 Paula Carasi1
Paula Carasi1 Ernesto Rodríguez1
Ernesto Rodríguez1 Valeria da Costa1
Valeria da Costa1 Sofía Frigerio1
Sofía Frigerio1 Natalie Brossard1
Natalie Brossard1 Verónica Noya1
Verónica Noya1 Carlos Robello2,3
Carlos Robello2,3 Ignacio Anegón4,5
Ignacio Anegón4,5 Teresa Freire1*
Teresa Freire1*
- 1Laboratorio de Inmunomodulación y Desarrollo de Vacunas, Facultad de Medicina, Departamento de Inmunobiología, Universidad de República, Montevideo, Uruguay
- 2Departamento de Bioquimica, Facultad de Medicina, Universidad de la República, Montevideo, Uruguay
- 3Unidad de Biología Molecular, Institut Pasteur de Montevideo, Montevideo, Uruguay
- 4Centre de Recherche en Transplantation et Immunologie UMR1064, INSERM, Université de Nantes, CHU Nantes, Nantes, France
- 5Institut de Transplantation Urologie Néphrologie (ITUN), CHU Nantes, Nantes, France
Fasciola hepatica, also known as the liver fluke, is a trematode that infects livestock and humans causing fasciolosis, a zoonotic disease of increasing importance due to its worldwide distribution and high economic losses. This parasite immunoregulates the host immune system by inducing a strong Th2 and regulatory T immune response by immunomodulating dendritic cell (DC) maturation and alternative activation of macrophages. In this paper, we show that F. hepatica infection in mice induces the upregulation of heme-oxygenase-1 (HO-1), the rate-limiting enzyme in the catabolism of free heme that regulates the host inflammatory response. We show and characterize two different populations of antigen presenting cells that express HO-1 during infection in the peritoneum of infected animals. Cells that expressed high levels of HO-1 expressed intermediate levels of F4/80 but high expression of CD11c, CD38, TGFβ, and IL-10 suggesting that they correspond to regulatory DCs. On the other hand, cells expressing intermediate levels of HO-1 expressed high levels of F4/80, CD68, Ly6C, and FIZZ-1, indicating that they might correspond to alternatively activated macrophages. Furthermore, the pharmacological induction of HO-1 with the synthetic metalloporphyrin CoPP promoted F. hepatica infection increasing the clinical signs associated with the disease. In contrast, treatment with the HO-1 inhibitor SnPP protected mice from parasite infection, indicating that HO-1 plays an essential role during F. hepatica infection. Finally, HO-1 expression during F. hepatica infection was associated with TGFβ and IL-10 levels in liver and peritoneum, suggesting that HO-1 controls the expression of these immunoregulatory cytokines during infection favoring parasite survival in the host. These results contribute to the elucidation of the immunoregulatory mechanisms induced by F. hepatica in the host and provide alternative checkpoints to control fasciolosis.
Introduction
Fasciolosis, a helminth infection caused by Fasciola hepatica, is of paramount importance due to its wide spectrum of definitive hosts (1) and its worldwide distribution (2) affecting both livestock and human health. World Health Organization (WHO) estimates that at least 2.4 million people are infected in more than 70 countries worldwide, with several million at risk. Several studies have independently demonstrated that F. hepatica-derived molecules inhibit or decrease dendritic cell (DC) activation, which results in the induction of a tolerogenic phenotype (3–7). Furthermore, we have demonstrated that DCs from mice infected with F. hepatica have a semi-mature phenotype that is characterized by low MHC II and CD40 expression, high secretion of the immunoregulatory cytokine IL-10, and the ability to differentiate and expand IL-10-producing CD4 T cells (8). In addition, different groups have reported that F. hepatica-derived molecules also modulate macrophage activation, inducing the alternative activation of IL-10-producing macrophages (9, 10) and inhibiting the production of pro-inflammatory cytokines, such as IL-1β (11), IL-10 (12), Arg-1, PDL-1 (13), and PDL-2 (14, 15). Thus, it has been hypothesized that F. hepatica may modulate both macrophages and DC function and fate as a mean to control its pathogenesis and survival in the infected hosts.
Heme-oxygenase-1 (HO-1), the rate-limiting enzyme in the catabolism of free heme, is involved in many physiological and pathophysiological processes, by affording cytoprotection (16) and regulating the host inflammatory response. Indeed, HO-1 is a stress-responsive enzyme important for defense against oxidant-induced injury during inflammatory processes and is highly inducible by a variety of stimuli, such as LPS, cytokines, heat shock, heavy metals, oxidants, and its substrate heme. Several works confirm that HO-1 plays a role in different infectious diseases, and can have both beneficial and detrimental consequences for the host immunity against pathogens (17). For instance, HO-1 is able to promote Plasmodium liver infection (18), whereas it plays a favorable role in the host during cerebral malaria (19). On the other hand, HO-1 controls a variety of infections in mice, including Mycobacterium avium (20), Listeria monocytogenes (21), Plasmodium falciparum (22), Salmonella typhimurium (23), Toxoplasma gondii (24), and respiratory syncytial virus (25).
Expression of HO-1 in monocyte-derived DC inhibits LPS-induced maturation and reactive oxygen species production (26). In addition, HO-1+ DCs express the anti-inflammatory cytokine IL-10 resulting in the inhibition of alloreactive T-cell proliferation (26). Also, IL-10-producing anti-inflammatory macrophages (M2) express HO-1 (27). Thus, HO-1 has been proposed to be key mediator of the anti-inflammatory effects of macrophages and DCs.
In the present study, we demonstrate that during infection with the trematode F. hepatica, HO-1 is upregulated by immune cells expressing F4/80 in the peritoneal cavity and liver. We also show that the pharmacological induction of HO-1 with the synthetic metalloporphyrin CoPP promotes F. hepatica infection increasing the clinical signs associated with the disease, such as liver damage. Moreover, treatment with the HO-1 inhibitor SnPP protected from parasite infection. The increase of HO-1 during F. hepatica infection was associated with the increase of TGFβ and IL-10 in liver and peritoneal exudate cells (PECs). Interestingly, we identified two different F4/80+ cell populations that expressed HO-1. HO-1hi F4/80int cells were characterized by the expression of CD11c, CD38, TGFβ, and IL-10 suggesting that they correspond to regulatory DCs. On the other hand, HO-1int F4/80hi cells expressed high levels of CD68, Ly6C, and FIZZ-1 indicating that they might be alternatively activated macrophages. Our results contribute to the elucidation of immunoregulatory mechanisms induced by F. hepatica in the host and could provide alternative checkpoints to control fasciolosis.
Materials and Methods
Ethics Statement
Mouse experiments were carried out in accordance with strict guidelines from the National Committee on Animal Research (Comisión Nacional de Experimentación Animal, CNEA, National Law 18.611, Uruguay) according to the international statements on animal use in biomedical research from the Pan American Health Organization and WHO. The protocol was approved by the Uruguayan Committee on Animal Research. Cattle’s livers were collected during the routine work of a local abattoir (Frigorífico Carrasco) in Montevideo (Uruguay).
Mice
Six- to eight-week-old female BALB/c mice were obtained from DILAVE Laboratories (Uruguay). Animals were kept in the animal house (URBE, Facultad de Medicina, UdelaR, Uruguay) with water and food supplied ad libitum. Mouse handling and experiments were carried out in accordance with strict guidelines from the National Committee on Animal Research (CNEA, Uruguay). All procedures involving animals were approved by the Universidad de la República’s Committee on Animal Research (CHEA Protocol Number: 070153-000180-16).
Infections and Cell Cultures
BALB/c mice were orally infected with 10 F. hepatica metacercariae (Baldwin Aquatics, USA) per animal. After 1, 2, or 3 weeks post-infection (wpi) mice were bled and PECs, spleens, and livers were removed. In order to evaluate the severity of the infection, a disease severity score was developed (Table 1), which was applied in blinded experiments by two independent experimenters. Alanine aminotransferase (ALT) activity in sera was determined by using a commercial kit (Spinreact, Spain) according to the manufacturers’ instructions. PECs from infected and non-infected mice were washed twice with PBS containing 2% FBS and 0.1% sodium azide. The following antibodies were used in these experiments anti-CD11c (N418), -I-A/I-E (2G9), CD40 (HM40-3), -F4/80 (BM8), -CD11b (M1/70), -CD172a (P84), -Ly6C (HK1.4), and -Siglec-F (E50-2440). Cells were then washed twice with PBS containing 2% FBS and 0.1% sodium azide and fixed with 1% formaldehyde. Cell populations were analyzed using a BD FACSCalibur (BD-Biosciences) or Cyan (Beckman Coulter). Expression of HO-1 (ab13248) and CD68 (FA-11) were analyzed by intracellular staining. Antibodies were obtained from Affymetrix (CA, USA), from BD-Biosciences (CA, USA), from Biolegend (CA, USA) or from Abcam.
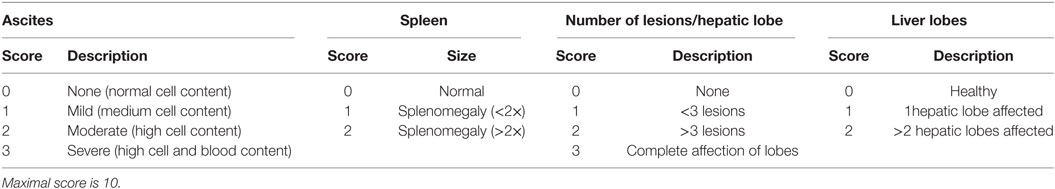
Table 1. Clinical score of Fasciola hepatica-infected mice.
Pharmacological Induction or Inhibition of HO-1
In order to modulate HO-1 activity, mice infected with five metacercariae also received intraperitoneal injections of either vehicle (PBS, 100 µL), CoPP (20 mg/kg), SnPP (40 mg/kg), or CoPP plus SnPP. The doses of CoPP and SnPP were within a range of doses used in studies describing upregulation of HO-1 by CoPP and inhibition of the enzyme’s activity by SnPP (28, 29). Mice were injected 1 day before infection, 1 day after infection and every 5 days until the end of the experimental protocol.
Quantitative Real-time RT-PCR
Total RNA was isolated by use of TRI-reagent (Sigma-Aldrich) from spleen, liver, PEC and purified F4/80int and F4/80hi cells from PEC. Samples were analyzed in an Eco real-time PCR System (Illumina) using Fast SYBR® Green Master Mix (Applied Biosystems). The reactions were performed according to the following settings: 95°C for 5 min for initial activation, followed by 40 thermal cycles of 10 s at 95°C and 30 s at 60°C. All reactions were performed with at least five biological replicates.
Microscopy Analyses
Livers from infected mice after 3 wpi or non-infected mice (control) were harvested, embedded in OCT, and snap frozen in nitrogen. Sections were cut at a thickness of 6 µm, fixed with cold acetone for 10 min and blocked with 5% BSA in 3% rat serum for 1 h at room temperature. Sections were then overnight incubated at 4°C with anti-HO-1 (ab13248) and -F4/80 (BM8), stained with DAPI and visualized in an epifluorencense microscope Olympus IX-81 and confocal microscope Leica TCS-SP5-II. The same procedure and the same antibody were used to evaluate HO-1 expression in bovine livers from naturally infected and non-infected cattle. In this case, livers were first examined by the veterinary inspector at the abattoir and determined to be infected by the presence of multiple parasites found in the bile ducts. Livers from non-infected animals were identified by absence of liver damage and flukes.
Statistical Analysis
Results were analyzed using GraphPad Prism software (GraphPad Software, San Diego, CA, USA). Normality distribution was evaluated using the D’Agostino-Pearson omnibus normality test followed by one-way ANOVA with Bonferroni Multiple Comparison test or a student’s T test was used. Results were considered to be significantly different when p < 0.05 (*), 0.01 (**), or 0.001 (***).
Results
HO-1 Expression Is Induced in F. hepatica-Infected Animals
We first evaluated whether HO-1 was expressed in F. hepatica-infected animals. To this end, mice were infected with 10 metacercariae and after 3 wpi, livers, spleens, and PECs were removed and HO-1 expression was analyzed by qRT-PCR, microscopy, and flow cytometry. Livers from infected mice expressed high levels of HO-1, both at the mRNA (Figure 1A) and protein levels (Figure 1B). Indeed, a 25-fold increase in the transcript levels was determined by qRT-PCR with respect to non-infected animals (Figure 1A). HO-1 expression was found both in the leukocyte infiltrates and the liver parenchyma (Figure 1B), while undetectable levels of HO-1 were found in control livers from naive mice (Figure 1B). HO-1 gene expression was also induced in PECs, revealing, similar to liver, a 25-fold increased in PECs from infected animals, comparing to control mice (Figure 1C). Moreover, HO-1+ cells were detected in the peritoneum both by flow cytometry and microscopy (Figures 1D–F). On the contrary, we failed to detect an increase in HO-1 transcript levels and protein expression by flow cytometry in spleens from infected- with respect to control animals (Figure S1 in Supplementary Material).
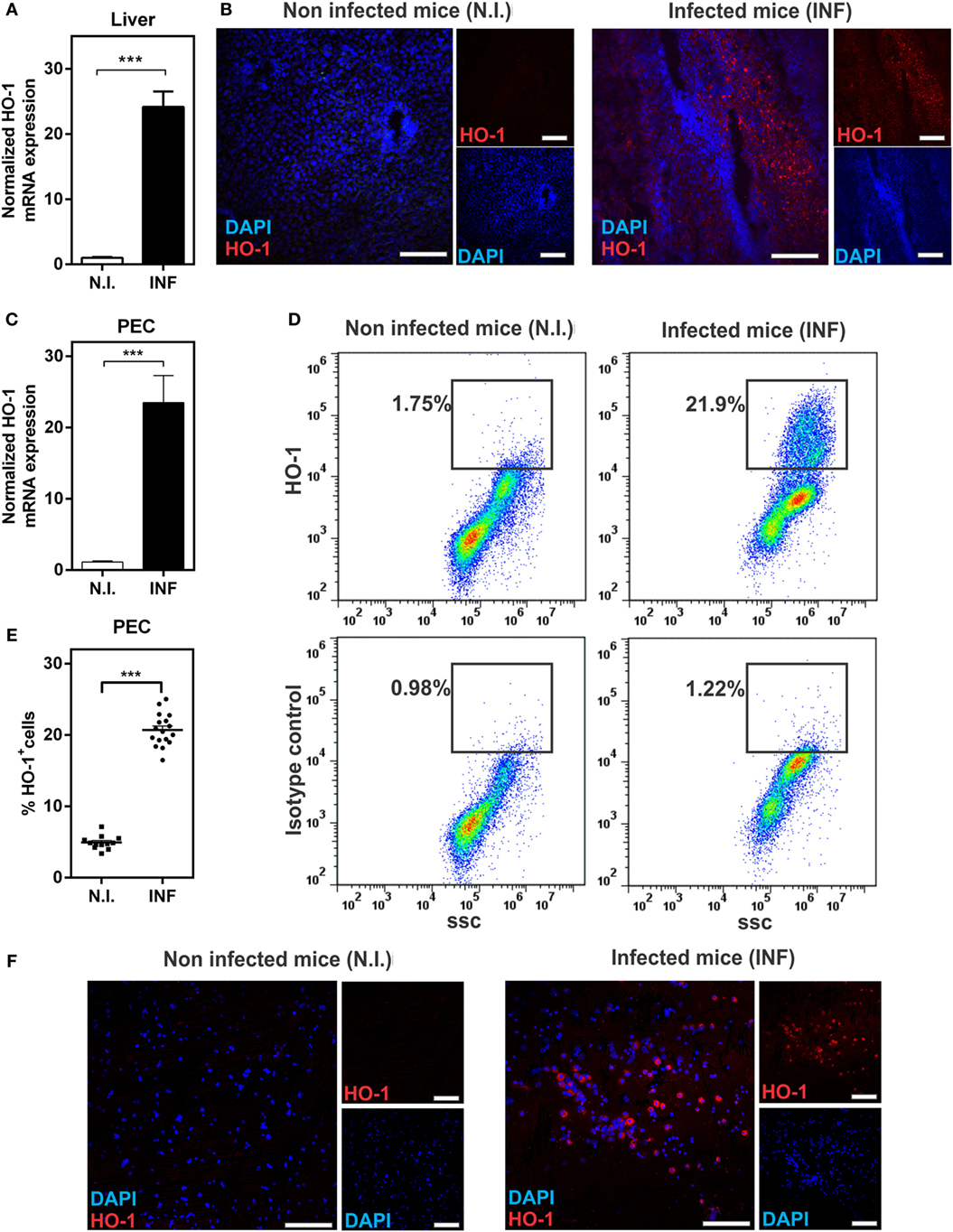
Figure 1. Heme-oxygenase-1 (HO-1) is induced during Fasciola hepatica experimental infection in mice. (A) mRNA expression of HO-1 in the liver from control and F. hepatica-infected mice at 3 wpi. (B) HO-1 expression in the liver from control and infected mice at 3 wpi by confocal microscopy. (C) mRNA expression of HO-1 in peritoneal exudate cell (PECs) from control and infected mice at 3 wpi. (D) HO-1+ cells in PECs from control and infected mice at 3 wpi by flow cytometry. (E) Percentage of HO-1+ cells in PECs from control and infected mice at 3 wpi by flow cytometry. (F) HO-1 expression in PECs from control and infected mice at 3 wpi by confocal microscopy. (F) mRNA expression of HO-1 in the spleen from control and F. hepatica-infected mice at 3 wpi. The figures represent the results of three independent experiments (±SEM, indicated by error bars). Mice were analyzed individually: control mice n = 12 and infected mice n = 17. Asterisks indicate statistically significant differences (***p < 0.001). The bar represents 100 µm.
The gene expression of HO-1 was also investigated in bovine livers (Figure 2) revealing an increase of HO-1 mRNA levels in livers from infected bovine with respect to non-infected animals (Figure 2A). This increase in HO-1 gene expression was confirmed at the protein level by microscopy (Figure 2B). HO-1 was expressed both in the hepatocytes (larger cells) and the infiltrated leukocytes (smaller cells) in livers from infected mice (Figure 2B). Altogether, these results indicate that HO-1 expression increases both in liver and PEC, but not in spleen, of F. hepatica-infected animals.
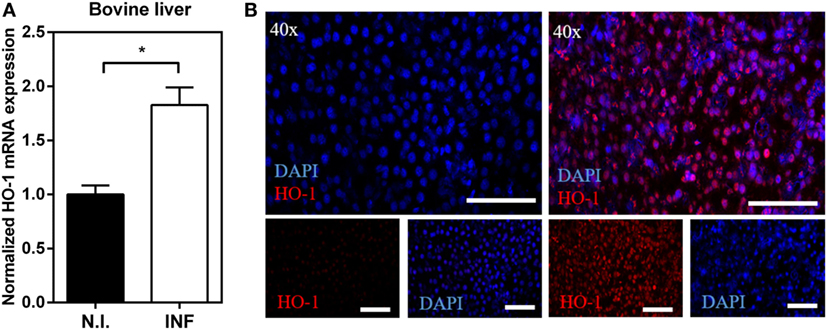
Figure 2. Upregulation of heme-oxygenase-1 (HO-1) expression in liver from Fasciola hepatica-infected bovines. (A) mRNA expression of HO-1 in the liver from F. hepatica-infected (n = 7) and control (n = 5) animals. (B) HO-1 expression in the liver from infected and control animals by confocal microscopy. Asterisks indicate statistically significant differences (*p < 0.05). The bar represents 100 µm.
The Pharmacological Inhibition or Induction of HO-1 Affects Clinical Signs Associated to F. hepatica Infection
The use of pharmacological agents and genetic probes to manipulate HO-1 has been widely used as a tool to explore the role of HO-1 in infections and other pathological systems, as well as its immune regulatory properties. Thus, we investigated whether the pharmacological induction or inhibition of HO-1, using cobalt (CoPP) and tin (SnPP) protoporphyrin IX, respectively, increased or ameliorated the clinical signs associated with by F. hepatica infection. The treatment consisted of five i.p. administrations of CoPP or SnPP at days −2, 2, 5, 12, and 17, with infection at day 0 (Figure 3A). Importantly, CoPP administration lead to a significant increase of HO-1 transcript levels, while SnPP administration did not change the HO-1 gene expression (Figures S2A,B in Supplementary Material) Clinical signs were determined by two different read outs: (i) hepatic damage followed by ALT activity in serum, a common marker to detect hepatic dysfunction (30), and (ii) general state of the animal by a defined clinical score (Figure 3). The clinical score was defined according the parameters described in Table 1. First, we evaluated the HO-1 transcript levels in livers and PECs from treated mice at 2 wpi, time were the highest differences in HO-1 expression were determined. Infected mice expressed high transcript levels of HO-1, both in liver (Figure 3B) and PEC (Figure 3C). Furthermore, when infected mice were treated with CoPP, they presented higher HO-1 transcript levels than infected mice in both biological samples, while SnPP-treatment dramatically reduced the gene expression of HO-1 in infected mice, both in liver (Figure 3B) and PEC (Figure 3C). Of note, when infected mice were treated with simultaneous administration of CoPP and SnPP, the HO-1 transcript levels both in PEC and liver were similar to those found for infected control mice (Figures 3B,C).
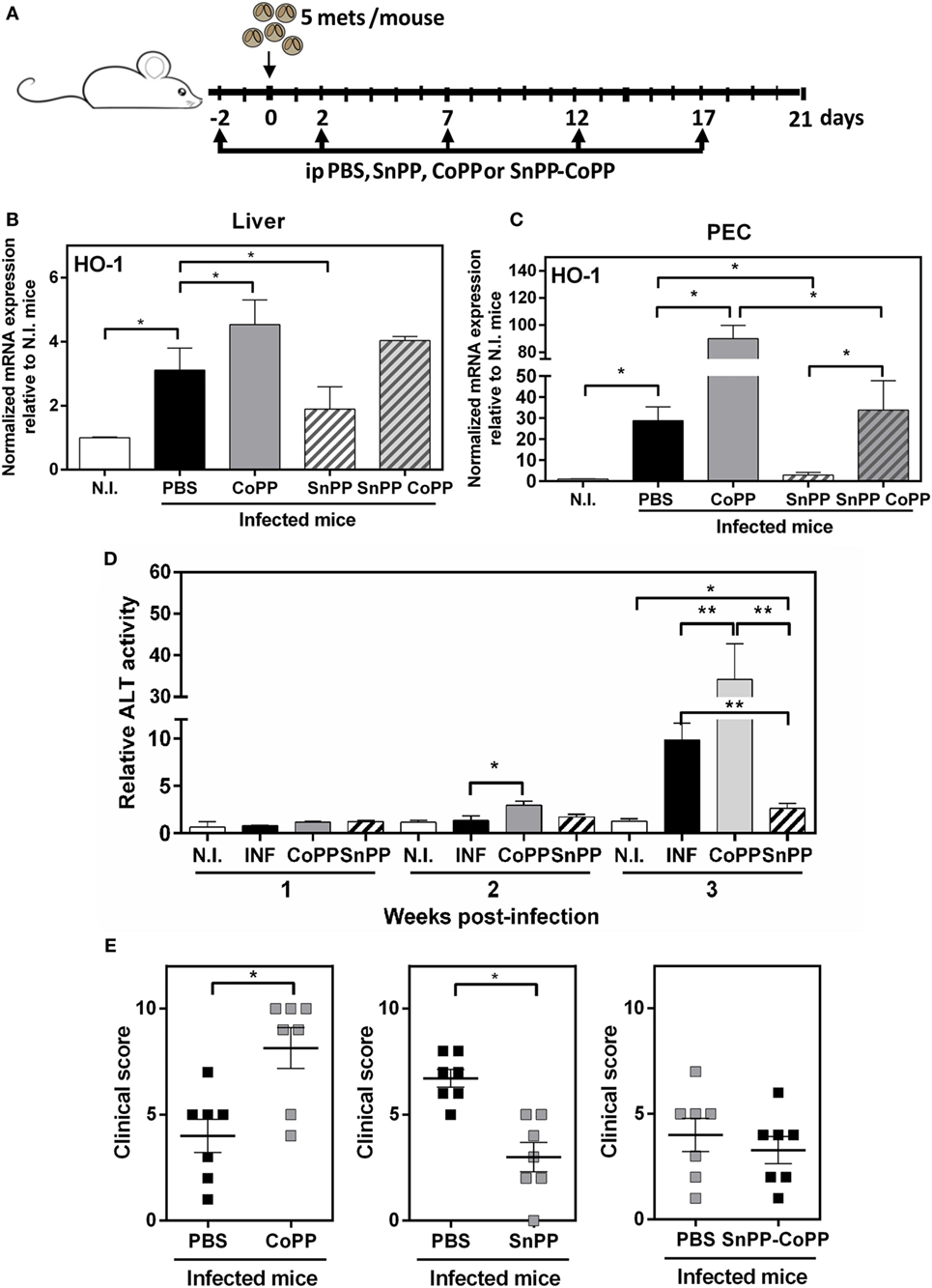
Figure 3. Pharmacological induction and inhibition of heme-oxygenase-1 (HO-1) alters the clinical signs associated with Fasciola hepatica infection. (A) Treatment of infected mice with CoPP, SnPP, SnPP/CoPP, or PBS (control). (B) mRNA expression of HO-1 in liver from CoPP-, SnPP-, and SnPP/CoPP-treated F. hepatica-infected mice at 3 wpi. (C) mRNA expression of HO-1 in peritoneal exudate cells (PECs) from CoPP-, SnPP-, and SnPP/CoPP-treated F. hepatica-infected mice at 3 wpi. (D) Alanine aminotransferase (ALT) activity was measured in sera from CoPP- and SnPP-treated infected and control mice. (E) Clinical score of CoPP- and SnPP-treated F. hepatica-infected mice at 3 wpi, according to Table 1. The figures represent the results of three independent experiments (±SEM, indicated by error bars). Mice were analyzed individually: CoPP (n = 7), SnPP (n = 7), SnPP/CoPP (n = 7), or PBS (n = 7). Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01).
Importantly, the expression of HO-1 correlated with the ALT activity levels found in sera. Indeed, CoPP-treated infected mice presented higher ALT activity levels in serum at 2 and 3 wpi (Figure 3D). On the contrary, SnPP-treated infected mice, had a remarkable decrease in ALT activity levels at 3 wpi with levels comparable to those of non-infected mice, although they were slightly increased (Figure 3D). The hepatic damage determined as ALT activity in serum found in CoPP-treated infected mice correlated with other clinical signs, such as hemorrhage, splenomegaly and increase in ascites and cells in the peritoneum (Figure 3E). In contrast, SnPP-treated infected mice, presented a decreased clinical score as compared to controls. Importantly, control infected mice treated with a mix of SnPP and CoPP, presented the same clinical score as infected mice not treated with protoporfirins (Figure 3E). Importantly, non-infected mice treated with CoPP did not show either liver damage, changes in ALT activity in sera nor any clinical symptom related to the infection with respect to non-treated mice (Figure S2C,D in Supplementary Material). These results suggest that an increase of HO-1 expression augments the susceptibility of F. hepatica infection, while a decrease in this enzyme provides mice resistance to the infection.
It has been reported that HO-1 regulates the expression of multiple cytokines, and has essentially anti-inflammatory properties (26, 31–34). In order to further study the immune response induced in the group of mice treated with protoporphyrins, we evaluated the transcript levels of a panel of Th2/regulatory molecules that are highly expressed during F hepatica infection: FIZZ-1, IL-4, IL-10, and TGFβ. Indeed, at 2 wpi, livers from infected mice expressed high transcript levels of IL-10, TGFβ and FIZZ-1 (Figure 4A). Interestingly, IL-10 and TGFβ transcript levels were even higher in CoPP-treated infected mice, than control infected mice (Figure 4B). Moreover, SnPP-treated infected mice presented lower mRNA levels of IL-10 and FIZZ-1 than infected control mice (Figure 4B). Consistent with these results, simultaneous treatment with CoPP and SnPP did not induce any change in the mRNA levels of these molecules with respect to control infected mice (Figure 4B). Of note, IL-4 gene expression in liver was not modified either with F. hepatica infection nor the treatment with metal protoporphyrins.
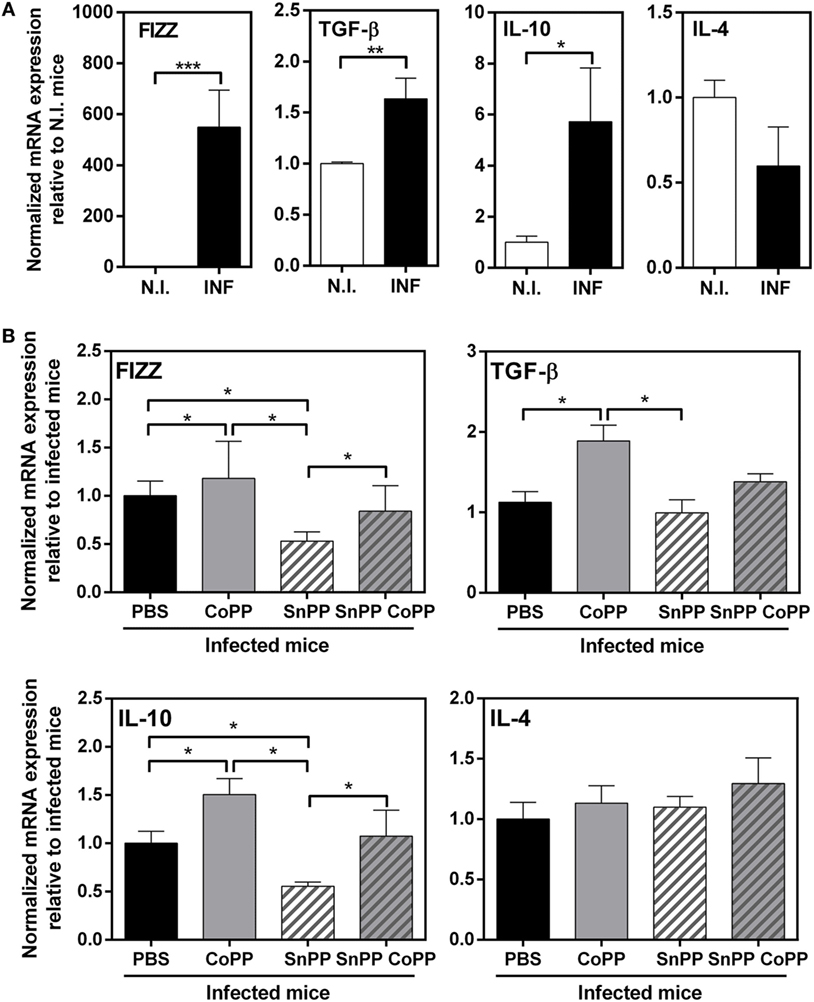
Figure 4. Pharmacological induction of heme-oxygenase-1 expression correlates with IL-10, TGFβ, and FIZZ-1 transcript levels in liver of infected animals. (A) mRNA expression of IL-4, IL-10, TGFβ, and FIZZ-1 in the liver from control and F. hepatica-infected mice at 2 wpi. (B) mRNA expression of IL-4, IL-10, TGFβ, and FIZZ-1 in the liver from CoPP-, SnPP- and SnPP/CoPP-treated F. hepatica-infected mice at 2 wpi. The figures represent the results of three independent experiments (±SEM, indicated by error bars). Mice were analyzed individually: CoPP (n = 7), SnPP (n = 7), SnPP/CoPP (n = 7), or PBS (n = 7). Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001).
Cells from the peritoneum of infected mice, on the other hand, expressed higher transcript levels of TGFβ, IL-4 and FIZZ-1 (Figure 5A), but not IL-10 as shown in liver. Surprisingly, PEC from CoPP- or SnPP-treated mice did not present any change in the expression of either TGFβ, IL-4, FIZZ-1 or IL-10, except for FIZZ-1 which was slightly decreased in SnPP-treated infected mice (Figure 5B), consistent with lower hepatic damage and clinical score.
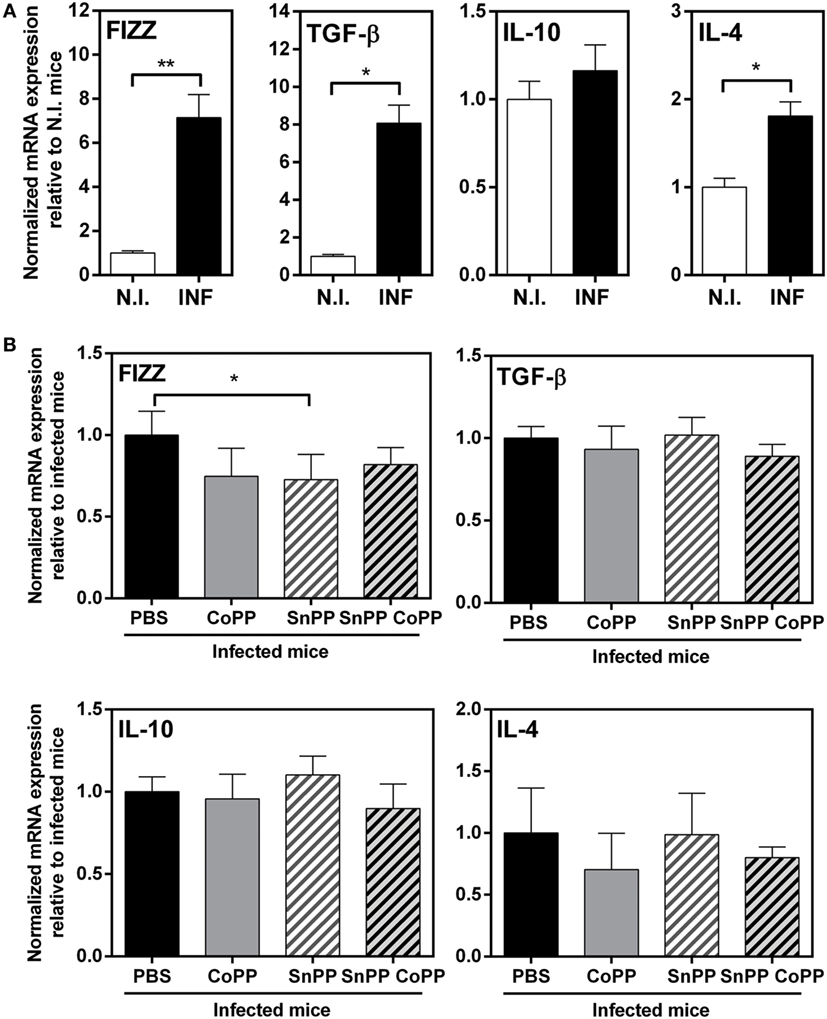
Figure 5. IL-4, IL-10, TGFβ, and FIZZ-1 mRNA levels in peritoneal exudate cells (PECs) from CoPP-, SnPP-, and SnPP/CoPP-infected animals. (A) mRNA expression of IL-4, IL-10, TGFβ, and FIZZ-1 in PEC from control and Fasciola hepatica-infected mice at 2 wpi. (B) mRNA expression of IL-4, IL-10, TGFβ, and FIZZ-1 in PEC from CoPP-, SnPP- and SnPP/CoPP-treated F. hepatica-infected mice at 2 wpi. The figures represent the results of three independent experiments (±SEM, indicated by error bars). Mice were analyzed individually: CoPP (n = 7), SnPP (n = 7), SnPP/CoPP (n = 7), or PBS (n = 7). Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01).
In summary, these results show that the induction of HO-1 is associated with higher levels of the immunoregulatory molecules IL-10 and TGFβ and the reparatory molecule FIZZ-1 in liver, while the inhibition of HO-1correlated with lower expression of TGFβ and FIZZ-1 in the liver of infected animals. However, although infected animals presented increased levels of TGFβ, IL-4 and FIZZ-1 on peritoneal cells, we could not find significant changes associated with the modulation of HO-1. Considering that the peritoneum is essential for F. hepatica juvenile maturation, we studied in further detail peritoneal cells and the expression of HO-1.
HO-1 Is Induced in Two Different Peritoneal F4/80+ Cell Populations
Considering previous reports demonstrating that: (i) F. hepatica-infected mice express high levels of IL-10 (8, 35), (ii) HO-1 expression is related to IL-10 signaling and viceversa (36, 37), (iii) alternatively activated macrophages are associated with F. hepatica infection (9, 38, 39), and (iv) HO-1 is highly expressed by M2 macrophages (40), we sought to evaluate whether the HO-1+ cells identified in F. hepatica-infected mice expressed the molecule F4/80, traditionally used to identify macrophages. As seen in Figure 6A, HO-1+ cells from PECs from infected animals expressed this surface marker. However, two HO-1+ populations were identified according to the expression of HO-1 and F4/80: HO-1int F4/80hi and HO-1hi F4/80int, which significantly augmented upon infection (Figure 6A). Interestingly, although the F4/80hi cell population was also detected in PECs from control mice (Figure 6A), the expression of HO-1 was induced upon infection (Figure 6B). On the contrary, the HO-1hi F4/80int cell population was absent in control mice (Figure 6A) and expressed higher HO-1 levels than HO-1int F4/80hi cells from infected mice (Figure 6B). Of note, the F4/80low population identified in infected mice were Siglec-F+ (could correspond to eosinophils) and did not express HO-1 as determined by the corresponding isotype staining (Figure S3 in Supplementary Material).
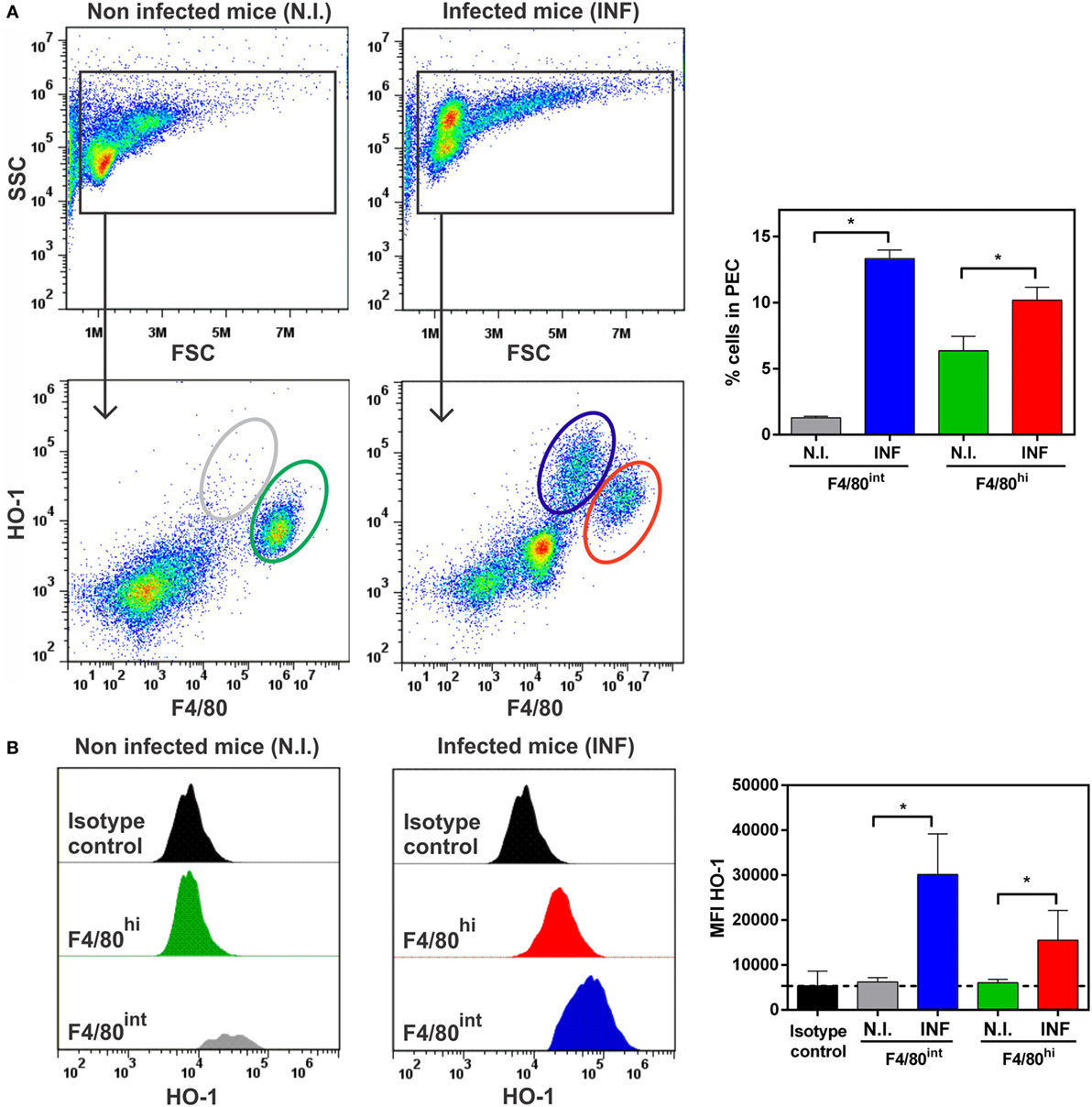
Figure 6. HO-1+ cells recruited in peritoneal exudate cell (PEC) during Fasciola hepatica infection are F4/80+. (A) HO-1+ and F4/80+ cells in PEC from control and infected mice at 3 wpi by flow cytometry. Percentage of HO-1hi F4/80int and HO-1low F4/80hi cells in the peritoneum from control and infected mice at 3 wpi. (B) heme-oxygenase-1 (HO-1) expression in different cell populations from PEC from control and infected mice at 3 wpi. MFI (Mean Fluorescence Intensity) of HO-1 in HO-1hi F4/80int and HO-1low F4/80hi cells from PECs by flow cytometry. The figures represent the results from at least three independent experiments (±SEM, indicated by error bars). Mice were analyzed individually: CoPP (n = 7), SnPP (n = 7), SnPP/CoPP (n = 7), or PBS (n = 7). Asterisks indicate statistically significant differences (*p < 0.05).
The presence of F4/80+ cells in PECs expressing different levels of this surface marker was also confirmed by microscopy, revealing co-localization with HO-1 (Figure 7A). Furthermore, F4/80+ HO-1+ cells were also identified in the leukocyte infiltrate present in livers from infected mice (Figure 7B), while these cells were undetected in control livers (data not shown). HO-1 was also expressed by F4/80- hepatocytes (Figure 7B) as mentioned earlier (Figure 1B).
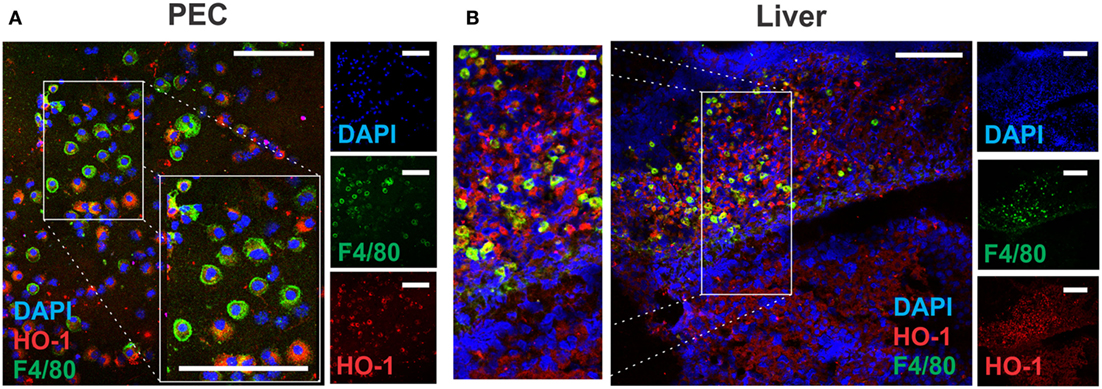
Figure 7. Identification of HO-1+ cells in peritoneal exudate cell (PEC) and liver by confocal microscopy. Heme-oxygenase-1 (HO-1) and F4/80 expression in PEC (A) and liver (B) from control (n = 5) and infected mice (n = 5) at 3 wpi by confocal microscopy. DAPI was used to stain cell nucleus. The bar represents 100 µm. The figures represent the results from at least three independent experiments.
HO-1int F4/80hi and HO-1hi F4/80int Cells from Infected Mice Have Different Phenotype
In order to further characterize HO-1+ cells, we evaluated both populations and HO-1 expression during the process of infection. To this end, PECs and livers were collected at 1, 2, and 3 wpi. Interestingly HO-1 transcript levels augmented progressively with the course of infection (Figure 8A). PECs from infected and control mice were labeled and analyzed by flow cytometry in order to identify both HO-1+ cell populations, HO-1hi F4/80int and HO-1int F4/80hi, and compare them with PECs from control mice. HO-1int F4/80hi cells were already present in control mice and its number doubled from the second week post infection (Figure 8B), time in which they presented increased levels of HO-1 expression (Figure 8B). On the other hand, HO-1hi F4/80int cells in PECs were detected as soon as 1 wpi, and remained constant up to 3 wpi (Figure 8C). The expression of HO-1 by these cells was induced from 2 wpi (Figure 8C). Both the cell number and the HO-1 expression by HO-1int F4/80hi cells remained constant or increased after 2 wpi (Figure 8C).
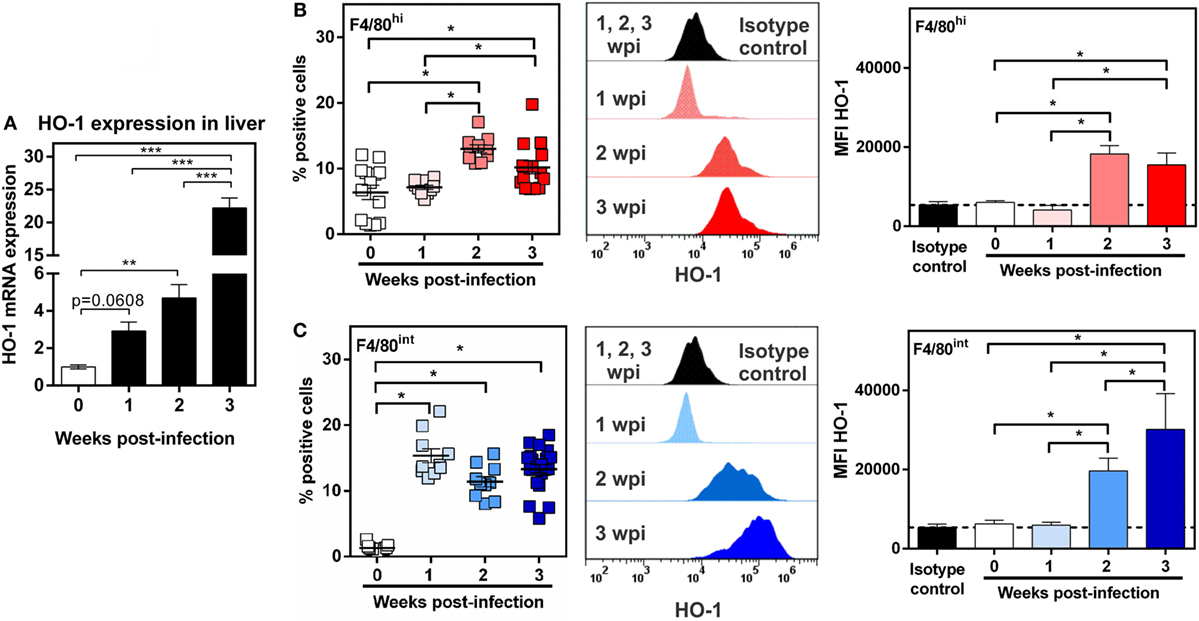
Figure 8. Identification of HO-1+ cells during Fasciola hepatica-infection. (A) mRNA expression of heme-oxygenase-1 (HO-1) in the liver from control (0 wpi) and infected mice (1, 2, and 3 wpi). (B) HO-1+ F4/80hi cells in peritoneal exudate cell (PEC) from control (0 wpi) and infected mice (1, 2, and 3 wpi) by flow cytometry. On the right the HO-1 expression as the MFI of HO-1+ F4/80hi cells is shown. (C) HO-1+ F4/80int cells in PEC from control (0 wpi) and infected mice (1, 2, and 3 wpi) by flow cytometry. On the right, the HO-1 expression as the MFI of HO-1+ F4/80int cells is shown. Mice were analyzed individually: control (n = 12), 1 wpi (n = 10), 2 wpi (n = 10) or 3 wpi (n = 16). Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001).
We then investigated the phenotype of HO-1+ cells by evaluating the expression of different molecules by flow cytometry and compared them to F4/80+ cells found in naïve mice (Figure 9; Figure S4 in Supplementary Material). HO-1int F4/80hi and HO-1hi F4/80int cells expressed CD11b, CD68 and CD172a (SIRPα, Figure 9), all molecules that are expressed by DCs or macrophages (41). However, HO-1int F4/80hi cells expressed higher levels of CD11b, CD68 and CD172a than HO-1hi F4/80int cells. Furthermore, HO-1hi F4/80int cells expressed CD11c while HO-1int F4/80hi expressed Ly6C (Figure 9). Finally, HO-1hi F4/80int cells expressed higher levels of MHC class II but lower expression of CD40 than HO-1int F4/80hi cells. Of note, both cells populations expressed very low levels of Siglec-F (Figure 9). Interestingly, the phenotype described for HO-1int F4/80hi cells resembled to that of peritoneal macrophages from naïve mice (Figure S4 in Supplementary Material). Altogether, these results suggest that HO-1hi F4/80int cells could constitute DCs while HO-1int F4/80hi cells would correspond to monocytes or macrophages.
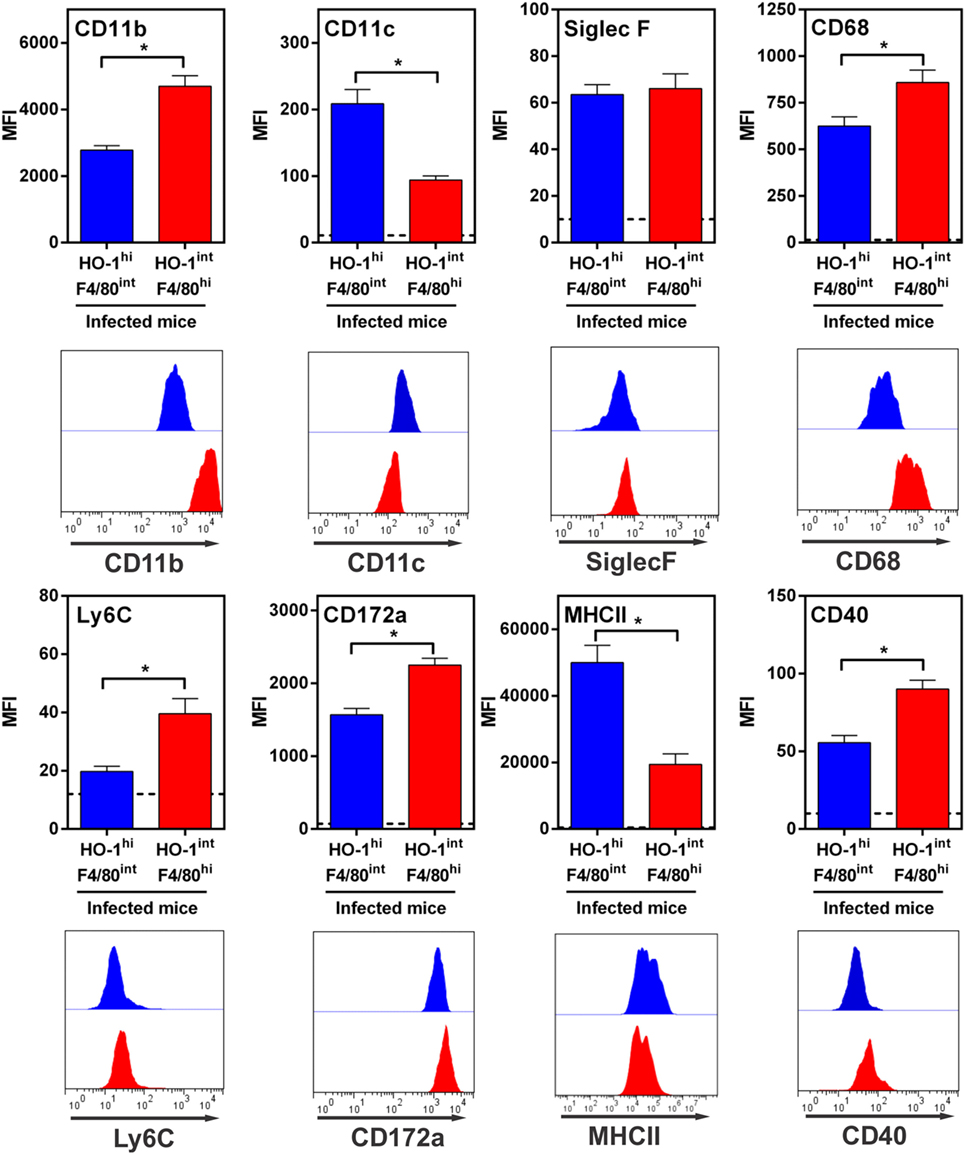
Figure 9. Immunophenotyping of HO-1hi F4/80int and HO-1int F4/80hi in the peritoneum of Fasciola hepatica-infected animals. HO-1hi F4/80int cells (blue), HO-1int F4/80hi cells (red) from peritoneal exudate cells of infected mice at 3 wpi were evaluated for the expression of different cell markers by flow cytometry. Cells from the peritoneal cavity from infected mice were stained with CD11b- CD11c-, MHCII, CD40, SIRPα-, CD68-, Ly6C-, or Siglec-F- specific antibodies and evaluated by flow cytometry. A representative figure of three independent experiments is shown. Mice were analyzed individually: control mice n = 5 and F. hepatica-infected mice n = 5. Asterisks indicate statistically significant differences (*p < 0.05).
To further characterize these cells, we sorted them by flow cytometry and analyzed the gene expression of other molecules by qRT-PCR. In agreement with flow cytometry analyses, HO-1hi F4/80int cells expressed higher transcript levels of HO-1 than HO-1int F4/80hi cells (Figure 10). Interestingly, both cells populations were very different in the set of expressed genes. Indeed, HO-1hi F4/80int cells expressed CD38 and Arg-1, while HO-1int F4/80hi cells did not. On the contrary, HO-1int F4/80hi cells expressed FIZZ-1 and IL-10. Finally, HO-1hi F4/80int cells expressed TGFβ and IL-10 (Figure 10). According to the expression of these markers, these results suggest that HO-1hi F4/80int cells correspond to regulatory or tolerogenic DCs, while HO-1int F4/80hi cells could constitute alternatively activated macrophages.
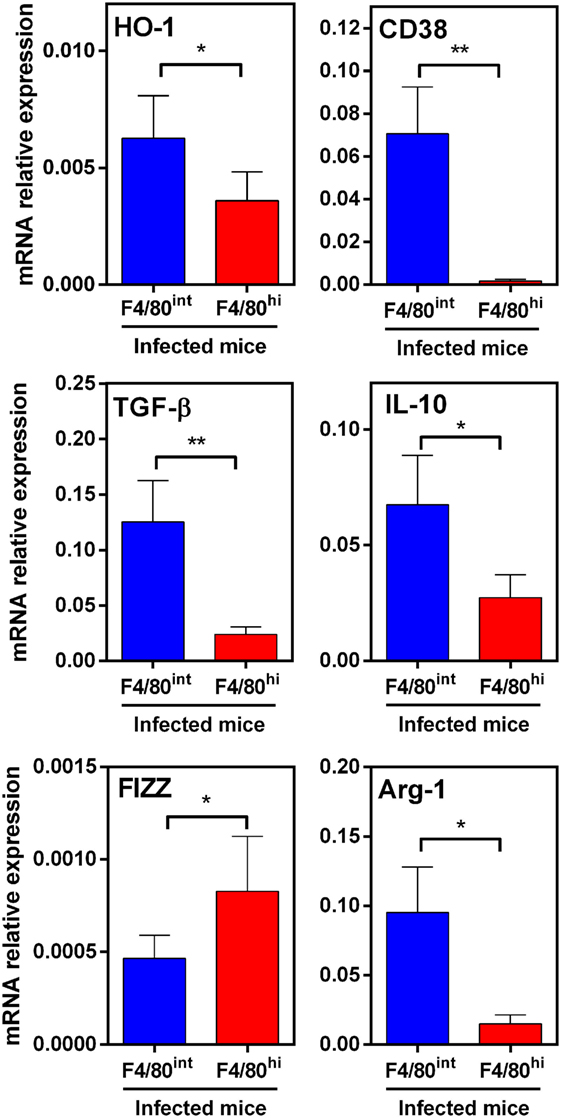
Figure 10. Expression of immunoregulatory molecules in HO-1hi F4/80int and HO-1int F4/80hi in peritoneal exudate cells (PECs) from Fasciola hepatica-infected animals. HO-1hi F4/80int cells (blue), HO-1int F4/80hi cells (red) from PECs of infected mice at 3 wpi were first sorted. Then, the expression of heme-oxygenase-1 (HO-1), TGFβ, IL-10, FIZZ-1, Arg-1, and CD38 was evaluated by qRT-PCR. Gene expression relative to GAPDH transcript levels is shown. Mice were analyzed individually: control mice n = 5 and F. hepatica-infected mice n = 5. Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01).
Discussion
In this work, we show that HO-1 is a key immunoregulatory molecule during F. hepatica infection and that promotes infection and liver damage. The role of HO-1 in infections by intracellular pathogens has been previously approached, demonstrating an upregulation of HO-1 mRNA and/or protein expression in response to viral (25), bacterial (23, 41–46), or protozoan parasitic (18, 19, 47) infections. Furthermore, overexpression or induction of HO-1 promotes persistence of other infectious agents, such as Leishmania chagasi and Plasmodium liver infection (18, 47). However, to our knowledge, this is the first report demonstrating the role of HO-1 in favoring a helminth infection.
The involvement of HO-1 in the anti-inflammatory immune response in F. hepatica-infected mice was confirmed using pharmacological approaches. We show that the pharmacological induction of HO-1 promoted clinical signs associated with F. hepatica infection, and it was correlated with an increase of IL-10 and TGFβ in liver, indicating that the induction of HO-1 is associated with the upregulation of these two immunoregulatory cytokines. The fact that the use of the enzymatic inhibitor of HO-1 SnPP significantly decreased the levels of IL-10, TGFβ, and FIZZ-1 in liver, even to lower levels to control infected mice (for IL-10 and FIZZ-1) strongly suggests that HO-1 is involved in the upregulation of IL-10, promoting parasite survival, and hence liver damage that leads to the upregulation of FIZZ-1 indicating liver fibrosis. Indeed, several studies have demonstrated that HO-1 mediates the anti-inflammatory effect of IL-10 (37, 48) showing that the use of competitive inhibitors or the knock down expression of HO-1 abrogated the suppressive effect of IL-10. In our model, this hypothesis is in agreement with the results obtained with the simultaneous administration of CoPP and SnPP, obtaining similar clinical signs and IL-10, TGFβ, and FIZZ-1 levels as non-treated mice. Further studies are needed to define which of the heme degradation products following the action of HO-1 activity iron, biliverdin, or CO, are responsible for these actions, as has been previously reported for other pathogens (23). Alternatively, cytokine induction may be due to direct interaction of HO-1 with other host molecules. Interestingly, HO-1 gene expression is regulated at the transcriptional level, by several transcriptional factors including activator protein-1 (49, 50), nuclear factor erythroid 2-related factor-2 (NRF2), nuclear factor-kappa B (50, 51), among others. Also, HO-1 expression is regulated by signaling cascades such as mitogen-activated protein kinase and phosphatidylinositol 3-kinase/Akt (49, 52). In our model, the identification of the molecular mechanisms that lead to HO-1 upregulation in F. hepatica-infected animals will eventually contribute to the development of molecular strategies to control the infection.
Apart from its immunoregulatory properties, HO-1 also plays a significant role in inhibiting oxidant-induced injury during inflammatory processes (53). In fact, an appropriate balance of the inflammatory and redox states is essential to resolve most infections and finally the infectious process (17). In this context, another possibility is that F. hepatica induces HO-1 expression not only to evade the host immune response, but also to inhibit oxidant production by macrophages or other cells. One immunologically relevant place in the host for F. hepatica, is the peritoneal cavity, where the production of oxygen or nitrogen derived molecules could limit and restrain juvenile parasites. Indeed, lower levels of liver damage have been suggested to be the consequence of effective killing of the invading parasites within the peritoneum or shortly after reaching the liver (54). In this context, F. hepatica-mediated HO-1 induction might support parasite survival, for instance by favoring its passage through the peritoneum to the liver.
Our data indicate that in the peritoneal cavity two different populations of antigen presenting cells express HO-1. Judged by the high expression of CD11c, CD38, MHCII and the immunoregulatory cytokines IL-10 and TGFβ, HO-1hi F4/80int cells could constitute tolerogenic myeloid-derived DCs (55) or regulatory DCs that potentially participate in the induction of specific regulatory or anergic T cells (8, 56). Indeed, DCs conditioned with parasite-derived molecules can induce T cell anergy (8, 14, 56, 57). It remains to be determined whether HO-1-expressing DCs can induce specific anergic or regulatory T cells in a HO-1 dependent mechanism. In contrast, HO-1int F4/80hi cells were characterized by the high expression of CD68, CD172a, Ly6C, CD11b, and FIZZ-1, as well as low levels of MHCII expression, indicating that they may correspond to alternatively activated macrophages (58). In this line, the alternative activation of macrophages by F. hepatica or its derived molecules has been previously described (10, 39, 59, 60). Macrophages play a central role in innate immune responses toward both extracellular and intracellular pathogens, particularly through the formation of reactive oxygen/nitrogen species (RO/NS) (61, 62). Indeed, oxidative stress can kill F. hepatica flukes by a mechanism that may involve oxidation of proteins or lipids from parasite tegument since peroxyntrite or superoxide radicals significantly diminished parasite viability in vitro (54, 63). Moreover, RO/NS can effectively target extracellular pathogens through the formation of extracellular traps (61). Taking into account that HO-1 in macrophages limits the production of reactive species (34) and induces IL-10 producing anti-inflammatory macrophages (64) and that F. hepatica favors the alternative activation of macrophages (65, 66), it is likely that HO-1+ macrophages at early stages allow F. hepatica survival in the peritoneum through ineffective free radical production.
Finally, the role of HO-1 in favoring F. hepatica infection in the natural host (e.g., livestock, human) remains unknown. Although we show preliminary data demonstrating an increase in HO-1 expression in livers from naturally infected cattle, further studies are necessary to determine whether HO-1 expression correlates with a certain stage of the infection or if participates in the immunoregulatory or anti-oxidant mechanisms during the infection in these hosts.
In conclusion, HO-1 overexpression benefits F. hepatica infection increasing clinical signs and liver damage. Upregulation of HO-1 leads to an increase of IL-10 which could promote and benefit parasite transport from the peritoneum to the liver. On the other hand, an enzymatic inhibitor of HO-1 provided mice with resistance to infection, decreasing IL-10 and FIZZ-1 transcript levels in liver. Although the mechanisms by which HO-1+ DCs or macrophages regulate the expression of IL-10 or oxidative responses during F. hepatica infection remain to be elucidated, targeting HO-1 to control fasciolosis could constitute an interesting alternative strategy to drugs or vaccines against fasciolosis.
Ethics Statement
Mouse experiments were carried out in accordance with strict guidelines from the National Committee on Animal Research (Comisión Nacional de Experimentación Animal, CNEA, National Law 18.611, Uruguay) according to the international statements on animal use in biomedical research from the Pan American Health Organization (PAHO) and World Health Organization (WHO). The protocol was approved by the Uruguayan Committee on Animal Research. Cow’s livers were collected during the routine work of a local abattoir (Frigorífico Carrasco) in Montevideo (Uruguay). All procedures involving animals were approved by the Universidad de la República’s Committee on Animal Research (Comisión Honoraria de Experimentación Animal, CHEA Protocol Number: 070153-000180-16).
Author Contributions
PC performed the experiments, analyzed data, and contributed with manuscript revision. ER, VC and SF contributed with mouse infections and flow cytometry experiments. VN and NB participated in obtention of flukes and extract preparation and detoxification. CR contributed with reactifs and participated in real-time RT-PCR experiments. IA contributed with reactifs, designed experiments with protoporphirn treatment and helped with manuscript revision. TF contributed to supervision and design of all experiments shown in this paper, analyzed data, and prepared the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are particularly grateful to abattoir “Frigorífico Carrasco” for their help with the collection of flukes and cow’s livers. We also thank Eduardo Osinaga and Marcelo Hill for their valuable advice.
Funding
PC was funded by “Agencia Nacional de Investigación e Innovación” (ANII) and “Asociación de Universidades Grupo Montevideo” from Uruguay. IA was funded by the Labex IGO project (no. ANR-11-LABX-0016-01) which is part of the «Investissements d’Avenir» French Government program managed by the ANR (ANR-11-LABX-0016-01) and by the IHU-Cesti project funded also by the «Investissements d’Avenir» French Government program, managed by the French ANR (ANR-10-IBHU-005). The IHU-Cesti project is also supported by Nantes Métropole and Région Pays de la Loire. TF received a grant from Comisión Sectorial de Investigación Científica (Uruguay), CSIC I+D 2016 ID114.
Supplementary Material
The Supplementary Material for this article can be found online at http://journal.frontiersin.org/article/10.3389/fimmu.2017.00883/full#supplementary-material.
References
1. Rondelaud D, Vignoles P, Abrous M, Dreyfuss G. The definitive and intermediate hosts of Fasciola hepatica in the natural watercress beds in central France. Parasitol Res (2001) 87:475–8. doi:10.1007/s004360100385
2. Dietrich C, Kabaalioglu A, Brunetti E, Richter J. Fasciolosis. Z Gastroenterol (2015) 53:285–90. doi:10.1055/s-0034-1385728
3. Vukman KV, Adams PN, O’Neill SM. Fasciola hepatica tegumental coat antigen suppresses MAPK signalling in dendritic cells and up-regulates the expression of SOCS3. Parasite Immunol (2013) 35:234–8. doi:10.1111/pim.12033
4. Dowling DJ, Hamilton CM, Donnelly S, La Course J, Brophy PM, Dalton J, et al. Major secretory antigens of the helminth Fasciola hepatica activate a suppressive dendritic cell phenotype that attenuates Th17 cells but fails to activate Th2 immune responses. Infect Immun (2010) 78:793–801. doi:10.1128/IAI.00573-09
5. Falcón C, Carranza F, Martínez FF, Knubel CP, Masih DT, Motrán CC, et al. Excretory-secretory products (ESP) from Fasciola hepatica induce tolerogenic properties in myeloid dendritic cells. Vet Immunol Immunopathol (2010) 137:36–46. doi:10.1016/j.vetimm.2010.04.007
6. Falcon CR, Carranza FA, Aoki P, Motran CC, Cervi L. Adoptive transfer of dendritic cells pulsed with Fasciola hepatica antigens and lipopolysaccharides confers protection against fasciolosis in mice. J Infect Dis (2012) 205:506–14. doi:10.1093/infdis/jir606
7. Hamilton CM, Dowling DJ, Loscher CE, Morphew RM, Brophy PM, O’Neill SM. The Fasciola hepatica tegumental antigen suppresses dendritic cell maturation and function. Infect Immun (2009) 77:2488–98. doi:10.1128/IAI.00919-08
8. Rodriguez E, Carasi P, Frigerio S, da Costa V, van Vliet S, Noya V, et al. Fasciola hepatica immune regulates CD11c+ cells by interacting with the macrophage Gal/GalNAc lectin. Front Immunol (2017) 8:264. doi:10.3389/fimmu.2017.00264
9. Adams PN, Aldridge A, Vukman KV, Donnelly S, O’Neill SM. Fasciola hepatica tegumental antigens indirectly induce an M2 macrophage-like phenotype in vivo. Parasite Immunol (2014) 36:531–9. doi:10.1111/pim.12127
10. Figueroa-Santiago O, Espino AM. Fasciola hepatica fatty acid binding protein induces the alternative activation of human macrophages. Infect Immun (2014) 82:5005–12. doi:10.1128/IAI.02541-14
11. Alvarado R, To J, Lund ME, Pinar A, Mansell A, Robinson MW, et al. The immune modulatory peptide FhHDM-1 secreted by the helminth Fasciola hepatica prevents NLRP3 inflammasome activation by inhibiting endolysosomal acidification in macrophages. FASEB J (2017) 31:85–95. doi:10.1096/fj.201500093R
12. Sulaiman AA, Zolnierczyk K, Japa O, Owen JP, Maddison BC, Emes RD, et al. A trematode parasite derived growth factor binds and exerts influences on host immune functions via host cytokine receptor complexes. PLoS Pathog (2016) 12:e1005991. doi:10.1371/journal.ppat.1005991
13. Haçarız O, Akgün M, Kavak P, Yüksel B, Sağıroğlu MŞ. Comparative transcriptome profiling approach to glean virulence and immunomodulation-related genes of Fasciola hepatica. BMC Genomics (2015) 16:366. doi:10.1186/s12864-015-1539-8
14. Guasconi L, Chiapello LS, Masih DT. Fasciola hepatica excretory-secretory products induce CD4+T cell anergy via selective up-regulation of PD-L2 expression on macrophages in a Dectin-1 dependent way. Immunobiology (2015) 220:934–9. doi:10.1016/j.imbio.2015.02.001
15. Stempin CC, Motrán CC, Aoki MP, Falcón CR, Cerbán FM, Cervi L. PD-L2 negatively regulates Th1-mediated immunopathology during Fasciola hepatica infection. Oncotarget (2016) 7:77721–31. doi:10.18632/oncotarget.12790
16. Brouard S, Otterbein LE, Anrather J, Tobiasch E, Bach FH, Choi AMK, et al. Carbon monoxide generated by heme oxygenase 1 suppresses endothelial cell apoptosis. J Exp Med (2000) 192:1015–26. doi:10.1084/jem.192.7.1015
17. Chung SW, Hall SR, Perrella MA. Role of haem oxygenase-1 in microbial host defence. Cell Microbiol (2009) 11:199–207. doi:10.1111/j.1462-5822.2008.01261.x
18. Epiphanio S, Mikolajczak SA, Gonçalves LA, Pamplona A, Portugal S, Albuquerque S, et al. Heme oxygenase-1 is an anti-inflammatory host factor that promotes murine plasmodium liver infection. Cell Host Microbe (2008) 3:331–8. doi:10.1016/j.chom.2008.04.003
19. Pamplona A, Ferreira A, Balla J, Jeney V, Balla G, Epiphanio S, et al. Heme oxygenase-1 and carbon monoxide suppress the pathogenesis of experimental cerebral malaria. Nat Med (2007) 13:703–10. doi:10.1038/nm1586
20. Silva-Gomes S, Appelberg R, Larsen R, Soares MP, Gomes MS. Heme catabolism by heme oxygenase-1 confers host resistance to Mycobacterium infection. Infect Immun (2013) 81:2536–45. doi:10.1128/IAI.00251-13
21. Tachibana M, Hashino M, Nishida T, Shimizu T, Watarai M. Protective role of heme oxygenase-1 in Listeria monocytogenes-induced abortion. PLoS One (2011) 6(9):e25046. doi:10.1371/journal.pone.0025046
22. Seixas E, Gozzelino R, Chora A, Ferreira A, Silva G, Larsen R, et al. Heme oxygenase-1 affords protection against noncerebral forms of severe malaria. Proc Natl Acad Sci U S A (2009) 106:15837–42. doi:10.1073/pnas.0903419106
23. Onyiah JC, Sheikh SZ, Maharshak N, Steinbach EC, Russo SM, Kobayashi T, et al. Carbon monoxide and heme oxygenase-1 prevent intestinal inflammation in mice by promoting bacterial clearance. Gastroenterology (2013) 144:789–98. doi:10.1053/j.gastro.2012.12.025
24. Araujo ECB, Barbosa BF, Coutinho LB, Barenco PVC, Sousa LA, Milanezi CM, et al. Heme oxygenase-1 activity is involved in the control of Toxoplasma gondii infection in the lung of BALB/c and C57BL/6 and in the small intestine of C57BL/6 mice. Vet Res (2013) 44:89. doi:10.1186/1297-9716-44-89
25. Espinoza J, León M, Céspedes P, Gómez P, Canedo-Marroquín G, Riquelme S, et al. Heme oxygenase-1 modulates human respiratory syncytial virus replication and lung pathogenesis during infection. J Immunol (2017) 199(1):212–23. doi:10.4049/jimmunol.1601414
26. Chauveau C, Rémy S, Royer PJP, Hill M, Tanguy-royer S, Hubert F-X, et al. Heme oxygenase-1 expression inhibits dendritic cell maturation and proinflammatory function but conserves IL-10 expression. Blood (2005) 106:1694–702. doi:10.1182/blood-2005-02-0494
27. Sierra-Filardi E, Vega MA, Sánchez-Mateos P, Corbí AL, Puig-Kröger A. Heme oxygenase-1 expression in M-CSF-polarized M2 macrophages contributes to LPS-induced IL-10 release. Immunobiology (2010) 215:788–95. doi:10.1016/j.imbio.2010.05.020
28. Paul G, Bataille F, Obermeier F, Bock J, Klebl F, Strauch U, et al. Analysis of intestinal haem-oxygenase-1 (HO-1) in clinical and experimental colitis. Clin Exp Immunol (2005) 140:547–55. doi:10.1111/j.1365-2249.2005.02775.x
29. Sardana MK, Kappas A. Dual control mechanism for heme oxygenase: tin(IV)-protoporphyrin potently inhibits enzyme activity while markedly increasing content of enzyme protein in liver. Proc Natl Acad Sci U S A (1987) 84:2464–8. doi:10.1073/pnas.84.8.2464
30. Liu Z, Que S, Xu J, Peng T. Alanine aminotransferase-old biomarker and new concept: a review. Int J Med Sci (2014) 11:925–35. doi:10.7150/ijms.8951
31. Nitti M, Piras S, Marinari U, Moretta L, Pronzato M, Furfaro A. HO-1 induction in cancer progression: a matter of cell adaptation. Antioxidants (Basel) (2017) 6:29. doi:10.3390/antiox6020029
32. Espinoza JA, González PA, Kalergis AM. Modulation of antiviral immunity by heme oxygenase-1. Am J Pathol (2017) 187(3):487–93. doi:10.1016/j.ajpath.2016.11.011
33. Loboda A, Damulewicz M, Pyza E, Jozkowicz A, Dulak J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases : an evolutionarily conserved mechanism. Cell Mol Life Sci (2016) 73:3221–47. doi:10.1007/s00018-016-2223-0
34. Blancou P, Tardif V, Simon T, Rémy S, Carreño L, Kalergis A, et al. Immunoregulatory properties of heme oxygenase-1. In: Cuturi MC, Anegon I, editors. Methods in Molecular Biology Methods in Molecular Biology. Totowa, NJ: Humana Press (2010). p. 247–68.
35. Rodríguez E, Noya V, Cervi L, Chiribao ML, Brossard N, Chiale C, et al. Glycans from Fasciola hepatica modulate the host immune response and TLR-induced maturation of dendritic cells. PLoS Negl Trop Dis (2015) 9:e0004234. doi:10.1371/journal.pntd.0004234
36. Chen S, Kapturczak MH, Wasserfall C, Glushakova OY, Campbell-Thompson M, Deshane JS, et al. Interleukin 10 attenuates neointimal proliferation and inflammation in aortic allografts by a heme oxygenase-dependent pathway. Proc Natl Acad Sci U S A (2005) 102:7251–6. doi:10.1073/pnas.0502407102
37. Lee T-S, Chau L-Y. Heme oxygenase-1 mediates the anti-inflammatory effect of interleukin-10 in mice. Nat Med (2002) 8:240–6. doi:10.1038/nm0302-240
38. Flynn RJ, Mulcahy G. Possible role for Toll-like receptors in interaction of Fasciola hepatica excretory/secretory products with bovine macrophages. Infect Immun (2008) 76:678–84. doi:10.1128/IAI.00732-07
39. Flynn RJ, Irwin JA, Olivier M, Sekiya M, Dalton JP, Mulcahy G. Alternative activation of ruminant macrophages by Fasciola hepatica. Vet Immunol Immunopathol (2007) 120:31–40. doi:10.1016/j.vetimm.2007.07.003
40. Weis N, Weigert A, von Knethen A, Brüne B. Heme oxygenase-1 contributes to an alternative macrophage activation profile induced by apoptotic cell supernatants. Mol Biol Cell (2009) 20:1280–8. doi:10.1091/mbc.E08-10-1005
41. Athale J, Ulrich A, Chou MacGarvey N, Bartz RR, Welty-Wolf KE, Suliman HB, et al. Nrf2 promotes alveolar mitochondrial biogenesis and resolution of lung injury in Staphylococcus aureus pneumonia in mice. Free Radic Biol Med (2012) 53:1584–94. doi:10.1016/j.freeradbiomed.2012.08.009
42. Kumar A, Deshane JS, Crossman DK, Bolisetty S, Yan BS, Kramnik I, et al. Heme oxygenase-1-derived carbon monoxide induces the Mycobacterium tuberculosis dormancy regulon. J Biol Chem (2008) 283(26):18032–9. doi:10.1074/jbc.M802274200
43. MacGarvey NC, Suliman HB, Bartz RR, Fu P, Withers CM, Welty-Wolf KE, et al. Activation of mitochondrial biogenesis by heme oxygenase-1-mediated NF-E2-related factor-2 induction rescues mice from lethal Staphylococcus aureus sepsis. Am J Respir Crit Care Med (2012) 185(8):851–61. doi:10.1164/rccm.201106-1152OC
44. Regev D, Surolia R, Karki S, Zolak J, Montes-Worboys A, Oliva O, et al. Heme oxygenase-1 promotes granuloma development and protects against dissemination of mycobacteria. Lab Investig (2012) 93:868–79. doi:10.1038/labinvest.2012.125
45. Shiloh MU, Manzanillo P, Cox JS. Mycobacterium tuberculosis senses host-derived carbon monoxide during macrophage infection. Cell Host Microbe (2008) 3:323–30. doi:10.1016/j.chom.2008.03.007
46. Nairz M, Theurl I, Ludwiczek S, Theurl M, Mair SM, Fritsche G, et al. The co-ordinated regulation of iron homeostasis in murine macrophages limits the availability of iron for intracellular Salmonella typhimurium. Cell Microbiol (2007) 9:2126–40. doi:10.1111/j.1462-5822.2007.00942.x
47. Luz NF, Andrade BB, Feijó DF, Araújo-Santos T, Quintela GC, Andrade D, et al. Heme oxygenase-1 promotes the persistence of Leishmania chagasi infection. J Immunol (2012) 188(9):4460–7. doi:10.4049/jimmunol.1103072
48. Piantadosi CA, Withers CM, Bartz RR, MacGarvey NC, Fu P, Sweeney TE, et al. Heme oxygenase-1 couples activation of mitochondrial biogenesis to anti-inflammatory cytokine expression. J Biol Chem (2011) 286:16374–85. doi:10.1074/jbc.M110.207738
49. Lu C-Y, Yang Y-C, Li C-C, Liu K-L, Lii C-K, Chen H-W. Andrographolide inhibits TNFα-induced ICAM-1 expression via suppression of NADPH oxidase activation and induction of HO-1 and GCLM expression through the PI3K/Akt/Nrf2 and PI3K/Akt/AP-1 pathways in human endothelial cells. Biochem Pharmacol (2014) 91:40–50. doi:10.1016/j.bcp.2014.06.024
50. Lavrovsky Y, Schwartzman ML, Levere RD, Kappas A, Abraham NG. Identification of binding sites for transcription factors NF-kappa B and AP-2 in the promoter region of the human heme oxygenase 1 gene. Proc Natl Acad Sci U S A (1994) 91:5987–91. doi:10.1073/pnas.91.13.5987
51. Wardyn JD, Ponsford AH, Sanderson CM. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem Soc Trans (2015) 43:621–6. doi:10.1042/BST20150014
52. Li W, Ma F, Zhang L, Huang Y, Li X, Zhang A, et al. Propargyl-cysteine exerts a novel protective effect on methionine and choline deficient diet-induced fatty liver via Akt/Nrf2/HO-1 pathway. Oxid Med Cell Longev (2016) 2016:1–17. doi:10.1155/2016/4690857
53. Chau L-Y. Heme oxygenase-1: emerging target of cancer therapy. J Biomed Sci (2015) 22:1–7. doi:10.1186/s12929-015-0128-0
54. Piedrafita D, Estuningsih E, Pleasance J, Prowse R, Raadsma HW, Meeusen ENT, et al. Peritoneal lavage cells of Indonesian thin-tail sheep mediate antibody-dependent superoxide radical cytotoxicity in vitro against newly excysted juvenile Fasciola gigantica but not juvenile Fasciola hepatica. Infect Immun (2007) 75:1954–63. doi:10.1128/IAI.01034-06
55. Simon T, Pogu J, Rémy S, Brau F, Pogu S, Maquigneau M, et al. Inhibition of effector antigen-specific T cells by intradermal administration of heme oxygenase-1 inducers. J Autoimmun (2017) 81:44–55. doi:10.1016/j.jaut.2017.03.005
56. Aldridge A, O’Neill SM. Fasciola hepatica tegumental antigens induce anergic-like T cells via dendritic cells in a mannose receptor-dependent manner. Eur J Immunol (2016) 46:1180–92. doi:10.1002/eji.201545905
57. Carranza F, Falcón CR, Nuñez N, Knubel C, Correa SG, Bianco I, et al. Helminth antigens enable CpG-activated dendritic cells to inhibit the symptoms of collagen-induced arthritis through Foxp3+ regulatory T cells. PLoS One (2012) 7:e40356. doi:10.1371/journal.pone.0040356
58. Gautier EL, Shay T, Miller J, Greter M, Jakubzick C, Ivanov S, et al. Gene-expression profiles and transcriptional regulatory pathways that underlie the identity and diversity of mouse tissue macrophages. Nat Immunol (2012) 13:1118–28. doi:10.1038/ni.2419
59. Donnelly S, O’Neill SM, Sekiya M, Mulcahy G, Dalton JP. Thioredoxin peroxidase secreted by Fasciola hepatica induces the alternative activation of macrophages. Infect Immun (2005) 73:166–73. doi:10.1128/IAI.73.1.166-173.2005
60. Donnelly S, Stack CM, Neill SMO, Sayed AA, Williams DL, Dalton JP. Helminth 2-Cys peroxiredoxin drives Th2 responses through a mechanism involving alternatively activated macrophages. FASEB J (2008) 22:4022–32. doi:10.1096/fj.08-106278
61. Gostner JM, Becker K, Fuchs D, Sucher R. Redox regulation of the immune response. Redox Rep (2013) 18:88–94. doi:10.1179/1351000213Y.0000000044
62. Singel KL, Segal BH. NOX2-dependent regulation of inflammation. Clin Sci (2016) 130:479–90. doi:10.1042/CS20150660
63. Sadeghi-Gashjin G, Naem S. Parasiticidal effects of peroxynitrite on ovine liver flukes in vitro. J Helminthol (2001) 75:73–6. doi:10.1079/JOH200032
64. Naito Y, Takagi T, Higashimura Y. Heme oxygenase-1 and anti-inflammatory M2 macrophages. Arch Biochem Biophys (2014) 564:83–8. doi:10.1016/j.abb.2014.09.005
65. Reyes JL, Terrazas LI. The divergent roles of alternatively activated macrophages in helminthic infections. Parasite Immunol (2007) 29:609–19. doi:10.1111/j.1365-3024.2007.00973.x
Keywords: helminth, heme-oxigenase-1, immune regulation, dendritic cell, macrophage
Citation: Carasi P, Rodríguez E, da Costa V, Frigerio S, Brossard N, Noya V, Robello C, Anegón I and Freire T (2017) Heme-Oxygenase-1 Expression Contributes to the Immunoregulation Induced by Fasciola hepatica and Promotes Infection. Front. Immunol. 8:883. doi: 10.3389/fimmu.2017.00883
Received: 29 May 2017; Accepted: 11 July 2017;
Published: 26 July 2017
Edited by:
Luciana Balboa, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), ArgentinaReviewed by:
Grace Mulcahy, University College Dublin, IrelandBeatrix Schumak, University of Bonn, Germany
Copyright: © 2017 Carasi, Rodríguez, da Costa, Frigerio, Brossard, Noya, Robello, Anegón and Freire. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Teresa Freire, dGZyZWlyZUBmbWVkLmVkdS51eQ==