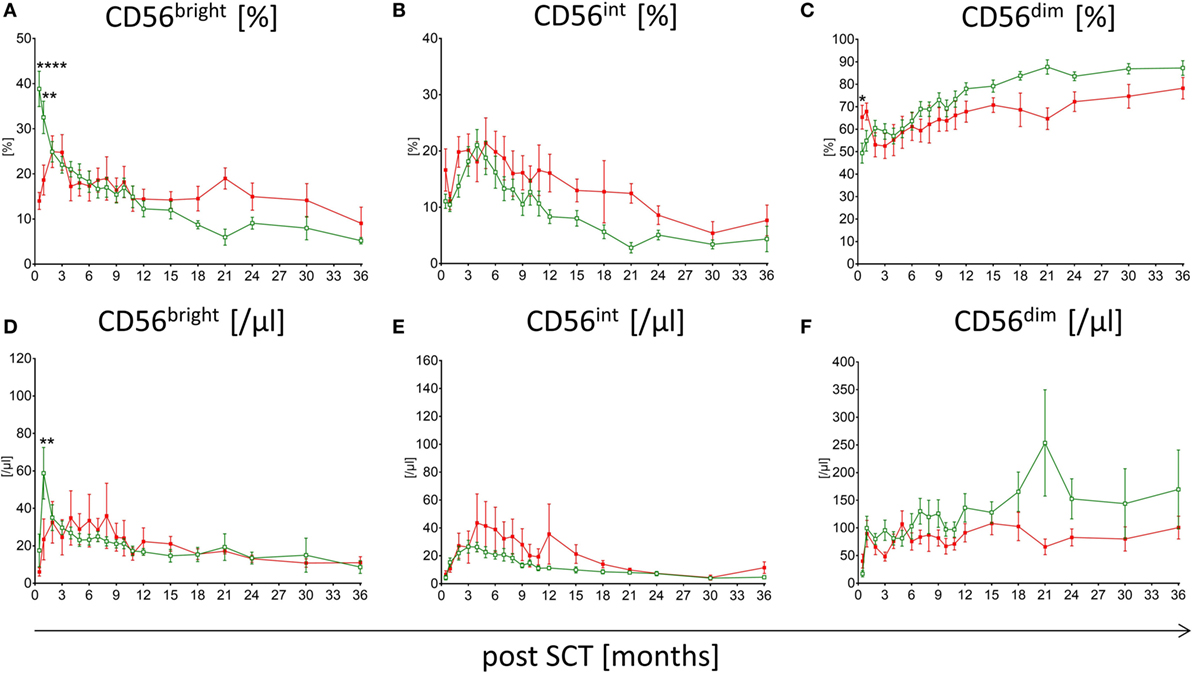
Figure 2. Comparison of natural killer (NK) cell reconstitution in patients suffering from acute graft versus host disease (aGvHD) versus patients without events. Regeneration of CD56bright, CD56int, and CD56dim NK cells in patients affected with severe aGvHD with grade III and IV ( , red squares, n = 19) and patients without events post-hematopoietic stem cell transplantation (HSCT) (
, red squares, n = 19) and patients without events post-hematopoietic stem cell transplantation (HSCT) ( , green rimmed squares, n = 23) at time point of 15 days, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 15, 18, 21, 14, 30, and 36 months after SCT. Especially, in the first 3 months, post-HSCT aGvHD patients showed significantly lower CD56bright NK cells in both percentage (A) (p < 0.0001) and absolute amounts (D) (p < 0.01). CD56int NK cells of aGvHD patients seem delayed in their development as patients without events show an increased intermediate population over a period of at least 18 months (B,E). Except in the beginning, CD56dim NK cell levels of GvHD patients remain below those of patients without events during the whole monitoring interval (C,F). Measurements were available at almost all time points, except for five patients of the aGvHD group who died 2, 3, 9, 9, and 18 months post-HSCT.
, green rimmed squares, n = 23) at time point of 15 days, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 15, 18, 21, 14, 30, and 36 months after SCT. Especially, in the first 3 months, post-HSCT aGvHD patients showed significantly lower CD56bright NK cells in both percentage (A) (p < 0.0001) and absolute amounts (D) (p < 0.01). CD56int NK cells of aGvHD patients seem delayed in their development as patients without events show an increased intermediate population over a period of at least 18 months (B,E). Except in the beginning, CD56dim NK cell levels of GvHD patients remain below those of patients without events during the whole monitoring interval (C,F). Measurements were available at almost all time points, except for five patients of the aGvHD group who died 2, 3, 9, 9, and 18 months post-HSCT.
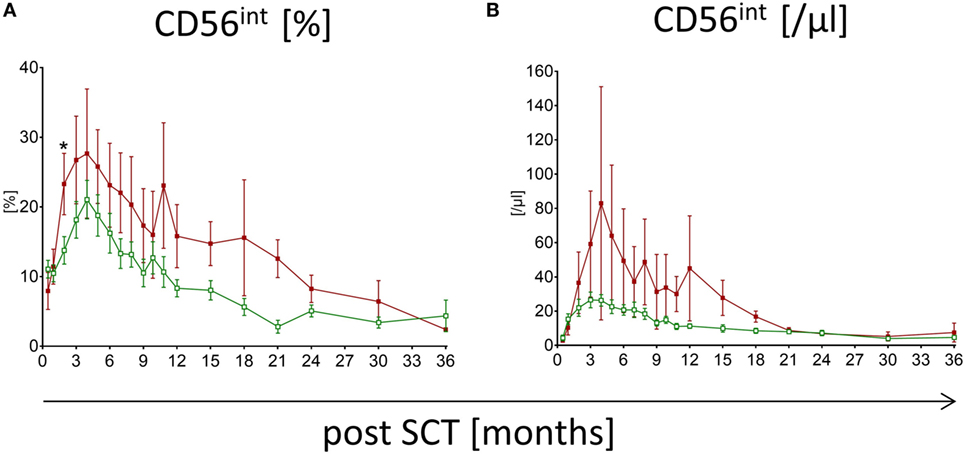
Figure 3. Natural killer (NK) cell reconstitution in patients suffering from chronic graft versus host disease (cGvHD). Development of CD56int NK cells of patients suffering from cGvHD after severe acute graft versus host disease (grade III or IV) is shown for frequency (A) and absolute count (B). Over a period of at least 18 months, a higher amount of CD56int NK cells for patients suffering from cGvHD ( , dark red squares, n = 8) was detectable compared to patients without severe events post-hematopoietic stem cell transplantation (
, dark red squares, n = 8) was detectable compared to patients without severe events post-hematopoietic stem cell transplantation ( , green rimmed squares, n = 23).
, green rimmed squares, n = 23).
Receptor Expression of CD56bright, CD56int, and CD56dim NK Cells Post-HSCT
As part of the patients had low absolute cell counts post-HSCT, a detailed phenotyping of the CD56bright, CD56int, and CD56dim NK cell populations was only possible for an elected cohort of patients. To get an understanding of the function of CD56int NK cells, we analyzed surface molecules linked to NK cell cytotoxicity, adhesion, and immune regulatory functions (e.g., chemokine and cytokine receptors) and compared the expression of KIRs, CD62L, NKG2A, CD127, CD117, CX3CR1, CD226, and CD57 on all three NK subpopulations. The CD56dim population showed a higher expression of KIRs, whereas the CD56int and CD56bright population did not. However, not all KIRs applied within the mix were expressed with equal density (Figure 4). Regarding the homing receptor CD62L, the CD56int, and the CD56bright fraction showed an increased expression compared to CD56dim population. The expression of NKG2A was highest on CD56int and CD56bright cells but bipartite in CD56dim NK cells. CX3CR1 is involved in adhesion and migration of NK cells and to a small extent higher presented on CD56dim NK cells. As already described, we could also show that CD57 was only detectable on the CD56dim subpopulation. Low expression of CD127 (IL7α chain), CD117 (c-Kit), and CD226 (DNAM-1) could be seen on all NK cell subpopulations; however, CD56bright intend to have higher expression than CD56dim, whereas CD56int was always lying in between. In summary, the expression profiles of CD56int and CD56bright NK cells were nearly congruent, but differed to CD56dim cells in KIR, CD62L, NKG2A, CX3CR1, and CD57 expression (Figure 4).
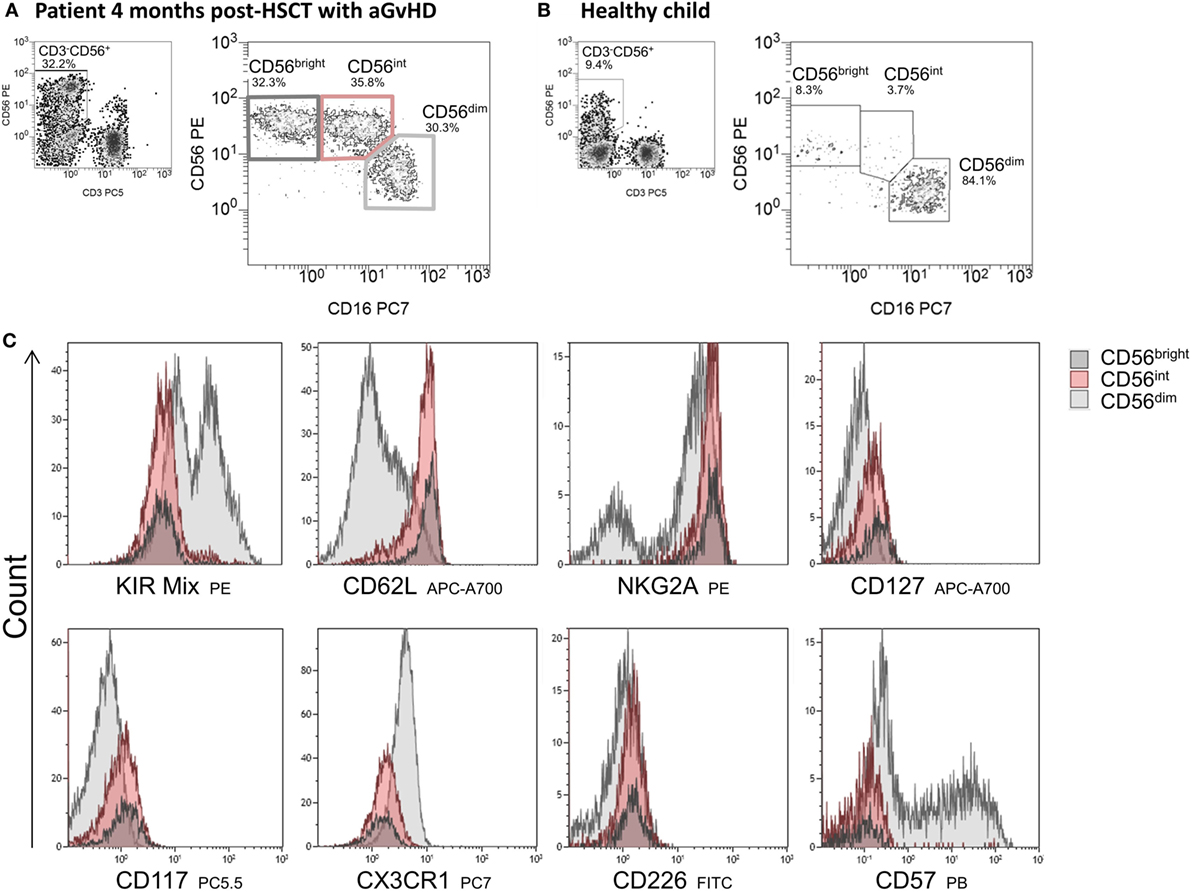
Figure 4. Immunophenotyping of CD56bright, CD56int, and CD56dim cells. Exemplary flow cytometric plots of natural killer (NK) cell subpopulations of a patient 4 months post-hematopoietic stem cell transplantation (HSCT) with GvHD showing an almost equal distribution of CD56bright, CD56int, and CD56dim cells (A) and of a healthy child aged 3 years with mainly CD56dim cytotoxic NK cells (B). NK cell subpopulations were characterized in detail by flow cytometric measurement of surface antigen expression of several ligands involved in adhesion, chemotaxis, and cytotoxicity. Expression of the corresponding receptors on NK cells is shown by overlay plots with MFI on the x-axis of respective antigen in allocation to CD56bright (dark gray), CD56int (red) compared to CD56dim NK cells (light gray). CD56bright and CD56int NK cells show comparable receptor expression, but differ to CD56dim cells in KIR-, CD62L, NKG2A, CX3CR1, and CD57 expression (C).
Influence of Viral Infection Post-HSCT on NK Cell Reconstitution
The immune reconstitution of NK cell subpopulations post-HSCT was analyzed in patients without events and patients suffering from ADV (n = 5), EBV (n = 5), and CMV infection (n = 8). Infection was detected by the routine analysis of DNA copies in peripheral blood. Patients with elevated viral load at the day of transplantation were excluded from the study, resulting in a cohort of patients with occurrence of a positive viral load between 30 and 90 days post-HSCT. Interestingly, we observed a slight reduction in CD56 and CD16 expression in patients suffering from viral infection in between day 30 and day 60 post-HSCT measured by mean fluorescence intensity (Figure 5). After viral clearance in most patients, a considerable loss in absolute CD56dim NK cell count occurred followed by continued regeneration of CD56int NK cells, which was lower in patients without events post-HSCT on day 150 post-HSCT (Figure 5).
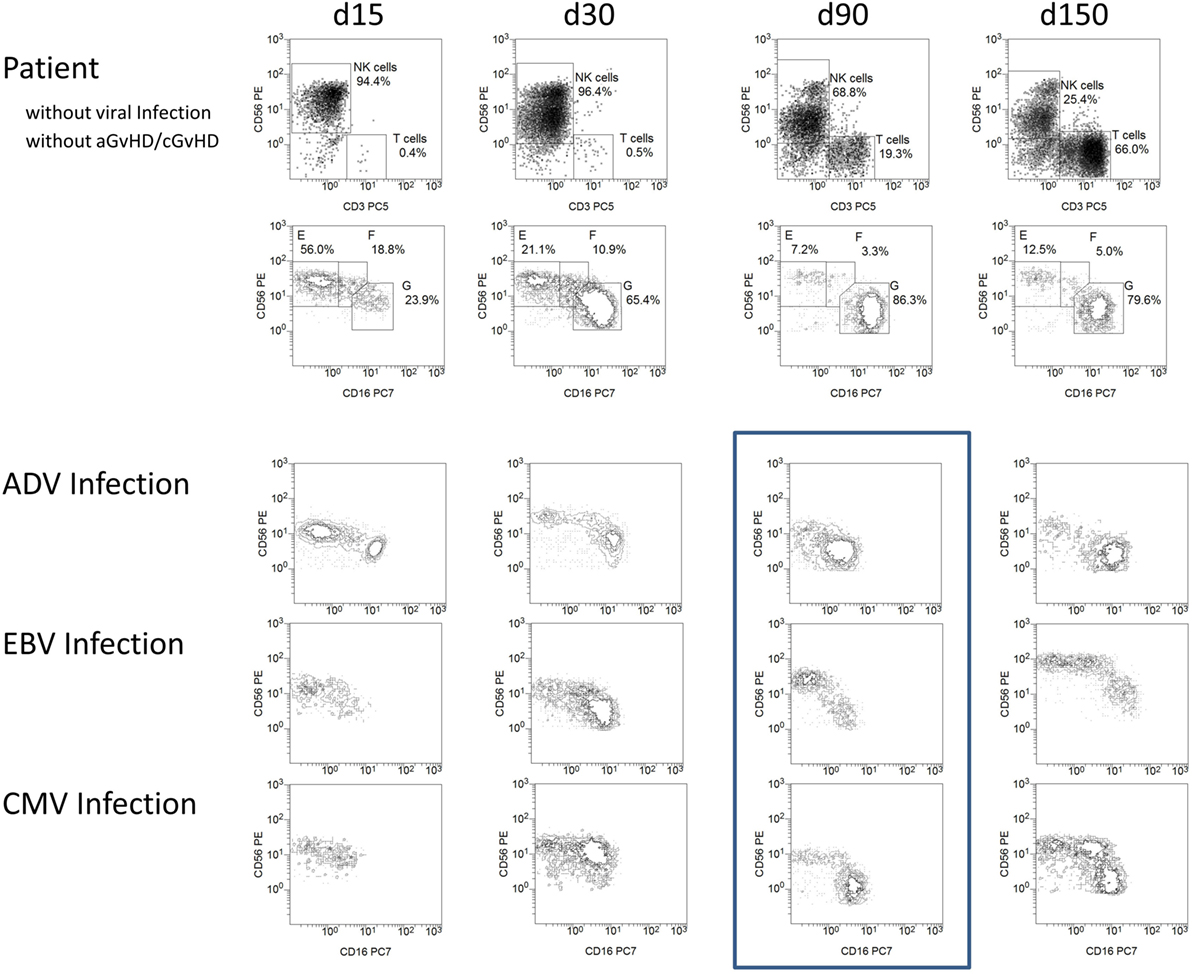
Figure 5. Viral infections influencing natural killer (NK) cell reconstitution and NK marker expression density. Exemplary flow cytometric plots of a patient without events and patients suffering from adenovirus (ADV), Epstein–Barr virus (EBV), and cytomegalovirus (CMV) infection between 15 and 150 days post-HSCT. Patients with high viral load (e.g., ADV above 40,000 genome equivalent/ml and CMV above 90,000 IU/ml) or qualitatively confirmed EBV infection with disease onset between day 30 and day 60 were exemplarily presented. NK cells of patients suffering from viral infection showed a slight reduction in CD56 and CD16 receptor expression 90 days post-HSCT (frame). After 150 days post-HSCT, the NK cells of the patient without events consisted of a major population of CD56dim, a small amount of CD56bright and only few CD56int cells whereas patients after EBV and CMV infection showed a distinct population of CD56int NK cells again. Patients suffering from infections: ADV (n = 5), EBV (n = 5), CMV (n = 8).
Discussion
Especially in the early phase following allogeneic HSCT, together with neutrophils, NK cells are the first line of immune defense. Their immune reconstitution is of crucial importance for transplantation outcome with special regard to the occurrence of GvHD and viral infections. In this project, we analyzed NK cell subpopulations in detail not only focusing on CD56bright and CD56dim cells but also the fraction in between those subsets. With regard to NK cell subpopulation development in young healthy children, we found that it takes around 12 months until CD56bright, CD56int, and CD56dim NK cells of patients post-HSCT reach the 50th percentile of age-matched reference range. Comparable results were published by Pical-Izard et al. describing that rapidly re-emerging NK cells remain immature for more than 6 months (8). Directly after HSCT we detected a highly increased frequency of CD56bright, whereas CD56int NK cells correspond to the reference range, but considerably expand within the first 3 months post-HSCT. In contrast, CD56dim NK cells deserve around 8 months to enter the reference range. These results confirm the hypothesis of sequential development of NK cells with CD56int NK cells representing an intermediate state from CD56bright to CD56dim NK cells (14, 15). These CD56bright NK cells in peripheral blood are closely related to those NK cells populating secondary lymphoid tissues (16). Further evidence supporting this hypothesis was published by Freud et al. describing that the CD56bright subset is the major NK cell population that is derived early in vitro when CD34+ HPC are cultured in NK development supportive conditions, whereas CD56dim NK cells develop later (17). Furthermore, CD56bright NK cells display longer telomeres than the CD56dim NK cells, indicating lower proliferation capacity (6).
We further characterized all three NK cell subpopulations with the finding, that CD56int presented antigen expressions among CD56bright and CD56dim NK cells, even so CD56bright and CD56int NK cells showed rather equal expression profiles and seemed related more to CD56bright. However, differential expression of KIRs, CD62L, NKG2A, and CD57 was observed on CD56dim NK cells. This is in parallel to other findings describing an increased expression of NKG2A, the IL-7 receptor (CD127) and the lymph node homing receptor CCR7 on CD56bright cells (2, 5, 8, 18, 19) whereas CD56dim NK cells acquire KIR, NKG2C, and CD57 expression (20).
Promoted by the IL-15 rich cytokine milieu post-transplant, NK cells are known to be one of the first lymphocyte subpopulation recovering post-HSCT (21). Therefore, NK cell reconstitution might be the basis for generating early prognostic markers regarding the occurrence of severe events and transplantation outcome. Kim et al. published that NK cell counts after allo-HSCT, especially on day 30, were predictive markers for GvHD, non-relapse mortality, and survival (22). Furthermore, there is evidence that the speed of NK cell reconstitution correlates with transplant outcome, suggesting their important role in the early period when specific T cell immunity is absent (7, 8). Our and other findings suggest that the monitoring of NK cell subsets in the early phase post-HSCT might provide first signs of aGvHD development (23). Interestingly, within the first 2 months post-HSCT patients without aGvHD or viral infections had significantly elevated levels of CD56bright NK cells compared to patients suffering from aGvHD. This might be an early prognostic factor regarding GvHD development; however, it needs to be confirmed in a prospective study. Likewise results were also published by Kheav et al. showing an impaired reconstitution of CD56dim NK cells 3 months post-HSCT (24). We also found a comparable trend for NK cell regeneration in patients suffering from cGvHD, although not significant (data not shown). This might be explained by the fact, that for aGvHD analysis, only patients suffering from GvHD grades III and IV were considered, whereas no differentiation was available regarding cGvHD (e.g., chronification of primary aGvHD grades I and II).
Literature is discordant whether steroids/immunosuppression have a negative impact on NK cell reconstitution. Giebel et al. proposed that the use of steroids for GvHD prophylaxis negatively affects quantitative reconstitution of NK cells after allo-HSCT (25). Although, patients suffering from GvHD grades III and IV normally receive steroids in our transplantation unit, we did not see any significant differences in the quantitative reconstitution of absolute NK cell numbers. Interestingly, Wang et al. described that CSA suppresses the in vitro proliferation of NK cells, especially the CD56dimCD16+KIR+ NK cells, resulting in a relative increase in the number of immature CD56brightCD16−KIR− NK cells (26). This might also contribute to the delayed NK cell development in patients suffering from higher grade aGvHD that we observed within this study. However, this remains controversial as other studies analyzed the effect of CSA on NK cell function in short-term cultures and their cytokine production without finding significant differences between NK cells with and without CSA treatment (27, 28).
In patients suffering from viral infection post-HSCT, we observed a slight reduction in CD56 and CD16 expression. Notably, other publications already described the existence of CD56−CD16+ NK cells (CD56negative) NK cells in viral infections (e.g., HIV, hepatitis C), where NK cells undergo numerous phenotypic and functional changes (29). This CD56negative subset has been associated with high HIV viral load and has been reported to have an impaired cytolytic function and cytokine production (30). This increase occurred primarily at the expense of CD56dim NK cells, whereas numbers of CD56bright NK cells remained stable (31). Furthermore, we observed a considerable loss in absolute CD56dim NK cells followed by continued regeneration of CD56int NK cells. Alteration of NK cells upon viral infection has already been shown by other research groups, for example Pical-Izard et al. showed that in patients being affected by CMV reactivation, NK cells showed lower degranulation and TNF-α production compared to patients without CMV reactivation post-HSCT (8). In addition, it was shown that CMV reactivation is followed by an increase in the proportion of NKG2C+ NK cells within 2–4 weeks, which persist for at least a year (32, 33).
In conclusion, only after around 12 months, NK cells post-HSCT reconstitute to a distribution of the subpopulations CD56bright, CD56int, and CD56dim comparable to age-matched healthy controls. The expression profiles of CD56int and CD56bright NK cells resemble each other but differed in KIR, CD62L, NKG2A, CX3CR1, and CD57 expression to CD56dim NK cells. We observed elevated levels of CD56bright directly after and CD56int NK cells 3 months post-HSCT accompanied by reduced CD56dim NK cells supporting the hypothesis of sequential NK cell development. Furthermore, we analyzed alterations in NK cell development in patients with severe viral infections and GvHD. Following viral infection, there was a slight reduction in CD56 and CD16 receptor expression followed by a considerable loss in absolute CD56dim NK cells and continued regeneration of CD56int NK cells. Most important, within the first 2 months, post-HSCT patients without severe events had significantly elevated levels of CD56bright NK cells compared to patients suffering from aGvHD. While first measurements performed as early as 15 days following HSCT revealed the most significant differences, clinical occurrence of aGvHD was observed in median on day 22 post-HSCT. Therefore, we recommend immunophenotyping of NK cell subpopulations directly following engraftment, which might be an early prognostic factor regarding GvHD development.
Author Contributions
Study design: MB, SH, RE, and EK. Performed the experiments: JB-D and SBe. Provided clinical data: JS, AJ, SBa, and PB. Analyzed the data: SH and EK. Coordinated the research: SH and CC. Contributed reagents/materials/analysis tools: MB, SH, VP, JB-D, and SBe. Performed statistical analyses: SH and ES-M. Wrote the manuscript: MB and SH. Discussed data and revised the manuscript: CC, PB, VP, EU, and CK. Supervised the research: PB and TK. All the authors read and approved the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank Stephanie Erben, Olga Zimmermann, Regine Müller, Andrea Quaiser, Tanja Gardlowski, and Michael Marburger for the excellent technical support.
Funding
This project was supported by “Frankfurter Stiftung für krebskranke Kinder,” “Hilfe für krebskranke Kinder e.V.,” Edith-von-Heyden-Vermächtnis-Stiftung, the German Cancer Aid (Max Eder Nachwuchsgruppe, Deutsche Krebshilfe; EU), and the LOEWE Center for Cell and Gene Therapy (to TK, PB, and EU), Frankfurt, funded by the Hessian Ministry of Higher Education, Research, and the Arts, Germany (III L 4-518/17.004). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at http://journal.frontiersin.org/article/10.3389/fimmu.2017.00109/full#supplementary-material.
References
1. Farag SS, Caligiuri MA. Human natural killer cell development and biology. Blood Rev (2006) 20(3):123–37. doi: 10.1016/j.blre.2005.10.001
2. Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S. Functions of natural killer cells. Nat Immunol (2008) 9(5):503–10. doi:10.1038/ni1582
3. Ullah MA, Hill GR, Tey SK. Functional reconstitution of natural killer cells in allogeneic hematopoietic stem cell transplantation. Front Immunol (2016) 7:144. doi:10.3389/fimmu.2016.00144
4. Jacobs R, Stoll M, Stratmann G, Leo R, Link H, Schmidt RE. CD16- CD56+ natural killer cells after bone marrow transplantation. Blood (1992) 79(12):3239–44.
5. Giebel S, Dziaczkowska J, Czerw T, Wojnar J, Krawczyk-Kulis M, Nowak I, et al. Sequential recovery of NK cell receptor repertoire after allogeneic hematopoietic SCT. Bone Marrow Transplant (2010) 45(6):1022–30. doi:10.1038/bmt.2009.384
6. Romagnani C, Juelke K, Falco M, Morandi B, D’Agostino A, Costa R, et al. CD56brightCD16- killer Ig-like receptor- NK cells display longer telomeres and acquire features of CD56dim NK cells upon activation. J Immunol (2007) 178(8):4947–55. doi:10.4049/jimmunol.178.8.4947
7. Savani BN, Mielke S, Adams S, Uribe M, Rezvani K, Yong AS, et al. Rapid natural killer cell recovery determines outcome after T-cell-depleted HLA-identical stem cell transplantation in patients with myeloid leukemias but not with acute lymphoblastic leukemia. Leukemia (2007) 21(10):2145–52. doi:10.1038/sj.leu.2404892
8. Pical-Izard C, Crocchiolo R, Granjeaud S, Kochbati E, Just-Landi S, Chabannon C, et al. Reconstitution of natural killer cells in HLA-matched HSCT after reduced-intensity conditioning: impact on clinical outcome. Biol Blood Marrow Transplant (2015) 21(3):429–39. doi:10.1016/j.bbmt.2014.11.681
9. Ruggeri L, Capanni M, Urbani E, Perruccio K, Shlomchik WD, Tosti A, et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science (2002) 295(5562):2097–100. doi:10.1126/science.1068440
10. Koehl U, Brehm C, Huenecke S, Zimmermann SY, Kloess S, Bremm M, et al. Clinical grade purification and expansion of NK cell products for an optimized manufacturing protocol. Front Oncol (2013) 3:118. doi:10.3389/fonc.2013.00118
11. Stern M, Passweg JR, Meyer-Monard S, Esser R, Tonn T, Soerensen J, et al. Pre-emptive immunotherapy with purified natural killer cells after haploidentical SCT: a prospective phase II study in two centers. Bone Marrow Transplant (2013) 48(3):433–8. doi:10.1038/bmt.2012.162
12. Meinhardt K, Kroeger I, Bauer R, Ganss F, Ovsiy I, Rothamer J, et al. Identification and characterization of the specific murine NK cell subset supporting graft-versus-leukemia- and reducing graft-versus-host-effects. Oncoimmunology (2015) 4(1):e981483. doi:10.4161/2162402X.2014.981483
13. Koenig M, Huenecke S, Salzmann-Manrique E, Esser R, Quaritsch R, Steinhilber D, et al. Multivariate analyses of immune reconstitution in children after allo-SCT: risk-estimation based on age-matched leukocyte sub-populations. Bone Marrow Transplant (2010) 45(4):613–21. doi:10.1038/bmt.2009.204
14. Beziat V, Duffy D, Quoc SN, Le Garff-Tavernier M, Decocq J, Combadiere B, et al. CD56brightCD16+ NK cells: a functional intermediate stage of NK cell differentiation. J Immunol (2011) 186(12):6753–61. doi:10.4049/jimmunol.1100330
15. Gismondi A, Stabile H, Nisti P, Santoni A. Effector functions of natural killer cell subsets in the control of hematological malignancies. Front Immunol (2015) 6:567. doi:10.3389/fimmu.2015.00567
16. Vukicevic M, Chalandon Y, Helg C, Matthes T, Dantin C, Huard B, et al. CD56bright NK cells after hematopoietic stem cell transplantation are activated mature NK cells that expand in patients with low numbers of T cells. Eur J Immunol (2010) 40(11):3246–54. doi:10.1002/eji.200940016
17. Freud AG, Yu J, Caligiuri MA. Human natural killer cell development in secondary lymphoid tissues. Semin Immunol (2014) 26(2):132–7. doi:10.1016/j.smim.2014.02.008
18. Cooper MA, Fehniger TA, Caligiuri MA. The biology of human natural killer-cell subsets. Trends Immunol (2001) 22(11):633–40. doi:10.1016/S1471-4906(01)02060-9
19. Nguyen S, Dhedin N, Vernant JP, Kuentz M, Al Jijakli A, Rouas-Freiss N, et al. NK-cell reconstitution after haploidentical hematopoietic stem-cell transplantations: immaturity of NK cells and inhibitory effect of NKG2A override GvL effect. Blood (2005) 105(10):4135–42. doi:10.1182/blood-2004-10-4113
20. Bjorkstrom NK, Riese P, Heuts F, Andersson S, Fauriat C, Ivarsson MA, et al. Expression patterns of NKG2A, KIR, and CD57 define a process of CD56dim NK-cell differentiation uncoupled from NK-cell education. Blood (2010) 116(19):3853–64. doi:10.1182/blood-2010-04-281675
21. Boyiadzis M, Memon S, Carson J, Allen K, Szczepanski MJ, Vance BA, et al. Up-regulation of NK cell activating receptors following allogeneic hematopoietic stem cell transplantation under a lymphodepleting reduced intensity regimen is associated with elevated IL-15 levels. Biol Blood Marrow Transplant (2008) 14(3):290–300. doi:10.1016/j.bbmt.2007.12.490
22. Kim SY, Lee H, Han MS, Shim H, Eom HS, Park B, et al. Post-transplantation natural killer cell count: a predictor of acute graft-versus-host disease and survival outcomes after allogeneic hematopoietic stem cell transplantation. Clin Lymphoma Myeloma Leuk (2016) 16(9):527.e–35.e. doi:10.1016/j.clml.2016.06.013
23. Ullrich E, Salzmann-Manrique E, Bakhtiar S, Bremm M, Gerstner S, Herrmann E, et al. Relation between acute GVHD and NK cell subset reconstitution following allogeneic stem cell transplantation. Front Immunol (2016) 7:595. doi:10.3389/fimmu.2016.00595
24. Kheav VD, Busson M, Scieux C, Peffault de Latour R, Maki G, Haas P, et al. Favorable impact of natural killer cell reconstitution on chronic graft-versus- host disease and cytomegalovirus reactivation after allogeneic hematopoietic stem cell transplantation. Haematologica (2014) 99(12):1860–7. doi:10.3324/haematol.2014.108407
25. Giebel S, Dziaczkowska J, Wojnar J, Krawczyk-Kulis M, Markiewicz M, Kruzel T, et al. The impact of immunosuppressive therapy on an early quantitative NK cell reconstitution after allogeneic haematopoietic cell transplantation. Ann Transplant (2005) 10(2):29–33.
26. Wang H, Grzywacz B, Sukovich D, McCullar V, Cao Q, Lee AB, et al. The unexpected effect of cyclosporin A on CD56+CD16- and CD56+CD16+ natural killer cell subpopulations. Blood (2007) 110(5):1530–9. doi:10.1182/blood-2006-10-048173
27. Shao-Hsien C, Lang I, Gunn H, Lydyard P. Effect of in vitro cyclosporin. A treatment on human natural and antibody-dependent cell-mediated cytotoxicity. Transplantation (1983) 35(2):127–9. doi:10.1097/00007890-198302000-00004
28. Derniame S, Perazzo J, Lee F, Domogala A, Escobedo-Cousin M, Alnabhan R, et al. Differential effects of mycophenolate mofetil and cyclosporine A on peripheral blood and cord blood natural killer cells activated with interleukin-2. Cytotherapy (2014) 16(10):1409–18. doi:10.1016/j.jcyt.2014.05.010
29. Fauci AS, Mavilio D, Kottilil S. NK cells in HIV infection: paradigm for protection or targets for ambush. Nat Rev Immunol (2005) 5(11):835–43. doi:10.1038/nri1711
30. Mavilio D, Lombardo G, Benjamin J, Kim D, Follman D, Marcenaro E, et al. Characterization of CD56-/CD16+ natural killer (NK) cells: a highly dysfunctional NK subset expanded in HIV-infected viremic individuals. Proc Natl Acad Sci U S A (2005) 102(8):2886–91. doi:10.1073/pnas.0409872102
31. Bjorkstrom NK, Ljunggren HG, Sandberg JK. CD56 negative NK cells: origin, function, and role in chronic viral disease. Trends Immunol (2010) 31(11):401–6. doi:10.1016/j.it.2010.08.003
32. Beziat V, Dalgard O, Asselah T, Halfon P, Bedossa P, Boudifa A, et al. CMV drives clonal expansion of NKG2C+ NK cells expressing self-specific KIRs in chronic hepatitis patients. Eur J Immunol (2012) 42(2):447–57. doi:10.1002/eji.201141826
Keywords: NK cells, immune reconstitution, CD56, CD16, allogeneic transplantation, children, reference values
Citation: Huenecke S, Cappel C, Esser R, Pfirrmann V, Salzmann-Manrique E, Betz S, Keitl E, Banisharif-Dehkordi J, Bakhtiar S, Königs C, Jarisch A, Soerensen J, Ullrich E, Klingebiel T, Bader P and Bremm M (2017) Development of Three Different NK Cell Subpopulations during Immune Reconstitution after Pediatric Allogeneic Hematopoietic Stem Cell Transplantation: Prognostic Markers in GvHD and Viral Infections. Front. Immunol. 8:109. doi: 10.3389/fimmu.2017.00109
Received: 01 November 2016; Accepted: 23 January 2017;
Published: 10 February 2017
Edited by:
Gianfranco Pittari, Hamad Medical Corporation, QatarReviewed by:
Nicholas D. Huntington, Walter and Eliza Hall Institute of Medical Research, AustraliaAnne Caignard, Institut National de la Santé et de la Recherche Médicale, France
Copyright: © 2017 Huenecke, Cappel, Esser, Pfirrmann, Salzmann-Manrique, Betz, Keitl, Banisharif-Dehkordi, Bakhtiar, Königs, Jarisch, Soerensen, Ullrich, Klingebiel, Bader and Bremm. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sabine Huenecke, c2FiaW5lLmh1ZW5lY2tlQGtndS5kZQ==